Introduction
One of the main objectives of paleoanthropology is the identification of the socio-reproductive organization and subsistence strategies that shaped human behavior. During the 1970s and 1980s, several models emerged to interpret socioeconomic behaviors of early hominins through the analysis of the early Pleistocene archaeological record in Africa. Some models emphasized behaviors that resembled those of some modern foraging populations (e.g., Leakey’s “living-floors” [Leakey, Reference Leakey1971]; and the “home base” or “food-sharing” models [Isaac, Reference Isaac1978, Reference Isaac1983]). Subsequent revisionist models (with a concern for site formation processes) during the 1980s produced ethological models in which hominins had social behaviors that were reconstructed as more similar to those of other nonhuman primates or inspired by them, such as the marginal or obligate scavenger (Binford, Reference Binford1981) and the passive scavenger (Blumenschine, Reference Blumenschine1986) models, the stone-cache model (Potts, Reference Potts1988), the “chimpanzee-nesting” model (Sept et al., Reference Sept, King, McGrew, Moore, Paterson and Strier1992), or the “refuge” model (Blumenschine, Reference Blumenschine1991; Blumenschine et al., Reference Blumenschine, Cavallo and Capaldo1994).
The information gap between the reconstruction of site formation (through the taphonomic analysis of materials preserved at sites) and the hominin socioeconomic organization required to sustain any of the above models led scholars to abandon social and functional interpretations based on the modeling of early sites. Isaac’s “central-place foraging” model (Isaac, Reference Isaac1983) de-emphasized social aspects of his previous “home base” model, and Cavallo (Reference Cavallo1997) even managed to reconcile it with passive scavenging models. Schick’s “favored place” model did not include any significant social components, and stressed that sites could simply be created by unintentional reuse of certain spaces (Shick, Reference Shick1987). These may even have served as secondary sources of raw material (Plummer et al., Reference Plummer, Ditchfield, Bishop, Kingston, Ferraro and Braun2009). The “near-kill location” model (O’Connell, Reference O’Connell1997) or the “male display” model (O’Connell et al., Reference O’Connell, Hawkes, Lupo and Blurton Jones2002) did not emphasize any specific social organization, despite depicting sites as carcass obtainment loci created through confrontational scavenging to increase male mating fitness. Even though some authors suggested an evolved version founded on general primate and mammal behavior to explain early sites (e.g., the “resource-defense” model) (Rose and Marshall, Reference Rose and Marshall1996), most models produced during the past four decades have approached Early Pleistocene hominin behavior by making it similar to those of other primates in an increasingly dehumanizing trend. One of the last models produced, the “obligate carnivory” model (Ferraro, Reference Ferraro2007) intentionally avoided any interpretation of the social behavior of hominins or of the functionality of sites beyond their reconstruction as places where hominins ate substantial amounts of meat. This avoidance of hominin social organization is surprising, since in ethology it is widely known that subsistence/adaptive behavior is strongly dependent on socio-reproductive strategies (Brooks and McLennan, Reference Brooks and McLennan1991). All this leads to the conclusion that the current interpretation of early archaeological sites is not only confusing, for the array of diverse and widely divergent views, but also highly speculative, because most of the models are disconnected from a hypothetical-deductive framework with which to test their main arguments. In addition, one could question that these interpretations should be called models, since models, as representation of systems, should be conceptual constructs; that is, an intertwined representation of entities (i.e., behavioral features) and their relationships (i.e., dynamics) articulated to a substantial degree on an empirical foundation (Domínguez-Rodrigo, Reference Domínguez-Rodrigo and Domínguez-Rodrigo2012a).
This increasing trend of focusing on technological and eco-dietary issues, framing them fundamentally within the primate order, has led to an incomplete understanding of what early sites represented in early human evolution. Although there is a substantial amount of information available about the subsistence of hominins at a small number of Early Pleistocene sites, it is fair to state that we know very little about early site functionality (i.e., what was done at those loci beyond carcass butchery), and about hominin socio-ecology. In addition, there is a potential confusion among the large diversity of interpretations of Early Pleistocene hominin subsistence strategies, as observed in the array of behavioral models produced. How can the heuristics of these models be empirically tested? This diversity of interpretations may be due to the controversial nature of an insufficient archaeological record and/or to flawed theoretical framing of these models (see critical discussion in Domínguez-Rodrigo, Reference Domínguez-Rodrigo2012a, b).
Another biasing factor to understand early sites is that paleoanthropologists have adopted a clearly homologous paradigm, by framing human evolution with continuous reference to the primate order, or more specifically, to extant apes. This has conditioned research in two ways: One, by backtracking hominin physical evolution towards the chimpanzee anatomy (taking chimpanzees as a model for the last common ancestor [LCA]) (Muller et al., Reference Muller, Wrangham and Pilbeam2017); and two, by taking greater apes, and more specifically chimpanzees, as homological models for the behavior of early humans (Carvalho et al., Reference Carvalho, Matsuzawa, McGrew, Sanz, Call and Boesch2013, Reference Carvalho, Cunha, Sousa and Matsuzawa2008; Carvalho and McGrew, Reference Carvalho, McGrew and Domínguez-Rodrigo2012; Haslam et al., Reference Haslam, Hernandez-Aguilar, Ling, Carvalho, de la Torre and DeStefano2009; Sept et al., Reference Sept, King, McGrew, Moore, Paterson and Strier1992; Thompson et al., Reference Thompson, Carvalho, Marean and Alemseged2019; Toth and Schick, Reference Toth and Schick2009). For the first point, there is a wealth of research suggesting that such a view may be misguided. Most Miocene apes did not anatomically resemble extant hominines, since their environments were less forested, much more seasonal, their diets were more varied, their dentition evolved thicker enamel, and their locomotion was different (Alba et al., Reference Alba, Almécija, DeMiguel, Fortuny, Pérez de los Ríos and Pina2015; Almécija et al., Reference Almécija, Hammond, Thompson, Pugh, Moyà-Solà and Alba2021; Andrews, Reference Andrews2016; Begun, Reference Begun2015). Some of the earliest hominins also contradict a chimp-like anatomy and locomotion for the LCA (White et al., Reference White, Lovejoy, Asfaw, Carlson and Suwa2015, Reference White, Asfaw, Beyene, Haile-Selassie, Lovejoy and Suwa2009). For the second point, although taking extant apes as the closest neuro-anatomical analogs may be reasonable, restricting the behavior of Early Pleistocene Homo to what is observed only in modern apes may also be biasing, since early humans at early sites were adopting behaviors that are not documented in chimpanzees nor in any other primate. These include, among others, dependence on stone-tool manufacture, butchery and consumption of large animals, long-distance transport of food and raw materials, and redundant selection of specific loci in the landscape to bring and process food. In several of these characteristics, early humans were more similar to other animals, such as several mammal carnivores, birds, and eusocial insects.
Isaac’s (Reference Isaac1978) diagnosis of humanness as the list of features that separate us from other primates emphasized the view of human uniqueness. This view has been highly popular because it connects with Judeo-Christian cultural anthropocentric cosmovisions, and also because conceptually it seems derived from the neoplatonic Great Chain of Being philosophical school, which so impacted Western cultures (Lovejoy, Reference Lovejoy2021). Indirectly, this perspective is at the core of several of the models listed above. However, this approach depends tightly on the analogical reference(s) chosen. If we take extant apes (or even all extant primates) as the reference, it is true that Isaac produced a list of features that can be observed uniquely in modern humans (Table 1.1). However, if we extend the comparison to the rest of the organic world, many of these features cease to be structurally unique. This brings forward the question of which analogical references should we select to understand human-purported uniqueness? Should we indeed keep focusing on our unique behavioral features (compared to primates) or would we gain a much better insight thereof and of the rest of what it means to be human if we frame Homo sapiens within as broad an organic framework as possible? As conceptually self-evident as it seems, investing the same effort in discerning connecting features with other organisms, as attempting to follow the evolutionary progress of features that may potentially be unique to humans, will result in a better understanding of human evolutionary trajectories. Although some efforts have been made in linking early humans with our closest relatives (see most research done on chimpanzee use of technology or what has been referred to as primate archaeology, for example), these approaches have been rather conceptually limited, because the goal was to show that early humans did not need to be substantially different from modern chimpanzees.
Here, we will argue that in order to understand the function that early sites played in the lifestyle and adaptation of early humans, we need to adopt an evolutionary biological approach in which we view hominins as part of the organic network, and subjected to the same constrictions, patterning, and adaptive solutions as many other species of organisms. This view may be unpopular, because it emphasizes what humans have in common with a plethora of organic life forms, and returns humans to the rather “unexceptional” animal realm. This perspective is holistic and integrative, and provides explanation for aspects of hominin adaptation, and the role played by early sites that have either been neglected or could not be interpreted in previous interpretations. This view stems from the use of evolutionary behavioral ecology. As Strier (Reference Strier2015) puts it:
Behavioral ecology helped to shift anthropocentric perspectives in primatology away from the focus on finding referential behavioral models for human social evolution. Instead, strategic models, which emphasized the evolutionary and ecological processes that affect behavior, were developed to provide an alternative way of understanding behavioral adaptations.
It should be emphasized that part of the reasons why so many interpretations about early hominin sites have been made has a conceptual basis. Most of the existing models make two indirect assumptions: Sites are unstructured (i.e., hominins [like chimpanzees] did not use the locus space in any structured manner), and site configuration is, thus, stochastic. The latter has major relevance, because it assumes that such a stochasticity is due also to palimpsestic intervention of post-depositional processes that impacted the original hominin-made assemblage configuration. It also assumes that hominins were disturbing agents by randomly intervening at different moments at sites, with probably each occupation being spaced along an extended time sequence. Therefore, taphonomic biases and time-averaging are the two main elements that prevent archaeologists from going beyond the interpretation that the only well-supported inference that can be made from the early archaeological record is that sites represent loci where hominins made some tools and processed animal carcasses. However, are these commonsensical premises really supported by the available evidence?
Recent Advances on Oldowan Site Integrity and Resolution for Interpreting Early Human Behavior
Key 1. Not All Oldowan Sites Are Equally Useful: The Importance of Selecting a High-Quality Record
Interpretations like the “stone cache” model (Potts, Reference Potts1988) or the “favored place” model (Shick, Reference Shick1987) projected hominin redundancy in site use in a long-term time scale represented by several years. Randomness in the timing and use of the space pervades these models. It is, thus, not surprising that site structure was not a concern, and that integrity and resolution,Footnote 1 especially the latter, were considered moderate to low. These assumptions force archaeologists to deal with the most difficult variable in archaeological research: The time span represented in any given archaeological record. How much time is commonly represented in the formation of the Oldowan bone and lithic assemblages? Obviously, there must exist a wide range of variation, with some assemblages being formed through long-term (i.e., multi-yearly) intervention of diverse agencies, and others formed by faster depositional and behavioral processes, with a more homogeneous agency and limitation of the number of activities dynamically contributing to the accretional formation of the deposit. Otherwise said, there must be sites with extremely low integrity and resolution (i.e., multi-layered palimpsests), and others where these variables qualify as high. Adding both types of records for meta-analysis, especially through a timeline, is bound to create biases and distort the empirical record to an extreme in which arguments supporting interpretations may lack heuristics, and may easily adapt to any given interpretation and its opposite at the same time. This distortion has recently led to what some describe as the anomalous “Olduvai effect” (Barr et al., Reference Barr, Pobiner, Rowan, Du and Faith2022). This refers to part of the archaeological record found at Olduvai Gorge in Tanzania that deviates from the typical hominin impact documented on most other archaeofaunal assemblages in Africa, by showing a substantially higher hominin modification signal. Are some selected sites from Olduvai “anomalous” in exhibiting a significantly higher frequency of hominin butchery-related modifications compared to most other sites or is this an artifact of method resulting from a predominant background of Oldowan sites in other locations lacking similar preservation properties and containing lower integrity and resolution? We argue that the latter argument is the reason. We emphasize that both sets of sites should not be analytically compared, because this introduces strong biases. We also argue that what some consider “anomalous,” could actually be “normative” when such sites show repeated structure and patterning. This view opposes a “patchwork” reconstruction of early human behavior based on aggregating information from diverse sites whose taphonomic histories and biases are varied and widely unknown.
We can concur with other researchers in assuming that several (probably most) Oldowan sites may be characterized by limited integrity and resolution; however, we stress that this needs to be empirically demonstrated instead of assumed (Domínguez-Rodrigo, Reference Domínguez-Rodrigo, Hovers and Braun2009a). A major problem with this is that preservation at many of these assemblages may be insufficient to test their integrative and resolutive properties. For example, this may be the case with most of the Koobi Fora Early Pleistocene record documented so far, which has been impacted by biostratinomic and diagenetic processes to a much larger extent than assemblages at Olduvai, leading to leeched bones and poor cortical preservation. This prevents proper taphonomic discernment of agency and the range of depositional events (Isaac and Isaac, Reference Isaac and Isaac1997). As a proof thereof, in the few well-preserved assemblages discovered at Koobi Fora, the anthropic signal spikes up compared to the background sample (Pobiner et al., Reference Pobiner, Rogers, Monahan and Harris2008).
We stress that because of the taphonomically impacted archaeofaunal fossil record, efficient and heuristically solid interpretations on Oldowan hominin behavior can only be made using exceptionally preserved anthropogenic assemblages. This should lead archaeologists to abandon the use of bulk site samples and focus on well-preserved and taphonomically supported anthropogenic records. This, unfortunately, may lead researchers to use very small samples of sites where confidence in their behavioral content and human agency is high. This practice is not new. As a matter of fact, we have been doing this during the past half century. Several of the models described above have been repeatedly tested against one single “anomalous” site (FLK Zinj, Bed I, Olduvai Gorge) (Binford, Reference Binford1981; Binford et al., Reference Binford, Bunn and Kroll1988; Blumenschine, Reference Blumenschine1995; Bunn, Reference Bunn1982; Bunn et al., Reference Bunn1986; Capaldo, Reference Capaldo1997; Domínguez-Rodrigo and Barba, Reference Domínguez-Rodrigo and Barba2007a, Reference Domínguez-Rodrigo and Barba2006; Pante et al., Reference Pante, Blumenschine, Capaldo and Scott2012; Potts, Reference Potts1988; Selvaggio, Reference Selvaggio1994). Only more recently have archaeologists expanded this to another well-preserved site: Kanjera (Kenya) (Ferraro et al., Reference Ferraro, Plummer, Pobiner, Oliver, Bishop and Braun2013; Oliver et al., Reference Oliver, Plummer, Hertel and Bishop2019; Parkinson, Reference Parkinson2013; Parkinson et al., Reference Parkinson, Plummer, Oliver and Bishop2022; Plummer et al., Reference Plummer, Ditchfield, Bishop, Kingston, Ferraro and Braun2009). This situation is not fortuitous. It is a reflection of generally poorly preserved faunal assemblages at Oldowan sites, and the awareness of the immense interpretive potential of the rather limited sample of exceptionally preserved anthropogenic sites. Our confidence in our interpretations will be thus boosted by the quality of the record used, and not by its sample size, given the large array of biasing processes and preservation issues impacting on the formation of sites.
How can we then define this exceptionally preserved anthropogenic record? Isaac’s (Reference Isaac1978) and Isaac and Crader’s (Reference Isaac, Crader, Harding and Teleki1981) nomenclature of site types showed that the integrity-resolution binomial was first determined by contextual sedimentary dynamics. These authors differentiated between sites clearly affected by abiotic post-depositional processes and time-averaging (in the form of vertically distributed deposits), and vertically discrete assemblages, suggestive of less tractive sedimentary processes and more time-restrictive depositional dynamics. Within the latter types of sites, Isaac further differentiated among diverse configurational and behavioral varieties, with variable impact on integrity and resolution. Contextual information was essential to this, because it provides evidence of site formation and paleoecological information supporting or discarding potential impacts by physical processes (e.g., hydraulic jumbles) or by behavioral processes (e.g., common-amenity scenarios) in site formation (Isaac, Reference Isaac1983). From this perspective, with the purpose of inferring site functionality and dealing more efficiently with time-averaging processes, we argue that research efforts should focus on vertically discrete assemblages, where contextual information exists on post-depositional sedimentary dynamics and the timing of the biostratinomic-diagenetic interface. It is important to remark that only in such contextual circumstances “time” can be approached efficiently by archaeologists.
Once restricted to such a contextually high-quality record, researchers should select taphonomically defined anthropogenic sites. These are defined as assemblages accumulated in specific locations by hominins, and modified either almost exclusively or primarily by them (with or without interaction with other biotic agents). This implies sites that preserve enough information to determine their integrity. For this purpose, good bone surface preservation and limited bone dry breakage are essential. The resource extractive processes (namely, defleshing, demarrowing, and grease consumption) by hominins and other carnivores leave their traces in the way that bones are cortically modified and dynamically broken. This information needs to be retrieved from such well-preserved assemblages. Only in these types of sites can integrity (i.e., agencies) be confidently determined.
Resolution (i.e., number of activities or depositional events) has also been argued to be a major problem for correctly inferring site functionality (Binford, Reference Binford1981). Single-event sites (e.g., single-carcass butchery and kills sites) are supposed to better preserve behavioral information because of lack of successive overlying depositional events; however, this should be nuanced. We must differentiate between sequential activity processes (SAP) and sequential depositional events (SDE). SAP involves different activities performed by the same or different agents overlapping in their depositions on the same spot. SDE can be defined as the same activity performed by the same agent carried out repeatedly on the same spot. SAP does not require accretional generation of materials, whereas SDE does. Contrary to what might be thought, SAP can lead to substantial modification of a previous depositional event, while SDE may actually preserve its properties. For example, carnivore ravaging of human-deposited bone assemblages may modify substantially their original compositional and spatial properties without adding more elements to them. In contrast, redundant butchery episodes by humans on the same location may generate an increasing amount of materials without necessarily distorting the compositional and spatial properties of previous depositional events. This is especially so if the process is carried out longitudinally without performing any cleaning, as some modern human foragers do when they stay in the same camps for weeks or months. In this case, SDE will be increasingly informative, whereas SAP will be biasing in terms of hominin behavior. In both cases, the sequential process will be taphonomically informative (Behrensmeyer et al., Reference Behrensmeyer, Kidwell and Gastaldo2000). One consequence of this is that low resolution (i.e., the impossibility of splitting different depositional events and their material byproducts) does not need to be detrimental, especially if the sequential process is single-agent (high integrity). The detrimental effects are more likely to be detected in cases of moderate to low integrity, especially in situations of multiple agent interactions. In this situation, for cases of time-limited accumulations, integrity and not resolution is the fundamental variable.
The traditional approach to this question on site formation is a quantitative description of resolution. Complementarily, there is a qualitative interpretation thereof. This is based not on the total number of depositional events, but on the number of activities performed according to agency. For example, in the absence of ecological constrictions, the same agent (e.g., humans) can carry out multiple activities in different immediate loci within the same occupational space, like modern foragers do. These do not need to impact one another unless cleaning is performed. Even when same-agent activities leading to SAP are sequentially performed on the same spot, these do not need to alter the properties of previous depositional events. Attempting to unravel the effects of single events and their material results in such circumstances may be certainly challenging. However, this is not required if targeting interpretations on the use of the space if the same type of activity is repeatedly performed on the same spots, and by extension, if determining the adaptive/behavioral functions of any given site. For these reasons, targeting sites with high integrity and high resolution (if referring to limited sequential activities by different agents [qualitative resolution]) or moderate to low resolution (if referring to more sequential activities by the same agent [quantitative resolution]) can still enable interpreting how space at any given site was used.
When we apply these requirements to the Early Pleistocene fossil record of East Africa, we have to come to the conclusion that there is a dearth of sites that are vertically constricted, in low-energy depositional settings, with good to optimal faunal preservation, with a clear anthropogenic predominant taphonomic signature, and preserving their original spatial configuration. Even without the latter feature, the limited number of existing Oldowan anthropogenic assemblages is a limiting factor in the interpretation of our origins, compensated only by their sometimes exceptionally preserved information (Table 1.2). It is our duty to be able to make this information intelligible.
| Locality | Stratigraphic unit | Number of lithic specimensFootnote * | Number of bone specimensFootnote * | TypeFootnote ** | Bone preservation | Anthropic origin of archaeofaunal assemblage demonstrated | |
|---|---|---|---|---|---|---|---|
| DK 1-3 | Olduvai | Bed I | 1.198 | 9.984 | D | good | |
| FLKNN1 | Olduvai | Bed I | 16 | 275 | C | good | |
| FLKNN3 | Olduvai | Bed I | 48 | 2.158 | C | good | |
| FLK Zinj | Olduvai | Bed I | 2.470 | 3.510 | C | good | x |
| DS | Olduvai | Bed I | 1229 | 3458 | C | good | x |
| PTK | Olduvai | Bed I | 1004 | 2589 | C | good | x |
| AGS | Olduvai | Bed I | In progress | In progress | C | good | x |
| FLKN 6 | Olduvai | Bed I | 123 | 614 | B | good | |
| FLKN 5 | Olduvai | Bed I | 151 | 2.210 | D | good | |
| FLKN 4 | Olduvai | Bed I | 67 | 929 | D | good | |
| FLKN 3 | Olduvai | Bed I | 171 | 1.254 | D | good | |
| FLKN 1-2 | Olduvai | Bed I | 1.205 | 3.294 | C | good | |
| HWK East 1 | Olduvai | Bed II | 154 | 425 | C | good | |
| HWK East 2 | Olduvai | Bed II | 313 | 631 | D | good | |
| HWK EE | Olduvai | Bed II | 4351 | 859 | D | good | |
| FLKN Deino | Olduvai | Bed II | 39 | - | B | good | |
| FLK West | Olduvai | Bed II | 2790 | 1042 | C | moderate-bad | |
| HWK East 3-5 | Olduvai | Bed II | 1.989 | 269 | D | good | |
| EF-HR | Olduvai | Bed II | 522 | 34 | A | good | |
| FC West Tuff | Olduvai | Bed II | 673 | 254 | A | moderate | |
| FC West Floor | Olduvai | Bed II | 1.184 | 127 | A | moderate | |
| MNK Main (all levels) | Olduvai | Bed II | 4.399 | 1.723 | D | good | |
| SHK | Olduvai | Bed II | 915 | 34 | D | good-moderate | |
| TK upper floor | Olduvai | Bed II | 5.180 | 230 | A | bad | |
| TK lower floor | Olduvai | Bed II | 2.153 | 147 | A | bad | |
| BK | Olduvai | Bed II | 6.801 | 2.957 | D | good | x (depending on level) |
| FxJj1 | Koobi Fora | KBS | 140 | 680 | D | bad | |
| FxJJ3 | Koobi Fora | KBS | 122 | 237 | B-D | bad-moderate | |
| FxJj20 | Koobi Fora | Karari | 4437 | 4574 | C | bad | |
| FxJj50 | Koobi Fora | Karari | 1535 | 2120 | C | bad-moderate | Footnote *** |
| FxJj64 | Koobi Fora | Okote | 45 | 578 | D | bad | |
| KS1 | Kanjera | Kanjera | 179 | 982 | D | good | x |
| KS2 | Kanjera | Kanjera | 2533 | 2153 | D | good | x |
| KS3 | Kanjera | Kanjera | 171 | 491 | D | good | x |
| AB-Lw/Up | Ain Boucherit | Ain Hanech | 17/834 | 296/277 | D | moderate | Footnote *** |
| EK | El-Kherba | Ain Hanech | 811 | 616 | D | moderate | Footnote *** |
Only sites with extensive excavations, preserved fauna, and clear stratigraphic context have been included.
* Data from Leakey (Reference Leakey1971, table 6) for sites discovered by Leakey at Olduvai. Data from Koobi Fora from Isaac and Isaac (Reference Isaac and Isaac1997). Data for Kanjera from Ferraro et al. (Reference Ferraro, Plummer, Pobiner, Oliver, Bishop and Braun2013). Data for Ain Boucherit and El Kherba from Sanhouni et al. (Reference Sanhouni, Rosell, van der Made, Verges, Ollé and Kandi2013, Reference Sanhouni, Pares, Duval, Cáceres, Harichane and van der Made2018).
** Isaac and Crader (Reference Isaac, Crader, Harding and Teleki1981)
*** Hominin input is detected in the form of some specimens bearing cut marks. Anthropic accumulation of assemblage is likely but needs to be confirmed taphonomically at the assemblage level.
Key 2. Taphonomic Constriction of Time: Placing a Boundary to Time-Averaging
Taphocoenoses (sensu Fernández-López, Reference Fernández-López2006, Reference Fernández-López1995) are dynamic entities subjected to constant change. Pioneering experimental/observational work in the 1980s (Behrensmeyer and Hill, Reference Behrensmeyer and Hill1988; Hill, Reference Hill1979a, Reference Hill1979b; Hill and Behrensmeyer, Reference Hill and Behrensmeyer1985, Reference Hill and Behrensmeyer1984), building upon the foundational neotaphonomy from the early 1900s (Weigelt, Reference Weigelt2009) demonstrated how anatomically associated elements are subjected to centrifugal processes that tend to disperse them over time. These phenomena can be taken as relative timers of the biostratinomic timeline. Although such a modification of the original associations of elements and element portions is not linear, with most of it taking place in the early stages of deposition, it certainly continues through time (Allison and Bottjer, Reference Allison, Bottjer, Allison and Bottjer2011, Reference Allison and Bottjer2010; Aslan and Behrensmeyer, Reference Aslan and Behrensmeyer1996; Flessa et al., Reference Flessa, Cutler and Meldahl1993; Gupta and Briggs, Reference Gupta, Briggs, Allison and Bottjer2011; Itoh et al., Reference Itoh, Fernández-Jalvo, Andrews and Inaba2019; Karr and Outram, Reference Karr and Outram2012; Kidwell and Behrensmeyer, Reference Kidwell and Behrensmeyer1993; Martin et al., Reference Martin and Autor Martin1999; Staff et al., Reference Staff, Stanton, Powell and Cummins1986). Biotic processes disperse elements, and even element-related fragments, in just a few weeks (Domínguez-Rodrigo and Lezana, Reference Domínguez-Rodrigo and Lezana1996). Abiotic processes do the same, but much more discontinuously (Rogers et al., Reference Rogers, Eberth and Fiorillo2010; Rogers and Kidwell, Reference Rogers, Kidwell, Rogers, Eberth and Fiorillo2007; Wood et al., Reference Wood, Thomas and Visser1988). Controlled ethnoarchaeological work shows how even long bone shaft fragments, devoid of nutritional resources after human demarrowing, can be dispersed by small carnivores and birds (Domínguez-Rodrigo and Lezana, Reference Domínguez-Rodrigo and Lezana1996). Therefore, spatial dispersal of elements and its constituent parts, as well as de-structuralization (i.e., loss of the spatial and compositional properties) of assemblages tend to increasingly occur naturally through time. Ethnoarchaeological work in East Africa, under the typical African savanna biome competitive conditions, shows that it may just take a few weeks for disaggregation and dispersal to split original depositional associations by biotic agents over distances of several meters between closely associated parts (e.g., antimeric elements) and hundreds of meters of same-individual anatomical elements (Domínguez-Rodrigo and Lezana, Reference Domínguez-Rodrigo and Lezana1996). Therefore, close structural and spatial association of elements that constitute originally joint depositional clusters is indicative of very limited impact of biostratinomic processes. Indirectly, it could also be a relative indicator of the time that any given archaeofaunal assemblage has been exposed prior to sedimentation.
A less relative approach to the timing of site formation comes from the way that bones dehydrate and weather through subaerial exposure (Behrensmeyer, Reference Behrensmeyer1978). In modern East Africa, this process has been timed through a controlled longitudinal study of animal carcasses. In such a study, involving thirty-five animal skeletons, it was shown that bones show gradual weathering in several stages (Behrensmeyer, Reference Behrensmeyer1978). Such a weathering process is conditioned by moisture and exposure to high temperatures. Bones exposed on the surface between 0–3 years showed as many as three different weathering stages, with stage 1 (some longitudinal cracking parallel to the bone collagen structure) being the most common. Bones exposed for longer periods showed more intensive cortical modification characterized in successive weathering stages. It is important to stress that habitat type (more or less wooded) did not play a significant variation in the timing of the expression of different weathering stages. It is the local conditions at a micro scale that determine within-habitat variations, with all habitats displaying similar stages of representation of different weathering stages.
Despite the potential variability in the occurrence of the different weathering stages, it should be stressed that “it is possible to distinguish carcasses exposed for less than 3 years with fairly certainty using weathering stages” (Behrensmeyer, Reference Behrensmeyer1978). Under the Amboseli environmental conditions, bones exhibit “significant weathering by the time they are 3–5 years old, and most disintegrate in less than 15 years” (Behrensmeyer, Reference Behrensmeyer1978). This was documented in subsequent studies in other East African locations with similar distribution of the different weathering stages (Lala, Reference Lala2011). A recent study of carcasses deposited over a four-year period in a leopard lair, with dense vegetation and lower temperature contrast than documented in Amboseli, showed that weathering stages 1–2 could appear in several bones early in time (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Organista, Baquedano, Cifuentes-alcobendas, Pizarro and Vegara2022b). Weathering rates within specific-stage ranges seem to be chronologically constricted for earlier stages of bone weathering (stages 0–2: 0–3 years), and more variable for subsequent stages (stages 3–5: 3–15+ years) (Behrensmeyer, Reference Behrensmeyer1978). Therefore, under similar equatorial climatological conditions, the early stages of bone weathering when considered at the assemblage level can provide a relative estimate of time since deposited on the ground. This is especially so in soil surfaces that do not frequently contain high doses of moisture. The effectiveness of bone weathering to infer timing decreases as one moves away from the equator and contrasts in temperature and moisture become less marked (Fernández-Jalvo et al., Reference Fernández-Jalvo, Andrews, Pesquero, Smith, Marín-Monfort and Sánchez2010; Itoh et al., Reference Itoh, Fernández-Jalvo, Andrews and Inaba2019).
Key 3. Selecting Exceptional Oldowan Sites
The Olduvai Bed I archaeofaunal assemblages have been considered as a good example of time-limited paleobiological records, because several of them occur in vertically discrete horizons (Leakey, Reference Leakey1971). The FLK Zinj-FLKNN-1 penecontemporaneous assemblages are specifically documented within a distinctive ~20 cm clay layer (Ashley et al., Reference Ashley, Barboni, Domínguez-Rodrigo, Bunn, Mabulla and Diez-Martin2010a, Reference Ashley, Barboni, Domínguez-Rodrigo, Bunn, Mabulla and Diez-Martin2010b; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Bunn, Mabulla, Baquedano and Pickering2010a; Hay, Reference Hay1976; Leakey, Reference Leakey1971). Research specifically focused on this thin clay stratum over the past fifteen years has detected that there are two clearly distinctive depositional periods documented within this unit: One occurring on the top of the clay, and the other one occurring at the bottom, with a clear sedimentary hiatus in between, further constricting both archaeological levels to ~5–10 cm each (Aramendi et al., Reference Aramendi, Uribelarrea, Arriaza, Arráiz, Barboni and Yravedra2017; Arráiz et al., Reference Arráiz, Barboni, Ashley, Mabulla, Baquedano and Domínguez-Rodrigo2017; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Bunn, Mabulla, Ashley, Diez-Martin and Barboni2010b; Domínguez-Rodrigo and Cobo-Sánchez, Reference Domínguez-Rodrigo and Cobo-Sánchez2017a; Martín-Perea et al., Reference Martín-Perea, Fesharaki, Rey Samper, Arroyo, Uribelarrea and Cobo-Sánchez2019; Uribelarrea et al., Reference Uribelarrea, Domínguez-Rodrigo, Pérez-González, Vegas Salamanca, Baquedano and Mabulla2014). This is as vertically constricted as can be. Additionally, over the past twelve years, new sites (PTK, AMK, DS, AGS) have been found, some of them containing archaeological levels both in the upper and lower portions of this clay unit (Aramendi et al., Reference Aramendi, Uribelarrea, Arriaza, Arráiz, Barboni and Yravedra2017; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo and Cobo-Sánchez2017a; Domínguez-Rodrigo and Cobo-Sánchez, Reference Domínguez-Rodrigo and Cobo-Sánchez2017a) (Figure 1.1). The upper level was in the process of sedimentation when a volcanic explosion creating Tuff IC sealed the paleo-landscape, simultaneously burying all these assemblages. Given the shallow morphology of the Olduvai lacustrine basin (Hay, Reference Hay1976), this would be indicative of the volcanic event occurring during a transgressive episode, most likely during the wet season. This synchronicity of sedimentary processes across different records on the same landscape is difficult to match. Both archaeological levels formed on top and at the bottom of the clay also suggest that hominin occupation must have taken place during the dry season, during regressive lake episodes. This would also explain why the bulk of the fauna exploited by hominins at these loci was local (Linares-Matás and Clark, Reference Linares-Matás and Clark2022).
Location of the Oldowan sites of Olduvai Bed I.

There is a wealth of taphonomic analyses on some of these assemblages showing that: (a) They are autochthonous (i.e., they were retrieved in their original depositional locus); (b) that they were impacted marginally by abiotic post-depositional processes; (c) that the impact of carnivore post-depositional ravaging was not important in affecting the overall site spatial configuration properties (see below); (d) that there was not a significant degree of sedimentary compaction to affect the paleo-relief where these sites were formed; and, most importantly, (e) that most of the sites (FLK Zinj, DS, PTK, AGS) are of anthropogenic origin (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Baquedano, Organista, Cobo-Sánchez, Mabulla and Maskara2021, Reference Domínguez-Rodrigo, Baquedano, Barba, Uribelarrea and Gidna2019a, Reference Domínguez-Rodrigo, Uribelarrea, Santonja, Bunn, García-Pérez and Pérez-González2014b, Reference Domínguez-Rodrigo2012, Reference Domínguez-Rodrigo, Barba and Egeland2007; Martín-Perea et al., Reference Martín-Perea, Fesharaki, Rey Samper, Arroyo, Uribelarrea and Cobo-Sánchez2019; Uribelarrea et al., Reference Uribelarrea, Domínguez-Rodrigo, Pérez-González, Vegas Salamanca, Baquedano and Mabulla2014). An additional proof of these statements, supporting this taphonomic evidence, comes from the spatial configuration of several of these sites, which shows virtually identical properties in spatial inhomogeneity, clustering, intensity, and composition (Domínguez-Rodrigo and Cobo-Sánchez, Reference Domínguez-Rodrigo and Cobo-Sánchez2017a). This contrasts with what would be expected had the assemblages undergone any significant modification by biostratinomic tractive and sedimentary processes, and had they resulted from stochastic physically induced resedimentation (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Baquedano, Barba, Uribelarrea and Gidna2019a, Reference Domínguez-Rodrigo and Cobo-Sánchez2017b). This also indicates that sites have patterning and, thus, similar structure. The behavioral implications thereof cannot be overemphasized (see below).
There are three additional elements that make this autochthonous record exceptional: Integrity, resolution, and formation time. All available taphonomic data indicate that hominins were the main/exclusive accumulating agents at these anthropogenic sites bringing together bones and stone artifacts in close functional association (Binford et al., Reference Binford, Bunn and Kroll1988; Blumenschine, Reference Blumenschine1991; Bunn, Reference Bunn1986, Reference Bunn1982; Bunn et al., Reference Bunn1986; Capaldo, Reference Capaldo1997; Domínguez-Rodrigo, Reference Domínguez-Rodrigo2015, Reference Domínguez-Rodrigo, Hovers and Braun2009a; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo and Cobo-Sánchez2017a, Reference Domínguez-Rodrigo, Barba and Egeland2007; Potts, Reference Potts1988). Taphonomic data also show that durophagous carnivores (namely, hyenas) intervened post-depositionally. Therefore, a dual interactive agency (hominin–hyenid) has been documented. The likely intervention of other less impacting agents remains unreported, with the exception of one carcass at DS which shows felid damage (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Courtenay, Cobo-Sánchez, Baquedano and Mabulla2022a), and two bones at FLK Zinj (Parkinson et al., Reference Parkinson, Plummer and Hartstone-Rose2015). The promising agent-detecting computer vision tools may elucidate the role of other agents in the future (Abellán et al., Reference Abellán, Jiménez-García, Aznarte, Baquedano and Domínguez-Rodrigo2021; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Cifuentes-Alcobendas, Jiménez-García, Abellán, Pizarro-Monzo and Organista2020; Jiménez-García et al., Reference Jiménez-García, Aznarte, Abellán, Baquedano and Domínguez-Rodrigo2020). So far, in the only assemblage where these tools have been applied (DS), the overwhelming majority of damage identified is confidently classified as hyena-made (Cobo-Sánchez et al., Reference Cobo-Sánchez, Pizarro-Monzo, Cifuentes-Alcobendas, Jiménez, Abellán and Courtenay2023). Given that the roles of both agents differ – hominins (accumulators and primary modifiers) and hyenas (secondary modifiers) – the integrity of these assemblages is high. Despite the predominant anthropic agency, it should also be mentioned that it is difficult to assess agency over 100% of the constituent elements of archaeofaunal assemblages. Sites formed in highly productive environments may incorporate naturally deposited background elements, produced by different agencies. These remain very marginal at these sites, because they are either undetected or marginally represented by isolated elements and/or by agent-specific ichnological traces (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Courtenay, Cobo-Sánchez, Baquedano and Mabulla2022a, Reference Domínguez-Rodrigo2015; Parkinson et al., Reference Parkinson, Plummer and Hartstone-Rose2015). Now that carnivore damage and biochemical modification of bones on these assemblages have been properly elucidated (Domínguez-Rodrigo and Barba, Reference Domínguez-Rodrigo and Barba2007b, Reference Domínguez-Rodrigo and Barba2006), overall carnivore damage is documented at very low rates, although their impact in deletion of elements can be substantially higher (Cobo-Sánchez, Reference Cobo-Sánchez2020).Footnote 2 In contrast, hominin-imparted butchery damage is significant and patterned across all anthropogenic sites (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Baquedano, Organista, Cobo-Sánchez, Mabulla and Maskara2021). No other site with the same or similar chronology documented until present shows a frequency of hominin-made modifications on faunal remains as high as these Olduvai assemblages.
Resolution at these assemblages can also be understood by either adopting the single-agency or the multiple-agency versions of this variable. Using the latter, carnivore impact has had a limited effect on the spatial configuration of the sites, and seems to have been limited to the deletion/modification of part of the assemblage, but not to the extensive distortion of its depositional properties (Figure 1.2). Carnivore damage in modern faunal assemblages is spatially reflected in the peripheral dispersal and scattering of remains (Arilla et al., Reference Arilla, Rosell and Blasco2020, Reference Arilla, Rosell and Blasco2019; Arriaza et al., Reference Arriaza, Organista, Yravedra, Santonja, Baquedano and Domínguez-Rodrigo2019; Camarós et al., Reference Camarós, Cueto, Teira, Tapia, Cubas and Blasco2013; Mondini, Reference Mondini1995; Young et al., Reference Young, Márquez-Grant, Stillman, Smith and Korstjens2015). Although this process is intensive in the first few days of deposition (Arriaza et al., Reference Arriaza, Organista, Yravedra, Santonja, Baquedano and Domínguez-Rodrigo2019), it continues through time (Arilla et al., Reference Arilla, Rosell and Blasco2020, Reference Arilla, Rosell and Blasco2019; Arriaza et al., Reference Arriaza, Organista, Yravedra, Santonja, Baquedano and Domínguez-Rodrigo2019; Camarós et al., Reference Camarós, Cueto, Teira, Tapia, Cubas and Blasco2013; Mondini, Reference Mondini1995; Young et al., Reference Young, Márquez-Grant, Stillman, Smith and Korstjens2015). In one of the most extensive and systematic experiments simulating a human camp, where several animal bones were butchered and some even fragmented (including defleshing and demarrowing) and exposed to wild carnivores for three years under controlled circumstances, the assemblage underwent serious modification of its original content and spatial configuration properties (Arilla et al., Reference Arilla, Rosell and Blasco2020). About 80% of the original elements disappeared. Arilla et al. describe it as follows:
A total of 60 stone tools were placed together with 466 skeletal elements. The bone assemblage consisted of large-sized (n = 131) and small-sized skeletal elements (n = 335) belonging to a MNI of 20 (MNI = 10 of size class 4 and MNI = 10 of size class 2). From the total sum, 388 (83.26%) elements disappeared and a set of 78 (16.74%) anatomical elements were recovered. The recovered MNI (n = 14) indicates a bias towards size class 4 (n = 10) in contrast to the recovered MNI of size class 2 (n = 4). The largest part of the recovered items belongs to large-sized animals (n = 57; 43.51% of placed bones from the same size class), specifically mid-shaft fragments (n = 51; 38.93%). Only 6.27% (n = 21) of recovered bones belong to size class 2. Of these recovered bones, 14 (17.95%) exhibited modifications produced by small carnivores, with percentages of 10.26% (n = 8) of size class 2 and 7.69% (n = 6) of size class 4, mostly located on mid-shaft fragments of long bones (n = 5; 6.41% related to recovered), followed by scapula (n = 3; 3.85%) and ulna (n = 2; 2.56%). The rest of modifications were located on proximal epiphysis of long bones (size class 4), humerus, radius, vertebrae and ribs (size class 2) with the same rates.
The percentage of disappearance of bone specimens according to the experimental sector ranges from 67.74% to 88.89%. This is a much higher impact of carnivores on human-simulated bone assemblages than documented in previous experiments (mostly with captive carnivores), probably because those involved large durophagous carnivores (such as hyenas, targeting immediate resource extraction), whereas Arilla et al.’s assemblages were impacted mostly by small wild carnivores (namely, foxes targeting resource caching). These figures are highlighted also by the presence in the experiments of not only fragmented bones, but also complete bones. Only long bone fragmented shaft specimens underwent a lower deletion and showed higher rates of preservation. The overall trend observed with both wild and captive carnivores is for bone clusters to diffuse substantially and for non-long bone shaft portions to disappear from these clusters (Arilla et al., Reference Arilla, Rosell and Blasco2020, Reference Arilla, Rosell and Blasco2019; Camarós et al., Reference Camarós, Cueto, Teira, Tapia, Cubas and Blasco2013).
Kernel maps of the intensity distribution of the archaeofaunal assemblages of the anthropogenic sites of Olduvai Bed I. A, FLK Zinj. B, PTK. C, DS 22a level. D, DS 22b level. The upper level of DS is highlighted to show its similar structure to FLK Zinj and PTK.

Figure 1.2 Long description
The gradient scales present the lowest to highest value represented by darker to lighter shades. Part A. The highest intensity is in the centre-right area. Part B. The highest intensity is located in the center. Part C. The right lower side has a high intensity, with lower intensity in the rest. Part D. The highest intensity is spread in the left and centre-right. The values of the scale are from 20 to 120 in A, 20 to 100 in B, 5 to 15 in C, and 10 to 70 in D.
In contrast with this situation, the Olduvai Bed I anthropogenic sites show intense clustering, with 80%–90% of the bone assemblage and the lithic assemblage being concentrated around small areas (~25–30 m2) (Figure 1.2). The surrounding scattering area may comprise hundreds of square meters. Such an intense clustering of materials over such a small space suggests a limited impact by large and small carnivores on the site’s original configuration. If anything, it could be argued that the density of these clusters must have been higher prior to carnivore intervention, and that carnivore ravaging was not substantial enough to de-cluster the area. This indicates that the original hominin bone-lithic clustering behavior has been preserved at these sites, given that the spatial inhomogeneity of both types of materials is not random and that they show spatial covariance and codependence (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Cobo-Sánchez, Baquedano, Mabulla, Gidna and Diez-Martin2024). The inter-agent resolution is, therefore, high enough to allow the preservation of the original clustering spatial properties of the hominin-created faunal assemblage.
Regarding the single-agent resolution approach, it is difficult to assess hominin impact on their sequential performance of activities at these sites. One striking feature of all of them is that butchery of carcasses, documented in higher numbers than in most modern hunter–gatherer camps (Bartram et al., Reference Bartram and Marean1999, Reference Bartram, Kroll and Bunn1991; Bunn et al., Reference Bunn, Bartram and Kroll1988; Lupo, Reference Lupo2001, Reference Lupo1995; Monahan, Reference Monahan1998; O’Connell et al., Reference O’Connell, Hawkes and Blurton-Jones1992; Yellen, Reference Yellen1991), was systematically performed in very small spaces, configuring the single-cluster nature of most of these sites (DS exceptionally shows three similarly sized clusters) (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo and Cobo-Sánchez2017a; Domínguez-Rodrigo and Cobo-Sánchez, Reference Domínguez-Rodrigo and Cobo-Sánchez2017a; Leakey, Reference Leakey1971; Cobo-Sánchez, Reference Cobo-Sánchez2020). This has major behavioral relevance (see below). We do not know if hominins engaged in other non-butchery activities at these locations, but in other Oldowan sites use-wear analysis has found evidence of diverse activities in addition to butchery, mostly related to plant processing and wood working (Keeley and Toth, Reference Keeley and Toth1981; Lemorini et al., Reference Lemorini, Plummer, Braun, Crittenden, Ditchfield and Bishop2014). What can be inferred from the disposition of the clustered materials at the Olduvai anthropogenic sites is that if such additional activities were performed on the exact location where sequential butchery was carried out, they certainly did not impact in a significant way the original spatial properties of the depositional bone clustering behavior by hominins. If those additional activities were carried out outside the main clustering area, they certainly did not generate any additional secondary clustering with different properties for the discarded lithic implements.
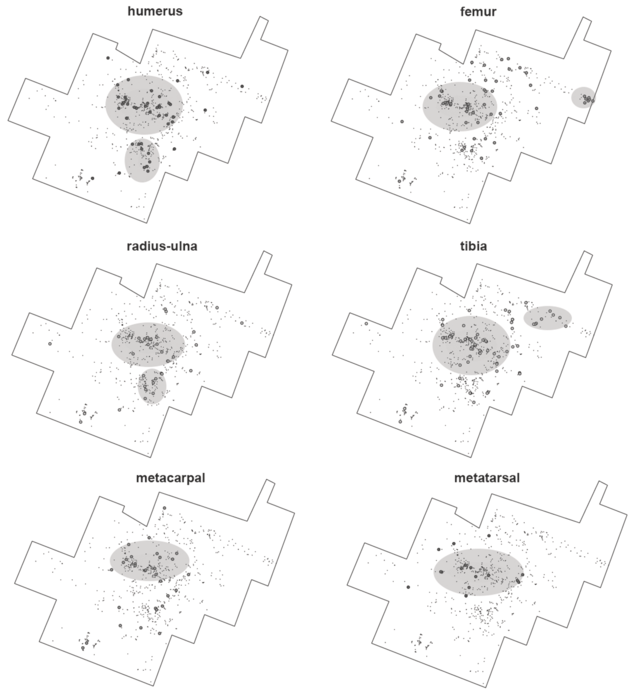
An analysis of the spatial relationships among bone specimens belonging to the same anatomical elements (for example, long bones) shows that the resolution of these assemblages may actually be better than assumed earlier. Experiments with hyenas have shown that these agents may not only generate more long bone shaft fragments than originally present at an assemblage, but they may significantly disperse them over a few meters (Marean and Bertino, Reference Marean and Bertino1994). In modern experiments, the degree of dispersion was significantly impacted by competition; the more hyenas the more dispersed shaft fragments appeared. Given the intense clustering documented at FLK Zinj, and the overlap in tooth-marked and percussion-marked bones, Marean and Bertino (Reference Marean and Bertino1994) concluded that the site “is consistent with the single hyena/low competition context.” This situation is more clearly documented in the new PTK and DS sites (Bed I, Olduvai), when focusing on the predominant medium-sized animals (Figures 1.3 and 1.4). For example, when considering only long bones, at PTK there is a central high-density cluster where most long bone elements are documented. For humeri and radii-ulnae, there is a second smaller cluster, and in both cases their locations coincide, underscoring that both elements underwent the same depositional process and the same limited post-depositional modification (Figure 1.3). Something similar is documented for femora and tibiae; both show second clusters, which although are not overlapping, generally occur in the same area. Metapodials are mostly clustered in the main high-density concentration. This very limited scattering of long bones would be suggestive of limited time since deposition to sedimentation.
Spatial distribution of the long limb bones from medium-sized carcasses at PTK. Notice the clustering of most elements in the same areas.
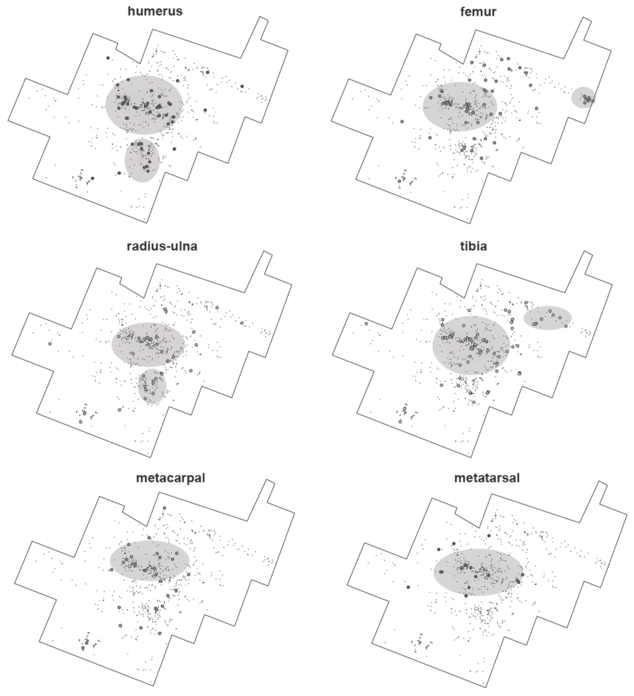
Figure 1.3 Long description
The data points are clustered within each bone outline. For all six, the cluster is dense in the centre. Humerus and radius-ulna also have the cluster closely below the centre, tibia has it on the right, and femur in the extreme right.
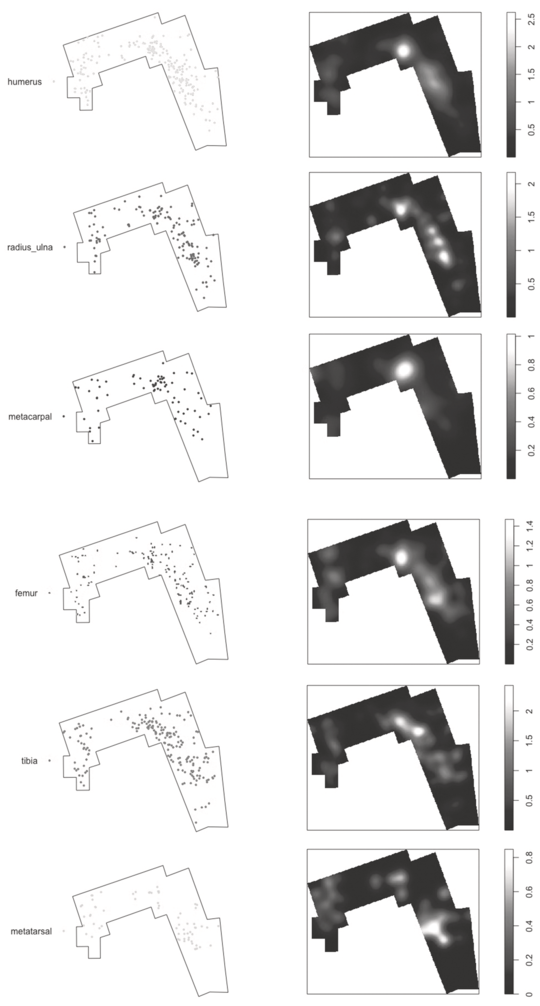
Spatial distribution of the long limb bones from medium-sized carcasses at DS. Notice the clustering of most elements in the same areas.
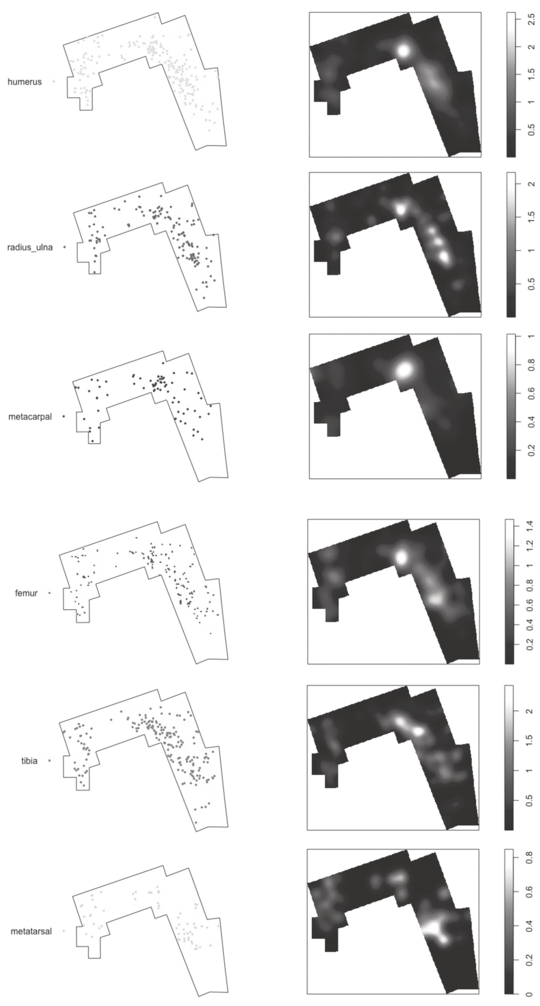
Figure 1.4 Long description
The bones are the humerus, radius or ulna, metacarpal, femur, tibia, and metatarsal. Each pair consists of a scatter plot on the left and a kernel density map on the right. The scatter plots show the spatial distribution of individual bone elements within a defined area, outlined by a polygon. The kernel density maps provide a visual representation of the concentration of these bone elements, with lighter areas indicating higher densities. Each kernel density map is accompanied by a color scale bar on the right, indicating the density values from low to high. The overall pattern suggests a clustering of most bone elements in similar areas within the defined space. The kernel maps present the highest intensity in the center for metacarpal, and the rest have the highest intensity in the center-right. Their gradient scales range from 0.5 to 2.5, 0.5 to 2, 0.2 to 1, 0.2 to 1.4, 0.5 to 2, and 0 to 0.8, respectively.
This intense overlap of all elements on the same areas is also documented at DS (Figure 1.4). If statistical clustering is defined at significantly intense areas, there are three clusters at DS; two with large density (B and C) and a small one with low density (A) (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Cobo-Sánchez, Baquedano, Mabulla, Gidna and Diez-Martin2024), although B and C are fused into a single large cluster (see discussion below). Taking into account that more than 20 carcasses were processed at DS, humeri appear very discreetly located in cluster B–C. Radii-ulnae overlap in this distribution. Metacarpals are even more intensively concentrated in cluster B. Femora and tibia show their more intense concentration in cluster B, and metatarsals appear heavily concentrated in cluster C. In all cases, dispersion outside the B–C cluster is nuanced and only contrasted by cluster A to the right (Figure 1.4). This coherence in the concentration of long bone specimens on exactly the same spots, with limited dispersion, is similar to that documented in the PTK assemblage and is suggestive of limited exposure; more so than at PTK because DS contains a substantially higher presence of epiphyseal ends, which usually disappear very early in the post-depositional stage, because of impact by scavenging carnivores.
Limited dispersal of anatomically associated elements is not the only factor suggesting that the Olduvai Bed I anthropogenic assemblages were subjected to very limited time-averaging. The degrees of bone modification caused by subaerial weathering also indicate that the faunal assemblages at these sites were accumulated over very short time spans.
Another consideration that is of utmost relevance to understand hominin site use is that in all the anthropogenic sites there is an abundance of axial elements. This is especially relevant regarding the presence of rib fragments. All available experiments of fresh bones exposed to the action of ravaging durophagous carnivores show that most axial elements disappear in the first two days of exposure (Arilla et al., Reference Arilla, Rosell and Blasco2020; Arriaza et al., Reference Arriaza, Organista, Yravedra, Santonja, Baquedano and Domínguez-Rodrigo2019; Marean, Reference Marean1991). The presence of substantial axial elements in the Oldowan anthropogenic sites indicates that they were not immediately exposed to carnivores, and that during a substantial amount of time (or time intervals), hominins must have occupied the space, which prevented hyenas and other ravaging carnivores from having prompt access to the hominin-deposited remains. This indicates prolonged use of sites by hominins, instead of sporadic or episodic occupation as suggested by several existing models (see Figures 1.5–1.7).
Aerial aspect of DS (Olduvai Gorge, Bed I) and some images showing the wealth of fossils, with special emphasis on the presence of axial elements in level 22B. Bottom right image shows the clear position of levels 22A and 22B separated by a hiatus.

PTK (Olduvai Gorge, Bed I), with detail of concentration of materials on level 22A.

AGS (Olduvai Gorge, Bed I), with detail of concentration of materials on level 22A.

Assuming similar environmental conditions in temperature–moisture contrasts in the Olduvai Bed I assemblages as documented in Amboseli for modern bones from small and medium-sized carcasses deposited on the same paleosurface, one could theoretically argue that these represent a potentially maximum time span of ~15 (20) years (Behrensmeyer, Reference Behrensmeyer1978). At a geochronological scale, 15–20 years may seem very little time, but for biostratinomic processes, this represents a large time span. If Oldowan sites contained depositional events represented by the preserved fauna prior to sedimentation, it could be argued that if deposition was continuously or intermittently operating during this time, the faunal assemblages should contain bones accordingly displaying all different weathering stages (unless deposited in swamps or wetlands, which would have slowed down weathering). In contrast, at the three anthropogenic assemblages already excavated from Bed I (FLK Zinj, DS, and PTK), this is not documented. We have observed in our actualistic assemblages that axial elements show weathering traces before long bones, whose denser cortical sections protect them better from dehydration and collagen deterioration. For this reason and because most of the Oldowan archaeofaunal assemblages are composed of limb bones, we will use only long bones as a weathering reference (since they may be more conservative in showing weathering stages). At PTK, almost 90% of long bone fragments show stages 0–1 (>75% of them display weathering stage 0). This indicates that the bulk of the assemblage was not subaerially exposed for more than 0–2 years, and most likely, less than a year (Behrensmeyer, Reference Behrensmeyer1978). At FLK Zinj, the sample of diagenetically unaffected long bone specimens that showed good cortical preservation (n = 595), display only weathering stages 0 (91.4%) and 1 (8.6%) (Table 1.3). If including the moderately preserved cortical sample (where about half of the original bone as modified biostratinomically is visible), which shows substantial impact by diagenetic bioerosion and chemical modification (n = 95), the extended sample still displays only stages 0 (79%) and 1 (21%). This also indicates a very short time span in the subaerial exposure of this assemblage. DS coincides in the patterning. Almost 97% of the long bone specimens display weathering stage 0. Stage 1 is documented in only 2.85% of bones. Only 0.45% of limb specimens display stage 2. The DS collection display a very limited amount of subaerial exposure time, probably limited to 0–2 years, and most likely not much more than a year. This would help explain why epiphyseal ends have survived better here than in other assemblages, like PTK. This information, taken together, underscores that the three archaeological sites seem to have been formed equally in very little time. The three sites coincide in displaying weathering stage 0 as the predominant type, and show that the bulk of their archaeofaunal assemblages was not exposed for more than just a few months, and accumulated probably in shorter time spans (Table 1.3). The challenge is, therefore, to explain such intense concentrations of archaeofaunal remains over such limited time periods, which limits by itself the process of accumulation by chance or through redundant processes along extensive number of years. This emphasizes that site formation is, in contrast, caused by specific immediately adaptive processes, and triggered by sites having had a focal function in the socio-reproductive organization and adaptation of hominins.
| Weathering stages | PTK (n = 380) | FLK Zinj (n = 595) | DS (n = 763) |
|---|---|---|---|
| 0 | 75.5 | 91.4 | 96.7 |
| 1 | 13.7 | 8.6 | 2.85 |
| 2 | 7.6 | 0 | 0.45 |
| 3 | 3.1 | 0 | 0 |
| 4 | 0 | 0 | 0 |
| 5 | 0 | 0 | 0 |
Early Humans as Eusocial Primates?
Eusociality is an unusual feature in the organic life of the planet; however, a diverse group of insects and mammals has adopted it. Originally defined as a hypersocial behavior resulting from intense group cooperation based on labor division (Crespi and Yanega, Reference Crespi and Yanega1995), it was subsequently expanded to behaviors that showed the combination of the following elements: Distribution of labor, group constitution based on overlapping generations, and collective breeding with prominence of alloparenting (Nowak et al., Reference Nowak, Tarnita and Wilson2010; Wilson, Reference Wilson2012; Wilson and Hölldobler, Reference Wilson and Hölldobler2005). Originally studied among the Hymenoptera (antes, bees, and wasps) and later on also in Isoptera (termites), it was also discovered in some mammals, like the naked mole rat and its hairy version (Damaraland mole rat). Some authors have posited that humans are an example of eusociality (Nowak et al., Reference Nowak, Tarnita and Wilson2010; Wilson, Reference Wilson2012), to the controversial chagrin of a plethora of evolutionary biologists (Abbot et al., Reference Abbot, Abe, Alcock, Alizon, Alpedrinha and Andersson2011; Boomsma et al., Reference Boomsma, Beekman, Cornwallis, Griffin, Holman and Hughes2011; Bourke, Reference Bourke2011; Marshall, Reference Marshall2019, Reference Marshall2015). These base their interpretation of any eusocial behavior as resulting from Hamilton’s inclusive fitness theory (Gardner and West, Reference Gardner and West2014; Hamilton, Reference Hamilton2017), although the limits of inclusive fitness have also been discussed (Allen et al., Reference Allen, Nowak and Wilson2013). Hamilton’s rule is strongly dependent on the “degree of relatedness,” commonly understood as “the probability that an allele is identical in the benefactor and the recipient by common descent” (Gintis, Reference Gintis2012). It has been argued that an in-depth examination of Hamilton’s rule does not actually explain eusociality without controversy (Bowles and Gintis, Reference Bowles and Gintis2013, Reference Bowles and Gintis2011), and it has been shown that “the cooperation of genes in metazoan cells and in social groups is therefore effected by selection at a higher level than the gene itself” (Gintis, Reference Gintis2012). The controversy arises from dual positioning in understanding selection. One, majoritarian, argues that selection operates only at the individual level. The other defends that there are several levels of selection (multilevel selection, MLS), and one of them is group selection. The discussion has even reached philosophical realms. In one of the most comprehensive approaches based on the application of formal logic to the arguments and the dissection of the mathematical foundations of each view, it has been argued that not only MLS exists, but that it is an understandable extension of individual selection (Okasha, Reference Okasha2006).
Irrespective of this debate, one crucial aspect of eusocial species is that only one individual or a small fraction of individuals reproduces to the expense of the rest of the group. This was actually one of the defining features of eusociality. A strict application of the term results in excluding humans as a eusocial taxon (Gintis, Reference Gintis2012). Although alloparenting behaviors and altruistic behaviors are abundant in the human species, they do not result in a significant fraction of the population giving up reproduction for the sake of just a few individuals, as is the rule in all eusocial species. This is a clear indication that kin selection operates in a multipolar way in our species, and that individual selection is an essential element of our socio-reproductive systems. This is not incompatible with MLS and, as a matter of fact, inclusive fitness can be combined with inter-group competition and selection when explaining human evolutionary biology (Bowles and Gintis, Reference Bowles and Gintis2013). Nevertheless, if we exclude this defining factor and focus on the other three listed above (collective breeding, overlapping intensive cooperative generations, and labor division), modern humans formally could qualify as a eusocial species (Nowak and Highfield, Reference Nowak and Highfield2011; Wilson, Reference Wilson2012). The combination of these features is novel in primate evolution. Although some primates adopt alloparenting (e.g., callitrichids) or marginal alloparenting (i.e., helping in occasional carrying of offspring or protection thereof) (Hrdy, Reference Hrdy2009; Kishimoto et al., Reference Kishimoto, Ando, Tatara, Yamada, Konishi and Kimura2014), none of the extant nonhuman primate taxa adopt a social structure in which group breeding involves continuous alloparenting, including feeding of young and older individuals, regardless of age. This is the case of modern humans. Foraging groups are organized around the collective obtainment of food, which is shared among all individuals, ultimately benefiting the subadult portion of the group. For this to occur, humans engage in the most intense cooperative system that any primate has developed. Here, cooperation is not limited to group defense, as in other primate species, but it permeates every single aspect of the behavior and adaptation of humans as an organic species. High degree of cooperation involves structured labor division, as in all eusocial species. Human cohesive groups, in contrast with other primate cooperative groups (Strier, Reference Strier2015), involve more than one family (i.e., several reproductive units) and, most commonly, these are unrelated. Humans are also the only primates that maintain cooperative and social networks with individuals residing in other groups, cooperating, relating, and intentionally reproducing with them (Hill et al., Reference Hill, Walker, Bozicević, Eder, Headland and Hewlett2011, Reference Hill, Barton and Hurtado2009). This is not observed in any other eusocial species either. In this sense, humans display unique adaptive features; however, it is the other ones shared with eusocial species that are of utmost interest, because they lead to common evolutionary biological strategies.
The key element in eusocial animals is what Wilson (Reference Wilson2012) defined as the “nest.” The nest is a referential locus in space where the community lives, reproduces, carries out variable degrees of feeding, takes care of the offspring, and organizes labor division. One structural feature of the nesting behavior is that food is transported into it. Nesting is not exclusive to eusocial species. We find it linked to other non-eusocial (mesosocial, genosocial?) animals that practice joint breeding and some degree of alloparenting. This may be the case of just the reproductive couple (male–female), as in most birds, or the reproductive couple plus extended kin, as in some mammal carnivores like wolves or brown hyenas, (David Mech, Reference David Mech2003; David Mech and Boitani, Reference David Mech and Boitani2010; Mills, Reference Mills2003), or association of related and unrelated individuals in the form of extensive clans, as in spotted hyenas (Kruuk and Kruuk, Reference Kruuk and Kruuk1972). In the latter case, the dynamics of relationship are competitive and within-clan females provision their offspring mostly (Holekamp et al., Reference Holekamp, Cooper, Katona, Berry, Frank and Smale1997; Holekamp and Smale, Reference Holekamp and Smale2010; Smith et al., Reference Smith, Swanson, Reed and Holekamp2012, Reference Smith, Van Horn, Powning, Cole, Graham and Memenis2010); however, in the other cases, provisioning is cooperative among all individuals involved, because of inclusive fitness. The breeding purpose of dens is so universal that even solitary predators (like leopards) use them to hide their cubs and bring food to them.
The only solid information that archaeologists have collected from early sites (Isaac’s type C site: Stone tools associated with faunal remains from different individuals, most commonly from diverse taxa) is that they acted as referential spots, where hominins repeatedly transported and processed food; namely, animal carcasses (Isaac and Isaac, Reference Isaac and Isaac1989). The “central-provisioning“ behavior is at the core of what has been universally identified as human behavior in modern foragers (Apicella et al., Reference Apicella, Marlowe, Fowler and Christakis2012; Marlowe, Reference Marlowe2010, Reference Marlowe, Hihmann, Robbins and Boesch2006; Wood and Marlowe, Reference Wood and Marlowe2013; Wood et al., Reference Wood, Harris, Raichlen, Pontzer, Sayre and Sancilio2021). Modern foragers’ home bases or camps are the loci where the group lives, reproduces, brings food, and consumes it, where the offspring is taken care of and where labor is organized; all these are the essential features of eusocial organisms as was mentioned above. Regardless of whether modern humans are seen as eusocial primates or not, their converging behavior with eusocial taxa should be applied to our understanding of early archaeological sites. If “central-provisioning” is essential to humanness, its roots may extend to the origin of focalization of food processing and consumption. Extensive taphonomic literature exists that emphasizes that resolving the amount of food that was transported to early sites is crucial for testing the diverse existing models. A summary of the taphonomic evidence is outside the scope of the present work. Suffice it to say that there has been increasingly a wealth of data showing that hominins gained early access to small and medium-sized carcasses and targeted bulk flesh exploitation (Bunn and Gurtov, Reference Bunn and Gurtov2014; Domínguez-Rodrigo, Reference Domínguez-Rodrigo2015, Reference Domínguez-Rodrigo2012; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Baquedano, Organista, Cobo-Sánchez, Mabulla and Maskara2021, Reference Domínguez-Rodrigo, Barba and Egeland2007; Ferraro, Reference Ferraro2007; Ferraro et al., Reference Ferraro, Plummer, Pobiner, Oliver, Bishop and Braun2013; Parkinson, Reference Parkinson2018, Reference Parkinson2013; Parkinson et al., Reference Parkinson, Plummer, Oliver and Bishop2022; Pickering, Reference Pickering2013; Pickering et al., Reference Pickering, Domínguez-Rodrigo, Egeland and Brain2004, n.d.), although some researchers still defend alternative scenarios (Barr et al., Reference Barr, Pobiner, Rowan, Du and Faith2022; Pante et al., Reference Pante, Blumenschine, Capaldo and Scott2012). So, why were hominins bringing substantial animal food resources repeatedly to the same loci (to a larger extend than is documented among modern African foragers) over limited periods of time, if their purpose was not their collective consumption by the group and the provisioning of the offspring?
Isaac’s original idea of explaining “humanness” from everything that made us different from other primates set a paradigm that was based on the idea of human uniqueness. This, fused with the idea of progressive (i.e., gradual) evolution, opened a big space for imagination and for possible models without any relevant modern counterpart. Otherwise said, as long as models could explain part of the material record and were logically feasible, they could be used as interpretive proxies for early sites if they were distinct enough from modern human foraging behavior. This is how interpretations like the “stone-cache” model (Potts, Reference Potts1988) or the “preferred site” model (Shick, Reference Shick1987) emerged. However, interpretations of models that emphasize human unique behavioral features, or strategies without modern reference, isolate humans from the organic mainstream and the selective criteria that shape behavior in its most basic form. These models are also bound to be non-testable for lack of an observable reference.
The redundancy in anatomical and behavioral convergence of the organic world repeatedly expresses the constrictive patterns of natural selection; this is very well documented among mammals (Clutton-Brock, Reference Clutton-Brock2016; Prothero, Reference Prothero2016, Reference Prothero2006; Prothero et al., Reference Prothero and Schoch2002), and among these, in primates (Clutton-Brock, Reference Clutton-Brock1974; Clutton-Brock and Harvey, Reference Clutton-Brock and Harvey1977, Reference Clutton-Brock, Harvey, Bateson and Hinde1976; Clutton-Brock and Janson, Reference Clutton-Brock and Janson2012; Strier, Reference Strier2015). Organisms must follow deep ecological principles that shape adaptation and affect all biological entities without exception. This is why evolutionary biology and comparative behavioral ecology exist; they seek the mechanisms that channel and prompt adaptive solutions. Organisms that adapt to similar environments must face these constrictions, and frequently adopt similar solutions fitted to their specific phenotypes (Bekoff and Byers, Reference Bekoff, Byers, Hecht, Wallace and Prance1986; Kundu, Reference Kundu2021; Losos, Reference Losos2011; Plutynski, Reference Plutynski2021; Sober, Reference Sober1994; Wroe and Milne, Reference Wroe and Milne2007). If we envision humans as just another organism subjected to the same rigors of natural selection as other organisms, then our interpretation of their behavior is no longer based on uniqueness, but on commonness. Comparative behavioral ecology shows that most organisms that create nests establish high levels of cooperation and vice versa. It also shows that organisms that use focal places to transport food do so because breeding and feeding offspring (and, when applicable, others) is the trigger. If we compare humans to those organisms, then we should view early archeological sites as the expression of “nesting” behavior in hominins, and we should expect high levels of cooperation. As a matter of fact, all available data indicate significantly higher levels of cooperation among the hominins responsible for the early archaeological record than documented among other nonhuman extant primate taxa (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Barba and Egeland2007; Isaac, Reference Isaac1978; Potts, Reference Potts1988; Stanish, Reference Stanish2017; Sterelny, Reference Sterelny2021). Early access to medium-sized carcasses would have implied collective participation in their acquisition, preprocessing, transport, and, probably, consumption at sites. The large amount of raw materials transported to sites, especially those formed over limited amounts of time (see above) also supports this (Potts, Reference Potts1988).
Following similar behavioral ecological principles, the existence of a “nest” would have implied also a universal gesture in many unrelated species with central provisioning: Postponement of food consumption upon attainment, and transport to the locus of collective consumption and offspring feeding. Under this view, early archeological sites would have been provisioning loci and breeding grounds. This contradicts previous ideas of sites having had a single-individual or a partial-group refuge function, and interpretations thereof assuming an expedient use of the locations for carcass processing only (Blumenschine, Reference Blumenschine1991; Blumenschine et al., Reference Blumenschine, Cavallo and Capaldo1994; Potts, Reference Potts1988). If hominins were “nesting” for other purposes, they would have been the only organism doing so, and major principles of behavioral ecology would not apply to them. The focalization of activities by early humans, involving stone tool making and use, and animal food transport and consumption, is so similar to the central-provisioning behaviors of eusocial taxa and several other mammal carnivorans that one must assume that the adaptive triggers and associated selective factors were similar if not the same. From a behavioral ecological perspective, it could be argued that such sites were already structural versions of modern foragers’ home bases where food provisioning and collective consumption took place. It is not known if other social activities usually performed at “nests” (e.g., sleeping, labor division) were also performed at sites. The limited evidence of use–wear analyses on lithic artifacts repeatedly show that tools at these sites were used for purposes other than carcass butchery, and plant and wood processing seem to be represented in these techno-assemblages (Keeley and Toth, Reference Keeley and Toth1981; Lemorini et al., Reference Lemorini, Plummer, Braun, Crittenden, Ditchfield and Bishop2014). Therefore, hominins seem to have engaged in several types of activities at those sites.
Current evidence indicates an overlap of the earliest traces of encephalization and the occurrence of the earliest archaeological record. We do not know if there is a link between both phenomena. This biological innovation, with important repercussions for the evolution of the human brain, may have started the process of altricial offspring typical of modern humans. This would have required a safe breeding ground, and sites may have provided this haven. Behavioral ecology indicates that the main function of “nests” is to keep the young offspring and provision them.
Conservative estimates of the number of hominins involved in site formation and use, based on the areas over which they generated debris and with modern foragers as a reference (Yellen, Reference Yellen1977), indicate that early hominins using sites were doing so in groups of a size substantially bigger than a single reproductive unit and their immediate kin (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Cobo-Sánchez, Aramendi and Gidna2019b). Eusocial species displaying aggregation of numerous individuals in a cooperative system are based on kin. Meso-social mammal carnivorans displaying similar behaviors also are based on a kin-centric structure. One could argue that such a structure may also have been the basis for early human sociality; probably a large portion of those hominin groups were genetically related.
The most visible behavior of hominins at early sites is their dietary change involving consumption of animal food. For this reason, in order to obtain these resources, essential for the central-provisioning system, they would have been required to overcome the same selective rigors that limit other animals whose diet relies on the same resources. This is why, we argue, primates are only a limited proxy for early hominins. We should seek a better proxy in those other animals; namely, mammal carnivorans adapted to the same environments. In this case, the analogy prevails over the homology.
Early Humans as Carnivorans
In a landmark paper published half a century ago, the idea with which we finished the previous paragraph was best defended; namely, that African savanna carnivores (and not primates) provide a better model for the behavior of early humans entering the carnivoran guild (Schaller and Lowther, Reference Schaller and Lowther1969). These authors specifically remarked that the aspects on which carnivoran behavior may be a better referent for modeling early hominin adaptation than primate behavior are: Social organization, cooperative hunting, food sharing, and the division of labor. Most of these topics have been abandoned in the past few decades by paleoanthropologists, not because they were superseded by better conceptual models, but because academic agendas switched. Social organization and food sharing are intimately related. For this reason, we will pay closer attention to these two elements from a carnivoran point of view. Social organization is more ecologically than phylogenetically determined. Schaller and Lowther (Reference Schaller and Lowther1969) noticed that phylogenetically closer species to humans, like gorillas and chimpanzees, exhibited more distant social structures than some carnivorans. They also remarked that even phylogenetically close taxa – like Papio ursinus or Papio hamadryas – or even the same species placed in three different environments displayed very different social systems. This plasticity and variability have an obvious ecological reason. Schaller and Lowther (Reference Schaller and Lowther1969) argued that primates have drastically different diets from humans, and restrict themselves to small home ranges. Humans, like most similarly sized carnivorans, enjoy a diet with regular inclusion of animal resources, and have much bigger home ranges. Carnivorans share these features and the commonality of their structural adaptation should provide a behavioral ecological basis to understand hominin adaptations once they enter the carnivoran guild.
Schaller and Lowther (Reference Schaller and Lowther1969) showed that wild dogs in the Serengeti are grouped in packs of an average of 11 individuals (range 2–32), spotted hyenas were grouped in clans of 10–100 individuals, and lion prides usually contained an average of 25 individuals, half of which were subadult individuals. These figures are not too far off the average group composition of modern human foragers in Africa (Jones, Reference Jones2016; Marlowe, Reference Marlowe2010; Turnbull, Reference Turnbull2015; Woodburn and British Museum, Reference Woodburn and Museum1970). Despite the basic structure of the group, composed of reproductive units and their extended kin, the composition of the group among wild dogs, spotted hyenas, and lions is flexible, with individuals leaving and returning (Creel and Creel, Reference Creel and Creel2002; Kruuk and Kruuk, Reference Kruuk and Kruuk1972; Schaller, Reference Schaller and Keane1972). Human foragers also maintain a similar flexible group structure (Apicella et al., Reference Apicella, Marlowe, Fowler and Christakis2012; Hill et al., Reference Hill, Walker, Bozicević, Eder, Headland and Hewlett2011; Marlowe, Reference Marlowe2010; Wood et al., Reference Wood, Harris, Raichlen, Pontzer, Sayre and Sancilio2021). An initial attempt to estimate group size among early Pleistocene hominins based on compositional and spatial data indicates that a minimum range of 18–28 individuals may have generated the FLK Zinj food debris, using modern forager data as a reference (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Cobo-Sánchez, Aramendi and Gidna2019b). Estimates for the penecontemporaneous site of DS yield similar estimates (Cobo-Sánchez, Reference Cobo-Sánchez2020). These figures are probably an underestimation, given the amount of food represented at these two sites, but they show similar group density to that reported among the main large mammal carnivorans in the same type of biome.
Schaller and Lowther (Reference Schaller and Lowther1969) selected the following forces as the main constrictive selective agents determining social structure in social predators:
a. Group predation is more effective than solitary predation, especially in environments where ambushing is not successful.
b. Group formation can target larger prey more efficiently.
c. Groups are able to use and defend resources more efficiently than solitary predators from scavengers.
d. Division of labor is possible within groups. For carnivorans this means that some individuals will guard and protect youngsters in dens while other individuals forage and hunt.
e. Group size determines to some extent the position of the species within the predatory guild. Larger groups tend to be associated with dominant taxa.
An additional element is that group size determines prey size. For the collective and cooperative effort that requires hunting game, the amount of resources obtained must satisfy the dietary demands of the group. This is why within the same species, the larger the group the bigger the prey. For example, mostly lions residing in prides >30 individuals target elephants and megafauna more successfully than others who live in smaller prides (Andreassen et al., Reference Andreassen, Neo-Mahupeleng, Flagstad, Wegge, Skarpe, du Toit and Moe2014; John Power and Shem Compion, Reference John Power and Shem Compion2009; Joubert, Reference Joubert2006).
All this would suggest that hominins would have been more adaptive if they had adopted a group strategy. If, as taphonomic research suggests (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Baquedano, Organista, Cobo-Sánchez, Mabulla and Maskara2021, Reference Domínguez-Rodrigo, Barba and Egeland2007), they were successfully acquiring medium-size game through hunting, this would only have been feasible if they were cooperating as a group. This also implies that young offspring would have had to stay at “nests,” which would have required some alloparenting and guarding. If group size determines the position in the carnivoran guild, this could also help interpret why hominin groups became increasingly bigger longitudinally over time. If site size is a proxy for group size, the moderate size of groups during the early Oldowan gave rise to substantially larger groups after the Acheulian at 1.8 Ma. This is suggested by the much bigger sizes of Acheulian sites as documented in the Olduvai Gorge archaeological record. Sites like FLK West (Diez-Martín et al., Reference Diez-Martín, Sánchez Yustos, Uribelarrea, Baquedano, Mark and Mabulla2015), SHK (Diez-Martín et al., Reference Diez-Martín, Sánchez-Yustos, Uribelarrea, Domínguez-Rodrigo, Fraile-Márquez and Obregón2014; Manuel Domínguez-Rodrigo et al., Reference Diez-Martín, Sánchez Yustos, Uribelarrea, Baquedano, Mark and Mabulla2014), TK (Panera et al., Reference Panera Gallego, Rubio Jara, Domínguez-Rodrigo, Yravedra Saínz de los Terreros and Méndez Quintas2019; Rubio-Jara et al., Reference Rubio-Jara, Panera, Santonja, Pérez-González, Yravedra and Domínguez-Rodrigo2017; Santonja et al., Reference Santonja, Panera, Rubio-Jara, Pérez-González, Uribelarrea and Domínguez-Rodrigo2014) or BK (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Bunn, Mabulla, Baquedano, Uribelarrea and Pérez-González2014a; Organista et al., Reference Organista, Domínguez-Rodrigo, Yravedra, Uribelarrea, Arriaza and Ortega2017, Reference Organista, Domínguez-Rodrigo, Egeland, Uribelarrea, Mabulla and Baquedano2016) are several times bigger than previous Oldowan sites. Only a small fraction of them have been excavated. It comes, thus, as no surprise that in such sites exploitation of megafauna is systematically documented, which would be expected if hominin group size was larger than during the Oldowan. Under this behavioral ecological referent, hominins would have increased their capacity of acquiring faunal resources through time, contra theoretical interpretations based on comparisons of a mix of taphonomically heterogenous sites (Barr et al., Reference Barr, Pobiner, Rowan, Du and Faith2022).
Although Schaller and Lowther’s paper has been used as evidence for the role of scavenging in human evolution, these authors clearly claim that “not a single large mammal has become solely adapted to the role of scavenger. One probable reason is that not enough meat is available to support a population of mammalian scavengers, mainly because each predator consumes most of its kill; there is no ecological room for a total scavenger” (Schaller and Lowther, Reference Schaller and Lowther1969). All current large carnivorans residing in the Serengeti–Nogongoro ecosystem base their survival on a hunting strategy. Hyenas consume more than 70% of their food through hunting (Kruuk and Kruuk, Reference Kruuk and Kruuk1972). Lions consume more than 90% of their food through hunting (Schaller, Reference Schaller and Keane1972). Cheetah consume all their food through hunting (Caro, Reference Caro1994). Wild dogs also rarely scavenge (Creel and Creel, Reference Creel and Creel2002). These percentages depend on ecosystems and degree of inter-carnivore competition, but hunting remains the bulk strategy of these predators (Creel and Creel, Reference Creel and Creel1996), including large scavengers like hyenas. This has major consequences for our understanding of early humans if using this behavioral ecological framework. If hominins entered the carnivoran realm, they also must have done so by essentially being predators. Some archaeologists will object to this because this interpretation leaves limited room for evolutionary trajectories (i.e., evolutionary narratives that anchor processes in a progressive evolutionary scale). Under this perspective, it would be more acceptable to conceptualize hunting as following a series of prior footsteps guided by opportunistic/scavenging strategies (Pobiner, Reference Pobiner2020; Thompson et al., Reference Thompson, Carvalho, Marean and Alemseged2019). However, a clear indicator that scavenging can only be a complementary and minor contributing strategy to hominin adaptive patterns comes not only from this comparative behavioral ecological approach (comparing hominins to carnivorans), but also from the fact that virtually all the consumption of meat by primates is carried out through hunting. Chimpanzees, our closest relatives, avoid scavenging and actively acquire their faunal food through hunting (Boesch, Reference Boesch2005, Reference Boesch2002; Boesch and Boesch, Reference Boesch and Boesch1989; Domínguez-Rodrigo and Gilby et al., Reference Gilby, Eberly, Pintea and Pusey2006; Gilby and Wrangham, Reference Gilby and Wrangham2007; Muller et al., Reference Muller, Wrangham and Pilbeam2017; Nakamura and Itoh, Reference Nakamura and Itoh2008; Newton-Fisher et al., Reference Newton-Fisher, Notman and Reynolds2003; Pickering, Reference Pickering, Domínguez-Rodrigo, Egeland and Brainn.d.; Stanford et al., Reference Stanford1998; Stanford and Bunn, Reference Stanford and Bunn2001; Watts and Mitani, Reference Watts and Mitani2002; Wilson, Reference Wilson2021). Bonobos also obtain all their faunal resources through hunting (Hohmann and Fruth, Reference Hohmann and Fruth2008; Ihobe, Reference Ihobe1992; Samuni et al., Reference Samuni, Wegdell and Surbeck2020; Surbeck and Hohmann, Reference Surbeck and Hohmann2008). The meat that baboons consume is also obtained through hunting (Butynski, Reference Butynski1982; Harding, Reference Harding and Tuttle1975; Morris and Goodall, Reference Morris and Goodall1977; Rhine et al., Reference Rhine, Norton, Wynn, Wynn and Rhine1986; Strum, Reference Strum, Harding and Teleki1981; Strum and Mitchell, Reference Strum, Mitchell and Kinzey1987). Both mammal carnivorans and primates indicate that when hominins started incorporating meat in their diets, they must have done so essentially through hunting. Scavenging probably was (as still is today among all carnivores, including humans) a secondary strategy that never accounted for the bulk of their faunal consumption. In words of Schaller and Lowther (Reference Schaller and Lowther1969: 326): “A carnivorous hominid would by necessity have had to be both a scavenger and a hunter, using every available method to obtain meat, as the predators do today.” This leaves the question of whether there was a scavenging phase in human evolution in the air, since hominins may have adopted chimpanzees’, bonobos’, and baboons’ strategies for meat acquisition, instead of having been the odd primates purportedly depending mostly or solely on scavenging for acquiring animal protein.
Another relevant issue is that social carnivores targeting large game are territorial. This may be all year round or seasonally, especially during the earliest stages of breeding. Kruuk (Reference Kruuk and Kruuk1972) documented that clans of spotted hyenas in Ngorongoro and Serengeti fed within their own territorial ranges. Lion prides also use permanent territorial ranges, like wolves. Hominins, behaving like carnivorans exploiting large game, may also have tenured land in a similar fashion. Given the clearly defined patterning in the composition, taphonomic properties, and spatial distribution of debris in the anthropogenic Bed I assemblages (see below), it could be argued if similar groups of hominins generated them. This could also explain why in lower Bed II, the HWK complex has such an intense deposition of archaeological materials over the landscape, probably resulting from a very intense use of the space (Sistiaga et al., Reference Sistiaga, Husain, Uribelarrea, Martín-Perea, Ferland and Freeman2020), as would be expected from redundant use of tenured land systems.
From the perspective of a social carnivoran, cooperation in carcass acquisition immediately leads to food sharing, even if this also involves competition and aggression (Schaller and Lowther, Reference Schaller and Lowther1969). Hominins, as carnivorans, would probably also have cooperated in carcass acquisition and sharing (see below discussion of the archaeological evidence thereof). Related to this, most social carnivores also bring food back to the den. Canids do so without physical evidence thereof by regurgitating meat. Felids bring carcasses back to their cubs; except lions, which move their cubs to the kills. Hominin social structure (extended groups) may also have been based on “den” provisioning (as pointed out in the previous section) if they behaved like most social carnivorans. It has been argued that spotted hyenas are of interest for the interpretation of cooperation and social organization in hominins, because they are some of the most complex mammal carnivorans, since they live in larger groups (up to a hundred individuals) and most of the individuals are unrelated (Smith et al., Reference Smith, Swanson, Reed and Holekamp2012). A multivariate analysis testing the factors that coevolve with cooperation among mammalian carnivorans shows that there are statistically meaningful factors that are related with increased levels of cooperation. These are: Reduced sexual dimorphism, increase in relative brain size, foraging adaptations (hunting of large game in open habitats), and life history elements (delayed weaning and growth). This analysis uncovered a positive relationship between hunting large game in open environments and the formation of social groups in predators. The study also found that carnivores using cursorial hunting or large game were most likely to cooperate. If early hominins followed this trend, it would be expected for them to exhibit larger body sizes, bigger brains, decreasing sexual dimorphism compared to previous hominins, and slower growth rates; all of this sees its onset at the time in which the archaeological record shows intense consumption of large animals by early humans, especially with the emergence of Homo erectus two million years ago (Aiello, Reference Aiello2007; Aiello and Wheeler, Reference Aiello and Wheeler1995; Antón, Reference Antón2012; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Baquedano, Organista, Cobo-Sánchez, Mabulla and Maskara2021; Faurby et al., Reference Faurby, Silvestro, Werdelin and Antonelli2020; Herries et al., Reference Herries, Martin, Leece, Adams, Boschian and Joannes-Boyau2020; Pontzer, Reference Pontzer2012; Roebroeks, Reference Roebroeks2007). The more intense cooperation interpreted from the early archaeological record coincides in time with the earliest evidence of the adoption of endurance running by hominins (Bramble and Lieberman, Reference Bramble and Lieberman2004; Lieberman et al., Reference Lieberman, Bramble, Raichlen, Shea, Grine, Fleagle and Leakey2009, Reference Lieberman, Bramble, Raichlen and Shea2007), which is another coincidental feature in the emergence of Homo erectus that signals to early humans closely following the constrictive precepts documented through the behavioral ecology of large mammal carnivorans.
The Structure of the Bed I Oldowan Sites
There is another relevant way in which large mammal carnivoran behavioral ecology can inform us about the social organization of early humans. Social carnivorans cooperate when hunting (pack predation), and they collectively consume their prey. Pack hunting is mostly associated with cooperative breeding and is mostly documented in tropical savanna biomes. Once an animal is killed, in social carnivorans organized around extended kinship, like lions or wild dogs (also wolves or coyotes in higher latitudes), the prey is collectively consumed on the spot. The group, acting as a cohesive pack, only marginally disaggregates the prey when inter-individual competition is too strong, to consume it just in the nearby periphery of the kill. Nevertheless, most of the time, carcasses are consumed at the kill. In non-related social carnivores, like spotted hyenas, the initial consumption of carcasses also occurs at the kill, but the strong inter-individual competition generates centrifuge forces that end up dismembering carcass parts, and carrying them away from the kill to much longer distances than in kin-related social predators. This is also documented at the collective dens. Spotted hyena dens created in multiple burrows offer multiple bone clusters, generated by separate individuals bringing in bones and accumulating them in their burrows. This is also a typical modern human forager behavior. Most modern human hunter–gatherer camps show multiple clusters, each belonging to separate households, as a reflection of the individual use of the space within the camp by each reproductive unit (Bartram et al., Reference Bartram, Kroll and Bunn1991; Domínguez-Rodrigo and Cobo-Sánchez, Reference Domínguez-Rodrigo and Cobo-Sánchez2017b; Kroll and Price, Reference Kroll and Price1991; Yellen, Reference Yellen1977). In contrast with this, the taphonomically supported anthropogenic sites of Olduvai Bed I (FLK Zinj, DS, PTK, AGS) all show an enigmatic patterning in the following elements: Spatial distribution of archaeofaunal remains, site size, faunal composition, and taphonomic properties (i.e., extent of carnivore damage, skeletal part representation, and butchery patterns).
Spatial distribution. The Olduvai Bed I anthropogenic sites display a higher density of remains than many modern forager home bases, but instead of showing multiple clusters and scatters, most of the remains are concentrated in a single ~20–25 m2 cluster (Domínguez-Rodrigo and Cobo-Sánchez, Reference Domínguez-Rodrigo and Cobo-Sánchez2017a) (Figure 1.2). This is also the average size of the area where the bulk of the carcass parts abandoned at the kill by gregarious carnivores remains after consumption. Carnivorans do not generate accumulations of carcass remains on the same spots because each kill varies according to location, and killing only rarely happens around the same place, unless in arenas of serial predation (e.g., around ponds). Even in the latter case, it is rare to find concentrations of more than one individual carcass on exactly the same spot (Domínguez-Rodrigo, Reference Domínguez-Rodrigo1993). If there were only one archaeological example, it could be argued that such clustering occurred around an exceptional habitat feature (e.g., a tree); however, the pattern is the same in all the archaeological levels documented so far: FLK Zinj (level 22a), PTK (levels 22a and 22b), DS (levels 22a), AGS (levels 22a,22b, Chapati). Only the 22b level of DS shows three intense clusters adjacent to each other. Therefore, the patterning has been documented so far in as many as seven different archaeological levels. In addition, the paleobotanical information available shows that such ecological constrictions did not exist, since the loci where these sites occurred were densely vegetated with abundant trees (Arráiz et al., Reference Arráiz, Barboni, Ashley, Mabulla, Baquedano and Domínguez-Rodrigo2017; Ashley et al., Reference Ashley, Barboni, Domínguez-Rodrigo, Bunn, Mabulla and Diez-Martin2010b). Therefore, by exclusion, one must conclude that such spatial patterning does not have an ecological, but a behavioral causation. Hominins transported their animal carcass resources to the same locus over and over again, without any apparent ecological constriction, and repeatedly processed them on the same area. This would be suggestive of use of that particular spot on the same occupational episode. Carcasses that reached the spot in a fairly complete state were not scattered, but processed, consumed, and accumulated on the same spot repeatedly. This indicates, using mammal carnivoran analogs, that the processing and consumption process took place (probably collectively) in a similar fashion as carcass consumption by gregarious kin-related carnivorans like lions. This has enormous consequences for our interpretation of the socio-reproductive structure of early hominin groups. If carcass consumption was communal, early humans behaved like some large carnivorans, and consumed their game without the typical secondary dismembering occurring in modern human forager home bases, which is based on distribution among different reproductive units (i.e., households). In this sense, early humans acted as a more cohesive single large group, probably because the structure of the group involved extensive kinship of at least a good part thereof. From this perspective, it could be argued that early human social organization was much more similar to modern lions than it was to modern humans. This would imply that although the increase of cooperation would have resulted in a decrease of sexual dimorphism, compared to previous and other sympatric hominins, body size differences would still be prominent, as documented among these social carnivorans (either lions or hyenas). This is evinced in the anatomy of early Homo erectus, which still shows a significant degree of sexual dimorphism, although more reduced than australopithecines (Antón, Reference Antón2012; Callan, Reference Callan2018; Plavcan, Reference Plavcan2012; Spoor et al., Reference Spoor, Leakey, Gathogo, Brown, Antón and McDougall2007). The enigma still remains as to why hominins within all these sites were continuously processing carcasses locally on the same spot (Figure 1.2).
Site size and faunal composition. At the moment of writing this, only three Bed I anthropogenic assemblages are fully taphonomically analyzed. Despite the different sizes of their excavation, the three coincide in showing that 90% of their archaeological materials are accumulated in less than 200 m2 (Table 1.4). Only one of them (DS) shows a bigger accumulation area, because it embodies two different cluster areas. If split in two, the area concentrating 90% of materials is ~200 m2 for each cluster (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Cobo-Sánchez, Baquedano, Mabulla, Gidna and Diez-Martin2024). If focusing on the main clusters in each site, with significantly intense clustering values, the patterning is also obvious; sites display intense clusters in areas between 18 m2 and 30 m2 (Table 1.4). It is worth commenting on the differences found at DS. This site contains two spatially discrete clusters with three high intensity spots; two of them form part of the same elongated cluster (Figure 1.2). If considering these two intensity spots as two hot spots within the same cluster (with intense clustering values of 25 m2 and 30 m2 respectively), since they are connected through more material intensity than the surrounding space, it could be argued that DS is a different version of the mono-cluster pattern, given that the accessory cluster is very small (showing intense clustering in only 8.5 m2). In general, site size, and lithic and bone densities in these clusters are similar (Table 1.4).
| Level | Excavated area (m2) | Area (m2) covering 90% of archaeological remains | Area (m2) of significant clusteringFootnote * | Number of lithic specimensFootnote ** | Number of bone specimensFootnote ** | |
|---|---|---|---|---|---|---|
| FLK Zinj | 22a | 300 | 183 | 18 | 2.470 | 3.510 |
| DS | 22b | 554 | 406 | 25-30Footnote * | 1229 | 3458 |
| PTK | 22a | 187 | 157 | 22 | 1004 | 2589 |
AGS and the other level assemblages are still in the process of study, and they are, thus, not documented here.
* After Cobo-Sánchez (Reference Cobo-Sánchez2020). The double figure in DS refers to the sizes of both concentrations inside the same elongated cluster.
** Numbers reported and drawn by Leakey (Reference Leakey1971).
Regarding faunal composition, the Bed I sites show a similar predominance of individuals according to taxon. In all of them, Kobus sigmoidalis is the most abundant species, followed by Antidorcas recki, and Parmularius altidens. These three taxa add up to 18 individuals in FLK Zinj, 13 individuals (21 if adding postcranial) at PTK (Organista et al., Reference Organista, Moiclán, Aramendi, Cobo-Sánchez, Egeland and Uribelarrea2023), and 20 individuals in DS (Cobo-Sánchez, Reference Cobo-Sánchez2020; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Cobo-Sánchez, Baquedano, Mabulla, Gidna and Diez-Martin2024) (Table 1.5). Their representation is so similar among sites that one is tempted to think that the three sites represent similar occupation times and similar hominin group sizes. The fact that the three predominant taxa are nonmigratory would suggest that occupation took place in moments in which the dry season was not drawing allochthonous fauna (including migratory species) to the lacustrine basin. If ecological dynamics were similar to those documented in the Serengeti–Nogorongo ecosystems today, that would mean a predominant occupation during the dry season (Linares-Matás and Clark, Reference Linares-Matás and Clark2022). Nevertheless, if wildebeests were migratory then as they are today, their marginal representation in all sites would indicate that hominins were around the lake basin part of the wet season or even all year-round, unless the presence of wildebeest remain at the sites, mostly identified by isolated dental remains, could also be a background scatter occurring on the landscape deposited through processes that did not involve a hominin agency. An argument against this is that the presence of Connochaetes is regular across all the archaeological samples, and they appear represented by a similar number of individuals. The inference of at least two different occupation episodes at some of these sites has been made based on the distinctive microwear of herbivore dental enamel (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Sánchez-Flores, Baquedano, Arriaza, Aramendi and Cobo-Sánchez2019c). The pit-scratch pattern documented in Antidorcas at FLK Zinj suggests deposition in at least two different times of the year. In contrast, the pattern retrieved from the more abundant Kobus remains indicates a single occupational episode. This is supported by dental microwear documented on Parmularius teeth. The same results from Parmularius were obtained from the small sample obtained at DS. Therefore, the bulk of the faunal remains at FLK Zinj and DS would indicate (limited by the small and probably non-representative nature of the sample) site occupation during the same season. This would be supported by the virtual lack of subaerial weathering on these assemblages. The Antidorcas data may be discordant at FLK Zinj because they were derived from only four individuals, and a case was made for the likely addition of some Antidorcas remains to the site by feline agency (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo2015), given the presence of a felid-modified complete element at the site that was not processed by hominins (Parkinson et al., Reference Parkinson, Plummer and Hartstone-Rose2015). These inferences must be sustained with more efforts to collect bigger samples for microwear analysis from all these sites.
| Taxa | FLK ZinjFootnote * | FLK ZinjFootnote ** | PTK | DS | averageFootnote *** |
|---|---|---|---|---|---|
| Antidorcas recki/antilopini | 7 | 7 | 3 | 5 | 5 |
| Kobus sigmoidalis | 9 | 7 | 8 | 11 | 9 |
| Parmularius altidens | 6 | 4 | 2 | 4 | 4 |
| Connochaetes sp. | 3 | 2 | 2 | 4 | 2 |
| Tragelaphus sp. | 1 | 2 | 1 | ||
| Equus sp. | 5 | 1 | 3 | 3 | |
| Kolpochoerus | 5 | 1 | 1 | 1 | 1 |
| Megalotragus | 2 | ||||
| Giraffidae | 2 |
Only taxa represented by more than one individual in at least one site appear.
* Bunn and Kroll (Reference Bunn and Kroll1986), using all dental remains
** Domínguez-Rodrigo et al. (Reference Domínguez-Rodrigo, Barba and Egeland2007) using only encased mandibular and maxillary dentition. Equid remains were not available at the Louis Leakey Memorial Institute in the National Museum in Nairobi when the study was conducted.
*** Using minimum estimates for FLK Zinj and rounding up to the closest integer. Only taxa with a minimum of 1 MNI on average are documented.
Taphonomic properties. All these assemblages are dominated by long limb bones, with cranio-axial elements following (Bunn, Reference Bunn1986; Bunn et al., Reference Bunn1986; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Barba and Egeland2007). Although some complete elements are marginally represented, most of the assemblages are green-broken with clear hammerstone percussion signs in the form of acute breakage planes and abundant notches (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Barba and Egeland2007). Bone surface modifications are abundant and they are mostly generated in the butchering process. Although there is some variability, frequencies of cut marks on long bones (the most abundant anatomical sections) are homogeneously represented in all the compared assemblages in concordance with carcass size (Table 1.6). Cut marks on limbs range from 10% to 15% in small carcasses and from 15% to 25% in larger carcasses, with estimates corrected for diagenetic poor cortical preservation and dry bone breakage. Percussion marks show more variance, but their frequencies range between 7–22% (small carcasses) and 12–24% (larger carcasses). Carnivore tooth mark damage is conspicuously low, being the least common bone surface modification type in the three assemblages (Table 1.6). Toothmarking is more abundant on long bone ends and near-epiphyseal portions, as would correspond to the impact of post-depositional scavenging agents.
| CM | PM | TM | ||
|---|---|---|---|---|
| Small carcasses | ||||
| FLK Zinj | 14.9 | 22.1 | 17.3 | |
| PTK | 10 | 7 | 3 | |
| DS | 11.7 | 8.41 | 5.56 | |
| Medium-sized carcasses | ||||
| FLK Zinj | 22.6 | 12.5 | 14 | |
| PTK | 15.7 | 16.1 | 5.5 | |
| DS | 25 | 24.14 | 4.31 | |
Estimates are corrected according to cortical bone surface preservation and green/dry breakage.
CM, cut marks; PM, percussion marks; TM, tooth marks.
The homogeneity in the high frequencies of butchery marks and lower frequencies of carnivore modifications is accompanied by the anatomical patterning where these marks occur. A recent study comparing the three-dimensional distribution of cut marks showed that in the three assemblages, the patterning in frequency and location of where cut marks occur is virtually identical, underscoring the same behavior behind it (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Baquedano, Organista, Cobo-Sánchez, Mabulla and Maskara2021).
In summary, the Bed I anthropogenic sites exhibit a common set of physical properties in size, spatial patterning, and composition, which indicate a specifically gregarious and cohesive group attitude during food processing and consumption. This differs from modern human foragers’ social structure at camps, and compares positively with the behavior of gregarious large carnivorans upon prey acquisition and consumption.
This situation also shows that the Bed I sites selected have a taphonomically fully anthropogenic signal in their accumulation and formation. What was termed the “Olduvai effect” (Barr et al., Reference Barr, Pobiner, Rowan, Du and Faith2022), as a euphemistic description of the outlying position of the Olduvai record when lumped with other African sites with poor preservation, or with a palimpsestic nature – where hominin agency is unknown, or diluted among other agencies – turns out to be a signature of truly Early Pleistocene anthropogenic assemblages.
Discussion
Models on early hominin behavior created around the premise that they should emphasize novel behaviors that are neither modern human nor nonhuman extant primate to fit an intermediate-stage evolutionary theoretical framework do not contribute in any scientific or heuristic way to the advancement of knowledge on early human behavior. The reasons are that: (a) They are not testable (for lack of modern analogs), (b) their heuristics cannot be confronted because there can be as many untestable models as paleoanthropologists willing to put their imagination to work, and (c) they do not need to abide by comparative behavioral ecological criteria, by depicting hominin behaviors that do not necessarily follow major adaptive trends and evolutionary constrictions. This is exactly what explains the stagnated status of the interpretation of early Oldowan sites for the past four decades. Several radically different models coexist without any of them having been convincingly tested, because they are in essence conceptual models, which selectively adapt to some of the empirical information from the archaeological record, but never to all or most of it. In addition, most of these models, especially those that emphasize the ethological framework, use primates as references (Haslam et al., Reference Haslam, Hernandez-Aguilar, Ling, Carvalho, de la Torre and DeStefano2009). This is also an additional problem, because although adaptation is a combination of anatomy and behavior, most of the socio-reproductive structures in the evolution of mammals are ecologically channeled. This explains why in completely unrelated mammal groups, evolution produced similar (in some cases) identical ecological niches and organisms adapted to them in a similar fashion. See for example the occupation of ecological niches in the evolution of mammals in the Americas and their counterpart in the African continent (Prothero, Reference Prothero2006; Prothero et al., Reference Prothero and Schoch2002). This is why a broader ecological reference that incorporates other similarly adapted taxa, even if phylogenetically unrelated, may be more useful than using only apes as analogs.
Models on hominin behavior like the “favored place” (Shick, Reference Shick1987) do not really explain what triggered the selection and redundant use of the same loci. Hominins in this model are presented as being dependent on raw materials for stone tool production. This would have conditioned the loci on the landscape where stone tools and bones were more likely to accumulate. According to Schick (Reference Shick1987), repeatedly visited sites with nearby lithic sources would have a net import of artifacts (and potentially food residue, like carcass parts) greater than on other parts of the landscape, because the need to transport stone was less immediate and artifact discard rates were increased, due both to the proximity of raw material sources and the use of tools on these spots (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Barba and Egeland2007). This assumption is logical, but has some flaws. If this is the case, one wonders why these sites were not created at or in the immediate vicinity of the raw material sources. Although the Bed I sites are not far from some of the raw materials used they are still a few kilometers away from them, not at their immediate source. We conceptually also understand that the site may accidentally be formed by incremental use of the location through reuse of lithic and other resources – in the same fashion as chimpanzee nut-cracking sites –, but we do not know what makes them “favored” over so many other potential locations. Chimpanzee nut-cracking sites do not compare to Oldowan sites in density of materials (Carvalho et al., Reference Carvalho, Cunha, Sousa and Matsuzawa2008; Mercader et al., Reference Mercader, Panger and Boesch2002). Chimpanzees reuse the rocks previously transported there, but do not seem to continue transporting rocks in any significant way over the years of reuse. Therefore, this hominin model lacks an analogical referent. In addition, one assumes that if reuse of lithic resources is a premium, these should have been stored instead of transported for just the required activities of each occupational moment. There are Oldowan sites, where the bulk of the lithic artifacts are just flakes (e.g., PTK) and these, after use, could only have been efficiently reused through renewing their edges; however, retouched flakes are virtually nonexistent in the assemblage, which suggests that if reoccupation existed, it was not aimed at reusing local lithic resources. A last observation is that this model conceptualizes its validity if site formation took place through reoccupation along large time intervals (i.e., years). However, as described above, the Bed I anthropogenic sites have integral taphonomic properties and weathering indicators suggesting that their formation took place in much shorter times.
This observation could also be made of the “stone cache” model (Potts, Reference Potts1988). Here, as explained in Domínguez-Rodrigo et al. (Reference Domínguez-Rodrigo, Barba and Egeland2007), according to Potts (Reference Potts1988), the co-occurrence of lithic material and bones on the landscape could be due to: (a) The transport of carcasses to raw material sources; (b) the movement of stones to carcass acquisition points; or (c) the transport of both stones and carcasses to referential places. Arguing from an optimal foraging standpoint, Potts concluded that the third option would have been the most energetically efficient. If encounter rates with carcasses increased, hominids were expected to establish several referential places in order to reduce transport distances. This stood in contrast to Isaac’s “single-place” strategy where hominids would transport carcasses to a single home base regardless of transport distance. It was therefore suggested that early sites represented “stone caches” – collections of stored lithic material – where carcasses would have been transported for quick processing. Such a strategy, like that employed by, for example, leopards, which brings prey to particular spots for consumption, was meant to avoid competition with other carnivores and to cut down on energetic costs in faunal processing. Thus, these caches would have been carcass processing loci only. According to Potts, this model was appealing because hominids could be understood wholly within an ape behavioral framework. They were not required to possess premeditation or a fundamentally different type of social organization that fostered extensive food sharing. Therefore, the model predicted an intermediate range of behaviors, somewhat different from extant apes, but not quite fully “human.” Testing this model would require: (a) That unmodified rocks at archaeological sites were not transported or deposited there by the action of natural abiotic processes, and (b) that there is a continuity in the reduction sequence represented at sites from the unmodified rocks to the discarded knapped products. Neither of these conditions are satisfied. In similar lacustrine basins, natural rainwash streams transport and eject similarly sized rocks in concentrations similar to that documented in some early sites during the rainy season (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Barba and Egeland2007). In addition, a reanalysis of the Bed I “manuports” suggested that their small size and poor quality (being porous basalts), together with the fact that most detached pieces are in a different raw material (quartz), identified these stones as ecofacts (de la Torre and Mora, Reference de la Torre and Mora2005). The discontinuity in the reduction chain of the unmodified and modified lithic part of the assemblages refutes the axial argument of this model. As an additional argument, the fast accumulation of the bulk of the faunal materials counters the longitudinal use of the site for years, as was originally posited in the model.
The refuge model (Blumenschine, Reference Blumenschine1991; Blumenschine et al., Reference Blumenschine, Cavallo and Capaldo1994) would be more similar to some of the behaviors exhibited by mammal carnivorans that may mobilize locally their prey depending on the ecological context (competition and vegetation). However, the model also fails to address why the accumulation in the same spot is redundant, given that all carnivores who mobilize prey to avoid competition do so in the periphery of the kill, which being always stochastic does not enable the repeated use of the same location. This applies even in those cases of serial predation around ponds, where the deposition of carcasses is less concentrated, much more scattered, and barely redundant on exactly the same spot (Domínguez-Rodrigo, Reference Domínguez-Rodrigo1993). Only in some cases, because of the limited presence of trees can several tree-stored carcasses be transported by leopards to the same tree (up to maximum of five have been documented), but their remains are either eaten or transported away by scavengers once they fall on the ground (Cavallo, Reference Cavallo1998). At the loci of the Bed I anthropogenic sites, as was described above, there was not a paucity of trees; quite the opposite, arboreal vegetation was dense to the point of allowing the presence of ferns in some places (Arráiz et al., Reference Arráiz, Barboni, Ashley, Mabulla, Baquedano and Domínguez-Rodrigo2017). Intense clustering as a result of reutilization of just one tree cannot, therefore, be taken as a valid interpretation in the light of the available paleobotanical data.
The “near-kill” location model attempts to address the issue of redundancy by explaining that sites could be similar to modern Hadza near-kill locations occurring in ambush spots near water sources, where they preprocess carcasses for transport to camps (O’Connell, Reference O’Connell1997). This model, however, has a conceptual and an empirical problem. The conceptual issue is that it contains its contradiction in its formulation. Modern forager near-kill locations, as spots where carcasses are prepared for transportation to home bases, are extensions of camps; that is, they owe their existence to the presence of home bases. Food is prepared in those spots because it is transported to camps for shared consumption. Without camps, near-kill locations lose their meaning. The model does not cancel the problem of “home bases” in the early Pleistocene, but transfers it to different unknown loci. We even applied it to interpret the space-delimited archaeological record of Type Section at Peninj (Tanzania), because it was formed by spatially close and small concentrations of lithic and faunal remains, with a reduced number of animals per location, and the presence of semi-articulated elements in some localities (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Hovers and Braun2009; Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, de La Torre, de Luque, Alcalá, Mora and Serrallonga2002). However, to date not a clear study containing all the raw data has been published of these modern forager hunting blinds. O’Connell (Reference O’Connell1997) admits that most of these hunting blinds contain small amounts of fauna, and that they can be scattered over more than 10.000 m2; however, he mentions – referring to Lupo (Reference Lupo1993) –, as an exception, one faunal assemblage from a hunting blind where five large mammals are represented by more than 500 NISP, more than 300 MNE, and 11 MNI, dominated by appendicular parts. Lupo (Reference Lupo1993) provides a table with ranking values in the representation of elements in presumably that hunting blind (Mugugu), but not with raw data. The first thing that is noticeable is the low element fragmentation, since each element is represented by less than 2 specimens. The second consideration is that this assemblage has been formed over an extensive period of time involving several years. As a result, bone weathering stage 0 is not predominant, and the collection contains specimens displaying weathering stages from 0 to 4 (Lupo, Reference Lupo1993). This situation contrasts with the faster accumulation rates documented at the Olduvai Bed I sites and their lower weathering. Although skeletal representation varies according to taxon, if we take impala as representative of small game, and “alcelaphine” as representative of medium-sized bovids, the Mugugu assemblage is dominated by skull, neck, radius-ulna, and tarsals, followed by tibiae and metapodials for small bovids, and skull, neck, tibiae, and metatarsals in medium-sized bovidae. There is asymmetry in limb representation, since front limbs have not been recorded. In both cases, it can be seen that the highest-yielding meat-bearing bones (humeri and femora) are underrepresented or non-existing, and that skull and feet are best represented, as would be expected if bone discard was carried out on the lowest-yielding bones to ease transport weight. This contrasts with the appendicular bones best represented at the Olduvai Bed I sites. Meat-bearing bones (humeri and femora and zygopodials) are better represented than or equally represented as feet (depending on carcass size) in the bovid samples from these sites, implying a very different transport and discarding behavior. Ribs and vertebrae are also more abundantly represented than at Mugugu for both carcass sizes (Cobo-Sánchez, Reference Cobo-Sánchez2020; Organista et al., Reference Organista, Moiclán, Aramendi, Cobo-Sánchez, Egeland and Uribelarrea2023). Therefore, qualitative assessment of the appendicular-dominated assemblage is required before making claims that the Olduvai sites could be near-kill locations because they are dominated by appendicular parts as the Hadza hunting blind assemblages are (O’Connell, Reference O’Connell1997). An interesting feature of Mugugu is that the alcelaphine carcass sample contains no front limbs. These are more easily transported than hindlimbs for carcasses this size. At DS, front limbs are overrepresented; this could indicate the opposite end of the transport process, from kill to central-consumption locus.
Finally, a point that is of greater concern than any of the above is that the anomalous high-density hunting blind assemblage (Mugugu) is not a controlled assemblage. It has not been monitored, especially along the years sampled by the preserved fauna. Given that several other hunting blinds are palimpsests, which have also been used as temporary camps (Lupo, Reference Lupo1993), one wonders if that was the case of Mugugu also. In essence, there is no justifiable hunting blind faunal assemblage published yet, which could be taken as prototypic for this type of faunal accumulations, and as referential for interpreting prehistoric sites. Therefore, no reliable modern assemblage can currently be presented, which could be used to sustain a “near-kill location" model of sorts.
These are just a few examples on how some of the most popular models produced in the past four decades fail to fit the available archaeological data properly, and present several conceptual as well as empirical contradictions that question whether we are on the right path to interpreting the earliest archaeological record.
Conclusions
Traditionally, Homo habilis hominins were interpreted as the creators of the Oldowan, simply because they co-occurred on the same strata containing Oldowan tools (Leakey, Reference Leakey1971). However, co-occurrence does not imply causation. For the same reason, Australopithecus garhi was argued to be the first stone tool user, because it appeared on the same landscape as the earliest tools, 2.6 million years ago (Asfaw et al.., Reference Asfaw, White, Lovejoy, Latimer, Simpson and Suwa1999). If these predicates are valid, even baboons could also be factored as potential stone tool users, given their fine manipulative skills and the fact that other primates (i.e., capuchine monkeys, chimpanzees) use and modify rocks as tools (Reeves et al., Reference Reeves, Proffitt, Malaivijitnon and Luncz2023). The discovery of Homo habilis-like hominin remains dated to 2.8 Ma (prior to the earliest securely identified archaeological record) (Villmoare et al., Reference Villmoare, Kimbel, Seyoum, Campisano, DiMaggio and Rowan2015), the discovery of Homo erectus remains dated to 2 Ma (Herries et al., Reference Herries, Martin, Leece, Adams, Boschian and Joannes-Boyau2020), and its association with 2 Ma Oldowan and Acheulian tools (Mussi et al., Reference Mussi, Skinner, Melis, Panera, Rubio-Jara and Davies2023), makes it a better candidate for the formation of those anthropogenic sites where early access/hunting to small and medium-sized carcasses can be taphonomically supported. This is especially true ever since Homo habilis has been shown to be a three-foot anatomically primitive hominin (Johanson et al., Reference Johanson, Masao, Eck, White, Walter and Kimbel1987), highly dependent on trees, with no clear anatomical evidence for any terrestrial adaptation involving cursoriality, running, throwing, or fine hand manipulation. If anatomy conditions adaptation, a hunting lifestyle in early humans must have been accompanied by strong anatomical modifications involving more terrestriality, larger size and better use of arms and legs than those inferred from the Homo habilis anatomy. Using a behavioral ecological proxy, hyenas are not adapted to hunt buffaloes because they lack an anatomy which would enable them to have the size, strength, and stealth to achieve what lions can do. Anatomy and behavior coevolve.
Increase of body and brain size, slower growth, decreased sexual dimorphism, and cursorial adaptations are typical of denning gregarious mammal carnivorans targeting large game (Smith et al., Reference Smith, Swanson, Reed and Holekamp2012). In human evolution, all these physical characteristics are detected seemingly synchronically with the emergence of Homo erectus two million years ago (Anton, Reference Anton2003; Bramble and Lieberman, Reference Bramble and Lieberman2004; Lieberman et al., Reference Lieberman, Bramble, Raichlen, Shea, Grine, Fleagle and Leakey2009). One may argue that they hide the same adaptive strategy in both cases, shaped by universal behavioral ecology selective factors. If so, this strategy comes with other dependent factors attached, which we could theoretically also infer for hominins. One particular case is that all these carnivorans produce altricial (i.e., helpless and immature at birth) offspring, which justifies their denning behaviors. Immature progeny must remain secluded during the parental foraging trips, and they must be fed regularly. Food repeatedly brought to these dens is mainly consumed by subadult individuals, and adult individuals who stayed at the den alloparenting. In this regard, mammal carnivorans are no different from most avian species and their nesting behavior. The expansion of this behavior by eusocial species, so that food reaches all adult individuals in the community is just an additional extension of this basic provisioning behavior.
When we look at modern human foraging groups, home bases also have a prominent role in protecting young individuals through alloparenting behaviors. Some evolutionary anthropologists have paid close attention to this:
Surprisingly little attention has been paid to why our ancestors began using central places; avoidance of predation is likely the initial reason, but I argue that taking food back to a central place may have been favored by the benefit gained from leaving young weaned children behinds with an adult babysitter while foraging…. Children who are too big to carry or too young to keep up with adults during foraging pose a problem that can be solved by leaving them behind in a central place where others can look after them. Those who go out foraging must bring food back to feed the people who stay in camp with young children. A crèche is the place and people who look after the young of others, something that is typical of lions as well as of the Hadza.
A detailed analysis of Hadza camps based on subadult individuals, using three cohorts (nurslings [under 2 years old], weanlings [2–3.9 years old], and older children [>4 years old]) showed that weanlings only leave the camp 1.9 hours per day, because women take nurslings with them during plant foraging but leave weanlings in camp; and most of the time they leave the camp, they do so only beyond the edge of the camp for playing (Marlowe, Reference Marlowe2010). Sick, injured, and older people may also stay in camp for longer periods than the rest of the adult cohort, and some of them play a prominent role in the alloparenting of the weanlings (Marlowe, Reference Marlowe2010). Given that modern human foragers maintain age cohorts virtually residing at camps most of the day, their central-provisioning behavior is an extended version of the central-place foraging behavior of most denning or nesting animals. Given that the latter and modern foraging humans share the same group of adaptive characteristics, it would be logical to think that these apply to earlier hominins. This is why the empirically documented central-provisioning behavior at the Olduvai Bed I Oldowan sites would be best explained as the sites having had also a prominent breeding/nursing role. This would have made Oldowan sites a sort of primate “nesting” (sensu eusocial organisms) ground. Does this mean that the essence of modern foragers’ camps emerged two million years ago? Probably, but this should not surprise anthropologists more than realizing that humans did not evolve this set of adaptive features as something novel compared to other mammals, but as a convergent path during their adaptation to similar ecological settings. If this set of features is interdependent, then their emergence with the earliest archaeological record in the Early Pleistocene should not be surprising. Only theoretical prejudices founded on an unjustified assumption that all evolutionary processes must be an aggregation of gradual (and progressive) features would find this interpretation problematic.
Beyond the physical novelties listed above, compared to previous and other penecontemporaneous hominins, Homo erectus (if this taxon is indeed responsible for the Oldowan and Acheulian archaeological records at Olduvai Gorge), was also consuming meat, and targeting large animals for that purpose. Its anatomy showed the earliest evidence of running, following similar adaptation as cursorial mammals targeting similar prey sizes (Smith et al., Reference Smith, Swanson, Reed and Holekamp2012). The most abundant animals represented at the Bed I Oldowan sites are waterbucks, weighing between 160–225 (females) and 200–270 kg (males). Medium-sized animals, if accessed when fleshed (as suggested by taphonomic research), constitute a large food surplus for several individuals. This implies that the food that entered sites was abundant, and likely to be shared with others, as a central-provisioning behavior would require. Until here, nothing is new from previous interpretations (Bunn et al., Reference Bunn1986; Isaac, Reference Isaac1983, Reference Isaac1978; Oliver, Reference Oliver1994). As mentioned in the previous paragraph, whether considering the behavioral ecology of tropical mammalian carnivorans or that of eusocial animals, this locus-centered (animal) food transport is better understood if sites acted as breeding/nursing grounds, where hominins engaged in central provisioning. Hominins used sites extensively, judging from the amount of food remains represented in them (Domínguez-Rodrigo et al., Reference Domínguez-Rodrigo, Cobo-Sánchez, Aramendi and Gidna2019b; Domínguez-Rodrigo and Pickering, Reference Domínguez-Rodrigo and Pickering2017), and from the fact that they accumulated over short time spans. So much food over so little time would be more easily understood if sites were food-sharing grounds, where nutrients were repeatedly transferred among individuals.
In the generally forested and wooded fluvio-lacustrine habitat where these sites were formed (Arráiz et al., Reference Arráiz, Barboni, Ashley, Mabulla, Baquedano and Domínguez-Rodrigo2017), hominins were not ecologically restricted to use exactly the same locations if sites were only temporary refuges for individual self-provisioning (Blumenschine, Reference Blumenschine1991; Blumenschine et al., Reference Blumenschine, Cavallo and Capaldo1994). They were loci that show so far two features in common: They were in wooded/forested habitats, and very close to fresh water sources. These features, especially the latter, are also shared by modern human foragers (Marlowe, Reference Marlowe2010).
The perspective presented here, following principles of comparative behavioral ecology, may seem that it leaves little room for intermediate progressive evolutionary steps towards the configuration of modern foraging human behavior. By defending the view that the basic function and dietary use of modern human foragers’ camps was structurally in place at least two million years ago, one could argue that there is some contradiction in assuming stasis in the behavioral structure, but evolution of the biology of the hominin organisms responsible for it. This is not correct. The basic anatomical structure (in size and morphology) of modern humans was first documented in Homo erectus. It should be no surprise from this perspective that the earliest evidence of a similar structural behavior accompanied the emergence of this anatomy/physiology and its evolution. The subsequent evolution of this species into its descendants is mainly determined by a process of robusticity and gracilization, accompanied by a punctuated or progressive encephalization process, despite the mosaic patterning (including more primitive features) documented in some side branches (e.g., Homo floresiensis or Homo naledi). This latter process also leaves room for assuming that the structural behavior for these side branches (still with strong arboreal components) may have been utterly different. Unfortunately, no securely associated archaeology is known for them. Considered from this perspective, the behavioral evolution of Homo erectus and its descendants is more of an accretional process, in which elements or layers are added to the basic structure, than a progressive (i.e., linear) evolution of the structure itself.
Despite that the basic human central-provisioning behavior is probably represented at the Olduvai anthropogenic Bed I sites, there is still ample space for the differences that existed compared to modern humans. Modern foragers have piled up several behavioral layers on top of the basic structure just described. They have managed to evolve an intensive social life and focus most of it at the camp. They have also evolved a symbolic cosmogenesis and recreate it at the camp. They have adopted different socio-reproductive strategies and have adapted them to the central-provisioning basic behavior. This is one of the major differences with early humans and Oldowan sites. The intensively cohesive group structure that could potentially be inferred from the densely packed mono-clusters that constituted most of the Oldowan anthropogenic sites is not documented among modern foragers. This indicates that their socio-reproductive strategy was very different. Early human groups acted as a single family instead of an aggregation of individually defined reproductive units as modern human foragers do (i.e., households). In this sense, early humans consumed their food (once it reached the site) like modern lion prides do (most individuals being highly related) and not like modern hyenas (most individuals being unrelated).
An inclusive theory of human evolution assumes that humans are part of the organic world and follow the same rules and are constrained by the same rigors of natural selection. Such a theory places a strong emphasis on ecology. This is the view adopted here. It has not been the traditional view on how to interpret early human behavior. The predominant paradigm still relies heavily on singling out humans from the rest of taxa, on a predominance of homology over analogy, and on the emphatic use of chimpanzees as a referent, since humans are viewed almost exclusively as primates instead of as mammals. From that perspective, humans are indeed very different in many evolutionary aspects from other primates, and the approach described here makes the quest for such differences understandable. This positioning also thrives on a view of progressive evolution that is not orthodox. Although mosaic evolution has been pervasive throughout the human evolutionary process, with many side branches exhibiting very primitive anatomies, adaptations, and very likely primitive accompanying behaviors until very recently, anthropology places a special focus on the evolution of one of those branches leading to Homo sapiens as a progressive process. Despite their efforts to assimilate early humans to apes, most of the models produced so far about the behavior of hominins at Oldowan sites have been forced into a paradigmatic view that requires them to fit a progressive path towards modern humans. This almost automatically requires anthropologists to think of the early stages of human evolution as necessarily primitive, equating this term with being closer to ape (or more exactly, chimpanzee) adaptation and behavior. However, chimpanzees (and all apes) evolved in very different environments from early Homo, and there is no epistemologically valid way of linking them, since a minimum of four million years separate chimpanzees from early Homo. From a chronological standpoint, from an anatomical and physiological point of view, and from a neurological point of view early Homo sensu lato (i.e., Homo aff. erectus) is much closer to Homo sapiens than to modern chimpanzees. Therefore, the behavior of these early humans conceptually should be assumed to be closer to us than to any other primate. However, this clashes against the progressive evolution perspective that still pervasively shapes many human evolutionary narratives.
Intermediate evolutionary models are intellectually challenging, and some of them provide fascinating interpretations. However, most of those models existing so far are not testable and do not fit empirical evidence well (see Discussion). Some are narratives stemming from inductive use of empirical information. Others provide stimulating evolutionary scenarios, which although feasible, provide uncertain heuristics given their limited testability. However, the main drawback of this type of modeling is that it is not carried out considering behavioral ecology. It is not considering humans and their ancestors as belonging to nature’s deep web of socio-ecological adaptations. It does not assume that organisms adapted to the same environments and displaying a similar set of adaptive features are better proxies for interpreting hominin adaptation and behavior than phylogenetically close primates, all of which are adapted not only to different environments, but also different biomes. The contextual gap of apes and early humans is too wide ecologically to be heuristically useful analogs. A proof of concept of the criticism addressed here to these “intermediate” models is that we are still where Glynn Isaac left it four decades ago (Isaac, Reference Isaac1983). Our knowledge of early human behavior and Oldowan sites has not only not advanced, but it is stagnated by a wide range of possible scenarios, most of which cannot be tested and conceptually oppose one another. No wonder some archaeologists became skeptical of them all and decided that the only premise of those models which can generate some consensus is that they were locations where hominins brought food and ate it (Ferraro, Reference Ferraro2007).
The behavioral ecological approach adopted here may also be considered testable. It can be logically structured as follows:
A: Increasing levels of cooperation in mammals require group formation.
B: The most cooperative mammal groups are kin related (as are also most eusocial especies).
C: Gregarious carnivorans are some of the more cooperative mammals, and their prey size is related to bulk group size.
D: Gregarious carnivorans produce altricial offspring.
E: Cooperation and altriciality require denning and central-provisioning behaviors.
F: Following these principles, highly cooperative hominins would most likely be kin-related groups with altricial offspring, with central-place provisioning, and dietarily dependent on meat consumption.
If making new predictions is the hallmark of scientific hypotheses and theories (Domínguez-Rodrigo, Reference Domínguez-Rodrigo and Domínguez-Rodrigo2012a, b), one that stems from the comparative behavioral ecological approach adopted here is that archaeological sites older than two million years, with the same properties of the Olduvai Bed I anthropogenic sites described here, would require evidence of hominin increasing body and brain size, with potentially altricial offspring. In effective terms, it would require some hominin like Homo erectus to have been present earlier than currently documented.
Following the premises above, the “testability” of the model would predict physical evidence of redundant spatial focalization of food consumption, and evidence of food surplus under the form of primary access to large game. We admit that using the archaeological evidence from Oldowan sites as confirmation for the model is somewhat spurious, even if the whole set of adaptive features could be inferred therefrom. Nevertheless, we argue that if any interpretation of the Oldowan record is to consider early humans as organisms (and more specifically, mammals) that can be understood through behavioral ecology principles, it must do so by taking the reconstruction provided in the present essay as a null hypothesis. Rejecting this baseline hypothesis would open the door to alternative scenarios. By adopting the baseline interpretation, we have a model that temporarily explains early human behavior. Refusing its adoption leaves us in the same current interpretive limbo as we now are, where Oldowan sites are a complete enigma. We should not let academic narratives control which option we take if our field is to have any meaningful impact in how we understand our evolution, and how we pass this knowledge to society.