


Introduction
Roads are essential for social and economic development but are commonly associated with deforestation, habitat loss and challenges for biodiversity conservation (Huijser et al., Reference Huijser, Abra and Duffield2013; Dornas et al., Reference Dornas, Teixeira, Gonsioroski and Nóbrega2019). In addition to fragmenting habitats, they create artificial edges, alter environmental conditions, and exacerbate anthropogenic impacts such as traffic and pollution (Fuentes-Montemayor et al., Reference Fuentes-Montemayor, Cuarón, Vázquez-Domínguez, Benítez-Malvido, Valenzuela-Galván and Andresen2009; Molbert et al., Reference Molbert, Alliot, Goutte and Hansson2025). Unlike some cultivated areas that can serve as habitats for various species (Magioli et al., Reference Magioli, Bovo, Huijser, Abra, Miotto and Andrade2019), roads are uninhabitable areas and act as high-impact fragmentation agents (Da Rosa et al., Reference Da Rosa, Secco, Carvalho, Maia and Bager2018). Habitat loss as a result of fragmentation results in two primary effects: edge effects and barrier effects. Edge effects involve changes in ecological characteristics, such as light and humidity, that influence the composition and structure of biological communities and facilitate the arrival of invasive species (Fahrig, Reference Fahrig2003; Hofmeister et al., Reference Hofmeister, Hošek, Brabec, Střalková, Mýlová and Bouda2019). Barrier effects occur when fragmentation involves physical obstacles such as urban areas and roads that hinder or prevent species movement between remaining fragments (Fahrig & Rytwinski, Reference Fahrig and Rytwinski2009; Laurance et al., Reference Laurance, Goosem and Laurance2009). These effects can reduce habitat connectivity, leading to population isolation, decreased genetic diversity and an increased risk of local extinctions (Jaeger et al., Reference Jaeger, Bowman, Brennan, Fahrig, Bert and Bouchard2005, Grilo et al., Reference Grilo, Borda-de-Água, Beja, Goolsby, Soanes and le Roux2021).
Vehicle traffic exacerbates habitat fragmentation by increasing isolation and edge effects, while also causing noise, light and chemical pollution, and wildlife mortality (Forman, Reference Forman and Alexander2003). Among these effects, there is evidence that roadkills are frequently the top cause of wildlife deaths globally. A systematic review showed that roadkill was the greatest source of mortality in 28% of studied mammal populations, and up to 80% of all known deaths in some cases, ahead of diseases and hunting (Moore et al., Reference Moore, Petrovan, Bates, Hicks, Baker and Perkins2023). In Latin America, > 346 bird species and 159 mammal species are affected by roadkill (Medrano-Vizcaíno et al., Reference Medrano-Vizcaíno, Grilo, Silva Pinto, Carvalho, Melinski, Schultz and González-Suárez2022). These pressures are of particular concern in neotropical forests, which harbor some of the highest levels of biodiversity (Gaston, Reference Gaston2000). In the Brazilian Atlantic Forest only 23% of the original forest cover remains, fragmented into patches of varying sizes and conservation stages (Vancine et al., Reference Vancine, Muylaert, Niebuhr, Oshima, Tonetti and Bernardo2024). This hotspot of endemism and biodiversity faces intense anthropogenic pressure (Myers et al., Reference Myers, Mittermeier, Mittermeier, da Fonseca and Kent2000; Trindade & Marques, Reference Trindade and Marques2024), exacerbating the impacts of roads on local wildlife. For example, in São Paulo state, Brazil, c. 39,600 medium-sized to large mammals die annually on roads (Abra et al., Reference Abra, Huijser, Magioli, Bovo and Ferraz2021).
In southern Bahia, cabrucas (cocoa plantations under the shade of forest fragments) help to mitigate habitat fragmentation, and support the presence of various animal taxa, including endemic and threatened mammal species (Cassano et al., Reference Cassano, Schroth, Faria, Delabie and Bede2009; Oliveira et al., Reference Oliveira, Costa, Sambuichi and Filho2011) such as Wied’s marmoset Callithrix kuhlii and the golden-headed lion tamarin Leontopithecus chrysomelas, categorized as Vulnerable and Endangered, respectively, on the IUCN Red List (IUCN, 2024). However, despite the region’s significance for biodiversity, the impact of roads on local fauna remains understudied.
Research on smaller roads in Brazil is limited (Magioli et al., Reference Magioli, Bovo, Huijser, Abra, Miotto and Andrade2019), with their impacts often overshadowed by those of larger roads (Abra et al., Reference Abra, Huijser, Magioli, Bovo and Ferraz2021). Collisions are more frequent on roads with intermediate traffic volumes, as low-traffic roads (< 2,500 vehicles/day) present minimal risk, and roads with high traffic volumes (> 10,000 vehicles/day) act as barriers, hindering wildlife movement (Seiler, Reference Seiler2003). Our principal aim was to investigate the influence of a small road within a cocoa agroforestry region. We surveyed vertebrates killed on the road, analysing the relationship with traffic volume, and identifying roadkill hotspots. We then focused on mammals as a model group to evaluate wildlife presence near the road and examine its relationship with the road edge. Additionally, we explored the overlap between mammal species in the surrounding habitats and those killed on the road.
Study area
The southern region of Bahia contains the largest remaining portion of Atlantic Forest in the state and is a biodiversity hotspot (Martini et al., Reference Martini, Fiaschi, Amorim and Paixão2007). This mosaic of landscapes includes primary and secondary forests, pastures, rubber and eucalyptus plantations, and cabrucas (MapBiomas Cacau, 2023). Cabrucas, which are created by thinning understorey vegetation for shaded cocoa planting, help maintain forest structure, mitigate deforestation (Ferreira et al., Reference Ferreira, Peres, Dodonov and Cassano2020), function as ecological corridors, and promote gene and biodiversity flow (Oliveira et al., Reference Oliveira, Costa, Sambuichi and Filho2011). Such complex agroforestry systems provide habitat for native flora and fauna, including forest-dependent species (Ferreira et al., Reference Ferreira, Peres, Dodonov and Cassano2020, Reference Ferreira, Arroyo-Rodríguez, Morante-Filho, Storck-Tonon, Somavilla and dos Santos-Silva2024; Cabral et al., Reference Cabral, Faria and Morante-Filho2021).
Our study was conducted along the BA-262 road, also known as the Cocoa Route, which links the municipalities of Ilhéus, on the coast, to Uruçuca, in the west of Bahia. This is a single carriageway surrounded by cabrucas (Fig. 1), and is also important for the transport of iron, connecting the interior of the state to the port complex of Porto Sul. The region is characterized by a tropical humid climate (Af, Köppen), with annual precipitation > 2,000 mm, minimal seasonal variation in rainfall and temperature, and a mean annual temperature of 24 °C (SEIA, 2022).
Distribution of survey sites along a section of the BA-262, a single carriageway road in a cocoa agroforestry region of southern Bahia, Brazil. Land cover is from MapBiomas Cacau (2023).

Methods
Rainfall data used were obtained from station 291360602A, located in Ilhéus (14.789158°W, 39.048858°S; SEIA, 2022).
Roadkill monitoring
We monitored deaths of wildlife from collisions with vehicles during May–December 2022. Given the region’s limited seasonality in temperature and rainfall, we expect this survey period to capture typical roadkill patterns (SEIA, 2022). We monitored wildlife collisions along a 24 km section, in eight surveys of 5 consecutive days, totalling 40 days of monitoring. On survey days, a continuous active search for carcasses was conducted by car at an average speed of 40 km/h, from 5.30 to 16.00. On the same days, we inspected live traps and camera traps on transects along the same road section. The same 24 km section was surveyed in both directions (outbound and return; 48 km in total), resulting in 240 km surveyed per campaign. To prevent the recounting of the same carcasses, all were removed from the road upon detection; well-preserved specimens were deposited in scientific collections, and deteriorated ones were discarded. Because of limitations in detecting small animals (< 1 kg) in vehicle surveys (Teixeira et al., Reference Teixeira, Coelho, Esperandio and Kindel2013), we also conducted walks along 3 km of the road, in six random segments of 500 m, re-randomized on each survey day. Total effort over the eight surveys was 120 km on foot and 1,800 km by car. At the beginning of each 5-day survey, any carcasses already on the road were recorded and then removed; they were included only in the hotspot analyses.
Species identification was validated by specialists, and nomenclature follows Abreu et al. (Reference Abreu, Casali, Garbino, Loss, Moras and Nascimento2025). Opportunistic records, including sightings during monitoring and carcasses found outside the monitoring period, were solely included to compile the species list. Threats were assessed using IUCN (2024), ICMBio (2018) and SEMA (2017).
Traffic volume estimation
To assess the volume of vehicle traffic on the BA-262, a camera trap (Bushnell Trophy Cam HD, Bushnell, USA) was positioned at a 45º angle on the edge of the road. During the eight wildlife monitoring surveys, the camera trap was installed for 6 days and 5 nights, with daily inspections to replace batteries and the memory card. The camera provided detailed information on traffic flow and composition.
The mean daily traffic volume was computed by averaging the total number of vehicles recorded over 24 h. For periods with missing data as a result of equipment malfunction, an average was calculated based on data from the corresponding time and day of the week, considering the observed traffic fluctuations across monitoring days. Traffic patterns over 24 h were elucidated using hourly mean values.
Mammal surveys
To survey small mammals, 60 live traps (30 Sherman, 25 × 8 × 9 cm; 30 Tomahawk, 18 × 18 × 39 cm) were placed in forest fragments along 10 transects perpendicular to the road, following a fishbone pattern covering both sides of the road, and spaced 2 km apart (Fig. 2). Each transect included three survey points: 50, 150 and 250 m from the road edge. At each point, one trap of each type was deployed on the ground. Traps were deployed for 6 nights each month during May–December 2022, totalling 2,880 trap-nights. Traps were baited with a mixture of banana, sardine, ground peanuts, cornmeal and palm oil, a mixture designed for the agroforestry setting and to attract local species. The traps were checked daily, and any captured animals were identified, marked with identification earrings, and released near the capture site in accordance with the guidelines of the American Society of Mammalogists (Sikes et al., Reference Sikes2016).
Design of the camera-trap and live-trap surveys for mammals along and beside the BA-262 road in southern Bahia, Brazil (Fig. 1).

For surveying medium-sized and large mammals, one camera trap (Bushnell Trophy Cam HD) was deployed on each of the 10 transects, spaced c. 2 km apart (Fig. 2). Camera placement alternated among points from 50 to 250 m from the road edge, with positions rotated monthly to avoid site-specific bias. Cameras were installed at knee height and remained continuously active for 30 days before being moved to the next location. The total sampling effort was 1,800 trap-days, corresponding to an average of 180 trap-days per transect. Records of the same species at the same station were considered independent if > 1 h apart. We defined small non-flying mammals as those weighing < 1 kg (Chiarello, Reference Chiarello2000).
Data analysis
To analyse the relationship between carcass numbers and traffic volume, we used generalized linear models (GLMs) with a Poisson distribution. We examined daily vehicle count, total carcass count, and carcass numbers by faunal group (amphibians, reptiles, birds and mammals). We used linear regression to assess the relationship between mean monthly roadkill and monthly precipitation. To prevent data overrepresentation, only daily records of fresh carcasses were considered. To investigate differences in roadkill between months, we used an ANOVA and the Tukey post-hoc test. We tested homogeneity and normality of residuals with Levene and Shapiro–Wilk tests, respectively.
The spatial distribution of roadkill incidents was analysed using KDE+ 2.2 (Bíl et al., Reference Bíl, Andrášik, Svoboda and Sedoník2016), which combines kernel density estimation with statistical testing to identify significant clusters of wildlife–vehicle collisions along linear networks. Cluster significance was evaluated using cluster strength, a dimensionless index used to rank high-risk road sections, determined by four factors: the number of collisions within a cluster, cluster length, the total number of collisions along the analysed road section and the length of that section (Bíl et al., Reference Bíl, Andrášik and Janoška2013, Reference Bíl, Andrášik, Svoboda and Sedoník2016). Analyses were based on GPS-recorded locations, using a bandwidth of 150 m, which defines the spatial window used to smooth and estimate local roadkill density along the road, and 800 Monte Carlo simulations, in which events are randomly redistributed along the road network to generate a null distribution to test the statistical significance of the observed clusters. Clusters were analysed for all vertebrates and per group (mammals, birds, reptiles, amphibians), using roadkill data from the entire monitoring period.
To evaluate any potential relationship between mammal richness and relative abundance and distance from the road edge, we used GLMs and generalized linear mixed models (GLMMs). Relative abundance was expressed as the number of independent records per sampling point for camera traps, and the number of captures per sampling point for live traps. We checked the adequacy of the data distribution and confirmed that it followed a Poisson distribution. We tested models both with and without transect as a random factor, and selected the best-fitting model based on the Akaike information criterion (AIC).
GLM analyses were performed using R 4.4.0 (R Core Team, 2024), and GLMM analyses with the R package lme4 (Bates et al., Reference Bates, Maechler, Bolker, Walker, Christensen and Singmann2015). Model assumptions were checked using the performance package (Lüdecke et al. Reference Lüdecke, Ben-Shachar, Patil, Waggoner and Makowski2021). Model selection was based on AIC, with models differing by < 2 from the lowest AIC considered the best fit. The best-fitting models for mammal abundance included transect as a random factor. Analyses focused on: (1) richness and relative abundance of small mammals captured in live traps in relation to distance from the road; (2) richness and relative abundance of medium-sized and large mammals recorded by camera traps, in relation to distance from the road; and (3) total richness and relative abundance of mammals recorded by camera traps (small, medium and large), in relation to distance from the road. In the latter analysis, small rodents recorded by camera traps were included only in the abundance analysis, because this method does not allow reliable species-level identification for this group.
Results
Mortality
We recorded 245 vertebrate roadkills totaling 42 species in 12 orders (Supplementary Table 1), with a roadkill rate of 0.12 individuals per km per day. The mean number of roadkills per 5-day survey was 30.63 ± SE 5.31. Amphibians were the most recorded, with 153 individuals (63% of the total), followed by reptiles (49, 20%), birds (28, 12%) and mammals (13, 5%). The most commonly recorded taxon in each group were toads Rhinella sp. (14), blind snake Amphisbaena alba (11), black vulture Coragyps atratus (6) and crab-eating fox Cerdocyon thous (3). Of the 153 amphibian carcasses recorded, only 32 (21%) could be identified to species (nine species in total), because of rapid decomposition. The months with the highest and lowest number of roadkills were November (60) and July (14), respectively. Variation in the monthly roadkill rates was greatest for amphibians (Fig. 3). There were significant differences in the number of roadkills between months (F (7,32) = 2.71, P = 0.02), specifically between November and July (P = 0.01) and November and September (P = 0.03). There were no significant differences between months in the number of roadkills of mammals, birds and reptiles. There was no relationship between mean number of vertebrate roadkills per month and precipitation (Pseudo-R2 = 0.04, slope = 0.001, P = 0.13).
Monthly roadkill rate (individuals/km) during May–December 2022 for amphibians, reptiles, birds and mammals on a 24-km section of the BA-262 road in southern Bahia, Brazil.

Mortality and traffic volume
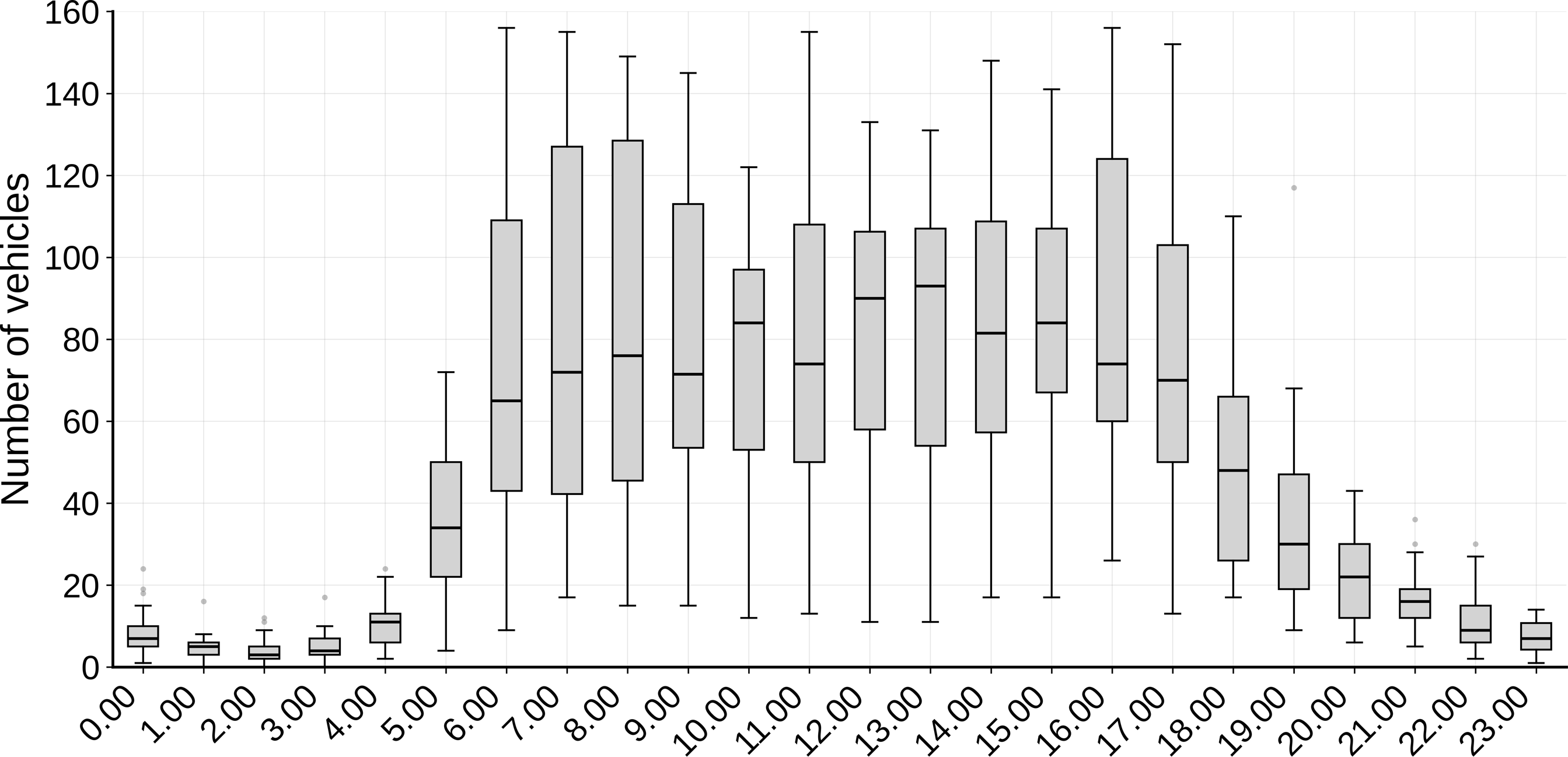
A total of 49,626 vehicles were recorded, an average daily traffic volume of 1,240 vehicles/day. There was little variation in traffic volume between days but significant variation within the day, with most vehicle records (80%) between 7.00 and 18.00 (82.61 ± SE 10.43 per h; Fig. 4). Traffic volume was significantly lower at night than during the day (U = 26.5, P < 0.05). Overall vertebrate roadkill slightly decreased as daily traffic volume increased (Pseudo-R² = 0.03, slope = −0.003, P = 0.04) (Fig. 5), although this was not the case for each vertebrate group considered separately (Supplementary Fig. 1).
Hourly traffic volume on the BA-262 road in southern Bahia, Brazil, over 24 h. Boxes show the interquartile range (25th–75th percentile), the horizontal line the median, whiskers the minimum and maximum values, and dots outliers. Average daily traffic volume was 1,240 vehicles/day). Approximately 80% of vehicles passed during 7.00–18.00 (mean 82 vehicles/h). During 18.00–7.00, the mean was 20 vehicles/h, with the lowest flow recorded during 1.00–3.00 (4 vehicles/h).

Relationship between number of daily roadkill events and mean daily traffic volume on the BA-262 road in southern Bahia, Brazil. Each point represents one survey day (n = 40). The central line shows the fitted generalized linear model with Poisson distribution, and the grey shaded area represents the 95% CI. The model indicates a weak negative relationship between roadkill and traffic volume.

Roadkill hotspots for all vertebrates combined and for amphibians, birds, mammals and reptiles along a 24 km section of the BA-262 road in southern Bahia, Brazil.

Roadkill clusters
Considering all vertebrate carcasses together (n = 245), hotspot analysis identified nine clusters, covering 18.7% of the surveyed road section (Fig. 6). When taxa were analysed separately, 13 clusters were identified for amphibians (covering 19.3% of the road section), 10 for reptiles, six for birds and two for mammals (Supplementary Table 2). Differences between pooled and taxon-specific analyses reflect distinct spatial patterns among taxonomic groups, and the partial spatial overlap among clusters indicates that some road sections are shared crossing areas for multiple taxa.
Mammal surveys
We obtained 409 mammal records using live traps and camera traps, totalling 21 species of eight orders (Supplementary Table 3). We captured 56 small mammals (with 21 recaptures), including eight native species, and one exotic (Rattus sp.). Hylaeamys seuanezi was the most captured species (Supplementary Table 3). Although many records (43%) occurred < 50 m from the road, some species (Oecomys catherinae, Oligoryzomys nigripes and Phyllomys pattoni) were recorded only 250 m from the road (Supplementary Fig. 2). With camera traps, we obtained 353 records, comprising 20 species of eight orders; C. thous was the most recorded species (Supplementary Table 3). Similarly to small mammals, 57% of the records were < 50 m from the road. As expected, most mammal records (73%) were obtained during the night (Supplementary Fig. 3). Fourteen species were detected at 50 m from the road and 13 species at 250 m. Some species (Herpailurus yagouaroundi, Coendou insidiosus and Didelphis aurita) were only detected 250 m from the road (Supplementary Fig. 4).
Mammal richness and relative abundance
For small mammals, neither richness nor relative abundance was significantly related to distance to the road. Richness of medium-sized and large mammals also showed no relationship with distance to the road but their abundance was significantly higher near the road (GLMM: Z = −2.16, P = 0.03; Supplementary Table 4).
Mammal diversity and roadkill
Of the 10 mammal species roadkilled, seven were also identified by camera trap or live trapping (Supplementary Table 5). One individual of Leontopithecus chrysomelas was also killed on the road, and this species was frequently observed during fieldwork. Rhipidomys mastacalis was only detected as roadkill.
Discussion
Wildlife roadkill and traffic volume
The relationship between wildlife roadkill and traffic volume has been widely debated, with studies reporting positive (e.g. Medeiros et al., Reference Medeiros, Cornelius, Costa, Venticinque, Atroari and Waimiri2025), negative (e.g. Teixeira et al., Reference Teixeira, Kindel, Hartz, Mitchell and Fahrig2017) and neutral effects (e.g. Bissonette & Kassar, Reference Bissonette and Kassar2008). Although findings are mixed, a positive relationship is most frequently reported (Pagany, Reference Pagany2020). In contrast, we detected a negative relationship between the number of vertebrates killed and daily traffic volume. Similar to the negative trends observed for some species in Europe (Grilo et al., Reference Grilo, Zanchetta Ferreira and Revilla2015), our results suggest that higher traffic intensity may act as a barrier, limiting animal crossings and consequently reducing the probability of collisions. These findings suggest the nature of the effect of traffic is not universal but dependent on context (Denneboom, Reference Denneboom, Bar-Massada and Shwartz2024).
The risk of collisions with wildlife can vary among regions, habitats and species as a result of morphological and ecological differences (Medrano-Vizcaíno et al., Reference Medrano-Vizcaíno, Grilo, Silva Pinto, Carvalho, Melinski, Schultz and González-Suárez2022). Of the four vertebrate groups we recorded killed on the BA-262, amphibians comprised 63% but their death rate was not correlated with traffic volume. Other factors, such as low mobility, seasonal reproductive behaviour and sensitivity to vehicle lights and noise, presumably played a part (Coelho et al., Reference Coelho, Teixeira, Colombo, Coelho and Kindel2012; Teixeira et al., Reference Teixeira, Coelho, Esperandio and Kindel2013; Dornas et al., Reference Dornas, Teixeira, Gonsioroski and Nóbrega2019). In addition, we may have underestimated amphibian mortality, as precipitation and temperature strongly influence the persistence of their carcasses. Precipitation above c. 4 mm increases carcass disappearance rates by up to 800%, potentially eliminating up to 87.5% of carcasses within short time periods (Boyle et al., Reference Boyle, Balsdon, Newediuk, Litzgus and Lesbarrères2025), consistent with studies indicating greater disappearance in humid environments (Boyle et al., Reference Boyle, Keevil, Litzgus, Tyerman and Lesbarrères2021; Bénard et al., Reference Bénard, Bonenfant and Lengagene2024). Given the high rainfall and temperatures in our study area, it is plausible that the mortality of amphibians and other small vertebrates, such as reptiles, birds and mammals, is underestimated (Teixeira et al., Reference Teixeira, Coelho, Esperandio and Kindel2013; Santos et al., Reference Santos, Mira, Salgueiro, Costa, Medinas and Beja2016; Barrientos et al., Reference Barrientos, Martins, Ascensão, D’Amico, Moreira and Borda-de-Água2018).
Reptiles, primarily snakes, were the second most recorded group in our study, with their mortality probably linked to low mobility, thermoregulatory behaviour and possibly intentional roadkill (Secco et al., Reference Secco, Ratton, Castro, da Lucas and Bager2014; Gonçalves et al., Reference Gonçalves, Alvares, Teixeira, Schuck, Coelho and Esperandio2018). For instance, Farias et al. (Reference Farias, Napoli, Dodonov and Forti2022) recorded 838 roadkills (827 amphibians and 11 reptiles) in 47 days along a 2.5 km section of a secondary road in Bahia during the rainy season, when animals move between habitats to feed and reproduce.
For birds, greater body weight, population size, wider habitat range, and generalist habits are important factors in explaining vulnerability to roadkill, and flight and foraging activity along roads further increase this risk (Medrano-Vizcaíno et al., Reference Medrano-Vizcaíno, Grilo, Silva Pinto, Carvalho, Melinski, Schultz and González-Suárez2022). In addition, flight height and body weight also influence mortality as a result of vehicle-induced air displacement (Clevenger et al., Reference Clevenger, Chruszcz and Gunson2003). Some birds avoid roads because of noise pollution, whereas others are attracted to food resources, such as garbage, insects and seeds (Erritzoe et al., Reference Erritzoe, Mazgajski and Rejt2003). The most common bird species killed in our study, the black vulture Coragyps atratus, frequently forages on mammal carcasses, supporting the suggestion that large body size and ground foraging behaviour are associated with higher collision risk (González-Suárez et al., Reference González-Suárez, Zanchetta Ferreira and Grilo2018). Small forest passerines are also affected, as a result of foraging in roadside shrubs (Santos et al., Reference Santos, Mira, Salgueiro, Costa, Medinas and Beja2016). Fourteen of the 28 bird species we recorded killed were passerines.
We recorded only 13 roadkills of native mammals, and there was no significant correlation with traffic volume, potentially because most of the species detected are predominantly nocturnal and traffic volume is lowest at night. Similar patterns have been reported elsewhere (Odden et al., Reference Odden, Athreya, Rattan and Linnell2014; Kite et al., Reference Kite, Nelson, Stenhouse and Darimont2016; Abra et al., Reference Abra, Granziera, Huijser, Ferraz, Haddad and Paolino2019, Reference Abra, Canena, Garbino and Medici2020). Alteration of activity patterns as a result of traffic volume may be more likely amongst species that are active during the day (Brett et al., Reference Brett, Mehner, Young, Lehnen and Kline2025).
Most mammal species recorded in our study (except H. yagouaroundi) were killed at night, with C. thous being the most frequently killed species, aligning with studies elsewhere in Bahia (Oliveira et al., Reference Oliveira, Costa, Sambuichi and Filho2011). The occurrence of roadkill involving endemic and threatened primates, such as Callithrix kuhlii and Leontopithecus chrysomelas, highlights the potential impacts of roads on species of conservation concern in southern Bahia, a region dominated by cabruca agroforestry systems that are important habitats for these primates (Oliveira et al., Reference Oliveira, Costa, Sambuichi and Filho2011; IUCN, 2024). In this context, the BA-262 road crosses landscapes that are directly relevant for regional conservation strategies, including those established by the Bahia Lion Tamarin Conservation Initiative in 2020–2029, and reinforces concerns about the effects of road infrastructure on the persistence and connectivity of primate populations in human-modified forest mosaics (Praill et al., Reference Praill, Eppley, Shanee, Cunneyworth, Abra and Allgas2023). Even low roadkill rates can have significant impacts on some species (Forman & Alexander, Reference Forman and Alexander1998; Abra et al., Reference Abra, Huijser, Magioli, Bovo and Ferraz2021).
Spatial analysis revealed roadkill hotspots along almost 20% of the road length surveyed. Overlapping roadkill hotspots across taxa, as observed in other studies, suggest a shared use of landscape features and ecological pathways (i.e. routes and habitats regularly used by multiple species for movement and access to resources), which reinforces the value of multi-species mitigation strategies (Carvalho, Reference Carvalho2014; Bueno et al., Reference Bueno, Sousa and Freitas2015) such as humps in the road to reduce vehicle speed. Identification of larger-scale hotspots combined with habitat connectivity mapping can support the optimization of mitigation measures (Clevenger et al., Reference Clevenger, Chruszcz and Gunson2003). For biodiverse regions, such approaches are crucial, especially as increased traffic from infrastructure developments could exacerbate roadkill rates or increase the barrier effect. Measures such as speed humps can serve as multi-species mitigation because they reduce vehicle speed and thus the likelihood of collisions. For example, the installation of four speed humps on a single-carriageway road in Zanzibar, Tanzania, resulted in an 80% reduction in the mortality of a threatened primate (Olgun et al., Reference Olgun, Mohammed, Mzee, Green, Davenport and Georgiev2021).
Patterns of mammal roadkill
Ecological changes caused by roads can affect species diversity and density hundreds of meters away from the roads (Forman & Alexander, Reference Forman and Alexander1998), with the zone of effect varying amongst taxa. For mammals, this zone can be variable in size, extending up to c. 5 km (Benitèz-Lopes et al., Reference Benítez-López, Alkemade and Verweij2010). However, the effect can also vary depending on the type of road, differing between secondary and main roads (Andrasi et al., Reference Andrasi, Jaeger, Heinicke, Metcalfe and Hockings2021). However, we did not detect significant variations in mammal richness in relation to distance from the road. Although the effects of roads are more pronounced within the first 100 m from the edge (Da Rosa et al., Reference Da Rosa, Secco, Carvalho, Maia and Bager2018), the maximum distance we considered (250 m) may not have been sufficient to detect any changes in species numbers. Road density may be more relevant than road presence for explaining differences in species richness, with increased road density being associated with decreased species richness and population persistence in some areas (Pinto et al., Reference Pinto, Bager, Clevenger and Grilo2018; Grilo et al., Reference Grilo, Borda-de-Água, Beja, Goolsby, Soanes and le Roux2021; Kent et al., Reference Kent, Schwartz and Perkins2021).
The diversity of mammals recorded along the road margins reinforces the relevance of cabruca-dominated landscapes in shaping species occurrence in our study area. Previous studies have indicated that forest cover favours medium-sized and large species, such as Subulo gouazoubira, Dicotyles tajacu, Eira barbara, and Tamandua tetradactyla, whereas C. thous and Procyon cancrivorus (species considered generalists in habitat and diet) responded negatively to it (Cassano et al., Reference Cassano, Schroth, Faria, Delabie, Bede, Oliveira and Mariano-Neto2014; Ferreira et al., Reference Ferreira, Peres, Dodonov, Mariano-Neto, Faria and Cassano2025). In our study area, Cervidae were more frequently recorded at the edges of the road, especially during periods of lower traffic, possibly attracted by the presence of grasses. In contrast, species more sensitive to disturbance, such as Leopardus wiedii and H. yagouaroundi, were not recorded near the road, suggesting avoidance behaviour even under low traffic volume. Thus, species occurrence near the road margins appears to be more strongly associated with sensitivity to traffic and associated disturbance than with vegetation cover per se. These findings reinforce the suggestion that the use of areas adjacent to the highway varies according to the sensitivity of the species, their biological characteristics, and the attributes of the environment (Da Rosa et al., Reference Da Rosa, Secco, Carvalho, Maia and Bager2018; Brett et al., Reference Brett, Mehner, Young, Lehnen and Kline2025).
The relative abundance of medium-sized and large mammals was significantly higher near the road, possibly a result of initial attraction to the road followed by retention because of the resources available in the surrounding area (Da Rosa et al., Reference Da Rosa, Secco, Carvalho, Maia and Bager2018; Ruiz-Capillas et al., Reference Ruiz-Capillas, Mata, Fernández, Fernandes and Malo2021). However, although proximity to the road may bring localized benefits, these effects vary between species, populations and the temporal scale evaluated, and may coexist with negative impacts, such as demographic changes and direct mortality (Hill et al., Reference Hill, DeVault and Belant2021; Brett et al., Reference Brett, Mehner, Young, Lehnen and Kline2025).
Monitoring data indicated that eight of the 10 mammal species killed by vehicles were also recorded near the road, suggesting a link between species presence and road proximity, consistent with evidence that roads can attract wildlife and increase collision risk (Da Rosa et al., Reference Da Rosa, Secco, Carvalho, Maia and Bager2018; Ruiz-Capillas et al., Reference Ruiz-Capillas, Mata, Fernández, Fernandes and Malo2021). Traits such as generalist habits, home ranges < 10 km², carrion- or invertebrate-based diets, short life cycles (< 10 years) and a body mass of 2–35 kg are predictors of road mortality (Medrano-Vizcaíno et al., Reference Medrano-Vizcaíno, Grilo, Silva Pinto, Carvalho, Melinski, Schultz and González-Suárez2022). The mammal species with the greatest number of roadkills in our study, C. thous, is one of the mammals most recorded as roadkill in the Atlantic Forest (Abra et al., Reference Abra, Huijser, Magioli, Bovo and Ferraz2021; Bernsdorf, Reference Bernsdorf2022). A highly adaptable species, it is frequently observed near roads (Tenorio et al., Reference Tenorio, Cruz, Silveira-Filho, Alexandre, Gripp and Araújo2023) and has an opportunistic diet of fruits, insects, crustaceans and small vertebrates, including amphibians (Gatti et al., Reference Gatti, Bianchi, Rosa and Mendes2006; Goebel et al., Reference Goebel, Longo, Oliveira, Fermiano, da Silva, Ignácio and dos Santos-Filho2023). Its opportunistic foraging, along with the presence of amphibian carcasses, may further attract individuals to roads (Tenorio et al., Reference Tenorio, Cruz, Silveira-Filho, Alexandre, Gripp and Araújo2023).
The relatively low number of collisions involving mammals, compared to other groups, may be related to their predominantly nocturnal activity, which coincides with lower traffic volumes and, consequently, a reduced risk. However, diurnal species are especially vulnerable to this risk. Among the species affected, for those threatened with extinction (Callithrix kuhlii and Leontopithecus chrysomelas), roadkill can aggravate population declines. Considering the 245 vertebrates recorded killed in the 40 days of our survey, we estimate that > 2,000 vertebrates (amphibians, reptiles, birds and mammals combined), are hit by vehicles annually on this 24 km section of the BA-262 road.
Unlike most studies focusing solely on roadkill, we integrated surveys of live fauna to identify species and activity patterns near the road. This approach revealed a diverse mammal community inhabiting areas directly affected by the road, indicating potential vulnerability even in the absence of frequent collisions. The low number of roadkill records may reflect a barrier effect, which limits animal movement and reduces connectivity. Our findings provide a baseline for future comparisons and emphasize the importance of integrating ecological knowledge into development planning, guiding the design of mitigation measures such as canopy bridges, wildlife underpasses, fencing, speed control and vegetation restoration. These measures should be considered not as additional costs, but as integral components of infrastructure projects, essential for maintaining habitat connectivity and road safety.
Author contributions
Study design: ARSR, FDA, RSB; fieldwork: ARSR; data analysis: ARSR, RSB, MM; writing and revision: all authors.
Acknowledgements
This work was supported by the Coordination for the Improvement of Higher Education Personnel (CAPES) (Funding Code 001, FAPESB–Foundation for Research Support of the State of Bahia), and the State University of Santa Cruz (PROPP No 073.6764.2023.0022931-63 awarded to the PPG Ecology and Biodiversity Conservation Program, and the Applied Ecology and Conservation Lab). MM thanks the São Paulo Research Foundation for a postdoctoral scholarship (#2022/06791-9).
Competing interests
None.
Ethical standards
This research abided by the Oryx guidelines on ethical standards. We obtained authorization from the Animal Ethics Committee of the State University of Santa Cruz and a licence for scientific activities from the Biodiversity Authorization and Information System of the Chico Mendes Institute for Biodiversity Conservation. Camera traps were installed exclusively to monitor wildlife activity, and positioned and oriented to avoid the intentional recording of people or vehicles. Any images or recordings that included identifiable people were deleted and not stored or analysed. Data were used solely for scientific purposes related to biodiversity monitoring. All procedures followed principles for the socially responsible use of conservation technology, following Sandbrook et al. (Reference Sandbrook, Clark, Toivonen, Simlai, O’Donnell, Cobbe and Adams2021) and Sharma et al. (Reference Sharma, Fiechter, George, Young, Alexander and Bijoor2020).
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Supplementary material
The supplementary material for this article is available at doi.org/10.1017/S0030605325102597








 Open access
Open access