


Introduction
Since the abolition of European Union milk quotas, the Irish dairy cow herd has expanded by 41.6 % to 1.6 million cows during the period 2013–2023. Within the same time-period the specialized beef cow herd has decreased by 24.2 % (CSO, 2023). This shift in the composition of the national cow herd has led to an increasing proportion of beef cattle in Ireland being of dairy-origin or ‘dairy-beef’ (AIM, 2024). This is not a unique Irish situation, with the dairy-origin beef making an increasingly important contribution to beef production in countries such as the USA (Boykin et al., Reference Boykin, Eastwood, Harris, Hale, Kerth, Griffin, Arnold, Hasty, Belk and Woerner2017) and New Zealand, where 700 000 dairy-beef animals were raised in 2022 (BLNZ, 2023). Dairy-beef systems have the potential to have greater (Farrell et al., Reference Farrell, Morris, Kenyon and Tozer2024) farm-level profitability than the most technically efficient beef cow-calf production systems (McGee et al., Reference McGee, Regan, Moloney, O’Riordan, Lenehan, Kelly and Crosson2024). Additionally dairy-beef has a lower carbon footprint than beef cow-calf production (Mogensen et al., Reference Mogensen, Kristensen, Nielsen, Spleth, Henriksson, Swensson, Hessle and Vestergaard2015; Taylor et al., Reference Taylor, McGee, Kelly and Crosson2020; van Selm et al., Reference van Selm, de Boer, Ledgard and van Middelaar2021), and grass-based dairy-beef systems are typically net producers of human edible protein (Kearney et al., Reference Kearney, O’Riordan, McGee, Breen and Crosson2022). A reduction in slaughter age of beef animals has been identified as a key greenhouse gas mitigation strategy within the Teagasc Marginal Abatement Cost Curve (Lanigan et al., Reference Lanigan, Black, Donnellan, Crosson, Beausang, Hanrahan, Buckley, Lahart, Herron and Redmond2023), therefore investigating dairy-beef systems which allow greenhouse gas mitigation to be achieved is desirable, however, remaining cognizant that such systems must remain profitable.
A key factor influencing the performance of dairy-beef systems is the beef genetic merit of animals produced from the dairy herd. However, dairy breeding objectives in temperate pasture-based systems tend to select females which are fertile, have a high forage intake (Delaby et al., Reference Delaby, Finn, Grange and Horan2020), and generally place negative emphasis on cow mature weight to reduce maintenance requirements (Berry et al., Reference Berry, Dunne, McHugh, McParland, O’Brien and Twomey2022). In Ireland this has resulted in a decrease in the genetic merit for beef production in dairy-beef animals (Mulhall et al., Reference Mulhall, Sleator, Evans and Twomey2023), with each 10 kg decrease in genetic merit for dairy cow mature body weight reducing progeny carcass weight by 3.8 kg (Mulhall et al., Reference Mulhall, Sleator, Evans, Berry and Twomey2024). To enhance the beef genetic merit of dairy-beef progeny in Ireland, a dairy-beef breeding index to identify beef sires for use on the dairy herd, the dairy-beef index (DBI), has been developed (Berry et al., Reference Berry, Amer, Evans, Byrne, Cromie and Hely2019). Previous studies have shown that selecting high beef genetic merit sires using the DBI, increases progeny carcass weight compared to lower beef genetic merit sires (O’Driscoll et al., Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025); however, a financial and environmental assessment of the potential impacts of this at farm level is required.
Farm systems models offer the opportunity to simulate the consequences of factor changes within a system, thus, determining the robustness of the system to external pressures and increasing producer confidence in new technologies (Janssen and Van Ittersum, Reference Janssen and Van Ittersum2007). The Grange Dairy Beef Systems Model (GDBSM; (Ashfield et al., Reference Ashfield, Crosson and Wallace2013)), is a simulation model of a dairy-beef farm, and has previously been used to investigate the effect of exploiting compensatory growth (Ashfield et al., Reference Ashfield, Wallace, McGee and Crosson2014a), differing Holstein-Friesian beef production systems (Murphy et al., Reference Murphy, Crosson, Kelly and Prendiville2017), and stocking rate (Kearney et al., Reference Kearney, O’Riordan, McGee, Breen, Dunne, French and Crosson2024) on dairy-beef system economic and environmental performance. However, no study to date has quantified the effect of pasture-based concentrate supplementation or sire beef genetic merit with the aim of reducing slaughter age on both the economic and environmental performance in dairy-beef systems.
The objective of this study was to model production data from an animal study involving the progeny of beef sires of divergent beef genetic merit and Holstein-Friesian dams, managed under three differing pasture-based feed treatments (O’Driscoll et al., Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025), to quantify the differences in whole farm profit. Additionally, greenhouse gas emissions from each system were quantified and a food-feed analysis conducted. Results from this analysis will guide dairy and beef producers on the potential profitability, and environmental benefits of using sires of superior beef genetic merit in the dairy herd.
Materials and methods
Bio-economic model
The Grange Dairy Beef Systems Model (GDBSM), is a static, steady-state, biophysical simulation model, developed in Microsoft Excel, which simulates pasture-based dairy-beef production systems and was developed and calibrated by Ashfield et al. (Reference Ashfield, Crosson and Wallace2013) and Kearney et al. (Reference Kearney, O’Riordan, McGee, Breen and Crosson2022). The model can be used to assess the impact of institutional, technical and environmental changes on economic, greenhouse gas emissions and food-feed competition performance of a production system. For the current study, the model was populated using animal intake, growth, carcass, and grassland production data from a multi-year farm systems experiment.
Production study details
The production study modelled in the present study was carried out at Teagasc, Animal & Grassland Research and Innovation Centre, Grange, Co. Meath, Ireland between March 2020 and March 2023, across two full dairy-beef production cycles, and is detailed in O’Driscoll et al. (Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025). In summary, spring born male calves were purchased directly from commercial dairy farms in spring 2020 and 2021. Calves were the progeny of Holstein-Friesian (HF) dams, mated to either Angus, or HF sires. Detailed information on the three genotypes investigated is available in O’Driscoll et al. (Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025). In brief three genotypes were compared, 1) calves produced from nine Angus sires above the sixtieth percentile of the DBI beef sub-index (HA); 2) calves produced from nine Angus sires below the sixtieth percentile of the DBI beef sub-index (LA); and 3) calves produced from thirteen Holstein-Friesian sires (HF). The mean sire DBI and DBI sub-index values for beef, calving and carbon, and predicted transmitting ability for carcass weight, conformation, fat, age at slaughter, dystocia and gestation length are available in supplementary Table S1.
Calves were artificially reared on site as described by O’Driscoll et al. (Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025). Calves were weaned from artificial milk replacer upon reaching a target body weight of 90 kg after which they were assigned, within genotype, to one of three experimental feed treatments (FT) with each treatment balanced for date of birth, body weight, genotype and sire thereby creating a 3 × 3 factorial arrangement. 1) The control (CTL) treatment was designed to represent ‘conventional’ management in Irish steer dairy-beef production systems. 2) Low concentrate (LC) was designed to investigate a strategy to increase growth rate during the first grazing season, and 3) high concentrate (HC) was designed to investigate a strategy aimed at finishing steers without a second housing period, therefore reducing slaughter age. Control calves received no concentrate supplementation during the first summer grazing period (June 1st to September 15th), with LC and HC calves offered 0.85 kg dry matter (DM) concentrate per day during this period. From September 15, all calves across all treatment groups received 0.85 kg DM concentrate/d until approximately November 1, with a feeding rate of 1.25 kg DM concentrate/d thereafter until housing. All steers received a common first winter period indoors, receiving grass silage ad-libitum and concentrate supplementation of 1.25 kg DM per day. Supplementation over the first winter period consisted of a coarse mixture; 740 g/kg rolled barley, 180 g/kg soybean meal, 50 g/kg molasses and 30 g/kg vitamins and minerals. Following turn-out to pasture in the spring (March 8) of the second grazing season, steers in CTL and LC treatments were rotationally grazed on pasture only (no supplementation) from turnout until housing for finishing. Steers in HC were rotationally grazed and offered concentrate supplementation at pasture as follows: from July 1 to August 14 HC steers were offered 2.6 kg DM concentrate per animal, and from August 15 concentrate supplementation increased to 4.2 kg DM kg concentrate per animal per day until finish or housing at the end of the grazing season. Body condition score (BCS) on a scale of 1–5 (1 = emaciated, 5 = obese) with increments of 0.25, was assessed fortnightly during the finishing period by a trained individual and steers were deemed fit for slaughter once they reached a BCS of 3.75 (Lowman et al., Reference Lowman, Scott and Somerville1976). Across all treatments, steers that were not slaughtered directly off pasture in the second grazing season having not met the target BCS of 3.75 were housed for indoor finishing (October 1). During this final indoor finishing period, grass silage was offered daily ad-libitum and supplemented with concentrate gradually increased over a seven-day period to 4.25 kg DM per animal per day. Finishing concentrate consisted of a coarse mixture; 862 g/kg rolled barley, 60 g/kg soybean meal, 50 g/kg molasses and 28 g/kg minerals and vitamins. Dry matter intake during the first winter and indoor finishing period was measured on a per pen basis in a randomized complete block design. Steers assigned to pens were balanced for feed treatment, genotype, sire, date of birth and body weight. With four animals per pen over the first winter period, and six animals per pen during the indoor finishing period. Grass silage was offered daily ad-libitum (0.1 in excess of intake). Refusals were weighed twice weekly and discarded once weekly. Feed intake was measured at pasture using the n-alkane technique (Dillon, Reference Dillon1993), twice during the steers second grazing season in May and July, in both 2021 and 2022. Intake data was used to populate the model to get an accurate measurement of the amount of forage consumed in each system, in-depth analysis of the difference in animal intake can be found in the supplementary material (Table S2).
Details of pasture management including pasture and concentrate sampling, processing and chemical analysis were reported by O’Driscoll et al. (Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025) and are available in the supplementary material (Table S3). Grass growth and herbage availability was recorded and decisions regarding grazing management were aided using the online decision support system, ‘PastureBase Ireland’ (Hanrahan et al., Reference Hanrahan, Geoghegan, O’Donovan, Griffith, Ruelle, Wallace and Shalloo2017).
Animal body weight (kg) was measured using calibrated scales (Tru-Test XR3000, load bars XHD 10 000, Auckland, New Zealand). Average daily gain (ADG) was calculated as the change in body weight over a period divided by the duration of the specified period. Immediately prior to transportation for slaughter, body weight was recorded on all animals. Cold carcass weight, dressing percentage, carcass fat and carcass conformation scores were measured (EUROP classification system, 15-point scale) (Conroy et al., Reference Conroy, Drennan, Kenny and McGee2010).
Model assumptions
Detailed animal and feeding information from O’Driscoll et al. (Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025) were used to populate the GDBSM model, such as animal growth rates, body weight, and slaughter characteristics (Supplementary Tables S4 and S5). The model utilizes production data, such as animal growth, diet offered, measured animal intake, and final slaughter age to calculate the daily feed demand of the system and alters animal numbers purchased at the start of the production cycle according to the forage supply (grazed pasture and grass silage) required to support each production system. Total forage (grazed + conserved) accumulation was 12.5t DM/ha and a farm area of 40 ha, an animal mortality rate of 5 % from 0–12 months and 1 % from 13–24 months was assumed. Feed availability, feed intake, and animal stocking rate were used to determine the output of animal numbers sold per scenario. Variable costs (fertilizer, contractor charges, veterinary fees, transport and feed costs) were based on experimental data from the production study. Price assumptions were based on prevailing market conditions and included straw (4 × 4 bale) at €18; urea fertilizer at €550/t; 0-10-20 (N-P-K compound fertilizer) at €400/t; ground limestone at €35/t; calves concentrate at €420/t; yearling concentrate a €350/t; finishing concentrate at €350/t; and milk replacer at €2100/t. Fixed costs (depreciation, machinery running and maintenance, farm maintenance, car, telephone, electricity and insurance) were based on prevailing costs for dairy-beef systems. The cost of labour was not included, as family labour is typical in Irish beef farm systems and was assumed to be available without cost. Likewise, farmland was assumed to be family-owned with no cost assigned. A calf purchase price of €60 and €200 was assumed for HF and Angus sired calves, respectively. The average beef price was assumed to be €4.85/kg carcass weight which reflected the prevailing price during the study and varied based on seasonality (i.e. date of slaughter), and carcass specifications in respect of carcass conformation and fat score (Kenny et al., Reference Kenny, Murphy, Sleator, Judge, Evans and Berry2020). Additionally, a breed bonus of €0.20/kg for Angus steers that met minimum specifications is typically available for Angus cattle slaughtered in Ireland and this was included to the Angus genotypes in the present study. Using information on input and output costs, the GDBSM calculates gross output (value livestock sales − value livestock purchases), gross margin (gross output − variable costs) and net margin (gross output − (variable + fixed costs)) for each scenario. This study used net margin as a proxy for overall farm profitability. To determine the potential effects of volatility in input costs and beef price, sensitivity analyses were completed including a fluctuation of ±€20/head in calf purchase price, ±€50/t in finishing concentrate price and ±€0.50/kg carcass weight in beef price.
Greenhouse gas emissions were quantified using on-farm (enteric fermentation, manure management and soils emissions) and off-farm (manufacture of inputs such as concentrate feed and fertilizers and indirect nitrous oxide emissions from nitrate leaching and ammonia volatilization) sources. Emissions generated post-farm gate, such as transport for slaughter, slaughter and carcass processing, and beef product distribution were omitted from the analysis, as well as emissions associated with farm buildings and machinery. Soil organic carbon was assumed to be in equilibrium. GHG emissions were quantified based on their CO2 equivalent factors over a 100-year time horizon (1 for CO2, 28 for CH4, and 128 for N2O: IPCC, 2013). The emissions factors used to calculate greenhouse gas emissions are shown in Table S6. Greenhouse gas emissions were expressed per kg of beef carcass weight in the current study.
The GDBSM used the approach of Mosnier et al. (Reference Mosnier, Jarousse, Madrange, Balouzat, Guillier, Pirlo, Mertens, ORiordan, Pahmeyer and Hennart2021) to assess ‘food-feed’ competition (Kearney et al., Reference Kearney, O’Riordan, McGee, Breen and Crosson2022). This was measured using three indicators, (1) the production of human edible protein and human edible energy from the farm system, (2) the conversion efficiency of human edible resources into animal edible products (the ratio of human-edible protein and energy in beef meat to the quantity of human-edible protein and energy in animal feed), whereby an efficiency ratio greater than one indicates a farm system is a net producer of protein/energy, a ratio of less than one indicates a system is a net consumer of protein/energy, and (3) the area of agricultural land required to produce beef carcass (m2/kg carcass). As per Mosnier et al. (Reference Mosnier, Jarousse, Madrange, Balouzat, Guillier, Pirlo, Mertens, ORiordan, Pahmeyer and Hennart2021), one kg of bovine meat was assumed to comprise of 158 g gross protein, and 10.9 mega joules (MJ) of gross energy.
Results
Feed consumed and stocking rate
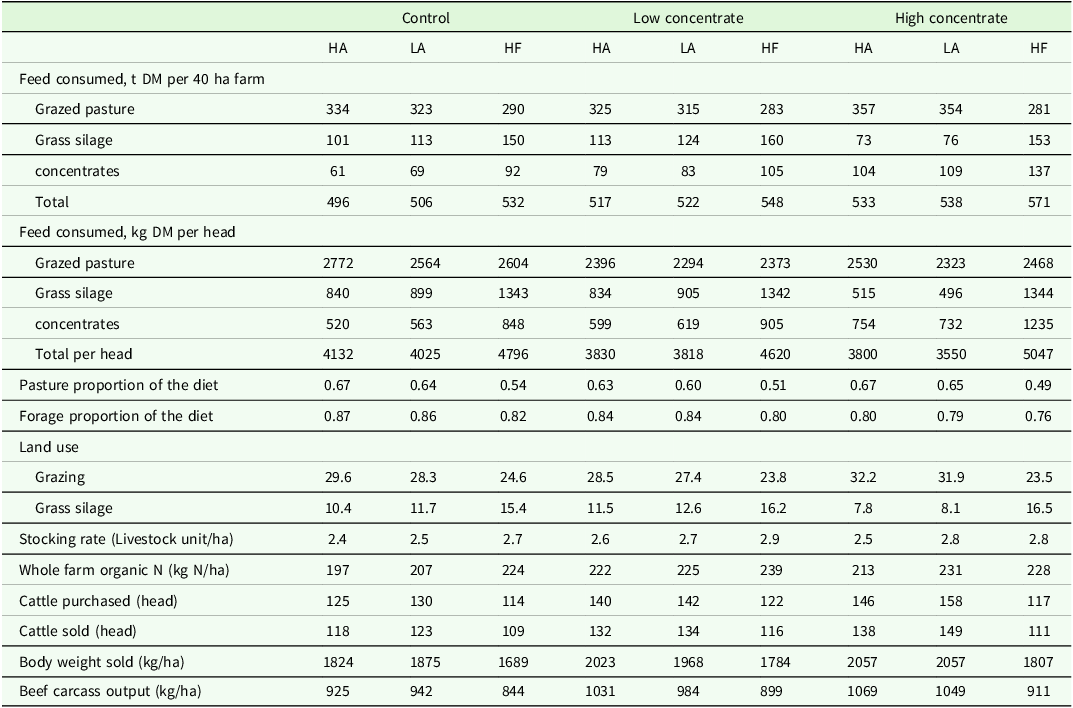
The feed consumed and stocking rate in each of the nine scenarios is shown in Table 1. Across all feed treatments HF steers consumed more feed per head and per farm than both HA and LA. High Angus steers consumed more feed per head than LA steers, but LA scenarios consumed more feed per farm. Based on the lifetime feed budget, HA steers consumed the most grazed forage, due to their greater body weight, and higher total individual feed intake during the first and second grazing season, with LA steers consuming the least grazed forage per head, and HF intermediary.
Beef farm systems results for dairy-beef production systems of contrasting genotype a and feed treatment modelled using the grange dairy beef systems model

a Genotype: HA (High Angus) = Top 40 % sire DBI beef sub-index, LA (Low Angus) = Bottom 60 % sire DBI beef sub-index, HF = Holstein-Friesian.
Across all genotypes, the CTL feed treatment consumed the greatest amount of grazed forage per head, with the proportion of forage (grazed and conserved) in CTL steers diet on average 3 and 7 % higher than LC and HC, respectively. However, the average total feed consumed per farm was lowest for the CTL treatment, 511 t DM per farm, compared to 529 and 548 t DM per farm for LC and HC, respectively, due to a lower SR in CTL than both LC and HC. Across all feed treatments, scenarios using the LA genotype purchased most calves with those utilizing HF genotypes purchasing the least, and HA intermediary. Stocking rate (SR) was lowest for the HA genotype across all FT, with HF having the highest SR across all feed treatments due to the older age at slaughter of HF steers. Stocking rate was lowest within the CTL feed treatment, due to a lower proportion of purchased concentrate within this scenario.
As expected, the greater number of calves purchased in the HA and LA scenarios, led to higher carcass output per hectare compared to HF scenarios (Table 1). Within the CTL scenarios, LA had a higher carcass output per hectare at 942 kg/ha, compared to 925 and 844 kg/ha for HA and HF, respectively. In contrast, under the LC and HC scenarios HA had the highest carcass output per hectare, 5 and 2 %, greater than LA, and 15 and 17 %, greater than HF, respectively (Table 1).
Economic performance
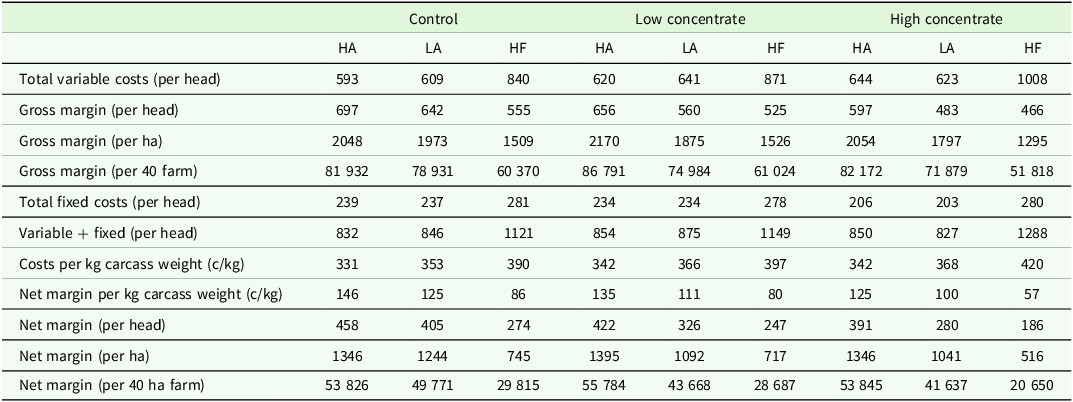
Across the scenarios, gross margin differed by genotype, with HA being greatest, HF being lowest and LA intermediate (Table 2). This ranking was unaffected by feed treatment. Within the CTL scenario, HA had a gross margin per ha of €2048, compared to €1973 for LA, and €1509 for HF, respectively. This trend was followed in both LC and HC, with HA having a 16 % greater gross margin per ha than LA, and a 42 % greater gross margin per ha than HF, in LC and within HC, HA had a 14 % greater gross margin per ha than LA, and a 59 % greater margin per ha than HF.
Financial performance of dairy-beef production systems of contrasting genotype a and feed treatment modelled using the grange dairy beef systems model

Price assumptions: Calf price (€200 for HA (High Angus) and LA (Low Angus), €60 for HF); Beef price €4.85/kg (Steer on EUROP system with a conformation and fat score of 8); Breed bonus €0.20/kg for Angus steers; Straw (4 × 4 bale) €18/bale; Urea fertilizer €550/t; 0-10-20 €400/t; Lime €35/t; Calves concentrate price €420/t; Yearling concentrate price €350/t; Finishing concentrate price €350/t; Milk replacer price €2100/t.
a Genotype: HA (High Angus) = Top 40 % sire DBI beef sub-index, LA (Low Angus) = Bottom 60 % sire DBI beef sub-index, HF = Holstein-Friesian.
On average, across feed treatment, the CTL scenarios had the greatest net margin expressed on both a per head and a per kg carcass weight basis and the lowest total cost per kg of carcass produced across all genotypes (Table 2). The greater net margin associated with the CTL scenario could be explained by the higher proportion of forage in the diet compared to either the LC and HC feed treatments (Table 2). The HA genotype had a similar net margin per hectare across all feed treatments, with both LA and HF genotypes having a greater net margin per ha and per farm within the CTL feed treatment compared to either LC or HC. The greatest differences in net margin per ha were observed within the HC feed treatment, with the HA genotype achieving a net margin per ha of €1346, compared to €1041 and €516 for LA and HF, respectively. The HF genotype in the HC feeding treatment achieved the lowest net margin, with a net margin of €20 650 per 40 ha farm; this resulted in a 37 % lower net margin compared to the most profitable scenario, the HA genotype in LC feeding treatment, which achieved a net margin of €55 784 per 40 ha farm (Table 2).
On average, variable costs accounted for 74 % of total costs across the nine scenarios, however this ranged between genotypes, with HF steer having on average 3 % higher variable costs than both the HA and LA genotypes (Table 2), due to higher concentrate input. Across all scenarios, concentrate feeds were the greatest contributor to variables costs, with concentrate feed accounting for 42 % of variable costs in the CTL feed treatment, 46 % within LC, and 53 % within the HC feed treatment, respectively. Angus genotypes within the HC feed treatment had lower variable costs associated with silage production compared to CTL and LC, as no conserved forage was required for an indoor finishing period. Only small differences in the fixed costs were observed across the three genotypes, although the HF genotypes had greater electricity and machinery operating and depreciation costs due to the longer indoor finishing period required.
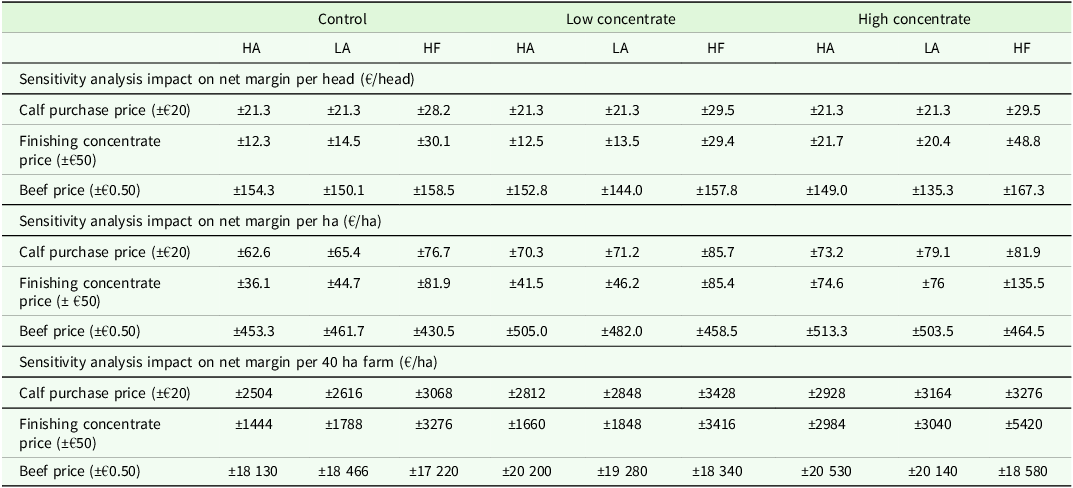
The effect of variations in calf purchase price, concentrate price, and beef price are outlined in Table 3. In all scenarios, the HA genotype remained the most profitable genotype, on both a per head and per hectare basis. Fluctuation in beef price had the largest impact on net margin across all scenarios, equating to an average change in net margin of ±€152 per head, ±€475 per hectare, and ±€18 987 per 40 ha farm (Table 3). In the modelled scenarios with a higher proportion of concentrates in the diet, i.e. all HF genotype scenarios, and across all genotypes in the HC feed treatment, the purchase price of finishing concentrates had a greater impact on net margin (Table 3).
The impact of fluctuations in calf purchase, concentrate and beef price on net margin per head, per ha and per 40 ha farm

Price assumptions: Calf price (€200 for HA (High Angus) and LA (Low Angus), €60 for HF); Beef price €4.85/kg (Steer on EUROP system with a conformation and fat score of 8); Breed bonus €0.20/kg for Angus steers; Straw (4 × 4 bale) €18 bale; Urea fertilizer €550/t; 0-10-20 €400/t; Lime €35/t; Calves concentrate price €420/t; Yearling concentrate price €350/t; Finishing concentrate price €350/t; Milk replacer price €2100/t.
1Genotype: HA (High Angus) = Top 40 % sire DBI beef sub-index, LA (Low Angus) = Bottom 60 % sire DBI beef sub-index, HF = Holstein-Friesian.
Greenhouse gas emissions, food competition and land use
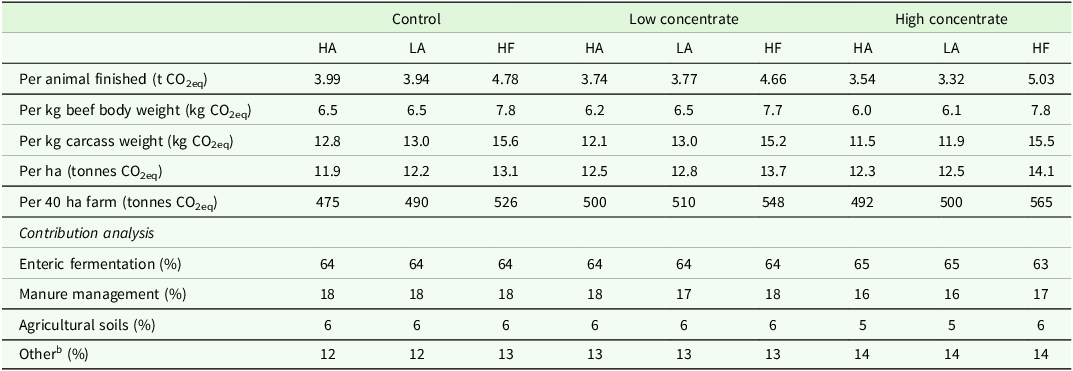
Across all feed treatments both the HA and LA genotypes had similar GHG emissions per animal finished, per kg of body weight and per kg of carcass weight with both being lower than the HF genotypes (Table 4). On average, the LA, HA and HF genotypes produced 3.68, 3.76 and 4.83 t CO2eq per animal finished, respectively. When expressed per 40 ha farm, HA scenarios produced the lowest total emissions, with HF being the highest, and LA intermediary. The higher SR of LA systems increased overall emissions per ha and per farm compared to HA. In addition, the higher carcass output per animal for HA steers reduced emissions per kg of carcass weight sold compared to LA.
Effect of genotype a and feed treatment on greenhouse gas emissions and contribution analysis of different sources

a Genotype: HA (High Angus) = Top 40 % sire DBI beef sub-index, LA (Low Angus) = Bottom 60 % sire DBI beef sub-index, HF = Holstein-Friesian.
b Other: Diesel use + nitrate leaching + ammonia volatilization + production of fertilizer, feed, electricity and diesel.
The CTL feed treatment had the highest GHG emissions per animal finished, per kg of body weight and per kg of carcass weight, with HC feed treatment being the lowest and LC intermediary. However, on a per hectare and per farm basis, the CTL feed treatment had the lowest GHG emissions produced due to lower animal numbers, with HC being intermediary and LC being the highest (Table 4).
For the Angus genotypes, GHG emissions per animal finished, per kg of body weight and per kg of carcass weight were lowest within the HC feed treatment, and highest within the CTL treatment. In contrast to the Angus steers, HF steers within HC had the highest overall GHG emissions per animal finished, with CTL having the lowest and LC intermediate, and can be attributed to the much greater consumption of concentrate per animal for HF steers in the HC feed treatment (Table 4). Nonetheless, GHG emissions per kg of body weight and per kg of carcass weight were similar across feed treatments for HF steers.
No difference was seen across genotypes or feed treatments for the contribution of each potential source of GHG emissions. Contribution analysis from this study showed that 63–65 % of each scenario’s emissions came from enteric fermentation (Table 4); manure management contributed an additional 16–18 % of GHG emissions, and agricultural soils contributed 5–6 % across scenarios. The remaining 12–14 % of GHG emissions was associated with ‘other’ factors, such as diesel use on-farm, nitrate leaching and ammonia volatilization, and the production of fertilizer, feed, electricity and diesel, resulting in the HC feed treatment having higher ‘other’ emissions due to the higher level of lifetime concentrate.
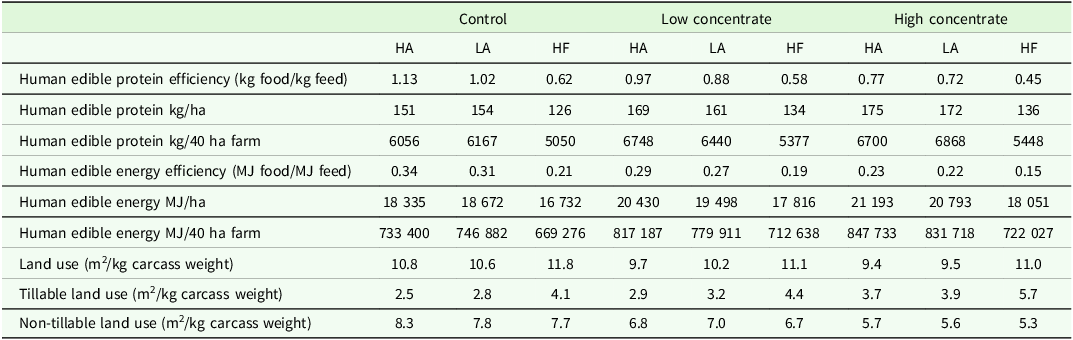
The quantity of human edible protein and energy produced within each scenario investigated is shown in Table 5. Both Angus genotypes produced similar quantities of human edible protein and energy across feed treatments. High Angus had a superior human edible protein efficiency and human edible energy efficiency ratio than the LA genotype. Across all feed treatments, the HF steers produced less protein and energy compared to both Angus genotypes and had lower human edible protein efficiency and human edible energy efficiency compared to both angus genotypes. Across all genotypes, the CTL feed treatment had a greater human edible protein efficiency and human edible energy efficiency ratio than either of the supplementation scenarios albeit the CTL feed treatment produced a lower quantity of human edible protein and energy than the other feed treatments. All scenarios investigated in the present study were net consumers of human edible energy, with all scenarios having an efficiency ratio <1.0, with the rank order of the scenarios consistent with the total human edible protein produced (Table 5). Only two of the scenarios, HA and LA genotypes in the CTL feed treatment, were net producers of human edible protein, with a human edible protein efficiency ratio of 1.13 and 1.02, respectively (Table 5). In contrast, all other scenarios were net consumers of human edible protein, with both Angus genotypes displaying more favourable efficiency ratios than HF steers within feed treatments.
The effect of genotype a and feed treatment on food-feed competition and land use

a Genotype: HA (High Angus) = Top 40 % sire DBI beef sub-index, LA (Low Angus) = Bottom 60 % sire DBI beef sub-index, HF = Holstein-Friesian.
Within genotypes, the HA and LA genotypes had similar land use per kg carcass weight produced, with the HF genotype requiring more land to produce the same amount of product. Across feed treatment, the HC feed treatment had the lowest land use per kg of carcass weight produced, with the CTL feed treatment having the greatest land use for across all genotypes per kg carcass weight.
Discussion
The positive impact of high beef merit sires on the body weight and carcass performance of progeny has been shown previously (O’Driscoll et al., Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025) and is an important component of profitable dairy-beef systems (Berry, Reference Berry2021). Previous studies in dairy (O’Sullivan et al., Reference O’Sullivan, Shalloo, Pierce and Buckley2020) and sheep (Fetherstone et al., Reference Fetherstone, McGovern, McHugh, Boland and Bohan2023) investigated the benefits of using total genetic merit indexes to increase farm profit; however, to-date, the effect of sire selection using a dairy-beef breeding objective, on the economic and environmental performance of dairy-beef systems, has not been quantified. Dairy-beef modelling studies have often relied on experimental data using genotypes that are no longer widely used (Ashfield et al., Reference Ashfield, Crosson and Wallace2013; Kearney et al., Reference Kearney, O’Riordan, McGee, Breen and Crosson2022), or experimental data where information on the beef merit of dairy-beef animals were unavailable (Kearney et al., Reference Kearney, O’Riordan, McGee, Breen, Dunne, French and Crosson2024). In the present study, results obtained from experimental data as outlined by O’Driscoll et al. (Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025) were used to populate a dairy-beef farm systems simulation model (Ashfield et al., Reference Ashfield, Crosson and Wallace2013; Kearney et al., Reference Kearney, O’Riordan, McGee, Breen and Crosson2022) to address this specific issue.
System physical and economic performance
This study was designed to quantify the differences in whole farm production, profit and environmental performance (greenhouse gas emissions and food-feed) of the progeny of sires of divergent beef genetic merit and Holstein-Friesian dams, managed under contrasting pasture-based concentrate supplementation strategies, which aimed to reduce the age at slaughter of the progeny. The use of high beef genetic merit (HA) sires improved progeny carcass weight, and both Angus genotypes had lower slaughter ages than HF. Consequently, HA steers had the greatest net margin per hectare, and produced less GHG emissions per kg beef product, than both lower beef genetic merit (LA) and HF steers. Strategic concentrate supplementation during the second grazing season (HC) reduced slaughter age of Angus sired steers but did not reduce the age at slaughter of HF steers. This reduced the emissions produced per animal finished and per kg of carcass weight produced of the Angus scenarios in HC. Concentrate supplementation over the first grazing season (LC) had no effect on slaughter age, however allowed a greater SR compared to CTL, increasing carcass output per hectare reducing GHG emissions per head and kg beef output. However, the CTL scenario was the most profitable scenario, due to lower concentrate usage and a greater proportion of forage in the diet. Additionally, the CTL scenario had the lowest overall emission produced per farm and per hectare.
Historically, the main challenge of beef production has been poor levels of profitability (Taylor et al., Reference Taylor, McGee, Kelly and Crosson2020a). In the case of dairy-beef systems, a 2023 survey in Ireland indicated these systems are returning a net margin of €162 per ha, excluding farm subsidies (Dillon, 2024). Additionally, large variations in profitability were found with farmers in the top 33 % achieving a net margin of €458/ha, compared to negative net margin −€134/ha for the bottom 33 % of farmers surveyed (Dillon, Reference Dillon, Donnellan, Moran and Lennon2024). Multiple causal factors are likely to underpin such variation at farm level, with differences in grassland management (Doyle et al.,Reference Doyle, McGee, Moloney, Kelly and O’Riordan2022), stocking rate (Kearney et al., Reference Kearney, O’Riordan, McGee, Breen, Dunne, French and Crosson2024), and animal genetics (Campion et al., Reference Campion, Keane, Kenny and Berry2009) all shown to effect animal performance. Therefore, information on the implications for farm profitability of genotype selection and feeding strategies is crucial. Systems and genotypes which make use of the most cost-effective feed resource in temperate climates, grazed pasture, are at an advantage in terms of minimizing the cost of production (Finneran et al., Reference Finneran, Crosson, O’kiely, Shalloo, Forristal and Wallace2012), with feed shown to account of 74 % of variable costs in pasture-based dairy-beef systems (Murphy et al., Reference Murphy, Crosson, Kelly and Prendiville2017). The use of pasture to reduce costs and increase beef farm profitability has been shown by Murphy et al. (Reference Murphy, Crosson, Kelly and Prendiville2017) in Irish dairy-beef steer and bull production systems.
Across all feed treatment scenarios, both HA and LA Angus genotypes had a lower cost per kg beef produced than HF steers. This is a result of their ability to reach the prerequisite slaughter BCS of 3.75 sooner than their HF counterparts, which reduced slaughter age, requiring fewer feed days on an expensive indoor finishing diet, as concentrate has consistently been found to be a more expensive feedstuff in temperate climates than grazed and conserved forage (Finneran et al., Reference Finneran, Crosson, O’kiely, Shalloo, Forristal and Wallace2012). Early maturing (Angus or Hereford) × dairy progeny are typically fatter than dairy × dairy progeny, at a given age (Berry and Ring, Reference Berry and Ring2020b), therefore in a study such as this, where slaughter age was determined by BCS, this reduction in slaughter age for Angus sired steers compared to HF, regardless of system is to be expected.
Carcass output per hectare has been identified as a key indicator of profit in pasture-based dairy-beef systems (Rutherford et al., Reference Rutherford, Lively and Arnott2021). Although many farmers aim to increase carcass output as a metric to increase profitability, the present study showed that compromising production costs to achieve high levels of carcass output per ha (i.e. by increasing the quantity of concentrates fed) can lead to reduced profitability, as shown by the results of the CTL scenarios, which had the highest proportion of forage in the diet and were the most profitable. These results corroborate the findings of Murphy et al. (Reference Murphy, Crosson, Kelly and Prendiville2017), which found HF steer and bull production systems, which had high levels of carcass output per hectare, and maximized the proportion of forage in the diet, to be more profitable than systems with high levels of carcass output, delivered through ad-libitum concentrate feeding indoors or at pasture. Within the CTL scenarios, the LA genotype had a higher carcass output per hectare than the HA genotype, but resulted in an increased concentrate input per head, thus reducing net margin for LA. Similarly, relative to the CTL feeding treatments, carcass output per hectare increased in the LC and HC feed treatment scenarios for both the LA and HF genotypes, primarily through increased livestock numbers, but higher concentrate input reduced net margin. Although the number of calves initially purchased within the LC and HC feed treatments was lower for HA, the higher carcass output per head compared to LA diluted costs associated with the increased carcass output per hectare achieved through supplementation, leading to greater net margin for HA than LA.
A decline in the beef genetic merit of animals of dairy origin in Ireland was previously reported by Mulhall et al. (Reference Mulhall, Sleator, Evans and Twomey2023), who found dairy and dairy-beef animals to have an annual yearly decline in estimated breeding values (EBVs) for carcass weight of 0.47 and 0.14 kg, respectively, from 2008–2018. In the current study, irrespective of feed treatment, the HF genotypes took longer to reach the prerequisite fat score for slaughter which in turn increased their concentrate and total feed requirements, limited the farm carrying capacity, reduced carcass output per hectare and resulted in a higher cost of production for HF steers compared to Angus. This is similar to previous studies on 24-month-old HF dairy-beef systems and 20-month-old early maturing steer systems (Ashfield et al., Reference Ashfield, Wallace, Prendiville and Crosson2014b; Kearney et al., Reference Kearney, O’Riordan, McGee, Breen and Crosson2022) which found steers slaughtered at 20 months consumed less feed over their lifetime than those finished at 24 months of age. The current study highlights the potential of replacing low beef genetic merit HF steers, with the progeny of high beef genetic merit bulls selected using a tool such as the DBI, in increasing profit at beef farm level. Such changes can be made without the detrimental effects of dystocia on the dam due to the DBI taking cognizance of both calving and beef production related traits (Berry and Ring, Reference Berry and Ring2020a).
The price received for the primary output of any system significantly impacts profit levels, and has been shown in sheep (Farrell et al., Reference Farrell, Kenyon, Tozer, Ramilan and Cranston2020; Fetherstone et al., Reference Fetherstone, McGovern, McHugh, Boland and Bohan2023), dairy (McClearn et al., Reference McClearn, Shalloo, Gilliland, Coughlan and McCarthy2020; O’Sullivan et al., Reference O’Sullivan, Shalloo, Pierce and Buckley2020) and beef cattle (Clariget et al., Reference Clariget, Kelly, Banchero, Keogh, Kenny and Crosson2024). The effect of the price received for the primary output (kg carcass weight) was also seen in the present study with the sensitivity analysis of ±€0.50c in base beef price, reflecting the changes typically seen in price received over a production cycle, resulting in ±€152 net margin per head and ±€475 per hectare. Calf purchase price had the next largest effect on farm profit and had a greater effect on the HF genotypes due to the lower number of calves purchased resulting in reduced total output. The greater sensitivity associated with the HF genotypes to differences in finishing concentrate purchase price is to be expected given the longer finishing duration and increased total finishing concentrate per head observed in these scenarios relative to Angus genotypes. The relative stability of the Angus genotypes in the present study to changes in input prices infers their increased resilience to changes in input prices, which is not unexpected given that maximizing the use of forage in the diet, whilst minimizing the use purchased feed has been shown to increase profitability across a range of input price scenarios (Hanrahan et al., Reference Hanrahan, McHugh, Hennessy, Moran, Kearney, Wallace and Shalloo2018).
Greenhouse gas emissions
Dairy-beef production systems have been shown to produce 29 % lower greenhouse gas emissions compared to traditional cow-calf production systems (van Selm et al., Reference van Selm, de Boer, Ledgard and van Middelaar2021), this is due to the fact emissions from suckler-beef production is dominated by the emissions produced by the beef cow, whereas the emissions output of the dairy cow is primarily attributed to milk production (De Vries et al., Reference De Vries, Van Middelaar and De Boer2015). However, multiple factors influence total system emissions. Genetic selection tools such as the DBI aim to produce more efficient animals through increased growth rates and improved carcass performance (Berry et al., Reference Berry, Amer, Evans, Byrne, Cromie and Hely2019; Berry and Ring, Reference Berry and Ring2020b).
One of the most effective GHG mitigation strategies identified in dairy-beef systems (Kearney et al., Reference Kearney, O’Riordan, Byrne, Breen and Crosson2023), and Irish agriculture as a whole (Lanigan et al., Reference Lanigan, Black, Donnellan, Crosson, Beausang, Hanrahan, Buckley, Lahart, Herron and Redmond2023) is reducing age at slaughter. This is due to animals consuming feed and emitting biogenic methane as a by-product of ruminal fermentation for fewer days leading to a reduction in total GHG emissions. Previous studies modelling Irish production systems found that early maturing dairy beef steers slaughtered at either 20 or 24 months produce 12.1 and 15.0 kg CO2eq/kg carcass weight, respectively (Kearney et al., Reference Kearney, O’Riordan, McGee, Breen and Crosson2022). In the present study, the CTL and LC feed treatments were most similar to the 24-month slaughtering system, requiring a winter housing period prior to slaughter. Angus steers from either genotype in these FT had a lower slaughter age thereby reducing GHG emissions per kg carcass weight in comparison to the HF genotype. Conversely, the HC feed treatment was most comparable to the 20 month slaughter system, with higher individual animal carcass weight in the HA genotype within the HC feed treatment reducing GHG emissions per kg of beef product compared to Kearney et al. (Reference Kearney, O’Riordan, McGee, Breen and Crosson2022). The higher growth potential of the HA genotype (O’Driscoll et al., Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025) enabled them to achieve a greater carcass weight at similar slaughter ages compared to the LA genotype. Higher lifetime ADG has been shown to be a key strategy to reduce emissions in pasture-based production settings but it is critical that this comes from a predominantly forage-based diet (Foley et al., Reference Foley, Crosson, Lovett, Boland, O’Mara and Kenny2011).
Food-feed competition and land use
Agricultural land is a limited resource, and beef production systems are often criticized for being inefficient at turning non-human edible feed sources into human edible feed (Gerber et al., Reference Gerber, Mottet, Opio, Falcucci and Teillard2015). Nonetheless, the demand for beef is rising globally (Henchion et al., Reference Henchion, Moloney, Hyland, Zimmermann and McCarthy2021), therefore beef production systems which efficiently utilize agricultural land typically unsuited to growing direct human edible feed are an important avenue of converting non-human edible food into useable product (Mosnier et al., Reference Mosnier, Jarousse, Madrange, Balouzat, Guillier, Pirlo, Mertens, ORiordan, Pahmeyer and Hennart2021). Land use per kg carcass weight produced in this study was lower than that seen previously in both early maturing and HF steer systems in Kearney et al. (Reference Kearney, O’Riordan, McGee, Breen and Crosson2022), and can be attributed to the greater carcass output per hectare due to a higher stocking rate in the current study. This is similar to results seen in Kearney et al. (Reference Kearney, O’Riordan, McGee, Breen, Dunne, French and Crosson2024), where a high carcass output per hectare due to high stocking rate reduced land use per kg carcass weight produced, due to higher animal numbers within the system producing more beef on a fixed area of land.
Ruminants have a unique ability to transform bulky, fibrous, non-human edible feeds such as grazed pasture to high quality human edible products (Mottet et al., Reference Mottet, de Haan, Falcucci, Tempio, Opio and Gerber2017). However, there is growing concern over the use of human-edible products in livestock production systems, as ruminants, monogastric livestock, and humans are all competing for a limited amount of cultivable land worldwide (Wilkinson and Lee, Reference Wilkinson and Lee2018). Beef production systems in temperate climates such as Ireland, the UK, and New Zealand, are based on the usage of human inedible feedstuffs such as grazed and conserved pasture as the primary feedstuff (80–90 % of an animals total lifetime feed requirement), grown on land typically not used for the production of human edible feed (Hennessy et al., Reference Hennessy, Shalloo, Van Zanten, Schop and De Boer2021). A positive human edible protein ratio (kg protein produced/kg protein fed), in combination with a high total human edible protein output per hectare is important to ensure that the agricultural land is used efficiently (Wilkinson and Lee, Reference Wilkinson and Lee2018). High and Low Angus genotypes in the CTL feed treatment were found to be net producers of human edible protein, this is not surprising given that both scenarios had the highest proportion of forage in the animals lifetime diet (87 %), and corroborates previous research which found beef systems that maximize the use of pasture in the diet have more favourable human edible protein efficiency (Mosnier et al., Reference Mosnier, Jarousse, Madrange, Balouzat, Guillier, Pirlo, Mertens, ORiordan, Pahmeyer and Hennart2021; Doyle et al., Reference Doyle, O’Riordan, McGee, Crosson, Kelly and Moloney2023).
Total human edible protein output was greater for both Angus genotypes across all FT (151–175 kg/ha) in this study than results seen in early maturing steers slaughtered within a 20 month (109 kg/ha) and 24 month (100 kg/ha) steer system in Kearney et al. (Reference Kearney, O’Riordan, McGee, Breen and Crosson2022) and within early maturing steer systems (117 kg/ha) differing in stocking rate seen in Kearney et al. (Reference Kearney, O’Riordan, McGee, Breen, Dunne, French and Crosson2024). The stocking rates within the aforementioned study were defined as low (2.6 Livestock units/ha) and high (3.2 Livestock units/ha), with only the ‘Low’ system within (Kearney et al., Reference Kearney, O’Riordan, McGee, Breen, Dunne, French and Crosson2024) comparable to the Angus scenarios in the present study. The age at slaughter for early maturing steers in Kearney et al. (Reference Kearney, O’Riordan, McGee, Breen, Dunne, French and Crosson2024) was 858 days, 228 days older than the average slaughter age of the Angus genotypes (average 630 days) within the present study. This limited overall carrying capacity as it necessitated steers to return to pasture for a third summer, reducing the number of animal units and carcass output per hectare; therefore, human edible protein per hectare was lower than those observed in the present study.
Conclusion
This study showed that progeny of high beef genetic merit Angus sires had superior carcass, economic and environmental performances than beef steers sired by lower genetic merit beef sires and HF sires. Results from this study show that farm systems that maximized carcass output per hectare and minimized the cost of production by utilizing a greater proportion of forage, were the most profitable systems. Concentrate supplementation does have the potential to reduce GHG emissions on a per head and per kg of product basis, however systems which utilized more forage produced less GHG emission per hectare and per farm. In addition, systems that utilized a higher proportion of forage were also net producers of human edible protein, in contrast to systems which utilized more concentrate, which were net consumers of human edible protein. Overall, this study shows that the improved phenotypic performance as described in O’Driscoll et al. (Reference O’Driscoll, Purfield, McHugh, Berry and Byrne2025) for high beef merit dairy-beef progeny, transpired into tangible economic and environmental benefits for the beef producer thereby providing them with greater confidence at calf purchase time when selecting progeny of sires of superior genetic merit.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0021859626100562.
Acknowledgements
The authors wish to acknowledge the technical expertise of Donall Fahy, Anthony Mulligan and Ronan Grealis in conducting this research. Particular thanks go to Brian Duffy for his care for the animals during this project.
Author contributions
Conceptualization, methodology, formal analysis and resources, J. O, D. C. P, N. M, P. C, and N. B.; investigation, J. O and N. B.; data curation, J. O, P. C, and N. B, writing – original draft preparation, J. O.; writing – review and editing, D. C. P, N. M, P. C and N. B.; supervision, N. B and D. C. P.; project administration, N. B.; funding acquisition, N. M.
Funding statement
This research was supported by the Department of Agriculture, Food and the Marine (Dublin, Ireland) Research Stimulus Fund 2019R553 (Dairy4Beef)
Competing interests
The author has no real or perceived conflicts of interest.
Ethical standards
Ethical approval was obtained from the Teagasc Animal Ethics Committee (TAEC2020-288) and the Health Protection Regulatory Authority (AE19132/P135) in adherence with the Cruelty to Animals’ Act 1876 and the European Communities Regulations 1994.



 Open access
Open access