


Introduction
Parasitic diseases remain a significant global health burden, particularly in low- and middle-income countries, where inadequate sanitation, poor healthcare infrastructure, and limited access to clean water facilitate their spread (Raval et al., Reference Raval, Jadav, Gohil and Vora2024). Helminthic infection is one of the most prevalent parasitic diseases, affecting both animals and human populations, with severe economic, health, and environmental consequences. In humans, soil-transmitted helminths (STHs), such as Ascaris lumbricoides, Trichuris trichiura, and hookworms, infect 1.5 billion people worldwide, particularly in developing regions, resulting in severe morbidity and impaired quality of life (Breman et al., Reference Breman, Egan and Keusch2001; Chen et al., Reference Chen, Gong, Chen, Li and Zhou2024; Hubbard et al., Reference Hubbard, Thompson, Else, Shears, Rollinson and Stothard2023). In parallel, gastrointestinal nematode (GIN) infections pose a significant threat to livestock health and productivity worldwide. These parasites reduce feed conversion efficiency, lower growth rates, and compromise animal welfare, thereby affecting global food security (Bricarello et al., Reference Bricarello, Longo, Da Rocha and Hotzel2023). In Europe alone, the combined annual cost of three major helminthic infections across 18 European countries has been estimated at €1.8 billion (Charlier et al., Reference Charlier, Rinaldi, Musella, Ploeger, Chartier, Vineer, Hinney, Von Samson-Himmelstjerna, Băcescu, Mickiewicz, Mateus, Martinez-Valladares, Quealy, Azaizeh, Sekovska, Akkari, Petkevicius, Hektoen, Höglund, Morgan, Bartley and Claerebout2020). The control of parasitic diseases has relied heavily on anthelmintic drugs, such as benzimidazoles, macrocyclic lactones, and tetrahydropyrimidines/imidazothiazoles. While these drugs have been effectively used to treat parasitic diseases, their widespread and often indiscriminate use has led to the emergence of anthelmintic resistance (Kaplan, Reference Kaplan2020; Kotze and Hunt, Reference Kotze and Hunt2023; Raza et al., Reference Raza, Qamar, Hayat, Ashraf and Williams2019). The growing demand for organic farming and chemical-free animal products has also intensified the search for alternative parasite control strategies. Hence, there is an urgent need to develop novel approaches for helminth control.
Bioactive plants containing compounds with anthelmintic activity demonstrate strong potential for controlling parasites (Thamsborg et al., Reference Thamsborg, Roepstorff, Nejsum and Mejer2010). Their use as nutraceuticals holds promise as sustainable anthelmintics, reducing drug resistance and residues in livestock while also offering safe and effective options for human parasite control. For example, seaweeds have shown promise due to their natural anthelmintic compounds. For instance, Digenea simplex, containing kainic acid, has long been used in traditional medicine to treat Ascaris infections in humans, while Notheia anomala and Nordic brown seaweeds such as Laminaria digitata and Saccharina latissima exhibit nematocidal activity against livestock parasites (Bonde et al., Reference Bonde, Bornancin, Lu, Simonsen, Martínez-Valladares, Peña-Espinoza, Mejer, Williams and Thamsborg2021, Reference Bonde, Mejer, Myhill, Zhu, Jensen, Büdeyri Gökgöz, Krych, Nielsen, Skovgaard, Thamsborg and Williams2023; Schwimmer and Schwimmer, Reference Schwimmer and Schwimmer1955). This activity of Nordic seaweed was associated with the presence of six polyunsaturated fatty acids (PUFAs) in the seaweed extracts.
PUFAs, particularly omega-3 (n-3) and omega-6 (n-6) fatty acids, are essential dietary lipids that play critical roles in human and animal health (Mititelu et al., Reference Mititelu, Lupuliasa, Neacșu, Olteanu, Busnatu, Mihai, Popovici, Măru, Boroghină, Mihai, Ioniță-Mîndrican and Scafa-Udriște2025). These fatty acids, obtained from sources such as fish oil, flaxseed, and walnuts (Rizzo and Baroni, Reference Rizzo and Baroni2023), are key components of cell membranes and precursors to bioactive lipid mediators, including eicosanoids, resolvins, and protectins, that regulate inflammation and immune responses (Kar et al., Reference Kar, Ghosh, Patra, Chini, Nath, Saha and Chandra Patra2023). Emerging evidence suggests that PUFAs may have both direct and indirect effects on parasites. By modulating host immunity and inflammation, PUFAs can enhance resistance to infection and reduce pathological damage (Rahman et al., Reference Rahman, Weng, Qadeer, Nawaz, Ullah and Chen2024). Additionally, they may disrupt parasite membranes and interfere with metabolic pathways, exhibiting direct anthelmintic effects (Leroux et al., Reference Leroux, Luquain-Costaz, Lawton, Azzouz-Maache and Delton2023). Unlike conventional anthelmintics, PUFAs are naturally occurring compounds with minimal environmental toxicity, making them an attractive alternative for sustainable parasite control (Manjusa and Pradeep, Reference Manjusa and Pradeep2022).
Helminthic infections are a classic One Health issue, as they often involve zoonotic transmission and are influenced by environmental factors such as climate change and land use (Thompson, Reference Thompson2023). Integrating PUFAs into parasite control strategies could simultaneously benefit human and animal health while mitigating environmental impacts. For example, PUFA-enriched diets for livestock may reduce parasite burdens, improve productivity, and decrease reliance on chemical anthelmintics, thereby reducing resistance and environmental contamination. This review critically examines the emerging evidence for PUFAs as multifunctional tools for controlling parasites in humans and animals. We evaluate their dual mechanisms, i.e., direct antiparasitic activity and immunomodulatory effects, while discussing challenges such as bioavailability and the limited number of clinical studies. We highlight the translational potential of PUFA-enriched diets, bioactive forages, and algal-derived compounds to reduce reliance on chemical anthelmintics, mitigate resistance, and promote ecological resilience.
PUFAs: biochemistry and sources
Classification of PUFAs
PUFAs are long-chain fatty acids with multiple cis double bonds (Kapoor et al., Reference Kapoor, Kapoor, Gautam, Singh and Bhardwaj2021). Fatty acids are constituted with hydrocarbon chains containing a carboxyl group at one end and a methyl group at the other end (also known as the ω end). One or more hydrogen-unsaturated double bonds are present in the chain of unsaturated fatty acids (Aldred et al., Reference Aldred, Buck, Vall, Aldred, Buck and Vall2009). PUFAs include multiple double bonds and are divided into two groups based on the position of double bonds along the carbon chain: omega-3 (n-3) and omega-6 (n-6). The identifier n-3 or n-6 denotes that the initial double bond on the hydrocarbon chain is at the third and sixth carbons from the n (or ω) end, respectively (Catalá, Reference Catalá2013). These multiple double bonds are the main reason for greater fluidity and lower melting points (Balić et al., Reference Balić, Vlašić, Žužul, Marinović and Bukvić Mokos2020). Omega-3 fatty acids include alpha-linolenic acid (ALA), eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA), whereas omega-6 fatty acids include linoleic acid (LA), calendic acid, and arachidonic acid (AA) (Kaur et al., Reference Kaur, Chugh and Gupta2014). These PUFAs are essential components of living systems and play an important role in the prevention and treatment of various diseases (Sampath and Ntambi, Reference Sampath and Ntambi2004).
Biosynthesis and metabolism of PUFAs
PUFAs are synthesized through a tightly regulated series of desaturation and elongation reactions, primarily occurring in the endoplasmic reticulum. Mammals lack the enzymes delta-12 (ω-6) and delta-15 (ω-3) desaturases, which are required to introduce double bonds at these positions in fatty acid chains. Therefore, LA (18:2n-6) and ALA (18:3n-3) are classified as essential fatty acids and must be obtained through dietary sources such as plant oils, nuts, seeds, and marine products (Abedi and Sahari, Reference Abedi and Sahari2014). Once ingested, these precursors undergo sequential elongation and desaturation to form long-chain PUFAs, which have distinct physiological roles.
LA is converted to AA (20:4n-6) via delta-6 desaturase (FADS2), elongase (ELOVL5), and delta-5 desaturase (FADS1). AA serves as a precursor to pro-inflammatory eicosanoids, including prostaglandins (PGs), thromboxanes, and leukotrienes (LTs), which mediate immune responses, vasoconstriction, and platelet aggregation (Calder, Reference Calder2013). ALA is metabolized to EPA (20:5n-3) and DHA (22:6n-3) through similar enzymatic steps. However, conversion efficiency in humans is limited (<10% for EPA and <1% for DHA), necessitating direct dietary intake of preformed EPA and DHA from marine and other sources (Brenna et al., Reference Brenna, Salem, Sinclair and Cunnane2009). EPA and DHA are precursors to specialized pro-resolving mediators (SPMs), such as resolvins, protectins, and maresins, which actively resolve inflammation and promote tissue repair (Serhan, Reference Serhan2014). The omega-3 and omega-6 pathways compete for the same enzymes (FADS2, ELOVL5), with the omega-6 pathway generally favoured due to higher substrate affinity (Burdge, Reference Burdge and Burdge2018). This competition highlights the importance of maintaining a balanced omega-6/omega-3 ratio in the diet.
Dietary sources of PUFAs
PUFAs are essential for human health, and their dietary sources span plant, marine, and animal origins. Each source contributes distinct fatty acid profiles, enabling diverse strategies to address global nutritional needs while aligning with sustainability goals. Plant-based oils and seeds are the primary sources of ALA and LA. Flaxseed oil (Linum usitatissimum) contains the highest concentration of ALA (53.37%), followed by canola oil (Brassica napus) (9.00%) and soya bean oil (Glycine max) (6.79%). LA is present in high amounts in soya bean oil (50.42%) and safflower oil (Carthamus tinctorius). Corn oil (Zea mays) is another significant source of omega-6 fatty acids, comprising 39.32% LA and only 1.16% ALA (Nosheen et al., Reference Nosheen, Bano, Yasmin, Keyani, Habib, Shah and Naz2016; Saini and Keum, Reference Saini and Keum2018).
Genetic modifications have enhanced fatty acid composition in various plants. CRISPR-Cas9 editing in rice (Oryza sativa) and soya beans (G. max) has targeted omega-6 desaturase, thereby increasing unsaturated oleic acid levels (Wang et al., Reference Wang, Wu, Chen, Dong, Chen, Wen, Hu, Fleming and Wang2021). Similarly, hairpin RNA-mediated post-transcriptional silencing of omega-6 desaturase in cotton (Gossypium hirsutum) has produced high levels of oleic and stearic acids, which may serve as replacements for hydrogenated fats in cooking (Liu et al., Reference Liu, Singh and Green2002). Saturated fats remain widespread in various cooking oils, with palm oil (Elaeis guineensis) containing 44% palmitic acid, cocoa butter (Theobroma cacao) 26%, olive oil (Olea europaea) 8–20%, and soya bean oil 10–12% (Saini and Keum, Reference Saini and Keum2018). However, breeding strategies can enhance PUFA content while reducing saturated fats (Coniglio and Shumskaya, Reference Coniglio and Shumskaya2023). Beyond plant oils, transgenic plants such as camelina (Camelina sativa) and microalgae cultivated on glucose have been engineered to produce omega-3 EPA and DHA, offering a sustainable alternative to fish-derived sources (Han et al., Reference Han, Usher, Sandgrind, Hassall, Sayanova, Michaelson, Haslam and Napier2020).
Preformed long-chain omega-3 PUFAs, such as EPA and DHA, are abundant in marine environments. Phytoplankton and zooplankton, wild-caught fatty fish such as salmon (Salmo salar), sardine (Sardina pilchardus), and herring (Clupea harengus) are rich sources of these vital lipids. In contrast, farmed fish primarily consume diets rich in omega-6 fatty acids derived from grain and vegetable oils. While both farmed and wild fish contain EPA and DHA, this dietary difference reduces the absolute levels of long-chain omega-3s in farmed fish and results in a lower omega-3:omega-6 ratio compared with wild-caught fish (Cladis et al., Reference Cladis, Kleiner, Freiser and Santerre2014). A variety of fish species naturally accumulate these fatty acids (Duan et al., Reference Duan, Song, Zhao and Yan2023; Rizzo and Baroni, Reference Rizzo and Baroni2023; Saini and Keum, Reference Saini and Keum2018). Fatty acids can be found in trace amounts in animal-derived foods, with organ meats such as liver and brain having the highest concentrations. Though not primary dietary sources, these tissues highlight the biological importance of PUFAs in neurological and metabolic functions across species (Saini and Keum, Reference Saini and Keum2018).
PUFAs and host–parasite interactions
PUFAs may play a multifaceted role in modulating host–parasite interactions, influencing the host’s immune response and the parasite’s survival. These effects are mediated through immunomodulatory, anti-inflammatory, and direct antiparasitic mechanisms, making PUFAs promising candidates for sustainable parasitic control (Fig. 1). Evidence from in vitro and in vivo studies shows that certain PUFAs can boost host defences, reduce harmful inflammation, and even act directly on parasites (Table 1). The following section explores the immunological, anti-inflammatory, and direct antiparasitic mechanisms of PUFAs across a range of parasites.
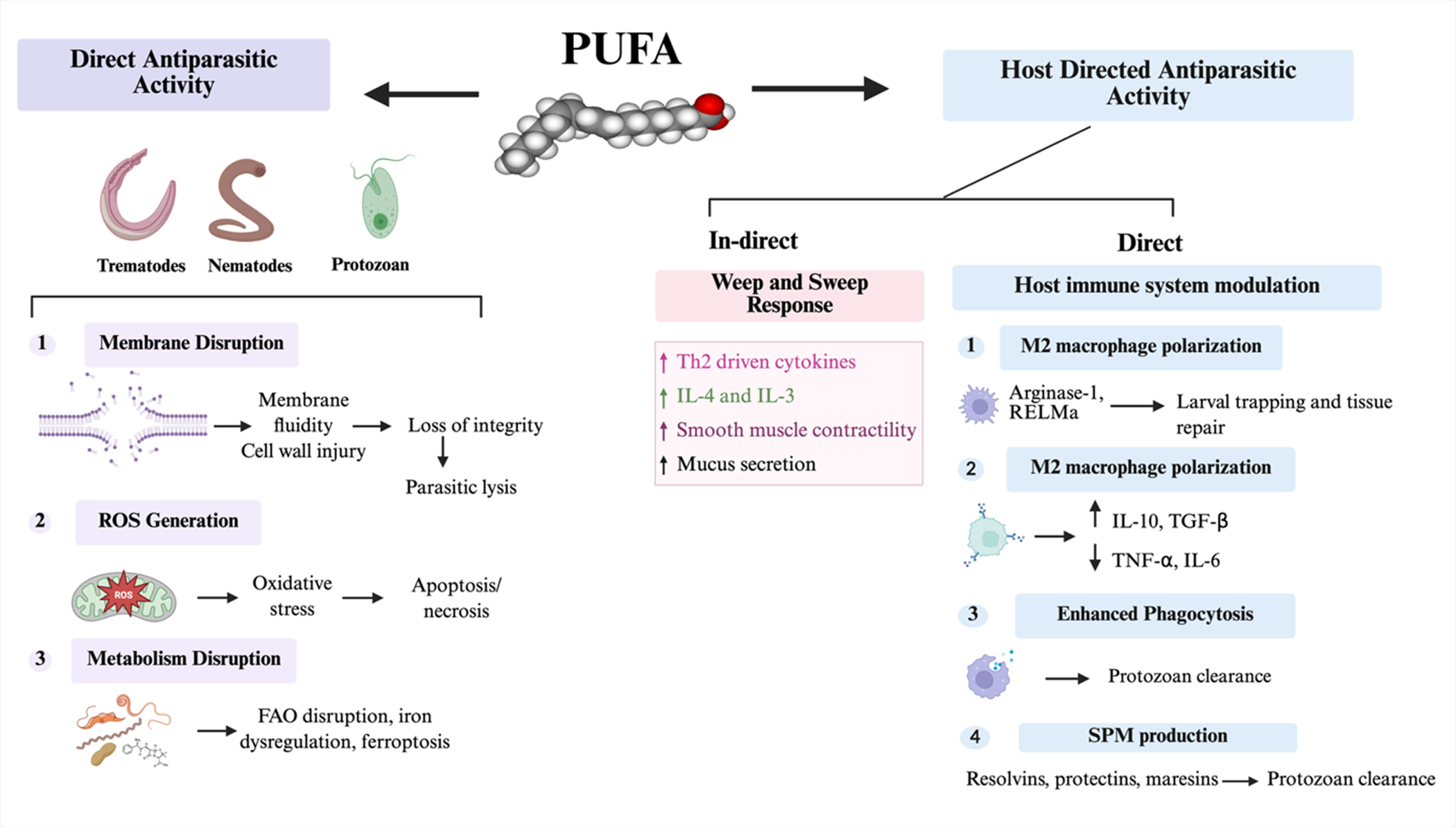
Dual antiparasitic actions of polyunsaturated fatty acids (PUFAs). PUFAs exhibit both direct and host-directed antiparasitic effects against helminths (trematodes, nematodes) and protozoa. Direct effects involve (1) membrane disruption, altering membrane fluidity and integrity leading to parasite lysis; (2) ROS generation, triggering oxidative stress and apoptosis/necrosis; and (3) metabolic disruption, including interference with fatty acid oxidation, iron regulation, and ferroptosis. Host-directed effects enhance mucosal and immune defences. Indirectly, PUFAs stimulate the ‘weep and sweep’ response through Th2 cytokines (IL-4, IL-13, IL-3), increased smooth muscle activity, and mucus secretion. Directly, they promote M2 macrophage polarization (arginase-1, resistin-like protein alpha) for larval trapping and tissue repair, increase anti-inflammatory cytokines (IL-10, TGF-β), reduce pro-inflammatory mediators (TNF-α, IL-6), enhance phagocytosis, and induce specialized pro-resolving mediators (resolvins, protectins, maresins) contributing to parasite clearance and tissue homeostasis. The figure was created with BioRender.com.

Figure 1 Long description
The diagram illustrates the dual antiparasitic actions of polyunsaturated fatty acids (PUFAs). On the left, direct antiparasitic activity is shown, affecting trematodes, nematodes and protozoa. This includes membrane disruption, leading to loss of integrity and parasite lysis, reactive oxygen species generation causing oxidative stress and apoptosis or necrosis and metabolism disruption involving fatty acid oxidation disruption, iron dysregulation and ferroptosis. On the right, host-directed antiparasitic activity is depicted, with indirect effects through the 'weep and sweep' response involving Th2 driven cytokines (IL-4 and IL-13), increased smooth muscle contractility and mucus secretion. Direct effects include host immune system modulation with M2 macrophage polarization (Arginase-1, RELM alpha) for larval trapping and tissue repair, increased anti-inflammatory cytokines (IL-10, TGF-beta), reduced pro-inflammatory mediators (TNF-alpha, IL-6), enhanced phagocytosis and specialized pro-resolving mediators production (resolvins, protectins, maresins) contributing to protozoan clearance.
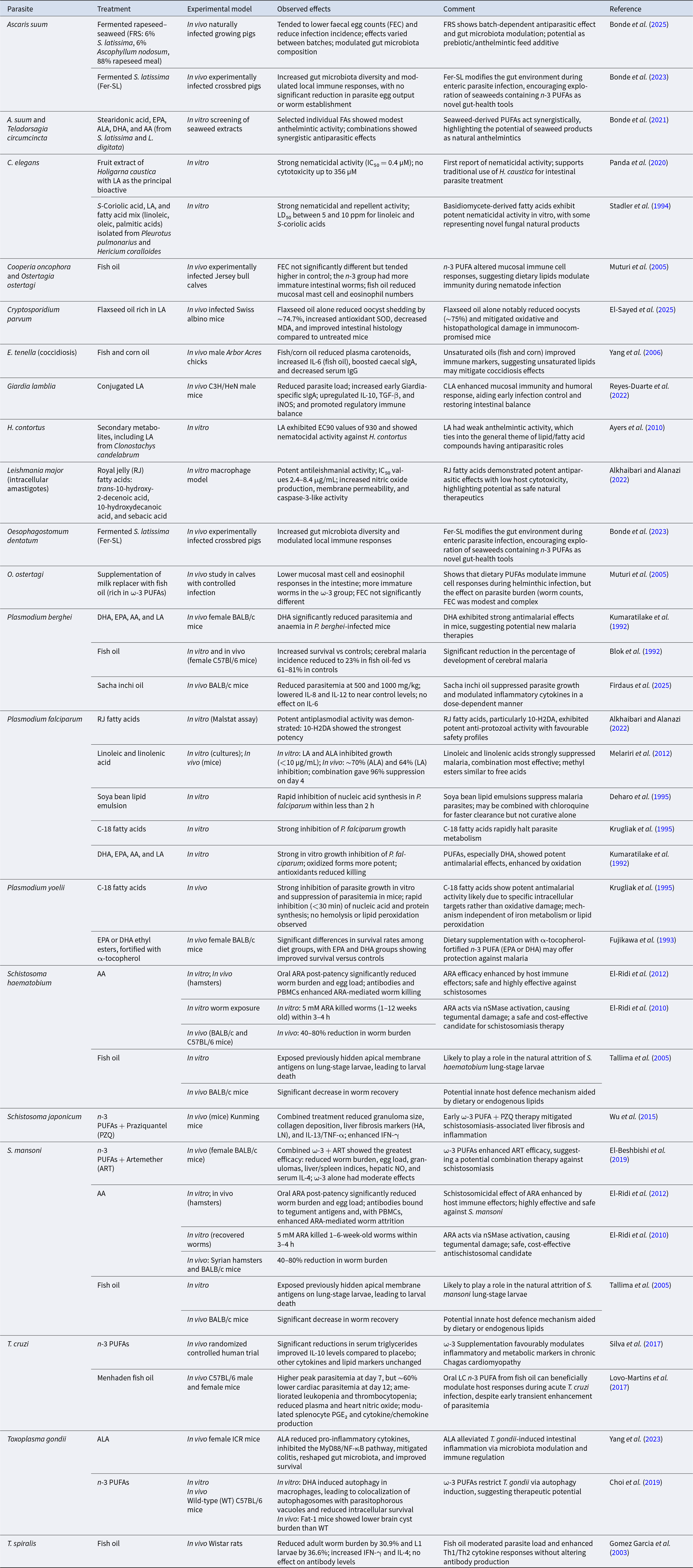
Antiparasitic and immunomodulatory effects of polyunsaturated fatty acids and related lipids: Summary of experimental, dietary, and in vitro studies across diverse parasite species

Table 1 Long description
The table presents a comprehensive overview of the antiparasitic and immunomodulatory effects of polyunsaturated fatty acids and related lipids across various experimental models and parasite species. Key findings include a reduction in fecal egg counts in pigs treated with fermented rapeseed–seaweed, the potent antileishmanial activity of royal jelly fatty acids, and a significant reduction in parasitemia in Plasmodium-infected mice treated with docosahexaenoic acid. Comparisons reveal that combinations of fatty acids often show synergistic effects, enhancing antiparasitic efficacy. Trends indicate that dietary lipids can modulate immune responses, as evidenced by increased gut microbiota diversity in pigs and modulation of inflammatory markers in Trypanosoma cruzi infections. Interpretation of results should consider the variability in effects due to different experimental conditions and the potential for batch-dependent outcomes.
PUFAs and anthelmintic immune response
One of the primary ways PUFAs may influence parasitic infections is by modulating the host immune system. Omega-3 PUFAs, such as ALA, EPA, and DHA, enhance the function of key immune cells, including macrophages, dendritic cells, and T cells (Gutiérrez et al., Reference Gutiérrez, Svahn and Johansson2019). As gut residents, GINs damage gut tissue and alter the composition of the gut microbiome, leading to impaired gut homeostasis and compromised host immunity. Anthelmintic immune responses are primarily dominated by type-2 immunity (Th2), which regulates the cellular and soluble effectors that disrupt the parasite niche by enhancing the physical mucosal barrier and ultimately promoting tissue repair (Sorobetea et al., Reference Sorobetea, Svensson-Frej and Grencis2018). Type-2 macrophages or alternatively activated macrophages (M2/AAMs) play a central role in trapping and killing helminth larvae, facilitating the expulsion of adult parasites, and contributing to tissue repair through the secretion of growth factors, chemokines, and molecules involved in collagen synthesis (Lechner et al., Reference Lechner, Bohnacker and Esser-Von Bieren2021). In addition, key AAM effector molecules, such as arginase-1 and resistin-like protein alpha, regulate Th2 cell activation and thus limit type-2 immunopathology during helminthic infection (Anthony et al., Reference Anthony, Urban, Alem, Hamed, Rozo, Boucher, Van Rooijen and Gause2006). The activation of AAMs with anthelmintic effector functions depends on the activation of multiple metabolic pathways, including increased oxidative phosphorylation, fatty acid oxidation (FAO), lipolysis, and glycolysis (Huang et al., Reference Huang, Smith, Everts, Colonna, Pearce, Schilling and Pearce2016). Therefore, external fatty acids could contribute to the fuelling of FAO in M2 macrophages. Omega-3 PUFAs have been reported to promote M2 polarization in vitro and in vivo (Haitz and Anandasabapathy, Reference Haitz and Anandasabapathy2015; Ohue-Kitano et al., Reference Ohue-Kitano, Yasuoka, Goto, Kitamura, Park, Kishino, Kimura, Kasubuchi, Takahashi, Li, Yeh, Jheng, Iwase, Tanaka, Masuda, Inoue, Yamakage, Kusakabe, Tani, Shimatsu, Takahashi, Ogawa, Satoh-Asahara and Kawada2018). Similarly, innate lymphoid cells (ILCs) play a crucial role in various immune processes, including the control of infections, inflammation, and tissue repair. During helminthic infection, ILC2s rely predominantly on fatty acid metabolism to sustain their immune functions. Notably, under conditions of nutrient limitation, such as vitamin A deficiency, ILC2s enhance fatty acid uptake and utilization to maintain IL-13 production, supporting barrier immunity and tissue repair (Wilhelm et al., Reference Wilhelm, Harrison, Schmitt, Pelletier, Spencer, Urban, Ploch, Ramalingam, Siegel and Belkaid2016). Hence, naturally occurring n-3 PUFAs may stimulate the anthelmintic immune response.
For example, EPA and DHA have been shown to increase the phagocytic activity of macrophages, which may enable them to more effectively engulf and destroy protozoan parasites (Gutiérrez et al., Reference Gutiérrez, Svahn and Johansson2019). This enhanced phagocytosis is particularly important in infections caused by intracellular parasites, such as Leishmania and Toxoplasma, where macrophage activity is critical for controlling parasite replication (Liu and Uzonna, Reference Liu and Uzonna2012). Unlike bacterial or protozoan pathogens, helminths are large parasites that cannot be engulfed and eliminated by phagocytosis; however, macrophages, owing to their remarkable plasticity, have developed diverse strategies to combat these parasites and mitigate parasite-induced tissue damage (Park et al., Reference Park, Silvin, Ginhoux and Merad2022). Overall, PUFAs represent a promising adjunct strategy to enhance anthelmintic activity by modulating host immunity and influencing parasite survival. However, more translational studies are needed to fully define their therapeutic potential.
Effects of PUFAs on the resolution of inflammation
Beyond immune modulation, PUFAs exert potent anti-inflammatory and pro-resolving effects that are particularly beneficial in parasitic diseases. Chronic inflammation is a hallmark of many parasitic infections, contributing to tissue damage and long-term morbidity (Blitz et al., Reference Blitz, Riddle and Porter2018). Omega-3 PUFAs serve as precursors to SPMs, including resolvins, protectins, and maresins, which actively resolve inflammation by reducing leukocyte infiltration, promoting macrophage clearance of cellular debris, and enhancing tissue regeneration (Serhan, Reference Serhan2014). These mediators are especially important in infections, particularly with tissue-migratory helminths, such as schistosomiasis, where chronic inflammation leads to liver fibrosis and granuloma formation. By contrast, omega-6 PUFAs, such as AA, are precursors to pro-inflammatory eicosanoids like PGs and LTs. However, the incorporation of omega-3 PUFAs into cell membranes competes with AA for enzymatic conversion, leading to the production of less inflammatory or anti-inflammatory eicosanoids (Simonetto and Infante, Reference Simonetto and Infante2019). This shift in eicosanoid balance helps to mitigate the inflammatory response and reduce tissue damage during parasitic infections.
PUFAs also regulate cytokine production, shifting the immune response towards a balanced state that avoids excessive inflammation while maintaining adequate parasite clearance (Calder, Reference Calder1998). In particular, omega-3 PUFAs promote the secretion of anti-inflammatory cytokines while suppressing pro-inflammatory cytokines. For example, in vitro exposure to ALA and its gut lactic acid metabolites induced M2 macrophage polarization in the presence of Th2 cytokines IL-4 and or IL-13 (Ohue-Kitano et al., Reference Ohue-Kitano, Yasuoka, Goto, Kitamura, Park, Kishino, Kimura, Kasubuchi, Takahashi, Li, Yeh, Jheng, Iwase, Tanaka, Masuda, Inoue, Yamakage, Kusakabe, Tani, Shimatsu, Takahashi, Ogawa, Satoh-Asahara and Kawada2018). Therefore, supplementation of ALA may drive early M2 polarization in the presence of helminthic infection through IL-4, thus developing an early anthelmintic response. Moreover, ALA’s ability to suppress pro-inflammatory cytokines (e.g., TNF-α, IL-1β) and increase anti-inflammatory cytokines (e.g., IL-10) makes it especially effective in restoring gut homeostasis (Song et al., Reference Song, Xia, Yang, Wang and Sun2018; Yan et al., Reference Yan, Jiang, Spinetti, Tardivel, Castillo, Bourquin, Guarda, Tian, Tschopp and Zhou2013). This cytokine modulation is critical in preventing immunopathology, which often exacerbates tissue damage in chronic parasitic infections such as schistosomiasis and filariasis.
Direct antiparasitic activity of PUFAs: in vitro and in vivo evidence
In addition to their effects on the host immune system, PUFAs have shown direct antiparasitic effects. Several in vitro studies have demonstrated the anthelmintic potential of PUFAs and related lipid compounds (Table 1). ALA has shown anthelmintic activity against the free-living nematode, Caenorhabditis elegans, as reported by Stadler et al. (Reference Stadler, Mayer, Anke and Sterner1994), who also noted similar effects from a fatty acid mixture containing linoleic, oleic, and palmitic acids. Additional research supports the role of medium-chain fatty acids and phenolic compounds in worm control. For instance, Kumarasingha et al. (Reference Kumarasingha, Karpe, Preston, Yeo, Lim, Tu, Luu, Simpson, Shaw and Gasser2016) reported the efficacy of a plant extract rich in medium-chain fatty acids and phenolic acids against C. elegans and Haemonchus contortus. Medium-chain fatty acids have also been reported to exert larvicidal activity against Toxocara canis and the monogenean, Heterobothrium okamotoi in vitro (Hirazawa et al., Reference Hirazawa, Oshima, Mitsuboshi and Hata2001; Kiuchi et al., Reference Kiuchi, Miyashita, Tsuda, Kondo and Yoshimura1987). The higher safety profile of ALA permits high-dose administration (up to 10 g/day) in humans and animals, highlighting its potential as a primary or complementary anthelmintic agent (Kim et al., Reference Kim, Nam, Kim, Hayes and Lee2014; Zehr and Walker, Reference Zehr and Walker2018).
Limited information is available on the potential of PUFAs in managing parasitic diseases in vivo. In helminth infections, such as those caused by Schistosoma spp., dietary supplementation with omega-3 PUFAs has been shown to reduce parasite burdens, liver fibrosis, and granuloma formation and mitigate associated tissue damage in schistosomiasis, highlighting their anti-inflammatory and pro-resolving properties (Wu et al., Reference Wu, Liang, Zhu, Gao, Chen, Zhang, Yin, Li, Wang and Xiao2015). In addition, the administration of AA and fish oil supplementation significantly reduced worm burdens, egg loads, and liver fibrosis in murine models of Schistosoma mansoni and S. haematobium infections, with combined therapies (e.g., fish oil plus praziquantel) enhancing survival and diminishing granuloma formation (El-Beshbishi et al., Reference El-Beshbishi, Saleh, Abd El-Mageed, El-Nemr, Abdalla, Shebl and Taman2019; El-Ridi et al., Reference El-Ridi, Tallima, Salah, Aboueldahab, Fahmy, Al-Halbosiy and Mahmoud2012; Wu et al., Reference Wu, Liang, Zhu, Gao, Chen, Zhang, Yin, Li, Wang and Xiao2015). Similarly, dietary supplementation with fish oil significantly reduced the parasite burden of Trichinella spiralis in rats, with a 30.9% reduction in adult worms and 36.6% reduction in L1 larvae, while enhancing both Th1 (IFN-γ) and Th2 (IL-4) immune responses during early infection (Gomez Garcia et al., Reference Gomez Garcia, Sanz Sampelayo, Fernandez Navarro, Carmona Lopez, Gil Extremera and Rodriguez Osorio2003).
In protozoan infections, omega-3 PUFAs, such as EPA and DHA, effectively reduced parasitaemia and improved survival rates in mice infected with Plasmodium species (Fujikawa et al., Reference Fujikawa, Kamitani, Tunru, Yamazaki and Hamazaki1993; Levander et al., Reference Levander, Ager, Morris and May1989b, Reference Levander, Ager, Morris and May1989a). In Trypanosoma cruzi infections, fish oil supplementation modulated immune responses, reduced cardiac parasitism and inflammatory markers, such as nitric oxide, while increasing protective cytokines such as IL-10 (Lovo-Martins et al., Reference Lovo-Martins, Malvezi, Da Silva, Zanluqui, Tatakihara, Câmara, De Oliveira, Peron, Martins-Pinge, Fritsche and Pinge-Filho2017; Silva et al., Reference Silva, Mediano, Silva, Brito, Cardoso, Almeida, Sangenis, Pinheiro, Hasslocher-Moreno, Brasil and Sousa2017). Poultry studies further highlight the benefits of PUFAs, with fish oil supplementation reducing caecal lesions and mortality in chickens infected with Eimeria tenella (Allen et al., Reference Allen, Danforth and Levander1996; Yang et al., Reference Yang, Guo, Wang and Nie2006). On the other hand, dietary supplementation with fish oil showed no benefit in leishmaniasis in susceptible BALB/c mice (Calabrese and Da Costa, Reference Calabrese and Da Costa1992). Recently, Rahman et al. (Reference Rahman, Weng, Qadeer, Nawaz, Ullah and Chen2024) reported that omega-3 PUFAs reduce the proliferation of T. gondii in vitro and in vivo, likely through a combination of immune modulation and direct antiparasitic mechanisms. For example, ALA ameliorated T. gondii-induced colitis in mice by modulating gut microbiota composition and suppressing pro-inflammatory cytokine production through inhibition of the MyD88/NF-κB signalling pathway. Notably, ALA administration or faecal microbiota transplantation from ALA-treated mice modified microbial communities and improved host survival, highlighting its potential as a therapeutic agent for parasite-induced intestinal inflammation (Yang et al., Reference Yang, Liu, Zhao, Li and Jiang2023).
Furthermore, PUFAs can integrate into parasite lipid bilayers, altering membrane fluidity, structural integrity, and permeability. For instance, Schistosoma, a trematode exposed to AA concentrations greater than 20 μM in vitro, experienced surface membrane breakdown and ultimately parasite mortality (Tallima et al., Reference Tallima, Salah and El-Ridi2005). These findings suggest that the PUFAs exert biophysical stress on parasites by destabilizing membranes and disrupting physiological functions, eventually resulting in parasite viability. Such dual mechanisms of action make PUFAs a promising adjunct therapy for parasitic infections, particularly in resource-limited settings where access to conventional antiparasitic drugs is limited.
Mechanistic insights into direct antiparasitic activity of PUFAs
The antiparasitic effects of PUFAs may be mediated by mechanisms including membrane disruption, induction of oxidative stress, and interference with parasite metabolism. Fatty acids are essential components of cell membranes and contribute to membrane fluidity; exposure to PUFAs may interact with and compromise the integrity of the cuticle, causing lethal damage to parasites. For example, in vitro exposure of Schistosoma to AA resulted in damage to the surface membranes, leading to parasite death (Tallima et al., Reference Tallima, Salah and El-Ridi2005). In addition, the peroxidation of PUFAs generates reactive oxygen species (ROS), which can induce oxidative stress in parasites. The elevated ROS levels can disrupt parasite membranes, enzymes, and mitochondrial function, ultimately leading to cell death through multiple redox-mediated pathways (illustrated in Fig. 2) (Goel et al., Reference Goel, Singla and Choudhury2020). Due to their limited antioxidant defences, helminths are vulnerable to oxidative damage upon exposure to ROS produced by host cells (Cancela et al., 2019; Roy et al., 2020; Su et al., 2019) or to chemicals, such as PUFAs.
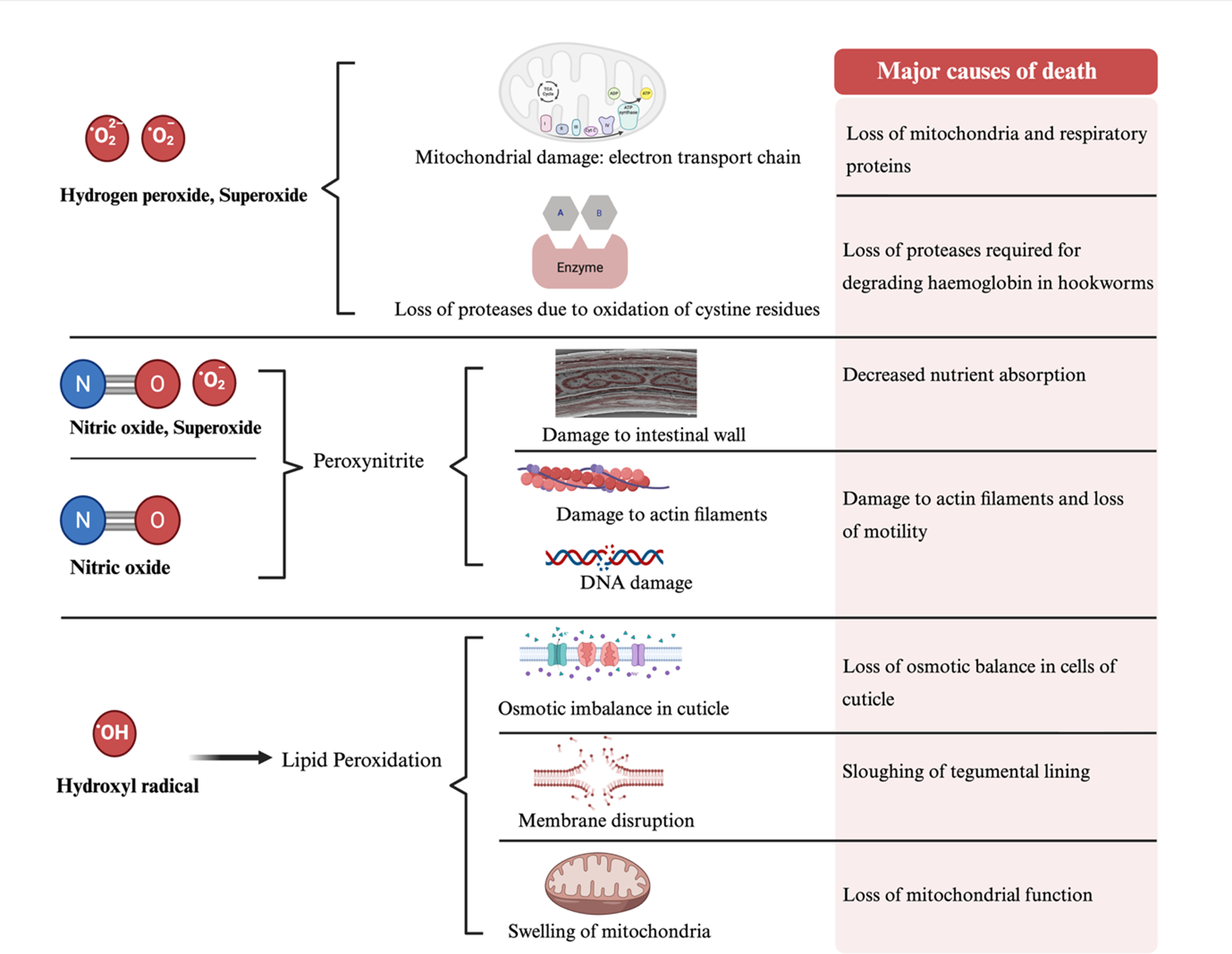
Potential mechanisms of reactive oxygen species (ROS)-mediated damage in parasitic worms. ROS, including hydrogen peroxide, superoxide, nitric oxide, peroxynitrite, and hydroxyl radicals, cause oxidative damage to multiple cellular targets within parasites. These effects include mitochondrial dysfunction and the oxidation of proteolytic enzymes essential for nutrient acquisition, as well as damage to structural components such as the intestinal wall, actin filaments, and DNA. Lipid peroxidation and hydroxyl radical formation disrupt membrane integrity, induce osmotic imbalance in the cuticle, and lead to mitochondrial swelling. Collectively, these ROS-driven processes compromise essential cellular functions, ultimately leading to parasite death. The figure was created with BioRender.com.

Figure 2 Long description
The diagram illustrates the effects of reactive oxygen species on parasites. Hydrogen peroxide and superoxide cause mitochondrial damage to the electron transport chain and loss of proteases due to cysteine residue oxidation. Nitric oxide and superoxide form peroxynitrite, leading to intestinal wall damage, actin filament damage and DNA damage. Hydroxyl radicals cause lipid peroxidation, resulting in osmotic imbalance in the cuticle, membrane disruption and mitochondrial swelling. These processes lead to major causes of death, including loss of mitochondria and respiratory proteins, decreased nutrient absorption, loss of motility and loss of mitochondrial function.
PUFAs can also interfere with essential metabolic pathways in nematodes, including iron homeostasis (Perez and Watts, Reference Perez and Watts2021). For example, dihomogamma-linolenic acid triggers germ-cell ferroptosis and sterility in C. elegans (Deline et al., Reference Deline, Keller, Rothe, Schunck, Menzel and Watts2015; Webster et al., Reference Webster, Deline and Watts2013). In addition, it was speculated that LA affects ionotropic neurotransmitter receptors in C. elegans, a target of some common anthelmintics. However, no studies have yet attempted to elucidate the biological processes disrupted in worms following PUFAs exposure.
Notably, studies on T. gondii have shown that DHA triggers autophagy in host cells via AMPK-mediated pathways, thereby significantly reducing intracellular parasite proliferation and brain cyst burdens (Choi et al., Reference Choi, Lee, Lee, Park, Lee, Shin, Cha, Lee, Lim and Yuk2019). On the other hand, the mechanisms of PUFA-mediated activity in parasitic nematodes remain uncharacterized. Key knowledge gaps persist regarding (1) the internalization and tissue-specific binding of PUFAs within helminths, (2) whether cuticular binding alone disrupts the nematode-environment interface sufficiently to induce metabolic damage, and (3) the necessity of intracellular PUFA penetration for direct inhibition of essential metabolic enzymes. Therefore, this necessitates the need for comprehensive multi-omics approaches (e.g., transcriptomic, proteomic, and metabolomic analyses) to systematically characterize PUFA-mediated anthelmintic effects in parasitic nematodes.
One Health approach: integrating PUFAs into sustainable anthelmintic strategies
The One Health approach views health as a shared outcome shaped by the interconnectedness of humans, animals, and the environment. This collaborative, cross-disciplinary perspective is crucial when tackling parasitic diseases, which often exploit ecological overlaps and shifts between host species. STHs, such as Ascaris, Trichuris, and hookworms, exemplify this interconnectedness, thriving in environments where poor sanitation and faecally contaminated soils facilitate transmission within host species and zoonotic transmission (Jourdan et al., Reference Jourdan, Lamberton, Fenwick and Addiss2018). Similarly, other zoonotic parasites, such as T. gondii and Echinococcus spp. depend on complex multi-host life cycles with reservoirs in wild and domestic animals, necessitating integrated surveillance and intervention strategies that exceed traditional disciplinary boundaries (Thompson, Reference Thompson2013). The environmental aspect of the One Health framework is vital, as human activities such as intensive livestock farming and climate-induced changes in habitat and land use significantly alter parasite transmission patterns (Short et al., Reference Short, Caminade and Thomas2017). The extensive use of synthetic anthelmintics in agriculture, for example, has led to the persistence of drug residues in the environment, thereby accelerating the development of anthelmintic resistance and posing risks to non-target aquatic and terrestrial species (Kaplan, Reference Kaplan2020). These practices highlight the urgent need for sustainable, eco-friendly alternatives to parasite control.
One such alternative approach is dietary interventions. Nutritional supplementation, for example, with protein, has been shown to enhance the host resistance and resilience against GIN infections (Atiba et al., Reference Atiba, Zewei and Qingzhen2020; Sweeny et al., Reference Sweeny, Clerc, Pontifes, Venkatesan, Babayan and Pedersen2021). Similarly, using a bolus of copper oxide wire particles can reduce H. contortus burdens and improve the efficacy of conventional anthelmintics (Quadros and Burke, Reference Quadros and Burke2024). Similarly, PUFAs represent a promising nutritional intervention as they have direct anthelmintic activity against a range of parasite species and immunomodulatory properties that can enhance host defence mechanisms (Table 1), offering a sustainable approach to parasite control in livestock and humans. In livestock, diets enriched with PUFAs may enhance innate resistance to GINs and improve feed efficiency, thereby decreasing the reliance on chemical anthelmintics. Similarly, in humans, parasitism has been linked to malnutrition, with individuals often experiencing both conditions simultaneously. Malnutrition can impair immune function and increase susceptibility to infection, while helminthic infections exacerbate nutritional deficiencies, creating a recurrent relationship that highlights the need for integrated nutritional and antiparasitic interventions (Cardona-Arias, Reference Cardona-Arias2018; Fauziah et al., Reference Fauziah, Aviani, Agrianfanny and Fatimah2022). Therefore, dietary interventions, including PUFA-rich diets, along with anthelminthic drugs, can improve treatment outcomes in helminthic infections, likely due to enhanced mucosal immunity and anti-inflammatory effects that complement the action of these drugs.
The environmental impact of chemical anthelmintics is a major concern, particularly in agricultural settings where these drugs are widely used. Residues of anthelmintics can persist in soil and water systems, posing risks to non-target organisms and contributing to the development of resistance in parasite populations (Beynon, Reference Beynon2012; Dimunová et al., Reference Dimunová, Matoušková, Navrátilová, Nguyen, Ambrož, Vokřál, Szotáková and Skálová2022). In contrast, PUFAs are naturally occurring compounds with minimal environmental toxicity, making them a safer alternative for parasite control. One of the key environmental benefits of PUFA-based interventions is their biodegradability. Unlike synthetic anthelmintics, which can persist in the environment for extended periods, PUFAs are rapidly broken down into non-toxic metabolites (Gong et al., Reference Gong, Fu, Liegl, Chen, Hellström and Smith2017; Qin et al., Reference Qin, Kurt, Lbassi, Sa and Xie2023). This reduces the risk of environmental contamination and minimizes the impact on non-target organisms, such as beneficial soil microbes and aquatic life. Another environmental advantage of PUFAs is their potential to enhance soil health and ecosystem resilience. For example, incorporating PUFA-rich plant residues into the soil can improve microbial diversity and nutrient cycling, promoting sustainable agricultural practices (Srivastava and Yadav, Reference Srivastava and Yadav2024). This is particularly important in regions where parasitic diseases are endemic, as improved soil health can reduce the transmission of STHs and other environmentally persistent parasites. Lastly, PUFAs are often derived from marine sources, such as seaweed or microalgae, which may result in a substantial reduction in the carbon footprint compared to terrestrial feed production (Hasselström and Thomas, Reference Hasselström and Thomas2022). Therefore, by integrating PUFAs into helminth control strategies, it is possible to achieve a balance between effective disease control and environmental sustainability.
Role of PUFAs in reducing anthelmintic resistance
Anthelmintic resistance is a growing global concern, particularly in veterinary medicine, where the overuse of anthelmintics has led to the emergence of resistant helminth populations. This resistance threatens the sustainability of current control programs and complicates efforts to manage important parasites in veterinary and public health (Raza et al., Reference Raza, Qamar, Hayat, Ashraf and Williams2019). PUFAs offer a promising alternative or adjunct to conventional anthelmintics, with the potential to reduce reliance on these drugs and slow the development of resistance. One of the key advantages of PUFAs is their potential to enhance the host’s natural resilience to parasites and to exert direct anthelmintic effects. As discussed earlier, omega-3 PUFAs contribute to parasite control by enhancing host type-2 immune responses and promoting macrophage activation, while also exerting direct antiparasitic effects such as membrane disruption and oxidative damage, together reducing parasite burdens and reliance on chemical anthelmintics. These mechanisms are distinct from synthetic anthelmintics, reducing the likelihood of cross-resistance and resistance development to PUFAs via single-gene mutations.
Combining PUFAs with synthetic anthelmintics offers another compelling opportunity to enhance therapeutic efficacy while mitigating the risk of resistance. PUFAs and conventional anthelmintics typically act via different mechanisms; anthelmintics target specific molecular pathways, such as neuromuscular junctions or microtubule formation (Martin, Reference Martin1997), whereas PUFAs may act through multiple mechanisms, including immune modulation, interference with membrane integrity, and induction of oxidative stress. Although studies in model organisms such as mice and poultry have shown that PUFAs can potentiate the effects of anthelmintic compounds (Allen et al., Reference Allen, Danforth and Levander1996; El-Ridi et al., Reference El-Ridi, Aboueldahab, Tallima, Salah, Mahana, Fawzi, Mohamed and Fahmy2010, Reference El-Ridi, Tallima, Salah, Aboueldahab, Fahmy, Al-Halbosiy and Mahmoud2012; Wu et al., Reference Wu, Liang, Zhu, Gao, Chen, Zhang, Yin, Li, Wang and Xiao2015; Yang et al., Reference Yang, Guo, Wang and Nie2006), the translation of this synergy into livestock animals remains unexplored. Studies reporting integrated approaches in veterinary helminth control that combine dietary or nutraceutical interventions with pharmacological treatments are limited. Recent work suggests that combining synthetic anthelmintics with plant-derived bioactives can enhance therapeutic efficacy. For example, co-administration of doramectin with cinnamaldehyde and pink grapefruit extract significantly improved efficacy against H. contortus without altering pharmacokinetics, likely through localized drug–phytochemical interactions that increase drug availability at the parasite interface (Miró et al., Reference Miró, Ichinose, Lloberas, Moriones, Lanusse, Virkel and Lifschitz2025). Despite the lack of evidence in livestock involving PUFA–anthelmintic combinations, evidence from rodent and in vitro models suggests that PUFAs may increase the toxicity of conventional drugs to helminth and protozoan parasites (El-Beshbishi et al., Reference El-Beshbishi, Saleh, Abd El-Mageed, El-Nemr, Abdalla, Shebl and Taman2019; Wu et al., Reference Wu, Liang, Zhu, Gao, Chen, Zhang, Yin, Li, Wang and Xiao2015), potentially by compromising parasite membranes or priming host immune responses. The combination of PUFAs with conventional anthelmintics also has the potential to reduce the dosage and frequency of chemical treatments, thereby reducing the risk of resistance and environmental contamination. For example, in regions where anthelmintic resistance is a concern, adding PUFAs to treatment regimens could enhance efficacy and extend the lifespan of existing drugs. This approach aligns with the principles of sustainable disease management, offering a cost-effective and environmentally friendly alternative to conventional strategies.
Challenges and future directions
Despite the promising role of PUFAs in modulating host immunity and exerting antiparasitic effects, several critical challenges hinder their broader application in clinical settings in human and veterinary medicine. These include issues related to bioavailability, oxidative stability, variable and low therapeutic response, and insufficient clinical and mechanistic data. Addressing these limitations is crucial for establishing PUFA-based therapies as viable and sustainable alternatives for controlling parasites.
Bioavailability and stability
One major obstacle to the efficacy of PUFAs is their limited bioavailability due to poor solubility, enzymatic degradation, and interactions with gut barriers. Although digestion by lipases and emulsification via bile salts facilitate absorption (Cansell et al., Reference Cansell, Nacka and Combe2003; Niot et al., Reference Niot, Poirier, Tran and Besnard2009; Singh et al., Reference Singh, Ye and Horne2009), factors such as meal composition and physiological conditions significantly influence uptake. Additionally, PUFAs are chemically unstable and prone to oxidative degradation, particularly in the presence of heme or metal ions, which can lead to cytotoxic by-products (Maestre et al., Reference Maestre, Douglass, Kodukula, Medina and Storch2013). To overcome these challenges, novel delivery systems, including microencapsulation, nanoemulsions, micellar matrices, and self-micro-emulsifying drug delivery systems, have been developed to enhance PUFA stability and tissue targeting (Cortés et al., Reference Cortés, Califano and Lorenzo2019; Hayashida et al., Reference Hayashida, Ishii, Adachi, Imai, Uotsu and Yui2022; Ibi et al., Reference Ibi, Chang, Kuo, Zhang, Du, Roh, Gahler, Hardy and Solnier2024; Joyce et al., Reference Joyce, Gustafsson and Prestidge2018; Kang et al., Reference Kang, Jeon, De, Hong and Park2023; Manusama et al., Reference Manusama, Balvers, Diepeveen-De Bruin, Headley, Bosi, Schwarm and Witkamp2021; Nunes et al., Reference Nunes, Pereira, Cerqueira, Silva, Pastrana, Vicente, Martins and Bourbon2020). Future research should focus on optimizing these technologies to ensure consistent delivery and effectiveness across diverse host systems.
Translational gaps and mechanistic uncertainty
Although numerous in vitro and experimental animal studies report PUFA-mediated antiparasitic effects, translation into clinical or field settings remains limited. The lack of standardized dosing, route-specific pharmacokinetics, and clinical trials poses a significant barrier. For example, while doses of 2–8 g/kg have been used in murine studies (Fasano et al., Reference Fasano, Serini, Cittadini and Calviello2017); comparable and adequate human or large-animal doses remain uncertain. Moreover, a mechanistic understanding, particularly regarding the interaction of PUFAs with the parasite membrane and parasite metabolic pathways, immune modulation (e.g., Th2-driven M2 macrophage activation), and ROS-mediated parasite killing, is still evolving (Zhao and Ma, Reference Zhao and Ma2014). Future studies utilising transcriptomic, proteomic, and metabolomic tools are essential for elucidating these pathways and informing rational drug design.
Socio-economic and implementation barriers
In developing countries where parasitic infections are most prevalent, challenges such as accessibility, affordability, and cultural acceptance of PUFA-based interventions further complicate implementation. While fish oil remains a key source of PUFAs, it is cost-prohibitive and ecologically unsustainable for widespread use. To address this, future research should prioritize plant-based and bioengineered PUFA sources, including genetically modified oilseeds and PUFA-rich microalgae or mosses (Duan et al., Reference Duan, Chen, Ma, Zhai, Hu, Li, Chen and Li2025; Ofori et al., Reference Ofori, Antoniello, English and Aryee2022). In parallel, community-level nutritional interventions using locally available PUFA-rich foods, e.g., seaweed, could provide an affordable and culturally appropriate strategy for parasite control, aligned with One Health principles (Fabjanowska and Kowalczuk-Vasilev, Reference Fabjanowska and Kowalczuk-Vasilev2023).
In summary, overcoming the pharmacological, translational, and socio-economic barriers to PUFA application will require an integrated, multidisciplinary research agenda. Combining advances in biotechnology, delivery systems, and mechanistic studies with public health strategies will be key to exploring the full potential of PUFAs as sustainable antiparasitic agents.
Conclusions
PUFAs offer a promising avenue for developing sustainable strategies for parasite control. Through their immunomodulatory, anti-inflammatory, and direct antiparasitic actions, PUFAs can strengthen host immunity, promote tissue repair, and impair parasite survival. These properties support effective disease control and also reduce dependence on conventional anthelmintics, thereby alleviating the dual challenges of drug resistance and environmental contamination. Importantly, PUFAs align strongly with the One Health framework: in humans, they may help mitigate the health and socio-economic burdens of parasitic diseases; in livestock, they can improve productivity and welfare while decreasing chemical inputs; and in the environment, their low toxicity and biodegradability make them a safer alternative to synthetic drugs.
At the same time, several barriers still constrain their translation into practice. Variability in therapeutic response, limited bioavailability, and a lack of robust clinical and field trials remain significant challenges. Addressing these gaps will require coordinated investment in research that bridges laboratory models with real-world settings. Advances in crop bioengineering and microalgal production may help secure scalable and sustainable PUFA sources, while novel delivery platforms, such as encapsulation and nanoemulsions, could improve stability, absorption, and targeted activity. Combination approaches that integrate PUFAs with conventional anthelmintics may also enhance efficacy while reducing drug pressure and slowing resistance development.
Future progress will depend on integrating mechanistic research with applied field studies to define optimal dosing strategies and delivery platforms across species and production systems. Interdisciplinary collaboration will be critical to ensure that PUFA-based strategies are practical, accessible, affordable, and scalable, particularly in resource-limited settings. By harnessing the unique properties of PUFAs and embedding them into holistic One Health frameworks, it may be possible to build more sustainable parasite control programs that address pressing global challenges of resistance, food security, and environmental stewardship.
Acknowledgements
Figures were created with www.BioRender.com.
Author contributions
AR: Funding acquisition, Conceptualization, Literature review, Investigation, Visualization, Writing – Original draft, and Writing – Review & editing. MB: Literature review, Data curation, and Writing – Review & editing. GF: Visualization and Writing – Review & editing. SMT: Supervision, Conceptual guidance, and Writing – Review & editing. ARW: Funding acquisition, Supervision, and Writing – Review & editing.
Funding statement
This work was supported by the European Commission through the ‘Marie Skłodowska-Curie Postdoctoral Fellowship program (Grant No. 101025249)’ and the Independent Research Fund Denmark (Grant 1127-00255B), which provided the primary funding for research forming the basis of this review.
Competing interests
The authors declare that they have no competing interests.

 Open access
Open access