


Non-technical Summary
Rocks in NW Scotland host abundant, well-preserved fossils of organisms that lived in a non-marine ecosystem one billion years ago. Our systematic investigation of these fossils reveals three new species including an unnamed form resembling a mycelial network, and Minimarmilla multicatenaria n. gen. n. sp., which may represent colonial cyanobacteria but also resembles some algae. These newly discovered microfossils shed new light on the diversification of complex life on Earth.
Introduction
Assemblages of organic-walled microfossils (OWM) preserved in sedimentary rocks provide a valuable archive of life in the Proterozoic, including the emergence and diversification of early eukaryotes (Butterfield, Reference Butterfield2015; Cohen and Macdonald, Reference Cohen and Macdonald2015; Agić and Cohen, Reference Agić and Cohen2021). Most of these assemblages are reported either from cherts or from shales, with a record reaching back to the late Paleoproterozoic (Miao et al., Reference Miao, Moczydłowska, Zhu and Zhu2019, Reference Miao, Yin, Knoll, Qu and Zhu2024; Riedman, et al., Reference Riedman, Porter, Lechte, dos Santos and Halverson2023). However, several Proterozoic phosphorites also host impressive Lagerstätten capturing a high diversity of eukaryotes compared to other Proterozoic assemblages (Muscente et al., Reference Muscente, Hawkins and Xiao2015). Most notably, the Ediacaran Weng’an biota from the lower Doushantuo Formation of South China (ca. 600–582 Ma) contains exceptionally preserved examples of complex organisms such as multicellular algae, tubular microfossils of putative metazoans, and possibly early-stage embryos (Xiao et al., Reference Xiao, Yuan and Knoll2000, Reference Xiao, Muscente, Chen, Zhou, Schiffbauer, Wood, Polys and Yuan2014). Evidence of multicellular algal thalli exceptionally preserved in phosphate, with some specimens even showing medulla–cortex thallus differentiation and individual cells, has also been found in the ca. 567–550-Ma Miaohe Member in China (Ye et al., Reference Ye, An, Yu, Zhou, Hu, Tong and Xiao2023). Thus, the fossil record hosted by Proterozoic phosphorites has proven to be of great value in understanding the fine-scale morphological traits of complex organisms, which can help to identify their affinities (e.g., Chen et al., Reference Chen, Xiao, Pang, Zhou and Yuan2014; Yin et al., Reference Yin, Zhu, Davidson, Bottjer, Zhao and Tafforeau2015).
The phosphatic lenses and laminae of the Torridon Group of NW Scotland are much older than these Ediacaran deposits and yield a less diverse assemblage, which is easily examined in petrographic thin sections (the phosphatic matrix is transparent and fine-grained). These fossils were first described early in the twentieth century (Geikie, Reference Geikie1900; Teall, Reference Teall and Geikie1907) and later examined more extensively by Strother et al. (Reference Strother, Battison, Brasier and Wellman2011) and by Battison and Brasier (Reference Battison and Brasier2012). An overlapping but distinct assemblage of OWMs has been obtained by the acid-maceration of the shale beds in the Torridon Group (Strother et al., Reference Strother, Battison, Brasier and Wellman2011; Strother and Wellman, Reference Strother and Wellman2016).
These fossils are paleontologically important for at least three reasons. Firstly, at ca.1 Ga they record the Mesoproterozoic–Neoproterozoic transition, which coincided with major innovations in the evolution of early eukaryotes, including the radiation of crown-group lineages, as indicated by phylogenetic studies (Chernikova et al., Reference Chernikova, Motamedi, Csürös, Koonin and Rogozin2011; Eme et al., Reference Eme, Sharpe, Brown and Roger2014), molecular-clock analyses (Parfrey et al., Reference Parfrey, Lahr, Knoll and Katz2011; Betts et al., Reference Betts, Puttick, Clark, Williams, Donoghue and Pisani2018), biomarkers (Brocks et al., Reference Brocks, Nettersheim, Adam, Schaeffer, Jarrett, Güneli, Liyanage, Van Maldegem, Hallmann and Hope2023), and unequivocal fossil eukaryotes (Butterfield, Reference Butterfield2000; Porter and Knoll, Reference Porter and Knoll2000; Loron et al., Reference Loron, Rainbird, Turner, Greenman and Javaux2019a; Tang et al., Reference Tang, Pang, Yuan and Xiao2020). Secondly, the Torridon microfossils include both prokaryotes and eukaryotes in an exceptional state of preservation, retaining sub-micron details and cellular structures in three dimensions (Strother et al., Reference Strother, Battison, Brasier and Wellman2011; Battison and Brasier, Reference Battison and Brasier2012; Wacey et al., Reference Wacey, Saunders, Roberts, Menon, Green, Kong, Culwick, Strother and Brasier2014). They include unique, evolutionarily important taxa such as Bicellum brasieri Strother and Wellman in Strother et al., Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021, which has been interpreted as a possible holozoan and is found nowhere else (Strother et al., Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021). Thirdly, these fossils represent an essentially non-marine aquatic environment. Proterozoic examples of such environments are rarely captured, and their structure and paleoecology remain poorly understood (Strother and Wellman, Reference Strother and Wellman2016; Nielson et al., Reference Nielson, Stüeken and Prave2024).
Despite this importance, the fossil assemblage hosted by the Torridon phosphates has not been systematically described in the published taxonomic literature, notwithstanding two important doctoral theses (Battison, Reference Battison2012; Sirantoine, Reference Sirantoine2021). We suggest that a formal systematic description, while always open to future amendment, is a necessary preamble for the assessment of the diversity and ecology of any fossil assemblage and for understanding the phylogenetic and ecological relationships of its constituent organisms, including any new or unique organisms. Systematic description includes commentary on the global occurrence of each taxon and thus places an assemblage in its penecontemporary global context.
Here we report an array of OWM from the phosphatic nodules and laminations of the lower Diabaig Formation of the Torridon Group, building on previous work (Battison and Brasier, Reference Battison and Brasier2012; Wacey et al., Reference Wacey, Saunders, Roberts, Menon, Green, Kong, Culwick, Strother and Brasier2014) while also reporting new taxa.
Geological setting
The Torridon Supergroup, informally designated “the Torridonian”, is an approximately 11 km-thick sequence of siliciclastic sedimentary rocks that comprises the Stoer, Sleat and Torridon groups (of which Stoer is the oldest and Torridon the youngest), which overlie the Archean–Paleoproterozoic Lewisian Gneiss in NW Scotland (Stewart, Reference Stewart1982, Reference Stewart2002). The Diabaig Formation is the lowermost unit of the Torridon Group (Figure 1), in which it is succeeded stratigraphically by the Applecross, Aultbea, and Cailleach Head formations (Stewart, Reference Stewart2002; Jones and Blake, Reference Jones and Blake2003). Although the Diabaig Formation consists mostly of red breccias, gray sandstones, and shales, its lower part hosts abundant laminae and thin lenses of whitish phosphorite (Stewart, Reference Stewart2002), which have been dated to 994 ± 48 Ma using the Rb–Sr system of radiometric dating (Turnbull et al., Reference Turnbull, Whitehouse and Moorbath1996). The succession is therefore interpreted as a latest Mesoproterozoic and/or earliest Neoproterozoic deposit (Kinnaird et al., Reference Kinnaird, Prave, Kirkland, Horstwood, Parrish and Batchelor2007).
Simplified regional geology. Sampling localities are arrowed (at this scale, the locality “Road cut above Lower Diabaig” is superimposed on Lower Diabaig). See SI Table 2 for GPS coordinates. Modified from Strother et al. (Reference Strother, Battison, Brasier and Wellman2011), Brasier et al. (Reference Brasier, Culwick, Battison, Callow and Brasier2017), and Wacey et al. (Reference Wacey, Brasier, Parnell, Culwick, Bowden, Spinks and Boyce2017).

The origin of the phosphatic horizons has been discussed by Battison and Brasier (Reference Battison and Brasier2012) and by Wacey et al. (Reference Wacey, Saunders, Roberts, Menon, Green, Kong, Culwick, Strother and Brasier2014). In their view, the phosphate ions were most likely sourced from the decay of organic matter, perhaps in an organic-rich ‘seston layer’ on the sediment–water interface. Phosphate mineralization then occurred in thin layers either just above or just below this interface, entombing large numbers of microorganisms and protecting their remains from subsequent decay. The co-precipitation of authigenic clay minerals appears to have enhanced the fidelity of morphological preservation (Wacey et al., Reference Wacey, Saunders, Roberts, Menon, Green, Kong, Culwick, Strother and Brasier2014).
The paleoenvironment of the Torridon succession has been debated since its discovery. The Diabaig Formation was long considered to represent a lacustrine depositional setting developed by constant infilling from rivers and streams in the surrounding valley systems; an absence of tidal and flow deposits was noted (see Brasier et al., Reference Brasier, Culwick, Battison, Callow and Brasier2017, and references therein). Recent sedimentological work (McMahon and Davies, Reference McMahon, Davies, Ghinassi, Colombera, Mountney, Reesink and Bateman2018, Reference McMahon and Davies2020) has largely upheld this view while also documenting several probable microbially induced sedimentary structures in the Diabaig shales. According to the alternative interpretation offered by Nielson et al. (Reference Nielson, Stüeken and Prave2024, p. 6), however, these rocks were deposited in a brackish estuarine/fluvial environment where “sedimentation occurred in mixed-energy, nearshore-marine settings fringed by fluvial networks.” Relevant features observed in outcrop include desiccation cracks, low-amplitude oscillation (wave) ripples, and ripple co-sets with bimodal flow directions indicative of tidal activity (Nielson et al., Reference Nielson, Stüeken and Prave2024). Geochemical proxies (e.g., Sr/Ba, Fe/Al) are consistent with the tidal/estuarine interpretation and imply that the Diabaig Formation was deposited under mostly freshwater, oxic conditions, with occasional seawater incursions (Nielson et al., Reference Nielson, Stüeken and Prave2024). Whether the Diabaig Formation represents an estuarine or a lacustrine deposit, its importance as a rare archive of a Proterozoic non-marine environment is undiminished. It should be added that the nature of lacustrine or estuarine deposits of this age is in general poorly understood and may differ in important ways from modern analogues (e.g., McMahon and Davies, Reference McMahon and Davies2020).
Materials and methods
Phosphatic horizons from the Diabaig Formation were sampled at eight localities by the authors in October 2022. Name conventions adopted for the localities were: Lower Diabaig (“LD"), a roadcut topographically above Lower Diabaig (“RCA”), a beach outcrop near a salmon farm at North Shieldaig (“SF”), a roadcut at Badachro (“BD”), and the beach at Badenscallie (“BC”). Sampling localities and horizons are shown on Figure 1. Their corresponding GPS coordinates and thin sections are listed in Supplementary Information (SI) Table 1.
Analysis of petrographic thin sections
Rock samples were processed into ~35 polished petrographic thin sections ~80 μm thick using a Logitech CS10 saw to cut the rocks, a Stuart Digital Hotplates to dry the samples, set the resin, and bond the samples to the glass slides, and a QATM Qpol 250 A1 machine for the final polishing at the Thin Section and Sample Preparation Facility in the School of GeoSciences, University of Edinburgh. Thin sections were studied using a Leica DM2700P optical microscope–camera system. Images were processed using the Leica Application Suite v. 4.0 and minimally enhanced with Adobe Photoshop v. 25.1. Measurements of fossil diameters were performed using both Leica Application Suite and the freeware ImageJ v. 1.53k. Final figures were made with Adobe Illustrator v. 27.8.
Morphometric analysis of Eohalothece
A morphometric analysis was performed to compare the cell shape of ellipsoidal unicellular forms in our thin sections and in the published literature. We used the free image processing software SHERPA (‘shape recognition, processing and analysis’), which was originally designed to identify and measure the outlines of diatom valves in brightfield microscopic images (Kloster et al., Reference Kloster, Kauer and Beszteri2014). In brief, SHERPA compares the outlines of specific objects with a set of templates of representative shapes by elliptical Fourier analysis. The software uses programming functions from the OpenCV 2.4.2 library, wrapped for the application platform .NET by Emgu CV 2.4.2 (OpenCV, 2012; Emgu, 2024). First, SHERPA detects the outline of a specific object applying noise reduction (Gaussian and median filters) and segmentation (Otsu’s method, histogram equalization plus Otsu’s method, Canny edge detector, or adaptive threshold), followed by shape detection and validation, contour extraction and optimization (“findContours” and “morphologyX” OpenCV functions), shape measurement (area, perimeter, width, and height), and finally template matching (elliptic Fourier descriptors).
We used a set of five templates created with Adobe Illustrator v. 27.8, representing a straight-sided “cylindrical” form, an “ellipsoidal” form, a smoothed “tapered” form, and two intermediate forms: “cylindrical tapered” and “ellipsoidal tapered”, all with a 2:1 aspect ratio. These templates were intended to capture reported morphological disparities between Eohalothece and Eosynechococcus cells. We tested 11 sets of cell outlines from 10 different studies reporting Eohalothece lacustrina Strother and Wellman, Reference Strother and Wellman2016, Eosynechococcus moorei Hofmann, Reference Hofmann1976, and Eosynecochoccus sp., including Hofmann (Reference Hofmann1976), Strother and Wellman (Reference Strother and Wellman2016), and Miao et al. (Reference Miao, Moczydłowska and Zhu2021). We traced each cell outline manually with Adobe Illustrator and saved them as individual images of 130 pixels in length for the analysis (Supplementary Information Figure 1). We followed the SHERPA workflow keeping by default the settings for noise reduction, segmentation, contour optimization, and validation parameters for contour because preliminary trials showed no difference in the results of our cell outlines when they were modified. Our templates showed convexity according to the software, so we selected the option “Use convexity” for the analysis. Final results (and templates) are shown in Supplementary Data 1 and SI Figure 2.
Comparison of Meso-Neoproterozoic organic-walled microfossils (OWM) assemblages using the data from SI Table 2. The Torridon Group presents 32 reported morphotaxa in total, with 8 eukaryotes, 11 prokaryotes, and 13 unknown specimens. This abundance is relatively high among Lagerstätte of comparable age, although surpassed by the Lower Shaler Supergroup, the Atar/El Mreïti Group, and the Mbuji-Mayi Supergroup with 53, 36, and 38 total OWM taxa reported, respectively. The present study adds the new taxon Minimarmilla multicatenaria n. gen. n. sp., and Unnamed species A and B microfossils (as shown in Figure 3). Abbreviations: Sg = supergroup, Gp = group, Fm = formation, Ma = million years.

List of organic-walled microfossils (OWM) from the Diabaig Formation (Torridon Group) described in this study, with their relative abundance, sizes, morphological attributes, and inferred domain. Morphotaxa classified as eukaryotes, Bicellum brasieri and Germinosphaera bispinosa, present at least two out of three morphology traits considered of eukaryotic origin in OWM (i.e., surface ornamentation, processes, and vesicle within a vesicle).

Repository and institutional abbreviation
Types, specimens, and other fossil material examined in this study are deposited in the Cockburn Geological Museum, Grant Institute, University of Edinburgh (UoE), Edinburgh, UK. Accession numbers are the same as the sample names shown in SI Table 1.
Systematic palaeontology
The International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code; ICBN) (Turland et al., Reference Turland, Wiersema, Barrie, Greuter and Hawksworth2018) was followed to describe the microfossils. All 11 taxa are presented and listed below in alphabetical order as ‘organic-walled microfossils’ (OWM), which is a more inclusive term for Precambrian microfossils of prokaryotic or eukaryotic origin than the artificial taxonomic identifier ‘acritarch’ (Colbath and Grenfell, Reference Colbath and Grenfell1995; Agić and Cohen, Reference Agić and Cohen2021). All illustrated specimens are identified in the figure captions by the locality and sample number as well as the England Finger coordinates indicated in SI Table 3.
Organic -walled microfossils
Genus Bicellum Strother and Wellman in Strother et al., Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021
Type species
Bicellum brasieri Strother and Wellman in Strother et al., Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021.
Remarks
This genus was erected by Strother and Wellman in Strother et al. (Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021) to accommodate specimens previously illustrated by Strother et al. (Reference Strother, Battison, Brasier and Wellman2011) and Wellman and Strother (Reference Wellman and Strother2015). Bicellum is described as a cell mass of spheroidal shape formed by two cell types found in the interior and exterior of the spheroid. Exterior cells forming the outer layer of about one cell thick are “elongate, sausage-shaped, thicker-walled, and tightly packed,” enclosing the interior cells that constitute a isodiametric mass of “thinner-walled cells closely packed in three dimensions.” and some interior cells may include a dark “organic bleb or spot” (Strother et al., Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021, p. 2659–2661).
Bicellum brasieri Strother and Wellman in Strother et al., Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021.
Photomicrographs of Bicellum brasieri specimens including different focal planes. (1, 5) Longitudinal view of a vesicle. (3, 4) Same specimen as (1) at different focal depths. (2, 6) Inner type-2 cells arranged inside the vesicle (arrows) with characteristic Y-shaped junction (circle). (7, 8) Same specimen as (5) at different focal depths. (9) Type-1 cells are here more easily resolved than the type-2 cells. (10–12) Same specimen as in (9) at different focal depths. (13) Specimen with very well-preserved type-1 cells in brick pattern. (14–16) Same specimen as (13) at different focal depths. Scale bars = (2, 6, 9, 13) 5 μm, (1, 5) 10 μm. (1–4) BC22-SH3-A. F26.4; (5–8) SF22-ENP. H18.1; (9–12) SF22-ENP. G35.3; (12–16) SF22-ENP. R8.2.

Reference Strother, Battison, Brasier and Wellman2011 “Spherical ball of cells enclosed within a complex wall”, Strother et al., p. 506, fig. 1h, suppl. info. p. 5, fig. 6.
Reference Wellman and Strother2015 “Unnamed multicellular ball of cells surrounded by a differentiated cell layer”, Wellman and Strother, p. 616, fig. 3G.
Reference Strother and Wellman2021 Bicellum brasieri Strother and Wellman in Strother et al., p. 2660, fig. 2–4, suppl. figs. S1, S2, S3.
Holotype
Specimen A in Strother and Wellman in Strother et al. (Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021, fig. 2).
Occurrence
Early Neoproterozoic Diabaig Formation, Scotland (Strother et al., Reference Strother, Battison, Brasier and Wellman2011, Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021; Wellman and Strother, Reference Wellman and Strother2015; this study).
Description
The type-1 cell-like vesicles are 0.9–1.6 μm in width (mean 1.1 μm) and 2.7–5.7 μm in length (mean 4 μm) (N = 20). Type-2 cell-like vesicles are 1.1–1.8 μm in width (mean 1.4 μm) and 1–2.4 μm in length (mean 1.8 μm) (N = 12). The diameter of the whole cluster (or main vesicle) is 15.5–20.7 μm in width (mean 17.6 μm) and 19.8–27.2 μm in length (mean 22.9 μm) (N = 5).
Materials
Seven specimens from Badenscallie and Salmon Farm localities in the Diabaig Formation (samples BC22-SH3-A, BC22-SH3-B, SF22-ENP).
Remarks
The exterior and interior cells found in our specimens correspond to those described by Strother and Wellman in Strother et al. (Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021), and we refer to them as type-1 and type-2 cells, respectively, in this study. Some specimens reported by Strother and colleagues (Strother et al., Reference Strother, Battison, Brasier and Wellman2011, Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021; Wellman and Strother, Reference Wellman and Strother2015) have a dark spot (“bleb”) in some type-2 vesicles. Such spots may represent condensed intracellular contents; similar organic inclusions are common in Proterozoic microfossils (Pang et al., Reference Pang, Tang, Wu, Li, Chen, Wan, Yuan, Bodnar and Xiao2020; Sforna et al., Reference Sforna, Loron, Demoulin, François and Cornet2022). Nevertheless, our specimens do not clearly show these inclusions. Similarly, the paucity of type-2 cells in some of our specimens may result from post-mortem degradation.
Strother et al. (Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021) reported two different types of cell-aggregate of this taxon: a ‘mature stage’ containing both type-1 (elongated, forming the outer layer) and type-2 (compacted and isodiametric, found in the interior) cells, and an ‘immature stage’ containing only type-2 cells. Although the formation dynamics of this multicellular structure are not yet determined, Strother et al. (Reference Strother, Brasier, Wacey, Timpe, Saunders and Wellman2021, p. 2663) regarded Bicellum as the “product of cell aggregation or cellularization of either a syncytium or coenobium.” They argued that the overall morphology (and lack of a rigid cell wall) supports a holozoan rather than algal or cyanobacterial affinity, and drew particular comparisons with early-branching holozoan groups such as Ichthyosporea and Corallochytrea (unicellular protists displaying a multicellular stage during their life cycle). According to those descriptions, our specimens comprise only the mature stage of B. brasieri (Figure 4) because they show a thick layer of peripheral type-1 cells surrounding a cluster of type-2 cells. Although the holozoan affinity remains uncertain, it is not incompatible with molecular clock estimates (Parfrey et al., Reference Parfrey, Lahr, Knoll and Katz2011; Eme et al., Reference Eme, Sharpe, Brown and Roger2014).
Genus Eohalothece Strother and Wellman, Reference Strother and Wellman2016
Type species
Eohalothece lacustrina Strother and Wellman, Reference Strother and Wellman2016.
Remarks
The genus Eohalothece was created by Strother and Wellman (Reference Strother and Wellman2016) for ellipsoidal to slightly fusiform cells found individually or in ‘colonies’, typically 5–7 μm in length and 3–4 μm in width. It was first described from the non-marine Nonesuch and Torridon deposits of ca. 1 Ga and interpreted as a cyanobacterium similar to the extant chroococcalean Aphanothece, and to the halotolerant Halothece–Euhalothece complex, which likewise exhibit a tapered to sub-elliptical cell morphology. Although the gross cell shape of Eohalothece clearly differs from some taxa that Strother and Wellman (Reference Strother and Wellman2016) mentioned by way of comparison, namely Palaeoanacystis (Schopf, Reference Schopf1968), Brachypleganon (Lo, Reference Lo1980), and Gloeotheceopsis (Zhang, Reference Zhang1988), it superficially resembles the genus Eosynechococcus (Hofmann, Reference Hofmann1976). Strother and Wellman (Reference Strother and Wellman2016) contended that Eohalothece cells represent a double-tapered, fusiform shape different from the “rod-like,” parallel-sided cells of Eosynechococcus. Our quantitative morphometric comparison between Eohalothece lacustrina and various Eosynechococcus morphospecies supports this distinction (analysis in Figure 6 and Supplementary Data 2). Eohalothece lacustrina cells previously from the Torridon, Nonesuch, and Xiamaling assemblages have aspect ratios of 1.5, 1.6, and 1.4, respectively, compared to Eosynechococcus moorei from the Belcher and Gouhou assemblages (2.1 and 2.3, respectively). New specimens measured for the present study also show an aspect ratio of 1.6. Elliptical Fourier analysis confirms consistent differences in shape between Eosynechococcus and Eohalothece, both in acid macerations and in thin sections (see Material and methods section, and Supplementary Data 1). Compared to Eosynechococcus, populations of Eohalothece lacustrina consistently contain more cells with strongly fusiform shape, and more cells with somewhat tapered elliptical or tapered cylindrical outlines (SI Figure 2). This analysis also shows that E. lacustrina reported by Miao et al. (Reference Miao, Moczydłowska and Zhu2021) from the putatively marine Xiamaling Formation indeed resembles E. lacustrina in the Torridon (both our specimens and those figured by Strother and Wellman, Reference Strother and Wellman2016). Thus, Eohalothece may be a cosmopolitan taxon not limited to non-marine settings. In any case, fossil taxa are properly defined by means of morphological distinctions between holotypes rather than extrinsic contextual factors (e.g., paleoenvironments) highly open to (re)interpretation.
Eohalothece lacustrina Strother and Wellman, Reference Strother and Wellman2016
Photomicrographs of Eohalothece lacustrina. (1, 8) Cells of various aspect ratios. (2–7) Individual cells with ellipsoidal to rounded shape. (9–11) Same dispersed cells at different focal depths within the phosphatic matrix. (12–15) Large clusters of E. lacustrina; (15) is a magnified region of (14). Organic material surrounds the clusters, and some Leiosphaeridia sp. can be found within the same area. Scale bars = (2–7; scale bar is in 5) 5 μm, (1, 8, 9) 20 μm, (12, 13, 15) 50 μm, (14) 100 μm. (1, 8–11) LDR3-D. M37.1; (2–7) LDR3-A. N7.1; (12, 13) LDR3-A. R6.4; (14, 15) LDR3-A. R9.4.

Cell sizes (length and width) of Eohalothece lacustrina in the Diabaig Formation compared to other similar taxa. Rectangles indicate the range of previously described E. lacustrina and E. moorei from multiple localities (including type localities), and E. amadeus Knoll and Golubic, Reference Knoll and Golubic1979, E. isolatus McMenamin et al., Reference McMenamin, Kumar and Awramik1983, E. thuleënsis Strother et al., Reference Strother, Knoll and Barghoorn1983, E. medius Hoffman, 1976, and E. grandis Hoffman, 1976, from their type localities. Reference column corresponds to the following publications: (1) Knoll and Golubic, Reference Knoll and Golubic1979; (2) McMenamin et al., Reference McMenamin, Kumar and Awramik1983; (3) Tang et al., Reference Tang, Pang, Yuan, Wan and Xiao2015; (4) Hoffman, 1976; (5) Strother et al., Reference Strother, Knoll and Barghoorn1983; (6) Miao et al., Reference Miao, Moczydłowska and Zhu2021; (7) Strother and Wellman, Reference Strother and Wellman2016.

Reference Strother and Wellman2016 Eohalothece lacustrina Strother and Wellman, p. 93, figs. 7A–G, 8.
Reference Strother and Wellman2021 Eohalothece lacustrinus [sic] Strother and Wellman, p. 8, fig. 5i.
Reference Miao, Moczydłowska and Zhu2021 Eohalothece lacustrina; Miao et al., p. 9, fig. 7c–e, h.
Reference Slotznick, Swanson-Hysell, Zhang, Clayton, Wellman, Tosca and Strother2023 Eohalothece lacustrina; Slotznick et al., fig. 8H.
Holotype
Specimen from thin section TS09-1 in Strother and Wellman (Reference Strother and Wellman2016, fig. 2A).
Occurrence
Xiamaling Formation (Miao et al., Reference Miao, Moczydłowska and Zhu2021) in China, the Nonesuch Formation in the USA (Strother and Wellman, Reference Strother and Wellman2016), the Diabaig Formation (Strother and Wellman, Reference Strother and Wellman2016; this study), and the Kinloch Formation (Strother and Wellman, Reference Strother and Wellman2016) in Scotland.
Description
Ellipsoidal vesicles occurring individually or in groups, which may exceed 200 units. Other fossils including Leiosphaeridia spp. and L. ternata (Timofeev, Reference Timofeev1966), can be found among the units of E. lacustrina, as well as amorphous organic material. The groups typically span several hundred μm in size. Individual cells are 2–4.8 μm in width (mean 3.5 μm) and 3.6–8.3 μm in length (mean 5.7 μm) (N = 120).
Materials
Three loose clusters of more than 100 individuals from the Diabaig Formation sampled at the Lower Diabaig locality (samples LDR3-A, LDR3-D).
Remarks
We identified the fossils described here as Eohalothece lacustrina, the same ‘colonial’ taxon erected by Strother and Wellman (Reference Strother and Wellman2016), and common to the Torridon Group, Nonesuch Formation, and the Xiamaling Formation (Miao et al., Reference Miao, Moczydłowska and Zhu2021). Our specimens are found in large groups and share similar cell shape to E. lacustrina (Strother and Wellman, Reference Strother and Wellman2016; Fig. 7A–D).
Genus Germinosphaera Mikhailova, Reference Mikhailova and Sokolov1986, emend. Butterfield, Knoll, and Swett, Reference Butterfield, Knoll, Swett and Bengston1994
Photomicrographs of Germinosphaera bispinosa and Leiosphaeridia spp. (1–4) Germinosphaera showing variable preservation of cell wall. (5–19) Leiosphaeridia of various sizes, some showing split or compromised cell wall. (20–22) Leiosphaeridia ternata. Scale bars = (3, 4, 6–18, 20–22) 10 μm, (1, 2, 5) 20 μm, (19) 50 μm. (1) BC22-SH3-B. S64.1; (2) SF22-ENP. N21.1; (3) BC22-SH3-B. K6.3; (4) SF22-ENP. Q19.4; (5) BC22-SH3-B. R8.4; (6) BC-ACH. G21.1; (7) BC22-SH3-A. X21; (8) BC-ACH. J30.2; (9) BC22-SH3-A. O23.4; (10) SF22-ENP. J20; (11) BC22-SH3-B. L5.4; (12) SF22-ENP. F11; (13) SF22-ENP. D37.3; (14) RCA-1.J28.3; (15) BC-ACH. H27. 3; (16) BC-ACH. N28.3; (17) BC22-SH3-A. M22; (18) SF22-ENP. Q16.3; (19) LDR3-A. A63; (20) LDR3-A. R13. 2; (21) LDR3-A. R12. 4; (22) LDR3-A. R13. 2.

Type species
Germinosphaera bispinosa Mikhailova, Reference Mikhailova and Sokolov1986, p. 33.
Remarks
Germinosphaera was erected to accommodate spheroidal vesicles bearing one or more tubular processes that open into the interior of the vesicle. These processes may commonly be distributed around the equatorial plane, but some non-equatorial attachments have been reported (e.g., Loron et al., Reference Loron, Rainbird, Turner, Greenman and Javaux2019a, fig. 8E). Mikhailova (Reference Mikhailova and Sokolov1986) originally named and distinguished two species by their number of processes: G. unispinosa and G. bispinosa. Butterfield et al. (Reference Butterfield, Knoll, Swett and Bengston1994) suggested that the variation in the number of processes was intraspecific and reported specimens with one to four equatorial processes; thus, the two genera of Mikhailova were unified into G. bispinosa (by name priority). Butterfield et al. (Reference Butterfield, Knoll, Swett and Bengston1994) also introduced G. jankauskasii Butterfield, Knoll, and Swett, Reference Butterfield, Knoll, Swett and Bengston1994, and G. fibrilla Butterfield, Knoll, and Swett, Reference Butterfield, Knoll, Swett and Bengston1994, based on material from the Svanbergfjellet Formation of Spitsbergen. A new species, G. alveolata, was erected by Miao et al. (Reference Miao, Moczydłowska, Zhu and Zhu2019) because of wall surface structures originally described as alveoli. Loron et al. (Reference Loron, Halverson, Rainbird, Skulski, Turner and Javaux2021) used scanning electron microscopy to show that these ornaments are made of little overlapping platelets (rather than alveoli) and emended the species accordingly.
The presence of processes and surface ornamentation suggests a eukaryotic origin for at least some Germinosphaera (Butterfield, Reference Butterfield2015; Loron et al., Reference Loron, Halverson, Rainbird, Skulski, Turner and Javaux2021). Although both prokaryotes and eukaryotes can generate protrusions (cilia in eukaryotes, and flagella in prokaryotes), they differ in composition and size (Rizzotti, Reference Rizzotti, Rizzotti and Evolution2000). Some examples of prokaryotes with long filamentous branching protrusions are known from the Planctomycetes–Verrucomicrobia–Chlamydiae (PVC) bacteria and from a species of Asgard archaeon (Imachi et al., Reference Imachi, Nobu, Nakahara, Morono and Ogawara2020). However, Germinosphaera is orders of magnitude larger in size. Also, prokaryotic ornaments and processes are usually proteinic in structure and prone to rapid degradation, and therefore unlikely to preserve as fossils (Javaux et al., Reference Javaux, Knoll and Walter2003).
Germinosphaera bispinosa Mikhailova, Reference Mikhailova and Sokolov1986, emend. Butterfield, Knoll and Swett, Reference Butterfield, Knoll, Swett and Bengston1994.
Reference Timofeev, Hermann and Mikhailova1976 Phycomycetes Timofeev et al., pl. VII, fig. 8.
Reference Mikhailova and Sokolov1986 Germinosphaera bispinosa Mikhailova, p. 36, fig. 6.
Reference Mikhailova and Sokolov1986 Germinosphaera unispinosa Mikhailova, p. 36, fig. 5.
Reference Jankauskas, Mikhailova and German1989 Germinosphaera bispinosa; Mikhailova in Jankauskas et al., p. 142, pl. XLVII, fig. 2.
Reference Jankauskas, Mikhailova and German1989 Germinosphaera tadasii Weiss in Jankauskas et al., p. 143, pl. XLVII, figs. 3–5.
Reference Jankauskas, Mikhailova and German1989 Germinosphaera unispinosa; Mikhailova in Jankauskas et al., p. 143, pl. XLVII, fig. 1.
Reference Knoll, Swett and Mark1991 Germinosphaera sp. Knoll et al., p. 557, fig. 19.6.
Reference Butterfield, Knoll, Swett and Bengston1994 Germinosphaera bispinosa; Butterfield et al., p. 39, fig. 16D, E.
Reference Zang1995 Germinosphaera sp. cf. G. unispinosa; Zang, p. 164, fig. 26K, L.
Reference Yin and and Guan1999 Germinosphaera unispinosa; Yin and Guan, p. 130, fig. 5.2, 5.4, 5.6, 5.9.
Reference Prasad and Asher2001 Germinosphaera unispinosa; Prasad and Asher, p. 121, pl. 13, fig. 11; p. 117, pl. 11, fig. 9.
Reference Prasad, Uniyal and Asher2005 Germinosphaera bispinosa; Prasad et al., p. 44, 46, pl. 11, fig. 3.
Reference Prasad, Uniyal and Asher2005 Germinosphaera unispinosa; Prasad et al., p. 44, pl. 11, fig. 2.
Reference Yin and Yuan2007 Germinosphaera unispinosa; Yin and Yuan, p. 353, fig. 2.11.
Reference Vorob’eva, Sergeev and Knoll2009 Germinosphaera sp. Vorob’eva et al., p. 189, fig. 13.13, 3.15, 13.17.
Reference Strother, Battison, Brasier and Wellman2011 Germinosphaera sp. Strother et al., suppl. info. p. 6, fig. 7d.
Reference Battison and Brasier2012 “Dark walled coccoid microfossils with broad tubular neck-like structure attached” Battison and Brasier, p. 213, fig. 8d.
Reference Loron and Moczydłowska2018 Germinosphaera bispinosa; Loron and Moczydłowska, p. 243, pl. 1, fig. 3.
Reference Loron, Rainbird, Turner, Greenman and Javaux2019a Germinosphaera bispinosa; Loron et al., p. 358, fig. 8E, F.
Reference Miao, Moczydłowska, Zhu and Zhu2019 Germinosphaera bispinosa; Miao et al., p. 187, fig. 5b–f.
Reference Loron, Halverson, Rainbird, Skulski, Turner and Javaux2021 Germinosphaera bispinosa; Loron et al., p. 1123, fig. 4.4, 4.5.
Reference Miao, Moczydłowska and Zhu2021 Germinosphaera bispinosa; Miao et al., p. 14, fig. 5d, 5e.
Reference Li, Luo, Tang, She and Xiao2023 Germinosphaera bispinosa; Li et al., p. 6, figs. 6a–d.
Reference Zheng, Tang, Xiao, Pang, Kang, Li, Zhou, Yuan and Wan2023 Germinosphaera bispinosa; Zheng et al., p. 7, fig. 5A–C.
Reference Wallet, Slater and Willman2023 Germinosphaera bispinosa; Wallet et al., p. 6, pl. 1, figs. 2–5.
Reference Riedman, Porter, Lechte, dos Santos and Halverson2023 Germinosphaera bispinosa; Riedman et al., p. 24, fig. 4F.
Holotype
Specimen number 882/2 in Mikhailova (Reference Mikhailova and Sokolov1986, fig. 6).
Occurrence
Mesoproterozoic to early Cambrian assemblages worldwide such as the Dashkin and Kamovsk formations, Krasnoyarsk region (Mikhailova, Reference Mikhailova and Sokolov1986); Omakhta and Neryuen formations, Uchur-Maya region (Jankauskas et al., Reference Jankauskas, Mikhailova and German1989); Miroedikha Formation, Turukhansk region (Timofeev et al., Reference Timofeev, Hermann and Mikhailova1976); Vychegda Formation, Timan Ridge (Vorob’eva et al., Reference Vorob’eva, Sergeev and Knoll2009) in the Russian Federation; the Draken Conglomerate Formation (Knoll et al., Reference Knoll, Swett and Mark1991) and Svanbergfjellet Formation (Butterfield et al., Reference Butterfield, Knoll, Swett and Bengston1994) in Svalbard; the Officer Basin (Zang, Reference Zang1995), and Limbunya Group, Birrindudu Basin (Riedman et al., Reference Riedman, Porter, Lechte, dos Santos and Halverson2023) in Australia; the Dongjia Formation (Yin and Guan, Reference Yin and and Guan1999; Yin and Yuan, Reference Yin and Yuan2007), Chuanlinggou Formation (Miao et al., Reference Miao, Moczydłowska, Zhu and Zhu2019), Xiamaling Formation (Miao et al., Reference Miao, Moczydłowska and Zhu2021), Diaoyutai Formation (Li et al., Reference Li, Luo, Tang, She and Xiao2023), and Qinggouzi Formation (Zheng et al., Reference Zheng, Tang, Xiao, Pang, Kang, Li, Zhou, Yuan and Wan2023) in China; the Sirbu Shale, Bhander Group (Prasad et al., Reference Prasad, Uniyal and Asher2005), and Avadh Formation, Ganga Basin (Prasad and Asher Reference Prasad and Asher2001) in India; the Visingsö Group (Loron and Moczydłowska, Reference Loron and Moczydłowska2018) in Sweden; the Mbuji-Mayi Supergroup (Baludikay et al., Reference Baludikay, Storme, François, Baudet and Javaux2016) in Democratic Republic of Congo; the Shaler Supergroup (Loron et al., Reference Loron, Rainbird, Turner, Greenman and Javaux2019a), and Dismal Lakes Group (Loron et al., Reference Loron, Halverson, Rainbird, Skulski, Turner and Javaux2021) in Canada; the Buen Formation (Wallet et al., Reference Wallet, Slater and Willman2023) in Greenland; and the Diabaig Formation, Torridon Group (Strother et al., Reference Strother, Battison, Brasier and Wellman2011; Battison and Brasier, Reference Battison and Brasier2012; this study) in Scotland.
Description
Spheroidal vesicle 19.5–51.0 μm in width (mean 32.6 μm), and a single unbranched process 2–9 μm in width (mean 5 μm) (N = 4) that opens into the interior of the vesicle.
Materials
Six specimens from the Lower Diabaig, Badenscallie, and Salmon Farm localities in the Diabaig Formation (samples LDR3-A, BC22-SH3-B, SF22-ENP).
Remarks
In some specimens the surface appears textured but no consistent pattern suggestive of original surface ornamentation is observed. Such a texture likely results from taphonomic degradation, mechanical compression, and mineral growth.
Genus Leiosphaeridia Eisenack, Reference Eisenack1958, emend. Downie and Sarjeant, Reference Downie and Sarjeant1963, emend. Turner, Reference Turner1984
Type species
Leiosphaeridia baltica Eisenack, Reference Eisenack1958.
Remarks
Leiosphaeridia comprises morphologically simple, smooth, spheroidal organic-walled microfossils of varying sizes and wall thicknesses. Because of this simple general morphology, morphospecies of Leiosphaeridia have been compared with various extant groups and this genus is likely to be polyphyletic (e.g., Javaux et al., Reference Javaux, Knoll and Walter2004; Willman, Reference Willman2009). Following Jankauskas et al. (Reference Jankauskas, Mikhailova and German1989), Butterfield et al. (Reference Butterfield, Knoll, Swett and Bengston1994), and Javaux and Knoll (Reference Javaux and Knoll2017), the majority of Leiosphaeridia specimens can be artificially divided into morphospecies based on the vesicle diameter and wall thickness (estimated from opacity under transmission light microscopy and the pattern of taphonomic folds). These are: L. crassa (Naumova, Reference Naumova1949) Jankauskas et al., Reference Jankauskas, Mikhailova and German1989 (thick wall, < 70 μm in diameter); L. jacutica (Timofev, Reference Timofeev1966) Jankauskas et al., Reference Jankauskas, Mikhailova and German1989 (thick wall, ≥ 70 μm in diameter); L. minutissima (Naumova, Reference Naumova1949) Jankauskas et al., Reference Jankauskas, Mikhailova and German1989 (thin wall, < 70 μm in diameter) and L. tenuissima Eisenack, Reference Eisenack1958 (thin wall, ≥ 70 μm in diameter). Many specimens fall between these categories and are difficult to classify (Porter and Riedman, Reference Porter and Riedman2016). For example, studies of wall ultrastructure with transmission electron microscopy (TEM) show that wall thickness and opacity are not always correlated (Javaux and Knoll, Reference Javaux and Knoll2017). Here, we place all specimens within the broad category of species plurimae, L. spp., except for L. ternata (Figure 8.20–8.22), which is readily distinguishable by the radial breakage pattern at the periphery.
Photomicrographs of Minimarmilla multicatenaria n. gen. n. sp. (1–16) Strap-like bundles comprising filamentous strands of cell-like units. (2–4) Magnified regions of (1) showing arrangement of units. (5) Magnified region of (4) showing thicker units (red arrow) and thinner units (black arrow) in linear arrangements (white arrows). (6–8) Additional examples of bundles. (7) Holotype of M. multicatenaria n. gen. n. sp. (SF22.ENP.FG6). (9) Thick units (black arrow) and thin units (red arrow) in the same bundle. (10) Unusually small units (black arrow) near a cluster of more typical units. (11) Poorly preserved bundle showing taphonomic variability. (12, 13) Arrangement of bundles showing coherence of adjacent strands. (14) Magnified region of (13). (15, 16) Knob-like structure attached to a short filamentous section of M. multicatenaria n. gen. n. sp. and consisting of similar cell-like units (black arrow), shown at two focal depths. Scale bars = (2, 3, 9–11, 14–16) 25 μm, (5–8) 50 μm, (1, 4, 13) 100 μm, and (12) 200 μm. Images 2, 5, 6, 8, 9, 11 are montages. (1–3) SF22-ENP. M14.3; (4, 5) SF22-ENP. M14. 3; (6) SF22-ENP. N62. 3; (7) holotype, SF22.ENP.FG6; (8) SF22-ENP. Q58. 0; (9) SF22-ENP. R47.3; (10) SF22-ENP. H24. 4; (11) SF22-ENP. Q58.0; (12) SF22-ENP. M63. 1; (13–16) SF22.ENP. L17.

Leiosphaeridia ternata (Timofeev, Reference Timofeev1966), emend. Mikhailova and Jankauskas in Jankauskas et al., Reference Jankauskas, Mikhailova and German1989
Reference Timofeev1966 Turuchanica ternata Timofeev, p. 45, pl. 9, fig. 8.
Reference Jankauskas, Mikhailova and German1989 Leiosphaeridia ternata (Timofeev, Reference Timofeev1966) Jankauskas et al., pl. XI, figs 2–4; pl. XII, figs. 4, 5, 8.
Reference Hofmann and Jackson1994 Leiosphaeridia ternata; Hofmann and Jackson, p. 26, fig. 17.5–17.7.
Reference Zang and Walter1992a Leiosphaeridia ternata; Zang and Walter, p. 296, pl. XII, fig. F–I.
Reference Zang and Walter1992b Leiosphaeridia ternata; Zang and Walter, p. 68, pl. 53, fig. A–E.
Reference Cotter1997 Leiosphaeridia ternata; Cotter, p. 264, fig. 7J.
Reference Ragozina, Weis and Afonin2003 Leiosphaeridia ternata; Ragozina et al., p. 57, pl. II, fig. 1.
Reference Strother and Wellman2016 Leiosphaeridia ternata; Strother and Wellman, p. 101. fig. 7A, F, G.
Reference Sergeev, Knoll, Vorob’eva and Sergeeva2016 Leiosphaeridia ternata; Sergeev et al., p. 92, fig. 4.3, 4.4.
Reference Javaux and Knoll2017 Leiosphaeridia ternata; Javaux and Knoll, p. 208, fig. 4.9.
Reference Beghin, Storme, Blanpied, Gueneli, Brocks, Poulton and Javaux2017 Leiosphaeridia ternata; Beghin et al., p. 70, pl. II, fig. k.
Reference Loron and Moczydłowska2018 Leiosphaeridia ternata; Loron and Moczydłowska, p. 232, pl. II, figs. 1, 2.
Reference Loron, Rainbird, Turner, Greenman and Javaux2019a Leiosphaeridia ternate; Loron et al., p. 352, fig. 2E.
Reference Shukla, Sharma and Sergeev2020 Leiosphaeridia ternate [sic]; Shukla et al., p. 501, fig. 6h.
Reference Arvestål and Willman2020 Leiosphaeridia ternata; Arvestål and Willman, p. 12, fig. 6H.
Reference Miao, Moczydłowska and Zhu2021 Leiosphaeridia ternata; Miao et al., p. 15, fig. 3c.
Reference Strother and Wellman2021 Leiosphaeridia ternata; Strother and Wellman, p. 4, fig. 2e.
Reference Zheng, Tang, Xiao, Pang, Kang, Li, Zhou, Yuan and Wan2023 Leiosphaeridia ternata; Zheng et al., p. 14, fig. 11B.
Holotype
Specimen shown in Timofeev (Reference Timofeev1966, pl. 9, fig. 8).
Occurrence
Widely distributed in Paleoproterozoic to Ediacaran successions worldwide.
Description
Specimens are spheroidal vesicles 12.9–15.6 μm in diameter (mean 14 μm) (N = 4), with an opaque smooth wall showing a maximum of 5 radial cracks. The cracks divide the periphery into trapezoidal portions and are of various lengths, 11–30% of the vesicle radius.
Materials
Seventeen specimens from road cut above Lower Diabaig and Badenscallie localities in the Diabaig Formation (samples LDR3-A, LDR3-D, BC22.SH3.A, BC22.SH3.B).
Remarks
Leiosphaeridia ternata possess a rigid wall structure and characteristic split vesicle interpreted as a possible artefact of compression (e.g., Hofmann et al., Reference Hofmann and Jackson1994). However, it has been suggested to be an expression of the primary structure of the organism given that the internal, central spheroidal portion of the vesicle is outlined and not affected by the radial cracking of the periphery (Loron and Moczydłowska, Reference Loron and Moczydłowska2018). Leiosphaeridia ternata is distributed worldwide in Paleoproterozoic to Ediacaran successions deposited in marine conditions (see Loron and Moczydłowska, Reference Loron and Moczydłowska2018). Its occurrence in the ca. 1-Ga Nonesuch Formation (USA) and in the Torridon Group (Scotland) is notable since both localities have been interpreted to record alluvial, fluvial/estuarine, and lacustrine depositional environments (Strother and Wellman Reference Strother and Wellman2016; Stüeken et al., Reference Stüeken, Bellefroid, Prave, Asael, Planavsky and Lyons2017; Nielson et al., Reference Nielson, Stüeken and Prave2024). Loron and Moczydłowska (Reference Loron and Moczydłowska2018) speculated that the occurrence of L. ternata in these successions might be explained by marine incursions.
Leiosphaeridia spp.
Description
Spheroidal vesicles with a smooth-walled surface. Major axis in each specimen is 15–141 μm in length (mean 42 μm), and minor axis is 14–123 μm (mean 37 μm) (N = 12). Openings by partial rupture or median split are observed in some specimens.
Materials
Specimens (1875) from the five studied localities in the Diabaig Formation (samples LDR3-A, LDR3-D, BD22-210-A, BD22-210-B2, BC-ACH, BC22-SH3-A, BC22-SH3-B, SF22-ENP, BD22-210, RCA-1, RCA-2).
Remarks
One specimen reported here (Figure 7.10) displays a medial split opening, which could be interpreted as an excystment structure (the opening in a cyst wall whereby the contents may be liberated), suggesting the vesicle was a cyst (dormant or reproductive). Complex excystment openings, like pylomes, can be considered a eukaryotic feature (Javaux et al., Reference Javaux, Knoll and Walter2003; Butterfield, Reference Butterfield2015) but simpler examples, like medial splitting, are also found in cyanobacteria (Waterbury and Stanier, Reference Waterbury and Stanier1978; Javaux and Knoll, Reference Javaux and Knoll2017).
Genus Minimarmilla new genus
Type species
Minimarmilla multicatenaria n. gen. n. sp., Figure 8.7 in this study, by monotypy.
Diagnosis
As per the type species.
Etymology
Derived from the Latin minima, meaning small, and armilla, -ae, f., meaning bracelet, referring to the small size of the fossil and characteristic cell alignment resembling a bracelet of beads.
Remarks
This genus is created to place new multi-element fossil specimens from the Diabaig Formation. Their morphology (bundle of small cell-like units with no sheath) has not been reported previously from the Torridon Group and differs from other Precambrian bundles of filaments (e.g., Polythrichoides) and filamentous colonies (e.g., Chlorogloeaopsis, Polysphaeroides, Arctacellularia). Minimarmilla n. gen. cell-like units are rectangular carbonaceous compressions; care should be taken to distinguish them from superficially similar euhedral crystals of pyrite found in chains in some fossil assemblages, for example those associated with the sheathed filamentous taxa Siphonophycus, Palaeolyngbya, and Rugosoopsis (e.g., Willman and Slater, Reference Willman and Slater2021). For these reasons we erect a new genus: Minimarmilla.
Minimarmilla multicatenaria new genus new species
Holotype
Specimen illustrated in Figure 8.7, from the Diabaig Formation, Torridon Group (Salmon Farm locality), sample SF22-ENP.O62.2 is selected as holotype.
Diagnosis
Approximately rectangular cell-like units arranged serially in strands. The units within well-preserved strands are oriented with their long axis in parallel and are separated from each other by a distance shorter than the length of each unit. Individual units are commonly slightly thicker at the ends. Strands typically contain several tens of units and are themselves organized into parallel or subparallel bundles. Bundles typically contain fewer than ten strands but may be several hundred micrometers in length. Cell-like units in adjacent strands within a bundle do not consistently align with each other transversely. Some bundles appear to radiate from a common origin.
Occurrence
Early Neoproterozoic Diabaig Formation, Torridon Group, Scotland (this study).
Description
Cell-like units organized into filament-like strands grouped together in bundles. Cell-like units occur in several varieties. Many are rectangular, measuring 1.6–2.6 μm in width (mean 2 μm), and 2.6–3.7 μm in length (mean 3.3 μm), with a spacing of less than 1 μm between units (N = 10). Some bundles are composed exclusively of strands with much smaller units (i.e., about 1 μm in characteristic dimension with proportionately tighter spacing; Figure 8.10). Other units are less than 1 μm in width; these “thin” units occur in discrete regions of strands and bundles that also contain the other, thicker variety. Although most units are clearly aligned in strands and bundles, some are better organized than others and a few irregular clusters can also be found. No branching of either strands or bundles has been observed and there is no clear evidence of a sheath. One knob-like grouping of cell-like units may represent a discrete structure (Figure 8.15, 8.16).
Etymology
Derived from the Latin multus, meaning many, and catena, -ae, f., meaning chain, in reference to the multiplicity of strands and the resemblance of the units to links in a chain.
Materials
More than 50 well-preserved specimens (bundles) from the Salmon Farm locality, and one poorly preserved specimen from the Lower Diabaig locality in the Diabaig Formation (samples SF22-ENP, and LDR3-D).
Remarks
Minimarmilla multicatenaria n. gen. n. sp. differs from other bundles of filaments reported in the Proterozoic: Polythrichoides lineatus (Hermann, Reference Hermann and Timofeev1974) are smooth-walled, filamentous sheaths, similar to Siphonophycus but occasionally septate (Tang et al., Reference Tang, Pang, Yuan and Xiao2020, fig. 14A) and the unnamed filaments reported by Loron et al. (Reference Loron, Rainbird, Turner, Greenman and Javaux2019a, fig. 3O–Q) are micrometer-scale filaments with a parallel alignment in a single plane.
Minimarmilla multicatenaria n. gen. n. sp. also differs from other Precambrian trichomes in several ways. It is made of consistent rectangular cell units, unlike the spheroidal to sub-spheroidal cells in Chlorogloeaopsis spp. or the barrel-shaped, taphonomically folded cells of Arctacellularia tetragonala (Maithy, Reference Maithy1973) Hofmann and Jackson, Reference Hofmann and Jackson1994 (e.g., Baludikay et al., Reference Baludikay, Storme, François, Baudet and Javaux2016 ; Beghin et al., Reference Beghin, Storme, Blanpied, Gueneli, Brocks, Poulton and Javaux2017). It lacks the surrounding sheath characteristic of Palaeolyngbya spp. (e.g., Butterfield et al., Reference Butterfield, Knoll, Swett and Bengston1994) or clear evidence of branching seen in Polysphaeroides filiformis Hermann in Timofeev et al., Reference Timofeev, Hermann and Mikhailova1976 (e.g., Baludikay et al., Reference Baludikay, Storme, François, Baudet and Javaux2016). Furthermore, none of these taxa (except A. tetragonala, see Sforna et al., Reference Sforna, Loron, Demoulin, François and Cornet2022) displays a multi-element body-plan with differentiated structure, which we suggest is evident from the knob-like grouping of units seen in Figure 8.15 and 8.16. Because of its unique and consistent morphology, we erect a new genus and species to accommodate our specimens. The variety in the shape and thickness of cell-like units may be at least partly taphonomic, although it is notable that entire bundles occur with consistently smaller, more closely packed units (Figure 8.10). Affinities to cyanobacteria, red algae, and green algae are considered in the Discussion section.
Genus Pterospermopsimorpha Timofeev, Reference Timofeev1966 (Mikhailova in Jankauskas et al., Reference Jankauskas, Mikhailova and German1989)
Type species
Pterospermopsimorpha pileiformis Timofeev, Reference Timofeev1966.
Remarks
Pterospermopsimorpha is described as a disphaeromorph (a sphere enclosing another sphere), with a central body showing a thick wall and a sharply outlined vesicle-like surrounding envelope, which may show a psilate texture, as in P. insolita (Timofeev, Reference Timofeev1969) Mikhailova in Jankauskas et al., Reference Jankauskas, Mikhailova and German1989, or a granular texture (P. pileiformis); see Loron and Moczydłowska (Reference Loron and Moczydłowska2018). Sixteen taxa were placed originally within Pterospermopsimorpha (Fensome et al., Reference Fensome, Williams, Barss, Freeman and Hill1990; Jachowicz-Zdanowska, Reference Jachowicz-Zdanowska2013), but Loron and Moczydłowska (Reference Loron and Moczydłowska2018) synonymized and relocated some species within P. insolita because of their characteristic smooth vesicle surface. Opening through a medial split has been reported in some specimens (Loron et al., Reference Loron, Rainbird, Turner, Greenman and Javaux2019a, fig. 8C). The body-plan of Pterospermopsimorpha resembles a phycoma, a vegetative, decay-resistant, organic-walled stage cyst-like structure produced by extant prasinophycean green algae (Tappan, Reference Tappan1980; Moczydłowska, Reference Moczydłowska2016). Hence, this genus has been proposed to be a possible eukaryote (e.g., Tappan, Reference Tappan1980; Inouye et al., Reference Inouye, Hori and Chihara1990; Guy-Ohlson, Reference Guy-Ohlson, Jansonius and McGregor1996; Samuelsson et al., Reference Samuelsson, Dawes and Vidal1999; Moczydłowska et al., Reference Moczydłowska, Landing, Zang and Palacios2011; Agić et al., Reference Agić, Moczydłowska and Yin2017; Loron et al., Reference Loron, Rainbird, Turner, Greenman and Javaux2019a).
Pterospermopsimorpha sp.
Photomicrographs of Pterospermopsimorpha sp., and Synsphaeridium spp. (1) Pterospermopsimorpha showing an envelope enclosing an amorphous vesicle. (2, 3) Magnified views of the fossil shown in (1) at two focal depths. (4–12) Synsphaeridium in different arrangements: tightly packed clusters (4–6), loose clusters (7, 8), duos (9, 10), or accompanied by Leiosphaeridia (11, 12; same specimen at two focal depths). Scale bars = (1–3, 5, 6) 20 μm, (7–10) 10 μm, (4, 11, 12) 40 μm. (1–3) BC22-SH3-A. W23.2; (4) BC22-SH3-A. E5.4; (5) SF22-ENP-E2. H56.3; (6) SF22-ENP-E2. Q69.4; (7) BC22-SH3-B. S57.3-4; (8) SF22-ENP. R18; (9) BC22-SH3-B. K7.2; (10) SF22-FLO-2. Q53.4; (11, 12) BC22-SH3-A. Q. 23.

Description
Spheroidal vesicle with smooth to shagrinate wall encapsulating a smaller smooth-walled vesicle. The inner vesicle is transparent, with a dark brown color in the center, and an apparent small irregular elongation. The outer vesicle is light brown and shows pronounced folds along the perimeter. The inner vesicle is 40 μm in diameter, while the outer vesicle is 88 μm in diameter.
Materials
One well-preserved specimen from the Diabaig Formation, Badenscallie locality (sample BC22-SH3-A).
Remarks
Wacey et al. (Reference Wacey, Saunders, Roberts, Menon, Green, Kong, Culwick, Strother and Brasier2014, fig. 1a) illustrated an extracted specimen of Pterospermopsimorpha from an unspecified formation the Torridon Group similar to those reported here, although with an opaquer inner vesicle. The shagrinate appearance of some of the wall in the specimen reported here is likely taphonomic (possibly connected with compression by mineral grains) rather than a true ornamentation. The simple morphology of this taxon (a vesicle enclosed within another vesicle) does not usefully discriminate between a eukaryotic and a prokaryotic origin. We refer to this specimen only by the genus to avoid overinterpretation.
Agić et al. (Reference Agić, Moczydłowska and Yin2017, fig. 10K) and Miao et al. (Reference Miao, Moczydłowska and Zhu2021, fig. 4K) illustrated similar specimens but did not name them. Baludikay et al. (Reference Baludikay, Storme, François, Baudet and Javaux2016, fig. 7J, K) described a similar taxon as P. insolita, interpreted as “dividing specimens.” Similar remarks can be made about the specimen illustrated in Loron and Moczydłowska (Reference Loron and Moczydłowska2018, pl. 4, figs. 7–9) and interpreted by them as P. pileiformis. Our specimen lacks ornamentation, processes, and medial splitting so it cannot be unambiguously recognized as a crown-group eukaryote, in contrast with P. insolita (e.g., Tappan, Reference Tappan1980; Moczydłowska, Reference Moczydłowska2016; Loron et al., Reference Loron, Rainbird, Turner, Greenman and Javaux2019a).
Genus Siphonophycus Schopf, Reference Schopf1968
Type species
Siphonophycus kestron Schopf, Reference Schopf1968.
Remarks
Siphonophycus includes smooth-walled, unbranched, non-septate tubular filaments. The filaments traditionally have been classified into six morphotaxa based on the filament width (Butterfield et al., Reference Butterfield, Knoll, Swett and Bengston1994; Tang et al., Reference Tang, Pang, Xiao, Yuan, Ou and Wan2013). Like Leiosphaeridia, the genus Siphonophycus is a waste-basket taxon likely to be highly polyphyletic. Nevertheless, the tubular structures of Siphonophycus are often interpreted as the empty sheaths of mat-forming cyanobacteria (Butterfield et al., Reference Butterfield, Knoll, Swett and Bengston1994). Some larger specimens, such as Siphonophycus gigas Tang et al., Reference Tang, Pang, Xiao, Yuan, Ou and Wan2013, may be remains of eukaryotes, but size alone cannot be a conclusive criterion.
Siphonophycus spp.
Photomicrographs of Siphonophycus spp. This form consists of filamentous sheaths of various sizes, probably of (cyano)bacterial origin. Most specimens are fragmentary but in rare examples rounded tips can be seen at one or both ends (1). Dense populations of aligned filaments are interpreted as in-situ microbial mats (2). Some individual filaments are large (1, 3, 4), some are broken into segments (5), and some are long, thin, and tangled (6–12). Scale bars = (3, 4) 25 μm, (1, 5–9, 12) 50 μm, (2, 10, 11) 100 μm. (1) SF22-ENP-N P39.0; (2) SF22-ENP-E2. O29.0; (3) SF22-ENP. K18. 3; (4) RCA-1. N12.1; (5) RCA-1. J38.2; (6) RCA-1. O11; (7) RCA-1. S9. 2; (8) BC-ACH. C34; (9) BC22-SH3-B. J6. 3; (10) BC-ACH. E30. 1; (11) SF22-ENP. N21. 2; (12) BC22-SH3-B. Q19. 4.

Description
Smooth-walled tubular non-septate filaments. Of those counted here, 279 are between 1.5 μm and 2 μm in width, 746 are between 3 μm and 5 μm wide, and 403 are between 5 μm and 10 μm. Measurements of length would not be instructive since few if any filaments are complete.
Materials
Specimens (1428) from the from the five studied localities in the Diabaig Formation (Samples LDR3-A, LDR3-D, BD22-210-A, BD22-210-B2, BC-ACH, BC22-SH3-A, BC22-SH3-B, SF22-ENP, BD22-210, RCA-1, RCA-2).
Remarks
In the present material, Siphonophycus specimens can be sorted into the following artificial morphotaxa (by width): S. solidum (Golub, Reference Golub and Sokolov1979) Butterfield et al., Reference Butterfield, Knoll, Swett and Bengston1994 (16–32 μm); S. septatum (Schopf, Reference Schopf1968) Knoll et al., Reference Knoll, Swett and Mark1991 (1–2 μm); and S. robustum (Schopf, Reference Schopf1968) Knoll et al., Reference Knoll, Swett and Mark1991 (2–4 μm). We leave them here under open nomenclature as Siphonophycus spp. This taxon is present as dense mats of sub-parallel filaments at some localities (e.g., Fig 10.2).
Genus Synsphaeridium Eisenack, Reference Eisenack1965
Type species
Synsphaeridium gotlandicum Eisenack, Reference Eisenack1965.
Remarks
Synsphaeridium consists of clusters of spheroidal vesicles formed by 3–15 units arranged irregularly, tightly or loosely packed, and with an opaque coloration. Some of the vesicles show opaque spots inside. Different taxa, such as Synsphaeridium Eisenack (Reference Eisenack1965), Myxococcoides Schopf (Reference Schopf1968), and Symplassosphaeridium Timofeev, 1959 ex Timofeev (Reference Timofeev1969), have been established to classify aggregates of psilate vesicles based on vesicle size, number, and aggregation style (i.e., rounded or irregular aggregates) (Jankauskas et al., Reference Jankauskas, Mikhailova and German1989; Hofmann and Jackson, Reference Hofmann and Jackson1994). Synsphaeridium shares a particularly similar morphology with Symplassosphaeridium, although the latter has vesicles of smaller size, with more units forming the clusters, and a more compacted and rounded arrangement (Tang et al., Reference Tang, Pang, Yuan, Wan and Xiao2015; Riedman and Porter, Reference Riedman and Porter2016).
Synsphaeridium spp.
Description
Clusters of spheroidal vesicles irregularly packed of 7.7–16.9 μm in diameter (mean 13.9 μm; N = 10). Some vesicles show folds on the vesicle membrane. The clusters are 37–48 μm in size.
Materials
Twenty-seven specimens from the five studied localities in the Diabaig Formation (Samples LDR3-A, LDR3-D, BC22-SH3-B, SF22-ENP, BC-ACH, BD22-210, BD22-210-B2).
Remarks
The relatively simple morphology of Synsphaeridium, essentially a cluster of leiospheres, does not allow a precise taxonomic assignment. Here, we considered as Synsphaeridum all closely packed clusters of more than three spheroids. Similarly, some specimens reported here are more or less closely packed but still fit within the genus description. Because of this ambiguity we leave the specimens under open nomenclature.
Unnamed species A
Photomicrographs of unclassified taxa, Unnamed species A, and Unnamed species B. (1) Unnamed species A, a mass of entangled filaments. (2, 3) Magnified regions of (1) showing filamentous texture. (4–9) Unnamed species B showing mycelium-like structure. (4, 5) Mycelium and associated vesicles (5 is a magnified region of 4). (6–9) Networks of filaments with variable preservation. Scale bars = (2, 3; scale bar is in 2) 25 μm, (1, 5) 50 μm, (4) 100 μm, (6–9) 200 μm. (1–3) LDR3-D. X30. 4; (4, 5) BC22-SH3a. R6.1; (6, 7, 9) SF22-ENP-Y. R51. 4; (8) BC22-SH3-A. T2.

Description
Large ball of tightly intertwined filaments of less than 1 μm in width each, twisted into a whorl. Diameters of the balls are 188.85 and 91 μm.
Materials
Two specimens from the Lower Diabaig locality, Diabaig Formation (sample LDR3-D).
Remarks
Clusters of filaments are common in the Precambrian, either in bundle-like assemblages (e.g., Polytrichoides lineatus Hermann, Reference Hermann and Timofeev1974), parallel-aligned in one plane (Loron et al., Reference Loron, Rainbird, Turner, Greenman and Javaux2019a), or simply in microbial-mat associations (e.g., Butterfield et al., Reference Butterfield, Knoll, Swett and Bengston1994). It is possible that Unnamed species A represents the remains of spheroidal filamentous cyanobacteria colonies like those formed by extant species of Nostocales or Oscillatoriales (Brehm et al., Reference Brehm, Krumbein and Palińska2003; Milferstedt et al., Reference Milferstedt, Kuo-Dahab, Butler, Hamelin and Abouhend2017; Willman and Slater, Reference Willman and Slater2021), although the possibility of passive post-mortem roll-up is not discarded. ‘Donut-shaped’ Siphonophycus bundled filaments comparable to our fossil have been described from the Hailuoto Island and Saarijärvi meteorite-impact crater in Finland, interpreted as originally Nostoc-ball-like masses in a torus shape that flattened overtime (Willman and Slater, Reference Willman and Slater2021). The very small width of some filaments observed in our specimens and their possibly colonial habit are suggestive of cyanobacteria although size is not sufficient by itself for such a diagnosis (see Javaux et al., Reference Javaux, Knoll and Walter2003).
Unnamed species B
Description
Networks of filaments of 2.7–5.2 μm in width (mean 4.5 μm) (N = 10), apparently hollow with Y-shaped connections. Possibly associated with a spheroidal vesicle about 21.3 μm in diameter (Figure 11.5). The complete network is several hundred μm across.
Materials
Two specimens from the Diabaig Formation, Badenscallie and Salmon Farm localities (samples BC22-SH3-a, and SF22-ENP).
Remarks
Several other branched networks have been reported from the Precambrian (Gan et al., Reference Gan, Luo, Pang, Zhou, Zhou, Wan, Li, Yi, Czaja and Xiao2021; Bonneville et al., Reference Bonneville, Delpomdor, Préat, Chevalier, Araki, Kazemian, Steele, Schreiber, Wirth and Benning2020), some as old as the early Paleoproterozoic (Bengtson et al., Reference Bengtson, Rasmussen, Ivarsson, Muhling, Broman, Marone, Stampanoni and Bekker2017). The biogenicity and/or syngenicity of these specimens can be questioned (e.g., preservation in karstic cracks for the Gan et al., Reference Gan, Luo, Pang, Zhou, Zhou, Wan, Li, Yi, Czaja and Xiao2021, specimen, or three-dimensionally in shale in the case of Bonneville et al., Reference Bonneville, Delpomdor, Préat, Chevalier, Araki, Kazemian, Steele, Schreiber, Wirth and Benning2020, whereas shale-hosted microfossils are usually compressed into two dimensions). Here, fully encased in the matrix and partly compressed, the filaments are undoubtedly syngenetic and biogenic. However, their degree of branching and conformation in space are difficult to resolve in the relatively coarse, granular phosphate layer in which they occur as partly compressed fossils. A network morphology coupled with associated vesicles is reminiscent of mycelial networks of fungi. Mycelium-like organization is also found in actinobacteria such as Corynebacterium, Streptomyces, and Mycobacterium, although these would typically be much smaller than the fossils described here (Barka et al., Reference Barka, Vatsa, Sanchez, Gaveau-Vaillant, Jacquard, Klenk, Clément, Ouhdouch and Van Wezel2016). Other Precambrian crown fungal candidates have been reported (Loron et al., Reference Loron, François, Rainbird, Turner, Borensztajn and Javaux2019b) but their affinities remain uncertain as molecular clock-based models suggest that filamentous and arbuscular fungi should be younger (Berbee et al., Reference Berbee, Strullu-Derrien, Delaux, Strother, Kenrick, Selosse and Taylor2020). Fungi are thought to have originated in freshwater environments in co-evolution with Archaeplastid algae based on molecular reconstructions (Lücking et al., Reference Lücking, Huhndorf, Pfister, Plata and Lumbsch2009; Berbee et al., Reference Berbee, Strullu-Derrien, Delaux, Strother, Kenrick, Selosse and Taylor2020). Hence, the low salinity of the Diabaig depositional environment (Nielson et al., Reference Nielson, Stüeken and Prave2024) makes this an attractive target in the search for new Proterozoic fungi. The green lineage of eukaryotic algae already radiated by this time (Tang et al., Reference Tang, Pang, Yuan and Xiao2020; Sforna et al., Reference Sforna, Loron, Demoulin, François and Cornet2022; Yang et al., Reference Yang, Ma, Wang, Tian, Sun, Zhang, Xiao, De Clerck, Leliaert and Zhong2023).
Results
Organic-walled microfossils (OWM) totaling 3367 were counted from samples of the Diabaig Formation, from which we report 11 taxa including new and previously described specimens (Figure 3). Thus, we increased the total number of reported fossil morphospecies in the Torridon Group from 29 to 32 (Figure 2 and SI Table 2). Our taxa include 2 unambiguous eukaryotes, Bicellium braseri and Germinosphaera bispinosa (Figures 4 and 7.1–7.4, respectively) and 2 probable but ambiguous eukaryotes (Pterospermopsimorpha sp., and Unnamed species B; Figures 9.1–9.3 and 11.4–11.9, respectively). We also describe a new multi-element, filament-forming taxon, Minimarmilla multicatenaria n. gen. n. sp. (Figure 8).
Simple, unornamented, spheroidal vesicles are the most abundant fossil morphotype (Leiosphaeridia ternata, Leiosphaeridia spp., and Synsphaeridium spp.), followed by filamentous forms of Siphonophycus spp. Rarer morphotaxa include large aggregates of the small, slightly tapered ellipsoids Eohalothece lacustrina, the multicellular Bicellum brasieri, few process-bearing Germinosphaera bispinosa, a double-walled vesicular Pterospermopsimorpha sp., ball-like bundles of thin filaments (Unnamed species A), and networks of branching filaments (Unnamed species B).
Discussion
Diversity of OWM in the Diabaig Formation
The Diabaig Formation provides a snapshot of a microbial community in an inland aquatic environment from the Mesoproterozoic–Neoproterozoic transition (e.g., McMahon and Davies Reference McMahon and Davies2020; Wei and Algeo, Reference Wei and Algeo2020; Nielson et al., Reference Nielson, Stüeken and Prave2024). Major geological events at that time, such as the Grenville Orogeny and the culmination of the amalgamation of the supercontinent Rodinia, produced the Torridonian succession of fluvial–alluvial and shallow-marine facies (Kinnaird et al., Reference Kinnaird, Prave, Kirkland, Horstwood, Parrish and Batchelor2007; Strachan et al., Reference Strachan, Holdsworth, Prave, Woodcock and Strachan2012; McMahon and Davies Reference McMahon and Davies2020; Krabbendam et al., Reference Krabbendam, Strachan and Prave2022).
Microfossils described herein, although not highly diverse, are among the oldest in Europe and include the oldest known aquatic eukaryotes from a non-marine ecosystem, as well as various prokaryotes (Figure 12). Some of these fossils, both new and previously described, are also common in contemporaneous assemblages (SI Table 2, Figure 2). Although this lack of diversity could be explained by different seasonal microbial communities in the paleo-ecosystem just as occurs in modern environments (Metz et al., Reference Metz, Lopes dos Santos, Berman, Bigeard, Licursi, Not, Lara and Unrein2019), the low preservation potential of microbial structures, diagenetic and taphonomic factors, and biases in rock-sample collection in the field should also be considered (Donoghue and Purnell, Reference Donoghue and Purnell2009; Javaux and Benzerara, Reference Javaux and Benzerara2009). All named taxa except for Bicellum brasieri and Minimarmilla multicatenaria n. gen. n. sp. are also found in marine settings of comparable age.
Diversity of organic-walled microfossils (OWM) in the Diabaig Formation. Across the five localities studied, Leiosphaeridia specimens were most abundant, followed by Siphonophycus and Synsphaeridium.

The 11 taxa described here include two undoubted eukaryotes (Bicellum brasieri and Germinosphaera bispinosa), and two likely eukaryotes (Pterospermopsimorpha sp. and Unnamed species B), or three if one includes Leiosphaeridia; we also consider a eukaryotic affinity for Minimarmilla multicatenaria n. gen. n. sp. but find the evidence insufficient. Previously reported taxa with inferred eukaryotic affinity in the Torridon are Bicellum brasieri (e.g., Strother and Wellman, Reference Strother and Wellman2016), Germinosphaera bispinosa (e.g., Battison, Reference Battison2012), Lophosphaeridium sp. (Strother et al., Reference Strother, Battison, Brasier and Wellman2011), “Microfossils with reticulated wall” (Strother et al., Reference Strother, Battison, Brasier and Wellman2011), Pterospermopsimorpha pileiformis (Battison, Reference Battison2012), Valeria lophostriata (Jankauskas, Reference Jankauskas1979) Jankauskas, Reference Jankauskas and Keller1982 (Battison, Reference Battison2012), Maculosphaera kingstonensis Licari, Reference Licari1978 (Battison, Reference Battison2012), and Stictosphaeridium sp. (Strother et al., Reference Strother, Battison, Brasier and Wellman2011).
It is estimated that crown-group eukaryotes evolved in the late Mesoproterozoic (Parfrey et al., Reference Parfrey, Lahr, Knoll and Katz2011; Eme et al., Reference Eme, Sharpe, Brown and Roger2014; Betts et al., Reference Betts, Puttick, Clark, Williams, Donoghue and Pisani2018) although the timing of eukaryogenesis is still poorly constrained (Porter, Reference Porter2020; Porter and Riedman, Reference Porter and Riedman2023). Morphologically complex fossils are evidently present in many Meso-Neoproterozoic assemblages, including the Torridon Group; further exploration will continue to provide key data to refine our overall picture of early eukaryotic evolution. Moreover, the Torridonian has been suggested as an ideal place to search for early fungal candidates (Berbee et al., Reference Berbee, Strullu-Derrien, Delaux, Strother, Kenrick, Selosse and Taylor2020), and the report of the mycelium-like specimen (Unnamed species B) in this study may be a first step in that direction. The environmental setting offers a plausible non-marine ecosystem for early fungi, including abundant carbon sources to support a parasitic or saprotrophic lifestyle, but also the possibility of mutualistic interactions with early free-living freshwater algae comparable to extant fungal species (Du et al., Reference Du, Zienkiewicz, Vande Pol, Ostrom, Benning and Bonito2019; Naranjo-Ortiz and Gabaldón, Reference Naranjo‐Ortiz and Gabaldón2019). Molecular-clock estimates indicate an emergence of the fungal lineage during the Tonian period (Lücking et al., Reference Lücking, Huhndorf, Pfister, Plata and Lumbsch2009; Berbee and Taylor, Reference Berbee and Taylor2010), so the present specimen would represent the earlier end of the estimated origin interval of filamentous fungi (Naranjo-Ortiz and Gabaldón, Reference Naranjo‐Ortiz and Gabaldón2019; Berbee et al., Reference Berbee, Strullu-Derrien, Delaux, Strother, Kenrick, Selosse and Taylor2020), similar to the other Neoproterozoic fungi-like fossils 1–0.89-Ga Ourasphaira giraldae Loron et al., Reference Loron, François, Rainbird, Turner, Borensztajn and Javaux2019b, and 1.02–1.03-Ga Mucorites ripheicus Hermann, Reference Hermann and Sokolov1979 (Hermann and Podkovyrov, Reference Hermann and Podkovyrov2006).
New filamentous microorganism
Minimarmilla multicatenaria n. gen. n. sp. is a new fossil form representing either a large cyanobacterial aggregate or possibly a microalga. Because of their uniformity in size and shape, we interpret the cell-like units as the remains of actual cells rather than fragments of a broken sheath. They are not a taphonomic variety of Siphonophycus, which is present in the same thin sections, but easily distinguishable (i.e., not bundled in strands, not opaque, and not broken into numerous regular segments, although some breakage is apparent as in Figure 10.5; nor do any of the Siphonophycus specimens preserve interior cell-like units).
Minimarmilla multicatenaria n. gen. n. sp. may represent aggregated bundles of uniseriate cyanobacterial trichomes (similar to the ‘rafts’ typical of the freshwater nostocalean Aphanizomenon, for example). To date, the oldest microfossils considered as undoubted cyanobacteria are Eoentophysalis belcherensis Hofmann, Reference Hofmann1976 (ca. 1.9 Ga; Hofmann, Reference Hofmann1976; Butterfield, Reference Butterfield2015), Polybessurus (1.35–1 Ga; Sergeev et al., Reference Sergeev, Knoll and Grotzinger1995), Eohyella (1.63 Ga; Zhang, Reference Zhang1988), and Polysphaeroides filiformis (ca. 1 Ga; Demoulin et al., Reference Demoulin, Lara, Cornet, François, Baurain, Wilmotte and Javaux2019), the latter considered an early member of Stigonemacea. Thus, M. multicatenaria n. gen. n. sp. could represent a contemporary of P. filiformis, showing the diversification of Stigonemacea during the Meso-Neoproterozoic and a new record of early complex cyanobacteria. However, M. multicatenaria lacks the right-angled branching pattern typical of Stigonemacea and indeed does not show any unambiguous branching in the specimens so far recovered.
Given the age of this material, a eukaryotic interpretation should not be favored without strong evidence, which is lacking. Nevertheless, we note that the aspect ratio and bone-like shape (slightly swollen ends) of the cell-like units comprising M. multicatenaria n. gen. n. sp. are reminiscent of several Ceramialean red algae (SI Figure 3). Speculatively, M. multicatenaria n. gen. n. sp. could be a rare example of a crown-group eukaryote lineage from the Meso-Neoproterozoic interval. The oldest accepted crown-group eukaryote to date is the 1047-Ma red alga Bangiomorpha pubescens Butterfield, Reference Butterfield2000 (Gibson et al., Reference Gibson, Shih, Cumming, Fischer and Crockford2018), followed by fossils interpreted as multicellular green algae, ca. 1000-Ma Proterocladus (Butterfield et al., Reference Butterfield, Knoll, Swett and Bengston1994; Tang et al., Reference Tang, Pang, Yuan and Xiao2020) and ca. 800-Ma Paleovaucheria (Butterfield, Reference Butterfield2004, Reference Butterfield2009).
Unlike Bangiomorpha, Minimarmilla n. gen. does not show any features unambiguously diagnostic of a red algal affinity, although a knob-like cell cluster observed in one specimen (Figure 8.15, 8.16 and SI Figure 4) is fairly reminiscent of the carposporangium in Ceramialean rhodophytes such as Polysiphonia (Stuercke and Freshwater, Reference Stuercke and Freshwater2008; Mamoozadeh and Freshwater, Reference Mamoozadeh and Freshwater2012; Baweja et al., Reference Baweja, Kumar, Sahoo, Levine, Fleurence and Levine2016). In any case, the cells of Minimarmilla n. gen. are not laterally aligned across the bundle, whereas those of the Ceramialean red algae generally are, giving a grid-like appearance to their thalli (it is possible that such an alignment may be lost taphonomically). The cells of extant red algae are also generally larger than those of Minimarmilla n. gen. Cells of Polysiphonia spp. are typically 20–30 μm in diameter (Díaz-Tapia et al., Reference Díaz-Tapia, Sook Kim, Secilla, Bárbara and Cremades2013), while those of Minimarmilla n. gen. are 3–4 μm in the best-preserved examples, notwithstanding the possibility of taphonomic shrinkage. Overall, unless and until more definitive algal characteristics are discovered, the balance of evidence provisionally favors a cyanobacterial interpretation.
Conclusions
The Torridon Group in Northwest Scotland hosts an important Lagerstätte with organic-walled microfossils (OWM) dated from the Mesoproterozoic–Neoproterozoic, an important interval for the early evolution and diversification of eukaryotes. This study focused on the exceptionally well-preserved microfossils found in phosphorites from the Diabaig Formation. We documented 11 morphospecies of eukaryotic and prokaryotic organisms, including the new taxon Minimarmilla multicatenaria n. gen. n. sp., and two unnamed taxa, thus complementing previous reports of fossil Torridon microbiota (i.e., Strother et al., Reference Strother, Battison, Brasier and Wellman2011, Battison, Reference Battison2012; Wacey et al., Reference Wacey, Saunders, Roberts, Menon, Green, Kong, Culwick, Strother and Brasier2014; Strother and Wellman, Reference Strother and Wellman2016; Sirantoine, Reference Sirantoine2021). Minimarmilla multicatenaria n. gen. n. sp. displays some features in common with extant red algae, although a cyanobacterial origin should be preferred without stronger evidence for eukaryotic affinity. Our unnamed species B appears to be a network-forming organism that bears tentative comparison with fungi. The biota of the Torridon Group opens a rare and important window on the evolutionary history of eukaryotes ca. 1 Ga ago in an inland aquatic paleoenvironment. Further study of these fossils will no doubt reveal additional interesting new fossil forms.
Acknowledgments
The authors thank J.J. Matthews, C. MacFadyen at NatureScot, and W. McMahon for advice regarding sampling, I. Febbrari at School of GeoSciences for preparing the thin sections used in this study, N.J. Butterfield for advice on M. multicatenaria n. gen. n. sp., B. Slater and an anonymous referee for their insightful criticisms, and assistant editor J. Botting for suggesting further useful revisions. ERD acknowledges the support of a PhD scholarship from the School of Physics of Astronomy, University of Edinburgh. CCL acknowledges support from the Royal Society and the Leverhulme Trust. HA acknowledges support from the Leverhulme Trust.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Data availability statement
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.9ghx3ffvp
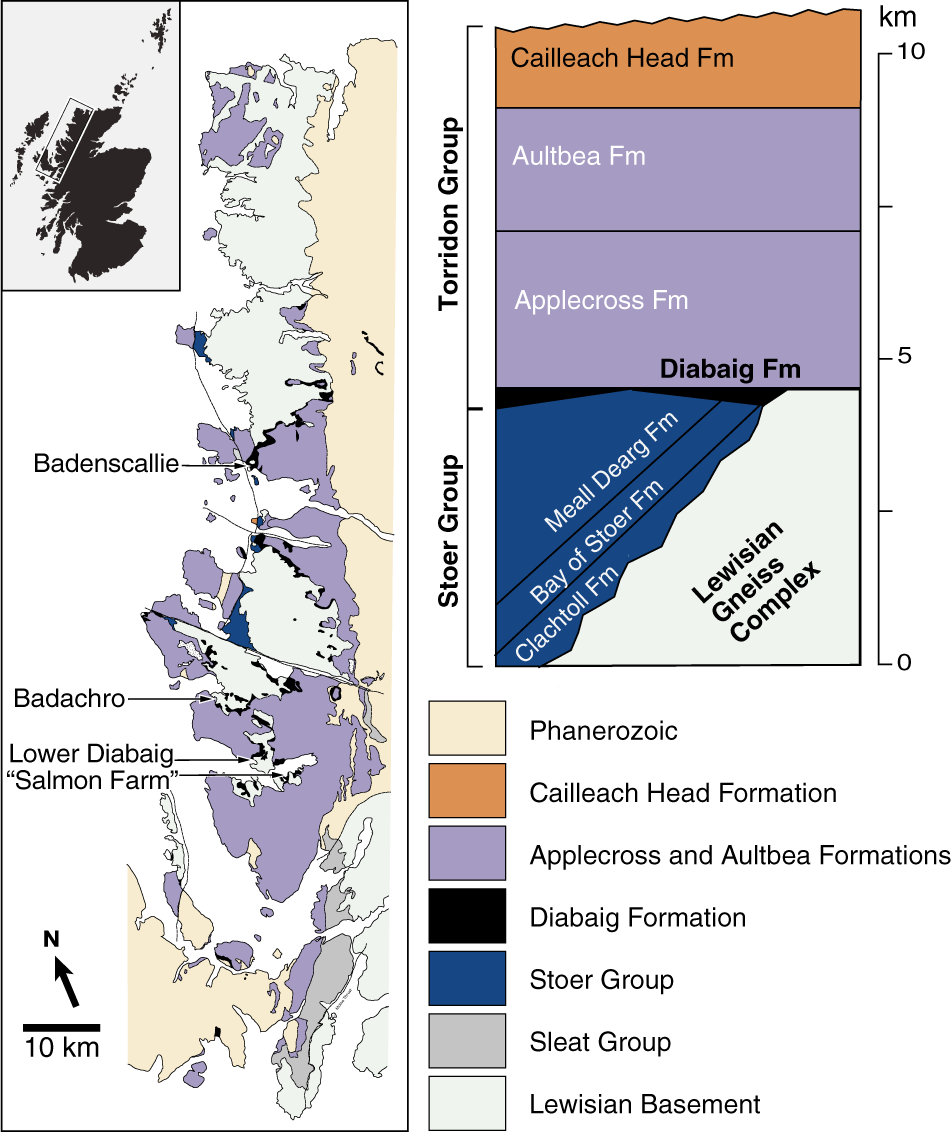
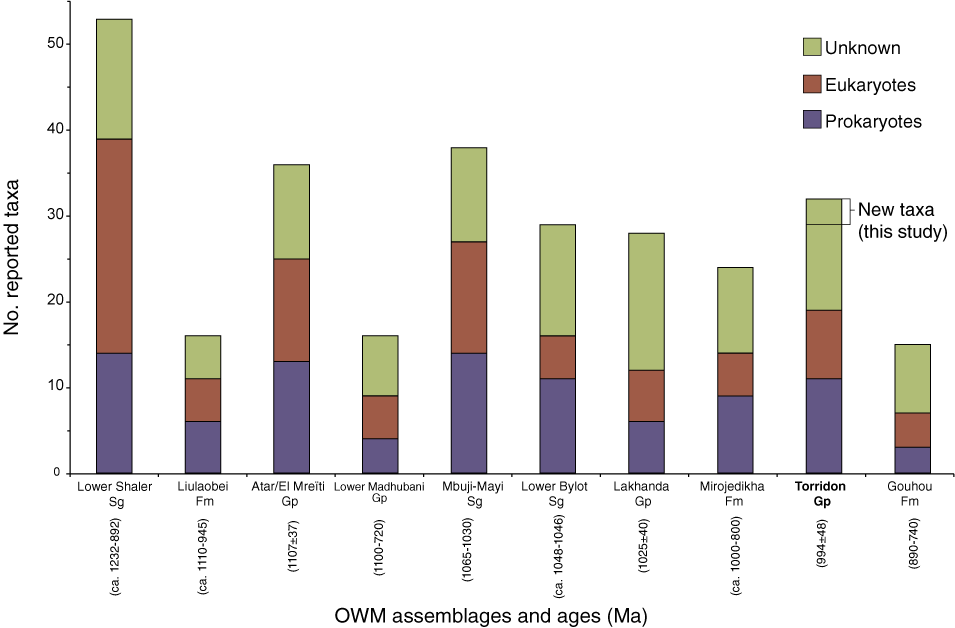









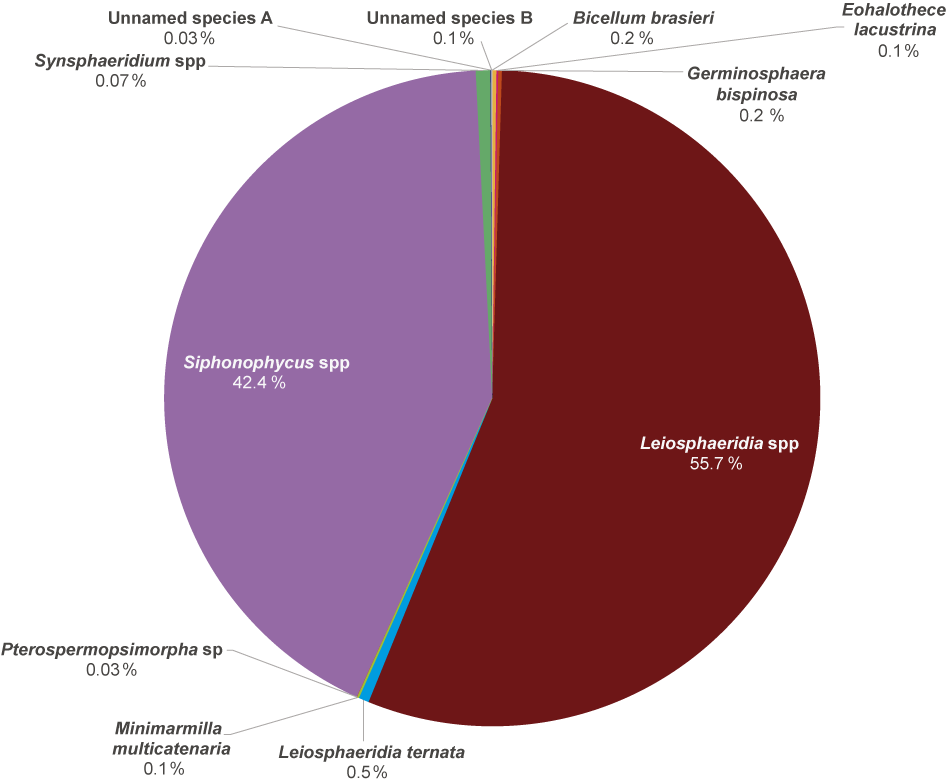

 Open access
Open access