


Introduction
The extinction of animal species not only reduces biodiversity but also leads to a reduction in ecological interactions (Galetti & Dirzo, Reference Galetti and Dirzo2013; Bello et al., Reference Bello, Galetti, Pizo, Magnago, Rocha and Lima2015), which in turn affects ecosystem functionality and services (Valiente-Banuet et al., Reference Valiente-Banuet, Aizen, Alcántara, Arroyo, Cocucci and Galetti2015). Reintroduction projects can restore locally extinct species and their associated ecological interactions by re-establishing viable populations within their original range (Soulé & Noss, Reference Soulé and Noss1998; Root-Bernstein & Svenning, Reference Root-Bernstein and Svenning2017). The success of reintroductions depends on extrinsic factors such as the habitat quality of the release area and intrinsic factors, including biology and behaviour of the species involved (Berger-Tal et al., Reference Berger-Tal, Blumstein and Swaisgood2020).
Since 2010 there has been a growing focus on understanding animal behaviour in reintroduction projects (Berger-Tal et al., Reference Berger-Tal, Blumstein, Carroll, Fisher, Mesnick and Owen2016), which has been linked to increasing success rates (Soorae, Reference Soorae2011). This knowledge is crucial for the reintroduction of captive-born animals, where success rates are lower compared to animals translocated from the wild because of behavioural differences in captivity (Griffith et al., Reference Griffith, Scott, Carpenter and Reed1989; Jule et al., Reference Jule, Leaver and Lea2008; Blanco et al., Reference Blanco, Spørring and Bitetti2016; Berger-Tal et al., Reference Berger-Tal, Blumstein and Swaisgood2020). Stress associated with the need to adjust behaviour to a new environment is a significant cause of post-release mortality (Teixeira et al., Reference Teixeira, de Azevedo, Mendl, Cipreste and Young2007). Behavioural adjustments can impact activity budgets, reducing time spent on foraging or resting, which may then compromise survival and reproductive success. Translocated animals must explore their new environment after release and identify suitable foraging and resting sites, whilst minimizing travel time and exposure to predators (Hurtado et al., Reference Hurtado, Beck and Thebpanya2018). Over time, new behaviour patterns can emerge post-release (Hurtado et al., Reference Hurtado, Beck and Thebpanya2018) and can be passed to young born within reintroduction projects (Kierulff et al., Reference Kierulff, Ruiz-Miranda, de Oliveira, Beck, Martins and Dietz2012; Parres et al., Reference Parres, Palazón, Afonso, Quenette, Batet and Camarra2020).
In the last 2 decades, efforts have been made to restore the Atlantic Forest (Fernandez et al., Reference Fernandez, Rheingantz, Genes, Kenup, Galliez and Cezimbra2017), a biodiversity hotspot covering just 12.4% of its original extent (Fundação SOS Mata Atlântica & INPE, 2024). A project was launched in 2017 to reintroduce the lowland tapir Tapirus terrestris to the Atlantic Forest of Rio de Janeiro state, south-east Brazil (Galliez et al., Reference Galliez, Zamboni, Macedo, Di Martino, Rosas, Melletti, Reyna-Hurtado and Medici2024). Although the lowland tapir has a wide distribution, extending from Venezuela to northern Argentina, the species has become extinct in many parts of its range (Medici et al., Reference Medici, Flesher, de Beisiegel, Keuroghlian, Desbiez and Gatti2012; Flesher & Medici, Reference Flesher and Medici2022), including in all forest fragments in Rio de Janeiro state (Macedo, Reference Macedo2017), and is categorized as Vulnerable on the IUCN Red List (Varela et al., Reference Varela, Flesher, Cartes, Chalukian, de Bustos, Ayala and Richard-Hansen2019). Tapirus terrestris is the largest terrestrial mammal in Brazil and is considered to be a surviving species of the Neotropical megafauna. Lowland tapirs are solitary animals (Tobler, Reference Tobler2008; Ferreguetti et al., Reference Ferreguetti, Tomas and Bergallo2017), with crepuscular and nocturnal activity patterns in wild populations (Medici, Reference Medici2010; Oliveira-Santos et al., Reference Oliveira-Santos, Machado-Filho, Tortato and Brusius2010; Cruz et al., Reference Cruz, Paviolo, Bó, Thompson and Di Bitetti2014). However, captivity affects tapirs by reducing the diversity of behaviour patterns and increasing diurnal activity (Arumugam et al., Reference Arumugam, Luan, Ibrahim, Toh, Buesching and Annavi2018). The reintroduction project in the Atlantic Forest used captive-born tapirs whose behaviour was likely to differ from wild tapirs.
We investigated the activity patterns of tapirs reintroduced to the Atlantic Forest to assess whether their behaviour post-release converged with that of wild tapirs. We tested the hypothesis that reintroduced tapirs would show a decline in diurnal activity and a reduction in overall activity over time as they adjusted to their natural environment and their behaviour became more closely aligned with that of wild tapirs. Additionally, we tested for effects of sex, season and group of origin on tapir activity patterns. Our objective was to identify the factors influencing tapir behavioural changes, to improve reintroduction strategies and the long-term success of reintroduction efforts.
Study area
Lowland tapirs were reintroduced to the Guapiaçu Ecological Reserve in Rio de Janeiro state, south-east Brazil (Fig. 1). The reserve is a 10,000 ha private protected area partially overlapping with Três Picos State Park, the largest protected area in the state (65,113 ha; INEA, 2015). It is located within a complex and varied landscape that includes secondary vegetation at various successional stages, reforested areas, agricultural fields, wetlands and pasture. The original vegetation of the region is classified as Dense Ombrophilous Forest and the climate is tropical, with a rainy summer and dry winter (Azevedo et al., Reference Azevedo, Camara, Francelino, Pereira and Leles2018). The dry season is May–September and the wet season October–April. Mean annual total rainfall is 2,050 mm, with monthly values of 59–338 mm. Mean annual temperature is 21.9 °C, January is the warmest month (25.3 °C) and July the coldest month (17.9 °C; Azevedo et al., Reference Azevedo, Camara, Francelino, Pereira and Leles2018). A 0.88 ha fenced enclosure was built in a reforested area of the Reserve to acclimatize the tapirs prior to their release (Galliez et al., Reference Galliez, Zamboni, Macedo, Di Martino, Rosas, Melletti, Reyna-Hurtado and Medici2024); it incorporated a small lake and a single shelter, which was occasionally used simultaneously by the individuals under controlled acclimatization conditions.
Location of protected areas and camera traps, and acclimatization pen and release points 1 and 2 for the reintroduced population of the lowland tapir Tapirus terrestris in the Guapiaçu Ecological Reserve, Rio de Janeiro, Brazil.

Methods
We selected 14 tapirs (7 males, 7 females) for reintroduction into the Guapiaçu Ecological Reserve from six ex situ conservation institutions (zoos and breeding facilities; Table 1) between December 2017 and October 2021. At the time of release, mean age was 7.0 ± SE 5.9 years (range 1.2–19.9 years). Tapirs were selected based on veterinary health screening, genetic and reproductive management criteria, behavioural suitability for release and institutional availability, following established reintroduction protocols (Galliez et al., Reference Galliez, Zamboni, Macedo, Di Martino, Rosas, Melletti, Reyna-Hurtado and Medici2024). All individuals were born in captivity in Brazilian ex situ institutions. We followed the soft release method whereby the tapirs were gradually acclimatized to their new environment in the enclosure, where they received supplemental food whilst learning to forage for themselves and survive in the wild (IUCN/SSC, 2013). The tapirs were protected in the enclosure for 88.3 ± SE 48.2 days. They were managed in six release groups (A–F; Table 1), with a maximum of four individuals acclimatized simultaneously. The acclimatization period varied dependent on an evaluation of the body condition, health and behaviour of each tapir by the team of biologists and veterinarians. Following the acclimatization period, each group was released together at a single release site. Groups were released directly from the acclimatization pen or were transported and released in one of three different areas of the Reserve, 2–4 km from the enclosure (Fig. 1; Galliez et al., Reference Galliez, Zamboni, Macedo, Di Martino, Rosas, Melletti, Reyna-Hurtado and Medici2024). Release sites were selected based on habitat suitability, water availability, low human disturbance and spatial planning considerations.
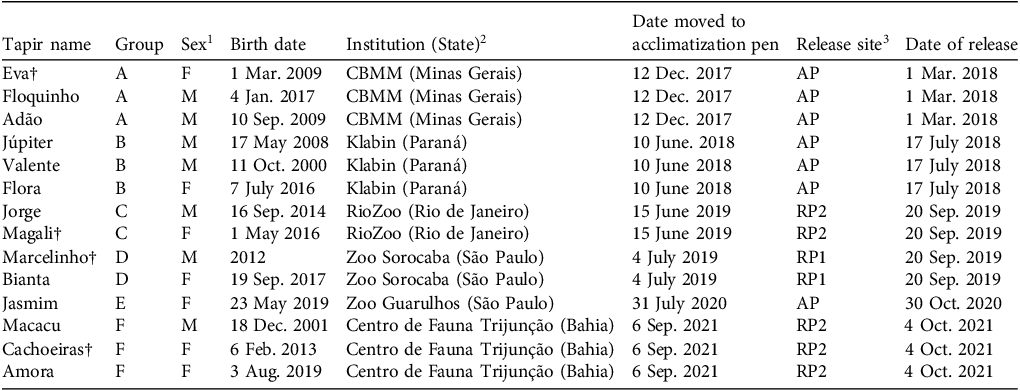
Lowland tapir Tapirus terrestris individuals reintroduced in six small groups to the Guapiaçu Ecological Reserve, Rio de Janeiro, Brazil (Fig. 1), from 2017 to 2021, with sex, birth date, conservation institution of origin, date moved to acclimatization pen, release site and date of release.

1 F, female; M, male.
2 CBMM, Companhia Brasileira de Metalurgia e Mineração.
3 AP, acclimatization pen site; RP1, release point 1; RP2, release point 2.
† Died during the reintroduction programme.
We monitored tapirs until February 2022 using camera traps distributed evenly across the Reserve and connecting landscape (Fig. 1). A camera trap was also located inside the fenced area to monitor tapirs during the acclimatization process. We established 38 survey points located > 500 m apart along predefined trails. We installed a single camera trap (Bushnell Core 119938C, Bushnell Core 119936C or Bushnell NatureView HD; Bushnell, USA) mounted on a tree 45 cm above the ground and facing the trail. We programmed the camera traps for continuous operation and no baits were used. The total sampling effort was 13,890 trap-days. We identified tapirs from individual characteristics such as size, scars, tail, collars and ear tags. We included records of unidentified tapirs in our analysis if they were separated by a minimum interval of 1 h and were considered to be independent (Carbajal-Borges et al., Reference Carbajal-Borges, Godínez-Gómez and Mendoza2014).
We tested the effect of length of time since release on the diurnality (proportion of records during daylight hours) and activity level (number of hours the animal was active over a 24 h period) of the released tapirs during the first 30 months following reintroduction. Daylight hours were defined according to local sunrise and sunset times for each date in the study area. Crepuscular periods corresponded to 1 h before and 1 h after sunrise and sunset, and all records were classified accordingly prior to analysis. We collated data over 6-month intervals to ensure reliable estimates, and excluded individuals with < 50 records in each 6-month period from the analysis. Because the tapir groups were not reintroduced simultaneously (Table 1), we included estimates of activity levels derived from camera-trap images up to 30 months post-release to facilitate comparisons between at least two different tapir groups. We estimated tapir activity levels using the package Activity (Rowcliffe et al., Reference Rowcliffe, Kays, Kranstauber, Carbone and Jansen2014) in R 4.3.0 (R Core Team, 2023). We applied generalized linear mixed models (GLMM) to test the effects of post-release time (6-month periods from date of release), group, sex and season on diurnal activity and activity level. We included individuals as random effects using the glmmPQL function in the package MASS (Ripley et al., Reference Ripley, Venables, Bates, Hornik, Gebhardt, Firth and Ripley2013) in R. Beginning with the full model, which included all explanatory variables, we used ANOVA to select the most parsimonious model by comparing it with reduced models and the null model.
We used circular kernel density functions to describe the activity patterns of reintroduced tapirs (Oliveira-Santos et al., Reference Oliveira-Santos, Zucco and Agostinelli2013). We performed 1,000 bootstraps for all analyses to determine accurate estimators (Ridout & Linkie, Reference Ridout and Linkie2009). We compared tapir activity patterns during acclimatization and during the post-release period, and between dry and wet seasons. We quantified activity patterns using the package overlap in R to generate the overlap coefficient (Δ), which varies from 0 (no overlap) to 1 (total overlap; Meredith & Ridout, Reference Meredith and Ridout2021). We applied the Δ4 estimator as recommended for samples > 75 records (Oliveira-Santos et al., Reference Oliveira-Santos, Zucco and Agostinelli2013). We used the Watson U² test to assess the homogeneity of tapir activity patterns during acclimatization and post-release, as well as between sexes and seasons (Jammalamadaka & Sengupta, Reference Jammalamadaka and Sengupta2001).
Results
We obtained 8,680 records over 4 years of monitoring reintroduced tapirs and we identified 73.1% of individuals (6,374 records) from the camera-trap images; 2,565 were recorded during the acclimatization period and 6,115 after release. There were four deaths during the study, three females and one male, from different groups (Table 1).
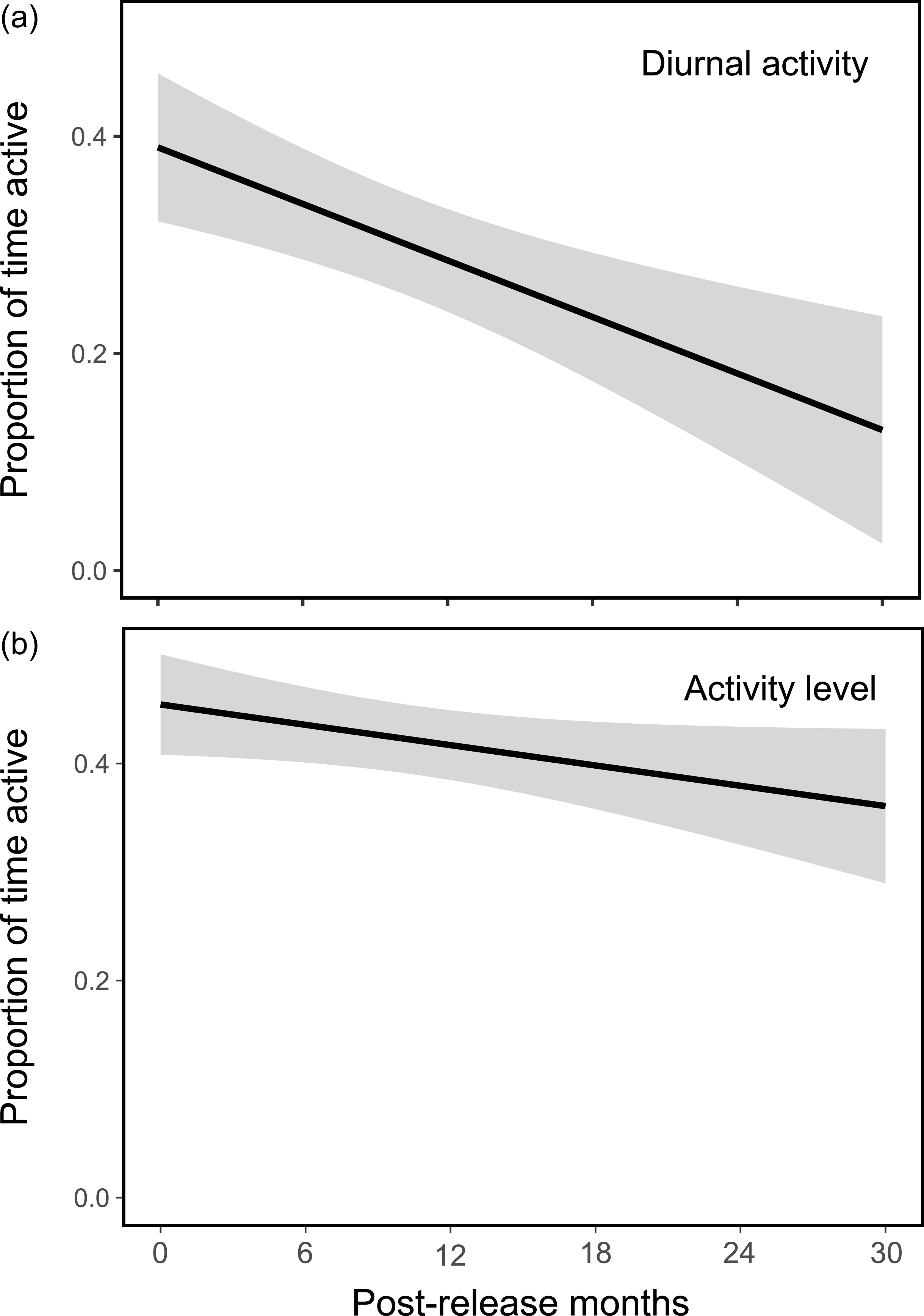
The diurnal activity of reintroduced tapirs decreased over time once they had been released from the acclimatization pen (β = −0.22, df = 33, P < 0.05) but varied amongst tapir groups (β = 0.11, df = 28, P < 0.05). During the acclimatization period, 36% of activity occurred during daylight hours but this proportion had decreased to 17% by 30 months post-release (Fig. 2a). Five out of six groups exhibited a reduction in diurnal activity after release compared to during their acclimatization period (Table 1). Sex and season had no significant effect on diurnal activity. Tapirs did not change their level of activity with time from release (β = −0.045, df = 33, P = 0.07), contrary to our initial hypothesis (Fig. 2b). Neither group identity (source of animals), sex or season had a significant effect on overall level of activity.
Estimated effect of time since release on (a) diurnal activity and (b) overall activity level of 14 reintroduced tapirs (Table 1) in the Guapiaçu Ecological Reserve (Fig. 1) between 2017 and 2022. Black lines represent model-predicted values derived from the fitted generalized linear mixed models, and shaded areas indicate the standard error of the estimates. The y-axis represents the proportion of time active (0 = no activity; 1 = theoretical maximum), with values < 0.5 reflecting the observed range of activity in the dataset

We found no significant difference in male and female activity patterns during the acclimatization period (94% overlap; U² = 0.09, P > 0.10; Fig. 3a) or after release (95% overlap; U² = 0.14, P > 0.10; Fig. 3b). However, activity patterns differed between the acclimatization and post-release periods (U² = 1.17, P < 0.001). We collected more images of tapirs during daylight hours during the acclimatization period compared to post-release, when tapirs were less active during daylight hours and more active at night (Fig. 3c). The overlap in activity levels between the acclimatization and post-release periods was 88%. The daily activity pattern also differed between the wet and dry seasons (U² = 0.13, df = 2, P < 0.001). The reintroduced tapirs were more active during the night-time in the wet season (October–April) and more active during the daytime in the dry season (May–September; Fig. 3d). The overlap in activity levels between the wet and dry season was 80%.
Activity patterns of reintroduced tapirs in the Guapiaçu Ecological Reserve showing overlap between: (a) females and males during acclimatization, (b) females and males post-release, (c) post-release and acclimatization periods, and (d) wet and dry seasons post-release. The y-axis represents kernel-estimated activity frequency over a 24 h period. The grey shaded area indicates the coefficient of overlap (Δ).

Discussion
The daily activity pattern of reintroduced tapirs varied across different stages of the reintroduction process, becoming more similar to natural patterns as time from release increased (Medici, Reference Medici2010; Cruz et al., Reference Cruz, Paviolo, Bó, Thompson and Di Bitetti2014). Post-release diurnal activity decreased over time but the overall activity levels remained constant. Activity patterns during the acclimatization period differed from those observed post-release, with no differences between sexes at any stage of the reintroduction. We found no effect of institution of origin or group identity on tapir activity pattern and conclude that the elevated levels of diurnal activity observed during the acclimatization period most likely reflected captive behaviour (Gilmore, Reference Gilmore2007).
We demonstrated that time since release was the primary factor influencing changes in the temporal behaviour of reintroduced tapirs. Whereas the time of day that the tapirs were active fluctuated, the number of active hours did not, as estimated from camera-trap records collected across different times of day. Despite significant behavioural modification during captivity, this was reversed in a relatively short time once tapirs were released and became free-living in natural conditions. The gradual shift in activity patterns towards those of natural populations highlights the profound impacts of reintroductions on routine behaviour and how animals adjust and learn throughout this process (Verzuh et al., Reference Verzuh, Heuer and Merkle2024). The increase in nocturnal and crepuscular behaviour patterns is thought to be a tactic to avoid contact with people (Bennie et al., Reference Bennie, Duffy, Inger and Gaston2014). Although the reintroduced tapirs in our study were accustomed to people during their periods of captivity and acclimatization, they were protected from domestic dogs. These dogs are known to chase wildlife and are predominantly diurnal so they may have disturbed the released tapirs during daylight hours (Young et al., Reference Young, Olson, Reading, Amgalanbaatar and Berger2011), leading the tapirs become active at other times to minimize the chances of an encounter (Silva-Rodríguez & Sieving, Reference Silva-Rodríguez and Sieving2012). These behavioural changes would have imposed a high energy cost on the reintroduced tapirs (Houston et al., Reference Houston, Prosser and Sans2012), particularly when predator avoidance is constant and therefore affects foraging behaviour (Leblond et al., Reference Leblond, Dussault and Ouellet2013).
In captivity, tapirs are accustomed to frequent interactions with their keepers and experience positive reinforcement associated with human presence, particularly during diurnal feeding routines. These stereotypical behaviours are imposed by captive conditions but may encourage reintroduced tapirs to search for people once released, which may increase the risk of roadkill and encounters with domestic dogs. These behaviours present short-term challenges to population establishment after reintroduction. (Carlstead et al., Reference Carlstead, Mellen and Kleiman1999; Kierulff, Reference Kierulff2000). In addition to predator avoidance, reintroduced animals from captive stock face other new challenges in learning how to survive in an unfamiliar environment, foraging and finding sufficient food, and interacting with conspecifics (Shier, Reference Shier, Berger-Tal and Saltz2016; Swaisgood & RuizMiranda, Reference Swaisgood, Ruiz-Miranda, Caro and Clutton-Brock2018). The shift in behaviour toward more natural patterns, specifically the decrease in diurnal activity between the acclimatization and post-release periods, indicates that the released tapirs successfully adjusted to their new environment and avoided daytime anthropogenic disturbances. Environmental enrichment activities conducted in captivity, such as providing food during crepuscular or nocturnal periods, ensuring access to water sources and wallows, and maintaining large enclosures that promote exploratory behaviour, can accelerate this process before reintroduction. A programme of negative reinforcement during preparation for reintroduction, including aversive conditioning to human presence (e.g. use of auditory stimuli or deterrent elements associated with people), while increasing the difficulty of handling during acclimation, could also be implemented to reduce problems related to contact with people post-release.
Thermoregulation is a key factor shaping the activity patterns of large mammals (Mourão & Medri, Reference Mourão and Medri2007). As a result of their size, tapirs are particularly affected by ambient temperature and avoid activity during the hottest hours of the day (Oliveira-Santos et al., Reference Oliveira-Santos, Machado-Filho, Tortato and Brusius2010). After release, reintroduced tapirs became more nocturnal, with activity peaks during twilight hours, similar to natural populations in other regions (Oliveira-Santos et al., Reference Oliveira-Santos, Machado-Filho, Tortato and Brusius2010; Wallace et al., Reference Wallace, Ayala and Viscarra2012; Medici & Fantacini, Reference Medici and Fantacini2022). This was more marked in the wet season when temperatures were higher: more camera-trap images were recorded in the hottest hours of the day (09.00–14.00) during the dry and coldest season, with more images recorded at night, particularly at twilight, in the warm and wet season. Although females are 20–50% larger than males (Medici, Reference Medici2010) and could therefore be more susceptible to thermal stress, this size difference did not influence tapir activity, consistent with other studies (Medici & Fantacini, Reference Medici and Fantacini2022). Males and females showed a high degree of overlap in activity patterns during the acclimatization period and post-release, indicating similar behavioural adjustments.
The timing of tapir activity changed post-release but overall levels of activity remained the same. The number of camera-trap images recorded during daylight hours, particularly during the hottest hours of the day, was higher in the dry season than in the wet season when temperatures were higher. However, there were more hours of daylight in the wet season, which compensated for this avoidance of the hottest time of day and resulted in no reduction in overall activity level. It is possible that the intrinsic stress of the reintroduction process could increase activity levels but we did not observe this in the reintroduced tapirs in our study. Despite the modulation of activity patterns, the consistency in activity levels demonstrates that acclimatization was important in reducing reintroduction stress.
Little is known about the ability of reintroduced animals to adjust their activity patterns post-release, although other studies have demonstrated a tendency for behaviour to converge with that shown by wild conspecifics (Converse et al., Reference Converse, Moore and Armstrong2013). This adaptability is important and serves as an indicator of short-term success in reintroduction projects. The behavioural plasticity of tapirs provides flexibility for management plans, especially those based on captive animals.
Author contributions
Fieldwork: MA; data collection: MA, JSM, MG; statistical analysis: MA; writing: MA, MG; revision: all authors.
Acknowledgements
We thank the Laboratório de Ecologia e Manejo de Animais Silvestres/Instituto Federal do Rio de Janeiro team for their help in designing the research, fieldwork, data analysis and discussions; J. Pires, A. Roque and A. M. Jansen for veterinary advice; J. Pires for sedating, veterinary screening and animal handling; Refauna and our partners Guapiaçu Ecological Reserve and Instituto de Ação Socio Ambiental for support; Companhia Brasileira de Metalurgia e Mineração, Klabin, Prefeitura do Rio de Janeiro, and RioZoo, Zoológico de Sorocaba, Zoológico de Guarulhos for providing the tapir; and Instituto Estadual do Ambiente/Rio de Janeiro, Instituto Chico Mendes de Conservação da Biodiversidade and Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis for environmental licences. This work was supported by Fundação Grupo Boticário de Proteção à Natureza (Program number: 0010/2014); Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (grant number: E26/010/001645/2014); Furnas Eletrobras (Antologia Project) and Petrobras (Guapiaçu Project), and was carried out by Ação Socioambiental, Instituto Federal do Rio de Janeiro, and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior.
Conflicts of interest
None.
Ethical standards
This research abided by the Oryx guidelines on ethical standards and those of the Institutional Animal Care and Use Committee of Instituto Federal do Rio de Janeiro (CEU-IFRJ 001/2016). Capture and handling of reintroduced tapirs were carried out in accordance with licences from the appropriate environmental agencies (INEA No. 002/2018; IBAMA No. 43795-9).
Data availability
The data used in this article are available upon reasonable request from the corresponding author.



 Open access
Open access