


Introduction
Common ragweed (Ambrosia artemisiifolia L.) is a ubiquitous, annual, herbaceous weed that can cause devastating impacts to crop production when left unmanaged (Chen et al. Reference Chen, Marusciac, Tamas, Valenta and Panaitescu2018; Leskovšek et al. Reference Leskovšek, Datta, Knežević and Simončič2012). Ambrosia artemisiifolia emerges earlier than many other summer annual weeds (Leskovšek et al. Reference Leskovšek, Datta, Knežević and Simončič2012), conferring an emergence advantage over its competitors. Its aggressive growth can reduce crop yield by 38% in corn (Zea mays L.) and 95% in soybean [Glycine max (L.) Merr.] (Barnes et al. Reference Barnes, Jhala, Knezevic, Sikkema and Lindquist2018). Ambrosia artemisiifolia is a prolific seed producer, a single plant can produce up to 62,000 seeds, and seeds remain viable for 39 years (Baskin and Baskin Reference Baskin and Baskin1980; Dickerson and Sweet Reference Dickerson and Sweet1971). Ambrosia artemisiifolia’s propensity to evolve resistance to commonly used herbicides in corn and soybean crop rotations further exacerbates control issues. Populations of A. artemisiifolia have been documented to have evolved resistance to five herbicide sites of action (SOA, WSSA group number) including: photosystem II (PSII, 5), acetolactate synthase (ALS, 2), protoporphyrinogen oxidase (PPO, 14), 5-enolpyruvoylshikimate 3-phosphate synthase (EPSPS, 9), and synthetic auxin inhibitors (4) (Heap Reference Heap2025). Overreliance on ALS, EPSPS, and PPO chemistries in row crops, particularly in soybean, has caused widespread resistance throughout the United States, with numerous populations developing multiple resistance. Interestingly, synthetic auxins have been used for weed control longer than the other SOAs but have far fewer documented cases of resistance (Busi et al. Reference Busi, Goggin, Heap, Horak, Jugulam, Masters, Napier, Riar, Satchivi and Torra2018; Peterson Reference Peterson1967). There has only been one reported case of synthetic auxin resistance in A. artemisiifolia (Gallina et al. Reference Gallina, Cregg, Patterson, Hill and Saha2023).
Clopyralid is a systemic synthetic auxin commonly used for postemergence weed control in agriculture and horticulture cropping systems. It is useful for its effectiveness on broadleaf species and selectivity against certain dicots (DiTomaso et al. Reference DiTomaso, Kyser, Miller, Garcia, Smith, Nader, Connor and Orloff2006). However, its broadleaf spectrum is unique from that of other synthetic auxins, being especially effective in controlling hard to manage weed species in the Asteraceae family but having less activity on critical weed species in the Chenopodiaceae (e.g., common lambsquarters [Chenopodium album L.]) and Amaranthaceae (e.g., waterhemp [Amaranthus tuberculatus (Moq.) Sauer]) families. This narrow spectrum indicates that clopyralid may have an SOA unique from other Group 4 synthetic auxin herbicides such as 2,4-D and dicamba. Since the registration of clopyralid in 1987, resistance to this herbicide has been rare, reported in only five species globally, including: C. album, A. artemisiifolia, lawn burweed (Soliva sessilis Ruiz & Pav.), spotted knapweed (Centaurea stoebe L.), and yellow starthistle (Centaurea solstitialis L.) (Gallina et al. Reference Gallina, Cregg, Patterson, Hill and Saha2023; Ghanizadeh and Harrington Reference Ghanizadeh and Harrington2017; Harrington et al. Reference Harrington, Ward and Wells2001; Sabba et al. Reference Sabba, Ray, Lownds and Sterling2003).
The inheritance pattern of herbicide-resistance mechanisms strongly influences the rate at which resistance evolves and spreads within weed populations (Hulme and Liu Reference Hulme and Liu2022). Dominant resistance alleles tend to increase in frequency more rapidly than recessive alleles, because both homozygous and heterozygous individuals express the resistant phenotype; however, they do not go to fixation nearly as quickly (Kreiner et al. Reference Kreiner, Stinchcombe and Wright2018). To date, the only inheritance study of clopyralid resistance has been conducted in C. solstitialis, where resistance was found to be controlled by a single nuclear recessive gene (Sabba et al. Reference Sabba, Ray, Lownds and Sterling2003).
Herbicide-resistance inheritance can also depend on the complexity of the resistance mechanism. Mechanisms that confer herbicide resistance are broadly categorized into two main categories, target-site resistance (TSR) and non–target site resistance (NTSR) (Heap Reference Heap2014). TSR is conferred by mutations directly affecting the herbicide’s target protein, including mutations that change the target protein amino acid sequence or those that cause the gene to be overexpressed, such as copy number variation. NTSR is conferred by a variety of mechanisms that prevent the herbicide from reaching the target site or reducing herbicide toxicity once it reaches the target site. TSR mechanisms that confer resistance to synthetic auxin herbicides include mutations in: transcriptional repressors (Aux/IAAs), auxin response factors (ARFs), and the Skp1-Cullin-F-box TIR1/AFB E3 ubiquitin ligase complex (SCFTIR1/AFB) (Todd et al. Reference Todd, Figueiredo, Morran, Soni, Preston, Kubeš, Napier and Gaines2020). Both TSR and NTSR mechanisms have been shown to confer resistance to synthetic auxin herbicides. For example, populations of kochia [Bassia scoparia (L.) A.J. Scott] have been found to have cross-resistance to dicamba, 2,4-D, and fluoxypyr caused by a mutation in an auxin gene, KsIAA16 (LeClere et al. Reference LeClere, Wu, Westra and Sammons2018). In many cases, the IAA protein subdomain known as the “degron” is particularly important for synthetic auxin binding (Todd et al. Reference Todd, Figueiredo, Morran, Soni, Preston, Kubeš, Napier and Gaines2020). Examples of NTSR mechanisms for synthetic auxin resistance include common poppy (Papaver rhoeas L.), which has evolved cross-resistance to imazamox and 2,4-D through cytochrome P450–induced metabolism (Torra et al. Reference Torra, Rojano-Delgado, Menéndez, Salas and de Prado2021). These findings underscore the need for further research to uncover the resistance mechanism endowing clopyralid resistance.
Ultimately, understanding herbicide-resistance mechanisms is critical for effective weed management, yet several gaps remain in the case of clopyralid and synthetic auxins in general. The specific genetic mutation(s) that confer resistance to various auxin herbicides such as clopyralid have not been fully elucidated, especially in species with complex resistance mechanisms. Likewise, we are still learning the exact binding site(s) for all the auxinic herbicides. The gaps in understanding auxinic herbicide resistance pathways can lead to ineffective control measures to prevent further resistance. Our study is guided by the hypothesis that a single-nucleotide polymorphism (SNP) in the degron domain of an AUX/IAA gene could confer clopyralid resistance, as this seems to be the most frequent mechanism for strong resistance to other synthetic auxins. With this in mind, the first objective of this research was to understand the mechanism(s) of clopyralid resistance. The second objective was to determine how the mechanism(s) of resistance are inherited. For Objective 1, we employed RNA-seq methods to simultaneously measure gene expression and to sequence the entire transcriptome. Whereas for Objective 2, we performed standard mendelian test crosses, phenotyping both the F1 and F2 generations. This investigation has broader implications in the field of resistance to herbicides, particularly auxinic herbicides, aiding in the development of strategies to prevent future instances of resistance, thereby contributing to the sustainable management of these hard to control weed populations.
Materials and Methods
Plant Material and Growth Conditions
In 2016, a field herbicide trial was performed by Bernard Zandstra in an 8-yr-old planting of Fraser fir [Abies fraseri (Pursh) Poir.] Christmas trees in Montcalm County in central lower Michigan. The grower had applied clopyralid (Stinger®, Corteva Agriscience, Indianapolis, IN 46268, USA) during each year of the planting to control A. artemisiifolia. In this field trial, A. artemisiifolia survived twice the field use rate (280 g ha−1) of clopyralid (data not shown). Mature plants that survived clopyralid application were collected from the 2016 field trial (AMBEL-40) and from a susceptible biotype (AMBEL-39) collected in a nearby field that did not receive clopyralid application. Seeds were hand threshed from the plant material and cleaned using an air-column separator. Cleaned seeds were placed in mesh bags (Outdoor Wilderness Fabrics, Caldwell, ID 83605, USA) and vernalized outside for a minimum of 6 wk during the winter months, under freeze–thaw conditions. After 6 wk, seeds were tested for germination, and the vernalization treatment was extended if necessary. After dormancy was broken, seeds were placed in a cold storage room (∼4 C) until the start of the experiment. Seeds were germinated in 25.4 by 25.4 by 7.6 cm flats and then transplanted once they had two completely unfurled leaves into a 10 by 10 by 12 cm pot with Suremix greenhouse potting media of peat moss with perlite (Michigan Grower Products, Inc™, Galesburg, MI 49053, USA). The greenhouse was maintained at 25 ± 5 C with a midday light intensity of 1,000 µmol m−2 s−1 photosynthetic photon flux and a 16-h photoperiod. Plants were watered and fertilized with Peters Excel pH Low 15-7-25 Hi Mag Hi K with black iron (ICL Fertilizers, St. Louis, MO 63141, USA) as needed to promote optimum plant growth.
Establishing F1 and F2 Populations
Two generations of recurrent group selection (approximately 50 plants per generation) with clopyralid were performed to remove any susceptible individuals from AMBEL-40. Homozygous clopyralid-resistant and clopyralid-susceptible plants from AMBEL-40 and AMBEL-39 were used as parents in crosses to generate F1 populations. Homozygosity of the parental plants was confirmed in AMBEL-40 by no injury from treatment with clopyralid applied at the field use rate (140 g ai ha−1) and susceptibility of AMBEL-39 by 100% injury from the same treatment. Clopyralid was applied when plants were 10 cm in height. Herbicide applications were made with the Generation 4 DeVries research track sprayer (DeVries Manufacturing, Inc™, Hollandale, MN 56045, USA), outfitted with an 8001EVS spray nozzle (TeeJet® Technologies, Wheaton, Glendale Heights, IL 60139, IL) calibrated to deliver 187 L ha−1 at 207 kPa. Three separate crosses were performed, AMBEL-40 served as the pollen donor and AMBEL-39 as the female parent; resulting seed was collected from AMBEL-39 plants. AMBEL-39 plants were emasculated daily to prevent pollen production, even though A. artemisiifolia has been shown to be an obligate cross-pollinated species (Friedman and Barrett Reference Friedman and Barrett2008). To ensure adequate seed production, one AMBEL-40 served as the pollen source for two AMBEL-39 plants. The three plants were kept in an enclosure made with mono-filament mesh with an aperture of 110 microns to prevent contamination from outside pollen sources; further enclosures were kept in separate greenhouse bays. Seeds collected from AMBEL-39 plants were kept separate to create each F1 family.
To confirm our crosses were successful, we identified an unrelated random polymorphic locus with three SNPs that were homozygous and polymorphic in AMBEL-40 and AMBEL-39 parental lines. Young leaf tissue from 10 plants from each F1 family and parental line was collected from the apical growing point of three completely unfurled leaves from each plant, and DNA was extracted using a DNeasy Plant Kit (Qiagen, Germany). We used PCR to amplify the sequence surrounding the SNP and sequenced the amplicon using Sanger sequencing performed by the Michigan State University Genomics Core (East Lansing, MI). For PCR reactions, the forward primer was: 5′ CAC GGG ATT ACA GAT TCA GGG 3′; and the reverse primer was: 5′ TGG CTT GTT CAG AAC AGG C3′, with a predicted amplicon size of 286 bp. PCR amplifications were performed in a total reaction volume of 50 μl, containing 1 × PCR buffer (10 mM Tris-HCl, pH 8.3; 50 mM KCl), 1.5 mM MgCl2, 200 μM of each dNTP, 0.5 μM of each primer, 1.25 units of Taq DNA polymerase (Thermo Fisher Scientific, Waltham, MA 02451, USA), and 100 ng of genomic DNA template. The thermal cycling conditions were as follows: an initial denaturation at 95 C for 2 min; followed by 35 cycles of denaturation at 95 C for 30 s, annealing at 55 C for 30 s, and extension at 72 C for 1 min; with a final extension at 72 C for 5 min. PCR products were analyzed by electrophoresis on a 10 g L-1 (w/v) agarose gel in 1 × TAE buffer, stained with GelRed Nucleic Acid Gel Stain (MilliporeSigma, Burlington, MA 01803, USA), and visualized under UV light. Crosses were confirmed to be successful by manually filtering the Sanger sequencing output and looking for heterozygosity in all three SNPs in this polymorphic locus.
Subsequently, two plants from each of the three F1 families were paired to create F1 sibling crosses to produce three segregating F2 populations. Each pair of F1 siblings were placed in the same pollination enclosure as previously described to prevent unwanted pollination events. Seeds were collected from both individuals in the cross and pooled from each F1 sibling to create each F2 family. Each F2 family was named using the M#F# convention, with M corresponding to the male, F corresponding to the female, and # corresponding to the individual. Ultimately the three crosses that were most successful and produced sufficient viable seed were the M3F1, M3F2, and M1F1 populations.
Herbicide Screening and Analysis
F1 families were subjected to a full dose–response assay to identify the rate causing 50% biomass reduction (ED50) and determine the dominance of the resistance trait. The assay consisted of 64 plants from each of the three F1 families grown beside the parental lines, AMBEL-40 and AMBEL-39, under the same greenhouse conditions described earlier. F1 families, AMBEL-40, and AMBEL-39 were treated with eight clopyralid doses including: 0, 35, 70, 140 (field use rate), 280, 560, 1,120, and 2,240 g ai ha−1, and each treatment was replicated eight times, conducted twice. Plant injury ratings on a scale of 0% to 100%, with 0% indicating no injury and 100 indicating complete death, were taken every 7 d. Aboveground biomass was collected 21 d after application and dried for 3 d at 66 C. F1 families, AMBEL-40, and AMBEL-39 dry weights were used to generate dose–response curves using the drc package in R (Ritz et al. Reference Ritz, Baty, Streibig and Gerhard2015). Three-parameter (Equation 1) and four-parameter (Equation 2) models were tested using the drc modelFit function, and the best model the one having the smallest residual error and highest P-value from a lack-of-fit test.
 $$Y = {{d}\over{{1 + {\rm{exp}}\left[ {b\left( {{\rm{log}}x - e} \right)} \right]}}}$$
$$Y = {{d}\over{{1 + {\rm{exp}}\left[ {b\left( {{\rm{log}}x - e} \right)} \right]}}}$$
Equation 1 is the three-parameter log-logistic model where Y is the response variable, x is the independent variable, d is the upper limit, b is the relative slope around e, and e is the inflection point.
 $$Y = {{{d - c}}\over{{1 + {\rm{exp}}\left[ {b\left( {{\rm{log}}\left( x \right) - {\rm{log}}\left( e \right)} \right)} \right]}}}$$
$$Y = {{{d - c}}\over{{1 + {\rm{exp}}\left[ {b\left( {{\rm{log}}\left( x \right) - {\rm{log}}\left( e \right)} \right)} \right]}}}$$
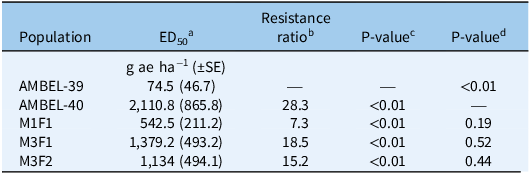
Equation 2 is the four-parameter log-logistic model where Y is the response variable and x is the independent variable. The parameter d represents the maximum response, c represents the minimum response, b describes the relative slope around the inflection point, and e is the inflection point at which the response is halfway between c and d. The resistance ratio (resistant/susceptible [R/S]) of ED50 values for AMBEL-40, AMBEL-39, and F1 families was calculated using numbers generated from the dose–response curves.
Additional dose–response studies were conducted with just the parental AMBEL-40 and AMBEL-39 lines with fluoxypyr (Starane® Ultra, Corteva Agriscience, Indianapolis, IN 46268, USA), triclopyr (Garlon®, Corteva Agriscience, Indianapolis, IN 46268, USA), dicamba (XtendiMax®, Bayer CropScience, Research Triangle Park, NC 27709, USA), and 2,4-D amine (Weedar® 64, Nufarm Agricultural Products, Alsip, IL 60803, USA) to explore whether AMBEL-40 was multiple or cross-resistant to other Group 4 auxinic herbicides. Within each herbicide, four dose treatments were used: 0, 0.5, 1, and 2 times the field recommended labeled rates of 0.16, 1.12, 0.56, and 1.17 kg ae ha−1, for fluoxypyr, triclopyr, dicamba, and 2,4-D amine respectively. There were five replications, and the experiment was conducted twice. At 28 DAT, replications were visually rated and harvested for dry biomass. Cross-resistance dose–response data were analyzed using the drc package in R (Knezevic et al. Reference Knezevic, Streibig and Ritz2007), and three-parameter log-logistic models (Equation 1) were fit to the data as selected by the drc modelFit function, and the best model was the one having the smallest residual error and highest P-value from a lack-of-fit test.
F2 families were subjected to a single-dose screen to test segregation of the clopyralid-resistance trait. F2 families and parental AMBEL-40 and AMBEL-39 were grown under the same greenhouse conditions outlined earlier. One hundred plants from each F2 family and parental AMBEL-40 and AMBEL-39 were treated with a discriminating dose of 280 g ha−1 clopyralid, which is lethal to all AMBEL-39 plants by 21 d after application, as determined by dose–response assays performed earlier. Plant injury ratings were taken every 7 d, and aboveground biomass was collected at 21 d after application. Survival ratings were collected and ranged between zero and one at 21 d after application. Chi-square tests were performed on visual survival ratings of F2 plants to determine the inheritance pattern of the resistance mechanism. Chi-square analysis was performed in R using the chi-square test function.
RNA-seq for Resistance Mechanism Prediction
An RNA-seq experiment was designed to investigate possible resistance mechanisms. Plant material for this experiment was collected from four AMBEL-40 and four AMBEL-39 plants grown from the same seed lot as the parental plants for the herbicide-screening experiments. Plants were grown under the same greenhouse conditions outlined earlier. Once plants reached 30 cm, leaf tissues from the three youngest, completely unfurled leaves were harvested from untreated plants, flash frozen in liquid nitrogen, and stored at −80 C for subsequent RNA extraction. RNA was extracted from leaf tissue using the RNeasy plant kit following the manufacturer’s protocol (Qiagen, Germany) with final RNA eluted in 50 μl of ddH2O. RNA concentration, purity, and quality were measured using a NanoDrop One spectrophotometer (Thermo Fisher, USA) and a Qubit 1.0 fluorometer (Invitrogen, Carlsbad, CA 92008, USA). RNA integrity was checked with an Agilent TapeStation 4150 using RNA screentape assay to validate that the RNA integrity number was >7.0. RNA was sequenced using Illumina NovaSeq6000, 150-bp, paired-end sequencing by Novogene. Raw FASTQ files were trimmed and filtered using fastp and default parameters (Chen et al. Reference Chen, Marusciac, Tamas, Valenta and Panaitescu2018). The reads were aligned to a transcriptome that was generated from the annotated reference genome (NCBI accession no. PRJNA929658) using HISAT2 with standard alignment parameters for paired-end Illumina alignment (Kim et al. Reference Kim, Paggi, Park, Bennett and Salzberg2019). Alignment files were subsequently used for differential expression analysis and potential target-site SNP investigations.
Samtools idxstats was used to generate read counts for each gene and to generate a read count table (Danecek et al. Reference Danecek, Bonfield, Liddle, Marshall, Ohan, Pollard, Whitwham, Keane, McCarthy and Davies2021). The libraries were normalized in edgeR using the trimmed mean of M-values method (Robinson et al. Reference Robinson, McCarthy and Smyth2010). Differences in gene expression between AMBEL-40 and AMBEL-39 were identified using pairwise comparison in edgeR following the recommendations from the edgeR handbook; specific code is available at https://github.com/PattersonWeedLab. To narrow our focus, transcripts with a P-value ≤ 0.01 and a log fold change ≥4 were selected for more detailed consideration.
Furthermore, we were specifically interested in sequence motifs known as “degrons,” which have previously been shown to be involved in TSR to other auxinic herbicides, including 2,4-D and dicamba, in other dicot species (LeClere et al. Reference LeClere, Wu, Westra and Sammons2018; Montgomery et al. Reference Montgomery, Soni, Marques Hill, Morran, Patterson, Edwards, Ratnayake, Hung, Pandesha and Slotkin2025; Todd et al. Reference Todd, Figueiredo, Morran, Soni, Preston, Kubeš, Napier and Gaines2020). We extracted all transcripts in the reference transcriptome that encoded an amino acid sequence, ‘GWPPV’, as this sequence has been shown to be essential for Aux/IAA to recognize auxin. We visually screened read alignments using the integrated genome viewer for polymorphisms (SNPs, InDels, etc.) that distinguished resistant from susceptible individuals to look for target-site polymorphisms that might explain the resistance trait.
Results and Discussion
Clopyralid Resistance in Ambrosia artemisiifolia
Dose–response assays were conducted to compare AMBEL-40 and AMBEL-39 responses to clopyralid (Figures 1 and 2). The ED50 values for AMBEL-40 and AMBEL-39 were 2,110.8 and 74.5 g ha−1, respectively, with a corresponding R/S ratio of 28.3 (Table 1). The standard field rate is 140 g ha−1, therefore AMBEL-40 has a resistance level 15 times greater than field rate, while AMBEL-39 is controlled at 0.5 times the field rate of clopyralid. When treated with clopyralid at rates below 1,120 g ha−1, AMBEL-40 remained unaffected, showing no symptoms of herbicide injury; while AMBEL-39 displayed symptoms including twisted and curled leaves, epinastic growth, and significant tissue damage, at clopyralid application rates as low as 35 g ha−1 (Figure 1). High levels of clopyralid resistance have been documented in other weed species. For example, a population of C. stoebe evolved resistance approximately 25,600-fold greater than the susceptible reference population (Mangin and Hall Reference Mangin and Hall2016), while a resistant C. solstitialis population exhibited about 3.7-fold greater resistance (Miller et al. Reference Miller, Shinn and Thill2001), and an S. sessilis population was found to be 225-fold more resistant to clopyralid than susceptible populations (Ghanizadeh and Harrington Reference Ghanizadeh and Harrington2017). The mechanism(s) of resistance in these populations are unknown; however, the wide range in resistance levels indicates different TSR and/or NTSR mechanisms.
Dose–response assay of a single representative replicate of resistant AMBEL-40 (A) and susceptible AMBEL-39 (B) Ambrosia artemisiifolia. Pots are labeled with corresponding clopyralid acid equivalent rates (g ha−1).

Dose–response assays of Ambrosia artemisiifolia biotypes AMBEL-40 (resistant), AMBEL-39 (susceptible), and F1 crosses (M1F1, M3F1, M3F2) treated with clopyralid. Error bars represent the standard error of the mean at each dose; biomass is expressed as a percentage of untreated AMBEL-39.

Mean clopyralid dose required for 50% biomass reduction (ED50) in Ambrosia artemisiifolia biotypes: AMBEL-40 (resistant), AMBEL-39 (susceptible), and F1 crosses (M1F1, M3F1, M3F2).

a Field recommended labeled rate for clopyralid is 140 g ha−1.
b Ratio calculated by dividing in-row population ED50 by AMBEL-39 ED50.
c P-value indicates level of significance of ED50 differences between in-row population and AMBEL-39.
d P-value indicates level of significance of ED50 differences between in row population and AMBEL-40.
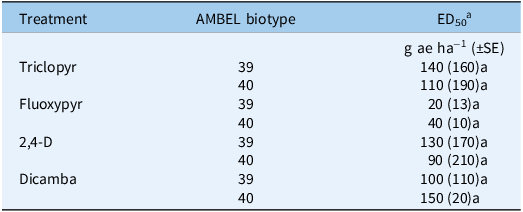
Cross-resistance dose–response results confirm that AMBEL-40 is not cross-resistant to the four other Group 4 herbicides (triclopyr, fluoxypyr, 2,4-D, and dicamba) tested in this study (Table 2). ED50 values predicted from relative biomass reduction data for triclopyr, fluoxypyr, and 2,4-D, and dicamba were not different between AMBEl-40 and AMBEL-39. Interestingly, the other reported clopyralid-resistant species: S. sessilis, C. album, and C. stoebe are cross-resistant to other Group 4 herbicides. Specifically, resistance to picloram and triclopyr in S. sessilis, aminopyralid and dicamba in C. album, and picloram in C. stoebe (Heap Reference Heap2025). The lack of cross-resistance in AMBEL-40 may be attributed to the diversity of auxin receptors in plants or specific metabolic pathways that can only act on clopyralid’s unique chemistry (Mithila et al. Reference Mithila, Hall, Johnson, Kelley and Riechers2011).
Mean herbicide doses required for 50% biomass reduction (ED50) in clopyralid- susceptible (AMBEL-39) and clopyralid-resistant (AMBEL-40) Ambrosia artemisiifolia plants.

a Mean ED50 (±SE) values within a treatment followed by the same letter are not significantly different (P ≤ 0.05). Field recommended labeled rates are 1,120, 160, 1,170, and 560 g ae ha−1, for triclopyr, fluoxypyr, 2,4-D amine, and dicamba respectively.
Dominance and Inheritance of Clopyralid Resistance in Ambrosia artemisiifolia
Dose–response assays on F1 populations were used to determine whether the trait was recessive, dominant, or incomplete dominant. These assays revealed large differences in clopyralid sensitivity between parental and F1 populations, suggesting incomplete dominance (Table 1; Figure 2). The resistant parent AMBEL-40 had the highest ED50 (2,110.8 g ha−1), while AMBEL-39, the susceptible parent, had the lowest (74.5 g ha−1). All F1 families had intermediate ED50 values between the parents. Among the three F1 families, M3F1, M3F2, and M1F1 were more resistant and significantly different compared with AMBEL-39 with ED50 values of 1,379.2, 1,134.0, and 542.5 g ha−1, corresponding to 18.5-, 15.2-, and 7.3-fold resistance, respectively.
Chi-square tests were performed on three segregating F2 families (M1F1, M3F1, and M3F2) to evaluate inheritance segregation ratios at the selection dose of 280 g ha−1. A single-gene 1:3 model did not adequately explain the segregation pattern, as M3F2 showed significant deviations from the expected ratio, and the combined analysis of all 300 plants strongly rejected this model (P < 0.05; Table 3). In contrast, none of the individual populations showed significant deviation from the two-gene 3:13 model (P = 0.8476, P = 0.6539, and P = 0.2236 for M1F1, M3F1, and M3F2, respectively), and the combined analysis was also not significant (P = 0.2835; Table 3). These results support a two-gene inheritance model for clopyralid resistance.
Chi-square analysis of inheritance segregation ratios for clopyralid resistance in F2 Ambrosia artemisiifolia a

a Observed and expected frequencies for resistant and susceptible phenotypes were compared under single-gene (1:3) and two-gene (3:13) inheritance models.
b Probability values ≥0.05 indicate that data do not differ significantly from the test ratio.
Given that the 3:13 ratio of resistant/susceptible phenotypes was a much better fit to the observed data compared with the 1:3 ratio, we concluded that the genetic pattern in these populations likely follows a more complex inheritance model. This finding supports the hypothesis that multiple genetic factors are involved in conferring high levels of resistance to clopyralid in these populations, consistent with other reports of multigenetic inheritance in Ambrosia spp. For example, glyphosate resistance in giant ragweed (Ambrosia trifida L.) involves complex genetic interactions among multiple genes (Moretti et al. Reference Moretti, Van Horn, Robertson, Segobye, Weller, Young, Johnson, Douglas Sammons, Wang and Ge2018), while ALS-inhibitor resistance in A. artemisiifolia has shown a two-gene pattern, reinforcing that herbicide resistance in A. artemisiifolia often arises through the interaction of multiple genetic factors (Patzoldt et al. Reference Patzoldt, Tranel, Alexander and Schmitzer2001). This complexity suggests that detailed genetic approaches such as quantitative trait locus (QTL) mapping, genome-wide association study (GWAS), or transcriptomic and enzyme assays will be required to identify the underlying genes and clarify the role of metabolic pathways (Rousonelos et al. Reference Rousonelos, Lee, Moreira, VanGessel and Tranel2012).
Mutation Analysis in AUX/IAA Degron Region
Some resistance cases for synthetic auxin herbicides have been shown to be caused by mutations in a multigene protein family known as AUX/IAAs, one of the proteins that bind with auxin and auxin-mimicking herbicides directly. AUX/IAAs are important transcriptional repressors of auxin responsive factors (ARFs). When AUX/IAAs bind auxins, they are degraded, releasing the ARFs from repression and increasing their expression. The short protein domain in AUX/IAAs that auxin typically binds is commonly called the “degron,” which is a specific, known amino acid sequence (Todd et al. Reference Todd, Figueiredo, Morran, Soni, Preston, Kubeš, Napier and Gaines2020). Mutations in degrons are known to confer auxinic herbicide resistance in several weed species by changing herbicide binding and thereby preventing herbicide-induced AUX/IAA degradation, ultimately allowing resistant biotypes to avoid growth abnormalities caused by these herbicides (De Figueiredo et al. Reference De Figueiredo, Küpper, Malone, Petrovic, De Figueiredo, Campagnola, Peersen, Prasad, Patterson, Reddy, Kubeš, Napier, Dayan, Preston and Gaines2022b; LeClere et al. Reference LeClere, Wu, Westra and Sammons2018).
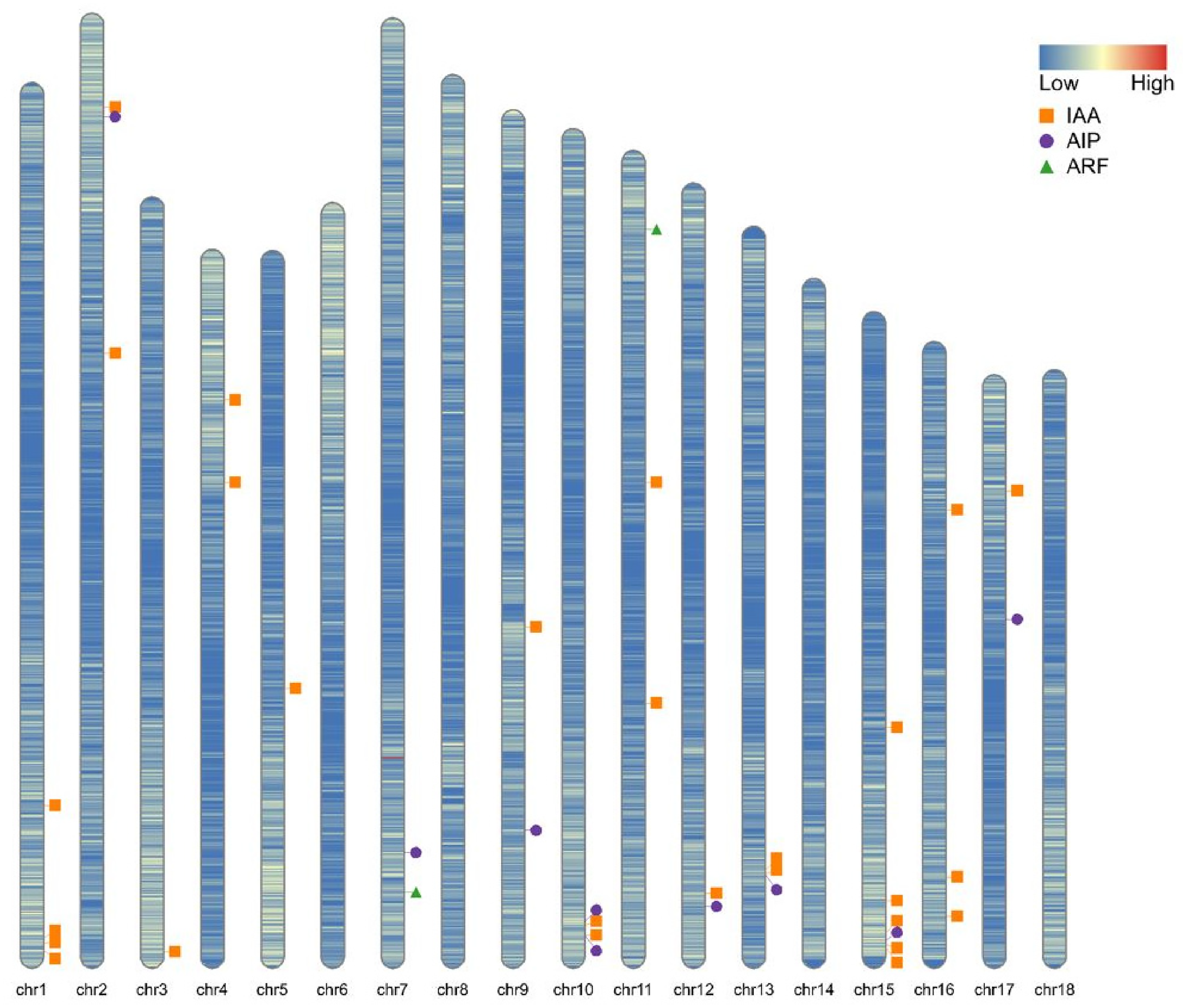
We searched for SNPs within degron regions of all annotated AUX/IAAs in the A. artemisiifolia genome to determine whether target-site mutation(s) could be the cause of clopyralid resistance. Out of the 23 unique AUX/IAA transcripts identified (Figure 3), we found three SNPs within a degron region. However, none of these SNPs led to consistent amino acid changes that clearly differentiated resistant from susceptible biotypes. Prior research has identified mutations in the degron domain as a direct cause of auxin resistance, as documented in weeds such as S. orientale and B. scoparia, where single amino acid substitutions disrupt herbicide binding and thereby confer resistance (LeClere et al. Reference LeClere, Wu, Westra and Sammons2018; Torra et al. Reference Torra, Rojano-Delgado, Menéndez, Salas and de Prado2021). Clopyralid most likely binds a different target AUX/IAA degron sequence than other auxinic herbicides such as 2,4-D and dicamba, indicated by the lack of cross-resistance in A. artemisiifolia lines as well as its distinct chemical structure as a 6-chloropicolinate compared with the phenoxycarboxylate and benzoate chemical classes as well as its unique spectrum of activity on Asteraceae and Fabaceae (and less so on Amaranthaceae and Chenopodiaceae). While we did not identify any SNPs in annotated AUX/IAA, until clopyralid’s exact target site is identified, TSR cannot be ruled out. It could also be that clopyralid’s target protein was incorrectly annotated or missing from the annotation, in which case, new and improved annotations will need to be reevaluated if and when they are generated.
Ideogram of Ambrosia artemisiifolia chromosomes displaying gene density per 100 kb as indicated by the color gradient. Positions of the auxin-related genes auxin/indole-3-acetic acids (AUX/IAAs) (orange squares), auxin-inducible proteins (AIPs) (purple circles), and auxin response factors (ARFs) (green triangles) are indicated based on genome annotations.

Transcriptomic Differentiation between Resistant and Susceptible Populations
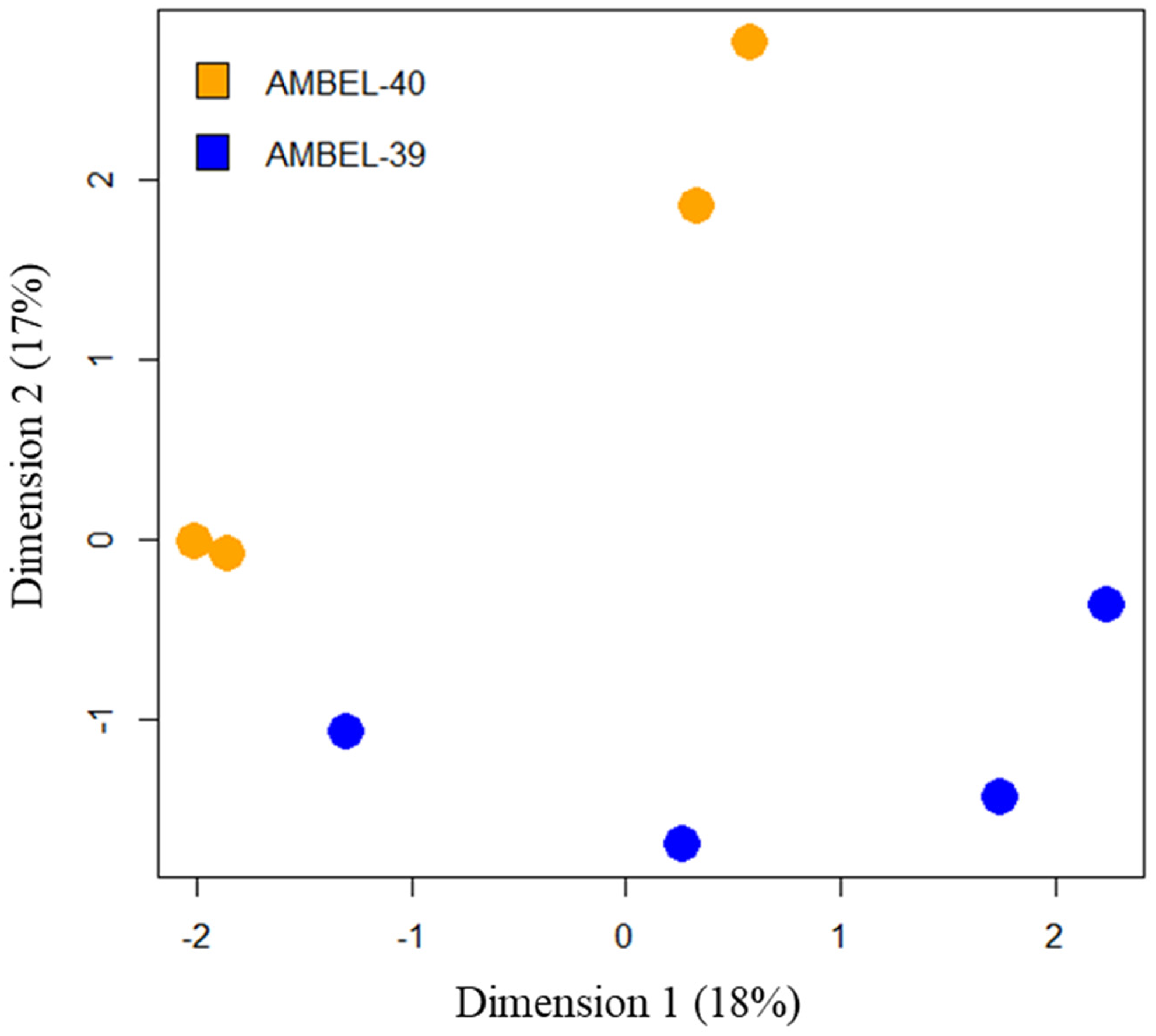
Resistance to auxinic herbicides in some cases has also been attributed to NTSR mechanisms (e.g., metabolism) (Todd et al. Reference Todd, Figueiredo, Morran, Soni, Preston, Kubeš, Napier and Gaines2020); therefore, we used our RNA-seq to perform a differential expression analysis to evaluate differentially expressed genes (DEGs) between untreated AMBEL-40 and AMBEL-39. First, we performed a multidimension scaling analysis on global gene expression data to evaluate variability between biological replicates and the two biotypes. Overall, AMBEL-40 and AMBEL-39 biotypes clearly clustered together, but there was considerable variation between biological replicates (Figure 4). This variability highlights the lack of inbreeding in these populations. Such variability in gene expression is somewhat useful for differential expression analysis, as genes with high variability between replicates will not have high levels of significance and will be removed from consideration with stringent P-value cutoffs. Only genes with consistent differential expression between the biotypes and correlation with the clopyralid-resistance phenotype will remain in the list of genes of interest. The biotypes segregate along the y axis, which accounts for only 17% of the genetic variation.
Multidimensional scaling analysis of normalized global gene expression profiles, highlighting the variability both within and between clopyralid-resistant (AMBEL-40) and clopyralid-susceptible (AMBEL-39) Ambrosia artemisiifolia populations.

The differential expression analysis identified 70 DEGs between AMBEL-40 and AMBEL-39, including 39 genes upregulated and 31 downregulated in AMBEL-40 with a fold change cutoff of 4 and a P-value cutoff of 0.01. For ease of discussion and simplicity, we will discuss these 70 DEGs in three general sections: (1) genes involved in hormone homeostasis, (2) genes involved in metabolism, and (3) genes involved in general defense response. We will also theorize about each gene’s potential involvement in clopyralid resistance.
Among the upregulated DEGs in AMBEL-40, there were several genes related to hormone signaling. The gene putative E3 ubiquitin-protein ligase LIN-1 exhibited 44.02 times increased expression in AMBEL-40 plants (Table 4). E3 ubiquitin ligases are known for their central role in modulating auxin signaling through targeted degradation of AUX/IAA proteins (Santner and Estelle Reference Santner and Estelle2010). Additionally, two critical genes in ethylene biosynthesis and regulation were overexpressed in AMBEL-40. The first, 1-aminocyclopropane-1-carboxylate oxidase (ACC oxidase), exhibiting 39.95 times increased expression in AMBEL-40, catalyzes the final step of ethylene biosynthesis, converting ACC into ethylene, a hormone involved with auxin signaling (Negi et al. Reference Negi, Ivanchenko and Muday2008). The second, ethylene-responsive transcription factor TINY, exhibiting 10.70 times increased expression in AMBEL-40, mediates stress responses triggered by hormonal signaling, particularly those involving ethylene and auxin interactions (Dubois et al. Reference Dubois, Van den Broeck and Inzé2018). Elevated TINY expression could enable AMBEL-40 to alter gene expression patterns in response to auxinic herbicide stress. Similar mechanisms involving stress-responsive transcription factors have been documented to be associated with resistance in various other auxin-resistant weeds; however, these relationships are only correlative and have not been shown to be causative (De Figueiredo et al. Reference De Figueiredo, Barnes, Boot, De Figueiredo, Nissen, Dayan and Gaines2022a; Todd et al. Reference Todd, Figueiredo, Morran, Soni, Preston, Kubeš, Napier and Gaines2020).
Summary of significantly differentially expressed genes (DEGs) of interest between clopyralid-resistant (AMBEL-40) and clopyralid-susceptible (AMBEL-39) Ambrosia artemisiifolia populations identified by RNA-sequencing analysisa

a Listed genes have a P-value < 0.01 and a differential expression greater than 4 × between the populations.
Interestingly, we observed significantly lower expression of the transcription factor RELATED TO APETALA 2 (RAP2-3) in AMBEL-40 (Table 4). RAP2-3 belongs to the APETALA2/Ethylene Responsive Factor (AP2/ERF) family, which mediates ethylene and auxin signaling pathways, especially during environmental stress (Ma et al. Reference Ma, Hu and Jiang2024). Previous studies have shown that excessive activation of ethylene signaling pathways under herbicide stress can cause severe growth inhibition, early senescence, and cell death, negatively impacting plant fitness (Grossmann Reference Grossmann2010).
Several genes involved in enhanced metabolic detoxification were also present in our significant gene of interest list. First, we saw significant overexpression of cytochrome P450 83B1 (CYP83B1) in AMBEL-40 (Table 4). Cytochrome P450 enzymes frequently mediate herbicide metabolism by catalyzing phase I oxidation reactions, which then facilitate their conjugation into large macromolecules which greatly reduce herbicide efficacy (Dimaano and Iwakami Reference Dimaano and Iwakami2021; Yu and Powles Reference Yu and Powles2014). This mechanism is becoming increasingly discovered in conjunction with auxinic herbicide resistance with examples from B. scoparia, P. rhoeas, and A. tuberculatus (De Figueiredo et al. Reference De Figueiredo, Barnes, Boot, De Figueiredo, Nissen, Dayan and Gaines2022a; Kumar et al. Reference Kumar, Jha, Jugulam, Yadav and Stahlman2019; Torra et al. Reference Torra, Rojano-Delgado, Menéndez, Salas and de Prado2021).
We also observed significantly lower expression of glutamate decarboxylase (GAD) in AMBEL-40 (Table 4). The enzyme GAD is responsible for converting glutamate into γ-aminobutyric acid, an important signaling molecule involved in plant responses to stress conditions, such as environmental challenges, mechanical injury, and herbicide exposure (Fromm Reference Fromm2020; Michaeli and Fromm Reference Michaeli and Fromm2015). Similar metabolic adjustments have been reported in glyphosate-resistant Palmer amaranth (Amaranthus palmeri S. Watson), in which changes in amino acid metabolism played a critical role in resistance mechanisms (Maroli et al. Reference Maroli, Nandula, Dayan, Duke, Gerard and Tharayil2015).
There was also increased expression of two very-long-chain aldehyde decarbonylase genes that are involved in cuticular wax biosynthesis, suggesting the potential for reduced herbicide uptake (Table 4). Cuticular wax acts as a protective barrier limiting water loss and herbicide penetration, with alterations in wax biosynthesis significantly impacting herbicide absorption rates (Bernard and Joubès Reference Bernard and Joubès2013). However, it is unlikely this is playing a role in our system, as AMBEL-40 is susceptible to the other auxinic herbicides screened.
While this experiment cannot directly implicate any one of these genes, the 3:13 segregation observed in the F2 crosses indicates that the high level of clopyralid resistance in AMBEL-40 is multigenic and that several of the genes described earlier could be involved in modulating the phenotype. It has been shown in other species that even when a strong TSR mechanism is sufficient to confer resistance, other modulating NTSR genes can influence the level of resistance expressed (Dalazen et al. Reference Dalazen, Pisoni, Rafaeli and Merotto2018; Laforest et al. Reference Laforest, Soufiane, Patterson, Vargas, Boggess, Houston, Trigiano and Brosnan2021). Similarly, dicamba-resistant B. scoparia populations exhibit metabolic-based mechanisms that suggest increased activity of detoxification pathways in addition to potential alterations in auxin signaling (Kumar et al. Reference Kumar, Jha, Jugulam, Yadav and Stahlman2019; LeClere et al. Reference LeClere, Wu, Westra and Sammons2018; Pettinga et al. Reference Pettinga, Ou, Patterson, Jugulam, Westra and Gaines2018). These examples highlight that auxin herbicide resistance often involves a combination of TSR and NTSR mechanisms, reinforcing that many resistance mechanisms are complex rather than straightforward.
Recognizing that herbicide resistance involves multiple interacting genes emphasizes the importance of integrating genetic, molecular, and biochemical approaches. In A. artemisiifolia, such combined methodologies have revealed both target-site and metabolic mechanisms. ALS resistance has been linked to the Trp-574-Leu substitution and enhanced cytochrome P450–mediated detoxification (Patzoldt et al. Reference Patzoldt, Tranel, Alexander and Schmitzer2001; Rousonelos et al. Reference Rousonelos, Lee, Moreira, VanGessel and Tranel2012). Glyphosate resistance in A. trifida has also been associated with EPSPS gene amplification and reduced herbicide translocation (Délye et al. Reference Délye, Duhoux, Pernin, Riggins and Tranel2015; Page et al. Reference Page, Martin, Meloche, Thibodeau, Nurse, Sikkema, Tardif, Cowbrough and Laforest2024). Overall, these studies illustrate how integrating multiple approaches can help in pinpointing the genetic basis of resistance. In our study, resistance followed a two-gene inheritance pattern with a 3:13 segregation ratio, indicating that multiple genes interact to confer clopyralid resistance. No obvious TSR mutations in the most likely target proteins were apparent, but DEG analysis presented several genes that may be good candidates for genes of major effect, including genes involved in ethylene homeostasis, small molecule metabolism, and general stress response. This outcome highlights the need for advanced genetic tools, such as GWAS or targeted mapping, to identify the specific genes and pathways underlying resistance as well as the need to clone the several putative clopyralid resistance genes we discovered into heterologous systems for resistance mechanism validation.
Acknowledgments
The authors would like to thank Bernard Zandstra, Jill O’Donnell, Tye Thompson, and colleagues at Michigan State University Diagnostic Services for their assistance in this research.
Funding statement
This research was funded by a Michigan State University Project GREEEN Grant.
Competing interests
The authors declare no conflicts of interest.





 Open access
Open access