


1. Introduction
As the survival rate of preterm infants increases, it is crucial to address their long-term neurodevelopmental outcomes (Alyward, Reference Aylward2014; Baralt & Darcy Mahoney, Reference Baralt and Darcy Mahoney2020; Hack, Reference Hack2007; Hee Chung et al., Reference Hee Chung, Chou and Brown2020; Soleimani et al., Reference Soleimani, Zaheri and Abdi2014; Inder et al., Reference Inder, Volpe and Anderson2023). The early years of life, particularly the period between birth and five years old, are marked by intense and rapid neuronal growth. However, due to shortened gestation and perinatal complications, many extremely preterm infants experience deficits in psychomotor, cognitive and attentional functions, as well as emotional vulnerability and underperformance in school (e.g., Hee Chung et al., Reference Hee Chung, Chou and Brown2020). Research suggests that these challenges stem from a central deficit in frontal lobe processing of complex information (Als et al., Reference Als, Duffy, McAnulty, Rivkin, Vajapeyam, Mulkern and Eichenwald2004). Children born prematurely are at a higher risk for disorders such as autism, attention-deficit disorder (ADD), attention-deficit hyperactivity disorder (ADHD) and impaired executive function (EF) (Hack, Reference Hack2007; Pettinger et al., Reference Pettinger, Copper, Boyle, Blower, Hewitt and Fraser2023).
1.1. Executive function and prematurity
EF refers to specific skills such as attention, emotional control, memory, information processing, cognitive flexibility, inhibitory control, problem solving and planning (Carlson et al., Reference Carlson, Zelazo, Faja and Zelazo2013; Zelazo & Carlson, Reference Zelazo and Carlson2020). The interrelated processes that make up EF are responsible for a child’s cognitive functioning, behavior, social interaction and emotional control. Inhibitory control, the capacity to suppress automatic or prepotent responses and filter out distracting information, is particularly critical for self-regulation and adaptive behavior. Deficits in EF, and inhibitory control in particular, can adversely affect a child’s academic readiness, social interactions and emotional stability. In preterm-born populations, these vulnerabilities are especially consequential: evidence from large-scale studies demonstrates that EF deficits associated with preterm birth may persist into adolescence and adulthood and are linked to long-term educational and functional outcomes (e.g., Kroll et al., Reference Kroll, Karolis, Brittain, Tseng, Froudist-Walsh, Murray and Nosarti2017; Mulder et al., Reference Mulder, Pitchford, Hagger and Marlow2009; Réveillon et al., Reference Réveillon, Borradori Tolsa, Monnier, Hüppi and Barisnikov2016; Sandoval et al., Reference Sandoval, Gaspardo and Linhares2022; Sun & Buys, Reference Sun and Buys2012). The relevance of these deficiencies lies in the way in which they affect a young child’s school readiness. Given the central role of EF in early learning and its enduring developmental consequences, it is essential to advance theoretical and empirical research focused on EF development in children born preterm.
1.2. Bilingualism as a potential enhancer of inhibitory control
A growing body of research indicates that bilingualism is associated with enhanced EF skills, including superior inhibitory control (Bialystok & Martin, Reference Bialystok and Martin2004; Gonzalez-Barrero & Nadig, Reference Gonzalez-Barrero and Nadig2019; Head et al., Reference Head, Baralt and Darcy Mahoney2015). Inhibitory control is a complex cognitive function that relies on a network of cortical regions, such as the inferior frontal cortex and pre-supplementary motor area, as well as subcortical structures, including the subthalamic nucleus and putamen (Aron & Poldrack, Reference Aron and Poldrack2006). Notably, research has demonstrated that term-born bilingual children consistently outperform their monolingual peers on nonlinguistic tasks that require filtering out distractions and resolving perceptual conflicts, both critical components of inhibitory control (e.g., Brito & Barr, Reference Brito and Barr2012; Poulin-Dubois et al., Reference Poulin-Dubois, Blaye, Coutya and Bialystok2011).
The theoretical rationale behind this proposed advantage is that bilinguals constantly engage in “intense processing” to monitor and control interference from simultaneously-active language systems, thereby enhancing the neural networks underpinning inhibitory control (Bialystok, Reference Bialystok2015). Neuroimaging research further supports this view: bilingual children and adults often demonstrate more efficient recruitment of frontal lobe regions associated with inhibition and conflict resolution (Abutalebi et al., Reference Abutalebi, Della Rosa, Green, Hernandez, Scifo, Keim, Cappa and Costa2012; Li et al., Reference Li, Wu, Yang, Xie, Chang and Luo2023; Luk et al., Reference Luk, Bialystok, Craik and Grady2011). In addition, research with infants suggests that exposure to multiple languages may accelerate the maturation of inhibitory control processes. For instance, Kovács and Mehler (Reference Kovács and Mehler2009) found that bilingual infants as young as seven months outperformed their monolingual peers on tasks requiring the inhibition of a dominant response. Further supporting this idea, subsequent studies have shown that bilingual infants and children exhibit heightened neural activation in regions linked to inhibitory control when engaged in tasks demanding conflict resolution (Engel de Abreu et al., Reference Engel de Abreu, Cruz-Santos, Tourinho, Martin and Bialystok2012; Morales et al., Reference Morales, Gómez-Ariza and Bajo2013).
1.3. Bilingualism, executive function and neural recruitment in preterm-born children
Given the vulnerability of preterm children to deficits in EF, a key question emerges: can bilingualism serve as a protective or enhancing factor for EF in this at-risk population? Both theoretical scholarship and emerging empirical evidence support this possibility (Baralt & Darcy Mahoney, Reference Baralt and Darcy Mahoney2020; Darcy Mahoney & Baralt, Reference Darcy Mahoney and Baralt2016; Gillenson et al., Reference Gillenson, Bagner, Darcy Mahoney and Baralt2023; Head et al., Reference Head, Baralt and Darcy Mahoney2015). In one of the first studies to examine this question, Darcy Mahoney and Baralt (Reference Darcy Mahoney and Baralt2016) examined inhibitory control in a sample of 22 preterm-born children using the Opposite Worlds task. Bilingual children demonstrated stronger performance on this task, suggesting an enhanced ability to suppress prepotent responses and flexibly shift attentional sets. Building on this initial work, Baralt and Darcy Mahoney (Reference Baralt and Darcy Mahoney2020) evaluated EF in 35 preterm-born children, comparing bilingual and monolingual groups. Results indicated that bilingual preterm-born children performed significantly better than their monolingual peers on the Flanker task, further supporting the hypothesis that bilingual experience may enhance inhibitory control in this population. More recently, Gillenson et al. (Reference Gillenson, Bagner, Darcy Mahoney and Baralt2023) assessed EF in bilingual and monolingual preterm-born children using a task from the Test of Everyday Attention for Children (TEA-Ch). Results indicated that bilingual preterm-born children outperformed their monolingual peers on measures of attentional control and cognitive flexibility. To the best of our knowledge, the only other study that explored a possible bilingual advantage in EF among preterm-born children is that by Loe and Feldman (Reference Loe and Feldman2016). They found no benefits of bilingualism on preterm children’s task performances measuring EF. However, their null findings may reflect methodological differences in the operationalization of bilingualism. In their study, bilingualism was defined broadly based on parent-reported language exposure, without verification of productive language use or proficiency. Subsequent work has demonstrated that imprecise definitions of bilingualism can obscure true group differences in EF outcomes (e.g., Baralt & Darcy Mahoney, Reference Baralt and Darcy Mahoney2020; Yurtsever et al., Reference Yurtsever, Anderson and Grundy2023), and that productive capacity in both languages may be the required threshold in order to reap the benefits of bilingualism (e.g., Baralt & Darcy Mahoney, Reference Baralt and Darcy Mahoney2020).
Taken together, the work summarized so far provides the first converging empirical evidence that bilingualism may support executive functioning in preterm-born children, particularly in domains of inhibitory control and attentional regulation. The findings also align with prior research in term-born populations, where bilingualism has been linked to enhanced cognitive flexibility and conflict resolution (e.g., Bialystok & Viswanathan, Reference Bialystok and Viswanathan2009). We therefore hypothesize that during early childhood – when neural systems supporting EF remain highly plastic – bilingual experience may act as a formative input that both enhances behavioral inhibitory control and promotes more efficient recruitment of the frontal networks that underlie EF in preterm-born children (Baralt & Darcy Mahoney, Reference Baralt and Darcy Mahoney2020).
To understand this relationship, it is essential to go beyond behavioral outcomes and examine the neural mechanisms that underlie bilingual effects on EF. Strong EF skills are foundational for academic success, social competence and overall well-being, which lead to successful and healthy lives. However, their development is deeply tied to neural processes, particularly in the frontal lobe regions responsible for inhibitory control. Brain imaging technologies offer a means to investigate how bilingual and monolingual preterm-born children recruit these neural networks differently. One promising tool for this investigation is functional near-infrared spectroscopy (fNIRS). This noninvasive imaging technique is ideal for studying young children because it allows for natural, active participation without requiring immobilization (Buss et al., Reference Buss, Fox, Boas and Spencer2014; Moriguchi & Hiraki, Reference Moriguchi and Hiraki2013). Recent research has increasingly employed fNIRS to explore how children neurally recruit EF (Buss et al., Reference Buss, Fox, Boas and Spencer2014; Fishburn et al., Reference Fishburn, Hlutkowsky, Bemis, Huppert, Wakschlag and Perlman2019), providing valuable insights into the brain activity underlying inhibitory control. Applying fNIRS to preterm-born bilingual children enables investigation of the neural mechanisms through which bilingual experience may shape EF, beyond what can be inferred from behavioral performance alone.
Building on this, the present study examines whether bilingual experience is associated with enhanced inhibitory control and more efficient neural recruitment in preterm-born children using a robust, triangulated assessment of bilingual competence (following Baralt & Darcy Mahoney, Reference Baralt and Darcy Mahoney2020). Inhibitory control was assessed behaviorally using a Go/No-Go task while simultaneous fNIRS measured frontal lobe activation associated with response inhibition. The study had two primary objectives: (1) to determine whether bilingual preterm-born children demonstrate superior inhibitory control relative to monolingual peers and (2) to characterize the differences in the temporal dynamics of hemodynamic responses underlying inhibitory control.
2. Methods
2.1. Participants
An original pool of 23 preterm-born children, aged between six and seven years, were recruited for the study to obtain both behavioral and fNIRS data. Simultaneous collection of the fNIRS and behavioral data was not completed in five participants because of attention span and gross motor skill challenges due to cerebral palsy, ADD and ADHD, or because of excess movement-related step-like noise during the tasks that rendered the fNIRS data unreadable. fNIRS data collection failed for two participants, so they were excluded from the study. Data are reported on from 16 out of the 23 participants recruited.
To minimize potential sources of variability, the following recruitment and eligibility criteria were employed: (1) all participants were drawn from a single hospital system to ensure consistency in neonatal care practices; (2) only right-handed children were included to control for potential confounding effects of hand dominance on task performance (Cuzzocreo, Reference Cuzzocreo, Yassa, Verduzco, Honeycutt, Scott and Bassett2009; see also Papadatou-Pastou et al., Reference Papadatou-Pastou, Ntolka, Schmitz, Martin, Munafò, Ocklenburg and Paracchini2020) and language lateralization (Szaflarski et al., Reference Szaflarski, Binder, Possing, McKiernan, Ward and Hammeke2002, Reference Szaflarski, Rajagopal, Altaye, Byars, Jacola, Schmithorst, Schapiro, Plante and Holland2012) and (3) each group was age- and acuity-matched based on key variables such as gestational age, birth weight, neonatal health conditions (e.g., intraventricular hemorrhage [IVH], retinopathy of prematurity, necrotizing enterocolitis) and length of stay in the neonatal intensive care unit (NICU) (Baralt & Darcy Mahoney, Reference Baralt and Darcy Mahoney2020; Darcy Mahoney & Baralt, Reference Darcy Mahoney and Baralt2016; Scharoun & Bryden, Reference Scharoun and Bryden2014).
Following prior work demonstrating that imprecise definitions of bilingualism may obscure EF differences (e.g., Yurtsever et al., Reference Yurtsever, Anderson and Grundy2023), bilingual status in the present study was operationalized using a multi-method approach emphasizing productive language ability. Specifically, bilingual classification followed the four-step procedure described by Baralt and Darcy Mahoney (Reference Baralt and Darcy Mahoney2020), ensuring that all children classified as bilingual demonstrated functional proficiency in both English and Spanish. These triangulated measures were: (1) responses to the Language Background Questionnaire (Gathercole et al., Reference Gathercole, Thomas, Roberts, Hughes, Hughes and Mueller Gathercole2013); (2) consistent responses in Spanish when addressed in Spanish as reported by parents; (3) positive attitudes toward Spanish as reported by both parents and children and (4) an interview conducted in Spanish with the child by a bilingual researcher. Additionally, all participants completed the Peabody Picture Vocabulary Test–Fourth Edition (PPVT-4; Dunn & Dunn, Reference Dunn and Dunn2007), and bilingual children also completed the Test de Vocabulario en Imágenes Peabody (TVIP; Dunn, Padilla, Lugo & Dunn, Reference Dunn, Padilla, Lugo and Dunn1986).
This level of methodological precision aligns with recent recommendations by Yurtsever et al. (Reference Yurtsever, Anderson and Grundy2023), whose meta-analysis underscores that insufficiently defined bilingual samples can obscure true group differences in EF outcomes. For full methodological details, please see Baralt and Darcy Mahoney (Reference Baralt and Darcy Mahoney2020).
2.1.1. Behavioral task stimuli and experimental design
The inhibition component of EF was measured with the Go/No-Go task, which assesses inhibitory control by asking children to respond quickly to frequent “Go” stimuli and to withhold responses on infrequent “No-Go” trials. Following the computerized Go/No-Go task for children by Fishburn et al. (Reference Fishburn, Hlutkowsky, Bemis, Huppert, Wakschlag and Perlman2019), the task was displayed on a computer monitor and showed cartoon images of tigers, elephants and giraffes. Participants were instructed to press the button as quickly as possible whenever they saw an image of a tiger or an elephant (the “Go” stimuli). However, they were told to refrain from pressing the button whenever they saw the giraffe (the “No-Go” stimulus). For integration with fNIRS, the task used a block design with six main blocks, each comprising 24-second stimulus phases (alternating between “Go” and “No-Go” conditions with three-second instruction periods) and a “Control” block during which children sat quietly. In the “Go” block, 12 images of elephants and 12 of tigers were randomly presented at a rate of 1 Hz (0.8 seconds display plus 0.2 seconds transition), while in the “No-Go” block, 12 tiger images and 12 giraffe images were shown, with behavioral accuracy and reaction time (RT) recorded from button responses. Participants’ responses during the Go/No-Go trials were accurately recorded using custom software. This software captures two key time stamps for each stimulation: the stimulus onset time (ts), which indicates when the image appears, and the button press time (tp), which marks when the participant responds. From each participant’s response record, a separate piece of software was used to calculate the RT (RT = tp−ts) and to assess the accuracy (success or failure) of its response to the corresponding stimulus. These two pieces of information were gathered from all participants and analyzed collectively. A schematic of the task structure is shown in Figure 1.
Block diagram illustrating the Go/No-Go study procedure. Note: Each control block (blue) is 24 seconds long. Each trial block (yellow) contains one Go (green) and one No-Go (orange) segments, with both lasting 24 seconds. The instruction blocks (black) within each Trial block last 8 seconds. Within the Go block, the tiger and elephant images are shown randomly for 0.8 s each, separated by 0.2 s transitions, for a total of 24 stimuli. Similarly, during the No-Go block, the tiger, elephant and giraffe (“No-Go”) images alternate randomly in the same timing pattern.

Figure 1. Long description
At the top, a horizontal sequence alternates blue CONTROL blocks and six yellow TRIAL blocks labeled Trial 1 through Trial 6. Below, a bracket groups the first three trials, leading to a detailed breakdown. Each trial contains an instruction segment (black rectangle labeled INST.), followed by either a green GO block or an orange NO-GO block. The GO block contains a grid of alternating tiger and elephant cartoon faces, while the NO-GO block contains a grid of tiger, elephant, and giraffe faces. The GO block is green and the NO-GO block is orange, matching their respective segment colors. The structure visually emphasizes the sequence: instruction, then Go or No-Go, with animal stimuli presented in randomized order within each block.
2.1.2. fNIRS data acquisition and processing
During the Go/No-Go task, brain activity data from each participant was collected using an fNIRS system (NIRx Scout 16–24, https://nirx.net/). To fit the head size of the participants and hence improve the signal quality, a child-sized cap was utilized in conjunction with the fNIRS system. The cap contained 8 sources and 7 detectors spaced approximately 1.5 centimeters apart, which were placed based on the international 10–20 coordinate system for frontal lobe investigation (Supplementary Figure 1). This combination of sources and detectors enabled the investigation of 22 unique local brain areas, referred to hereafter as “channels.”
In each recording session, which lasted about 450 seconds, the fNIRS system continuously recorded the responses of all detectors to two different illumination wavelengths (760 nm and 850 nm) at a sampling rate of approximately 4 Hz. During this period, a separate piece of software called NIRxStim (from NIRX Medical Technologies, Glen Head, NY) was used to insert time stamps marking the beginning and end of each activity type. These time stamps were essential for organizing and segmenting the data in the subsequent fNIRS data analysis.
The raw fNIRS data from each channel of a study subject were preprocessed to remove motion artifacts and then converted to the ∆OD signals using the NIRS toolbox developed by Santosa et al. (Reference Santosa, Zhai, Fishburn and Huppert2018). Next, the ∆OD signals were converted to relative changes in oxygenated hemoglobin (HbO) and deoxygenated hemoglobin (HbR) concentrations (i.e., ∆[HbO] and ∆[HbR]) using modified Beer–Lambert law. The ∆[HbO] and ∆[HbR] signals were segmented into several subsections based on the activity label. Subsections with the same activity label were collected and averaged, resulting in the averaged hemodynamic responses (i.e., Δ[HbO]mean and Δ[HbR]mean) for a given activity. In order to quantify the level of brain activities, Δ[HbO]mean and Δ[HbR]mean were fitted to the canonical hemodynamic response functions (cHRF, see Figure 2) using a general linear model (Santosa et al., Reference Santosa, Zhai, Fishburn and Huppert2018). Since the fitting coefficient is also considered as the “magnitude of brain activity,” it was used in the subsequent statistical analyses. This process was applied to the fNIRS raw data from all channels of all participants of the study; see Figure 2.
Segmentation of the processed fNIRS signal recorded from a single channel. The x-axis displays colored blocks representing various experimental conditions: “CL” (baseline) is shown at both the beginning and end of the recording session, while “GO” (Go trial) and “NG” (No-Go trial) alternate in 24-second blocks, with each block followed by an 8-second instruction period (shown in black).

Figure 3. Long description
The x-axis is labeled Time in seconds, ranging from 0 to 450. The y-axis is labeled delta Conc. in milli Molar per centimeter, spanning negative 5 to positive 4. Along the x-axis, colored blocks indicate experimental conditions: cyan for C L at the start and end, alternating green for G O and orange for N G, each separated by thin black blocks representing instruction periods. Above these blocks, two lines are plotted: a red line for H b O and a blue line for H b R, as indicated by the legend in the upper right. Both lines fluctuate with multiple peaks and troughs, generally increasing toward the right. Peaks in both lines often align with transitions between G O and N G blocks. The red H b O line consistently trends higher than the blue H b R line throughout the session.
2.1.3. Statistical analyses: Behavioral data
From the RT and Correct/Incorrect records of each participant, the average RT (RTave) and accuracy for both Go and No-Go stimuli were calculated. RTave was determined by considering only those RTs associated with “Correct” responses. Accuracy was calculated as the ratio of the number of “Correct” responses to the total number of stimuli presented. RTave and accuracy of monolingual participants were compared to those of bilingual participants using statistical methods to assess any differences between the two groups for both Go and No-Go stimuli.
2.1.4. Statistical analyses: fNIRS data
The main objective of the statistical analysis was to determine the differences in brain activities, measured using the β values, between bilingual and monolingual groups when performing Go and No-Go tasks. For this, β values from the bilingual group were compared statistically with those from the monolingual group for a specific channel and task. The method of statistical comparison used was either a t-test or Mann–Whitney U test, depending on the distribution characteristics of the β values, with a significance level of 0.05. This comparison was carried out for all channels and both tasks; channels showing statistically significant differences between the bilingual and monolingual groups were identified. For each channel exhibiting a statistically significant difference (i.e., P < 0.05) in the previous between-group analysis, the relative difference in brain activity level in this channel was further assessed using the Δβ value. Here Δβ = βmono−βbi; βmono and βbi represent the average β values of the monolingual and bilingual groups, respectively. When Δβ > 0, it would imply that the monolingual group had a stronger HbO/HbR response at the corresponding channel while performing a specified task. When Δβ < 0, the opposite conclusion would be reached; see Figure 3.
cHRFs used in the fNIRS data analysis. These functions are mathematical models used to describe the time-dependent changes in regional concentrations of HbO and HbR in response to neural activities.

3. Results
3.1. Descriptive statistics
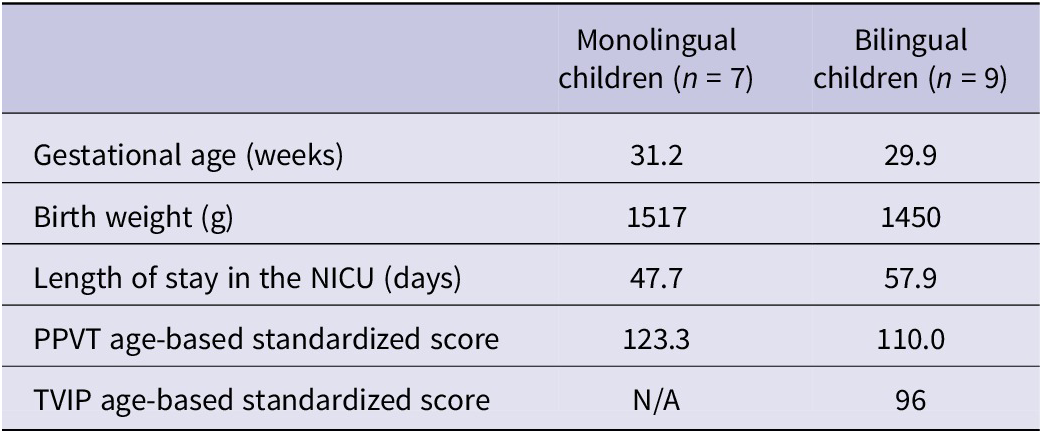
Data on children’s gestational age, birth weight, length of stay in the NICU and vocabulary test scores (PPVT-4 and TVIP) are presented in Table 1 by group. On average, the bilingual group was born earlier than the monolingual group, weighed less and was in the NICU for more time (an average of ten days more). The bilingual group also had two children on oxygen at day 28 and three children with IVH. However, there were no statistically significant differences between the monolingual versus bilingual preterm-born children in any of these variables.
Monolingual and bilingual preterm children’s neonatal health metrics

Table 1. Long description
Starting from the top row, the table lists health metrics for monolingual children, n equals 7, and bilingual children, n equals 9. Gestational age in weeks is 31.2 for monolingual and 29.9 for bilingual. Birth weight in grams is 1517 for monolingual and 1450 for bilingual. Length of stay in the N I C U in days is 47.7 for monolingual and 57.9 for bilingual. P P V T age-based standardized score is 123.3 for monolingual and 110.0 for bilingual. T V I P age-based standardized score is not available for monolingual and 96 for bilingual.
3.1.1. Results: Go/No-Go behavioral data
Descriptive statistics for Go/No-Go task RT and accuracy for both trial types are provided in Table 2. A Mann–Whitney U test confirmed no statistical difference in accuracy between groups on the Go trial (U = 7.0, p = .352) or No-Go trial (U = 8.5, p = .476). However, statistical differences were found in RT where inhibition was required; Go trial (U = 3.0, p = .057) and No-Go trial (U = 0.0, p = .010). This is visually demonstrated in Figure 4.
Accuracy and RT for Go/No-Go Task

Table 2. Long description
The table is divided into two sections. The first section, reaction time, lists GO and NO-GO tasks in rows. For GO, bilinguals have a mean reaction time of 398.8067 milliseconds with a standard deviation of 19.02925, while monolinguals have a mean of 486.0612 and standard deviation of 59.52814. For NO-GO, bilinguals have a mean of 504.7096 and standard deviation of 43.58556, monolinguals have a mean of 615.31 and standard deviation of 73.03976. The second section, accuracy, also lists GO and NO-GO tasks. For GO, bilinguals have a mean accuracy of 0.8875 with a standard deviation of 0.11392, monolinguals have a mean of 0.8515 and standard deviation of 0.06559. For NO-GO, bilinguals have a mean of 0.8783 and standard deviation of 0.0867, monolinguals have a mean of 0.8815 and standard deviation of 0.06501. All values are presented in the same order: bilingual mean, bilingual standard deviation, monolingual mean, monolingual standard deviation.
GNG accuracy and RT for bilinguals versus monolinguals.

Table 1. Long description
Starting from the top row, the table lists health metrics for monolingual children, n equals 7, and bilingual children, n equals 9. Gestational age in weeks is 31.2 for monolingual and 29.9 for bilingual. Birth weight in grams is 1517 for monolingual and 1450 for bilingual. Length of stay in the N I C U in days is 47.7 for monolingual and 57.9 for bilingual. P P V T age-based standardized score is 123.3 for monolingual and 110.0 for bilingual. T V I P age-based standardized score is not available for monolingual and 96 for bilingual.
3.1.2. Results: fNIRS data
Supplementary Figure 2 illustrates the statistically significant differences in cortical hemodynamic responses as quantified βHbO and βHbR between monolingual and bilingual participants during both Go and No-Go trials.
Spatial analysis of hemodynamic response differentials, juxtaposing the channels exhibiting statistically significant between-group differences (Supplementary Figure 2) with their corresponding neuroanatomical substrates, revealed systematic localization within frontal and prefrontal networks integral to executive control processing. Specifically, significant activation differentials were observed in Brodmann Area (BA) 9/46 (dorsolateral prefrontal cortex; fNIRS channels 6, 9, 10 and 17), BA 10 (frontopolar/anterior prefrontal cortex; fNIRS channels 9 and 10), BA 44/45 (inferior frontal gyrus; fNIRS channels 1, 2, 3 and 4) and BA 46/47 (lateral prefrontal cortex/orbital inferior frontal gyrus; fNIRS channels 5 and 22).
Supplementary Figure 2 also presents the quantitative Δβ analysis between the monolingual and bilingual groups. Notably, all channels exhibiting statistically significant differences displayed positive Δβ values, indicating a consistent directional pattern in neurofunctional disparities. This systematic pattern reveals that monolingual participants exhibited significantly elevated cortical recruitment, as indexed by ΔβHbO, across multiple prefrontal channels during both Go and No-Go experimental conditions. The bilingual cohort achieved equivalent or superior task performance while demonstrating substantially and significantly, reduced metabolic demand.
4. Discussion
The present study examined whether bilingual experience is associated with enhanced inhibitory control, a core component EF, in preterm-born children, a population at elevated risk for long-term difficulties in cognitive regulation. Using a Go/No-Go task, we found that bilingual preterm-born children responded significantly faster than their monolingual peers while maintaining comparable levels of accuracy. Converging with these behavioral findings, fNIRS data revealed lower task-evoked HbO responses in frontal regions among bilingual children during inhibitory control demands. Together, this pattern suggests that bilingual children achieved effective inhibitory control with reduced neural effort. To our knowledge, this is the first study to demonstrate converging behavioral and neurophysiological evidence of more efficient inhibitory control in a preterm-born bilingual population.
In cognitive neuroscience, neural “recruitment” refers to the engagement of additional neuronal populations as task demands increase (Petajan, Reference Petajan1991). A substantial body of work using both functional magnetic resonance imaging (fMRI) and fNIRS methodologies indicates that greater task-evoked HbO responses often reflect increased cognitive effort, compensatory processing or less efficient neural engagement, particularly under conditions of high task demand, novelty or lower proficiency. In contrast, reduced HbO responses are frequently observed in individuals who perform tasks more proficiently – such as experts, extensively trained participants or those with more optimized neural networks – and are interpreted as reflecting more efficient neural recruitment (e.g., Kelly & Garavan, Reference Kelly and Garavan2005; Neubauer & Fink, Reference Neubauer and Fink2009; Poldrack, Reference Poldrack2015). Importantly, this interpretation is appropriate when lower HbO responses occur alongside preserved or superior behavioral performance, as was observed in the present study.
Within this framework, the reduced frontal HbO responses observed in bilingual children are particularly compelling given their faster response latencies without accuracy costs. This pattern is consistent with experience-dependent neural adaptation, whereby bilingual children required the engagement of fewer or more focal neural resources to achieve effective inhibitory control. The spatial distribution of group differences across prefrontal regions implicated in response inhibition, interference suppression and cognitive flexibility further supports the interpretation that bilingual experience may shape the functional organization of executive control networks. In contrast, monolingual children appeared to recruit greater neural resources to achieve comparable levels of task accuracy, consistent with less efficient neural processing during inhibitory demands.
This interpretation aligns closely with prior developmental and bilingualism research. Studies comparing children and adults on Go/No-Go and related tasks consistently demonstrate that children exhibit more diffuse and stronger frontal activation, whereas adults show more focal and reduced activation, reflecting a developmental shift from inefficient to efficient neural recruitment (e.g., Durston et al., Reference Durston, Davidson, Tottenham, Galvan, Spicer, Fossella and Casey2006; Mehnert et al., Reference Mehnert, Akhrif, Telkemeyer, Wartenburger, Obrig and Neufang2013). Similarly, work in bilingual populations has shown that bilinguals often exhibit reduced frontal activation relative to monolinguals while achieving equal or superior performance, a pattern interpreted as more efficient tuning of executive control networks (e.g., Abutalebi et al., Reference Abutalebi, Della Rosa, Green, Hernandez, Scifo, Keim, Cappa and Costa2012). Recent fNIRS research in children further supports this account, showing that monolinguals may recruit more channels than bilinguals to achieve comparable success on executive control tasks (Li et al., Reference Li, Wu, Yang, Xie, Chang and Luo2023). Finally, higher HbO responses during inhibitory control have been associated with poorer regulation or increased compensatory demand in pediatric samples (Fishburn et al., Reference Fishburn, Hlutkowsky, Bemis, Huppert, Wakschlag and Perlman2019). The present findings extend this literature by demonstrating a similar efficiency pattern in a preterm-born bilingual population.
Importantly, these group differences emerged in the context of broadly comparable neonatal and developmental profiles. Monolingual and bilingual children did not differ significantly in gestational age, birth weight or length of NICU stay, reducing the likelihood that observed effects were driven by differences in early medical risk. In addition, receptive vocabulary skills were within age-expected norms for both groups. Monolingual children demonstrated high-average performance on the PPVT, while bilingual children scored in the average-to-high-average range on the PPVT and in the average range on the TVIP. Thus, the observed behavioral and neural differences in inhibitory control cannot be attributed to disparities in general language ability or neonatal health status.
From a psycholinguistic perspective, the combined behavioral and neuroimaging findings are consistent with theoretical accounts positing that bilingual language use puts continuous demands on executive control systems because both languages remain jointly active during comprehension and production. Managing this co-activation requires selective attention to task-relevant representations and suppression of competing alternatives, thereby repeatedly engaging core components of executive functioning. Crucially, contemporary frameworks emphasize that the bilingual advantage does not arise from inhibition alone, but from the repeated recruitment and fine-tuning of a unified executive control system supporting attention regulation, conflict monitoring and goal maintenance (Bialystok, Reference Bialystok2015). For preterm-born children, whose frontal networks supporting executive control are both vulnerable and highly plastic, sustained bilingual language engagement may contribute to the shaping and optimization of these control systems over time.
A notable strength of the present study lies in the rigorous, multi-method operationalization of bilingualism, which enhances confidence in the interpretation of these findings. Unlike prior work relying solely on parent-reported exposure, bilingualism here was defined by simultaneous bilingualism from birth with demonstrated productive proficiency in both English and Spanish. Classification was confirmed through a triangulated approach including parent report, behavioral verification of Spanish use, positive child and parent language attitudes toward Spanish and an interview conducted entirely in Spanish by a bilingual researcher. All participants additionally completed standardized vocabulary assessments (PPVT-4 for all children and TVIP for bilinguals). This comprehensive approach reduces misclassification risk and aligns with evidence suggesting that productive capacity may represent a threshold for observing bilingual effects on EF (Baralt & Darcy Mahoney, Reference Baralt and Darcy Mahoney2020; Head et al., Reference Head, Baralt and Darcy Mahoney2015). Taken together, the behavioral and neuroimaging results suggest that bilingual experience in preterm-born children may support the development of a more efficient inhibitory control system. This efficiency may serve as a protective factor against EF vulnerabilities commonly associated with prematurity and highlight the importance of integrating behavioral performance with neural measures, such as fNIRS, to more fully characterize the mechanisms underlying individual differences in EF development.
5. Clinical & policy implications
Given the wide variability in neurodevelopmental outcomes among children born preterm, as well as the central role of inhibitory control in school readiness and adaptive functioning, these findings carry meaningful clinical and educational implications. Early-life experiences that leverage heightened neural plasticity may help support EF development in this at-risk population. In this context, supporting bilingual language environments may represent one such experience-dependent input with potential cognitive benefits.
These findings carry important implications for education policy and clinical practice. For instance, clinicians and educators in the U.S. should encourage home language use alongside English for preterm-born children, as the cognitive benefits associated with bilingualism could translate into better long-term outcomes. Future research should continue to examine inhibitory control using developmentally sensitive tasks and neuroimaging methods in bilingual preterm populations, while employing robust, multidimensional measures of language proficiency. Such work will be critical for determining whether and how bilingual experience can moderate the long-term cognitive consequences of prematurity.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S1366728926101370.
Data availability statement
The data that support the findings of this study are available from the corresponding author upon reasonable request. Consistent with institutional policies and applicable regulations, study data will be retained and available for a minimum of five years following publication of the study.
Competing interests
The authors declare that they have no competing interests.
Funding statement
This work was supported by Robert Wood Johnson Foundation and Baptist Health South Florida.








 Open access
Open access