


Highlights
-
• The gradient values in the right calcarine gyrus and the left supramarginal gyrus in global gradient networks were associated with L2 proficiency.
-
• MVPA and ROI analysis indicated that L2 proficiency may not significantly influence the gradient values of language and cognitive control networks in the global brain activity flow.
-
• The high-proficiency L2 learners showed enhanced inhibitory connections from the language network to the cognitive control network, which might indicate that the development of L2 is accompanied by improved cognitive control abilities.
1. Introduction
Language processing and development rely on an extensive and distributed hierarchical network within the human brain (Kotz, Reference Kotz2009; Perani & Abutalebi, Reference Perani and Abutalebi2005). In particular, the key language processing circuit and the cognitive control network may play an essential role in the development of second language (L2) proficiency. Yet, it was poorly understood whether the acquisition of L2 proficiency would modify the global brain activity flow, which may indicate the spread of activation amplitude and underlie the information propagation flow (Cole et al., Reference Cole, Ito, Bassett and Schultz2016) across the brain, of key brain regions in language and cognitive control networks within large gradient networks. Meanwhile, it also remains unclear how L2 proficiency might influence the local brain activity flow among the key brain regions in these two networks, which may reveal the underlying directed interactions between these two networks related to L2 proficiency. From the perspective of the Adaptive Control Hypothesis (Green & Abutalebi, Reference Green and Abutalebi2013), bilingual language use places recurrent demands on a set of cognitive control processes, such as goal maintenance, conflict monitoring, and interference suppression, and these processes adapt according to the interactional context. This theoretical account predicts that L2 learning not only engages the classic language network but also drives adaptive changes in the cognitive control network, depending on whether speakers operate in single-language, dual-language or code-switching contexts. Such adaptation may be reflected in the functional organization and directed interactions of large-scale brain networks. Thus, our current study aimed to examine whether L2 proficiency would modify the global brain activity flow of language and cognitive control networks in large gradient networks. Furthermore, we investigated how L2 proficiency influences the local brain activity flow among key brain regions of language and cognitive control networks during the resting state, given that the functional interaction patterns of the resting state are stable and underlie the brain activity patterns during task performance (Cole et al., Reference Cole, Ito, Bassett and Schultz2016).
Successful L2 learners demonstrate improved cognitive functions in attention, working memory, and cognitive flexibility (Bialystok, Reference Bialystok2021; Kroll et al., Reference Kroll, Dussias, Bogulski, Kroff and Ross2012). These improvements are associated with enhanced neural efficiency and adaptability, accompanied by heightened activity in cognitive control regions of the brain, such as the prefrontal cortex (PFC) and anterior cingulate cortex (ACC) for the management of bilingual systems (Li et al., Reference Li, Abutalebi, Zou, Yan, Liu, Feng, Wang, Guo and Ding2015; Rafeekh et al., Reference Rafeekh, Krishna, Kapiley and Mishra2021; Tao et al., Reference Tao, Wang, Zhu and Cai2021). Extensive research demonstrated the critical role of the cognitive control network in L2 development besides the key language network (Li et al., Reference Li, Abutalebi, Zou, Yan, Liu, Feng, Wang, Guo and Ding2015; Liu et al., Reference Liu, de Bruin, Jiao, Li and Wang2021; Pliatsikas & Luk, Reference Pliatsikas and Luk2016; Tao et al., Reference Tao, Wang, Zhu and Cai2021). However, it remains unknown how the cognitive control network interacts with the classic language network to establish a smooth bilingual system, which may manifest efficient language switching and inhibition to access native language during L2 use (Darcy et al., Reference Darcy, Mora and Daidone2016; Linck et al., Reference Linck, Kroll and Sunderman2009; Liu et al., Reference Liu, Fan, Rossi, Yao and Chen2016; Xia et al., Reference Xia, An and Guo2022). While some studies revealed that L2 proficiency would influence the functional connectivity between brain regions in language and cognitive control networks (Li & Grant, Reference Li and Grant2016; Liu et al., Reference Liu, de Bruin, Jiao, Li and Wang2020; Sun et al., Reference Sun, Li, Ding, Wang and Li2019; Wang et al., Reference Wang, Ke, Zhang, Zhou, Li and Yang2020), most have not addressed interaction directionality, which is critical for understanding the activity flow across brain regions. In particular, network gradient approaches (Huntenburg et al., Reference Huntenburg, Bazin and Margulies2018; Margulies et al., Reference Margulies, Ghosh, Goulas, Falkiewicz, Huntenburg, Langs, Bezgin, Eickhoff, Castellanos, Petrides, Jefferies and Smallwood2016) can reveal the global brain activity flow pattern in each network gradient and the gradient values of particular brain regions, which may represent the relative position of brain regions in a global brain activity flow in gradient networks. In addition, dynamic causal modelling (DCM; Friston et al., Reference Friston, Harrison and Penny2003, Reference Friston, Kahan, Biswal and Razi2014) can quantify the strength of directed connections among brain regions, which provides important information about the local brain activity flow. Thus, our current study made use of gradient network and DCM to investigate how L2 proficiency influences global and local brain activity flows in language and cognitive control networks.
The large-scale brain network exhibits the gradient architecture (Huntenburg et al., Reference Huntenburg, Bazin and Margulies2018; Margulies et al., Reference Margulies, Ghosh, Goulas, Falkiewicz, Huntenburg, Langs, Bezgin, Eickhoff, Castellanos, Petrides, Jefferies and Smallwood2016), which hierarchically organizes the brain activity flow as a whole to support various cognitive functions. Meanwhile, it was suggested that language functions rely on distributed gradient networks (Huth et al., Reference Huth, de Heer, Griffiths, Theunissen and Gallant2016), which indicates that the global brain activity flow is involved in language processing. However, it is still poorly understood whether the brain activity flow is related to L2 proficiency. Specifically, the gradient values of particular regions, as revealed by the network gradient approach, represent the relative position in the whole brain activity flow of a particular gradient network. Such a network property allows us to examine whether the relative position of a brain region within the global brain activity flow in a gradient network is associated with L2 proficiency, particularly in regions such as language and cognitive control networks.
Nevertheless, the gradient network cannot probe the directionality or strength of interactions among brain regions, which are essential for characterizing the brain activity flow. By contrast, DCM allows the estimation of the strength of directed interactions (Friston et al., Reference Friston, Harrison and Penny2003, Reference Friston, Kahan, Biswal and Razi2014). This approach can enable us to examine how L2 proficiency is related to the locally detailed brain activity flow among language and cognitive control networks. Importantly, language function is supported by distributed brain networks (Huth et al., Reference Huth, de Heer, Griffiths, Theunissen and Gallant2016), among which the interactions between language and cognitive control networks may play a critical role in establishing a smooth bilingual system (Li et al., Reference Li, Abutalebi, Zou, Yan, Liu, Feng, Wang, Guo and Ding2015; Liu et al., Reference Liu, de Bruin, Jiao, Li and Wang2021; Pliatsikas & Luk, Reference Pliatsikas and Luk2016; Tao et al., Reference Tao, Wang, Zhu and Cai2021). Yet, only a few studies contrived to explore how the interactions among the brain regions of language and cognitive control networks are related to L2 learning. A recent study revealed how the directed connectivity in the fronto-striatal circuit is related to L2 learning (Yamasaki et al., Reference Yamasaki, Stocco, Liu and Prat2019), employing DCM to uncover how L2 acquisition shapes causal interactions supporting cognitive control. In parallel, research on functional connectivity has linked L2 proficiency to network changes, although this method cannot specify interaction direction (Chai et al., Reference Chai, Berken, Barbeau, Soles, Callahan, Chen and Klein2016; Ghazi Saidi et al., Reference Ghazi Saidi, Perlbarg, Marrelec, Pelegrini-Issac, Benali and Ansaldo2013; Sun et al., Reference Sun, Li, Ding, Wang and Li2019). For instance, Sun et al. (Reference Sun, Li, Ding, Wang and Li2019) revealed reduced connectivity between ACC and other language brain regions in participants with high L2 proficiency when compared to those with low L2 proficiency, a finding that implies inhibitory connections between language and cognitive control networks. Together with prior findings that L2 learning enhances cognitive control abilities (Bialystok, Reference Bialystok2021; Kroll et al., Reference Kroll, Dussias, Bogulski, Kroff and Ross2012), we hypothesized that there are inhibitory connections from the language network to the cognitive network, and these inhibitory influences are more pronounced in individuals with high L2 proficiency compared to those with low proficiency. Meanwhile, despite the heterogeneity in these findings, they consistently highlighted the involvement of brain regions in language and cognitive control networks. Further examining the brain activity flow among these key brain regions would advance our understanding of how the cognitive control network interacts with the classic language network to support efficient bilingual processing. To the best of our knowledge, no studies have systematically inspected the detailed pattern of directed interactions among the key brain regions in language and cognitive control networks as a function of L2 proficiency. Thus, our current study aimed to address this research gap.
In summary, language processing engages a large distributed brain network across the brain, which involves global and local brain activity flows to coordinate segmented brain regions to achieve language function. Network gradient can examine the global brain activity flow pattern, while DCM can examine the directed and quantified local brain activity flow. Thus, these two approaches can complement each other to enable us to 1) investigate whether L2 proficiency is related to the alteration of the relative position (gradient value) of language and cognitive control networks in the global brain activity flow and 2) explore how L2 proficiency is related to the profile of directed interactions among brain regions in language and cognitive control networks. We hypothesized that there are inhibitory connections from the language network to the cognitive network and that inhibitory connections might be stronger in individuals with high L2 proficiency compared to those with low L2 proficiency. This study aims to deepen our understanding of the interactions between the cognitive control network and the classic language network in establishing an efficient bilingual system. In line with the Adaptive Control Hypothesis (Green & Abutalebi, Reference Green and Abutalebi2013), which posits that bilingual experience adaptively tunes the engagement of cognitive control processes depending on interactional contexts, we further expect that variations in L2 proficiency will be associated with adaptive modifications in language and cognitive control networks in both global and local brain activity flows.
2. Materials and methods
2.1. Participants
Seventy-two college students aged between 18 and 30 years were recruited from the University of Macau campus. All participants were Mandarin Chinese native speakers and learned English as a foreign language through formal instruction after 6 years of age. None of them stayed in an English-speaking country for more than 6 months. All participants reported no physical or mental diseases during the time they took the functional magnetic resonance imaging (fMRI) scan. Participants were excluded if they met any of the following criteria: 1) a history of drug or alcohol abuse, 2) a history of severe brain damage and 3) left-handedness. All participants took the LexTALE English test (Lemhöfer & Broersma, Reference Lemhöfer and Broersma2012), based on which those who scored equal to or below 60 were categorized as the low-proficiency group, while those who scored above 60 were categorized as the high-proficiency group. This study was approved by the Ethics Committee at the University of Macau, and each participant gave their written consent before taking part in the study. The procedures of data acquisition and processing were visualized in Figure 1.
Flow diagram of data acquisition and analysis. Participants with low or high L2 proficiency were recruited and invited to take a resting-state fMRI scan. Two network approaches, including network gradient and dynamic causal modelling (DCM), were adopted to examine the characteristics of the brain activity flow related to L2 proficiency. Regarding the network gradient approach, it mainly included correlational analysis (general linear modelling) to reveal network gradients related to L2 proficiency, between-group tests to examine the potential changes in network gradients in language and cognitive control networks between low- and high-L2-proficiency groups, and multivariate pattern analysis (MVPA) to examine the predictive power for L2 proficiency from network gradients. DCM mainly examined the directed interactions between language and cognitive control networks related to L2 proficiency.

2.2. MRI data acquisition
MRI data were collected with a 3-T Siemens Magnetom Prisma scanner in the Centre for Cognitive and Brain Sciences at the University of Macau. Every participant took a structural MRI scan for approximately 4 minutes and a resting-state functional MRI scan for 8 minutes. The parameters for the structural scan included: FOV = 256 mm; slice number = 176; slice thickness = 1 mm; TR = 2300 msec; TE = 2.26 msec. During the resting state, the participants were instructed to fixate on a white cross against a black background, focusing on no particular thing. The parameters for the resting-state fMRI included: FOV = 192 mm; slice number = 65; slice thickness = 2 mm; voxel size: 2*2*2 mm3; TR = 1 s; TE = 30 msec; phase encoding direction = anterior to posterior; flip angle = 60.
2.3. MRI data preprocessing
The preprocessing procedures of fMRI data were conducted by DPABI (Yan et al., Reference Yan, Wang, Zuo and Zang2016), a package based on MATLAB statistical parametric mapping (SPM). It consists of several main sections, i.e., slice timing, realignment, normalization and smoothing. First, five volumes were removed from the original 600 volumes of resting-state fMRI data for each participant to eliminate the effects of unstable factors at the beginning of the scan. A total of 595 volumes of data were fed to perform slice timing correction to correct differences in image acquisition time between slices. Then, realignment was performed for head motion, which is a serious interference. In addition, confounding effects were removed for further procedures, including constant, linear and quadratic trends, 24 head motion parameters, effects originating from the white matter and cerebral fluid, as well as regressors indicating bad head motion time points. Before smoothing the data with a 4-mm Gaussian kernel, the data were normalized to a standard MNI space, using each participant’s structural scan. Of note, the images were resliced to 4*4*4 mm3 for network gradient analysis, while the images with voxel size 2*2*2 mm3 were used for DCM analysis.
2.4. Network gradient
BrainSpace toolbox was used to calculate network gradients with 10% sparsity of the correlation matrix, with normalized angle similarity as the kernel function and diffusion mapping as the dimension reduction method following previous studies (Nguyen et al., Reference Nguyen, Qian, Ng, Ong, Ngoh, Yeo, Lau, Tan, Broekman, Law, Gluckman, Chong, Cortese, Meaney and Zhou2023; Vos de Wael et al., Reference Vos de Wael, Benkarim, Paquola, Lariviere, Royer, Tavakol, Xu, Hong, Langs, Valk, Misic, Milham, Margulies, Smallwood and Bernhardt2020). Specifically, a 17,881 by 17,881 correlation matrix was obtained by calculating the Pearson correlation for each pair of voxels in an individual. Normalized angle similarity, which can circumvent negative similarities, was used to obtain the square non-negative affinity matrix between voxels with the correlation matrix. Diffusion mapping, with the anisotropic diffusion parameter α set as 0.5, is a nonlinear data mapping and was used to perform dimensionality reduction. Around 132 gradients were obtained in each gradient calculation. However, only the first 10 gradients were retained, which accounted for around 79% variances. Of note, individuals’ Fisher z-transformed correlation matrices were averaged across participants and then back-transformed to the correlation matrix, so as to estimate group-averaged network gradients. This was followed by calculating individual’s network gradients with the same procedure but with group-averaged gradients as reference using Procrustes alignment.
2.5. Explorative whole-brain regression analysis for network gradients
General linear regression (GLM) analysis was conducted with SPM (https://www.fil.ion.ucl.ac.uk/spm/) to unveil brain clusters associated with L2 proficiency in the first 10 gradients. The covariances of gender and age were included as covariates in the GLM. All significant results survived the family-wise error (FWE) correction for multiple comparisons (FWE cluster level p < 0.05, cluster-forming threshold p < 0.001). It is worth noting that in this explorative analysis, LexTALE scores were treated as continuous predictors to reveal L2-proficiency-related brain regions. In subsequent analyses, however, participants were instead divided into two proficiency groups based on their LexTALE scores to examine potential alterations in the brain activity flow linked to L2 proficiency. While the cross-sectional nature of our study does not allow us to directly trace longitudinal development, we consider that the observed group differences may still provide valuable insights into the developmental trajectory.
2.6. Selection of regions of interest for language and cognitive control networks
The online meta-analysis website Neurosynth (https://neurosynth.org/) was used to define language and cognitive control networks (detailed in Supplementary Table S1). Regarding the language network, we selected the top five largest full brain clusters revealed by the association test as our original regions of interest (ROI). Among them, the left inferior frontal gyrus (IFG) is a paramount brain region for language production (Hagoort, Reference Hagoort2005), the left posterior temporal gyrus (pTG) is essential for language comprehension (Binder, Reference Binder2015; Bogen & Bogen, Reference Bogen and Bogen1976), and the left temporal pole plays a key role in semantic processing and has been implicated in various disorders (Patterson et al., Reference Patterson, Nestor and Rogers2007). Additionally, the left supramarginal gyrus (SMG) and right calcarine area, regions frequently linked to semantic processing (Huth et al., Reference Huth, de Heer, Griffiths, Theunissen and Gallant2016), were also identified in our gradient network analyses (see Section 3.2) as associated with language proficiency. Thus, the left IFG, left posterior MTG, left temporal pole, left SMG and right calcarine area were selected as the ROI of the language network.
Regarding the cognitive control network, we selected the top five largest full brain clusters revealed by Neurosynth, including the right posterior middle frontal gyrus (MFG), right ACC, left MFG, right anterior MFG and right insula. These brain regions were also reported in previous studies to be involved in cognitive control (Cai et al., Reference Cai, Chen, Ryali, Kochalka, Li and Menon2016; Niendam et al., Reference Niendam, Laird, Ray, Dean, Glahn and Carter2012; Zhang et al., Reference Zhang, Pines, Stetz, Goldstein-Piekarski, Xiao, Lv, Tozzi, Lavori, Snowden, Venditti, Smyth, Suppes, Ajilore, Ma and Williams2024).
2.7. Multivariate pattern analysis
Multivariate pattern analysis (MVPA) makes use of activation patterns of multiple brain nodes to predict the information processing of brain signals (Haxby, Reference Haxby2012; Treder, Reference Treder2020). The time series of the average gradient value for each ROI was extracted by averaging the gradient values across all voxels within an ROI at each time point. An MVPA toolbox, MVPA-light (Treder, Reference Treder2020), was used to predict L2 proficiency levels (high or low) with averaged gradient values of brain regions in language and cognitive control networks as listed above. The prediction was performed with a linear discriminant analysis algorithm, dividing data into five folds for training and prediction. This analysis was intended to probe whether the gradient values in language and control networks could predict L2 proficiency.
2.8. ROI-based between-group analysis for network gradients
The average gradient value for each ROI in each of the first 10 gradients was extracted for each participant. Two-sample t-tests were performed to examine whether there were between-group differences in the gradient values of each ROI in each network gradient.
2.9. Dynamic causal modelling
The DCM algorithm in SPM 12 (version: 7771) was used to estimate the effective connections among brain nodes of language and cognitive control networks. A fully connected model was firstly built, which indicates that bidirectional connections were between each pair of brain regions. To determine the optimal model for each individual, we applied Bayesian model comparison based on negative variational free energy (Friston et al., Reference Friston, Mattout, Trujillo-Barreto, Ashburner and Penny2007). This approach selects the model with the greatest free energy, thereby achieving a better trade-off between explained variance and model complexity (i.e., number of directed connections; Friston et al., Reference Friston, Kahan, Biswal and Razi2014; Zeidman, Jafarian, Corbin, et al., Reference Zeidman, Jafarian, Corbin, Seghier, Razi, Price and Friston2019). After choosing the best model, Bayesian model inversion was performed to estimate the parameters based on the observed cross-spectral density (Zeidman, Jafarian, Corbin, et al., Reference Zeidman, Jafarian, Corbin, Seghier, Razi, Price and Friston2019). In the initial check after the model estimation at the individual level, it was found that the DCM model for each participant explained variance more than 60%. In the group-level analysis, a parametric empirical Bayes (PEB) framework (Zeidman, Jafarian, Seghier, et al., Reference Zeidman, Jafarian, Seghier, Litvak, Cagnan, Price and Friston2019) was used to estimate the relationship between effective connectivity (EC) and regressors, including proficiency groups, age and gender. Group-averaged common and differential effects were calculated with the Bayesian average. A posterior probability larger than 95% (Pp > 95%) indicates strong evidence for the connection strength or group effects. Of note, the positive (negative) EC between brain regions indicates an excitatory (inhibitory) effect between brain regions, while the positive (negative) EC within brain regions indicates increased self-inhibition (reduced self-inhibition; Zeidman, Jafarian, Corbin, et al., Reference Zeidman, Jafarian, Corbin, Seghier, Razi, Price and Friston2019).
To summarize network-level effective connections, Bayesian contrasts instead of arithmetic average were used, so as to represent network-averaged effects (Zhou et al., Reference Zhou, He, Yuan, Zhou, Yin, Chark, Fong, Fong and Wu2023; Zhou et al., Reference Zhou, Friston, Zeidman, Chen, Li and Razi2018; Zhou et al., Reference Zhou, He, Jin, Zeidman, Gao, Rong, Huang, Feng, Cui, Zhang, Wang, Wang, Xiang and Wang2023). Specifically, it extracted the expected value and the variance for all group-average parameters (EC) related to a regressor, i.e., the group common effect. Then, the cumulative distribution function was used to combine the sets of expected values and variances within a network or between networks to estimate the probability in which the expected value of EC within a network or between networks would differ from zero.
2.10. Test–retest reliability
To validate whether the gradients revealed by this study have stable test–retest reliability, we used a different dataset with 50 participants, using the same scanning parameters (except a different duration of scan: 600 time points) with the same scanner, to repeat the abovementioned gradient methods and calculate group-level gradients. This was followed by summarizing gradient values into Yeo’s 17-network and then comparing the spatial correlation (i.e., Spearman correlation) between each pair of gradient components in these two datasets.
3. Results
3.1. Demographic and behavioural data
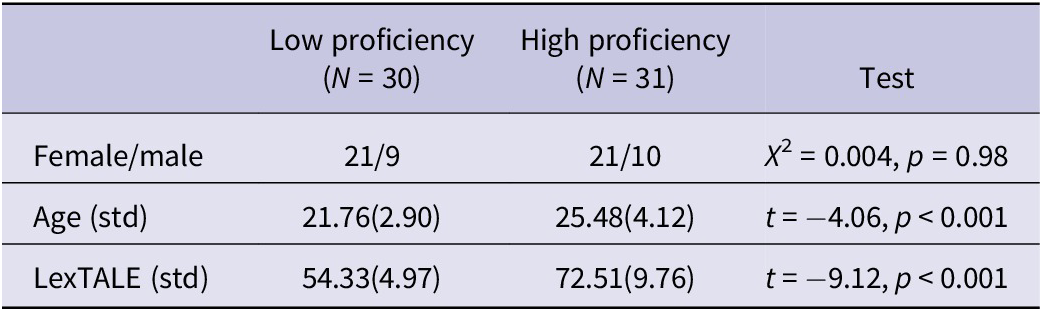
As shown in Table 1, 61 participants were included in further analysis after 11 participants were removed due to the missing of behavioural data or excessive head motion during the MRI scan. These two groups of participants had comparable gender distribution, but age difference was significant. Further analyses included gender and age as covariates to remove the confounding effects brought by these demographic data. The high-proficiency group (72.51 ± 9.76, N = 31) showed significantly higher LexTALE scores than the low-proficiency group (54.33 ± 4.97, N = 30).
Demographic information and proficiency scores of the included sample

3.2. Predictive power for L2 proficiency from network gradients
The first three network gradients revealed in our study (Supplementary Figure S1) were similar to previous findings. The first gradient separates sensory (unimodal) and transmodal brain regions, the second gradient separates somatosensory regions from visual and other regions, and the third gradient places frontoparietal and attentional networks at one end and the default mode network and other regions at the other end. Linear regression analysis did not reveal any cluster showing a significant correlation with L2 proficiency scores in the first three network gradients.
As shown in Supplementary Figure S2A, the explained variances between neighbouring gradients gradually became indistinguishable after the first 10 gradients, suggesting that these gradients contributed most of the variances to the time series of brain networks. As reported in the supplementary analysis (Section 2.10), eight of the ten gradients, with the exception of the eighth and ninth gradients, demonstrated high test–retest reliability in an independent dataset (Bonferroni correction, p < 0.05). Thus, we further conducted whole-brain linear regression analysis in the fourth–tenth gradients.
The fifth gradient revealed the semantic network in low gradient values (Figure 2A, 2B & 2C). More specifically, it showed low gradient values in default mode (DMN) and attention (AN) networks, while high gradient values in somatosensory (SMN), occipital (OCC) and frontoparietal (FPN) networks (Figure 2D). Regarding the seventh gradient (Figure 2G, 2H, & 2L), it depicted the closely aligned gradients among OCC, DAN, FPN, and DMN, showing the executive network in low gradient values (Figure 2J). Further whole-brain explorative analysis revealed that the gradient values of the right calcarine gyrus in the fifth gradient were positively correlated with L2 proficiency (Figure 2E & 2F, cluster-level FWE p < 0.05, cluster-forming threshold p < 0.001), while the gradient values of the left SMG in the seventh gradient were also positively correlated with L2 proficiency (Figure 2K & 2L, cluster-level FWE p < 0.05, cluster-forming threshold p < 0.001).
Gradient networks and L2 proficiency. (A/G) Average network gradient 5/7. (B/H) Average network gradient in the low-proficiency group in gradient 5/7. (C/I) Average network gradient in the high-proficiency group in gradient 5/7. (D/J) Radar plot showing the average gradient value in Yeo’s 7-network. (E) The right calcarine area, of which gradient 5 revealed a significant correlation with L2 proficiency (cluster-level FWE p < 0.05, cluster-forming threshold p < 0.001). (F) Scatter and line plot depicting the relationship between averaged gradient 5 values in the cluster of the right calcarine area and L2 proficiency. (K) The left supramarginal area, of which gradient 7 revealed a significant correlation with L2 proficiency (cluster-level FWE p < 0.05, cluster-forming threshold p < 0.001). (L) Scatter and line plot depicting the relationship between averaged gradient 7 values in the cluster of left supramarginal area and L2 proficiency.

Common and differential effects of directed connections between low- and high-L2-proficiency groups in language and cognitive control networks. (A) Directed connections showing common effects across groups (connections in blue indicate the inhibitory effect, while connections in red indicate the excitatory effect). (B) Matrix plot showing directed connections from common effects across groups (positive values in the main diagonal indicate increased self-inhibition, while negative values in the main diagonal indicate reduced self-inhibition; positive values in the non-main diagonal indicate an excitatory effect, while negative values in the non-main diagonal indicate an inhibitory effect). (C) The common directed interaction pattern between language and cognitive control networks at the network level (grey connections indicate the corresponding Pp was smaller than 95%; the number above the Pp is the expected effective connectivity displayed in Hz; the positive values of effective connectivity indicate excitatory effect, and the negative values of effective connectivity indicate inhibitory effect). (D) Directed connections showing differential effects between groups (connections in blue indicate negative values, while connections in red indicate positive values). (E) The matrix plot showing directed connections from group-different effects. Positive values indicate a stronger excitatory (or weaker inhibitory) effect, and negative values indicate a weaker excitatory (or stronger inhibitory) effect in the high-L2-proficiency group compared with the low-L2-proficiency group, depending on the common effects across groups. (F) The differential directed interaction pattern between language and cognitive control networks between groups at the network level (the grey connections indicate the corresponding Pp was smaller than 95%, and there were no directed connections from any brain regions of the cognitive control network to any brain regions of the language network; the indications of the signs of values above Pp are the same as panel E). Note: Pp: posterior probability; the unit of the expected effective connectivity in panels C and F is Hz; L (R): left (right); calc: calcarine area; SMG: supramarginal gyrus; pMTG: posterior middle temporal gyrus; IFG: inferior frontal gyrus; pMFG: posterior middle frontal gyrus; dACC: dorsal anterior cingulate cortex; MFG: middle frontal gyrus; aMFG: anterior middle frontal gyrus.

MVPA revealed that the gradient values in language and cognitive control networks in the first ten gradients had relatively low predictive power for proficiency levels (Supplementary Figure S2B). Four gradients had above-chance-level accuracy, including the highest predictive power around 62%, which may indicate that the between-subject variances of the global brain activity flow in these two networks were not sufficient to predict high or low proficiency.
3.3. Between-group differences in gradient values in language and cognitive control networks
As shown in Supplementary Figures S3–S12, no significant between-group differences in gradient values in the first 10 gradients survived multiple corrections (false discovery rate – FDR, q < 0.05). Several brain regions, including right calcarine area, left temporal pole and right anterior MFG, had different gradient values in either the fifth or ninth gradient (p < 0.05). These results might suggest that L2 proficiency would not modulate the relative position of language and cognitive control networks in the global brain activity flow.
3.4. Directed interactions among language and cognitive control networks
As shown in Figure 3A & 3B (Pp > 95%), there were various directed interactions within and between language and cognitive control networks, which were common across proficiency groups. For example, there was an excitatory effect from the left IFG to the left pMTG, while there was an inhibitory effect from the left temporal pole to the left MFG. In summary, it revealed many directed interactions across proficiency groups from the right insula to other brain regions in the cognitive control network, from dACC to brain regions in both language and cognitive control networks and from the left temporal pole to brain regions in both language and cognitive control networks. At the network level, it was observed that the language network has overall inhibitory effects on the cognitive control network (Pp > 95%, Figure 3C), while other network-level interactions were not supported by robust evidence (Pp < 95%).
The directed interactions modified by L2 proficiency within and between language and cognitive control networks were observed when comparing high- and low-L2-proficiency groups (Pp > 95%, Figure 3D & 3E). For example, there was an increased excitatory effect from the left IFG to the left pMTG, while there was an increased inhibitory effect (more negative) from the left temporal pole to the right pMFG. There was a reduced excitatory effect from the left IFG to the left SMG, and there was a reduced inhibitory effect from the dACC to the left MFG. In summary, L2 proficiency modulated directed interactions within language and cognitive networks, and from language to cognitive control networks, but no directed interactions from the cognitive control network to the language network were modulated by L2 proficiency. At the network level, it was observed that there were overall increased inhibitory effects (more negative) from the language network to the cognitive control network (Pp > 0.95, Figure 3F). This finding suggests that higher L2 proficiency enhances the inhibitory influence of the language network on the cognitive control network.
4. Discussion
Our study investigated the relationship between L2 proficiency and the brain activity flow in language and cognitive control networks. Whole-brain linear regression analysis revealed that gradient values in the right calcarine and left SMG in the language network were associated with L2 proficiency. However, the subsequent MVPA and ROI analysis indicated that gradient values in language and cognitive control networks had low predictive power to distinguish different proficiency levels. Meanwhile, our study revealed a consistent inhibitory connection from the language network to the cognitive control network. Notably, individuals with high L2 proficiency exhibited stronger inhibitory connections compared to those with low proficiency, thereby deepening our understanding of the local interactive patterns between these two networks.
In our study, the gradient values of the right calcarine gyrus in the fifth gradient and the left SMG in the seventh gradient were significantly associated with L2 proficiency scores. Although the right calcarine gyrus is not a core language region, it supports the visual processing of written language, including orthographic analysis and perceptual aspects of lexical learning, which are particularly relevant for L2 development (Jiang, Reference Jiang2000; Price, Reference Price2012; Wandell et al., Reference Wandell, Dumoulin and Brewer2007). Its interaction with language and attention networks facilitates the efficient acquisition of written forms in an L2. Brain regions with lower gradient values in the fifth gradient were similar to the semantic network revealed by a previous study (Huth et al., Reference Huth, de Heer, Griffiths, Theunissen and Gallant2016), suggesting that the adaptation of a relative position in the right calcarine in the global brain activity flow might also be related to the semantic processes in proficiency improvement.
Meanwhile, the left SMG is a key area of the inferior parietal lobe (IPL) that contributes to various cognitive functions, especially related to language comprehension, auditory processing and sensory integration. The left IPL, including left SMG and angular gyrus, has been widely recognized as related to L2 learning across differing developmental stages and bilingual types (Barbeau et al., Reference Barbeau, Chai, Chen, Soles, Berken, Baum, Watkins and Klein2017; Li et al., Reference Li, Zhang and Ding2021; Zhang et al., Reference Zhang, Li, Zhang, Wang, Ao, Jian, Wei, Liu, Ding and Meng2023). A recent meta-analysis by Li et al. (Reference Li, Zhang and Ding2021) revealed that enhanced activations in the left IPL were detected in L2, relative to L1, among both L1-L2-distant bilinguals (e.g., Chinese-English) and L1-L2-similar bilinguals (e.g., English-French). Zhang et al. (Reference Zhang, Li, Zhang, Wang, Ao, Jian, Wei, Liu, Ding and Meng2023) further unveiled that the functional preference of the left IPL in L2 learning might manifest the special demands of phonological processing. In addition, the left angular gyrus is likely to play a role in hosting both phonological and semantic representations, acting as a mediator between the non-lexical reading network and the semantic system (Junker et al., Reference Junker, Schlaffke, Lange and Schmidt-Wilcke2024). The seventh gradient’s emphasis on executive network regions with lower gradient values, along with prior evidence linking these areas to language comprehension and cross-component integration, suggests that the left SMG observed in our study may play a role in semantic switching between languages. While recognizing the role of left SMG underlying L2 comprehension, our results also highlighted the association between left SMG and overall L2 proficiency, independent of specific tasks.
Nevertheless, our results showed that the gradient values of the brain regions of language and cognitive control networks of the first ten gradients had limited predictive power over the classification of low versus high L2 proficiency. Meanwhile, ROI analysis did not reveal significant between-group differences in gradient values in the first ten gradients. Previous studies, focusing on specific language processes including lexical representation and discourse processing (Eisenhauer et al., Reference Eisenhauer, Gonzalez Alam, Cornelissen, Smallwood and Jefferies2024; Wu et al., Reference Wu, Morales, Patel, Pickering and Hoffman2022) by utilizing task-based fMRI, indicated that network gradients can support language processing. The current study instead investigated whether L2 proficiency influences the global brain activity flow in language and cognitive control networks in network gradients during the resting state. Our results provided evidence supporting that the between-subject variances of the global brain activity flow in these two networks are not sufficient to predict proficiency levels, even though network gradients play an important role in general language processing.
Furthermore, our study depicts directed interactions among brain regions within and between language and cognitive control networks, revealing the local brain activity flow among these two brain networks. Many studies have examined the close relationship between L2 acquisition and cognitive control ability (Bialystok, Reference Bialystok2009; Bialystok, Reference Bialystok2021; Li et al., Reference Li, Legault and Litcofsky2014; Tao et al., Reference Tao, Wang, Zhu and Cai2021). Meanwhile, some studies revealed the functional connectivity among brain regions in these two brain networks modulated by L2 development (Chai et al., Reference Chai, Berken, Barbeau, Soles, Callahan, Chen and Klein2016; Ghazi Saidi et al., Reference Ghazi Saidi, Perlbarg, Marrelec, Pelegrini-Issac, Benali and Ansaldo2013; Sun et al., Reference Sun, Li, Ding, Wang and Li2019). However, these findings on functional connectivity were not consistent. In our study, it was observed that individuals with high L2 proficiency manifested stronger inhibitory connections from language to cognitive control networks compared with individuals with low L2 proficiency. Such findings might indicate that the development of an L2 can have causal effects on the cognitive control system. L2 learning and the resultant bilingualism have been found to promote cognitive control, including switching and inhibition, which was supported by both behavioural and neural investigations (Bialystok, Reference Bialystok2009; Bialystok, Reference Bialystok2021; Li et al., Reference Li, Legault and Litcofsky2014; Tao et al., Reference Tao, Wang, Zhu and Cai2021). Gold et al. (Reference Gold, Kim, Johnson, Kryscio and Smith2013) demonstrated that brain activation levels in cognitive control regions were inversely related to task performance, with lower activation corresponding to better performance. Sun et al. (Reference Sun, Li, Ding, Wang and Li2019) further examined the modulation of L2 proficiency on the subcomponents of cognitive control networks, including inhibition, cognitive flexibility (switching) and working memory. Our study provided further evidence to support that the acquisition of L2 proficiency modulates the cognitive control system. These results are in line with the Adaptive Control Hypothesis (Green & Abutalebi, Reference Green and Abutalebi2013), which argues that bilingual experience adaptively shapes cognitive control processes depending on interactional demands. The stronger inhibitory influences observed in high-proficiency learners may thus reflect the adaptive tuning of the control network to support efficient bilingual language use.
Our study also has some limitations. First, we did not include behavioural measurements regarding the cognitive control function. Thus, we cannot directly associate the changes in the local brain activity flow between language and cognitive control networks with the improvement in cognitive control function while comparing L2 proficiency groups. Our study can only demonstrate that L2 proficiency modulates the interaction from the language network to the cognitive control network. Future studies can examine whether the changes in the interaction between language and cognitive control networks are associated with the improvement in cognitive control function. Second, we relied solely on resting-state fMRI and did not include specific tasks that reflect semantic, phonological or orthographic processing. As a result, we were unable to explore the characteristics of the brain activity flow of these specific processing mechanisms and language components. Future research could incorporate these language tasks, along with tasks related to cognitive control (e.g., inhibition, switching, working memory), to provide a more comprehensive depiction of the full process of L2 development.
5. Conclusion
To our knowledge, our study is among the first fMRI studies investigating global (gradient network) and local (directed connectivity) brain activity flows underlying L2 proficiency by combining gradient analysis and DCM. The gradient values in the right calcarine gyrus in the fifth gradient and the left SMG in the seventh gradient were positively associated with L2 proficiency, potentially indicating their roles in visual-orthographic and phonological/semantic processing during L2 learning. However, MVPA and ROI analysis indicated that L2 proficiency may not significantly modulate the relative position of brain regions in language and cognitive control networks in the global brain activity flow. Additionally, DCM results showed an overall increase in inhibitory effects (more negative) from the language network to the cognitive control network in high-proficiency L2 users compared to low-proficiency L2 users. This finding suggests that higher L2 proficiency may enhance these inhibitory interactions, which could underlie the improved cognitive control performance accompanied with L2 learning. Our findings contribute to a deeper understanding of the brain’s dynamic and interactive profile between language and cognitive control networks supporting the acquisition of L2 proficiency, which may be the critical neural signatures of how language and cognitive control networks interact to form an efficient bilingual system.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S1366728926101163.
Data availability statement
The datasets generated during and/or analysed in the current study are available from the corresponding author on reasonable request.
Funding statement
This work was supported by the National Social Science Foundation of China (24CYY088), the University of Macau (MYRG2022-00054-FHS, MYRG-GRG2023-00038-FHS-UMDF, and MYRG-GRG2024-00259-FHS), and the Macao Science and Technology Development Fund (FDCT 0014/2024/RIB1). In addition, this work was performed in part at SICC, which is supported by SKL-IOTSC, University of Macau.
Competing interests
The authors declare that they have no competing interests.
Ethics statement
This study was approved by the Ethics Committee at the University of Macau. Informed consent was obtained from all participants.






 Open access
Open access