


Introduction
Graphidaceae is the second largest family of lichenized fungi after Parmeliaceae, with c. 2300 species in over 80 genera (Lücking et al. Reference Lücking, Johnston, Aptroot, Kraichak, Lendemer, Boonpragob, Caceres, Ertz, Ferraro and Jia2014, Reference Lücking, Hodkinson and Leavitt2017; Kalb et al. Reference Kalb, Lücking and Kalb2018; Weerakoon et al. Reference Weerakoon, Aptroot, Lücking, Arachchige and Wijesundara2019; Feuerstein et al. Reference Feuerstein, Lücking and da Silveira2022a, Reference Feuerstein, Aptroot, da Silveira, Lücking and Cáceresb; Aptroot et al. Reference Aptroot, Lücking and Cáceres2023, Reference Aptroot, Lücking and Cáceres2024). It is most diverse in tropical regions, with a small number of species extending into or found in temperate zones (Lücking & Rivas Plata Reference Lücking and Rivas Plata2008; Lücking et al. Reference Lücking, Johnston, Aptroot, Kraichak, Lendemer, Boonpragob, Caceres, Ertz, Ferraro and Jia2014). In Colombia, c. 250 species of Graphidaceae in 44 genera – out of a total of over 3000 lichenized species in c. 370 genera known from the country – are currently known, representing more than 10% of the world’s diversity of this family (Lücking et al. Reference Lücking, Moncada, Soto-Medina, Simijaca and Sipman2021, Reference Lücking, Álvaro-Alba, Moncada, Marín-Canchala, Tunjano and Cárdenas-López2023; Moncada et al. Reference Moncada, Rincón-Murillo and Lücking2023; Soto-Medina et al. Reference Soto-Medina, Aptroot and Lücking2023; Rincón-Murillo et al. Reference Rincón-Murillo, Simijaca, Esquivel, Lücking and Moncada2024). Many species of Graphidaceae have been described as new or reported as new records from Colombia in recent years (Peláez et al. Reference Peláez, Moncada and Lücking2014; Ardila-Rios et al. Reference Ardila-Rios, Moncada and Lücking2015; Soto-Medina et al. Reference Soto-Medina, Lücking and Torres2017, Reference Soto-Medina, Aptroot and Lücking2023; Lücking et al. Reference Lücking, Moncada, Martínez-Habibe, Salgado-Negret, Celis, Rojas-Zamora, Rodríguez-M, Brokamp and Borsch2019, Reference Lücking, Álvaro-Alba, Moncada, Marín-Canchala, Tunjano and Cárdenas-López2023; Motta et al. Reference Motta, Amórtegui, Moncada and Lücking2019).
Graphidaceae exhibits similarities in evolutionary patterns with its sister family, Gomphillaceae. Whereas both families are primarily lichenized, some early diverging lineages include peculiar, non-lichenized fungi. In Graphidaceae, these are found in the subfamily Rubikioideae, with the genera Furcaspora Boar and Rubikia H. C. Evans & Minter (Cáceres et al. Reference Cáceres, Lücking, Schumm and Aptroot2020); in Gomphillaceae, these are represented by five genera in the subfamily Cladosterigmoideae, namely Cladosterigma Pat., Cytosporella Sacc., Neoacrodontiella Crous & M. J. Wingf., Nothoramularia Crous et al., and Vanderaaea Crous (Guterres et al. Reference Guterres, dos Santos, Silva, Souza, Soares, Pinho and Dianese2020; Piątek et al. Reference Piątek, Stryjak-Bogacka and Czachura2025).
In addition to lichenized and non-lichenized (saprotrophic or plant pathogenic) lineages, both families include a third lifestyle, lichenicolous fungi growing on other lichens. In Gomphillaceae, these include Adelphomyces Xavier-Leite et al., Corticifraga D. Hawksw. & R. Sant., Paragyalideopsis Etayo and Taitaia Suija et al., plus a small number of species with uncertain generic placement currently kept in Gyalideopsis Vězda (Pino-Bodas et al. Reference Pino-Bodas, Zhurbenko and Stenroos2017; Suija et al. Reference Suija, Kaasalainen, Kirika and Rikkinen2018; Roux Reference Roux2022; Xavier-Leite et al. Reference Xavier-Leite, Cáceres, Aptroot, Moncada, Lücking and Goto2022, Reference Xavier-Leite, Goto, Lücking and Cáceres2023; Lebreton et al. Reference Lebreton, Ertz, Lücking, Aptroot, Carriconde, Ah-Peng, Huang, Chen, Stenger and Cáceres2025). The best-known lichenicolous taxa in Graphidaceae are certain species of Diploschistes Norman, growing as youth parasites on Cladonia P. Browne, Stereocaulon Hoffm. and other genera, as well as Chroodiscus (Müll. Arg.) Müll. Arg, with several of its species growing on foliicolous Porina Müll. Arg. (Richardson Reference Richardson1999; Lücking & Grube Reference Lücking and Grube2002; Fernández-Brime et al. Reference Fernández-Brime, Llimona, Lutzoni and Gaya2013; Lumbsch et al. Reference Lumbsch, McCune and Morris2023; Diederich et al. Reference Diederich, Lawrey and Ertz2025).
As part of a broader study on Colombian dry forest lichens (Lara-Rojas Reference Lara-Rojas2025), we describe here a new and unique lichenicolous species of Graphis Adans., found as an adelphoparasite on lichens of the same genus, encountered in the centre of Colombia in an area of tropical dry forest, one of the most threatened ecosystems in Colombia and the tropics (Gonzáles-M. et al. Reference González-M, García, Isaacs, Cuadros, López-Camacho, Rodríguez, Pérez, Mijares, Castaño-Naranjo and Jurado2018; Kattan et al. Reference Kattan, Sánchez, Vélez, Ramírez and Celis2019). The vegetation cover in the area consists of secondary remnants of tropical dry forest, which remain under ongoing threat from agricultural activities, extensive livestock farming, and urban construction (Concejo del Municipio Girardot 2020). Due to these threats, biodiversity is likely to decrease as the region’s forests continue to disappear. This situation is not only critical due to the lack of biological knowledge about the area, but also because it is representative of the situation in Colombia and other biodiversity-rich regions (Pereira et al. Reference Pereira, Passos, Santos, Lücking and Cáceres2018; Ministerio de Ambiente 2021; Lara-Rojas Reference Lara-Rojas2025).
Together with the description of the new species, we provide a survey on lichenicolous taxa in Graphidaceae and discuss their phylogenetic relationships, evolutionary patterns, and life strategies as compared to its sister family, Gomphillaceae.
Material and Methods
The material was collected in the municipality of Girardot, Cundinamarca, Colombia, which is located in the Alto Magdalena Province, at a vertex of the Eastern Cordillera, at an elevation ranging from 290 to 1000 m above sea level (Lara-Rojas Reference Lara-Rojas2025). To the south, it borders the Tolima department, and to the north, the municipality of Tocaima. Girardot is characterized by a predominance of tropical dry forest, although only small areas of original vegetation remain and access to these is difficult. Besides agriculture, pastures and urban areas, secondary vegetation is more abundant and dominates many of the forest remnants (Gobernación de Cundinamarca 2002). Lichens with a focus on Graphidaceae were collected in the rural areas of Guabinal, north of the city of Girardot (Lara-Rojas Reference Lara-Rojas2025; Fig. 1).
Map of the collection site in the south-west of the department of Cundinamarca, Colombia, showing the location of transect 3, where the Graphis epigraphis specimens were collected. This transect was the most remote site from the city of Girardot. In colour online.
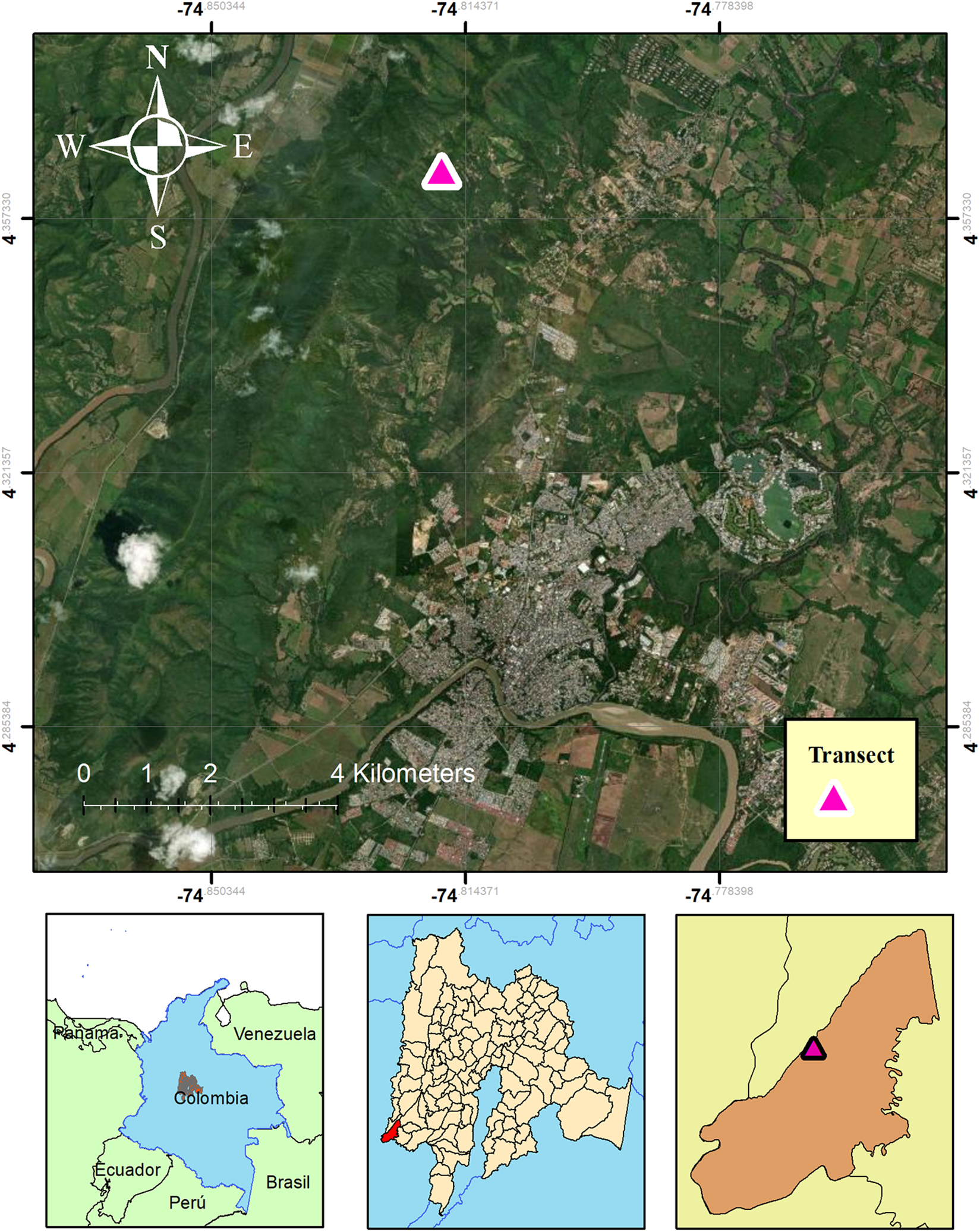
Figure 1. Long description
The top panel is a satellite map oriented with North at the top, showing a green mountainous region with a river and urban area in the lower half. A pink triangle marks transect 3 in the northwest quadrant, far from the main city. A yellow box at the lower right contains the label Transect and a pink triangle. The map includes a compass rose, latitude and longitude grid, and a 4 kilometers scale bar. Below, three smaller maps are arranged left to right. The leftmost shows northern South America with Colombia highlighted. The center map shows Cundinamarca department with the southwest region marked in red. The rightmost map zooms into the local area, showing the transect location with a pink triangle inside a brown region. All maps use consistent color coding to indicate the transect site.
Morphological and anatomical examination was carried out in the laboratory of the Herbario Forestal ‘Gilberto Emilio Mahecha Vega’, Sección Criptógamas, of the Universidad Distrital Francisco José de Caldas (UDBC), using a Leica S6D dissecting microscope and a Zeiss Axiostar Plus compound microscope. Anatomical measurements were taken in water mounts. Sections and squash mounts were examined in distilled water, 10% KOH solution (K) and Lugol’s iodine solution (I). Ascospore measurements were based on observations from hand-cut sections of two collections. All specimens examined are deposited in the Herbario Forestal ‘Gilberto Emilio Mahecha Vega’ (UDBC), Universidad Distrital Francisco José de Caldas, Bogotá, Colombia.
To assess the systematic placement of the new species and of Allographa pauciloculata (Coppins & P. James) Aptroot & Weerakoon, we performed morphology-based phylogenetic binning as described in Berger et al. (Reference Berger, Stamatakis and Lücking2011), Lücking & Kalb (Reference Lücking and Kalb2018) and Wilk & Lücking (Reference Wilk and Lücking2024). For that purpose, we expanded the character matrix used in Lücking & Kalb (Reference Lücking and Kalb2018) by the two aforementioned species (see Supplementary Material File S1, available online) and used the same molecular reference tree, composed of 38 taxa, 23 representing Graphis and 15 Allographa. The binning algorithm, implemented in RAxML v. 7 (Berger et al. Reference Berger, Stamatakis and Lücking2011), first computes individual weights for each character, based on the distribution of corresponding states on the molecular reference tree, and then assesses the placement of each species within the most likely clades of the reference tree using non-parametric bootstrapping of the morphological character matrix.
Taxonomy
Graphis epigraphis Lara-Rojas, Lücking & B. Moncada sp. nov.
MycoBank No.: MB 862274
Differing from the only other genuinely lichenicolous species known in the Graphis-Allographa complex, Allographa pauciloculata, in the lirellae forming regularly stellate (vs irregular) clusters covered by a lateral thalline margin formed by the host thallus (vs lacking a thalline margin), the laterally (vs completely) carbonized excipulum, and the transversely septate (vs partially submuriform) ascospores.
Type: Colombia, Cundinamarca, Girardot, Vereda Guabinal, Cerros de Guabinal, adjacent to Finca ‘Ancestral’, 4°21′49.6″N, 74°49′04.2″W, 527–600 m, secondary remnants of tropical dry forest, on thallus of Graphis leptoclada growing on tree bark, 15 April 2022, Carlos Lara-Rojas M75b-16 (UDBC C-00010531—holotype!).
(Fig. 2)
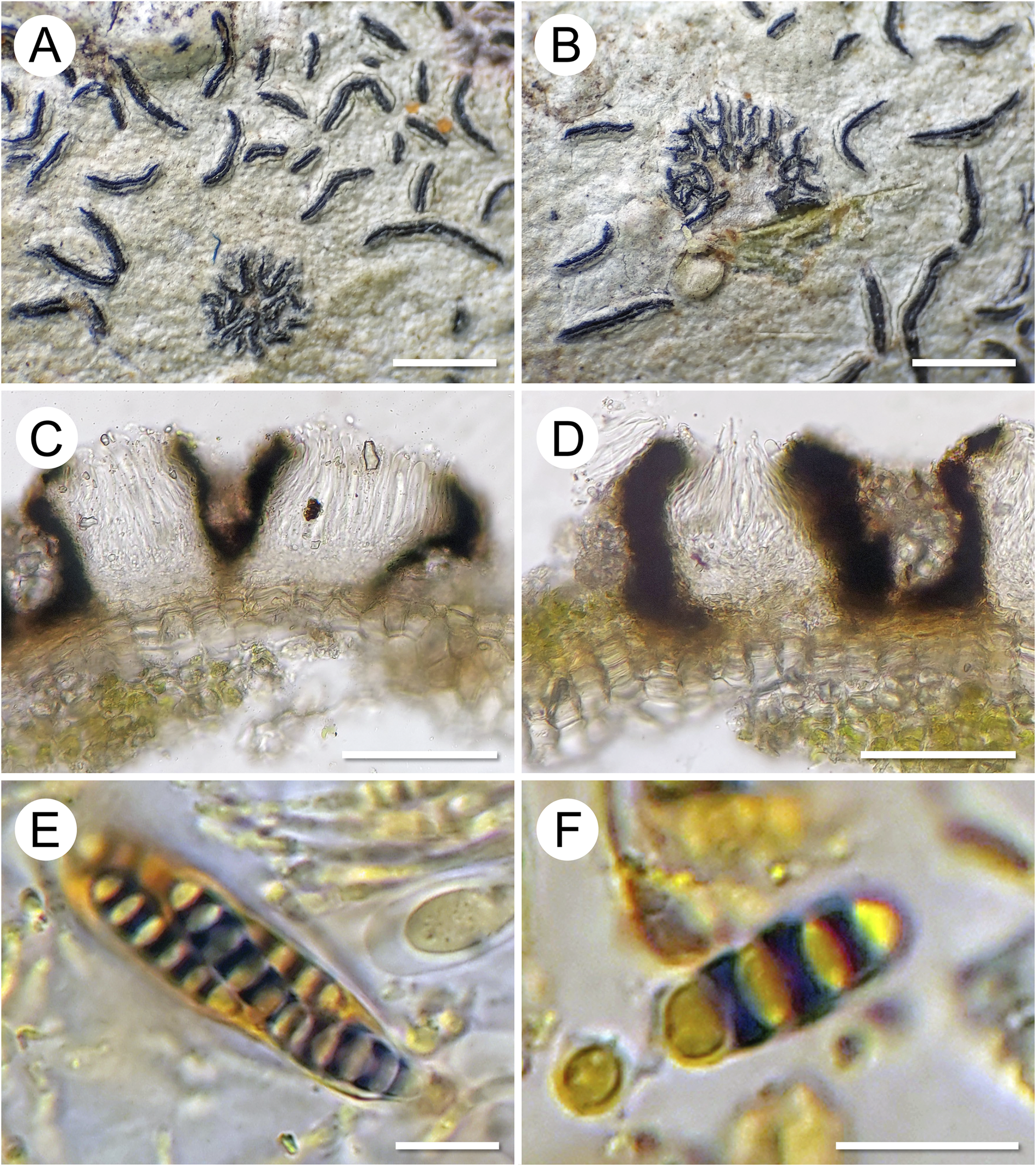
Morphology and anatomy of Graphis epigraphis. A & B, morphology of the two specimens collected (A, on G. pinicola, holotype; B, on G. leptoclada, paratype). C & D, lirellae (holotype) in section showing a non-inspersed (clear) hymenium and laterally carbonized excipulum, in D after applying KOH solution. E, ascus with ascospores (five ascospores visible, the others covered or already ejected), showing amyloid reaction of the ascospore septa in Lugol’s iodine solution (I). F, ascospore showing amyloid reaction. Scales: A & B = 1 mm; C & D = 100 μm; E & F = 10 μm. In colour online.
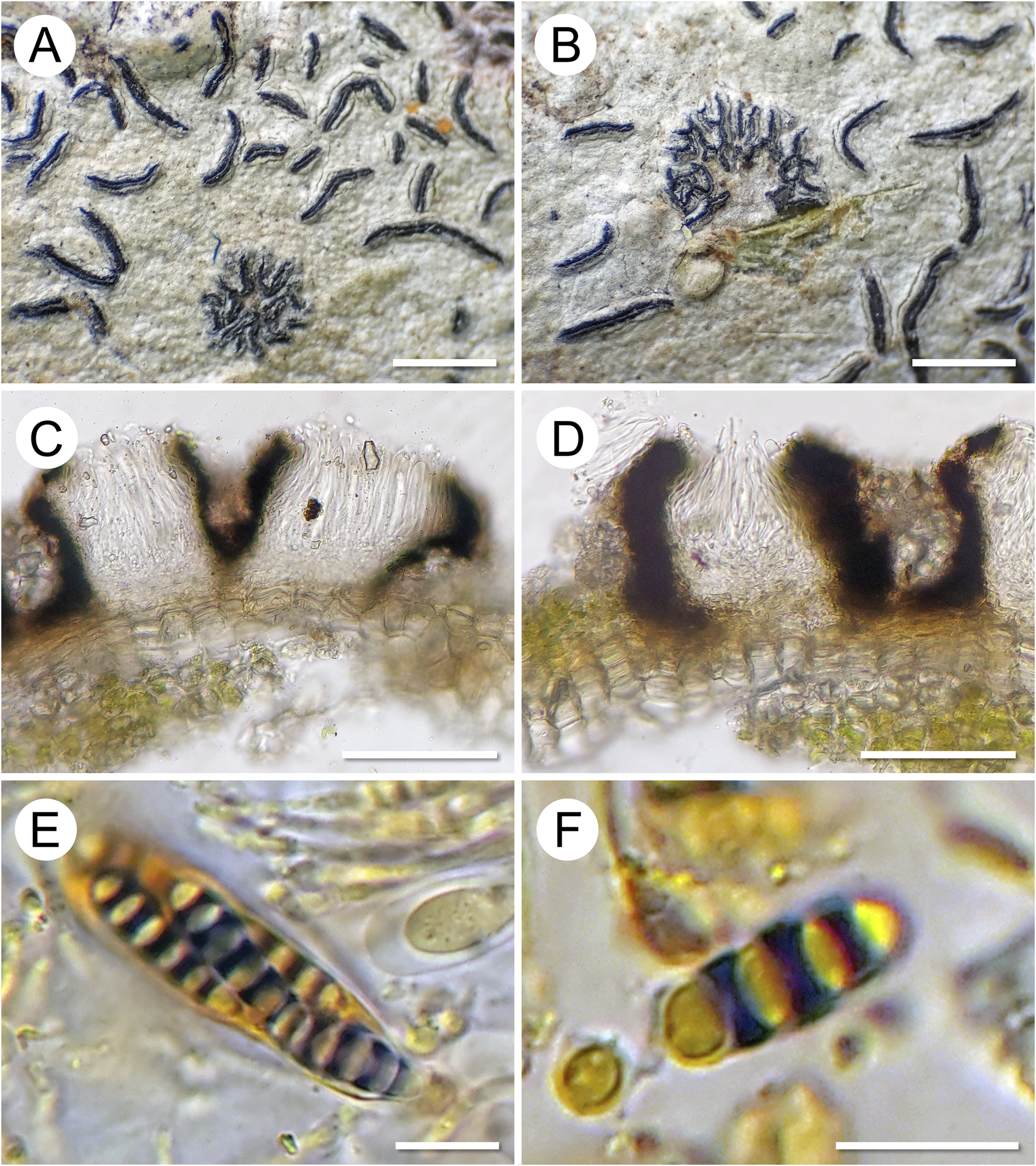
Figure 2. Long description
Top row, leftmost panel A shows numerous dark, curved lirellae scattered on a pale substrate, representing the holotype on G. pinicola. Center panel B displays a similar arrangement with a cluster of lirellae on G. leptoclada, the paratype. Bottom row, leftmost panel C presents a cross-section of a lirella from the holotype, revealing a clear hymenium and laterally carbonized excipulum. Center panel D shows a similar section after K O H application, highlighting the carbonized excipulum. Bottom row, leftmost panel E depicts an ascus containing five visible ascospores, with septa showing an amyloid reaction in Lugol’s iodine. Rightmost panel F focuses on a single ascospore with a pronounced amyloid reaction. Scale bars: panels A and B equal 1 millimeter, C and D equal 100 micrometers, E and F equal 10 micrometers.
Lichenicolous on the thallus of Graphis leptoclada Müll. Arg. (holotype) and G. pinicola Vain. (paratype) based on two specimens, without forming its own thallus.
Ascomata lirellate, forming stellately branched patches, very delicate, patches up to 2 mm diam., individual lirellae up to 0.5 mm long, 0.1–0.15 mm wide, dichotomously branched, becoming disconnected in the centre of the patches. Labia entire, black, laterally covered by the thin, somewhat irregular, thalline margin of the host thallus. Excipulum in section laterally carbonized, 15–25 μm thick, between individual lirellae sometimes with clusters of calcium oxalate crystals from the host thallus. Hymenium colourless, not inspersed (clear), 70–80 μm high; paraphyses unbranched, distinctly septate, apically slightly thickened, 1.5–2 μm thick; epithecium indistinct; hypothecium prosoplectenchymatous, 10–15 μm high. Asci clavate, 50–60 × 10–12 μm. Ascospores (4–)8 per ascus, oblong, with the distal end rounded and the proximal end somewhat acute, (3–)5-septate, (14–)17–20(–22) × (4.5–)5.0–5.5(–6.0) μm, length/width ratio c. (3.0–)3.5–4.0(–4.5) (n = 24), with slightly thickened outer walls, strongly thickened septa (0.5–1.5 μm) and lens-shaped lumina, colourless, septa I+ dark violet-blue (Fig. 2).
Secondary chemistry
No lichen substances detected with standard spot tests in microscopic sections (K−, C−, P−); TLC not performed due to the minute ascomata.
Etymology
From the Greek epi- (= upon), and Graphis, the host lichens.
Distribution and ecology
Thus far known only from the type locality, a secondary dry forest remnant near Girardot in the department of Cundinamarca in Colombia, growing on corticolous thalli of Graphis leptoclada and G. pinicola. The new species was found only in one of the three sampled transects, the one most conserved and most remote from the city of Girardot (Fig. 1). The host lichens are widespread tropical species (Lücking et al. Reference Lücking, Archer and Aptroot2009) but so far no lichenicolous Graphis has been reported from these or other Graphis species.
Remarks
This is only the second truly lichenicolous species of Graphis s. lat. known. The other species, originally described as Graphina pauciloculata Coppins & P. James from Great Britain (Coppins & James Reference Coppins and James1978) and recently recombined as Allographa pauciloculata (Coppins & P. James) Aptroot & Weerakoon (Aptroot et al. Reference Aptroot, Lücking and Cáceres2023), was at first considered an autonomous lichen, but later observed growing lichenicolous on Allographa ruiziana auct., now recognized as A. anomala (Leight.) Aptroot & Weerakoon (Aptroot et al. Reference Aptroot, Lücking and Cáceres2023).
The new species, Graphis epigraphis, shares with Allographa pauciloculata the formation of the lichenicolous ascomata as patches of irregular to stellate clusters on the host thalli, as well as the very small ascospores, at the lower end of the size variation found in both Graphis and Allographa (Lücking et al. Reference Lücking, Archer and Aptroot2009; Lücking & Kalb Reference Lücking and Kalb2018). In A. pauciloculata, the ascospores are partly 3-septate, partly submuriform and somewhat shorter and hence relatively wider (13–15 × 5–6 μm; length/width ratio 2.5–3). Also, the individual lirellae lack a thalline margin, the lirellae in the irregular rather than stellate clusters are secondarily divided by irregular transverse sections and do not become disconnected in the centre, and the excipulum is completely carbonized (Fig. 3).
External morphology of Allographa pauciloculata from the UK for comparison. A & B, clusters of lirellae growing on the thallus of Allographa anomala (holotype; Rose & Coppins s. n.), in B enlarged. C, numerous, partly clustered lirellae growing between partly dead lirellae of A. anomala (Coppins s. n., BM001242329). D, specimen with predominantly 3-septate ascospores, with lirellae separate to clustered (P.W. James s. n., BM001242330). Scales = 1 mm. In colour online.

Figure 3. Long description
Panel A at the top left shows numerous small, black lirellae scattered and clustered across a pale, cracked thallus surface. Panel B at the top right presents a closer view with fewer, larger clusters of lirellae, some adjacent to a woody fragment. Panel C at the bottom left displays many lirellae, some in dense groups, interspersed among pale, partly dead lirellae and reddish patches. Panel D at the bottom right features mostly separated lirellae with some clusters, and a predominance of 3-septate ascospores, visible as elongated black structures. All panels include a white scale bar indicating 1 millimeter for reference.
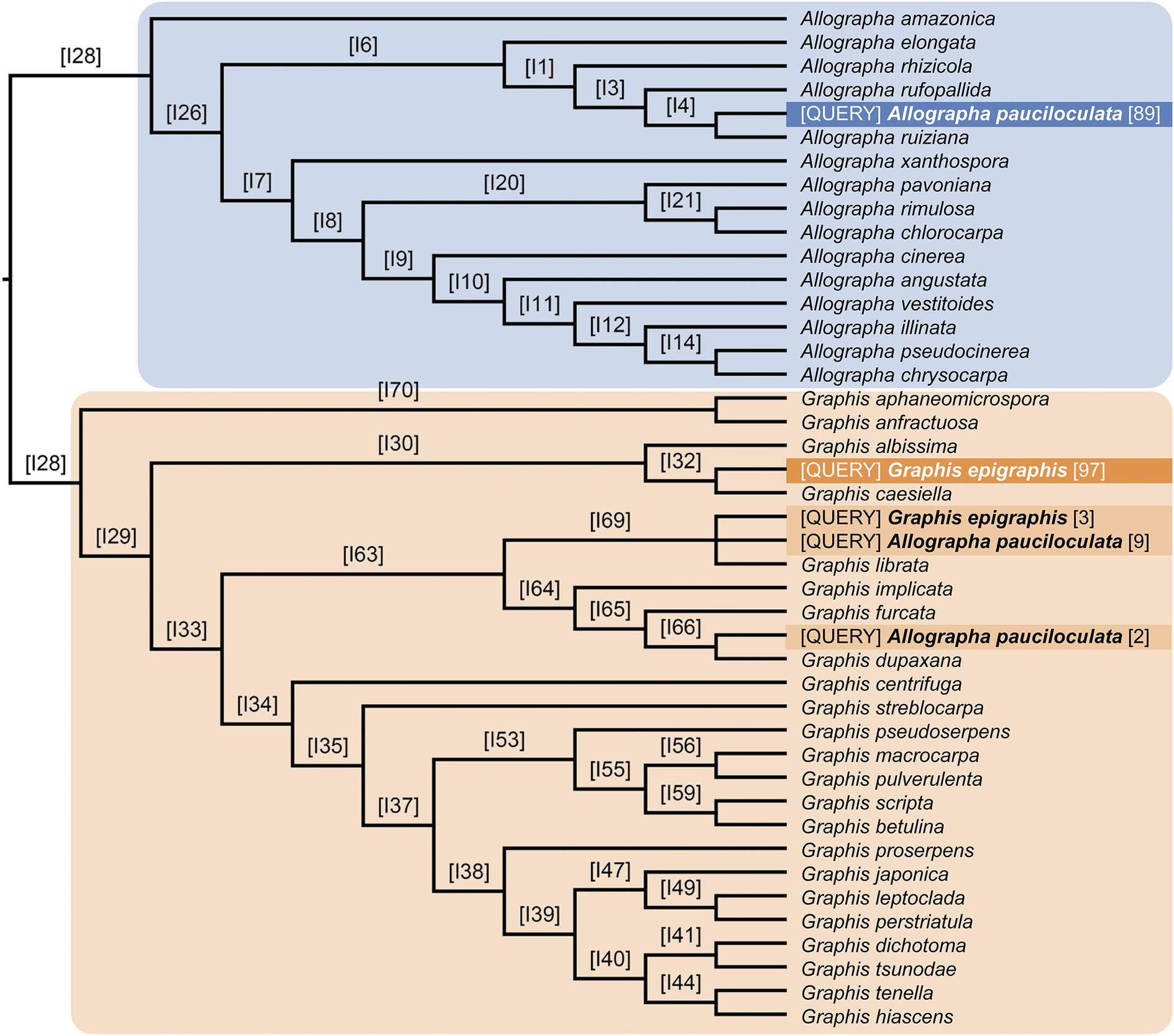
Lücking et al. (Reference Lücking, Archer and Aptroot2009) doubted the placement of Graphina pauciloculata within Graphis s. lat., which has recently been divided into Graphis s. str. and Allographa, based on molecular data (Rivas Plata et al. Reference Rivas Plata, Hernández-M, Lücking, Staiger, Kalb and Cáceres2011; Kalb et al. Reference Kalb, Lücking and Kalb2018; Lücking & Kalb Reference Lücking and Kalb2018). The placement of Graphis pauciloculata within Allographa, as suggested by Aptroot et al. (Reference Aptroot, Lücking and Cáceres2023), was supported by our binning analysis (Fig. 4). It was recovered in three different positions: with 89% support in the Allographa clade, sister to A. ruiziana (Fée) Lücking & Kalb, and with 11% in the Graphis clade, sister to either G. librata C. Knight (9%) or G. dupaxana Vain. (2%). In contrast, the new species, G. epigraphis, was recovered with 100% support in the Graphis clade, either sister to G. caesiella Vain. (97%) or sister to G. librata (3%).
Results of the morphology-based phylogenetic binning analysis of Graphis epigraphis (two alternative placements) and Allographa pauciloculata (three alternative placements). [QUERY] = query taxa binned on the molecular reference tree based on their morphological features. Letters and numbers above branches, e.g. [I4], indicate node numbers of the conserved topology of the molecular reference tree, and numbers in brackets after the query taxa, e.g. [89], indicate placement support for the two binned query taxa. In both cases, the query taxa have more than one alternative placement, but the mosty likely placement is strongly supported in each case.
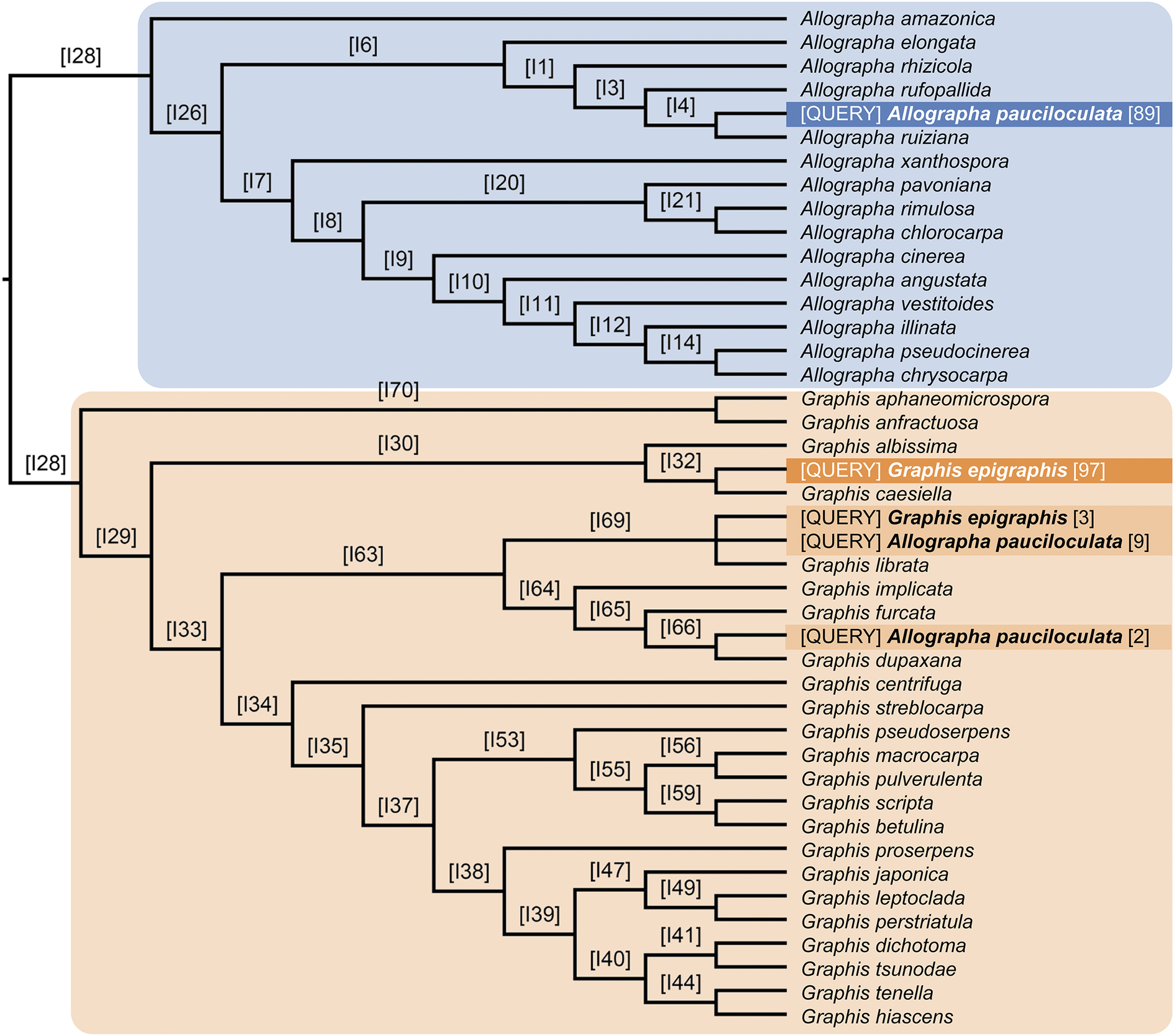
Figure 4. Long description
Starting at the left root labeled I28, the tree splits into two main clades. The upper clade, shaded blue, contains Allographa species, with Allographa pauciloculata placed as a query taxon at three positions: near Allographa rufopallida with support 89, and in the lower clade at two positions with supports 9 and 2. The lower clade, shaded tan, contains Graphis species. Graphis epigraphis is placed as a query taxon at two positions: near Graphis albissima with support 97, and near Graphis caesiella with support 3. Each branch is labeled with a node number in brackets, such as I6, I1, I3, and so on. Query taxa are highlighted in blue or orange, with their support values in brackets. The tree structure shows that both query taxa have multiple placements, but the highest support values indicate the most likely positions. All species names and node numbers are listed exactly as shown, with query placements clearly marked.
As outlined in previous studies (Wirth & Hale Reference Wirth and Hale1963, Reference Wirth and Hale1978; Staiger Reference Staiger2002; Berger et al. Reference Berger, Stamatakis and Lücking2011; Lücking & Kalb Reference Lücking and Kalb2018), typical representatives of Allographa can be distinguished from Graphis by their tendency to produce prominent, robust lirellae lacking or with a complete thalline margin and often massively and completely carbonized excipulum, an often densely inspersed hymenium with inspersion of type B, making the asci difficult to discern, anastomosing paraphyses in the lateral part of the hymenium, large ascospores, and a frequent lack of secondary compounds, with the exception of pigments on the lirellae. Typical species of Graphis s. str. (‘Eugraphis’ according to Wirth & Hale (Reference Wirth and Hale1963, Reference Wirth and Hale1978)) instead have immersed to erumpent, delicate lirellae with a lateral thalline margin and laterally carbonized excipulum, a fine (type A) inspersion of the hymenium when present, never anastomosing paraphyses, small ascospores, and often produce secondary compounds in the thallus, particularly norstictic, stictic, salazinic and/or protocetraric acids. However, many species display an intermediate morphology and so are difficult to assign to either genus without molecular data. In such cases, the binning approach has proved useful, since it predicts assignments based on the most similar sequenced species (Lücking & Kalb Reference Lücking and Kalb2018). The two characters that probably predict placement of the two lichenicolous species are the erumpent lirellae with lateral thalline margin and the laterally carbonized excipulum in Graphis epigraphis versus the prominent lirellae lacking thalline margin and with a completely carbonized excipulum in Allographa pauciloculata.
The similarities between both taxa in terms of the clustered ascomata and comparatively small ascospores may thus represent an analogy. Indeed, lichenicolous species in other, unrelated lineages, such as the Arthoniales, may also form clusters, for example Arthonia tavaresii Grube & Hafellner (Grube et al. Reference Grube, Matzer and Hafellner1995), Opegrapha placidiicola (Zhurb.) Nav.-Ros. & Romero (Navarro-Rosinés & Romero Reference Navarro-Rosinés and Romero2020) or species of Opegrapha Ach. on foliicolous lichens (Matzer Reference Matzer1996); this could therefore represent a convergent morphological response associated with the lichenicolous growth habit, rather than providing evidence for a close phylogenetic relationship between Graphis epigraphis and Allographa pauciloculata.
Additional specimen examined (paratype)
Colombia: Cundinamarca: Girardot, Vereda Guabinal, Cerros de Guabinal, adjacent to Finca ‘Ancestral’, 4°21′49.6″N, 74°49′04.2″W, 527–600 m, secondary remnants of tropical dry forest, on thallus of Graphis pinicola growing on tree bark, 15 iv 2022, Carlos Lara-Rojas M75b-17 (UDBC).
Results and Discussion
Lichenicolous species in Graphidaceae and Gomphillaceae
In their previous version of the global checklist of lichenicolous fungi, Diederich et al. (Reference Diederich, Lawrey and Ertz2018) mention members of the genus Diploschistes Norman as lichenicolous members of Graphidaceae, specifically D. bisporus (Bagl.) J. Steiner, D. caesioplumbeus (Nyl.) Vain., D. euganeus (A. Massal.) J. Steiner, and D. muscorum (Scop.) R. Sant. A fifth species, D. stereocaulorum Lumbsch et al., was recently described (Lumbsch et al. Reference Lumbsch, McCune and Morris2023). All are lichenicolous lichens that initially start their growth on other lichens (Cladoniaceae, Lecanoraceae, Ochrolechiaceae, Rhizocarpaceae, Stereocaulaceae) but eventually develop autonomous thalli. A similar youth parasitism has also been reported from the genus Chroodiscus (Müll. Arg.) Müll. Arg., another member of Graphidaceae, with six species facultatively lichenicolous on Porina species (Lücking & Grube Reference Lücking and Grube2002), namely C. australiensis Vězda & Lumbsch, C. coccineus (Leight.) Müll. Arg., C. neotropicus Kalb & Vězda, C. rubentiicola Lücking et al., C. submuralis Lücking, and C. verrucosus R. Sant. et al. (Fig. 5). These species have been included in the second version of the checklist of lichenicolous fungi (Diederich et al. Reference Diederich, Lawrey and Ertz2025). A careful literature search revealed no further lichenicolous taxa in this family other than Allographa pauciloculata and Graphis epigraphis.
Examples of youth parasitism in foliicolous species of Chroodiscus on thalli of Porina (Porinaceae). A, C. australiensis on P. alba (R. Sant.) Lücking. B, C. coccineus on P. karnatakensis Makhija et al. C, C. submuralis on P. epiphylla Fée. D, C. verrucosus on P. epiphylla. Scales = 1 mm. In colour online.

Figure 5. Long description
Panel A at the top left shows C. australiensis on P. alba, with a pale thallus densely covered by small, round, orange-centered spots and a central larger bump. Panel B at the top right displays C. coccineus on P. karnatakensis, with a yellowish surface featuring numerous raised, orange-margined pustules of varying sizes. Panel C at the bottom left presents C. submuralis on P. epiphylla, where the thallus is greenish with scattered orange-centered spots and smaller whitish bumps. Panel D at the bottom right shows C. verrucosus on P. epiphylla, characterized by a rough, yellowish surface with many pale, rounded warts and fewer orange centers. All panels include a white scale bar indicating 1 millimeter at the lower right.
Diederich et al. (Reference Diederich, Lawrey and Ertz2025) placed Diploschistes in its own family, Diploschistaceae, in the order Graphidales, whereas Chroodiscus is placed in Graphidaceae in the order Ostropales, an apparent inconsistency. Placement of Diploschistes in its own family is probably based on Kraichak et al. (Reference Kraichak, Huang, Nelsen, Leavitt and Lumbsch2018). However, such a classification does not reflect the phylogeny of Graphidaceae, which does allow the maintenance of previous, morphology-based classifications, such as Diploschistaceae or Thelotremataceae (Lücking Reference Lücking2019). Indeed, several supported subclades in Graphidaceae, such as the subfamily Fissurinoideae (e.g. Fissurina Fée vs Clandestinotrema Rivas Plata et al.) and tribes Graphideae (e.g. Allographa vs Glyphis Ach.), Ocellularieae (e.g. Ocellularia G. Mey. vs Sarcographina Müll. Arg.) and Wirthiotremateae (e.g. Carbacanthographis Staiger & Kalb vs Asteristion Leight.), contain both graphidoid and thelotremoid members, and Diploschistes is clearly nested within the subfamily Graphidoideae (Rivas Plata et al. Reference Rivas Plata, Parnmen, Staiger, Mangold, Frisch, Weerakoon, Hernández, Cáceres, Kalb and Sipman2013; Lumbsch et al. Reference Lumbsch, Kraichak, Parnmen, Rivas Plata, Aptroot, Cáceres, Ertz, Feuerstein, Mercado-Díaz and Staiger2014).
The evolutionary history of lichenicolous lineages in Graphidaceae differs from that of its sister family, Gomphillaceae. Most lichenicolous Graphidaceae are facultative or youth parasites found in two genera, Chroodiscus and Diploschistes, which also include genuinely lichenized species. This type of facultatively or partially lichenicolous habit that eventually develops into autonomous lichens is not (yet) known from Gomphillaceae. Lichenicolous forms in both Chroodiscus and Diploschistes grow on unrelated host lichens (Diederich et al. Reference Diederich, Lawrey and Ertz2018, Reference Diederich, Lawrey and Ertz2025). In contrast, lichenicolous lineages in Gomphillaceae either represent early diverging, separate lineages, such as Corticifraga D. Hawksw. & R. Sant. or Taitaia Suija et al. (Hawksworth & Santesson Reference Hawksworth and Santesson1990; Suija et al. Reference Suija, Kaasalainen, Kirika and Rikkinen2018; Roux Reference Roux2022), or are nested within the largely foliicolous and lichenized subfamily Gomphilloideae, such as Adelphomyces Xavier-Leite et al. or Paragyalideopsis Etayo (Etayo Reference Etayo2017; Xavier-Leite et al. Reference Xavier-Leite, Cáceres, Aptroot, Moncada, Lücking and Goto2022, Reference Xavier-Leite, Goto, Lücking and Cáceres2023; Lebreton et al. Reference Lebreton, Ertz, Lücking, Aptroot, Carriconde, Ah-Peng, Huang, Chen, Stenger and Cáceres2025).
The host spectrum of lichenicolous lineages in the two families also differs. Species of Corticifraga are mostly found on Lobariaceae, Peltigeraceae and Nephromataceae, but also on Melanohalea O. Blanco et al. and Ramalina Ach. in the Parmeliaceae and Ramalinaceae, respectively (Diederich et al. Reference Diederich, Lawrey and Ertz2018, Reference Diederich, Lawrey and Ertz2025). Taitaia is also found on Lobariaceae, namely the genus Crocodia (Suija et al. Reference Suija, Kaasalainen, Kirika and Rikkinen2018; Diederich et al. Reference Diederich, Lawrey and Ertz2025). Adelphomyces grows on various Gomphillaceae, whereas species currently classified in Gyalideopsis s. lat. and Paragyalideopsis are found on diverse hosts, mostly Parmeliaceae but also Ectolechiaceae, Stereocaulon, and even Trypethelium Spreng. and Dictyonema C. Agardh (Diederich et al. Reference Diederich, Lawrey and Ertz2018, Reference Diederich, Lawrey and Ertz2025). Thus, the only shared host family and genus are Stereocaulaceae (Stereocaulon).
Although the two lichenicolous species of Graphis and Allographa have not yet been sequenced, their current classification characterizes both as adelphoparasites on host lichens of the same genus, respectively, mirroring the adelphoparasitism of Adelphomyces on host lichens of its own family. The two cases also have in common that both the ascomata and the ascospores of the lichenicolous forms are unusually small compared to genuinely lichenized members of the same genera (Lücking Reference Lücking1997; present paper). Such ‘dwarfism’ is also known from other lichenicolous fungi and is seemingly a general consequence of this lifestyle (Matzer Reference Matzer1996; Etayo Reference Etayo2002, Reference Etayo2017; Lawrey & Diederich Reference Lawrey and Diederich2003). A potential explanation for the clustered growth of ascomata in lichenicolous species, together with the tendency to produce small ascospores, may be a strategy to confine reproductive structures to small portions of the host thallus, to minimize energy investment and antagonistic interactions, while at the same time maximizing ascospore production.
Overall, the evolutionary histories of lichenicolous forms in Graphidaceae and Gomphillaceae are different, suggesting that the evolution of this lifestyle in either family was opportunistic or driven by different evolutionary constraints. The near-zero overlap in host lichens underlines that the evolution of lichenicolous forms may have been triggered by ecological factors, including niche overlap between host lichens and ancestors of lichenicolous lineages, and possibly also photobiont relationships.
Both the lichenicolous and the non-lichenized (saprotrophic or plant pathogenic) lifestyle are extremely rare in the two families. In Graphidaceae, of the currently known c. 2300 species, seven are genuinely non-lichenized, clustered in one lineage in the subfamily Rubikioideae (Cáceres et al. Reference Cáceres, Lücking, Schumm and Aptroot2020), and 13 are lichenicolous, representing four genera (Diederich et al. Reference Diederich, Lawrey and Ertz2025), together making up less than one percent of all species in the family. In Gomphillaceae, of the currently known 460 species (Lebreton et al. Reference Lebreton, Ertz, Lücking, Aptroot, Carriconde, Ah-Peng, Huang, Chen, Stenger and Cáceres2025), eight are non-lichenized (clustered in one or possibly two lineages in the subfamily Cladosterigmoideae; Piątek et al. Reference Piątek, Stryjak-Bogacka and Czachura2025) and 17 are lichenicolous (Rivas Plata et al. Reference Rivas Plata, Lücking and Lumbsch2012, Reference Rivas Plata, Parnmen, Staiger, Mangold, Frisch, Weerakoon, Hernández, Cáceres, Kalb and Sipman2013; Xavier-Leite et al. Reference Xavier-Leite, Cáceres, Aptroot, Moncada, Lücking and Goto2022, Reference Xavier-Leite, Goto, Lücking and Cáceres2023; Diederich et al. Reference Diederich, Lawrey and Ertz2025; Lebreton et al. Reference Lebreton, Ertz, Lücking, Aptroot, Carriconde, Ah-Peng, Huang, Chen, Stenger and Cáceres2025), representing five percent of the overall species richness in the family. Hence, the non-lichenized and lichenicolous lifestyles contribute little to the species richness of Graphidales but do substantially to the phylogenetic and biological diversity of this ascomycete order.
Supplementary Material
The Supplementary Material for this article can be found at http://doi.org/10.1017/S0024282926101455.
Acknowledgements
We gratefully acknowledge Ángel María Lara and the late Noel Lara for their crucial field support. The workshop at the UDBC herbarium during which the material was taxonomically assessed was supported by the DRYLICH project ‘Establishing a Network of Expertise for the Lichen Biota of the Tropical Dry Forest’, a bilateral project between the Botanical Garden and Botanical Museum of Berlin and the Universidad del Norte, Barranquilla, funded by the Federal Ministry of Education and Research (Bundesministerium für Bildung und Forschung – BMBF; Förderkennzeichen 01DN23013) and the Ministry of Science, Technology and Innovation (MinCiencias; contract no. 80740-032-2023), within the program ‘Promoting Scientific and Technological Cooperation Projects with Colombia’ (Förderung von Projekten der wissenschaftlich-technologischen Zusammenarbeit mit Kolumbien). We thank two anonymous reviewers for constructive comments that helped to improve the manuscript considerably.
Author ORCIDs
Carlos Lara, 0009-0002-2447-2208; Robert Lücking, 0000-0002-3431-4636.





 Open access
Open access