


Introduction
Various types of coelomocytes in echinoderms have been described and are regarded as multi-functional cells, mostly serving as part of an arsenal of immune cells. The identification of these coelomocytes has conventionally been based on morphology as viewed through microscopy (Chia and Xing, Reference Chia and Xing1996; Queiroz et al., Reference Queiroz, Mauro, Arizza, Custódio and Vazzana2021; Queiroz and Custódio, Reference Queiroz, Custódio, Mercier, Hamel, Suhrbier and Pearce2024). Among the different classes in the phylum, there is a relatively high diversity of coelomocytes in holothuroids with at least nine recognised types: phagocytes (also known as leukocytes), spherule cells (also known as spherulocytes, amoebocytes, morula cells, or granulocytes), fusiform cells, hemocytes, giant cells, progenitor cells, minute corpuscles (fragments of cells), lymphocytes (small spherical cells), and crystal cells (crystalline cells or crystal corpuscles) (Endean, Reference Endean and Boolootian1966; Smith, Reference Smith, Ratcliffe and Rowley1981; Chia and Xing, Reference Chia and Xing1996; Smith et al., Reference Smith, Arizza, Barela Hudgell, Barone, Bodnar, Buckley, Cunsolo, Dheilly, Franchi, Fugmann, Furukawa, Garcia-Arraras, Henson, Hibino, Irons, Li, Lun, Majeske, Oren, Pagliara, Pinsino, Raftos, Rast, Samasa, Schillaci, Schrankel, Stabili, Stensväg, Sutton and Cooper2018; Queiroz et al., Reference Queiroz, Mauro, Arizza, Custódio and Vazzana2021; Queiroz and Custódio, Reference Queiroz, Custódio, Mercier, Hamel, Suhrbier and Pearce2024). Previously, vibratile cells were considered a type of coelomocytes in holothuroids until a recent analysis demonstrated that they are in fact male gametes at different stages of development (Caulier et al., Reference Caulier, Jobson, Wambreuse, Borrello, Delroisse, Eeckhaut, Mercier, Hamel, Mercier, Hamel, Suhrbier and Pearce2024).
Among the coelomocytes, crystal cells have mostly been described in holothuroids (Smith, Reference Smith, Ratcliffe and Rowley1981; Smith et al., Reference Smith, Arizza, Barela Hudgell, Barone, Bodnar, Buckley, Cunsolo, Dheilly, Franchi, Fugmann, Furukawa, Garcia-Arraras, Henson, Hibino, Irons, Li, Lun, Majeske, Oren, Pagliara, Pinsino, Raftos, Rast, Samasa, Schillaci, Schrankel, Stabili, Stensväg, Sutton and Cooper2018) and have also been mentioned in echinoids and asteroids (Johnson and Beeson, Reference Johnson and Beeson1966; Smith, Reference Smith, Ratcliffe and Rowley1981; Chia and Xing, Reference Chia and Xing1996; Queiroz et al., Reference Queiroz, Arizza, Vazzana and Custódio2022a). They were observed in 40% of ∼80 studied species of holothuroids, including in up to 49% of the Dendrochirotida investigated (Queiroz and Custódio, Reference Queiroz, Custódio, Mercier, Hamel, Suhrbier and Pearce2024). In this context, crystal cells are thought to be highly differentiated coelomocytes (Zavalnaya et al., Reference Zavalnaya, Shamshurina and Eliseikina2020). In most echinoderm taxa, crystal cells typically comprise between 0.5 and 5% of total coelomocyte cell counts (Endean, Reference Endean1958; Smith, Reference Smith, Ratcliffe and Rowley1981; Chia and Xing, Reference Chia and Xing1996; Smith et al., Reference Smith, Arizza, Barela Hudgell, Barone, Bodnar, Buckley, Cunsolo, Dheilly, Franchi, Fugmann, Furukawa, Garcia-Arraras, Henson, Hibino, Irons, Li, Lun, Majeske, Oren, Pagliara, Pinsino, Raftos, Rast, Samasa, Schillaci, Schrankel, Stabili, Stensväg, Sutton and Cooper2018; Queiroz et al., Reference Queiroz, Mauro, Arizza, Custódio and Vazzana2021, Reference Queiroz, Arizza, Vazzana and Custódio2022a, b; Jobson et al., Reference Jobson, Hamel and Mercier2022; Xue et al., Reference Xue, Peng and Wang2024). In some instances, the proportion of crystal cells can be as high as ∼10% of all coelomocytes (Caulier et al., Reference Caulier, Hamel and Mercier2020). Although not always present in a given individual, their densities in the coelomic fluid of holothuroids average around 600 cells mL−1 (Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002) but can reach upwards of 13,500 cells mL−1 (Jobson et al., Reference Jobson, Hamel, Hughes and Mercier2021) and 143,000 cell mL−1 (Li et al., Reference Li, Ren, Liang, Qiao, Wang, Ye and Li2018). Crystal cells are usually ascribed a crescent-shaped, heterochromatic nucleus and an intracellular vacuole containing the crystalline structure (Xing et al., Reference Xing, Yang and Chen2008; Smith et al., Reference Smith, Arizza, Barela Hudgell, Barone, Bodnar, Buckley, Cunsolo, Dheilly, Franchi, Fugmann, Furukawa, Garcia-Arraras, Henson, Hibino, Irons, Li, Lun, Majeske, Oren, Pagliara, Pinsino, Raftos, Rast, Samasa, Schillaci, Schrankel, Stabili, Stensväg, Sutton and Cooper2018; Caulier et al., Reference Caulier, Hamel and Mercier2020; Jobson et al., Reference Jobson, Hamel and Mercier2022; Queiroz and Custódio, Reference Queiroz, Custódio, Mercier, Hamel, Suhrbier and Pearce2024; Xue et al., Reference Xue, Peng and Wang2024). However, the nucleus is often indistinguishable, which suggests that it is either absent or, if present, challenging to identify (Hetzel, Reference Hetzel1963; Chia and Xing, Reference Chia and Xing1996; Chia and Xing, Reference Chia and Xing1996; Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002; Caulier et al., Reference Caulier, Hamel and Mercier2020).
While the function of crystal cells in echinoderms was generally considered unknown (Ratcliffe and Rowley, Reference Ratcliffe and Rowley1979; Smith, Reference Smith, Ratcliffe and Rowley1981; Karp and Coffaro, Reference Karp, Coffaro, Cohen and Sigel1982; Smiley, Reference Smiley, Harrison and Chia1994; Smith et al., Reference Smith, Arizza, Barela Hudgell, Barone, Bodnar, Buckley, Cunsolo, Dheilly, Franchi, Fugmann, Furukawa, Garcia-Arraras, Henson, Hibino, Irons, Li, Lun, Majeske, Oren, Pagliara, Pinsino, Raftos, Rast, Samasa, Schillaci, Schrankel, Stabili, Stensväg, Sutton and Cooper2018), some authors have proposed they relate to mesenchymal ossicle-forming cells (Hetzel, Reference Hetzel1963) or play a role in osmoregulation (Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002; Xing et al., Reference Xing, Yang and Chen2008; Ramírez-Gómez and García-Arrarás, Reference Ramírez-Gómez and García-Arrarás2010). Hetzel (Reference Hetzel1963) noted that a slight change in osmotic pressures can cause the disintegration of the intracellular crystalline structure in crystal cells (Hetzel, Reference Hetzel1963; Endean, Reference Endean and Boolootian1966; Chia and Xing, Reference Chia and Xing1996). Upon dissolution of the crystalline structure, the remaining cell collapses (Hetzel, Reference Hetzel1963). Eliseikina and Magarlamov (Reference Eliseikina and Magarlamov2002) observed that an increase or normalisation in osmotic pressure could result in the transformation of crystal cells into structurally similar cells (i.e., cells with a centrally located spherical vacuole), suggesting that the crystallisation process inside the cell was reversible (also see Xing et al., Reference Xing, Yang and Chen2008). However, any role with osmoregulation has been recently challenged when the density of crystal cells did not change under experimental exposure to low salinities (Jobson et al., Reference Jobson, Hamel, Hughes and Mercier2021). The solubility of the crystal-like structure under a slight change in osmotic pressure and its refractive properties have led several authors to conclude that the structure is crystalline in nature (Endean, Reference Endean and Boolootian1966; Karp and Coffaro, Reference Karp, Coffaro, Cohen and Sigel1982; Chia and Xing, Reference Chia and Xing1996) and, more recently, that its chemical identity is that of uric acid (KCK et al., Reference Ma, Gao, Hamel and Mercier2025). Briefly, crystals of uric acid in holothuroids were identified based on their morphology (cuboidal–barrel-, rhomb-, and rosette-shaped crystals) and size ranges, solubility in alkaline solution or upon heating but not in alcohol, and diagnostic Raman peaks at 839, 995, 1036, 1158, and 1649 cm−1 (KCK et al., Reference Ma, Gao, Hamel and Mercier2025).
In line with emerging questions surrounding some coelomocyte types (Karp and Coffaro, Reference Karp, Coffaro, Cohen and Sigel1982; Taguchi et al., Reference Taguchi, Tsutsui and Nakamura2016; Caulier et al., Reference Caulier, Jobson, Wambreuse, Borrello, Delroisse, Eeckhaut, Mercier, Hamel, Mercier, Hamel, Suhrbier and Pearce2024; Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025), the present review revisits the identity of crystal cells as a distinct type of coelomocyte. First, it summarises available information on the so-called crystal cells among coelomocytes of holothuroids and other echinoderms (e.g., asteroids, echinoids) and, second, reassesses this notion on the basis of a recent discovery and analysis of microscopic crystalline structures (i.e., microcrystals) in holothuroids (KCK et al., Reference Ma, Gao, Hamel and Mercier2025).
Review of crystal-cell coelomocytes in echinoderms
Morphology of crystal cells
Rhomb-shaped crystal cells
Crystal cells with a single rhomb-shaped crystalline structure have been documented in asteroids and holothuroids (Table 1). The shape of this crystal has been described as rhomboid(al), rhomb(ic), lozenge-shaped, or diamond-shaped (Table 1). In particular, cells have been described with two distinct rhomboidal crystals (Endean, Reference Endean1958), three distinct crystals (Endean, Reference Endean and Boolootian1966; Smith, Reference Smith, Ratcliffe and Rowley1981; Smiley, Reference Smiley, Harrison and Chia1994), and an arrangement of three individual crystals overlayed at offsetted angles that can give an appearance of a six-pointed star-shaped morphology (Hetzel, Reference Hetzel1963).
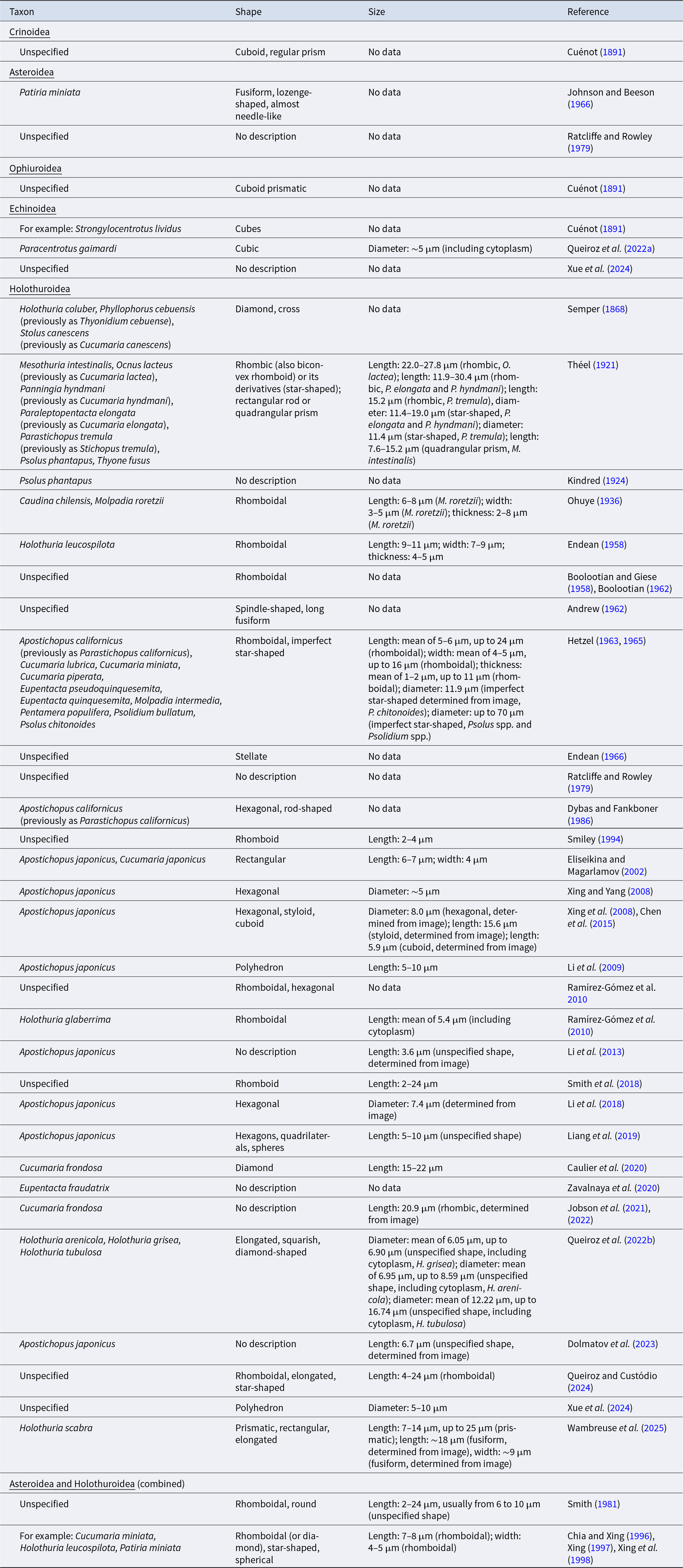
Summary of the diversity of morphological descriptions of crystal cells identified as a type of coelomocyte in echinoderms, organised by class and then by year of publication

The sizes of these crystal cells in holothuroids vary, with studies reporting lengths of 2–4 μm (Smiley, Reference Smiley, Harrison and Chia1994) and larger sizes of 7–8 μm in length and 4–5 μm in width (Chia and Xing, Reference Chia and Xing1996; Xing, Reference Xing1997; Xing et al., Reference Xing, Leung and Chia1998) as well as 5–6 μm in length (maximum of 24 μm), 4–5 μm in width (maximum of 16 μm), and 1–2 μm in thickness (maximum of 11 μm) (Hetzel, Reference Hetzel1963, Reference Hetzel1965) (Table 2). Species-specific measurements include lengths of 12–30 μm in Paraleptopentacta elongata and Panningia hyndmani, 15 μm in Parastichopus tremula, and 22–28 μm in Ocnus lactea (Théel, Reference Théel1921; Table 2). In Molpadia roretzii, they measured 6–8 μm in length, 3–5 μm in width, and 2–8 μm in thickness (Ohuye, Reference Ohuye1936; Table 2). In Holothuria glaberrima, they had an average length of 5 μm (Ramírez-Gómez et al., Reference Ramírez-Gómez, Aponte-Rivera, Méndez-Castaner and García-Arrarás2010; Table 2). In Holothuria leucospilota, they measured 9–11 μm long, 7–9 μm wide, and 4–5 μm thick (Endean, Reference Endean1958; Table 2). In Cucumaria frondosa, they ranged between 15 and 22 μm in length (Caulier et al., Reference Caulier, Hamel and Mercier2020; also see illustrations by Jobson et al., Reference Jobson, Hamel, Hughes and Mercier2021, Reference Jobson, Hamel and Mercier2022; Table 2). Length measurements of crystal cells in holothuroids, likely to closely represent the length of the crystal, may or may not include the cytoplasm and/or nucleus described in some publications; reports have ranged from 2 to 24 or 4 to 24 μm (Smith et al., Reference Smith, Arizza, Barela Hudgell, Barone, Bodnar, Buckley, Cunsolo, Dheilly, Franchi, Fugmann, Furukawa, Garcia-Arraras, Henson, Hibino, Irons, Li, Lun, Majeske, Oren, Pagliara, Pinsino, Raftos, Rast, Samasa, Schillaci, Schrankel, Stabili, Stensväg, Sutton and Cooper2018; Queiroz and Custódio, Reference Queiroz, Custódio, Mercier, Hamel, Suhrbier and Pearce2024; Tables 1 and 2). The size of rhomb-shaped crystal cells in asteroids has not been reported.
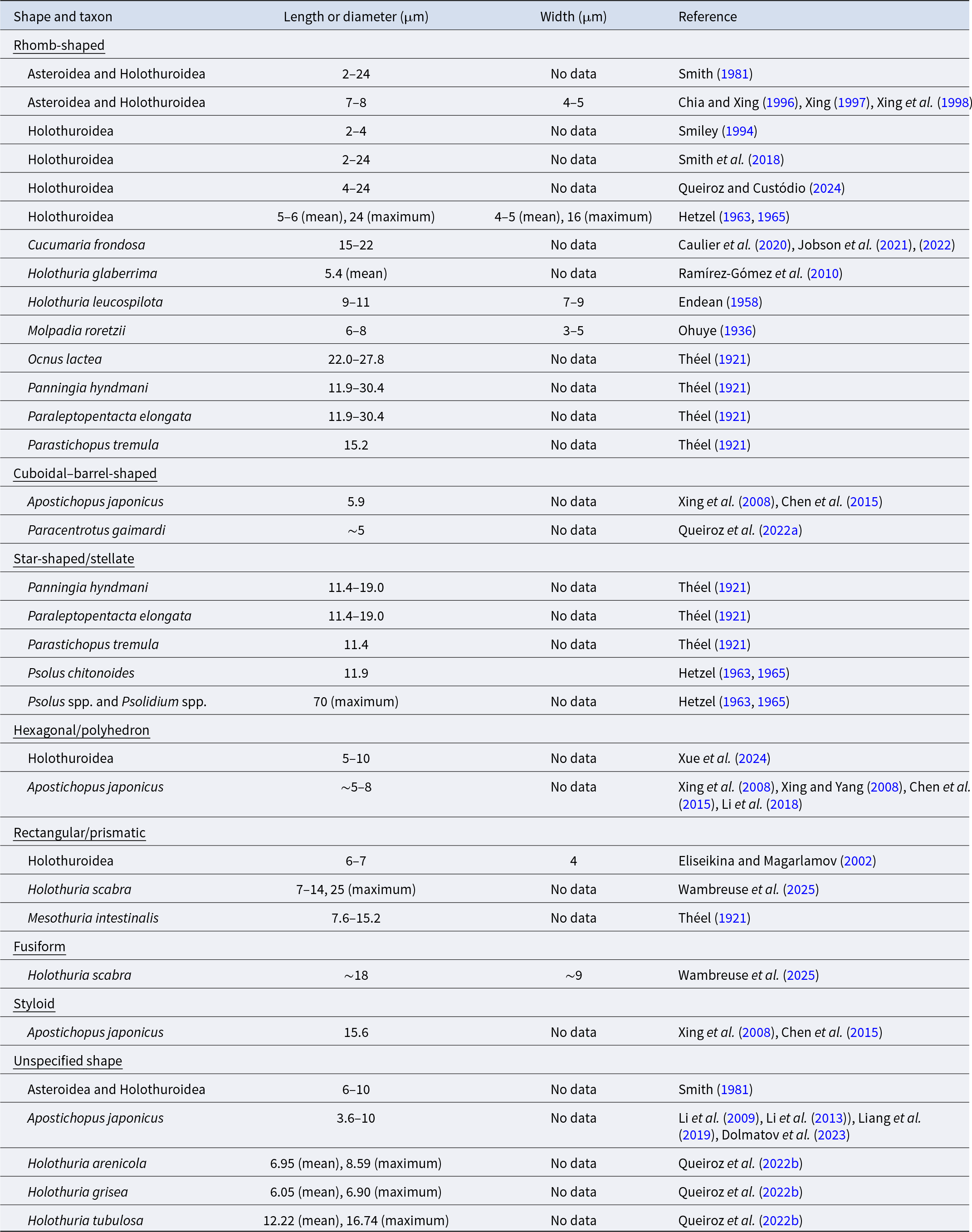
Size ranges of crystal cells identified as a type of coelomocyte in echinoderms, re-configured from the morphological summary presented in Table 1 and grouped by crystal shape

Cuboidal crystal cells
Crystal cells with cuboidal crystalline structures have been reported in crinoids, echinoids, ophiuroids, and holothuroids (Cuénot, Reference Cuénot1891; Xing et al., Reference Xing, Yang and Chen2008; Chen et al., Reference Chen, Wang, Xing, Zhu and Zhang2015; Queiroz et al., Reference Queiroz, Arizza, Vazzana and Custódio2022a). Among holothuroids, these crystals are less frequently observed compared to rhomb-shaped ones (described above). In the literature, this morphology has been described variously as cuboid(al), cubic, or cube-shaped (Table 1). In one instance, the shape was referred to as cuboid prismatic (Cuénot, Reference Cuénot1891), but it is unclear if other mentions of prisms/prismatic forms also refer to the cuboidal shape. Notably, the surfaces of some cuboidal crystals were slightly curved (Cuénot, Reference Cuénot1891). In the echinoid Paracentrotus gaimardi, live cells with this type of crystalline structure, including the surrounding cytoplasm, measured approximately 5 μm in diameter (Queiroz et al., Reference Queiroz, Arizza, Vazzana and Custódio2022a; Table 2). The sizes of those observed in echinoids by Cuénot (Reference Cuénot1891) were only described as variable. Based on a published image, the cuboidal crystal in the holothuroid Apostichopus japonicus was ∼6 μm in length (Xing et al., Reference Xing, Yang and Chen2008; Chen et al., Reference Chen, Wang, Xing, Zhu and Zhang2015; Table 2).
Other morphologies of crystal cells
Star-shaped/stellate, rosette-shaped, cross-shaped, hexagonal (also referred to as polyhedrons), rod-shaped/elongated/styloid/needle-like, rectangular, squarish, and spherical/round crystalline structures have been reported in asteroids and holothuroids (Table 1). Regular prisms and prismatic shapes in crinoids, echinoids, and holothuroids are too generic to distinguish them from or classify them with the other named morphologies (Cuénot, Reference Cuénot1891; Xue et al., Reference Xue, Peng and Wang2024; Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025). Théel (Reference Théel1921) remarked that rectangular crystals may be the side view of rhomb-shaped crystals and that cubic crystals may be the side view of star-shaped crystals. There are also reservations of the actual morphology of quadrilateral crystals in Apostichopus japonicus, as they may be either rhombic or rectangular crystals (Liang et al., Reference Liang, Ren, Wang, Li, Yang and Li2019). There are no instances of multiple crystals within a cell reported for any of these shapes (Table 1).
Star-shaped/stellate crystalline structures are generally not as common compared to rhomb-shaped ones (Endean, Reference Endean and Boolootian1966). They have been reported in the holothuroid genera Psolus and Psolidium with diameters up to 70 μm (Hetzel, Reference Hetzel1963, Reference Hetzel1965; Endean, Reference Endean and Boolootian1966; Table 2). In Psolus chitonoides, this crystalline structure, based on an illustration, measures 12 μm in diameter (Hetzel, Reference Hetzel1963, Reference Hetzel1965; Table 2). Additionally, this crystal shape is known in other holothuroid taxa – namely, Panningia hyndmani, Paraleptopentacta elongata, and Parastichopus tremula – with diameters ranging from 11 to 19 μm (Théel, Reference Théel1921; Table 2). The cross-shaped crystal isolated from a holothuroid and described by Semper (Reference Semper1868) may be a less elaborate form of the star-shaped crystal.
Hexagonal/polyhedron crystals are well documented in a few holothuroids in the genus Apostichopus (Dybas and Fankboner, Reference Dybas and Fankboner1986; Xing and Yang, Reference Xing and Yang2008; Xing et al., Reference Xing, Yang and Chen2008; Chen et al., Reference Chen, Wang, Xing, Zhu and Zhang2015; Li et al., Reference Li, Chen, Li and Park2009, Reference Li, Ren, Liang, Qiao, Wang, Ye and Li2018; Liang et al., Reference Liang, Ren, Wang, Li, Yang and Li2019; Xue et al., Reference Xue, Peng and Wang2024). Notably, polyhedrons are hexagonal crystals when viewed from the top and rectangular from the side (Li et al., Reference Li, Chen, Li and Park2009; Xue et al., Reference Xue, Peng and Wang2024). In A. japonicus, this is the more commonly encountered crystal shape (Xing et al., Reference Xing, Yang and Chen2008; Chen et al., Reference Chen, Wang, Xing, Zhu and Zhang2015; Li et al., Reference Li, Ren, Liang, Qiao, Wang, Ye and Li2018). The diameter of these cells/crystals measure between ∼5 and 10 μm (Xing et al., Reference Xing, Yang and Chen2008; Xing and Yang, Reference Xing and Yang2008; Li et al., Reference Li, Chen, Li and Park2009, Reference Li, Ren, Liang, Qiao, Wang, Ye and Li2018; Chen et al., Reference Chen, Wang, Xing, Zhu and Zhang2015; Xue et al., Reference Xue, Peng and Wang2024; Table 2).
Biconvex rhomboid/fusiform/spindle-shaped crystals are known in asteroids (e.g., Patiria minata) and holothuroids (Théel, Reference Théel1921; Andrew, Reference Andrew1962; Johnson and Beeson, Reference Johnson and Beeson1966). An image taken by Wambreuse et al. (Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025) may be depicting the fusiform morphology that was referenced by Andrew (Reference Andrew1962). This morphology of crystalline structure, isolated from Holothuria scabra, measures ∼18 μm long and ∼9 μm wide (Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025; Table 2).
Crystal cells with rod-shaped/elongated/styloid/needle-like crystals have been noted in asteroids (e.g., P. minata) and holothuroids (e.g., the genera Apostichopus and Holothuria) (Dybas and Fankboner, Reference Dybas and Fankboner1986; Johnson and Beeson, Reference Johnson and Beeson1966; Xing et al., Reference Xing, Yang and Chen2008; Chen et al., Reference Chen, Wang, Xing, Zhu and Zhang2015; Queiroz et al., Reference Queiroz, Mauro, Arizza, Custódio and Vazzana2022b; Queiroz and Custódio, Reference Queiroz, Custódio, Mercier, Hamel, Suhrbier and Pearce2024; Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025). In A. japonicus, crystal cells with this shape are 16 μm in length (Xing et al., Reference Xing, Yang and Chen2008; Chen et al., Reference Chen, Wang, Xing, Zhu and Zhang2015; Table 2). Rectangular crystalline structures have been observed in the holothuroids A. japonicus and H. scabra (Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002; Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025). Eliseikina and Magarlamov (Reference Eliseikina and Magarlamov2002) reported crystal cells with this form as having lengths of 6–7 μm and widths of 4 μm in holothuroids (e.g., A. japonicus, Cucumaria japonicus; Table 2). Possibly related to either the rod-shaped or the rectangular morphology, the rectangular rod/quadrangular prism crystal isolated from Mesothuria intestinalis measured 8–15 μm long (Théel, Reference Théel1921; Table 2). Furthermore, squarish crystals are found in the genus Holothuria (Queiroz et al., Reference Queiroz, Mauro, Arizza, Custódio and Vazzana2022b). This morphology could be a variant of the rectangular crystal or another term referring to the cuboidal shape (e.g., side view of the crystal).
Spherical/round crystalline structures inside crystal cells are among the least known morphologies. They are found in asteroids and holothuroids (Smith, Reference Smith, Ratcliffe and Rowley1981; Chia and Xing, Reference Chia and Xing1996; Xing, Reference Xing1997; Xing et al., Reference Xing, Leung and Chia1998; Liang et al., Reference Liang, Ren, Wang, Li, Yang and Li2019). To our knowledge, there are no images or information on their size (Table 1).
The sizes of unspecified morphologies of crystalline structures (perhaps several shapes pooled together) have been documented. Smith (Reference Smith, Ratcliffe and Rowley1981) reported the lengths of crystal cells to be 2–24 μm with typical sizes between 6 and 10 μm in length (Table 2). This was probably referring to the rhomb-shaped crystals because a later study by a different investigator reported the same range for this morphology (Smith et al., Reference Smith, Arizza, Barela Hudgell, Barone, Bodnar, Buckley, Cunsolo, Dheilly, Franchi, Fugmann, Furukawa, Garcia-Arraras, Henson, Hibino, Irons, Li, Lun, Majeske, Oren, Pagliara, Pinsino, Raftos, Rast, Samasa, Schillaci, Schrankel, Stabili, Stensväg, Sutton and Cooper2018). In holothuroids, crystal cells with crystals of unknown morphology measure 6 μm in diameter on average (up to 7 μm) in Holothuria grisea, 7 μm in diameter on average (up to 9 μm) in Holothuria arenicola, and 12 μm in diameter on average (up to 17 μm) in Holothuria tubulosa (Queiroz et al., Reference Queiroz, Mauro, Arizza, Custódio and Vazzana2022b; Table 2). In H. scabra, prismatic crystals with lengths 7–14 μm (up to 25 μm) are reported (Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025; Table 2). Liang et al. (Reference Liang, Ren, Wang, Li, Yang and Li2019) reported crystal cells of A. japonicus to be 5–10 μm in length, which was most likely referring to hexagonal crystals (Table 2). Moreover, images of undetermined morphologies of crystalline structures, isolated from A. japonicus, ranged from 4 to 7 μm in length (Li et al., Reference Li, Ri, Wang, Ye, Qiao and Li2013; Dolmatov et al., Reference Dolmatov, Ginanova, Eliseikina and Frolova2023; Table 2).
Cellular components of crystal cells
Cytoplasm and cell membrane of crystal cells
The hyaline cytoplasm and cell membrane surrounding the crystalline structure have been reported by several authors (Semper, Reference Semper1868; Théel, Reference Théel1921; Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002; Xing et al., Reference Xing, Yang and Chen2008; Queiroz et al., Reference Queiroz, Mauro, Arizza, Custódio and Vazzana2022b). Hetzel (Reference Hetzel1963, Reference Hetzel1965) further described the cytoplasm as a thin layer around the crystal and only visible when viewed from a ‘favourable angle’. Moreover, the cytoplasm of cells on a prepared microscope slide may have a tendency to spread a little (Hetzel, Reference Hetzel1963; Xing et al., Reference Xing, Yang and Chen2008; Queiroz et al., Reference Queiroz, Arizza, Vazzana and Custódio2022a; Queiroz and Custódio, Reference Queiroz, Custódio, Mercier, Hamel, Suhrbier and Pearce2024). Crystal cells with pseudopodia were noted by Hetzel (Reference Hetzel1963, Reference Hetzel1965).
Nucleus of crystal cells
Although a single nucleus enclosed within the crystal cell was first observed by Semper (Reference Semper1868), later investigators remarked that it is not always visible, describing it as altogether absent, often absent, or sometimes absent from the cell (Smith, Reference Smith, Ratcliffe and Rowley1981; Chia and Xing, Reference Chia and Xing1996; Xing, Reference Xing1997; Xing et al., Reference Xing, Leung and Chia1998; Queiroz et al., Reference Queiroz, Arizza, Vazzana and Custódio2022a, b). Furthermore, Théel (Reference Théel1921) noted that the nucleus was detectable after fixation and staining but not in freshly prepared samples. Hetzel (Reference Hetzel1963, Reference Hetzel1965) further added that the nucleus was difficult to see even after fixation. When detected, the nucleus is described as small, heterochromatic/hyperchromatic, crescent-shaped, round, and or flattened (Ohuye, Reference Ohuye1936; Hetzel, Reference Hetzel1963, Reference Hetzel1965; Smith, Reference Smith, Ratcliffe and Rowley1981; Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002; Xing et al., Reference Xing, Yang and Chen2008; Li et al., Reference Li, Chen, Li and Park2009; Chen et al., Reference Chen, Wang, Xing, Zhu and Zhang2015; Smith et al., Reference Smith, Arizza, Barela Hudgell, Barone, Bodnar, Buckley, Cunsolo, Dheilly, Franchi, Fugmann, Furukawa, Garcia-Arraras, Henson, Hibino, Irons, Li, Lun, Majeske, Oren, Pagliara, Pinsino, Raftos, Rast, Samasa, Schillaci, Schrankel, Stabili, Stensväg, Sutton and Cooper2018; Caulier et al., Reference Caulier, Hamel and Mercier2020; Dolmatov et al., Reference Dolmatov, Ginanova, Eliseikina and Frolova2023). Several authors have commented that the nucleus was often located on one side of the crystalline structure, i.e., ex-centrical position (Ohuye, Reference Ohuye1936; Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002; Li et al., Reference Li, Chen, Li and Park2009; Liang et al., Reference Liang, Ren, Wang, Li, Yang and Li2019; Dolmatov et al., Reference Dolmatov, Ginanova, Eliseikina and Frolova2023). Moreover, nuclei of crystal cells have been identified on SEM imagery (Xing et al., Reference Xing, Yang and Chen2008; Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025).
Other organelles of crystal cells
Organelles other than the nucleus have only occasionally been documented in crystal-cell coelomocytes (Smith, Reference Smith, Ratcliffe and Rowley1981). For instance, rare small vacuoles between 0.03 and 0.25 μm in diameter, occurring in the cytoplasm and next to the cell membrane, were reported by Eliseikina and Magarlamov (Reference Eliseikina and Magarlamov2002). A rectangular vacuole (presumably containing a crystalline structure of the same shape) was reported to take up most of the space within the cell (Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002; Smith et al., Reference Smith, Arizza, Barela Hudgell, Barone, Bodnar, Buckley, Cunsolo, Dheilly, Franchi, Fugmann, Furukawa, Garcia-Arraras, Henson, Hibino, Irons, Li, Lun, Majeske, Oren, Pagliara, Pinsino, Raftos, Rast, Samasa, Schillaci, Schrankel, Stabili, Stensväg, Sutton and Cooper2018). One to two yellow refractile granules, as large as 4 μm in diameter, were observed in crystal cells studied by Hetzel (Reference Hetzel1963, Reference Hetzel1965).
Colour, staining, and micrography of crystal cells
Colour of crystal cells
The colour appearance of crystalline features in the crystal-cell coelomocytes of holothuroids was described as white by Théel (Reference Théel1921). In the holothuroid Holothuria leucospilota, the ‘central part’ of the cells was reported as green (Chia and Xing, Reference Chia and Xing1996; Xing, Reference Xing1997; Xing et al., Reference Xing, Leung and Chia1998). In the asteroid Patiria minata, crystals were pale, golden, and somewhat opalescent (Johnson and Beeson, Reference Johnson and Beeson1966). Cubic crystals of crinoids were described as refringent and colourless or slightly yellow, those of echinoids (e.g., Strongylocentrotus lividus) were yellow, and those of ophiuroids were refringent yellow (Cuénot, Reference Cuénot1891).
Staining of crystal cells
Unlike the nucleus and other cellular components, crystalline structures are reported to be resistant to vital/cytochemical staining (Théel, Reference Théel1921; Ohuye, Reference Ohuye1936; Hetzel, Reference Hetzel1963, Reference Hetzel1965; Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002; Queiroz et al., Reference Queiroz, Arizza, Vazzana and Custódio2022a, b; Xue et al., Reference Xue, Peng and Wang2024).
Micrography of crystal cells
Although some investigators report difficulties (Queiroz et al., Reference Queiroz, Arizza, Vazzana and Custódio2022a), crystal cells have been successfully imaged using SEM (Xing et al., Reference Xing, Yang and Chen2008; Xing and Yang, Reference Xing and Yang2008; Li et al., Reference Li, Chen, Li and Park2009; Chen et al., Reference Chen, Wang, Xing, Zhu and Zhang2015; Queiroz and Custódio, Reference Queiroz, Custódio, Mercier, Hamel, Suhrbier and Pearce2024; Xue et al., Reference Xue, Peng and Wang2024). Using SEM, crystal cells appear to have notches on their surfaces (Li et al., Reference Li, Chen, Li and Park2009; Xue et al., Reference Xue, Peng and Wang2024). To date, attempts to image crystal cells using TEM procedures have been largely unsuccessful (Xing et al., Reference Xing, Yang and Chen2008; Li et al., Reference Li, Chen, Li and Park2009; Chen et al., Reference Chen, Wang, Xing, Zhu and Zhang2015; Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025), with successful visualisation reported in only two studies (Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002; Dolmatov et al., Reference Dolmatov, Ginanova, Eliseikina and Frolova2023).
Behaviour of crystal cells
Mobility of crystal cells
The lack of mobility of crystal cells has been widely documented (Ohuye, Reference Ohuye1936; Chia and Xing, Reference Chia and Xing1996; Xing, Reference Xing1997; Xing et al., Reference Xing, Leung and Chia1998), though some observations of slight movements have also been reported (Théel, Reference Théel1921).
Osmoregulation by crystal cells
The formation of crystals inside a vacuole in living cells of holothuroids under conditions of increasing osmotic pressure was described by Eliseikina and Magarlamov (Reference Eliseikina and Magarlamov2002). They concluded that this process was reversible, supporting the role of crystal cells in osmoregulation, i.e., normalising the osmotic pressure in the body. Under slight osmotic stress, the crystalline structure and the cell containing it, however, had been observed to dissolve and collapse, respectively (Hetzel, Reference Hetzel1963). Under laboratory conditions, the concentration of crystal cells did not change when the holothuroid Cucumaria frondosa was exposed to low salinities; however, there was an uptick in cell numbers when the holothuroid was exposed to cold air (Jobson et al., Reference Jobson, Hamel, Hughes and Mercier2021). These experimental results downplay the role that these cells play in regulating osmotic pressure (Jobson et al., Reference Jobson, Hamel, Hughes and Mercier2021).
Presence of crystal cells in response to stress
The abundance of crystal cells increased by 100 ± 122% in the Polian vesicle in response to exposure to cold air of 5 °C experienced by the holothuroid Cucumaria frondosa (Jobson et al., Reference Jobson, Hamel, Hughes and Mercier2021). Furthermore, the number of crystal cells increased slightly in sea cucumbers exposed to cold air of 17 °C and low salinity of 22 psu, but not to low salinity of 15 psu (Jobson et al., Reference Jobson, Hamel, Hughes and Mercier2021).
Chemistry of crystal cells
Solubility of crystal cells
Crystal cells were observed to be insoluble in potassium hydroxide, which is a strong base with a pH from 10 to 13; the exposure time, however, is not specified (Semper, Reference Semper1868).
Chemical composition of crystalline structures inside crystal cells
Based on the morphology of crystalline structures, Semper (Reference Semper1868) initially hypothesised that these structures were crystals of uric acid. However, he concluded that their insolubility in potassium hydroxide suggested otherwise. In the more recent literature, the chemical composition of crystalline structures inside crystal cells is reported as being largely unknown (Endean, Reference Endean and Boolootian1966; Smiley, Reference Smiley, Harrison and Chia1994; Queiroz and Custódio, Reference Queiroz, Custódio, Mercier, Hamel, Suhrbier and Pearce2024).
Bodily distribution of crystal cells
Derivation of crystal cells
Théel (Reference Théel1921) and Ohuye (Reference Ohuye1938) posited that crystal cells were derived from lymphocytes, which in turn were derived from coelomic epithelium (also see Kindred, Reference Kindred1924; Hetzel, Reference Hetzel1965). In the asteroid Patiria miniata, production of ‘crystal bodies’ was observed from tissue representing the coelomic epithelium; however, it is unknown which cell type from which crystal cells were derived (Johnson and Beeson, Reference Johnson and Beeson1966). Unlike other types of coelomocytes, crystal cells did not incorporate any injected carbon particles inside the cell, suggesting that they are not capable of phagocytosis of foreign particles and are already highly differentiated (Hetzel, Reference Hetzel1965; Zavalnaya et al., Reference Zavalnaya, Shamshurina and Eliseikina2020).
Occurrence of crystal cells
In holothuroids, crystal cells are commonly found in coelomic fluids (i.e., in the perivisceral and hydrovascular coeloms) and entirely absent from the connective tissue compartment of the body wall (Smiley, Reference Smiley, Harrison and Chia1994; Eliseikina and Magarlamov, Reference Eliseikina and Magarlamov2002; Caulier et al., Reference Caulier, Hamel and Mercier2020; but see Ramírez-Gómez et al., Reference Ramírez-Gómez, Aponte-Rivera, Méndez-Castaner and García-Arrarás2010). They are also present in aggregates of coelomocytes in the holothuroid Cucumaria frondosa (Caulier et al., Reference Caulier, Hamel and Mercier2020); however, Dybas and Fankboner (Reference Dybas and Fankboner1986) did not find any crystal cells trapped in aggregates (or brown bodies) of the holothuroid Apostichopus japonicus. Crystal cells are also known from the coelomic fluids of the asteroid Patiria miniata (Johnson and Beeson, Reference Johnson and Beeson1966). In crinoids, they are found near the oesophagus and in the part of the visceral sac beneath the oral disc (Cuénot, Reference Cuénot1891).
Proportion of crystal cells among coelomocytes
The proportion of crystal cells relative to other types of coelomocytes is generally low in holothuroids, ranging from 0.4 to 4.6% (Hetzel, Reference Hetzel1963; Queiroz et al., Reference Queiroz, Mauro, Arizza, Custódio and Vazzana2022b; Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025). In particular, densities are reportedly greater in the perivisceral than in the hydrovascular fluids even though not all individuals contain crystal cells (Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025). However, in the holothuroid Apostichopus japonicus, no difference in crystal cell abundance was detected between perivisceral and hydrovascular fluids (Li et al., Reference Li, Ri, Wang, Ye, Qiao and Li2013).
Factors affecting abundance and size of crystal cells
Density and size of crystal cells were reportedly the same between small (∼1 cm long) and larger individuals of the holothuroid Psolus phantapus (Théel, Reference Théel1921). Moreover, the size of crystal cells reportedly varied within the same individual (Hetzel, Reference Hetzel1963, Reference Hetzel1965), while their density tended to decrease over the course of the day (Hetzel, Reference Hetzel1963). In Apostichopus japonicus, densities of crystal cells did not vary between wild and cultured individuals or among individuals of different sizes (Li et al., Reference Li, Ri, Wang, Ye, Qiao and Li2013; Liang et al., Reference Liang, Ren, Wang, Li, Yang and Li2019).
Reassessment of crystal cells in holothuroids
Resemblance with microcrystals of uric acid
Upon re-examination of crystal-bearing structures, traditionally defined as crystal cells, isolated from coelomic cavities of the holothuroids Cucumaria frondosa and Psolus fabricii, a variety of microcrystals were recorded (Figure 1A–I) and convincingly identified as uric acid based on their morphology, and optical and chemical properties (Ma et al., Reference Ma, Gao, Hamel and Mercier2025). A summary of the parallels between these uric acid microcrystals and so-called crystal cells is provided below.
Microcrystals of uric acid isolated from holothuroids: (A) a rhombic crystal, (B) a rhombohedral crystal, (C) two rhombic crystals forming a multi-layered structure, (D) a smaller rhombic crystal growing from a larger one, (E) an irregular star-shaped crystal, (F) a cuboidal crystal, (G) barrel-shaped crystal, (H) a rhombic crystal, (I) three rhombic crystals overlayed at offsetted angles forming a six-pointed star-shaped structure, (J) a rhombic crystal under polarised light, (K) a rhombic crystal imaged using SEM, (L) two rhombic crystals and a foreign particle (arrow) encapsulated in a coelomocyte, (M) a rhombic crystal and amorphous substance (arrow) encapsulated in a coelomocyte, (N) two rhombic crystals encapsulated in a coelomocyte, (O) a single rhombic crystal encapsulated in a coelomocyte, (P) a single cuboidal crystal encapsulated in a coelomocyte, (Q) four cuboidal crystals encapsulated in a coelomocyte, (R) a rhombic crystal encapsulated in a coelomocyte with filipodia (arrow), (S) a rhombohedral crystal encapsulated in a coelomocyte with filipodia (arrow), and (T) a rhombic crystal trapped in an aggregate of haemocyte coelomocytes. Microcrystals illustrated in panels A–G, J–M, and O–T were isolated from Cucumaria frondosa and those in panels H, I, and N were from Psolus fabricii. Scale bars represent 10 μm.

Morphological properties
The shapes and sizes of uric acid microcrystals are consistent with those of the crystalline structures ascribed to ‘crystal cells’ (Tables 1 and 2). For instance, the lengths of rhomb-shaped microcrystals in Cucumaria frondosa (i.e., 5–29 μm; Ma et al., Reference Ma, Gao, Hamel and Mercier2025) and Psolus fabricii, and the diameter of a thin star-shaped in P. fabricii (a variant of the rhombic shape; e.g., Hetzel, Reference Hetzel1963) remains largely within the reported range of rhomb-shaped crystal-cell coelomocytes (Table 2). Furthermore, the width of these crystals in C. frondosa (i.e., 4–20 μm; Ma et al., Reference Ma, Gao, Hamel and Mercier2025) also aligns closely with the reported range of ‘crystal cells’ (Table 2).
Additionally, the width-to-length ratio of rhomb-shaped microcrystals of uric acid in C. frondosa had a mean of 0.60 and a maximum of 0.69 (derived from data collected by Ma et al., Reference Ma, Gao, Hamel and Mercier2025), in line with the width-to-length ratios of rhomb-shaped crystal-cell coelomocytes (described above) estimated to average 0.66, based on an analysis of published values, i.e., 0.50–0.63 (Molpadia roretzii; Ohuye, Reference Ohuye1936); 0.57–0.63 (Chia and Xing, Reference Chia and Xing1996; Xing, Reference Xing1997; Xing et al., Reference Xing, Leung and Chia1998); 0.60 (P. fabricii); 0.67, using maximum values (Hetzel, Reference Hetzel1963, Reference Hetzel1965); and 0.78–0.82 (Holothuria leucospilota; Endean, Reference Endean1958, Reference Endean and Boolootian1966).
Optical properties
Rhomb-shaped microcrystals of uric acid are colourless in the holothuroid Cucumaria frondosa (Ma et al., Reference Ma, Gao, Hamel and Mercier2025), but they can exhibit some colouration depending on the solution and surrounding material. Accordingly, the variable colours of ‘crystal cells’ reported in the literature (see above) may also be influenced by the tint of the solution in which they were prepared or the lighting conditions during observation. Moreover, rhombic crystalline structures isolated from the holothuroid C. frondosa were optically active under polarised light (Figure 1J), which is a known property of uric acid microcrystals. Finally, difficulties reportedly encountered when trying to image rhombic forms of uric acid crystals with SEM (Figure 1K) find echoes in some studies of ‘crystal cells’ (e.g., Queiroz et al., Reference Queiroz, Arizza, Vazzana and Custódio2022a).
Chemical properties
Solubility tests and Raman spectroscopy of cuboidal–barrel-shaped and rhomb-shaped microcrystals isolated from Cucumaria frondosa (Ma et al., Reference Ma, Gao, Hamel and Mercier2025), which were previously identified as crystal-cell coelomocytes (Caulier et al., Reference Caulier, Hamel and Mercier2020; Jobson et al., Reference Jobson, Hamel, Hughes and Mercier2021), provided more precise information on their chemical properties, which are consistent with those of uric acid. Although chemical analyses have so far focused on these two common morphologies, other crystal forms observed in echinoderms may represent additional morphological variants of uric acid (e.g., rosette-shaped; Kleeberg et al., Reference Kleeberg, Gordon, Kedar and Dobler1981) crystallised under different physicochemical conditions or, possibly, crystals composed of other chemical compounds.
Evidence of phagocytosis of uric acid microcrystals by coelomocytes
Similar to reports of ‘crystal cells’, rhomb-shaped, cuboidal, and barrel-shaped microcrystals of uric acid in the holothuroid Cucumaria frondosa were found in the perivisceral and hydrovascular fluids; however, the same microcrystals were also detected in the tissues of some internal organs (Ma et al., Reference Ma, Gao, Hamel and Mercier2025). In addition, cuboidal and rhombic microcrystals of uric acid were found encapsulated (i.e., in the ‘crystal cell’ appearance), suggesting that they were phagocytised by a coelomocyte. Small foreign particles may be trapped in the cell (Figure 1L), possibly giving the impression of a nucleus, and an amorphous substance may be present at one or both ends of a rhombic crystal (Figure 1M). Up to two rhomb-shaped crystals of uric acid can be packaged within a single coelomocyte (Figure 1L and N), which matches the reports of crystal-cell coelomocytes with two or more crystals in them (Endean, Reference Endean1958, Reference Endean and Boolootian1966; Smith, Reference Smith, Ratcliffe and Rowley1981; Smiley, Reference Smiley, Harrison and Chia1994). Although usually a single crystal is packaged inside a cell (Figure 1O and P), multiple cuboidal crystals, with up to four within a single cell, have been observed in C. frondosa (Figure 1Q; Ma et al., Reference Ma, Gao, Hamel and Mercier2025). Filipodia were observed on the cell membrane surrounding rhombic (Figure 1R) and rhombohedral microcrystals of uric acid (Figure 1S) (Ma et al., Reference Ma, Gao, Hamel and Mercier2025), which is similar to the description of ‘crystal cells’ with pseudopodia by Hetzel (Reference Hetzel1963, Reference Hetzel1965). Although many of these packaged microcrystals were stationary, a few displayed subtle vibratory/wobbly movements (K. C. K. Ma, personal observation).
Encapsulated microcrystals have been detected in the perivisceral coelomic fluid and the water expelled out of the respiratory tree, which is presumably how they get released from the body (Ma et al., Reference Ma, Gao, Hamel and Mercier2025). Another excretion pathway was detected in the form of non-encapsulated crystals of uric acid trapped in the brown-coloured aggregates (Figure 1T), likely in the process of being eliminated through transrectal coelomoducts in the cloaca (Dybas and Fankboner, Reference Dybas and Fankboner1986; Caulier et al., Reference Caulier, Hamel and Mercier2020; Jobson et al., Reference Jobson, Hamel and Mercier2022). The presence of these excretory naked microcrystals circles back to an observation made a century ago suggesting that the growth of crystalloids in phagocytic coelomocytes caused the degeneration and, eventually, the collapse of the cell (Kindred, Reference Kindred1924).
Conclusion
The detection of crystalline structures with a geometric shape (i.e., prismatic), refringent properties, and resistance to vital staining among coelomocytes of echinoderms gave rise long ago to the definition of so-called crystal cells in the literature. At one point, they were speculated to represent crystalline structures engulfed by phagocytic coelomocytes in asteroids, but direct observations of the process were lacking to support the hypothesis (Johnson and Beeson, Reference Johnson and Beeson1966) and it was largely overlooked. Since then, the validity of classifying ‘crystal cells’ as a distinct type of coelomocyte in echinoderms has been anecdotally called into question a number of times, but never formally examined. Furthermore, the putative role of ‘crystal cells’ in osmoregulation and ossicle formation has been downplayed due to the lack of direct evidence (Endean, Reference Endean and Boolootian1966; Ramírez-Gómez and García-Arrarás, Reference Ramírez-Gómez and García-Arrarás2010; Jobson et al., Reference Jobson, Hamel, Hughes and Mercier2021; see also Diehl Reference Diehl1986, for a review on osmoregulation in echinoderms). Some authors had started referring to them as ‘assumed crystal cells’ or excluding them entirely from analyses (Carter et al., Reference Carter, Jobson, Hamel and Mercier2024; Wambreuse et al., Reference Wambreuse, Guillaume, Eeckhaut, Borrello, Bureau, Fievez and Delroisse2025). Following further investigation of their morphology and polymorphism, optical activity under polarised light, and solubility, these crystalline structures were determined to be uric acid in holothuroids (Ma et al., Reference Ma, Gao, Hamel and Mercier2025), as described in many other phyla. The distribution of free and encapsulated microcrystals sheds further light on their genesis and elimination from the sea cucumber body (Ma et al., Reference Ma, Gao, Hamel and Mercier2025). Taken together, the lines of evidence strongly support that what has been termed ‘crystal cells’ is better understood as a functional state of phagocytic coelomocytes dedicated to the sequestration and transport of crystalline uric acid wastes. This shift in interpretation not only resolves inconsistencies in the roles attributed to this type of coelomocyte but also provides a clearer understanding of metabolic waste management in echinoderms.
Acknowledgements
We extend our thanks to Fei Gao and Sara Jobson for their insights throughout the research process.
Author contributions
KCKM: Conceptualisation, Investigation, Writing – original draft, Writing – review & editing. J-FH: Conceptualisation, Investigation, Writing – review & editing. AM: Conceptualisation, Funding Acquisition, Investigation, Supervision, Writing – review & editing.
Financial support
This study was supported by Mitacs Accelerate internships (IT15224 and IT29594) and grants from the Natural Sciences and Engineering Research Council of Canada (NSERC) to AM.
Conflict of interest
The authors declare no conflict of interest.
Data availability
The authors confirm that the data supporting the findings of this study are available within the article.
Ethical standards
Not applicable. This work is based on previously published data, and no new experiments involving sea cucumbers were carried out.


 Open access
Open access