


Introduction
Branched broomrape is an obligate holoparasitic plant that is threatening the processing tomato industry in California. This billion-dollar sector produced roughly 10 million metric tons of tomato in 2024, with an average productivity of 112 metric tons per hectare (USDA-NASS 2025). Although processing tomato production in California faces a broad suite of pest challenges, broomrapes (Phelipanche spp.) stand out as the most urgent threat (Martin et al. Reference Martin, Swett, Vinchesi-Vahl and Parreira2021). Branched broomrape is listed as an “A”-rated pest subject to regulatory actions by the California Department of Food and Agriculture (CDFA), imposing substantial economic and operational burdens on local tomato growers (Kelch Reference Kelch2017).
Native to the Mediterranean basin, branched broomrape lacks chlorophyll or typical root systems and depends entirely on a host for water, carbon, and mineral nutrients (Osipitan et al. Reference Osipitan, Hanson, Goldwasser, Fatino and Mesgaran2021; Joel Reference Joel2009). Like a majority of parasitic plants, branched broomrape seed germination requires a preconditioning process that includes incubation in a moist, warm environment for 5 to 21 d and a germination stimulant from a host plant (Murdoch and Kebreab Reference Murdoch, Kebreab, Joel, Gressel and Musselman2013; Xie et al. Reference Xie, Yoneyama and Yoneyama2010). A wide range of valuable crops in several families, including Alliaceae, Asteraceae, Brassicaceae, Cannabaceae, Cucurbitaceae, Fabaceae, and Solanaceae, can produce the germination stimulant, strigolactone, that branched broomrape needs for germination (Miyao Reference Miyao2017; Osipitan et al. Reference Osipitan, Hanson, Goldwasser, Fatino and Mesgaran2021).
Branched broomrape was first detected in California in the early 1900s and was presumed to have been eradicated by the late 1980s following fumigation programs with methyl bromide (Gaimari and O’ Donnell Reference Gaimari and O’Donnell2008; Jain and Foy Reference Jain and Foy1989). Although eradication programs based on fumigating the soil with methyl bromide can be effective, their current feasibility is limited by regulatory and economic constraints. Alternative soil fumigants are available; however, to our knowledge, empirical data supporting their efficacy under California environmental and production conditions do not exist (Osipitan et al. Reference Osipitan, Hanson, Goldwasser, Fatino and Mesgaran2021). Compared with the 1900s, the processing tomato industry has changed significantly, most notably with the introduction of mechanized harvest operations and greater movement of machinery between fields in larger farm operations, which leads to greater risk of spreading seed from infested to noninfested fields (Geissler and Horwath Reference Geissler and Horwath2016; Hershenhorn et al. Reference Hershenhorn, Eizenberg, Dor, Kapulnik and Goldwasser2009; Hosseini et al. Reference Hosseini, Swett and Hanson2025).
Managing branched broomrape in California processing tomato in the 2020s requires integrated weed management strategies (Hosseini et al. Reference Hosseini, Swett and Hanson2025; Osipitan et al. Reference Osipitan, Hanson, Goldwasser, Fatino and Mesgaran2021). In recent years, broomrape management research in California has focused on the use of herbicides (e.g., Fatino and Hanson Reference Fatino and Hanson2025; Fatino et al. Reference Fatino, Galaz and Hanson2025), equipment sanitation to reduce seed movement among fields (e.g., Hosseini et al. Reference Hosseini, Osipitan and Mesgaran2022, Reference Hosseini, Swett and Hanson2025), and predictive modeling to guide management efforts (e.g., Hosseini et al. Reference Hosseini, Hanson, Fatino and Mesgaran2026). However, long-term integrated management of branched broomrape in processing tomato and other potential host crops in California will require testing and development of additional approaches, including cultural practices that can be applied to multiple crops and to organic processing tomato. Previous research on Egyptian broomrape (Phelipanche aegypiaca [Pers.] Pomel) demonstrated that flooding and nitrogen fertilizer can reduce broomrape seed germination (Goldwasser and Rodenburg Reference Goldwasser, Rodenburg, Joel, Gressel and Musselman2013), but this has not been evaluated on branched broomrape or under California production conditions.
Fertilizers could affect branched broomrape and other parasitic weeds both directly and indirectly. Fernández-Aparicio et al. (Reference Fernández-Aparicio, Reboud and Gibot-Leclerc2016) reported that a well-fertilized host plant produced less germination stimulant, which led to reduced initiation of parasitism. Conversely, previous studies have shown that nitrogen and phosphate deficiencies in tobacco (Nicotiana tabacum L.) increased strigolactone exudation, and ammonium inhibited strigolactone exudation in red clover (Trifolium pratense L.) (Yoneyama et al. Reference Yoneyama, Takeuchi and Yokota2001; Zhang et al. Reference Zhang, Ma, Zhong, Jia, Wu, Yu and Ye2015). Heavy infestations of crenate broomrape (Orobanche crenata Forsk.) on faba beans have been associated with low soil fertility, and similar patterns have been observed for more Egyptian broomrape attachments under low-nutrient conditions (Jain and Foy Reference Jain and Foy1992; Trabelsi et al. Reference Trabelsi, Yoneyama, Abbes, Amri, Xie, Kisugi, Kim and Kharrat2017).
Fertilizers could also directly reduce branched broomrape seed germination and attachment to the host plant through phytotoxic effects. The application of fertilizers has been shown to suppress other parasitic plants, such as purple witchweed (Striga hermonthica (Delile) Benth) and Egyptian broomrape (Jain and Foy Reference Jain and Foy1992; Jamil et al. Reference Jamil, Charnikhova, Cardoso, Jamil, Ueno, Verstappen, Asami and Bouwmeester2011). In particular, increasing the amount of soil nitrogen and phosphorus through fertilizer application can reduce the germination and subsequent infestation rates of parasitic weeds (Jamil et al. Reference Jamil, Charnikhova, Cardoso, Jamil, Ueno, Verstappen, Asami and Bouwmeester2011). For example, ammonium nitrate combined with potassium phosphate or ammonium phosphate applied alone effectively reduced broomrape parasitism and enhanced tomato growth compared to applications of potassium sulfate (Jain and Foy Reference Jain and Foy1992).
Direct inhibitory effects of fertilizers on branched broomrape seeds could occur during preconditioning, germination stimulation, and seedling elongation stages. In one study, preconditioning Egyptian broomrape seeds in the presence of ammonium salts and urea resulted in a significantly reduction in germination; in contrast, nitrate fertilizer did not have the same inhibitory effect (Jain and Foy Reference Jain and Foy1992). Similarly, in another study, an increase in nitrogen rate (as ammonium nitrate) reduced branched broomrape seed germination and radicle length (Irmaileh Reference Irmaileh1994). Another experiment demonstrated that nitrogen in the ammonium form resulted in greater inhibition than nitrate nitrogen, and the primary inhibition mechanism was a reduction in branched broomrape and Egyptian broomrape radicle elongation rather than inhibition of germination (Westwood and Foy Reference Westwood and Foy1999).
Flooding is another cultural management approach that may help reduce branched broomrape pressure, but research evaluating its effects on this species is limited. Goldwasser and Rodenburg (Reference Goldwasser, Rodenburg, Joel, Gressel and Musselman2013) reported that flooding may be effective in reducing the seed bank of the parasitic weed Rhamphicarpa fistulosa. Several tomato growers in northern California anecdotally observed less broomrape among tomatoes grown in rotation with flooded rice; however, this has not been formally evaluated to date (B.D. Hanson, personal observation).
Despite the potential of nitrogen fertilization and flooding as cultural management practices, research on their effects on branched broomrape remains limited, particularly under California production practices. To begin addressing this gap, controlled environment experiments were conducted to evaluate branched broomrape responses to nitrogen fertilizer and flooding. The specific objectives of this research were to 1) evaluate the effects of nitrogen fertilizer type and concentration on branched broomrape seed germination and radicle elongation when fertilizers are applied at three germination stages (preconditioning, germination stimulation, and radicle elongation) and 2) evaluate the effects of flooding duration and temperature on branched broomrape seed germination in a petri dish study and attachments to the host plant in a pot study.
Materials and Methods
Source of Branched Broomrape Seeds
Branched broomrape seeds used in all experiments were collected from mature plants in an infested field of processing tomatoes near Woodland, California (38.757°N, 121.768°W) in September 2023 and 2024. Mature seed capsules were collected and transported in double containers under appropriate CDFA permit conditions. The seeds were sieved with 400-μm and then 100-μm mesh screens, air-dried at room temperature, and stored at 4 C until use.
General Germination Assay
All laboratory experiments were conducted using the germination assay described by Hosseini et al. (Reference Hosseini, Swett and Hanson2025) with minor adjustments to evaluate seed germination and measure radicle length. Branched broomrape seeds were surface-sterilized by washing in 10 mL L−1 quaternary ammonium sanitizer (Mg4-Quat; Morgan-Gallacher, Inc., Santa Fe Springs, CA) solution for 1 min and rinsed with deionized water three times. Seeds were then kept in the deionized water until transfer. Sterilized seeds were transferred using a pipette onto Whatman filter paper (55 mm diameter) in 60-mm-diam petri dishes. Each petri dish contained 0.5 mL of deionized water and seeds. Petri dishes were sealed with Parafilm and preconditioned in an incubator at 25 C in the dark for 10 d. After preconditioning, the petri dishes were unsealed, and the seeds were air-dried for 2 h. Then, for germination stimulation, 0.5 mL of GR24 (PHY0GR24; PhytoAB, San Jose, CA) solution (10−5 M in Milli-Q water) was added to each petri dish, which were then resealed and incubated at 25 C in the dark for an additional 10 d. After incubation, the petri dishes were unsealed, and the filter paper was transferred onto another filter paper to absorb excess moisture to improve image quality. Images were captured using a CMOS camera (Sony E3ISPM Series) and ToupView software (ToupTek Photonics, Hangzhou, China). The size of a branched broomrape seed is usually less than 0.4 mm (Figure 1) (Hosseini et al. Reference Hosseini, Swett and Hanson2025). Due to the extremely small size of the branched broomrape seeds, the number of seeds per petri dish varied. To evaluate germination percentage, a random area containing 20 seeds was selected on each filter paper; germinated and nongerminated seeds were counted manually from the images, and percent germination was calculated. From the same area, five seedlings were randomly selected to measure the radicle length using Fiji software (Schindelin et al. Reference Schindelin, Arganda-Carreras, Frise, Kaynig, Longair, Pietzsch, Preibisch, Rueden, Saalfeld, Schmid and Tinevez2012).
Branched broomrape seeds from a single cluster on a piece of printing paper (210 × 297 mm).

Fertilizer Study
Seven fertilizers were evaluated in separate experiments for their potential effects on broomrape seed germination and seedling radicle elongation at several germination stages and concentrations. Diammonium hydrogen phosphate (CAS No. 7783-28-0), calcium nitrate tetrahydrate (CAS No. 13477-34-4), potassium chloride (CAS No. 7447-40-7), potassium nitrate (CAS No. 7757-79-1), ammonium nitrate (CAS No. 6484-52-2), ammonium sulfate (CAS No. 7783-20-2), and urea (CAS No. 57-13-6) were obtained from Thermo Fisher Scientific (Waltham, MA) in solid form. In each experiment, an individual fertilizer was examined at six concentrations (50, 25, 12.5, 6.25, 3.125, and 1.56 mM), with 0 mM as the nontreated control. Within an experiment, fertilizer solutions were applied at each of the three stages of the germination process as described below.
Preconditioning Stage
Each petri dish started with 0.25 mL of deionized water and seeds. Then, fertilizer solution (0.25 mL) was added to reach a final volume of 0.5 mL of one of the six concentrations described above, except for the untreated control, which received 0.5 mL of deionized water and seeds. After the 10-d preconditioning process, seeds were rinsed with deionized water three times in a cell strainer (pluriStrainer Mini, 70 μm; pluriSelect USA, El Cajon, CA) and then transferred onto a new filter paper with 0.5 mL of GR24 solution (10−5 M in Milli-Q water) for the germination process.
Germination Stimulation Stage
Preconditioning was performed as described in the General Germination Assay section above. At the time of germination stimulant application, 0.5 mL of solution containing GR24 (10−5 M) and one fertilizer at one concentration described above was added to each petri dish.
Radicle Elongation Stage
The preconditioning and germination stimulation processes were performed as described in the General Germination Assay section. Then, at 5 d after germination stimulant application, petri dishes were unsealed, and the solution was allowed to evaporate by about 50% of the original volume to prevent complete drying, which would halt radicle elongation. The drying time was determined by weighing a new petri dish lined with filter paper and 0.5 mL of deionized water, then monitoring the weight on an analytical balance. Once only 50% of water remained in the petri dish, fertilizer solutions (0.25 mL) were applied to each petri dish to reach one of the six concentrations described above.
Each fertilizer was run separately in a two-factor factorial experiment with seven levels of concentration and applications at three germination stages. There were three replicates of each concentration × germination stage combination for each fertilizer. Germination rate and radicle length were measured 10 d after GR24 application, following the imaging and counting procedures described above. All experiments were conducted twice.
Flooding Study
Germination Study
A laboratory experiment was conducted to evaluate the germination response of branch broomrape seeds to flooding under simulated seasonal conditions in the Sacramento Valley, California. Seeds (∼100 per replicate) were weighed and sealed in fine-mesh nylon bags (Dulytek; 100-µm pore size and 51 mm by 89 mm bag size) and placed in one-quart glass jars containing 500 g of sieved field soil (2 mm pore size) collected from the University of California (UC) Davis Plant Sciences Field Facility (38.539°N, 121.784°W). Seed packets were covered with soil and water was added to each jar and maintained at 2.5 cm above the soil surface throughout the flooding period by periodically adding tap water. Jars were incubated at 10 C and 28 C to simulate winter and summer flooding conditions, respectively, for 3, 7, 14, 28, 42, and 56 d. Initiation of flooding treatments was staggered such that all treatments were terminated on the same day to ensure consistent post-treatment conditions. After the flooding treatments, seeds were retrieved from the bags, surface-sterilized, and transferred to petri dishes for the germination assay described above. This experiment was conducted twice.
Attachment Study
A greenhouse experiment was conducted to evaluate how flooding under two temperature regimes (10 C and 28 C) affected subsequent branched broomrape attachment to tomato roots. Branched broomrape seeds were flooded in the glass jars using the same treatment conditions described in the Germination Study. After flooding, branched broomrape seeds were removed from the bags and mixed into a dual-pot system filled with Redi-Gro Rose and Flower mix (Redi-Gro Corporation, Sacramento, CA), a dark, perlite-free potting medium. The system consisted of a transparent inner pot (1,242 mL and 5.3 cm radius) nested within a black-painted pot of the same size and the perlite-free medium facilitated regular nondestructive observation of broomrape root attachments (Figure 2). The inner pot was filled halfway with potting medium, followed by a layer thoroughly mixed with branched broomrape seed from one seed packet from the simulated flood treatments, and then topped with an additional layer of potting medium. A 14-d-old tomato seedling (Heinz 5508) was transplanted into each pot. All plants were maintained in a CDFA-permitted greenhouse at UC-Davis and fertigated three times per week using the UC-Davis standard fertilization program. The number of branched broomrape attachments was recorded 2 mo after transplanting. This experiment was conducted twice.
Branched broomrape attachments on a tomato plant observed in the dual-cup system in the greenhouse.

Figure 2. Long description
A clear plastic cup houses a tomato plant with an extensive root system visible through the transparent walls. The plant exhibits several branched broomrape attachments, which are parasitic growths. The broomrape appears as thin, thread-like structures with small, yellowish flowers at the base. The plant is placed on a black grid-like surface, likely part of a greenhouse dual-cup system. Other tomato plants can be seen in the background, indicating a controlled environment for study or cultivation.
Data Analysis
All analyses were conducted in R and RStudio software (v.4.4.2; Posit Team 2025; R Core Team 2023).
Fertilizer Study
Germination was analyzed as a binomial count outcome using generalized linear mixed models (GLMMs) fitted separately for each fertilizer using the glmmTMB package (Brooks et al. Reference Brooks, Kristensen, Van Benthem, Magnusson, Berg, Nielsen, Skaug, Mächler and Bolker2017). A beta-binomial GLMM with logit link was used to account for overdispersion, with germination stage, log-transformed concentration, their interaction as the fixed effects, and replication included as a random effect. The radicle length analyses were restricted to germinated seeds and modeled separately for each fertilizer using a Gamma GLMM with log link, including germination stage, concentration, their interaction as the fixed effects, and replication as a random effect. Model assumptions were assessed using simulation-based residual diagnostics using the dharma package (Hartig Reference Hartig2025). Estimated marginal means at observed concentration levels were obtained with the emmeans package (Lenth and Piaskowski Reference Lenth and Piaskowski2025) and reported with 95% confidence intervals.
Flooding Study
Branched broomrape seed germination rates and number of attachments from the flooding study were analyzed by dose-response analyses with the drc package (Ritz et al. Reference Ritz, Baty, Streibig and Gerhard2015). Data from the two experimental runs were pooled because there was no significant difference between them. Branched broomrape seed germination and attachments in relation to flooding duration were best described using a three-parameter log-logistic function as follows(Ritz Reference Ritz2010):
 $Y = \;{u \over {\left\{ {1 + \exp \left[ {b\left( {log\left( x \right) - \log \left( e \right)} \right)} \right]} \right\}}}$
$Y = \;{u \over {\left\{ {1 + \exp \left[ {b\left( {log\left( x \right) - \log \left( e \right)} \right)} \right]} \right\}}}$
where Y is the total germination rate or attachments for branched broomrapes, e reflects the effective concentration (duration) required to achieve a 50% response (ED50), u represents the upper limit, and b is the relative slope at the inflection point (e).
Data processing and plotting were performed using dplyr and ggplot2 packages (Wickham Reference Wickham2016; Wickham et al. Reference Wickham, François, Henry, Müller and Vaughan2025).
Results and Discussion
Fertilizer Study
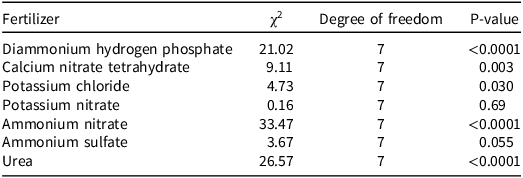
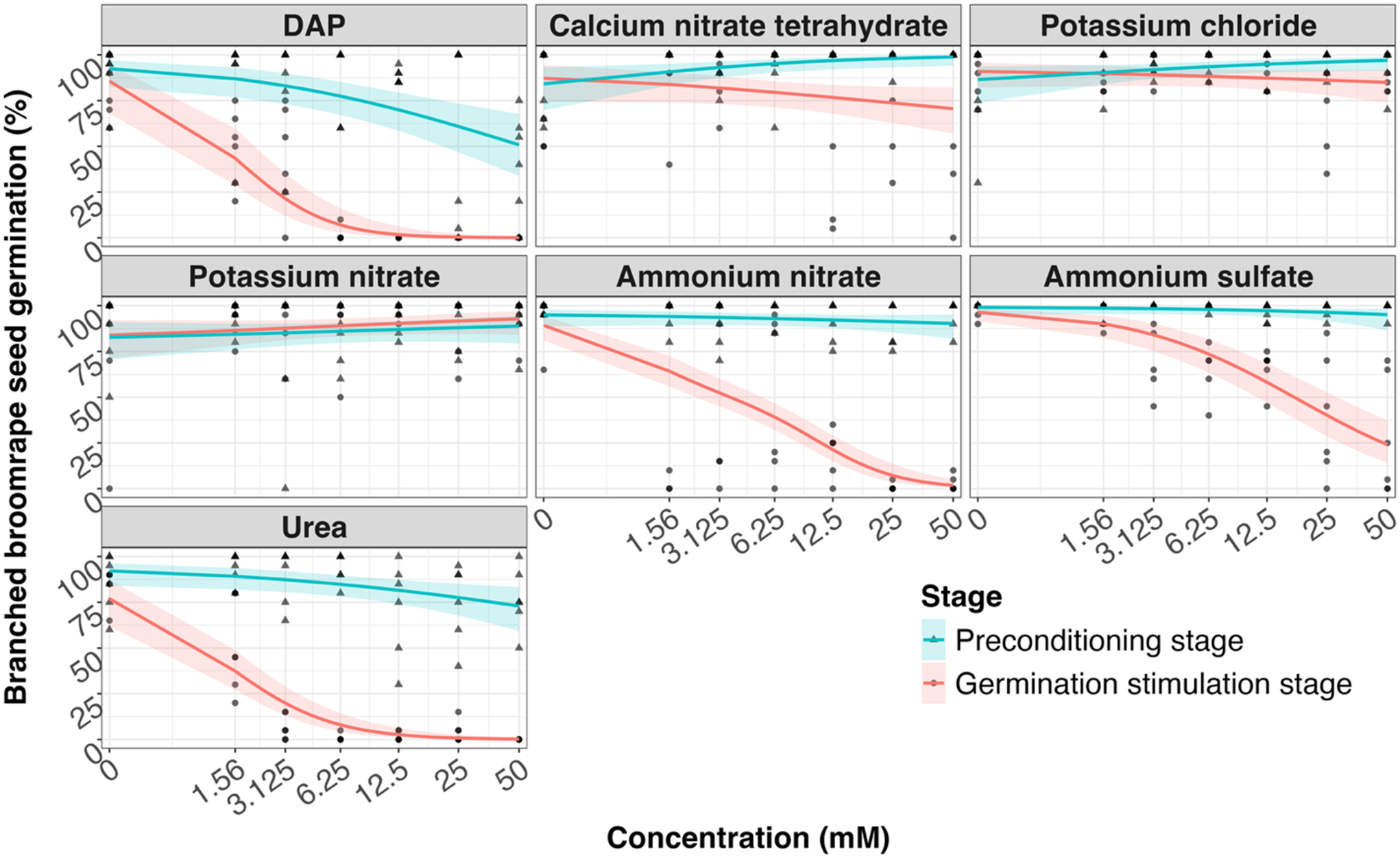
The average germination rate of plants in the control plots in the fertilizer study was 86%. The radicle lengths of the untreated branched broomrape seedlings ranged from 0.5 mm to 1.5 mm (data not shown). A likelihood ratio test comparing models with and without the interaction between stage and concentration indicated that the response differed between the preconditioning stage and the germination stimulation stage for several fertilizers (Table 1). The interaction significantly improved the model fit for diammonium hydrogen phosphate (χ2 = 21.02, P < 0.0001), calcium nitrate tetrahydrate (χ2 = 9.11, P = 0.003), potassium chloride (χ2 = 4.73, P = 0.03), ammonium nitrate (χ2 = 33.47, P < 0.0001), and urea (χ2 = 26.57, P < 0.0001). Among the fertilizers, the trials with calcium nitrate and potassium chloride a slight increase in seed germination occurred when treatments were applied at the preconditioning stage, whereas diammonium hydrogen phosphate, ammonium nitrate, and urea treatments resulted in a dramatic decline in broomrape germination as their concentrations increased (Figure 3). The interaction was marginal for ammonium sulfate (χ2 = 3.67, P = 0.055), and there was no interaction when potassium nitrate was applied (χ2 = 0.16, P = 0.69). When fertilizers were applied during the preconditioning stage, none had negative effects on seed germination.
Likelihood ratio tests for branched broomrape seed germination, comparing beta-binomial generalized linear mixed models with and without the stage × concentration interaction.

Table 1. Long description
The table presents likelihood ratio tests for branched broomrape seed germination, comparing beta-binomial generalized linear mixed models with and without the stage and concentration interaction. It includes data for eight fertilizers: diammonium hydrogen phosphate, calcium nitrate tetrahydrate, potassium chloride, potassium nitrate, ammonium nitrate, ammonium sulfate, and urea. The table has eight rows and four columns. The columns are labeled Fertilizer, Chi-squared, Degree of freedom, and P-value. Each row lists a different fertilizer along with its corresponding chi-squared value, degree of freedom, and p-value. Notable trends include significant interactions for diammonium hydrogen phosphate, calcium nitrate tetrahydrate, potassium chloride, ammonium nitrate, and urea, indicating that the response differed between the preconditioning stage and the germination stimulation stage. The interaction was marginal for ammonium sulfate and non-significant for potassium nitrate.
Relationship between fertilizer concentration (0–50 mM) and branched broomrape seed germination (%), shown separately for each fertilizer. Points show observed germination at each concentration; symbols indicate experimental stage (preconditioning stage vs. germination stimulation stage). Solid lines show predictions from generalized linear mixed models fitted separately for each fertilizer (beta-binomial with logit link) with fixed effects of stage, log-transformed concentration, their interaction, and experiment run, and a random effect for replication. Shaded ribbons indicate 95% confidence intervals around the predictions. Concentration is displayed on a pseudo-log x-axis to include 0 mM. Abbreviation: DAP, diammonium hydrogen phosphate.

Figure 3. Long description
The image contains six line graphs showing the relationship between fertilizer concentration and branched broomrape seed germination for different fertilizers. Each graph represents a different fertilizer: DAP, Calcium nitrate tetrahydrate, Potassium chloride, Potassium nitrate, Ammonium nitrate, and Urea. The x-axis represents the concentration in millimolar (mM) on a pseudo-log scale, ranging from 0 to 50 mM. The y-axis represents the percentage of branched broomrape seed germination, ranging from 0 to 100 percent. Points on the graphs show observed germination at each concentration, with symbols indicating the experimental stage: preconditioning stage (blue triangles) and germination stimulation stage (red circles). Solid lines show predictions from generalized linear mixed models fitted separately for each fertilizer, with shaded ribbons indicating 95 percent confidence intervals around the predictions. The graphs illustrate how different fertilizers affect seed germination at varying concentrations.
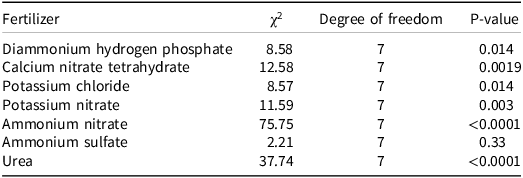
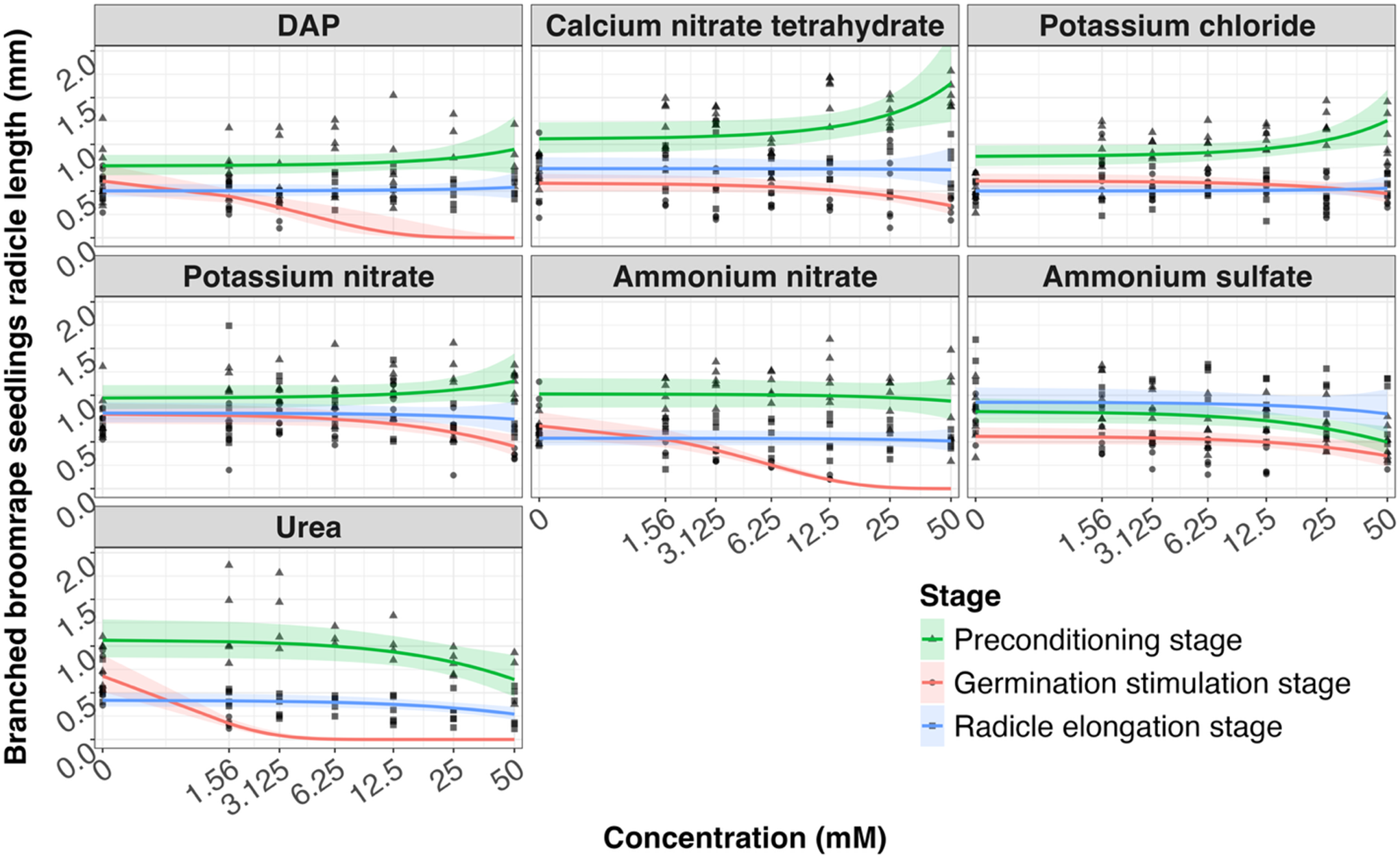
For branched broomrape radicle length, likelihood ratio tests were conducted to compare gamma GLMMs with and without the interaction between stage and concentration. The patterns differed among stages for several fertilizers as follows: diammonium hydrogen phosphate (χ2 = 8.58, P = 0.0137), calcium nitrate (χ2 = 12.58, P = 0.0019), potassium chloride (χ2 = 8.57, P = 0.0138), potassium nitrate (χ2 = 11.59, P = 0.0030), ammonium nitrate (χ2 = 75.75, P < 0.0001), and urea (χ2 = 37.74, P < 0.0001). The interaction was not supported for ammonium sulfate (χ2 = 2.21, P = 0.331) (Table 2). In terms of biological response, only diammonium hydrogen phosphate, ammonium nitrate, and urea inhibited radicle elongation when they were applied during the germination stimulation stage (Figure 4). The shortest radicle length was observed when 50 mM ammonium nitrate was applied (0.26 mm), compared with an untreated radicle length of approximately 0.5 mm to 1 mm, indicating at least a 50% reduction. When fertilizers were applied at the preconditioning stage or after germination had initiated (i.e., to emerged radicles), none of the seven fertilizers resulted in visibly observable damage to radicles or inhibited elongation (Figure 4).
Likelihood ratio tests for branched broomrape seed radicle length, comparing gamma generalized linear mixed models with and without the stage × concentration interaction.

Table 2. Long description
The table presents likelihood ratio tests for branched broomrape seed radicle length, comparing gamma generalized linear mixed models with and without the interaction between stage and concentration. It includes data for seven fertilizers: diammonium hydrogen phosphate, calcium nitrate tetrahydrate, potassium chloride, potassium nitrate, ammonium nitrate, ammonium sulfate, and urea. Each row lists the chi-squared value, degree of freedom, and p-value for the respective fertilizer. Notable trends include significant interactions for diammonium hydrogen phosphate, calcium nitrate tetrahydrate, potassium chloride, potassium nitrate, ammonium nitrate, and urea, with p-values indicating strong statistical significance. Ammonium sulfate shows no significant interaction with a p-value of 0.33. The table highlights that diammonium hydrogen phosphate, ammonium nitrate, and urea inhibit radicle elongation when applied during the germination stimulation stage, with ammonium nitrate showing the shortest radicle length at 0.26 millimeters when applied at 50 millimolar concentration.
Relationship between fertilizer concentration (0–50 mM) and radicle length (mm) for germinated branched broomrape seeds only (length >0), shown separately for each fertilizer. Points show observed radicle lengths; symbols indicate stage (preconditioning stage, germination stimulation stage, and radicle elongation stage). Solid lines show predictions from gamma generalized linear mixed models fitted separately for each fertilizer, with fixed effects of stage, concentration, and their interaction, and a random effect for replication. Shaded ribbons indicate 95% confidence intervals around the predictions. Concentration is displayed on a pseudo-log x-axis to include 0 mM. Abbreviation: DAP, diammonium hydrogen phosphate.

Figure 4. Long description
The image contains nine line graphs, each representing a different fertilizer: DAP, Calcium nitrate tetrahydrate, Potassium chloride, Potassium nitrate, Ammonium nitrate, and Urea. The x-axis shows the concentration in millimolar (mM) on a pseudo-log scale ranging from 0 to 50, while the y-axis shows the radicle length in millimeters (mm). Each graph includes points indicating observed radicle lengths and solid lines representing predictions from gamma generalized linear mixed models. The lines are color-coded to indicate different stages: green for the preconditioning stage, red for the germination stimulation stage, and blue for the radicle elongation stage. Shaded ribbons around the lines indicate 95% confidence intervals. The graphs illustrate how radicle length varies with different concentrations of each fertilizer and across different stages of germination. All values are approximated.
Westwood and Foy (Reference Westwood and Foy1999) reported that radicle length was reduced by about half after branched broomrape and Egyptian broomrape were exposed to 25 mM ammonium sulfate after only 4 to 8 h. In contrast, in the current study, ammonium sulfate had no significant negative effect on radicle elongation when it was applied at any of the three stages.
The results of the fertilizer study indicated that diammonium hydrogen phosphate, ammonium nitrate, and urea inhibited branched broomrape seed germination and seedling radicle elongation only when the fertilizers were applied at the germination stimulation stage. Once the radicle started elongating (5 d after exposure to the germination stimulants), there was no apparent inhibition of radicle elongation from any of the seven fertilizers.
Flooding Study
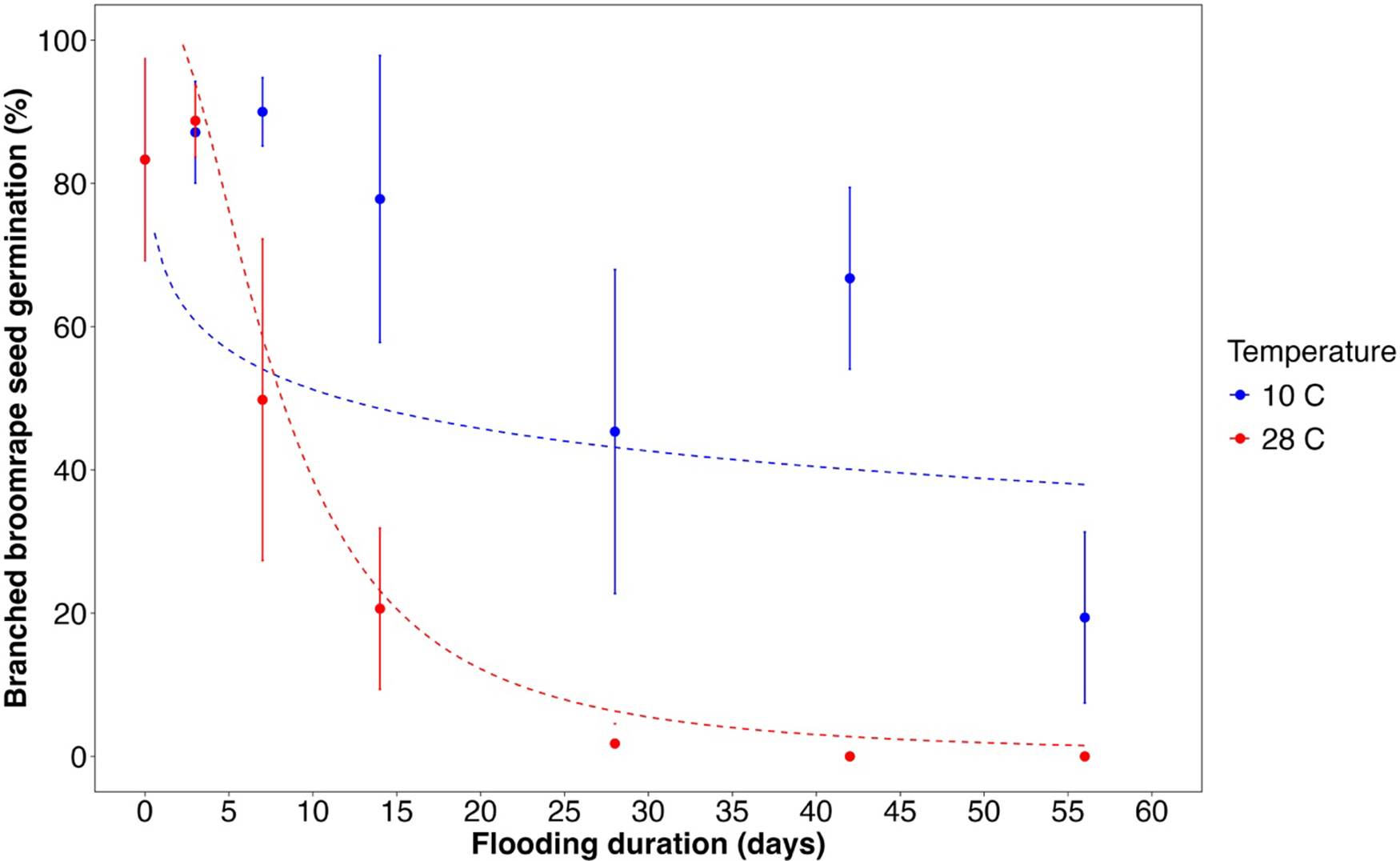
Seed germinability of branched broomrape was substantially reduced with flooding, with the magnitude of suppression depending on both flooding duration and temperature (Figure 5). The two temperatures, 10 C and 28 C, were intended to correspond to two potential flood timings in northern California: winter flooding with no crop, and summer flooding with rice in a crop rotation. The laboratory experiment data indicated that branched broomrape seed germination declined sharply as the flooding duration increased, and this effect is more substantial under warm-temperature conditions (Figure 5, Table 3). During short flooding periods (3 d and 7 d), seed germinability was unaffected. However, as the flooding period extended to 14 d or 28 d, germination rates decreased substantially at both temperature regimes. The 56-d flooding treatments at both temperature regimes almost eliminated subsequent broomrape seed germination.
Effect of flooding duration and temperature on branched broomrape seed germination under laboratory conditions. A three-parameter logistic model (
 $Y = \;{u \over {\left\{ {1 + \exp \left[ {b\left( {log\left( x \right) - \log \left( e \right)} \right)} \right]} \right\}}}$
) was fit to all data. Dotted lines are fitted values, and solid circles indicate observed germination (n = 6). Error bars indicate 95% confidence intervals.
$Y = \;{u \over {\left\{ {1 + \exp \left[ {b\left( {log\left( x \right) - \log \left( e \right)} \right)} \right]} \right\}}}$
) was fit to all data. Dotted lines are fitted values, and solid circles indicate observed germination (n = 6). Error bars indicate 95% confidence intervals.

Figure 5. Long description
A line graph showing the effect of flooding duration and temperature on branched broomrape seed germination under laboratory conditions. The x axis represents flooding duration in days, ranging from 0 to 60 days. The y axis represents branched broomrape seed germination in percentage, ranging from 0 to 100 percentage. Two data series are plotted: one for a temperature of 10 degrees Celsius and another for 28 degrees Celsius. The blue line represents the data for 10 degrees Celsius, and the red line represents the data for 28 degrees Celsius. Dotted lines indicate fitted values, and solid circles indicate observed germination with error bars showing 95 percentage confidence intervals. The graph shows that as flooding duration increases, seed germination decreases more rapidly at 28 degrees Celsius compared to 10 degrees Celsius. All values are approximated.
Estimated parameter values for the three-parameter log-logistic models used to describe the branched broomrape seed germination responses to flooding duration and temperature under laboratory conditions.

Table 3. Long description
The table presents estimated parameter values for the three-parameter log-logistic models used to describe branched broomrape seed germination responses to flooding duration and temperature under laboratory conditions. It includes four rows and five columns. The columns are labeled Parameter, Estimate, Standard error, t-value, and P-value. The parameters are measured at two different temperatures, 10 degrees Celsius and 28 degrees Celsius. The table shows the estimates, standard errors, t-values, and P-values for each parameter at these temperatures. Notable trends include higher estimates and t-values at 28 degrees Celsius compared to 10 degrees Celsius, indicating a more substantial effect of flooding duration on seed germination at higher temperatures.
a In the model, b is the slope at the inflection point; u is the upper limit; and e is the concentration that produces a germination response half the u value (ED50).
b Data from 10 C and 28 C flooding were separated into two models.
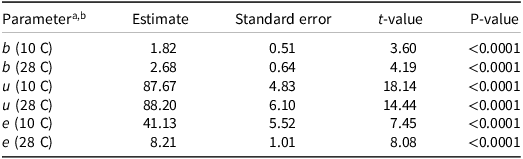
A similar pattern was observed in the pot study with host plants. Flooding duration of more than 14 d resulted in a dramatic decrease in the number of branched broomrape attachments on tomato plants at both flooding temperature regimes (Figure 6, Table 4). In the flooding regime at 28 C, the number of branched broomrape attachments dropped sharply after 7 d of flooding. Overall, these results indicated that flooding may suppress branched broomrapes by preventing seed germination and, ultimately, reducing attachment to subsequently planted tomatoes.
Effect of flooding duration and temperature on branched broomrape seed attachments to the host crop. A three-parameter logistic model (
 $Y = \;{u \over {\left\{ {1 + \exp \left[ {b\left( {log\left( x \right) - \log \left( e \right)} \right)} \right]} \right\}}}$
) was fit to all data. Dotted lines are fitted values, and solid circles indicate observed germination (n = 6). Error bars indicate 95% confidence interval.
$Y = \;{u \over {\left\{ {1 + \exp \left[ {b\left( {log\left( x \right) - \log \left( e \right)} \right)} \right]} \right\}}}$
) was fit to all data. Dotted lines are fitted values, and solid circles indicate observed germination (n = 6). Error bars indicate 95% confidence interval.

Figure 6. Long description
The line graph illustrates the effect of flooding duration and temperature on branched broomrape seed attachments to the host crop. The x-axis represents the flooding duration in days, ranging from 0 to 60 days. The y-axis represents the number of branched broomrape attachments, ranging from 0 to 20. Two data series are presented: one for a temperature of 10 degrees Celsius, indicated by blue circles and a dashed blue line, and another for a temperature of 28 degrees Celsius, indicated by red circles and a dashed red line. The dotted lines represent fitted values using a three-parameter logistic model, while the solid circles indicate observed germination with error bars showing the 95% confidence interval. The graph shows that as the flooding duration increases, the number of branched broomrape attachments decreases for both temperatures, with a more rapid decline observed at 28 degrees Celsius compared to 10 degrees Celsius. All values are approximated.
Estimated parameter values for the three-parameter log-logistic models used to describe the number of branched broomrape attachments to a single host plant, as a function of flooding duration and temperature.

Table 4. Long description
The table presents estimated parameter values for the three-parameter log-logistic models used to describe the number of branched broomrape attachments to a single host plant. The table includes five parameters measured at two different temperatures, 10 degrees Celsius and 28 degrees Celsius. Each parameter has an estimate, standard error, t-value, and P-value. The parameters are b, u, and e, each measured at the two temperatures. Notable trends include higher estimates and t-values for parameters measured at 28 degrees Celsius compared to 10 degrees Celsius, indicating a more significant impact of flooding duration at higher temperatures.
a In the model, b is the slope at the inflection point; u is the upper limit; and e is the duration that produces the number of attachment half the u value (ED50).
b Data from 10 C and 28 C flooding were separated into two models.
Practical Implications
In the processing tomato industry in Sacramento Valley, the most common fertilization regime is to apply a starter liquid nitrogen fertilizer before transplanting the crop, followed by three to four injections of UAN-32, which includes ammonium nitrate and urea, through the subsurface drip irrigation system over the growing season (Aegerter et al. Reference Aegerter, Lazicki, Miyao, Stewart, Murdock and Goodrich2023). The results from the fertilizer study indicated that branched broomrape seed is susceptible to low concentrations of diammonium hydrogen phosphate, ammonium nitrate, and urea within a relatively narrow application window during broomrape germination and radicle elongation. This suggests that increasing the number of injections and reducing the amount of fertilizer in each injection may affect branched broomrape parasitism. The other component of the nitrogen fertilizer duality effect, which affects strigolactone exudation from the host, was not studied in this experiment. Moreover, another challenge that may hinder field application is that positively charged ammonium readily binds to soil particles, which may reduce the likelihood of contact with branched broomrape seeds. Therefore, the effectiveness of nitrogen fertilization to control branched broomrape may be variable under field conditions, and field trials are needed to determine application rates and frequencies throughout the season.
Results from the flooding experiment suggested that flooding duration and temperature strongly influenced branch broomrape germinability and attachment to the host in both laboratory and greenhouse assays. In the pot experiment, cool-temperature flooding for longer periods had a similar suppression effect on branched broomrape seedling attachments, compared with warm-temperature flooding, suggesting that winter flooding could have practical value. From a grower perspective, winter flooding may be more feasible because it can be implemented during fallow periods with minimal risk to the cash crop, potentially at a lower opportunity cost than summer flooding, which is often tied to rice rotations and occurs when water demand and operational constraints can be higher and is not feasible in all tomato production areas (Hill et al. Reference Hill, Williams, Mutters and Greer2006). The mechanism of flooding inhibition on broomrape seed germinability remains unknown and may be associated with anaerobic soil conditions that disrupt seed metabolism (Conte et al. Reference Conte, Morrissey, Hanson and Simmons2026). The overall results suggested that refining nutrient management and irrigation programs might be the nonchemical methods growers can use alongside chemical controls to manage branched broomrape.
Acknowledgments
We gratefully thank Magdalene Burgess and Ni Tang for their assistance.
Funding
This research was supported by funding from the California Tomato Research Institute, the California League of Food Processors, and the California Department of Food and Agriculture Specialty Crop Block Grant Program (CDFA#22-0001-031-SF).
Competing Interests
The authors declare they have no conflicts of interest.





 Open access
Open access