


Introduction
The domestic cat Felis catus has a long history with humans, originating as a commensal relationship 11,000–10,500 years ago (Vigne et al., Reference Vigne, Carrere, Briois and Guilaine2011). Much appreciated as self-sustaining pest controllers (Krajcarz et al., Reference Krajcarz, Krajcarz, Baca, Baumann, Van Neer and Popović2020), cats were employed in the storehouses of emerging civilizations and for rodent control on trade ships (Ottoni et al., Reference Ottoni, Van Neer, de Cupere, Daligault, Guimaraes and Peters2017), thereby spreading worldwide (Long, Reference Long2003). Over time, cats have been partially domesticated, developing an increasing dependency on anthropogenic resources for food and shelter, and some behavioural adaptations to human interactions (Crowley et al., Reference Crowley, Cecchetti and McDonald2020; Krajcarz et al., Reference Krajcarz, Krajcarz, Baca, Baumann, Van Neer and Popović2020). They have become one of the most popular pets, with the global population being estimated at c. 600 million in 2009 (Driscoll et al., Reference Driscoll, Clutton-Brock, Kitchener and O’Brien2009). Nevertheless, cats have maintained a carnivorous diet and a fully functional hunting repertoire (Cecchetti et al., Reference Cecchetti, Crowley and McDonald2021c), enabling them to kill prey and survive independently from human assistance (Bradshaw, Reference Bradshaw2006). In contemporary times, the ubiquity, abundance and free-roaming habits of cats are associated with conspicuous ecological impacts (e.g. Loss et al., Reference Loss and Marra2017; Woinarski et al., Reference Woinarski, Murphy, Legge, Garnett, Lawes and Comer2017; Kays et al., Reference Kays, Dunn, Parsons, McDonald, Perkins and Powers2020; Loss et al., Reference Loss, Boughton, Cady, Londe, McKinney and O’Connell2022). Additionally, the presence of free-roaming cats has indirect effects on wild prey populations, such as inducing fear, which affect prey physiology, behaviour, movements and activity schedules (Bonnington et al., Reference Bonnington, Gaston, Evans and Whittingham2013; Webster et al., Reference Webster, Massaro, Michael, Bambrick, Riley and Nimmo2018).
The impacts of cats are particularly severe on island ecosystems (Bonnaud et al., Reference Bonnaud, Medina, Vidal, Nogales, Tershy and Zavaleta2011; Doherty et al., Reference Doherty, Glen, Nimmo, Ritchie and Dickman2016), where native species tend to lack defensive traits and behaviours to deal with novel predators (Wallach et al., Reference Wallach, Ramp, Benítez-López, Wooster, Carroll and Carthey2023). On islands, cats often become the main predator, contributing to the decline and extinction of insular endemic birds, mammals, reptiles and breeding seabirds (Doherty et al., Reference Doherty, Glen, Nimmo, Ritchie and Dickman2016). The latter are particularly vulnerable to cat predation because of their limited mobility on land and their exposed nests (Doherty et al., Reference Doherty, Glen, Nimmo, Ritchie and Dickman2016). In addition, islands are important stop-over sites for migratory birds, where they can rebuild energy reserves, and exercise other behaviours such as recovery, sleep, information gathering and interactions (Linscott & Senner, Reference Linscott and Senner2021). These birds are highly susceptible to cat predation (Medina et al., Reference Medina, Bonnaud, Vidal and Nogales2014). The diet of free-roaming cats on islands has been extensively studied, showing they are generalist predators that depredate whichever species are available (e.g. Bonnaud et al., Reference Bonnaud, Medina, Vidal, Nogales, Tershy and Zavaleta2011; Ozella et al., Reference Ozella, Cecchetti and Pessani2016; Galão et al., Reference Galão, Soto, Nunes, Pedroso, Rocha and Rato2025).
Although the ecological impacts of cats on islands are well known, the intrinsic and extrinsic factors that influence individual hunting behaviour, such as age, sex, neutering status, time spent outdoors and type of food provided, have been less studied (Lilith et al., Reference Lilith, Calver, Styles and Garkaklis2006; McDonald et al., Reference McDonald, Maclean, Evans and Hodgson2015; Cecchetti et al., Reference Cecchetti, Crowley, Goodwin and McDonald2021b; Mella-Méndez et al., Reference Mella-Méndez, Flores-Pered, Amaya-Espinel, Bolívar-Cimé, Mac Swinet and Martínez2022). Free-roaming cats on islands are most often characterized as pests, even where they were originally introduced to control rodent pests (Driscoll et al., Reference Driscoll, Clutton-Brock, Kitchener and O’Brien2009). On some islands, such as Little Barrier, New Zealand, their role in pest control is still important for reducing predation by non-native rodents on breeding seabirds (Rayner et al., Reference Rayner, Hauber, Imber, Stamp and Clout2007). A key factor in cat population management is to avoid the mesopredator release effect (Courchamp et al., Reference Courchamp, Langlais and Sugihara1999), in which the removal of a top predator, such as a cat, may be followed by an irruption of mesopredators, such as rats, magnifying detrimental effects on endemic species, as documented on Marion Island, South Africa (Courchamp et al., Reference Courchamp, Langlais and Sugihara1999; Huntley et al., Reference Huntley, Huntley, Lindsey and Zwaenepoel2023).
Cats elicit human emotional responses, and this is likely to have contributed historically to the establishment of populations through the provisioning of resources (Jarić et al., Reference Jarić, Courchamp, Correia, Crowley, Essl and Fischer2020). For instance, the provision of food enables the persistence of large cat populations that are less regulated, if at all, by prey availability (Sims et al., Reference Sims, Evans, Newson, Tratalos and Gaston2008; Maeda et al., Reference Maeda, Nakashita, Shionosaki, Yamada and Watari2019; Crowley et al., Reference Crowley, Cecchetti and McDonald2020). Previous studies on inhabited islands, such as Florida Keys, USA, and Tokunoshima Island, Japan, have shown that cats subsidized by humans exacerbate predation pressure on vulnerable species through hyperpredation (Cove et al., Reference Cove, Gardner, Simons, Kays and O’Connell2018; Maeda et al., Reference Maeda, Nakashita, Shionosaki, Yamada and Watari2019). This occurs when hunting motivation and prey consumption are decoupled, leading to surplus killing when opportunities arise (Adamec, Reference Adamec1976; Macdonald & Rogers, Reference Macdonald and Rogers1984). Even pet cats that kill wild prey often consume only small amounts and rely predominantly on food provided by people (Cecchetti et al., Reference Cecchetti, Crowley, Goodwin, Cole, McDonald, Bearhop and McDonald2021a). Hyperpredation is further facilitated by the availability of abundant introduced prey, such as European rabbits Oryctolagus cuniculus, rats Rattus spp. and house mice Mus musculus (Courchamp et al., Reference Courchamp, Langlais and Sugihara2000). On Macquarie Island, Australia, the introduction of rabbits provided an additional food source for cats, leading to population growth and intensified predation on native birds, ultimately contributing to local extinctions (Taylor, Reference Taylor1979).
A range of management strategies are used to control cat populations (Cecchetti et al., Reference Cecchetti, Crowley and McDonald2021c), and those commonly adopted to control free-roaming, feral and unowned cats on islands often involve lethal methods, which have been successfully applied for some cat eradications (Nogales et al., Reference Nogales, Martin, Tershy, Donlan, Veitch, Puerta, Wood and Alonso2004). However, the outcomes of culling can be affected by compensatory mechanisms (Lazenby et al., Reference Lazenby, Mooney and Dickman2015; Palmas et al., Reference Palmas, Gouyet, Oedin, Millon, Cassan and Kowi2020), increase in the risk of mesopredator release, and potential lack of support from local people (Campbell et al., Reference Campbell, Harper, Algar, Hanson, Keitt, Robinson, Veitch, Clout and Towns2011; Mameno et al., Reference Mameno, Kubo and Suzuki2017).
The trap–neuter–return method, which involves trapping, neutering and releasing cats at the capture site, is an alternative for managing unowned cat populations (Slater et al., Reference Slater, Di Nardo, Pediconi, Dalla Villa, Candeloro, Alessandrini and Del Papa2008; Wolf & Schaffner, Reference Wolf and Schaffner2019). However, its effectiveness has been debated, especially on large islands and mainlands, where substantial effort is required (Benka et al., Reference Benka, Boone, Miller, Briggs, Anderson and Slootmaker2022; Gunther et al., Reference Gunther, Hawlena, Azriel, Gibor, Berke and Klement2022; Lepczyk et al., Reference Lepczyk, Duffy, Bird, Calver, Cherkassky and Cherkassky2022) and population dynamics are influenced by the immigration of unneutered cats (Gunther et al., Reference Gunther, Finkler and Terkel2011). Feeding stations associated with trap–neuter–return may attract rodents and scavengers, exacerbating predation on wildlife (Grant & Longnecker, Reference Grant and Longnecker1999; Cove et al., Reference Cove, Gardner, Simons, Kays and O’Connell2018; Herrera et al., Reference Herrera, Cove, McShea, Flockhart, Decker, Moore and Gallo2022). Although neutered cats show reduced home ranges (Ferreira et al., Reference Ferreira, Machado, Nakano-Oliveira, Andriolo and Genaro2020), the impact of neutering on reducing wildlife predation remains uncertain. Trap–neuter–return programmes can achieve long-term population control if performed intensively and combined with measures such as the removal and adoption of sociable cats, reduction in abandonment, early desexing, and euthanasia of ill individuals (Gunther et al., Reference Gunther, Raz and Klement2018, Reference Gunther, Hawlena, Azriel, Gibor, Berke and Klement2022; Benka et al., Reference Benka, Boone, Miller, Briggs, Anderson and Slootmaker2022; McDonald et al., Reference McDonald, Finka, Foreman-Worsley, Skillings and Hodgson2023). Additionally, because of ethical considerations and public support, trap–neuter–return has become increasingly widespread and, in some European countries, remains the only permitted approach, following the adoption of no-kill policies for unowned cats (e.g. Italy: Law 281/91; Spain: Law 7/2023). This approach may be a viable option for managing cats humanely on small, isolated islands without immigration (Cecchetti & Nelli, Reference Cecchetti and Nelli2025).
Here we investigate the foraging ecology of free-roaming cats on the Mediterranean island of Linosa, estimating the contributions of wild animals vs provisioned foods to their diets, using stable isotope analysis of whiskers and by testing differences between unneutered and neutered individuals. The stable isotope technique provides information on the long-term contributions of major food groups and is less affected by differences in digestibility compared to scat analysis (Barrett et al., Reference Barrett, Camphuysen, Anker-Nilssen, Chardine, Furness and Garthe2007), and is non-invasive. A high dependency on provisioned foods would provide potential evidence of human-driven hyperpredation (e.g. Courchamp et al., Reference Courchamp, Langlais and Sugihara2000; Cove et al., Reference Cove, Gardner, Simons, Kays and O’Connell2018). We aimed specifically to investigate assimilated diet rather than overall predation patterns. Although carcass recovery and prey observations can provide useful insights into which species are targeted or killed, they do not necessarily reflect consumption or nutrient assimilation (Cecchetti et al., Reference Cecchetti, Crowley, Goodwin, Cole, McDonald, Bearhop and McDonald2021a).
We also hypothesized that variation in the cat–human relationship, specifically control over reproduction and feeding, would be associated with variation in the dependence of cats on wildlife. Such findings would provide a basis for increasing public awareness of the cat–wildlife issue and, potentially, for collaborating with residents in sterilization activities and promoting responsible cat ownership and better management of cat feeding stations.
Study area
Linosa is a 5.45 km2 volcanic island in the Sicilian Channel, Mediterranean Sea, between Italy and Tunisia, part of the Pelagie Archipelago (Fig. 1). It is a biodiversity hotspot and part of the Natura 2000 Network (EEA, 2023). It has a semi-arid Mediterranean climate, with mild winters (13–16 °C), hot summers (25–32 °C) and annual rainfall of 300–400 mm.
Locations of all free-roaming cats Felis catus in Linosa in 2022, and of Scopoli’s shearwater Calonectris diomedea nests monitored each year in the north of the island. The satellite image was generated using the Esri (Redlands, USA) world imagery basemap.

Linosa hosts endemic species such as the Filfola lizard Podarcis filfolensis, ocellated skink Chalcides ocellatus and big-head cricket Brachytrupes megacephalus, and the largest European colony of Scopoli’s shearwater Calonectris diomedea (c. 10,000 pairs; Massa & Valvo, Reference Massa and Valvo1986). These pelagic seabirds nest in natural cavities and caves in volcanic rock along the coast, mostly in the north of the island, where annual monitoring of c. 400 breeding pairs is conducted. Females lay a single egg in late May, chicks hatch mid July to early August, and fledglings leave by late October (Müller et al., Reference Müller, Massa, Phillips and Dell’Omo2015). The diet of Scopoli’s shearwater in the Mediterranean is dominated by pelagic fish and crustaceans, with non-pelagic fish and cephalopods occasionally taken during egg incubation (Thabet et al., Reference Thabet, Bourgeois, Le Loc’h, Abdennadher, Munaron and Gharsalli2019). Stable isotope analyses also indicate trophic segregation, suggesting that adults feed more on inshore resources while provisioning chicks with a greater proportion of oceanic prey (Thabet et al., Reference Thabet, Bourgeois, Le Loc’h, Abdennadher, Munaron and Gharsalli2019). In Linosa the diet may also include demersal fish discarded from fishing vessels (Cianchetti-Benedetti et al., Reference Cianchetti-Benedetti, Catoni, Kato, Massa and Quillfeldt2017).
Linosa is a stopover for many migratory birds, particularly passerines; c. 200 species stop over in spring (maximum peak in April–May), and c. 150 in autumn (maximum peak in October; eBird, 2021). Many of these species are considered Species of European Conservation Concern (SPEC) and/or are included in Annex 1 of the Birds Directive; e.g. the Eurasian thick-knee Burhinus oedicnemus (SPEC 3, Annex 1), great snipe Gallinago media (SPEC 1, Annex 1) and Eurasian dotterel Eudromias morinellus (SPEC 3, Annex 1).
Since its colonization by people, Linosa has been exposed to the introduction of invasive species such as the black rat Rattus rattus, house mouse and European rabbit. Inevitably, companion animals such as domestic dogs Canis familiaris, mainly kept for hunting, and domestic cats have also been introduced.
Cat population and management
Linosa has a large population of domestic cats, probably introduced as pest controllers. Although still appreciated for their rodent-hunting abilities, cats are also kept as companion and commensal animals, making their management complex and contingent on human beliefs and perceptions.
For terminology, we follow the classification proposed by the Australian Cat Action Plan (Getting to Zero, 2018). Owned cats are directly cared for by specific people and are generally sociable, though sociability varies. They include both responsibly owned cats (e.g. desexed, confined) and casually owned cats (e.g. unconfined or not desexed because of owner beliefs, resources or knowledge). Unowned cats live independently in human environments, surviving by scavenging or being fed by carers, and may form colonies but lack formal ownership. Feral cats are unsocialized, unowned and fully independent of people, living and reproducing in the wild. The terms free-ranging and free-roaming refer to any cats allowed to move freely in the environment, regardless of ownership status. There are no feral cats on Linosa, as all cats can be traced back to an owner or caretaker (Cecchetti & Nelli, Reference Cecchetti and Nelli2025). Therefore, we refer to owned and unowned cats collectively as ‘free-roaming’.
Cats on Linosa also receive supplemental food, mainly in the form of cat food from tourists, particularly during summer. On Linosa, under Italian Law 280/91, cat population control involves trap–neuter–return for unsocialized cats, alongside the neutering of sociable cats.
Preliminary surveys in 2014 estimated a cat population of c. 300 (Ozella et al., Reference Ozella, Cecchetti and Pessani2016). Scat analyses from cats near the shearwater colony revealed diets primarily composed of introduced rats and rabbits, and Scopoli’s shearwaters and migratory birds (Ozella et al., Reference Ozella, Cecchetti and Pessani2016). However, faecal analysis underestimates highly digestible anthropogenic foods, which likely account for a significant dietary component. Observations in 2021–2022 documented predation of at least 44 migratory bird species, such as the European pied flycatcher Ficedula hypoleuca and goldcrest Regulus regulus at two free-roaming cat colonies (Cecchetti et al., Reference Cecchetti, Janni, Corso and Dell’Omo2023).
In September–October 2022, a door-to-door survey of all island residents identified 485 free-roaming cats and six indoor-only cats, an average density of 89 cats/km2. Each cat was traced to an owner or caretaker, and for each cat we recorded details (e.g. age, sex) and type of food provided (e.g. cat food, pasta). Multiple photographs were taken, and a unique ID assigned to each cat. Each cat’s house or colony location was recorded using a GPS (Fig. 1). Additional details of the census and trap–neuter–return sessions are described elsewhere (Cecchetti & Nelli, Reference Cecchetti and Nelli2025).
Methods
Diet analysis
Stable isotope analysis of metabolically inert tissues, such as whiskers, hair and feathers, have been widely used to study the diet of animals. Ratios of carbon (13C/12C), reported as δ13C, in consumer tissues indicate its relative reliance on primary carbon sources. Carbon enrichment varies according to plant photosynthetic pathway, such as C3 vs C4. The ratios of nitrogen (15N/14N), reported as δ15N, can also be used as an indicator of trophic level, or the amount of animal protein consumed. Cecchetti et al. (Reference Cecchetti, Crowley, Goodwin, Cole, McDonald, Bearhop and McDonald2021a) were able to discern provisioned cat foods from wild prey, which differed in both their δ13C and δ15N ratios, reflecting the use of plant-derived ingredients in commercial cat foods.
We obtained cat whiskers during 8–13 November 2022. A whisker was plucked from each cat under anaesthesia during surgery (mainly neutering, but also other surgeries). Prior to surgery, each cat underwent a clinical examination by the veterinarian and was tested for feline leukaemia virus (FeLV) and feline immunodeficiency virus (FIV) when deemed appropriate. Whisker samples were collected only from cats assessed as healthy at the time of the procedure. A whisker was also clipped from a random sample of cats for which owners/carers gave their consent. Whiskers were stored in a paper envelope in ambient conditions.
The previous work on the diet of a small sample of cats in Linosa was used as a starting reference for the wild prey cats may consume (Ozella et al., Reference Ozella, Cecchetti and Pessani2016). During September–November 2022, people were requested, via messages, calls or direct encounters, to collect wild prey found dead or brought home by their cats. These were collected by author MC and stored at −20 °C. To maximize wild prey collection, we also visited areas of cat activity to collect any wild prey items. Samples of rabbits were provided by hunters, and fish by fishers. Scopoli’s shearwater juveniles were collected when found dead during the fledging period. As cats prey primarily on chicks, fledgling muscle tissue provided the most appropriate isotopic proxy for our study. This choice is supported by evidence of dietary segregation between adults and chicks, with the latter receiving more oceanic prey resources (Thabet et al., Reference Thabet, Bourgeois, Le Loc’h, Abdennadher, Munaron and Gharsalli2019). Muscle tissue was also analysed for other prey.
From the cat census, we knew that cats were fed with a variety of purpose-made commercial cat foods and a range of human foods, primarily pasta-based meals (Plate 1), which were sampled from the cat owners/caretakers directly. To sample cat food, both dry and wet foods were bought in the shops on the island or, when possible, collected directly from the owner/caretaker.
Cat colony fed with pasta and leftovers in Linosa (Fig. 1). Some individuals have one ear with the tip clipped, to identify them as having been neutered (Photograph: Martina Cecchetti).

Isotope ratios were calculated for two whisker sections. The period represented by each whisker section was calculated by dividing the length of the section by the daily average whisker growth rate: 0.48 mm (range 0.4–0.5 mm/day) (Cecchetti et al., Reference Cecchetti, Crowley, Goodwin, Cole, McDonald, Bearhop and McDonald2021a). Cat whiskers were rinsed in distilled water, scraped to remove surface contaminants, and freeze-dried for 24 h. For each whisker, total weight and total length were measured. Then, each whisker was divided into sections based on its total weight, with the basal section representative of the most recent diet and the tip the oldest diet. Each whisker was cut into 0.4–0.7 mg sections and sealed in a tin cup for analysis. Before analysis, food sources were freeze-dried and homogenized, and then 0.6–0.8 mg was weighed into a tin cup.
Samples were analysed for δ13C and δ15N using a Sercon Integra-2 E A-isotope ratio mass spectrometer (Sercon, UK). Stable isotope ratios were expressed as δ values in ‰ (per mil), the ratio of heavy to light isotopes, relative to the isotopic ratios of an international standard for each element: the Vienna Pee Dee Belemnite (VPDB) for δ13C and atmospheric N2 for δ15N. The estimated mean precision between sample runs was 0.07 ± SD 0.03 for δ15N and 0.10 ± SD 0.06 for δ13C, based on standards run within sample batches. We applied mathematical lipid normalization to δ13C values of samples with high lipid content (Kiljunen et al., Reference Kiljunen, Grey, Sinisalo, Harrod, Immonen and Jones2006; Post et al., Reference Post, Layman, Arrington, Takimoto, Quattrochi and Montaña2007; Cecchetti et al., Reference Cecchetti, Crowley, Goodwin, Cole, McDonald, Bearhop and McDonald2021a).
Statistical analyses and isotope mixing models
All statistical analyses were conducted in R 4.4 (R Core Team, 2024). To test whether the cats’ diet was affected by cat characteristics and extrinsic factors, we analysed variation in δ13C and δ15N of individual cats using two linear mixed models (LMM). Fixed factors were sex, age range (< 6 months, 6–11 months, 1–5 years, 6–10 years), neuter status (neutered, unneutered) and season (summer, autumn). Cat identity was set as a random effect. Explanatory variables were checked for correlations using Spearman’s correlation test. Given the high negative correlation between age range and neuter status, we inserted only neuter status in the global models.
Model selection was performed using the dredge function in the MuMIn package (Bartoń, Reference Bartoń2025), based on the Akaike information criterion (AIC). Model fit was assessed with the performance package (Lüdecke et al., Reference Lüdecke, Ben-Shachar, Patil, Waggoner and Makowski2021). Only cats with two whisker segments were included.
Subsequently, to estimate the relative contributions of wild prey and provisioned food source to the cats’ diet, we ran a Bayesian isotopic mixing model built with 1,000,000 iterations, a burn-in of 50,000, and a thinning rate of 50, using the package SIMMR 0.4.5 (Parnell et al., Reference Parnell, Inger, Bearhop and Jackson2010). We included the type of segment (basal and distal, reflecting different periods), as a grouping variable. The same structure was used to run a second and third model, to investigate variations in diets of neutered and unneutered cats during the two seasons. Gelman diagnostics were used to check model convergence. Trophic discrimination factors for cats were 2.9 ± SD 1.3‰ for δ13C and 1.9 ± SD 0.7‰ for δ15N (Cecchetti et al., Reference Cecchetti, Crowley, Goodwin, Cole, McDonald, Bearhop and McDonald2021a).
Bayesian estimates of diet composition by season and neuter status should be viewed as a means of gauging the relative importance of food sources across groups, rather than deriving precise estimates, because of the increased influence of uninformative priors in the models with lower numbers of observations (Swan et al., Reference Swan, Bearhop, Redpath, Silk, Goodwin, Inger and McDonald2020).
Results
Whiskers were taken from a sample of 87 cats, of which 22 were neutered and 65 unneutered. The sample comprised 53 females and 34 males, of which six were < 6 months old, 36 aged 6–11 months, 39 adults of 1–5 years, and six aged 6–10 years. The age of each cat was estimated during neutering based on physical characteristics such as size and dental development, or using information provided by owners or caretakers if available. The mean length of the basal section of whiskers was 17 mm (95% CI 14–20 mm; 87 cats), corresponding to growth over c. 1 month (35 days), and the distal section was 30 mm (95% CI 26–34 mm; 70 cats), indicating c. 2 months of diet (63 days; Fig. 2). The mean ratios of stable carbon isotopes of the basal and distal whisker sections were −21.2 ± SD 1.98‰ and −18.70 ± SD 1.00‰, and those of nitrogen were 7.98 ± SD 1.31‰ and 7.20 ± SD 1.03‰, respectively.
Timeline showing the breeding of Scopoli’s shearwater, passerine migratory periods, the tourist season and seasonality of the cat dietary analysis. Sampling of the cats’ whiskers took place during 8–13 November; the basal section corresponds to the peak of passerines stopping in Linosa during the autumn migration, and the distal section extends to the beginning of August, during early chick rearing by Scopoli’s shearwater.

The best LMM explaining variation in δ15N included neuter status and season as fixed effects. Neutered cats showed a reduction in δ15N compared to unneutered cats (estimate = −0.86, 95% CI −1.47–−0.24, P = 0.008). δ15N was lower in summer compared to autumn (estimate = −0.91, 95% CI −1.1–−0.72, P < 0.001). The total variance (R 2 c) explained by the model was 0.8. Regarding variation in δ13C, the initial LMM fit was singular, indicating a negligible influence of random effects. Therefore, the model was refitted as a linear model. The best linear model, which again included neuter status and season, revealed higher δ13C values in neutered cats (estimate = 0.72, 95% CI 0.10–1.33, P = 0.023) and during summer (estimate = 2.77, 95% CI 2.27–3.28, P < 0.001). R2c for this model was 0.5.
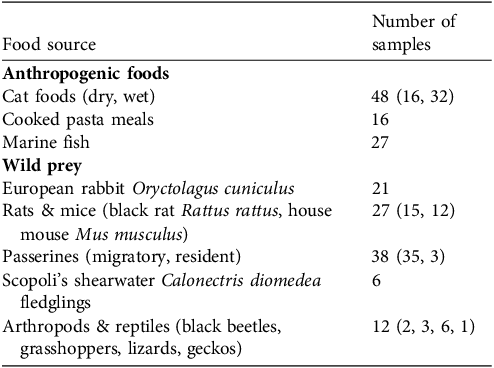
Food sources were grouped into eight broad, isotopically distinct categories for which we collected 195 samples (Tables 1 & 2): cat food (dry and wet); cooked pasta dishes (e.g. pasta with oil and/or sauces); marine fish (e.g. tuna, swordfish); passerines (e.g. European robin Erithacus rubecula, Eurasian skylark Alauda arvensis, European stonechat Saxicola rubicola); Scopoli’s shearwaters; arthropods and reptiles (e.g. grasshoppers, lizards); rabbits; and rats and mice.
Number of samples of each food source collected for stable isotope analysis. Food sources include anthropogenic foods (commercial cat food, pasta meals and fish) and wild prey (birds, arthropods, reptiles and mammals).

Wild prey species, found dead or brought home by cats, collected on Linosa Island (Fig. 1).

Stable isotope mixing models showed that in both periods cat diet comprised predominantly anthropogenic foods (Fig. 3, Table 3). In summer, cat food represented the bulk of the diet; fish and pasta comprised a smaller proportion. Consumption of Scopoli’s shearwaters was high compared to the consumption of passerines, arthropods and invasive mammals (Fig. 3a, Table 3). In autumn, cats showed decreased consumption of cat foods and Scopoli’s shearwaters, and an increased consumption of all other items; fish consumption remained broadly similar (Table 3, Fig. 3b).
Stable isotope ratios (δ15N and δ13C) of cats’ whiskers and their putative food sources (Table 1) in (a) summer, corresponding to the distal sections of cats’ whiskers (N = 70), and (b) autumn, corresponding to the basal sections of cats’ whiskers (N = 87). Food sources are shown as the mean ± SD, adjusted by the trophic discrimination factors. Higher δ15N and δ13C values indicate higher trophic levels and a greater contribution of marine- or C4-derived resources, respectively.

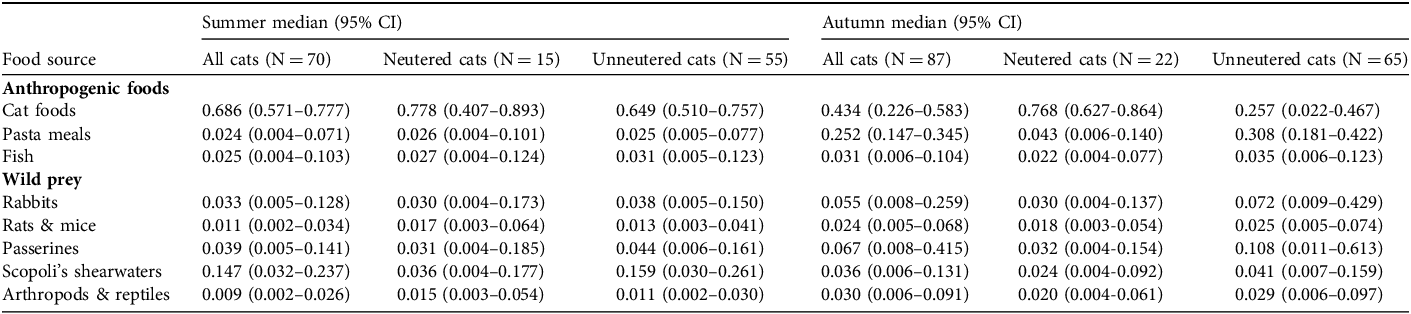
Estimates of the composition of the diet of free-roaming domestic cats on Linosa Island in summer and autumn, for all cats combined and separately for neutered and unneutered individuals. Estimates are proportions from Bayesian stable isotope mixing models, based on analysis of cat whiskers and eight putative food sources, including anthropogenic foods and wild prey. Diet composition was estimated for both summer and autumn, corresponding to the distal and basal sections of the cats’ whiskers, respectively.

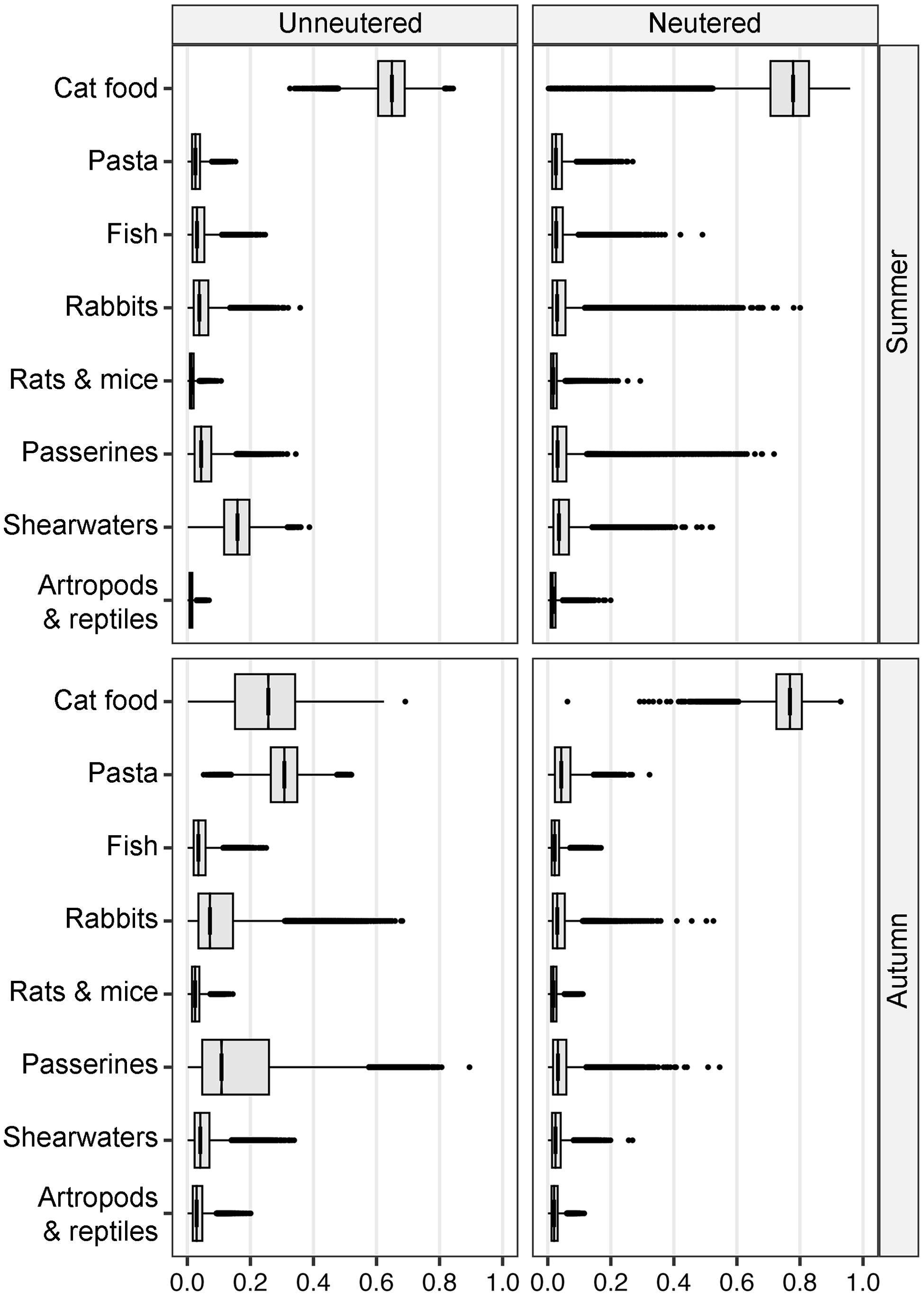
Unneutered cats had higher consumption of Scopoli’s shearwaters in summer, but the diets of neutered and unneutered cats were otherwise similar (Table 3, Fig. 4). In autumn, they differed in the consumption of anthropogenic foods, with cat foods more important for the diet of neutered cats, and pasta meals for unneutered cats. Consumption of wild animals, particularly passerines and rabbits, was higher in unneutered compared to neutered cats (Table 3, Fig. 4).
Estimates of the proportional composition of the diet of unneutered and neutered free-roaming cats in summer (August–early October) and autumn (early October–mid November), determined by Bayesian stable isotope mixing models of cat whiskers and their potential food sources on Linosa. The diet includes anthropogenic (cat foods, pasta meals, and provisioned fish) and wild food (rabbits, rats and mice, passerines, Scopoli’s shearwaters, and arthropods and reptiles). In the box plots, the horizontal line represents the median (Table 3), the box spans the 25th to 75th percentiles, the whiskers extend to 1.5 times the interquartile range, and individual points represent outliers.

Discussion
Stable isotope analysis confirmed that cats in Linosa relied significantly on foods provided by people, mainly cat foods and pasta, and small amounts of fish. Such regular availability of food across seasons is probably sustaining hyperpredation on endemic species, as reported on other islands (Cove et al., Reference Cove, Gardner, Simons, Kays and O’Connell2018; Maeda et al., Reference Maeda, Nakashita, Shionosaki, Yamada and Watari2019). Provision of cat foods was higher during summer, probably because of tourists feeding the cats roaming around rented houses. In both periods, introduced mammals (primarily rabbits, with fewer rats and mice) also contributed to the cats’ diet, potentially facilitating hyperpredation on wild species (Medina et al., Reference Medina, Bonnaud, Vidal, Tershy, Zavaleta and Donian2011; Palmas et al., Reference Palmas, Jourdan, Ringler, Cucherousset, Villeger and Oldeland2017).
Free-roaming cats in Linosa consumed a significant amount of wildlife. During the shearwater chick-rearing period, there was a high ingestion of these seabirds, probably reflecting the consumption of chicks, which are left alone during the day while parents are foraging at sea. As chicks grew and became juveniles, the per cent of shearwaters in cat diet decreased. Although shearwater population dynamics are particularly sensitive to changes in adult survival, even low rates of chick predation can have severe consequences for the survival of the colony in the long term (Le Corre, Reference Le Corre2008). Loss of chicks and juveniles may also cause separation of mates and nest abandonment, disrupting reproductive success in subsequent years (Greenwell et al., Reference Greenwell, Calver and Loneragan2019).
Consumption of passerines increased during the peak of migratory passerines stopping in Linosa in mid October. Vulnerability to cat predation is considered to be higher for lean birds during stopovers because of more intense foraging and displacement to suboptimal habitats (Dierschke, Reference Dierschke2003). Moreover, the risk of predation could modify stopover behaviours, inducing birds to spend less time foraging or resting, indirectly decreasing bird survival during migration (Lank & Ydenberg, Reference Lank and Ydenberg2003).
Cats in Linosa consumed small numbers of native arthropods and reptiles; stable isotope analysis could not distinguish these groups, although all reptiles and most arthropods on Linosa are endemic. The use of more taxonomically precise techniques for identifying dietary components, such as DNA metabarcoding, could be a complementary approach for identifying consumed prey in finer detail (e.g. Galão, et al., Reference Galão, Soto, Nunes, Pedroso, Rocha and Rato2025).
As far as we are aware, Linosa hosts the highest cat density (89 cats/km2, the average density across the whole island, being high in the village and lower in the surrounding areas) recorded on an oceanic island. A similar density (73.4 cats/km2) has been recorded in the settled area of Corvo Island in the Azores Archipelago (Oppel et al., Reference Oppel, Beaven, Bolton, Vickery and Bodey2011). Such densities are similar to those in urban areas (Soto et al., Reference Soto, Nunes, Nóbrega, Palmeirim and Rocha2023). As cats on Linosa are subsidized by human-provided food, their high population is maintained by anthropogenic resources rather than the availability of local prey (Sims et al., Reference Sims, Evans, Newson, Tratalos and Gaston2008). Consequently, even if per capita consumption of native arthropods and reptiles appears low, the cumulative predation pressure resulting from such high cat densities could still severely affect these endemic species.
The consumption of wild prey, particularly passerines and Scopoli’s shearwaters, varied significantly between unneutered and neutered cats, with unneutered cats exhibiting a higher per cent of wild prey in their diets. Although both groups had equal outdoor access, this difference may stem from behavioural variations linked to neutered status, such as reduced home range and lower activity levels in neutered cats (Ferreira et al., Reference Ferreira, Machado, Nakano-Oliveira, Andriolo and Genaro2020), and association with humans. Additionally, dietary quality may influence consumption of wild prey. Cats are obligate carnivores requiring protein-rich diets with essential amino acids, and poor-quality diets, such as the pasta provided by people, may compel unneutered cats to hunt more frequently to compensate for nutritional deficiencies (Macdonald & Rogers, Reference Macdonald and Rogers1984; Cecchetti et al., Reference Cecchetti, Crowley, Goodwin and McDonald2021b). Protein quality, determined by digestibility and the abundance and bioavailability of amino acids, is generally higher in animal sources, whereas plant proteins (e.g. in grain, corn, soybean) have lower digestibility (Kakanubo et al., Reference Kanakubo, Fascetti and Larsen2015), reduced bioavailability (Zafalon et al., Reference Zafalon, Risolia, Vendramini, Rodrigues, Pedrinelli and Teixeira2020), and a less complete amino acid profile (Donadelli et al., Reference Donadelli, Jones and Beyer2019). Pasta provides poor-quality protein and is dominated by carbohydrates, a macronutrient that does not meet feline nutritional requirements and should represent only c. 12% of their target intake (Hewson-Hughes et al., Reference Hewson-Hughes, Hewson-Hughes, Miller, Hall, Simpson and Raubenheimer2011).
Although neutered cats consumed less wild prey than unneutered individuals, stable isotope data reflect diet rather than the total number of prey killed. Nonetheless, any reduction in wildlife consumption should be considered ecologically beneficial. However, dietary differences may not result directly from neutering but instead reflect a capture bias, as sociable cats are easier to capture and sterilize (although less sociable individuals were also trapped).
Management of the cat population, and cat husbandry, are key in this system. Given the legal protection of cats, management strategies on Linosa have to rely exclusively on sterilization or non-lethal removal through adoption. Effective trap–neuter–return programmes should be more frequent and more intensive (Cecchetti & Nelli, Reference Cecchetti and Nelli2025), supported by policies such as mandatory neutering and restrictions on introductions of new cats from the mainland. Introduction or enforcement of laws without first gaining community acceptance and increasing public awareness of issues caused by free-roaming cats would likely result in limited compliance (Crowley et al., Reference Crowley, Hinchliffe and McDonald2017). Therefore, we recommend implementation of structured public campaigns to encourage active community participation and support. The role of irresponsible ownership and lack of participation in sterilization campaigns is recognized as a driver of increasing cat populations (Zito et al., Reference Zito, Vankan, Bennett, Paterson and Phillips2015). On Linosa, limited engagement by some caregivers and misconceptions about neutering have probably contributed to the maintenance of the high cat density, highlighting the importance of addressing human behaviours alongside other management efforts.
A limiting factor in the effectiveness of trap–neuter–return programmes is that neutered free-roaming cats have reduced morbidity and improved body condition, favouring cat welfare and increasing cat survival (Gunther et al., Reference Gunther, Raz and Klement2018, Reference Gunther, Hawlena, Azriel, Gibor, Berke and Klement2022). To achieve a faster reduction of the population over time, sterilizations could be coupled with the relocation of socialized kittens and adults (Gunther et al., Reference Gunther, Hawlena, Azriel, Gibor, Berke and Klement2022) to the mainland.
Our recent population modelling for Linosa (Cecchetti & Nelli, Reference Cecchetti and Nelli2025) has further shown that, even under consistent neutering, an initial increase in cat numbers is expected, largely as a result of improved survival among sterilized individuals, before the population stabilizes and begins to decline. The model suggests that only high-intensity, female-focused trap–neuter–return over c. 5 years achieves meaningful population reductions. In contrast, low-intensity or irregular campaigns risk temporary declines followed by rebounds. Recognizing these dynamics is critical for setting realistic expectations and designing effective long-term management strategies.
Permanent confinement or strategic space–time restrictions, although effective in eliminating or reducing predation of wildlife, are impractical given local reliance on cats for pest control and limited public awareness of their ecological impacts. Therefore, neutering could be coupled with improving the quality of the provided cat food (Cecchetti et al., Reference Cecchetti, Crowley, Goodwin and McDonald2021b) and equipping sociable cats with effective deterrents such as a BirdBeSafe collar during the passerine migratory peak. These collars disrupt the cat’s camouflage and have proven effective in reducing killing of birds (Pemberton & Ruxton, Reference Pemberton and Ruxton2019; Cecchetti, et al., Reference Cecchetti, Crowley, Goodwin and McDonald2021b), and of prey with good colour vision, including herpetofauna (Hall et al., Reference Hall, Fontaine, Bryant and Calver2015). However, this would not be effective in reducing predation of Scopoli’s shearwaters as cats mainly prey on chicks inside burrows, and adults return to the colony at night, or for cats hunting at night. Future studies could integrate data on the seasonal richness and abundance of prey across taxonomic groups to provide a more detailed understanding of how temporal fluctuations in prey availability influence cat predation patterns and associated impacts on the island ecosystem.
In conclusion, our study shows a free-roaming cat population living at extremely high density, sustained by the food provided by local residents and tourists. Neutering and feeding practices are key to reducing consumption of wildlife. However, it is essential to recognize that several additional factors may shape predatory behaviour, including the proximity of cats’ home ranges to wildlife habitats and their level of habituation to humans. Trap–neuter–return programmes appear to unintentionally select for cats already accustomed to human contact, potentially creating a misleading correlation between neutering and reduced predation rates. Addressing these confounding factors is vital for enhancing the design and efficacy of cat population management strategies.
Author contributions
Study design: MC, RAM, LO, LB; fieldwork: MC, with the help of GDO, GG, PS; data analysis: MC; writing: MC with input from RAM, GDO, LO.
Acknowledgements
MC was supported by the L’Oréal-UNESCO ‘for Women in Science’ award 2022. We thank all the cat caretakers and owners of Linosa; the Malati di Isolitudine allo Stadio Cronico birdwatchers for their insights and help; and Chris Mitchell for support in the stable isotope lab at the University of Exeter. Cat sterilization in 2022 was funded by Action Project Animal.
Competing interests
None.
Ethical standards
This study adhered to the Oryx ethical guidelines. The protocol for whisker plucking and clipping was approved by the Animal Ethics and Welfare Committee of the University of Turin (Protocol no. 0004124).
Data availability
The data that support the findings of this study are not publicly available, because the information could compromise the privacy of cat owners/caretakers, but are available from the corresponding author upon reasonable request.





 Open access
Open access