


Introduction
Seabirds are the most threatened bird group worldwide as a result of human-induced threats, including invasive species at nesting sites, fisheries bycatch, climate change impacts and infectious diseases (Dias et al., Reference Dias, Martin, Pearmain, Burfield, Small and Phillips2019; FAO, 2024). Fifty-six per cent of the global total of 314 seabird species are experiencing population declines, with 31% categorized as threatened (Critically Endangered, Endangered or Vulnerable) on the IUCN Red List (Phillips et al., Reference Phillips, Jérome, Dias, Young and VanderWerf2023). In Mexico, 16% of the seabird species occurring in the country (Berlanga et al., Reference Berlanga, Rodríguez-Contreras, Oliveras de ita, Escobar, Rodríguez, Vieyra and Vargas2008) are categorized as threatened on the IUCN Red List (IUCN, 2024). This is of concern as Mexico currently ranks third globally in terms of seabird diversity and second in terms of the number of endemic seabird species that breed within its territory (Croxall et al., Reference Croxall, Butchart, Lascelles, Stattersfield, Sullivan, Symes and Taylor2012). At present, 19% of the country’s seabird species are protected under Mexican law (NOM-059-SEMARNAT-2010), including Townsend’s shearwater Puffinus auricularis (Berlanga et al., Reference Berlanga, Rodríguez-Contreras, Oliveras de ita, Escobar, Rodríguez, Vieyra and Vargas2008).
Townsend’s shearwater is categorized as Critically Endangered on the IUCN Red List (Birdlife International, 2018). Although it breeds only on the Revillagigedo Archipelago in Mexico (Birdlife International, 2018; Fig. 1), its non-breeding distribution, which is restricted to the Pacific Ocean, ranges from Mexico to Central America (Ainley et al., Reference Ainley, Telfer and Reynolds2020). In Mexico, Townsend’s shearwater historically used the islands of Socorro, San Benedicto and Clarión as breeding sites (Anthony, Reference Anthony1898; Hanna, Reference Hanna1926). Like other seabird species, it is subject to several anthropogenic and natural threats that have decimated its breeding sites and resulted in population declines and the extirpation of multiple seabird colonies in the region (Wolf et al., Reference Wolf, Keitt, Aguirre-Muñoz, Tershy, Palacios and Croll2006; Bedolla-Guzmán et al., Reference Bedolla-Guzmán, Méndez-Sánchez, Aguirre-Muñoz, Félix-Lizarraga, Fabila-Blanco and Bravo-Hernández2019).
Revillagigedo Archipelago off the coast of Mexico, showing the areas on (a) Socorro and (b) Clarión Islands where we carried out acoustic surveys.

The introduction of sheep Ovis aries to Socorro in 1869 (Brattstrom, Reference Brattstrom2015) resulted in a loss of breeding habitat that persisted for > 100 years (Ortiz-Alcaraz et al., Reference Ortiz-Alcaraz, Maya-Delgado, Cortés-Calva, Aguirre-Muñoz, Rojas-mayoral, Córdoba-Matson and Ortega-Rubio2016). In addition, human settlements on Socorro, which were first established in 1957, have resulted in the introduction of invasive predators such as domestic cats Felis catus and house mice Mus musculus (Jehl & Parkes, Reference Jehl and Parkes1982). In San Benedicto, volcanic eruptions that occurred in 1952 and 1953 extirpated the breeding population (Brattstrom & Howell, Reference Brattstrom and Howell1956). In recent decades, individuals have occasionally been reported near San Benedicto (Pitman & Ballance, Reference Pitman and Ballance2002), but a Townsend’s shearwater colony on the island has not been confirmed. On Clarión, introduced species, such as sheep, pigs Sus scrofa and rabbits Oryctolagus cuniculus, have deleteriously affected the ecosystem, with feral pigs causing the extirpation of Townsend’s shearwater by the 1980s (Santaella & Sada, Reference Santaella and Sada1991).
Accurately estimating the remaining breeding population size of Townsend’s shearwater is challenging because of the inaccessibility of nesting colonies and the lack of systematic surveys, resulting in variable estimates over time, ranging from tens to thousands of pairs. Using 15 years of pelagic survey data (1980–1994), the average global breeding population was estimated to comprise 10,600 pairs (Spear et al., Reference Spear, Ainley, Nur and Howell1995). In 1981, the first quantitative estimate for Socorro suggested the breeding population comprised 1,000 pairs (Jehl, Reference Jehl1982). Martínez-Gómez & Jacobsen (Reference Martínez-Gómez and Jacobsen2004) combined the maximum counts from 1993 and 1997 and estimated that the breeding population on Socorro comprised 550 pairs. They also predicted a rapid decline in the breeding population as a result of the presence of feral cats, and reported no evidence of nesting activity on Clarión (Martínez-Gómez & Jacobsen, Reference Martínez-Gómez and Jacobsen2004). The recent estimates of a global population of 200–999 individuals (Birdlife International, 2018) and 75 breeding pairs on Socorro indicate a severe decline in abundance (Martínez-Gómez et al., Reference Martínez-Gómez, Matías-Ferrer, Sehgal and Escalante2015). The status of Townsend’s shearwater on San Benedicto remains unknown because of the inaccessibility of the island.
Invasive mammals have been removed from numerous Mexican islands over the past 25 years (Aguirre-Muñoz et al., Reference Aguirre-Muñoz, Bedolla-Guzmán, Hernández-Montoya, Latofski-Robles, Luna-Mendoza, Méndez-Sánchez, López-Hoffman, Chávez-Ortiz, Seminoff and Carpenter2018), allowing some seabird populations to recover (Méndez Sánchez et al., Reference Méndez Sánchez, Bedolla Guzmán, Rojas Mayoral, Aguirre-Muñoz, Koleff and Vargas2022). Grupo de Ecología y Conservación de Islas, in coordination with the Mexican federal government, eradicated pigs from Clarión in 2002 (Aguirre-Muñoz et al., Reference Aguirre-Muñoz, Bedolla-Guzmán, Hernández-Montoya, Latofski-Robles, Luna-Mendoza, Méndez-Sánchez, López-Hoffman, Chávez-Ortiz, Seminoff and Carpenter2018) and sheep from Socorro in 2012 (Ortíz-Alcaraz et al., Reference Ortiz-Alcaraz, Maya-Delgado, Cortés-Calva, Aguirre-Muñoz, Rojas-mayoral, Córdoba-Matson and Ortega-Rubio2016). In 2014, the group began to eradicate cats on Socorro (Ortíz-Alcaraz et al., Reference Ortiz-Alcaraz, Aguirre-Muñoz, Arnaud, Galina-Tessaro, Rojas-Mayoral, Méndez-Sánchez and Ortega-Rubio2017). These actions allowed vegetation to recover (Ortíz-Alcaraz et al., Reference Ortiz-Alcaraz, Maya-Delgado, Cortés-Calva, Aguirre-Muñoz, Rojas-mayoral, Córdoba-Matson and Ortega-Rubio2016) and land birds and reptiles to increase (Ortíz-Alcaraz et al., Reference Ortiz-Alcaraz, Aguirre-Muñoz, Arnaud, Galina-Tessaro, Rojas-Mayoral, Méndez-Sánchez and Ortega-Rubio2017), although the status of the Townsend’s shearwater population has not been assessed.
To document the current status of Townsend’s shearwater on Socorro and Clarión, we conducted the first systematic monitoring (2016–2024) of its remaining breeding populations. We aimed to (1) map the current distribution of breeding colonies, (2) estimate breeding population size, (3) evaluate reproductive success, (4) model population trends under different conservation scenarios, (5) identify current threats at breeding sites and (6) propose novel conservation and restoration actions. The results of this study will be essential for guiding future conservation actions for this Critically Endangered species.
Study area
Fieldwork was conducted on the islands of Socorro and Clarión during the breeding season (November–June, occasionally extending to mid July) from 2016 to 2024. These islands are > 300 km from mainland Mexico in the Eastern Tropical Pacific and form part of the Revillagigedo Archipelago National Park (Fig. 1; SEMARNAT–CONANP, 2019), which is also a World Heritage Site. The California Current and North Equatorial Current systems converge in the region, creating conditions that support high biodiversity (SEMARNAT–CONANP, 2019). Twelve pelagic seabird species are known to breed in the region, including Townsend’s shearwater (SEMARNAT, CONANP & GECI., 2015).
Socorro (13,039 ha) is the largest island in the Revillagigedo Archipelago, with a maximum elevation of 1,050 m (SEMARNAT–CONANP, 2019), semiarid (0–400 m) and subtropical (400–1,050 m) climate zones, and mean daily temperature of 11.2–19.7 °C (SEMARNAT–CONANP, 2019). Potential native predators of Townsend’s shearwater adults and chicks include the red-tailed hawk Buteo jamaicensis socorroensis and land crab Johngarthia oceanica (Whetje et al., Reference Wehtje, Walter, Rodriguez Estrella, Llinas and Castellanos1993; GECI, unpubl. data).
Clarión is the second largest island (1,925 ha) in the Revillagigedo Archipelago, with a maximum elevation of 320 m (CONANP, 2017), and grasslands, scrublands and coastal vegetation, including 58 species of xerophilous plants (CONANP, 2017). A large fire in 1984 and the introduction of mammals have severely modified the vegetation (Everett, Reference Everett1988). Potential native predators of Townsend’s shearwater include the endemic Clarión raven Corvus corax clarionensis and the Clarión snake Masticophis anthonyi (McLelland, Reference Mclelland1926; CONANP, 2017).
Methods
Distribution of breeding colonies
To determine the current distribution of Townsend’s shearwater colonies on Socorro and Clarión, we conducted auditory surveys, acoustic surveys and burrow searches. Auditory surveys were performed during November–April, 2016–2023, on new moon days at dusk (19.00–21.00) and, occasionally, dawn (3.30–5.30). At each site, we recorded the number of calls, call direction, approximate distance and whether the birds emitted calls in flight or from the ground (Raine et al., Reference Raine, Holmes, Travers, Cooper and Day2017). Survey sites were selected based on historical records (Martínez-Gómez & Jacobsen, Reference Martínez-Gómez and Jacobsen2004), mapped, and analysed to identify potential nest areas for subsequent burrow searches.
Acoustic surveys were conducted during November–April, 2019–2023. All call counts were recorded with Song Meter (SM2, SM3 and SM4) Acoustic Recorders (Wildlife Acoustics, USA). On Socorro, acoustic recorders were deployed over a historical nesting area of 1,819 ha at an altitude of 600 m (Jehl, Reference Jehl1982). The acoustic recorders were deployed every 300 m to avoid double counting (Raine et al., Reference Raine, Holmes, Travers, Cooper and Day2017), with two recording schedules: (1) 10 min on, 20 min off (2019–2022) and (2) 1 min on, 5 min off (2022–2023). Recorders operated from 19.00 to 5.30, independent of the moon phase. Recording mode was mono, with a sampling rate of 24–92 KHz, gain of 16.0 dB and preamp of 26 dB. On Clarión, the acoustic recorders (SM4) were deployed every 500 m using the same recording programme and specifications as those deployed on Socorro.
Burrow searches were conducted during November–February and June–July, 2016–2023, in sites with historical records of burrows, high nocturnal activity, circling birds or ground calls. Field personnel conducted exhaustive searches and were accompanied by wildlife detection dogs (pointers and shepherds trained in passive detection) on four occasions (Grymm-Seyfarth et al., Reference Grimm-Seyfarth, Harms and Berger2021). Search tracks were recorded using a GPS device (either handheld or mounted on dog collars).
Reproductive success
Infrared Trophy Cam HD trail cameras (Bushnell, USA) recorded data on burrow status (active or inactive), activity, predation events, fledgling success and cat presence. The cameras operated in night mode and were checked every 15 days. Predation signs were recorded between camera checks.
We visually assessed burrow occupancy (Eq. 1) by the presence of feathers or guano, and cleanliness, and confirmed burrow visits with photographs:
 $Occupancy = {{Number\;of\;active\;burrows} \over {Total\;number\;of\;burrows}}$
$Occupancy = {{Number\;of\;active\;burrows} \over {Total\;number\;of\;burrows}}$
Nest contents were examined with a borescope (NTS 300, 8 mm lens, Teslong Technology, USA). The burrows were checked once per week, and the reproductive metrics of clutch size, hatching success (proportion of eggs that hatch), fledgling success (proportion of chicks that fledge), reproductive success (proportion of eggs that result in fledglings) and reproductive rate (proportion of pairs breeding) were calculated. The breeding seasons of 2016 and 2017 were omitted from the analysis because of the lack of borescope data. In 2019 and 2020, the borescope failed, and breeding signs (e.g. eggshells, down remains and chick carcasses) were used to determine the number of breeders. Reproductive success was estimated considering only confirmed breeders. We included the breeding success results of one occupied artificial burrow from 2019 to 2023 and two of these burrows in 2024.
Population size and dynamic model
We used call rates as a proxy of burrow density (Oppel et al., Reference Oppel, Hervías, Oliveira, Pipa, Silva and Geraldes2014; Hart et al., Reference Hart, Oppel, Humphries, Blackburn and Nam2021). Four fixed recorders were deployed at sites with known burrow abundance during November–May, 2017–2024, using recording schedule 1. Population size was projected with a Leslie matrix (Leslie, Reference Leslie1945; Martínez-Gómez & Jacobsen, Reference Martínez-Gómez and Jacobsen2004), a widely used tool for evaluating age-structured population dynamics when data on habitat distribution, local population structure, occupancy and extinction patterns, and temporal or spatial variation are limited (Akçakaya & Sjögren-Gulve, Reference Akçakaya and Sjögren-Gulve2000). We chose the Leslie matrix to facilitate comparison with the results of Martínez-Gómez & Jacobsen (Reference Martínez-Gómez and Jacobsen2004) and evaluate the effectiveness of conservation actions over time. This enabled us to simulate population trends for 2020–2035 under three scenarios: (1) current (chick predation by land crabs and no cat predation), (2) worst-case (no management interventions and high predation by cats) and (3) optimal management (reduced chick predation by removing land crabs from burrows through trapping, fencing or translocating chicks). Population and reproductive parameters were estimated from monitoring data (2018–2023; Table 1); survival probability (for each age class: 1–6 years) and the cat predation rate were taken from Martínez-Gómez & Jacobsen (Reference Martínez-Gómez and Jacobsen2004).
Reproductive performance of Townsend’s shearwater Puffinus auricularis on Socorro Island, Revillagigedo Archipelago (Fig. 1), during the breeding seasons of 2016–2024. Numbers in parenthesis are breeders in artificial burrows. Blank cells indicate data were not obtained.

1 Based on number of fledglings.
2 No content checks. Egg shell remains used to estimate breeding pairs.
Data analysis
We manually reviewed recordings (Hart et al., Reference Hart, Oppel, Humphries, Blackburn and Nam2021) using Raven Pro 1.5 (Cornell Lab of Ornithology, USA) and counted all Townsend’s shearwater calls that were visible in spectrograms. Calls that were obscured or overlapped were counted by listening to the recording. Recordings that contained calls that were highly obscured by heavy rain or strong winds were excluded. Townsend’s shearwater calls were classified as broad-band information with low fundamentals (< 1.4 kHz) and harmonics reaching 4 kHz (Baptista & Martínez-Gómez, Reference Baptista and Martínez-Gómez2002). The duration of each call was c. 5 s; spectrograms longer than 5 s were considered more than one call (e.g. 10 s = 2 calls). Townsend’s shearwater presence or absence and the number of calls per site were plotted and classified by natural breaks, using QGIS 3.0 (QGIS, 2018). Pictures and videos from camera traps were reviewed manually to confirm the presence or absence of birds, fledglings and predators.
We estimated breeding population size from the call rates in a sample consisting of the three darkest days of each month in each nesting area with a known number of burrows. An average call rate from all recorders was calculated for each year (2019–2021). The density of burrows and call rates from the known nesting areas on Socorro were used to estimate the number of burrows in the surveyed area (including sites without calls; Fig. 2). Through a linear equation, we estimated the total number of breeding pairs (Nbreeding).
Acoustic recording sites on Socorro Island, Revillagigedo Archipelago (Fig. 1), showing the number of calls per site and Townsend’s shearwater Puffinus auricularis burrow locations.

The total population size (
 ${N_{total}}$
; number of pairs) was calculated from the number of breeding pairs (
${N_{total}}$
; number of pairs) was calculated from the number of breeding pairs (
 ${N_{breeding}}$
) and the breeding proportion (
${N_{breeding}}$
) and the breeding proportion (
 ${r_{breeding}}$
) of the population with Eq. (2):
${r_{breeding}}$
) of the population with Eq. (2):
 ${N_{total}} = {{{N_{breeding}}} \over {{r_{breeding}}}}$
${N_{total}} = {{{N_{breeding}}} \over {{r_{breeding}}}}$
The proportion of breeding pairs was calculated with Eq. (3):
 ${r_{breeding}} = {r_{mature}} \times {\rho _{breeding}}$
${r_{breeding}} = {r_{mature}} \times {\rho _{breeding}}$
where
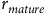 ${r_{mature}}$
is the proportion of sexually mature individuals in the population, and
${r_{mature}}$
is the proportion of sexually mature individuals in the population, and
 ${\rho _{breeding}}$
is the breeding probability of a mature individual (i.e. 0.55), which was calculated with data for 2016–2023. The number of non-breeding pairs (
${\rho _{breeding}}$
is the breeding probability of a mature individual (i.e. 0.55), which was calculated with data for 2016–2023. The number of non-breeding pairs (
 ${{N_{non{\hyphen}breeding}}}$
) was calculated from the total population size and the proportion of non-breeding pairs with Eq. (4):
${{N_{non{\hyphen}breeding}}}$
) was calculated from the total population size and the proportion of non-breeding pairs with Eq. (4):
 ${{N_{non{\hyphen}breeding}}} = {N_{total}} \times {r_{{non{\hyphen}breeding}}}$
${{N_{non{\hyphen}breeding}}} = {N_{total}} \times {r_{{non{\hyphen}breeding}}}$
The proportion of non-breeding pairs (
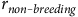 ${r_{{non{\hyphen}breeding}}}$
) was calculated with Eq. (5):
${r_{{non{\hyphen}breeding}}}$
) was calculated with Eq. (5):
 ${{r_{non{\hyphen}breeding}}} = {r_{mature}} \times {\rho _{{non{\hyphen}breeding}}},$
${{r_{non{\hyphen}breeding}}} = {r_{mature}} \times {\rho _{{non{\hyphen}breeding}}},$
where
 ${\rho _{{non{\hyphen}breeding}}} = 1 - {\rho _{breeding}}$
, i.e. the non-breeding probability of a mature individual. Lastly, the number of sexually immature pairs (
${\rho _{{non{\hyphen}breeding}}} = 1 - {\rho _{breeding}}$
, i.e. the non-breeding probability of a mature individual. Lastly, the number of sexually immature pairs (
 ${N_{immature}}$
) was calculated from the total population size and the number of mature non-breeding and breeding pairs with Eq. (6):
${N_{immature}}$
) was calculated from the total population size and the number of mature non-breeding and breeding pairs with Eq. (6):
 ${N_{immature}} = {N_{total}} - \left( {{N_{breeding}} + {{N_{non{\hyphen}breeding}}}} \right)$
${N_{immature}} = {N_{total}} - \left( {{N_{breeding}} + {{N_{non{\hyphen}breeding}}}} \right)$
Results
Distribution of breeding colonies
On Socorro, we recorded Townsend’s shearwater calls in 87 (48%) of the 183 surveyed sites (Fig. 2). We identified 29 (16%) high-potential nesting sites within c. 400 ha, equivalent to 22% of the surveyed area and 3% of the surface area of the island. The other sites were considered to be flyway paths for shearwaters entering or exiting the island. Through burrow searches, we covered c. 145 ha (36%) of the potential nesting area and identified 27 active burrows above an altitude of 800 m that were distributed in small colonies comprising 2–9 burrows. Some isolated burrows were found at sites without calls or with few recorded calls. Eight burrows were occupied by new pairs (i.e. pairs that were not identified in previous surveys or pairs occupying newly dug burrows). In 2023, a new burrow was identified through acoustic recordings in the north-east of the island (600 m altitude).
On Clarión, we identified a breeding pair at Cerro Gallego in 2016. Four new burrows were found from 2017 to 2019. By 2020, we had surveyed 95% of the island with acoustic recorders and identified nine sites with calls and two sites with active burrows (Fig. 3). Calls were located in the east of the island in the La Mujer Dormida site, in the north of the island and in the Cerro Gallego site. Five active burrows were found only on rocky substrates with sparse or no vegetation.
Acoustic recording sites on Clarión Island, Revillagigedo Archipelago), showing the number of calls per site and Townsend’s shearwater burrow locations.

Reproductive success
Reproductive success on Socorro varied among years (mean 43%, range 0–63%). No fledglings were detected in 2018, and only one fledgling, from an artificial burrow, was detected in 2019 (Table 1). The average burrow occupancy during the study period was 92% (76–100%), and the average reproductive rate was 59% (31–82%). Assuming the same birds returned to the same burrows each year, all pairs except two were confirmed to be breeders at least once during the survey period. Most breeding pairs (79%) produced a fledgling at least once, whereas only 28% succeeded in producing fledglings more than once in consecutive years; 21% never produced fledglings. Breeding success in artificial burrows (100% hatching success, 66.6% fledgling success) was higher than in natural burrows over 6 consecutive years. However, in 2024, a new nesting pair in an artificial burrow failed to hatch a chick.
Land crabs predated 24.3% of chicks, which was visually confirmed by direct observations or chick remains in nests (Plate 1). Although crab attacks on fledglings and adults were recorded, crab predation occurred only with chicks aged ≤ 20 days. In addition, 16.2% of chicks disappeared without evidence, although these disappearances coincided with increased crab activity. In 2020 and 2024, the failure of two chicks to fledge in an artificial burrow coincided with the presence of land crabs inside the nest chamber. In 2022, we found evidence of land crabs consuming a broken egg. We did not find any evidence of cat predation. Egg failures were due to disappearances (21.6%), as well as hatching failure (2.7%) and broken (2.7%), non-viable (2.7%) or abandoned eggs (2.7%). In addition, 27% of egg or chick losses could not be explained because the failure occurred between the incubation and hatching stages.
Land crabs Johngarthia oceanica predating Townsend’s shearwater Puffinus auricularis chick in 2021 on Socorro Island, Revillagigedo Archipelago (Fig. 1). Photo: Grupo de Ecología y Conservación de Islas.

On Clarión, average burrow occupancy was 81%. Reproductive success was not estimated because of the depth and complexity of the burrows. However, egg laying at the entrance of a burrow was recorded on video in 2019. Auditory surveys and camera traps confirmed breeding attempts in three burrows, but no fledglings were recorded. Photographic evidence (n = 147 records) showed frequent visits by Clarión snakes (48.3%; Plate 2), land crabs or ravens (21%) and burrowing owls Athene cunicularia rostrata (2.7%). Unlike snakes, land crabs only reached the burrow tunnel, and potentially the nest chamber, 11 times (7.5%); owls reached the tunnel only once (0.7%), and ravens never did.
Clarión snake Masticophis anthonyi visiting an active burrow of Townsend’s shearwater on Clarion Island. Photo: Grupo de Ecología y Conservación de Islas.

Population size and dynamic model
We estimated the Townsend’s shearwater breeding population on Socorro Island was 175 pairs (90%, CI = 94–386 pairs). Considering all other non-breeders and young birds at sea, the global population was estimated to be 481 pairs (962 individuals). The breeding population on Clarión was not estimated because of insufficient data. However, with the available data from field observations and surveys, we conservatively estimated that no more than 10 breeding pairs inhabit the island.
The total population of Socorro, including breeders, non-breeders and young birds, was modelled according to the current scenario, which indicated that the population will continue to decline at a rate of 1% over the next 15 years. The worst-case scenario model, which included high predation by cats, indicated that without cat eradication, the breeding population would decrease by 63% over the next 15 years. The optimal management model (reducing chick predation) predicted a 30% increase in population size over the next 15 years (Fig. 4), considering an increase of 0.17 in breeding success (60%).
Population trends of Townsend’s shearwater in the Revillagigedo Archipelago, projected with a Leslie matrix for 2020–2035, under three conservation scenarios: (1) current (chick predation by land crabs and no cat predation), (2) worst-case (no management interventions and high predation by cats), and (3) optimal management (reduced chick predation and a moderate increase in reproductive success).

Discussion
Distribution of breeding colonies
We have provided evidence of the remaining Townsend’s shearwater breeding colonies in northern Socorro after 7 years. However, no evidence of colonies was found in the south, although a few scattered burrows were present to the north-west of Mount Evermann, which differs from previous records (Martínez-Gómez & Jacobsen, Reference Martínez-Gómez and Jacobsen2004). Calling activity and presence of feathers and faeces indicated that Townsend’s shearwater has recovered some of its historical nesting areas (Wehtje et al., Reference Wehtje, Walter, Rodriguez Estrella, Llinas and Castellanos1993) on Socorro.
On Clarión, we confirmed active burrows in the two highest points of the island, Cerro el Gallego and La Mujer Dormida. However, we found no evidence of colonies in sandy soil on grasslands, as reported by Anthony (Reference Anthony1900) and Hanna (Reference Hanna1926). Townsend’s shearwater burrows were confined to rocky areas above 200 m. The absence of colonies on grasslands may have been because of ravens, which dig into sandy burrows to access chicks and eggs (Hayward et al., Reference Hayward, Atkins, Reichert and Henson2015). In addition, competition with invasive rabbits for habitat (Ávila-Guerrero et al., Reference Ávila-Guerrero, Castillo-Guerrero and Hernández-Vázquez2016) may have also resulted in the absence of colonies. Habitat destruction and predation by invasive pigs (Howell & Webb, Reference Howell and Webb1989), and a fire in 1984 (Everett, Reference Everett1988), may have also negatively affected Townsend’s shearwater nesting habitat.
Reproductive success
Reproductive success on Socorro was highly variable between years and lower (43%) than that of similar species such as Newell’s shearwater Puffinus newelli (66%; Ainley et al., Reference Ainley, Podolsky, Deforest, Spencer and Nur2001), Manx shearwater Puffinus puffinus (62–75%; Lee et al., Reference Lee, Haney, Carboneras, Jutglar and Kirwan2023) and wedge-tailed shearwater Ardenna pacifica (68.5%; Hyrenbach & Hester, Reference Hyrenbach and Hester2022), but similar to that of the black-vented shearwater Puffinus opisthomelas (36%) during El Niño conditions and when subject to predation (Keitt et al., Reference Keitt, Tershy, Croll and Sciences2003). The reproductive failures of Townsend’s shearwater in 2018, 2019, 2021 and 2024 coincided with La Niña-to-neutral and neutral-to-El Niño transitions (NOAA Climate Prediction Center, n.d.). Under neutral and El Niño conditions, sea surface temperatures, frequency of tropical storms (CONANP, 2017), humidity and land crab activity increase (Pérez-Chi, Reference Pérez-Chi2005), which may have been responsible for high chick predation. On Clarión, no Townsend’s shearwater fledglings were recorded during the study period, and the main predators of chicks were likely snakes, which also predate Laysan albatross Phoebastria immutabilis chicks (Wanless et al., Reference Wanless, Aguirre-Muñoz, Angel, Jacobsen, Keitt and Mccann2009). Ravens were unable to access rocky burrows, and land crabs and owls had fewer opportunities to reach chicks because of their nocturnal habits and the presence of adults.
Population size and dynamic model
Our population estimate for Socorro (175 pairs) is more optimistic than the previous estimate of c. 75 pairs (Martínez-Gómez et al., Reference Martínez-Gómez, Matías-Ferrer, Sehgal and Escalante2015). However, the current scenario model predicted a sustained population decline despite invasive mammal management and the increased availability of breeding habitat. This suggests that native predators, particularly land crabs, hinder population recovery. The impacts of land crabs on seabirds have been documented in other regions, such as Cayo Ratón in Puerto Rico, where loss of roseate tern Sterna dougallii chicks was probably a result of predation by the land crab Gecarcinus ruricola (Shealer & Burger, Reference Shealer and Burger1992). Similarly, on Malpelo Island in Colombia, the land crab Johngarthia malpilensis was found to predate Nazca booby Sula granti eggs and chicks (López-Victoria & Werding, Reference López-Victoria and Werding2008).
Kaeding (Reference Kaeding1905) suggested that crab predation may have prevented nesting on Socorro, whereas few crabs were observed on San Benedicto (Anthony, Reference Anthony1900) where thousands of shearwaters were found nesting. Hanna (Reference Hanna1926) also noted low land crab abundance and larger shearwater nesting colonies on Clarión. Thus, Townsend’s shearwater breeding populations on Clarión and San Benedicto may have been historically larger (Anthony, Reference Anthony1898) than on Socorro as a result of predation and the poor available habitat. Previous estimates of the breeding population on Socorro (Jehl, Reference Jehl1982; Whetje et al., Reference Wehtje, Walter, Rodriguez Estrella, Llinas and Castellanos1993; Martínez-Gómez & Jacobsen, Reference Martínez-Gómez and Jacobsen2004) may have been overestimated as a result of the inclusion of immigrants from remnant populations on San Benedicto and Clarión, where nesting habitats were lost (Santaella & Sada, Reference Santaella and Sada1991), or because of shearwater behaviour that led to double counting; e.g. flying in circles while arriving at colonies (Griesemer & Holmes, Reference Griesemer and Holmes2011).
On Clarión, human activities and invasive rabbits have changed food availability for native ravens and snakes (Ávila-Guerrero et al., Reference Ávila-Guerrero, Castillo-Guerrero and Hernández-Vázquez2016). Rabbits, which serve as prey for snakes, inhibit the recovery of vegetation, including cactus (Opuntia sp.; Brattstrom, Reference Brattstrom2015), a food item for ravens. Thus, the reduction in the availability of Opuntia sp. is likely to indirectly increase seabird predation by ravens (Ávila-Guerrero et al., Reference Ávila-Guerrero, Castillo-Guerrero and Hernández-Vázquez2016). The decline of the Townsend’s shearwater population is probably the result of synergistic effects as a result of habitat loss and hyperpredation (Spatz et al., Reference Spatz, Jones, Bonnaud, Kappes, Holmes, Bedolla-Guzmán and Lindsay Young2023a).
Conservation implications and future research needs
On Socorro, removal of cats has significantly reduced the extinction risk of Townsend’s shearwater (Croll et al., Reference Croll, Tershy and Zilliacus2021), and eradication is now 90% completed. Until full eradication, cat incursions can be managed by a permanent field team and interventions during the breeding season. Mitigating chick predation by land crabs is also critical, as highlighted in the optimal management model, which predicts population growth only when predation is reduced. Social attraction techniques have proven to be effective in increasing breeding success, resulting in a positive response (79%) when implemented for 49 procellariid species (Schreiber & Burger, Reference Schreiber and Burger2002). Translocating chicks to safer breeding areas may also improve breeding success, as evidenced by actions in the Hawaiian Islands for Newell’s shearwater (Young & VanderWerf, Reference Young and VanderWerf2024).
Although barn owls Tyto alba have been recorded on Socorro since the 1960s (Jehl & Parkes, Reference Jehl and Parkes1982), their potential impact on Townsend’s shearwater has probably been underestimated given their ability to catch birds in mid-air (Raine et al., Reference Raine, Driskill, Vynne, Harvey and Pias2020). Thus, the ecology of the barn owl on Socorro must be understood to implement management actions successfully during key periods of the Townsend’s shearwater breeding season and to improve fledgling success (Raine et al., Reference Raine, Driskill, Rothe and Travers2022).
On Clarión Island, future restoration actions must prioritize eradication of invasive rabbits, establishing artificial shearwater colonies within predator-proof fences, properly managing human garbage and wastewater, and restoring vegetation. Rabbit eradication will reduce food subsidies for ravens and snakes, and vegetation recovery will promote the availability of native food sources for native fauna (Brattstrom, Reference Brattstrom2015). Predator-proof fences are a cost-effective, short-term measure to protect shearwaters, and should consist of small enclosures (≤ 500 m2) to safeguard island connectivity and the movements of native fauna, which is particularly important for the Critically Endangered Clarión snake Masticophis anthonyi (Ponce-Campos & García-Aguayo, Reference Ponce-Campos and García Aguayo2007).
Management actions for Townsend’s shearwater must be accompanied by long-term monitoring programmes (Raine et al., Reference Raine, Driskill, Vynne, Harvey and Pias2020) that provide real-time information for adaptive management. In addition, the presence of Townsend’s shearwater, potential predation by land crabs, potential competition with the wedge-tailed shearwater for nesting habitat (Villard et al., Reference Villard, Dano and Bretagnolle2006) and habitat erosion (Brattstrom, Reference Brattstrom2015) must be evaluated on San Benedicto. Finally, vocal behaviours must be assessed to improve estimates of population size and dynamics (Arneill et al., Reference Arneill, Perrins, Wood, Murphy, Pisani, Jessopp and Quinn2019), and post-breeding studies are needed to identify potential threats at sea, such as bycatch and the effects of climate change (Días et al., Reference Dias, Martin, Pearmain, Burfield, Small and Phillips2019; Rodríguez et al., Reference Rodríguez, Arcos, Bretagnolle, Dias, Holmes, Louzao and Chiaradia2019).
Author contributions
Study design: AOA, FSC; funding acquisition: FMS, AAM, AOA; fieldwork: FSC, IML, EPV, YBG, AFB, AAO, JGS, NCH, BRM; data analysis: BRM, FSC; writing: FSC, YBG, AFB; revision: all authors.
Acknowledgements
We are grateful for the institutional support from Secretaría de Gobernación, Secretaría de Marina, Secretaría de Medio Ambiente y Recursos Naturales, and Comisión Nacional de Áreas Naturales Protegidas, and the funding provided by the National Fish and Wildlife Foundation, the David and Lucile Packard Foundation, Marisla Foundation, American Bird Conservancy, and the Mohamed Bin Zayed Species Conservation Fund. The study was conducted with permits from Secretaría de Gobernación, Dirección General de Vida Silvestre-Secretaría de Medio Ambiente y Recursos Naturales, and Dirección del Parque Nacional Revillagigedo. We thank all individuals involved in fieldwork.
Conflicts of interest
None.
Ethical standards
This research abided by the Oryx guidelines on ethical standards, approved by Comisión Nacional de Áreas Naturales Protegidas under Mexico’s laws by Secretaría de Medio Ambiente y Recursos Naturales. This study did not involve human subjects, experimentation with animals, or specimen collection, and was approved by Dirección General de Vida Silvestre. Non-invasive camera monitoring was conducted following ethical guidelines to avoid disturbance to wildlife or people.
Data availability
Raw data are available from the corresponding author upon reasonable request.








 Open access
Open access