Obesity and its complications are a global health concern. The mother’s ability to accumulate nutrients rises during pregnancy. However, nutritional excess during this period can easily cause obesity(Reference Koletzko, Godfrey and Poston1). Obesity during pregnancy raises the risk of type II diabetes mellitus and gestational diabetes, and obese females during pregnancy are three to four times more likely to acquire gestational diabetes than normal females(Reference Kong, Chen and Gissler2,Reference Catalano and Shankar3) . According to the International Diabetes Federation, as of 2024, the global population of individuals with diabetes is estimated to be approximately 589 million. Notably, gestational diabetes has a prevalence such that it affects 1 in 5 live births(Reference Magliano and Boyko4). Gestational diabetes causes abnormal glucose and lipid metabolism in females during pregnancy, increasing the risk of obstructed labour, caesarean section and stillbirth(Reference Alejandro, Mamerto and Chung5,Reference Creanga, Catalano and Bateman6) . In the long term, gestational diabetes can even have adverse effects on offspring across generations. Numerous studies have demonstrated that offsprings born to mothers with gestational diabetes are more likely to acquire obesity and diabetes in childhood or adulthood than those born to healthy during pregnancy(Reference Lowe, Scholtens and Kuang7,Reference Mantzorou, Papandreou and Pavlidou8) . Reduced maternal fat deposition and improved insulin sensitivity may thus be an effective method to improve maternal and offspring health. Currently, pioglitazone, rosiglitazone and metformin are widely used to treat obesity and its related complications, however, these drugs are often associated with adverse effects such as cardiopulmonary insufficiency, kidney or heart failure and diarrhoea, and nutritional interventions may be one of the effective measures to treat various types of pregnancy disorders(Reference Johns, Denison and Norman9–Reference Arya, Hansen and Wild11).
Recent studies have demonstrated that consuming fiber-rich medicinal mushrooms on a regular basis can help avoid obesity and complications and maintain health(Reference Ishara, Buzera and Mushagalusa12,Reference Chaturvedi, Agarwal and Gupta13) . Lentinus edodes, Ganoderma lucidum and Agrocybe cylindracea are medicinal mushrooms that reduce obesity and inflammation and improve insulin sensitivity in mice fed a high-fat diet (HFD)(Reference Maschio, Gentil and Caetano14–Reference Zhu, Huang and Liu16). Lentinus edodes is one of the most widely sourced fungi, that is rich in dietary fiber, polysaccharides, ergosterin and other bioactive substances, with hypoglycemic, antioxidant, antibacterial and other effect(Reference Du, Fang and Zhang17,Reference Gong, Wang and Liu18) . Gong et al.(Reference Gong, Wang and Liu18). reported that the infusion of Lentinus edodes polysaccharide in diabetic mice reduced fasting blood glucose and improved oxidative stress by activating the nuclear factor erythroid 2-related factor 2 (Nrf2)/haeme oxygenase-1 (HO-1) signaling pathway, thus, improving their insulin sensitivity. However, whether the supplementation of Lentinus edodes and its fermentation products during pregnancy and lactation can improve maternal and offspring insulin sensitivity remains elusive. Fermented Lentinus edodes (FLE) is a product of the fermentation of Lentinus edodes using cellulase, alkaline protease and a complex of probiotics. Studies have demonstrated that mushrooms can reduce the content of anti-nutritional factors and increase the content of nutrients such as crude protein, crude polysaccharide, and lactic acid via the synergistic fermentation of probiotics and protease(Reference Jabłońska-Ryś, Sławińska and Skrzypczak19–Reference Zhang, Wang and Wang21). In this study, we constructed maternal mice gestation and lactation overnutrition model using HFD to clarify whether FLE alleviates insulin resistance in maternal mice during gestation and lactation and in offspring, and to explore the mechanism.
Materials and methods
Experimental design and feeding management
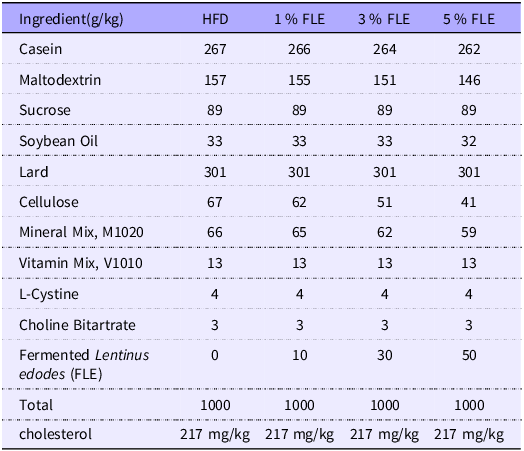
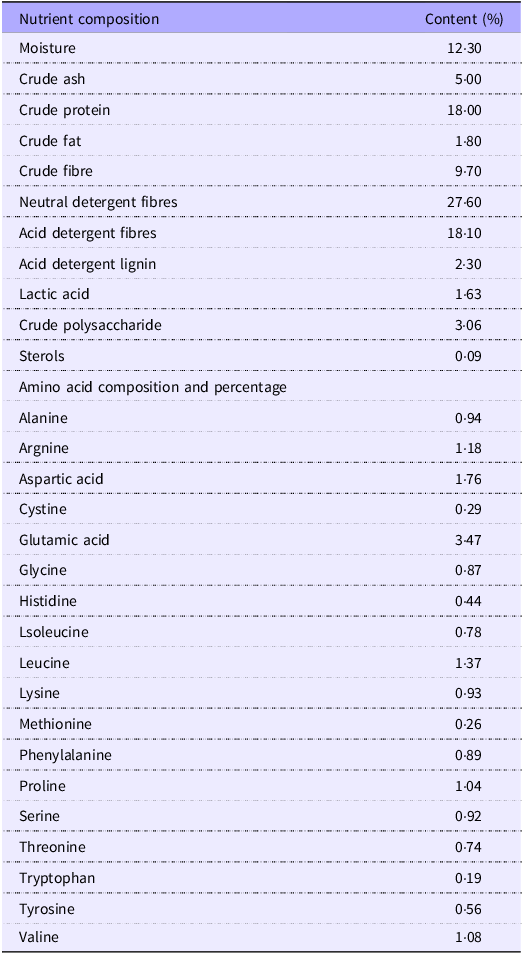
In this experiment, eighty 10-week-old female C57BL6/J mice of the specific pathogen free (SPF) inbred strain and forty 10-week-old male C57BL6/J mice were randomly selected, and the males and females were mated together in cages in a ratio of 1:2 at 20.00 in the evening, and the females were observed for the formation of vaginal plugs at 08.00 am the next day. If a vaginal plug was present, the female mice were recorded as 0·5 d of gestation (G 0·5 d), and at the same time the body weight of the mice at the beginning of gestation was recorded. The experiment procedure was illustrated in online Supplementary Figure S1. Pregnant mice were randomly divided into four groups (n 20 mice/group): HFD group (fed a 60 % kcal HFD); 1 %FLE group (fed the HFD and 1 % FLE); 3 % FLE group (fed the HFD and 3 % FLE); 5 % FLE group (fed the HFD and 5 % FLE). HFD was commissioned from, TROPHIC Animal Feed High-Tech Co. Ltd. under product number TP23302 and the composition of the diet is shown in Table 1. FLE is a self-developed product of Yuehao Biotechnology (Guangzhou) Co., Ltd. and is not an in-house laboratory produced sample. The preparation process is as follows: firstly, the cap and foot of Lentinus edodes are properly crushed and mixed well, and then lactobacilli, cellulase and alkaline protease are added to the pre-treated Lentinus edodes residue for fermentation. Finally, the fermentation products were dried and sterilised to obtain FLE. In this study, FLE moisture determination was referred to GB/T 6435-2014, ash crude fraction determination was referred to GB/T 6438-2007, crude protein determination was referred to GB/T 6432-2018, crude fat determination was referred to GB/T 6434-2006, crude fibre determination was referred to GB/T 6434-2006, neutral detergent fibre was referred to GB/T 20806-2006, acid detergent fibre reference NY/T 1459-2007, acid detergent lignin determination reference GB/T 20805-2006, lactic acid determination reference GB/T 23877-2009, sterol determination reference GB/T 25223-2010, crude polysaccharide determination reference SN/T 4260-2015, amino acid Determination referred to GB/T 18246-2019. Detailed nutritional composition of FLE is shown in Table 2.
Experimental diet composition

Nutritional composition of fermented Lentinus edodes

During the experimental period, each pregnant mice was housed in an individual cage and was provided with adequate diet and water. The indoor temperature of the animal house was always 24 ± 1°C, the relative humidity was 55–65 %, and a light-dark cycle of 12 h. During gestation, the body weights and feed intake of dams were recorded at gestation 7·5 d (G 7·5 d), G 14·5 d, and G 19·5 d. During lactation, the body weights and feed intake of the dams were recorded at lactation 7·5d (L 7·5 d), L 14·5 d, and L 21·5 d. The litter weights and individual weights of the pups were recorded at L 0·5 d, L 7·5 d, L 14·5 d, and L 21·5 d, without differentiating between male and female pups.
Experiment A: At G 15·5 d, after fasting for 12 h, the pregnant female mice were followed by an oral glucose tolerance test (OGTT) (n 6–9 mice/group). Additionally, At G 15·5 d, after fasting for 8 h, the pregnant female mice were followed by an intraperitoneal insulin tolerance test (IPITT) (n 6–8 mice/group). At G 18·5 d, after fasting for 12 h, the OGTT pregnant mice were anesthetized to collect blood from the retro-orbital and then euthanized by cervical dislocation(Reference Peng, Xiong and Cui22) (n 7 mice/group).
Experiment B: At L 15·5 d (delivery diary was L 0·5 d), after fasting for 12 h, the lactating female mice were followed by an OGTT (n 6–9 mice/group). At L 16·5 d, after fasting for 8 h, the lactating female mice were followed by an IPITT (n 6–8 mice/group). At L 21·5 d, after fasting for 12 h, the mice were anesthetized to collect blood from the retro-orbital and then euthanized by cervical dislocation (n 7 mice/group).
Experiment C: At L 21·5 d, after fasting for 12 h, the offspring female mice were followed by an OGTT (n 7–8 mice/group). Additionally, at L 21·5 d, after fasting for 6 h, the offspring female mice were followed by an IPITT (n 6–7 mice/group). At L 21·5 d, after fasting for 6 h, the offspring male mice were anesthetized to collect blood from the retro-orbital and then euthanized by cervical dislocation (n 7 mice/group).
Oral glucose tolerance test (OGTT) and intraperitoneal insulin tolerance test (IPITT)
Blood glucose was measured using a glucometer (ACCU-CHEK, Roche, Germany). For the oral glucose tolerance test (OGTT), the mice were fasted for 8 h and given 2 g/kg glucose orally. For the intraperitoneal insulin tolerance test (IPITT), the mice were fasted for 8 h and injected intraperitoneally with 1 IU/kg of insulin (offspring mice were fasted for 6 h)(Reference Tukhovskaya, Shaykhutdinova and Pakhomova23).
Sample collection
The placenta and fetus were collected from pregnant mice on gestational day 18·5 and then weighed subsequently. The liver, brown fat, and perirenal fat of the pregnant mice, and the jejunum of both the maternal and offspring mice were harvested and fixed in 4 % paraformaldehyde solution and then embedded in paraffin. Liver and visceral fat (brown fat, gonadal fat) were collected and stored at −80°C. The fetal homogenization protocol used here was derived from Peng et al.(Reference Peng, Xiong and Cui22), The fetus and placenta were ground into a fine powder in a porcelain mortar pre-cooled with liquid nitrogen. Subsequently, the powdered sample was homogenized in 500 µl of RIPA protein lysis buffer. Finally, the homogenate was centrifuged at 18 000 revolutions per minute (rpm) for 5 min.
Biochemical analysis
Serum and tissues levels of glucose (GLU) were detected using a commercial diagnostic kit (Rongsheng Biotechnology Co. Ltd, Shanghai, China). Triacylglycerol (TG), total cholesterol (TCHO), high-density lipoprotein cholesterol (HDL-C), and low-density lipoprotein cholesterol (LDL-C) levels in serum and tissues were measured using a commercial enzyme kits (Jiancheng Biological Engineering Research Institute, Nanjing, China). insulin levels in Serum were tested using a commercial ELISA kit (MS100, EZassay Biotech Ltd. Shenzhen, China). The formula for calculating Homeostatic Model Assessment of Insulin Resistance (HOMA-IR). was equal to fasting blood glucose multiplied by fasting blood insulin, and except for 22·5(Reference Tukhovskaya, Shaykhutdinova and Pakhomova23,Reference Kearney, Perron and Marc24) .
Quantitative real-time PCR
The total RNA of liver and adipose tissues were extracted by the Transzol reagent (TransGen Biotech Co. Ltd, Beijing, China). The RNA concentration was determined using a Micro UV-Vis spectrophotometer. A total of 500 ng of RNA was reversed and synthesized to cDNA by the Transcript reagent (TransGen Biotech Co. Ltd, Beijing, China). The mRNA gene expression was detected using a CFX Real-Time PCR System (Bio-Rad, USA)(Reference Gong, Wang and Liu18,Reference Peng, Zhou and Hong25) . β-actin was used as the internal reference gene, and the relative mRNA expression was quantified by the 2–ΔΔct method. The specific gene primer sequences are shown in online Supplementary Table S1.
Western blotting
The total proteins of maternal and offspring Liver were extracted using a RIPA protein lysate (Servicebio technology Co.Ltd, Wuhan, China). The protein concentrations were determined with BCA assay kit(Reference Gong, Wang and Liu18) (Beyotime Biotechnology Co. Ltd, Shanghai, China). Tissue sample proteins were separated by SDS-PAGE gels and repositioned to a polyvinylidene fluoride (PVDF) membranes. After 2 h of incubation in the blocking solution, PVDF membranes were transferred with primary antibodies (1:1000) and kept overnight at 4℃. The primary antibodies were included: PI3K (Affinity, AF6241, China), P-PI3K [(1:500) Affinity, AF3241, China], AKT (Affinity, AF6259, China), P-AKT (Affinity, AF0016, China) and β-actin (Affinity, AF7018, China). The PVDF membranes were incubated with the anti-rabbit IgG-HRP as the secondary antibody [(1:5000), Abclonal, AS014, China] for 1 h at room temperature. The protein bands were analyzed using Image J software.
Histological analysis
Paraffin sections of white fat, brown fat and jejunum were stained with hematoxylin-eosin (H&E), while frozen sections of liver were stained with Oil Red O. These staining procedures were completed by Powerful Biology Co. Ltd, (Wuhan, China). A microscopic (Leica DM500, Germany) were observed the size of white adipose adipocytes, brown adipose adipocytes, and liver lipid dropletsunder(Reference Chen, Lu and Li26). The jejunum villi height, crypt depth, villi width, epithelial cell thickness, and intestinal wall thickness were measured using an Image View software(Reference Özdoğan, Wellmann and Paksuz27).
Statistical analysis
Figures were drawn by One-way ANOVA using GraphPad Prism version 8.0.1 software. Table data were analyzed with IBM SPSS Statistics 26.0 software, assessing significance through one-way ANOVA and linear regression (using the Ordinary Least Squares method). All data are presented as means (SEM). P < 0·05 were considered statistically significant. R values denoting correlation coefficients.
Results
FLE improves insulin sensitivity in pregnant mice
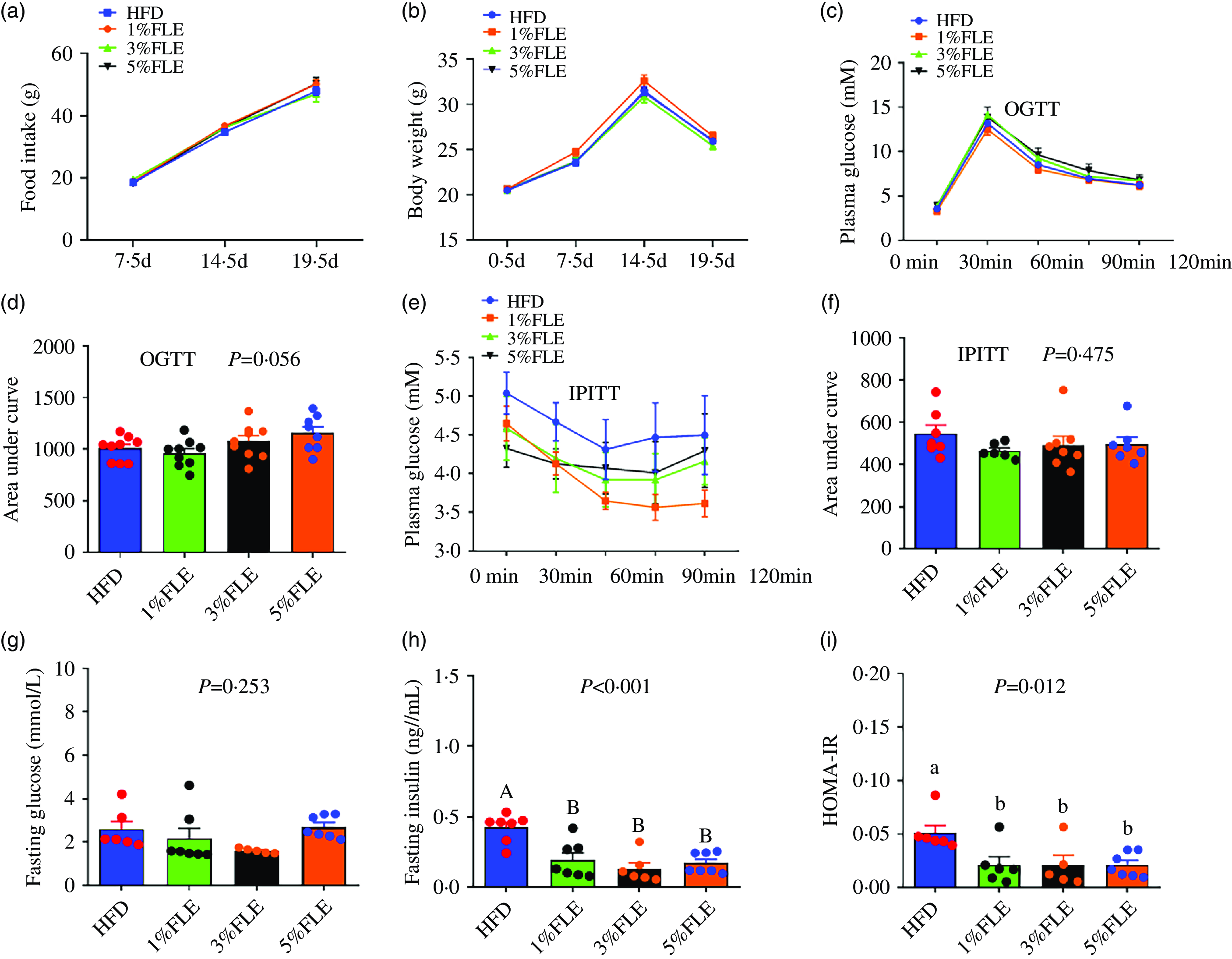
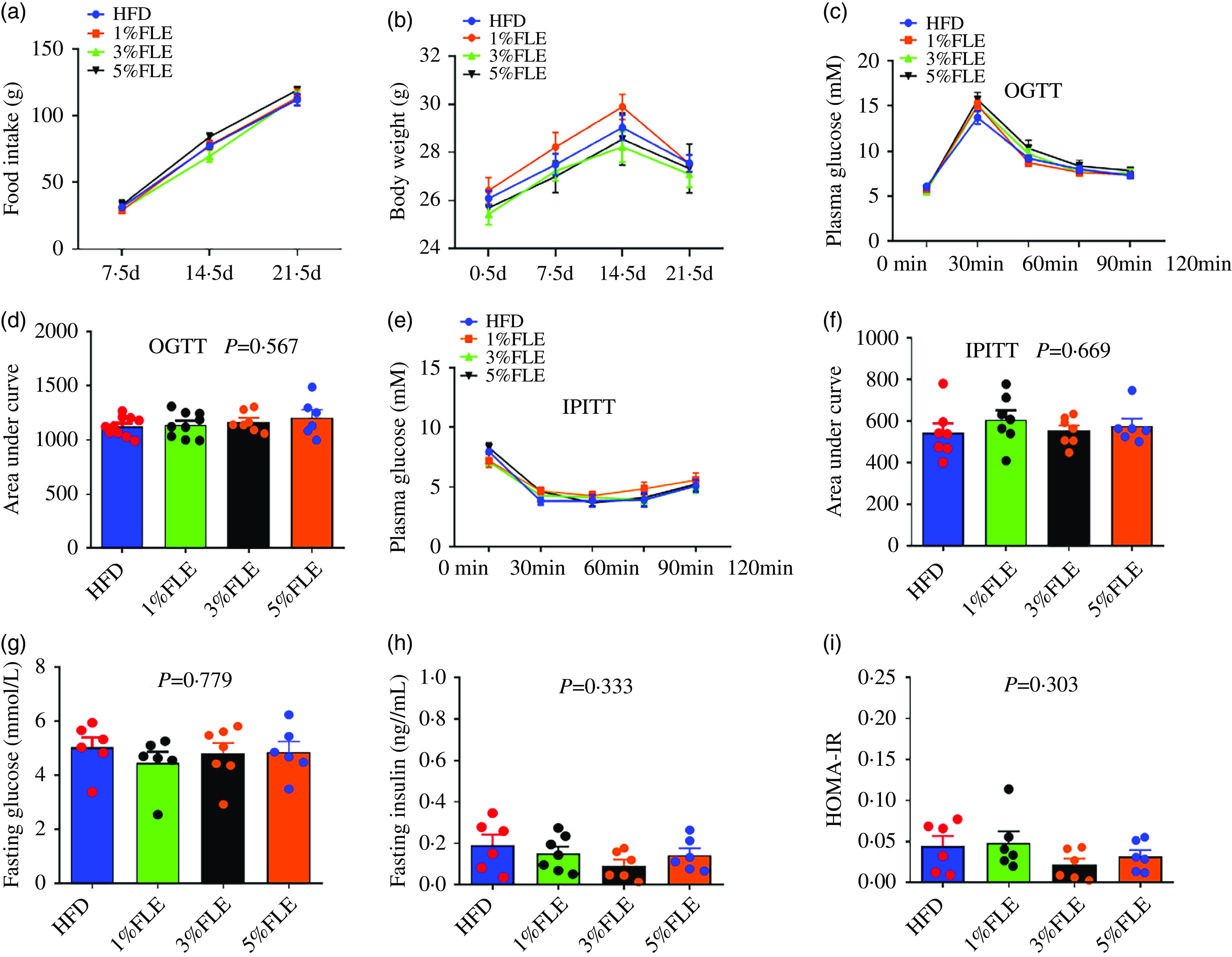
To investigate the effects of FLE on body weight, food intake and insulin sensitivity in pregnant mice, they were fed with different doses of FLE from P 0·5 d. In pregnant mice, the FLE had no significant effect on food intake, body weight, oral glucose tolerance test (OGTT), intraperitoneal insulin tolerance test (IPITT), area under the curve and fasting blood glucose compared to HFD (Figure 1(a)–(g)); however, it significantly reduced fasting insulin levels and HOMA-IR in pregnant mice (P < 0·05) (Figure 1(h)–(i)).
Effect of FLE on weight, food intake and insulin sensitivity of pregnant mice (a): Food intake; (b): Body weight; (c): Oral glucose tolerance test (OGTT); (d): Area under the curve from OGTT; (e): Intraperitoneal insulin tolerance test (IPITT); (f): Area under the curve from IPITT; (g): Fasting blood glucose; (h): Fasting insulin; (i): HOMA-IR; One-way ANOVA, Different capital letters represent extremely significant differences, P < 0·05, n 7–9/group.

FLE reduces maternal lipid deposition during pregnancy and promotes fetal development
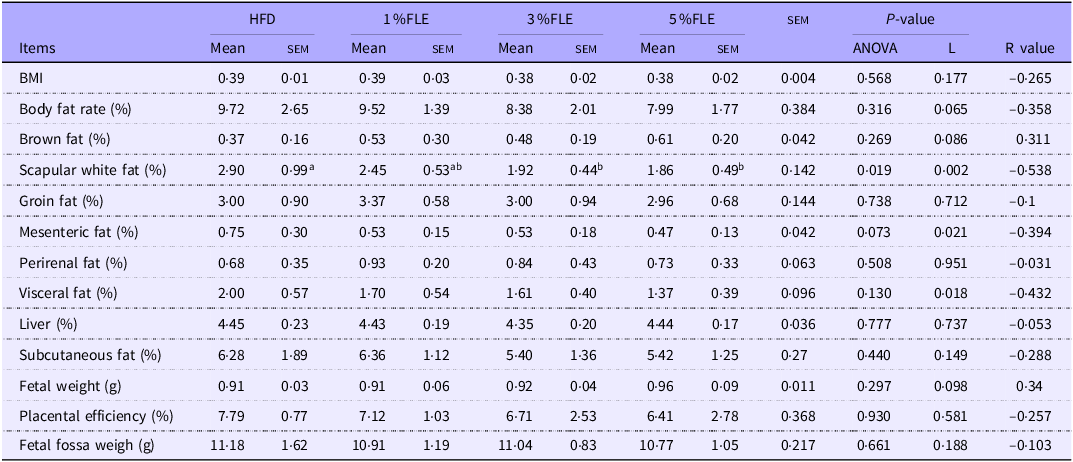
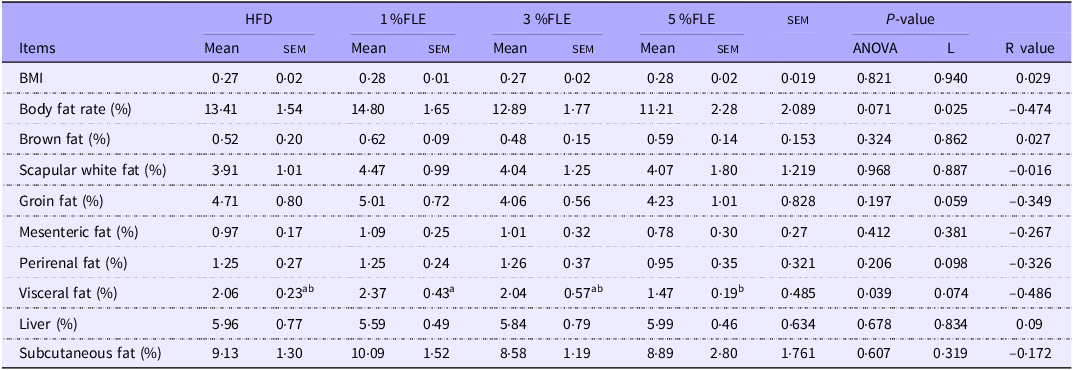
To analyse the effect of FLE on maternal health and fetal development, pregnant mice were euthanized at P 18·5 d. In FLE-fed mice, scapular white fat was linearly reduced (P < 0·05), and mesenteric fat exhibited a linear decreasing trend (P = 0·073) (Table 3). In addition, the effect of FLE on intestinal morphology in pregnant mice was assessed. It was observed that 5 % FLE had no significant effect on jejunal crypt depth, villus height, villus height/crypt depth, villus width, epithelial thickness and intestinal wall thickness in pregnant mice (online Supplementary Figure S3A).
Effects of FLE in pregnant mice on maternal lipid deposition and foetal development

Different lowercase letters in the table indicate P < 0·05, n 7/group, L stand for Linear Regression, R value represents the correlation coefficient in linear regression.
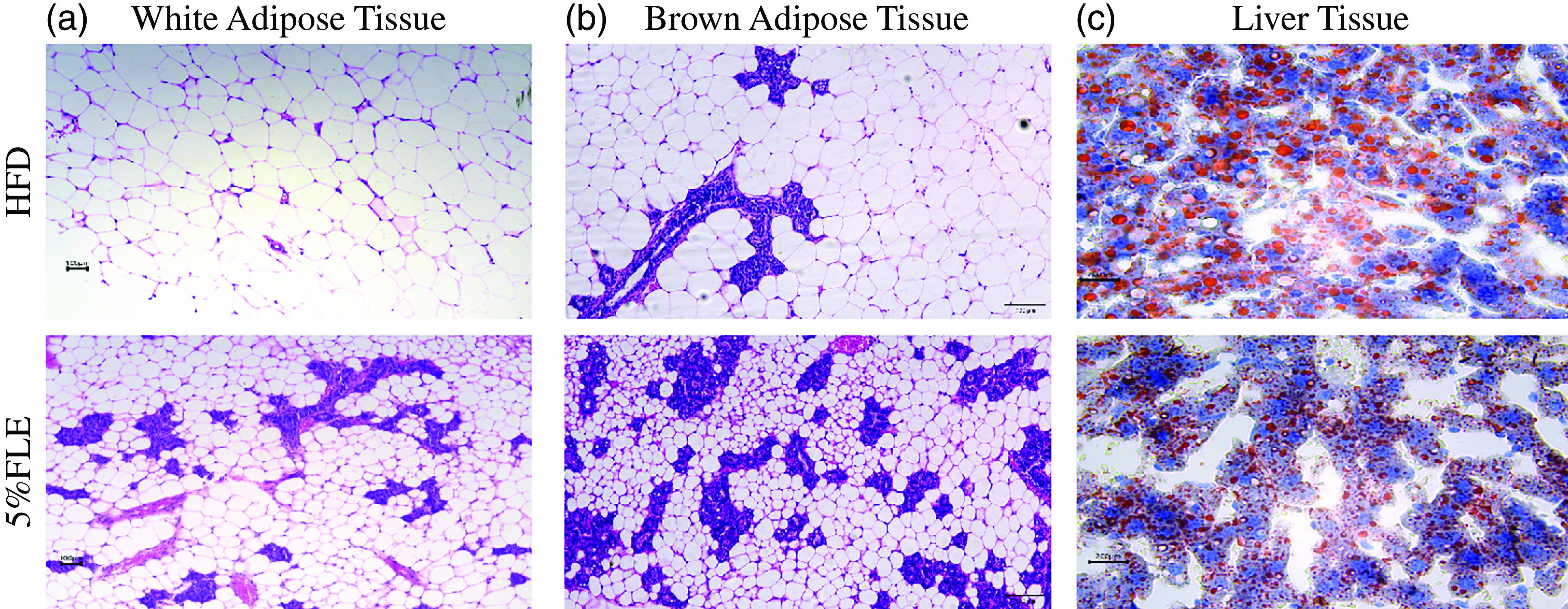
FLE reduces white fat size and inhibits ectopic deposition of lipids in the liver
Histological analysis of the fat and liver of pregnant mice revealed that 5 % FLE reduced the size of perirenal white and brown adipocytes and the size and number of lipid droplets in the liver (Figure 2(a)–(d)). Further testing of the serum lipid content of pregnant mice revealed that 5 % FLE had no significant effect on serum TG, TCHO, HDL-C and LDL-C (online Supplementary Figure S2A). In addition, testing of genes related to adipose tissue inflammation in pregnant mice revealed that 5 % FLE had no significant effect on white adipose inflammatory genes [interleukin (IL)-1β, IL-6, IL-10, tumour necrosis factor-α (TNF-α)] and macrophage makes [mouse EGF-like module-containing mucin-like hormone receptor-like 1(F4/80), monocyte chemoattractant protein-1 (MCP1)]mRNA expression (online Supplementary Figure S4A). Furthermore, 5 % FLE did not impact the expression of the brown adipose inflammatory factors (IL-1β, IL-6, IL-10 and TNF-α) and macrophage makes (MCP1, F4/80) mRNA in pregnant mice (online Supplementary Figure S4B).These results indicate that 5 % FLE has potential positive effects on regulating fat metabolism and does not cause significant inflammatory responses or other adverse effects.
Effect of FLE on lipid deposition of pregnant mice. (a)–(b): Hematoxylin and eosin staining of perirenal adipose and brown adipose tissue; (c): Oil red-O staining of liver, Scale bar, 50 µm(400X).

The activation of the hepatic phosphoinositide-3-kinase-protein kinase (PI3K/AKT) signaling pathway in the liver of pregnant mice by FLE improves hepatic lipid metabolism
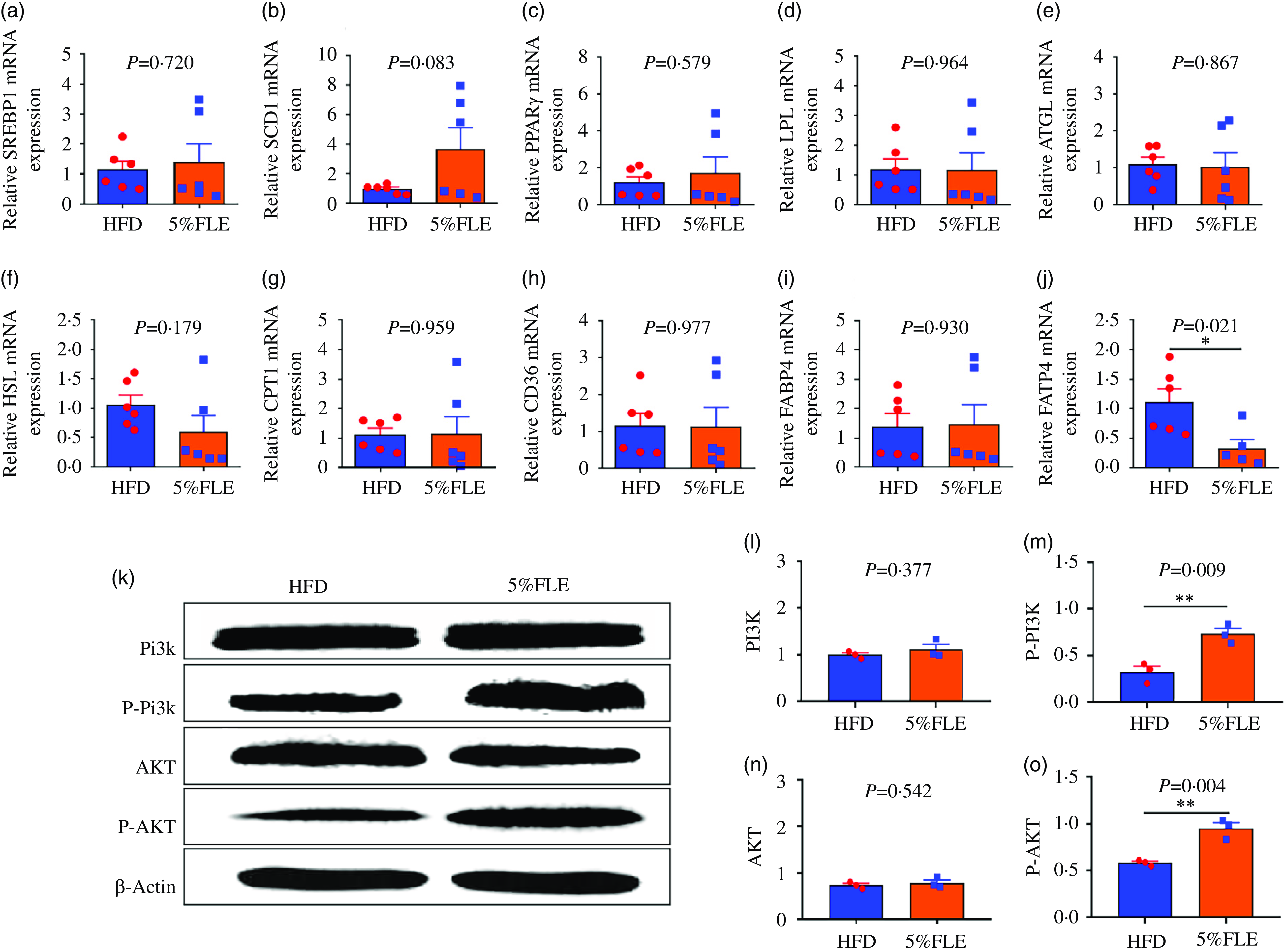
Genes related to hepatic lipid metabolism were assessed, and it was revealed that the addition of FLE during pregnancy had no significant effect on lipid synthesis genes [Sterol reg-ulatory element binding protein-1c (SREBP1c), stearoyl coenzyme A desaturase-1 (SCD1) and peroxisome proliferator-activated receptorγ (PPARγ)], lipid catabolic genes [adipose TG lipase (ATGL), lipoprotein lipase (LPL), hormone-sensitive lipase (HSL)] and fatty acid oxidation gene [palmitoyltransferase 1A (CPT1)] and fatty acid transporter genes [fatty acid translocaseCD36 (CD36), fatty acid binding protein 4 (FABP4)] mRNA (Figure 3(a)–(i)) but significantly down-regulated the liver fatty acid transporter gene fatty acid transporter 4 (FATP4) mRNA (P < 0·05) (Figure 3(j)). PI3K/AKT is a key signaling pathway that regulates insulin sensitivity. Next, the expression of PI3K/AKT signaling pathway proteins was examined, and it was observed that FLE did not significantly alter the total PI3K and AKT protein levels in the livers of pregnant mice; however, the phosphorylation levels of PI3K and AKT proteins were significantly increased (P < 0·05) (Figure 3(k)–(m)).
Effect of FLE on lipid metabolism and PI3K/AKT signaling pathway in the liver of pregnant mice. (a)–(c): The mRNA expression of lipid synthesis related genes; (d)–(f): The mRNA expression of lipolysis genes; (g): The mRNA expression of fatty acid oxidase; (h)–(j): The mRNA expression of fatty acid transport genes; n 6/group, k: Western blot protein bands; n 3/group, l-o: PI3K, P-PI3K, AKT and P-AKT protein expression in the liver of pregnant, n 3/group. Student’s t test, *P < 0·05, **P < 0·01, HFD v. FLE.

FLE improves fetal glucolipid metabolism
Pregnant maternal mice were euthanized at G 18·5 d, and the fetus was isolated for homogenization and biochemical measurements. Five percent FLE reduced fetal homogenized glucose content (P < 0·05) (online Supplementary Figure S5A), and increased fetal homogenized TCHO content (P < 0·01) (online Supplementary Figure S5B), but had no significant effect on TG, HDL-C and LDL-C content (online Supplementary Figure S5C–E).
FLE does not significantly affect insulin sensitivity in lactating mice
During lactation, FLE had no significant influence on food intake, body weight, OGTT, IPITT, the area under the curve, fasting glucose, fasting insulin and HOMA-IR in lactating mice (Figure 4(a)–(i)).
Effect of FLE on weight, food intake and insulin sensitivity of lactating mice (a): Food intake; (b): weight; (c): OGTT; (d): Area under the curve from OGTT; (e): IPITT; (f): Area under the curve from IPITT; (g): Fasting blood glucose; (h): Fasting insulin; (i): HOMA-IR. P < 0·05, ns, no significant, n 6–9/group.

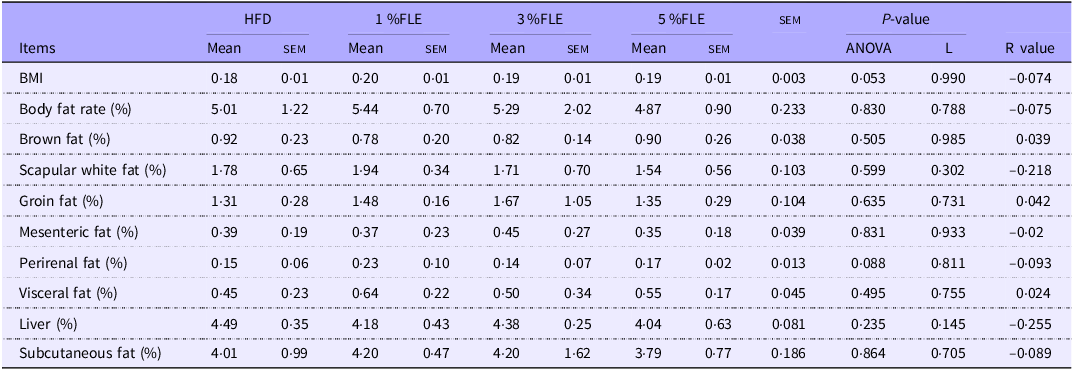
FLE reduces maternal lipid deposition during lactation
Lactating mice were executed at L 21·5 d. Analysis of body composition revealed a linear decrease in the visceral fat content of lactating mice with the addition of FLE (P < 0·05), and there was a trend of linear decrease in Body fat rate, with no significant effect on Body Mass index (BMI), brown fat, groin fat, perirenal fat, white scapular fat, mesenteric fat, subcutaneous fat and liver (Table 4). The serum lipid content of lactating mice indicated that FLE had no significant effect on TCHO, TG, HDL-C and LDL-C (online Supplementary Figure S2B). The effect of FLE on the intestinal health of lactating mice was further evaluated. It was observed that 5 % FLE had no significant effect on jejunal crypt depth, villus height, villus height/crypt depth, villus width, epithelial thickness and intestinal wall thickness in lactating maternal mice (online Supplementary Figure S3B).
Effect of FLE on lipid deposition in lactating mice

Different lowercase letters in the table indicate P < 0·05 and different uppercase letters indicate P < 0·01, n 7/group, L stand for Linear Regression, R value represents the correlation coefficient in linear regression.
FLE improves insulin sensitivity and promotes offspring growth
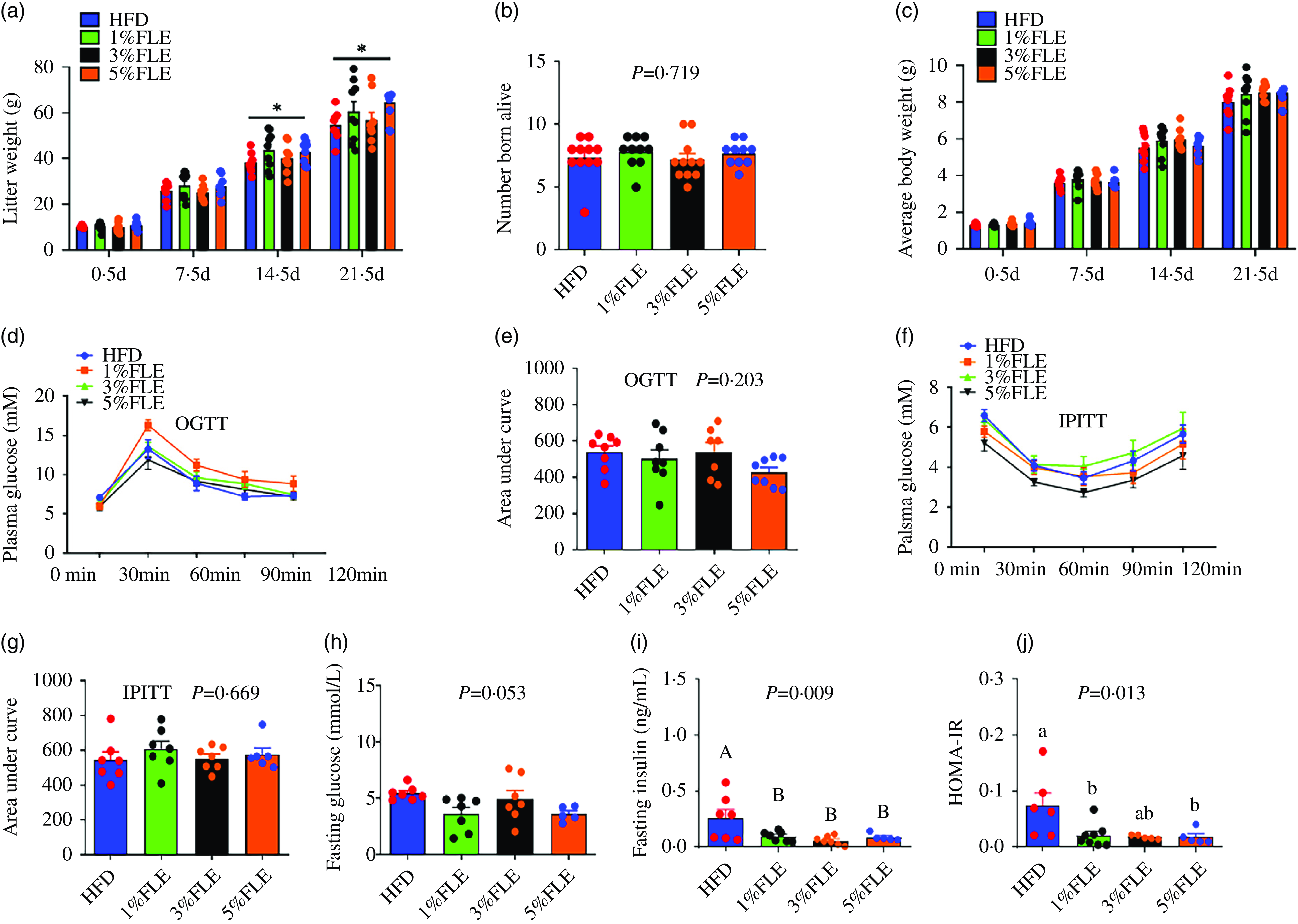
Analysis of the effect of FLE on offspring demonstrated that 5 % FLE significantly increased the weaning litter weight of offspring (P < 0·05) (Figure 5(a)), whereas that of the number of live born, 21·5 d offspring weight, OGTT, IPITT and area under the curve were unaffected (Figure 5(b)–(g)), however, there was a tendency to reduce fasting glucose (P = 0·053) (Figure 5(h)), and significantly reduced fasting insulin and HOMA-IR (P < 0·05) (Figure 5(i)–(j)).
Effect of FLE on growth performance and insulin sensitivity of offspring (a): litter weight; (b): number born alive; (c): average weight; (d): body weight of female litters; (e): body weight of female litters; (f): female offspring weight 21·5 d; (g): male offspring weight 21·5 days; (h): Fasting blood glucose; (i): Fasting insulin; (j): HOMA-IR. One-way ANOVA, Different lowercase letters represent significant differences, P < 0·05; different capital letters represent extremely significant differences, P < 0·01, *Student’s t test, P < 0·05, ***Student’s t-test, P < 0·01, ns, no significant, n 5–8/group.

FLE improves lipid metabolism in offspring
Male offspring mice were executed at L 21·5 d. Analysis of their body composition revealed that FLE with no significant effects on offspring BMI, body fat percentage, brown fat, white fat, scapular fat, groin fat, mesenteric fat, perirenal fat, subcutaneous fat, and liver content (Table 5). The detection of serum lipid indexes in offspring mice revealed that FLE had no significant effect on TCHO, TG, HDL-C and LDL-C (online Supplementary Figure S2C). The effect of FLE on the intestinal health of offspring was further evaluated. It was observed that 5 % FLE had no significant effect on the jejunal crypt depth, villus height, villus height/crypt depth, villus width, epithelial thickness and intestinal wall thickness in the littermates (online Supplementary Figure S3C).
Effects of FLE on lipid deposition of offspring

FLE activates PI3K/AKT signaling pathway in offspring and improves liver lipid metabolism
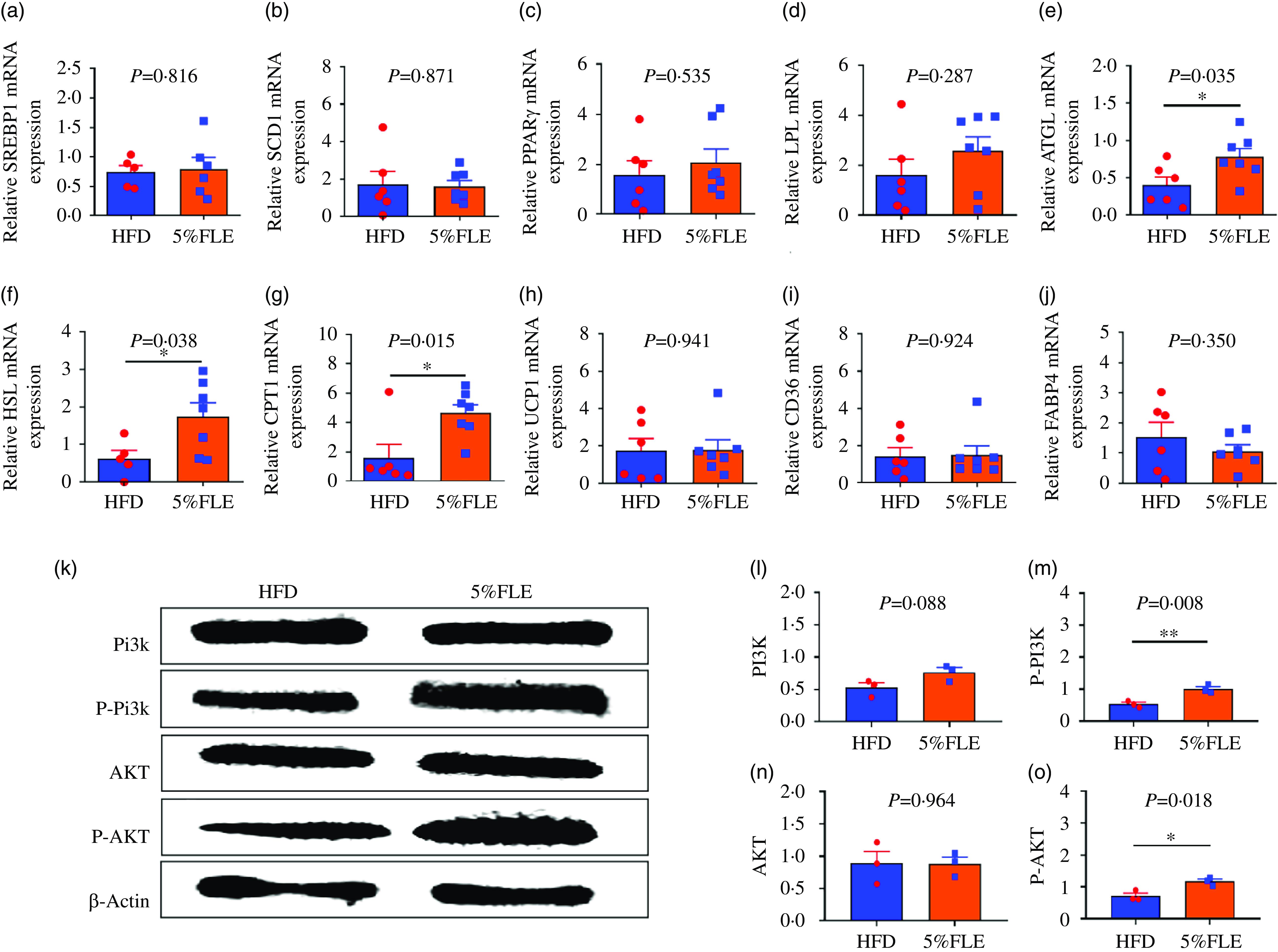
Evaluation of genes related to lipid metabolism in the offspring liver revealed that 5 % FLE had no significant effect on lipid synthesis genes SREBP1c, SCD1 and PPARγ (Figure 6(a)–(c)) and lipolysis genes LPL, but upregulated ATGL, HSL and fatty acid oxidation gene CPT1 mRNA (P < 0·05) (Figure 6(d)–(g)). Furthermore, no significant effect on UCP1 and fatty acid transport genes CD36 and FABP4 was observed (Figure 6(h)–(j)). PI3K/AKT signaling pathway protein expression was examined, and it was observed that total PI3K and AKT protein levels did not significantly change and PI3K and AKT protein phosphorylation levels were significantly increased (P < 0·05) in the livers of 5 % FLE offspring (Figure 6(k)–(m)).
Effect of FLE on lipid metabolism and PI3K/AKT signaling pathway in the liver of offspring (a)–(c): The mRNA expression of lipid synthesis related genes; (d)–(f): The mRNA expression of lipolysis genes; (g): The mRNA expression of fatty acid oxidase; (h): The mRNA expression of Uncoupling protein 1; (i)–(j): The mRNA expression of fatty acid transport genes, n 6–7/group; (k)–(o): PI3K, P-PI3K, AKT and P-AKT protein expression in the liver of offspring, n 3/group, Student’ s t-test, *P < 0·05, **P < 0·01, HFD v. FLE.

Discussion
Gestational obesity or excessive energy intake during pregnancy is the leading cause of the occurrence of gestational diabetes(Reference Marshall, Abrams and Barbour28). Excessive fat accumulation in the abdomen and liver can cause chronic systemic inflammation in the body, which leads to insulin resistance in peripheral tissues, thus, increasing the risk of diabetes(Reference Dinda, Dinda and Roy29,Reference Pulgaron and Delamater30) . Most of the current medications used to treat diabetes have side effects, and diet regulation is considered a safety measure to reduce the risk of occurrence of gestational diabetes(Reference Reynolds, Akerman and Mann31,Reference Boden, Sargrad and Homko32) . Asians have a long history of consuming mushrooms, the most common of which is Lentinus edodes, widely farmed worldwide(Reference Bisen, Baghel and Sanodiya33). Current research depicts that moderate consumption of mushrooms has no negative impact on animal health(Reference Mhd Omar, Abdullah and Abdullah34,Reference Abdullah, Lau and Ismail35) . Lentinus edodes has been reported to have various bioactive substances with antioxidant, anti-inflammatory and anti-diabetic properties(Reference Bisen, Baghel and Sanodiya33). It has been demonstrated that the addition of Lentinus edodes to the diet can reduce high blood lipids and blood glucose caused by HFD in mice and reduce liver lipid deposition(Reference Yang, Hwang and Kim36). Supplemental intake of Lentinus edodes in a rat model of streptozotocin-induced diabetes during pregnancy reduces serum glutamic-pyruvic transaminase and glutamic oxaloacetic transaminase and protects the liver from streptozotocin-induced damage(Reference Laurino, Viroel and Caetano37). This study observed that the addition of FLE to the gestational diet significantly reduced body fat deposition and fasting serum insulin levels and improved systemic insulin sensitivity in pregnant and lactating mice.
The liver is the center of the body’s energy metabolism and one of the most important insulin target organs. The liver regulates the body’s lipid metabolism by regulating the intake, synthesis, catabolism and transport of fatty acids in response to physiological fluctuations of nutrients. Studies have reported that inhibiting hepatic lipid uptake, synthesis and transport and promoting hepatic lipolysis are effective measures to alleviate fatty liver and obesity-related metabolic diseases(Reference Ipsen, Lykkesfeldt and Tveden-Nyborg38,Reference Badmus, Hillhouse and Anderson39) . Hepatic lipid metabolism is regulated by acetyl coenzyme A carboxylase (ACC), fatty acid synthase (FAS) and SCD1 and co-regulated by CPT1 and SREBP1c transcription factors(Reference Ravaut, Légiot and Bergeron40,Reference Fang, Pan and Qu41) . A study involving rats has observed that inhibition of protein and mRNA expression of fatty acid production genes ACC, SREBP1c and FAS and promotion of mRNA expression of fatty acid oxidation gene CPT1 inhibited the synthesis of cholesterol and fatty acids in the liver and promoted fatty acid oxidation, thus, alleviating fatty liver in mice(Reference Lin, Zeng and Liu42). Another study has reported that inhibition of FATP4 reduced free fatty acid content, cholesterol content and triglyceride deposition in the liver of mice(Reference Göcebe, Jansakun and Zhang43). This study observed that FLE inhibited the expression of the liver fatty acid transport gene FATP4 mRNA in pregnant mice, suggesting that FLE can reduce ectopic deposition of lipids in the liver by inhibiting hepatic lipid transport. In addition, FLE increased the mRNA expression of liver lipolysis and fatty acid oxidation genes (ATGL, HSL and CPT1) in mice, suggesting that it can also reduce liver lipid deposition by promoting lipolysis. This is similar to the findings of previous studies in which supplementation of 60 g/kg of Lentinus edodes powder or 5 % Lentinus edodes in a HFD prevented weight gain and total fat deposition and reduced plasma and liver triglyceride levels in rats(Reference Yoon, Alam and Lee44,Reference Handayani, Chen and Meyer45) . However, the substances in FLE or Lentinus edodes that play a role in reducing lipid deposition have not been discovered. A study has reported that the addition of mushroom-derived β glucan and ergosterol to HFD in mice reduced serum lipid levels(Reference Kanagasabapathy, Malek and Mahmood46,Reference Morales, Tejedor-Calvo and Jurado-Chivato47) . Similarly, Gong et al. demonstrated that administration of Lentinus edodes polysaccharides for 7 weeks to mice on a high-sugar, HFD and streptozotocin-induced diabetic reduced serum triglyceride and cholesterol levels(Reference Gong, Wang and Liu18). Reducing plasma cholesterol levels with mushroom-derived beta glucan may be associated with the inhibition of intestinal absorption of bile salts, fat and cholesterol and stimulation of hepatic resynthesis of bile salts to increase cholesterol utilisation(Reference Cerletti, Esposito and Iacoviello48,Reference Sima, Vannucci and Vetvicka49) . In addition, intestinal flora ferments β-glucan in the intestine to produce short-chain fatty acids, which inhibit cholesterol synthesis by inhibiting reductase, and inhibit cholesterol levels by increasing LDL cholesterol catabolism(Reference Cerletti, Esposito and Iacoviello48,Reference Rondanelli, Opizzi and Monteferrario50) . However, in this study, although FLE significantly reduced body fat deposition in mice during pregnancy and lactation, there was no significant reduction in maternal and offspring lipid levels, which may be associated with the short duration of the experimental treatment. The total duration of treatment during gestation and lactation was 6 weeks in this study, and previous studies have reported that feeding HFD during gestation and lactation alone did not cause very significant obesity and elevated blood lipids(Reference Cardenas-Perez, Fuentes-Mera and de la Garza51). In addition, it has been reported that the maternal body synthesises and absorbs large amounts of cholesterol and transports it to the fetus through the placenta to meet the foetal cholesterol requirements during the later stages of gestation(Reference Yañez and Leiva52), and maintaining a relatively high level of cholesterol is more conducive to its growth and development(Reference Wang, Chen and Zhang53,Reference Mauri, Calmarza and Ibarretxe54) , which might explain why FLE increases foetal cholesterol levels and promotes offspring growth.
Insulin is an important hormone, which regulates postprandial glucose and lipid metabolism, facilitating the transfer of glucose to the liver, muscle and adipose tissue and inhibiting gluconeogenesis in the liver to maintain metabolic homeostasis(Reference Lennicke and Cochemé55). Previous studies have indicated that enhancing maternal insulin sensitivity can optimize nutrient metabolism and transport, thereby promoting fetal growth. In this study, supplementation with FLE during pregnancy and lactation significantly improved maternal insulin sensitivity, lowered fasting insulin levels and the HOMA-IR index, and this contributed to an increase in body weight post-weaning. Among them, the hepatic PI3K/AKT signaling pathway is important in glucose uptake(Reference Huang, Liu and Guo56). Insulin interacts with insulin receptors, increasing PI3K/AKT phosphorylation. By activating downstream signaling molecules, the phosphorylated AKT mediates glucose transport into cells, inhibits hepatic glycogen synthesis, increases glucose uptake and lipid metabolism and hence maintains basal glucose homeostasis(Reference Huang, Liu and Guo56,Reference Li, Tang and Shi57) . In this study, FLE significantly increased the phosphorylation levels of proteins of PI3K and AKT in the liver of pregnant mice and offspring, suggesting that FLE may be an important molecular mechanism for improving maternal and offspring insulin sensitivity. Among these, Lentinus edodes polysaccharide and ergosterin in FLE may be the key active ingredients in activating PI3K/AKT signaling pathway. It has been demonstrated that dietary polysaccharides can improve insulin sensitivity by activating the Nrf2/HO-1 or PI3K/AKT pathways(Reference Gong, Wang and Liu18,Reference Liao, Zhang and Liu58) , and ergosterol can also improve insulin resistance and reduce blood lipid levels by activating PI3K/AKT signaling(Reference Xiong, Huang and Liu59), but this study did not further evaluate the active ingredients in FLE for maternal and offspring insulin sensitivity and mechanism of action.
Previous studies have confirmed that administration of a HFD during pregnancy and lactation can lead to certain abnormalities in maternal insulin sensitivity. Notably, in most animal models of insulin metabolic disorders, HFD treatment usually lasts for over 8 weeks. Mice subjected to longer-term HFD treatment generally exhibit a more pronounced insulin-resistant phenotype. Therefore, in future studies on insulin metabolism-related diseases during pregnancy and lactation, it is feasible to use HFD for pretreatment and prolong the HFD treatment period. This approach will enable the HFD-induced pregnancy metabolic disorder model to better reproduce clinical pathological features and enhance the observability of experimental interventions. Shiitake mushroom fermentation products are rich in polysaccharides. In animals, the utilization of polysaccharides is typically mediated by gut microbiota. In this study, we found that shiitake mushroom fermentation products can improve maternal lipid metabolism and insulin sensitivity in both mothers and fetuses by activating the hepatic PI3K/AKT signaling pathway. However, it remains unclear whether shiitake mushroom fermentation products exert their effects directly or indirectly by influencing the structure or metabolites of gut microbiota. Future studies can employ metagenomics, 16s RNA, and metabolomics technologies to further explore the mechanisms underlying the interactions between shiitake mushroom fermentation products, microbiota, and the host. This research is of great significance for the future treatment and application of shiitake mushroom fermentation products and similar polysaccharides in gestational obesity and gestational diabetes.
Conclusion
In summary, FLE plays an important role in regulating maternal and offspring lipid metabolism, reducing maternal fat deposition during pregnancy and lactation by promoting lipolysis and activating the hepatic PI3K/AKT signaling pathway to improve maternal and offspring insulin sensitivity during pregnancy.
Supplementary material
For supplementary material/s referred to in this article, please visit https://doi.org/10.1017/S0007114525103796
Acknowledgements
Not applicable.
This work was supported by the National Natural Science Foundation of China (Grant No.32202755), the Guangdong Basic and Applied Basic Research Foundation (2021A1515012417), the Guangzhou Science and Technology Project (202201010528, 2025A04J5335), the Special Projects in Key Areas of Ordinary Universities in Guangdong Province (2022ZDZX4021), the YueHao Biotechnology (Guangzhou) Co, Ltd., the Guangdong Provincial College Student Innovation and Entrepreneurship Training Program Project(S202311347012).
J. P., R. L.: conceptualization, investigation. Y. H.: Funding acquisition. M. Y., S. Z., Y. Z. and W. H.: Data curation. Y. T., H. W. and G. Z.: resources. W. W. and J. W.: project administration, supervision. Z. L. and W. Z.: writing-original draft. Z. L.: writing-review & editing. All authors reviewed and endorsed the final manuscript.
The authors declare that they have no financial/commercial conflicts of interest.
The experimental procedure was approved by the Animal Ethics Committee of ZhongKai College of Agricultural Engineering, with the ethical approval number is ZHKU-2020–0905.
The experimental data are included in the manuscript and is available from the corresponding author upon request.
Prior to the commencement of this study, a detailed protocol was prepared, but it was not registered on any public platform. All experimental details and analysis plans were documented and archived internally.