


Non-technical Summary
Our knowledge about the life appearance of extinct marine reptiles is limited by the rarity of preserved soft tissues in the fossil record. Therefore, we developed a new framework to predict the tail fin shape of mosasaurians—a major group of aquatic lizards that thrived in the world’s oceans some 98 to 66 million years ago—based on skeletal data and comparisons with living sharks. By analyzing how the backbone and associated muscles of shark tails relate to the fleshy fin outline, and by verifying that this relationship is also applicable to mosasaurians with preserved body outlines, we built a predictive model capable of reconstructing the tail fins of mosasaurian species lacking direct soft-tissue evidence. Our novel approach combines two-block partial least squares (2B-PLS) regression and geometric analysis and reveals that mosasaurians evolved a variety of tail fin shapes that likely reflect different lifestyles. Coastal species had long and almost eel-like tails ideal for maneuvering in shallow waters, while species living in open-ocean habitats evolved semi-lunate flukes similar to those of pelagic sharks. We also found that a bilobed, downward bent (hypocercal) tail evolved independently at least four times in Mosasauria, which highlights the evolutionary complexity of their secondary adaptation to aquatic life during the Late Cretaceous. This study demonstrates the potential of combining geometric analysis with data from modern analogues when reconstructing the life appearance of extinct animals.
Introduction
Mosasaurians (Mosasauria) are a group of squamates that became the dominant predators in the marine ecosystems of the Late Cretaceous, some 98 to 66 million years ago (Jacobs et al. Reference Jacobs, Ferguson, Polcyn and Rennison2005; Augusta et al. Reference Augusta, Zaher, Polcyn, Fiorillo, Jacobs, Gower and Zaher2022). During their relatively brief geological existence, they evolved from meter-long, semiaquatic forms to giant ocean-going leviathans (Lindgren et al. Reference Lindgren, Polcyn and Young2011; Cross et al. Reference Cross, Moon, Stubbs, Rayfield and Benton2022; MacLaren et al. Reference MacLaren, Bennion, Bardet and Fischer2022). Although Wiman (Reference Wiman1920) proposed the presence of an ichthyosaur-like tail fin in derived mosasaurians already in the early twentieth century, the traditional view of these ancient reptiles as serpentine creatures equipped with a paddle-shaped tail remained virtually unchanged until the early twenty-first century (e.g., Russell Reference Russell1967; Massare Reference Massare1988, Reference Massare, Maddock, Bone and Rayner1994). Over the past two decades, however, an array of osteological and soft-tissue evidence has revealed that fully aquatic forms possessed a well-developed hypocercal fluke akin to the tail fin of pelagic sharks (albeit being turned upside down), which suggests more specialized swimming capabilities (Lindgren et al. Reference Lindgren, Jagt and Caldwell2007, Reference Lindgren, Caldwell, Konishi and Chiappe2010, Reference Lindgren, Polcyn and Young2011, Reference Lindgren, Kaddumi and Polcyn2013; Harrell et al. Reference Harrell, Pérez-Huerta and Suarez2016).
The locomotor performance of axial-undulatory swimming vertebrates correlates with the morphology of their tail fins (Westneat and Wainwright Reference Westneat and Wainwright2001; Ferrón et al. Reference Ferrón, Martínez-Pérez and Botella2017; Sternes and Shimada Reference Sternes and Shimada2020). While previous research has established the presence of a bilobed fluke in certain hydropedal mosasaurians (Lindgren et al. Reference Lindgren, Kaddumi and Polcyn2013), our comprehension of the evolution of this propulsive surface remains limited due to the rarity of soft-tissue fossils across this clade. Furthermore, knowledge about the diversity of tail fin morphologies both within and among the different evolutionary lineages of mosasaurians remains virtually nonexistent.
The two-block partial least squares (2B-PLS) regression method was initially developed to explore the covariance between two or more sets of shape variables (Rohlf and Corti Reference Rohlf and Corti2000), but has since shown promise in the prediction of absent elements: an anatomical unit can be predicted from another one, under the condition that these exhibit high covariance (Archer et al. Reference Archer, Pop, Rezek, Schlager, Lin, Weiss, Dogandžić, Desta and McPherron2018). In recent years, the application of predictive analysis to anatomical system reconstructions has gained increasing attraction, particularly in the fields of orthognathic surgery and paleoanthropology (e.g., Bastir et al. Reference Bastir, Torres-Tamayo, Palancar, Lois-Zlolniski, García-Martínez, Riesco-López, Vidal, Been, Gómez-Olivencia and Ann Kramer2019; Torres-Tamayo et al. Reference Torres-Tamayo, Schlager, García-Martínez, Sanchis-Gimeno, Nalla, Ogihara, Oishi, Martelli and Bastir2020; Niño-Sandoval et al. Reference Niño-Sandoval, Jaque, González and Vasconcelos2022).
Despite being dorsoventrally inverted, the skeletal support of the tail fin in extant sharks shows remarkable similarity to that of derived mosasaurians, suggesting that the former can serve as a reasonable modern analogue for reconstructions of the fluke in these ancient marine lizards (Lindgren et al. Reference Lindgren, Jagt and Caldwell2007, Reference Lindgren, Polcyn and Young2011, Reference Lindgren, Kaddumi and Polcyn2013). Therefore, we created a predictive model designed to reconstruct the tail fin morphology of extant sharks using the underlying caudal skeleton and its associated musculature. This was done by employing geometric morphometrics in combination with 2B-PLS analysis. We then used this framework to predict the soft-tissue outline of the tail fin in a number of mosasaurians, ranging from basally branching to derived forms.
Material and Methods
Construction of Our Predictive Model
The sample dataset (Table 1) comprises the tail fin region of 29 specimens; collectively, these represent 21 species of extant sharks and 4 species of extinct marine reptiles that are known from soft-tissue fossils. Seven fixed landmarks and 52 semi-landmarks were digitized from lateral-view images of the tails using tpsDig v. 2.32 (Rohlf Reference Rohlf2015). The data from each specimen were divided into two subsets: the Tail Fin Outline (TFO), consisting of 4 fixed landmarks and 31 semi-landmarks, and the Caudal Fin Skeleton/Musculature (CFS/M), consisting of 3 fixed landmarks and 21 semi-landmarks (Fig. 1A). We refer to the part of the fluke in mosasaurians lacking skeletal support as the Tail Fin Expanded Soft Tissue (TFEST). It broadly corresponds to the caudal fin web of extant sharks (Kim et al. Reference Kim, Shimada and Rigsby2013), and its shape is defined by combining all landmarks and semi-landmarks of the TFO and CFS/M (Fig. 1A). Because the TFEST provides information about the relationship between the CFS/M and TFO, we used the TFEST (instead of TFO) when constructing our predictive model.
Extant sharks and extinct marine reptiles with known soft-tissue outlines used in this study

Landmarks (red) and semi-landmarks (gray) applied to: A, a generalized shark tail fin turned upside down (modified from Crofts et al. Reference Crofts, Shehata and Flammang2019: fig. 7A); and B, the posteriormost portion of the vertebral column of the derived mosasaurine Plotosaurus bennisoni (modified from Lindgren et al. Reference Lindgren, Caldwell and Jagt2008: fig. 4A). The tail fin region is subdivided into three main units: TFO, tail fin outline; CFS/M, caudal fin skeleton/musculature (blue area; and TFEST, tail fin expanded soft tissue (green area). Fixed landmarks of mosasaurians (with the arrangement reversed in sharks): landmark 1, CFS/M upper origin; landmark 2, CFS/M lower origin; landmark 3, CFS/M posterior tip; landmark 4, TFO upper origin; landmark 5, TFO lower origin; landmark 6, distal tip of tail fin ventral lobe; landmark 7, distal tip of the dorsal lobe (in mosasaurians lacking a dorsal lobe, this landmark corresponds to the uppermost point of the TFEST).

We performed generalized Procrustes analysis on the raw coordinates obtained from each shark and marine reptile. This method aligns landmarks and semi-landmarks by standardizing and eliminating variations that might arise from differences in the position, size, and orientation of the specimens, thus enabling direct comparison of shape (Cooke and Terhune Reference Cooke and Terhune2015). Thereafter, we employed 2B-PLS analysis on the shape coordinates to evaluate the covariance between the tail fin outline and underlying skeleton/musculature. These analyses were carried out using the Morpho package v. 2.11 in R Studio v. 4.1.1 (Schlager Reference Schlager, Zheng, Li and Székely2017).
To avoid overfitting in the 2B-PLS predictive model, we had to find the most suitable number of latent variables (LVs) with minimum prediction error (i.e., mean square error [MSE] of the Procrustes distance between each individual specimen and its prediction). Leave-one-out cross-validation (LOOCV) analysis was therefore employed to determine the optimal number of LVs. In this method (for a dataset containing N samples), one sample was selected as a test set and the remaining (N − 1) samples were used as a training set to build the predictive model. This process was then repeated N times, thereby ensuring that each sample had served as a test set once. After all tests were completed, the MSE was calculated to evaluate the model’s predictive accuracy. This ensured that the prediction for each specimen was done independently by preventing the excluded specimen from influencing the model through self-inference (Schlager Reference Schlager2013; Archer et al. Reference Archer, Pop, Rezek, Schlager, Lin, Weiss, Dogandžić, Desta and McPherron2018; Torres-Tamayo et al. Reference Torres-Tamayo, Schlager, García-Martínez, Sanchis-Gimeno, Nalla, Ogihara, Oishi, Martelli and Bastir2020).
Rationale for Mosasaurian Fluke Shape Prediction
The skeletons of nine species of mosasaurians (Pannoniasaurus inexpectatus, Platecarpus tympaniticus, Tylosaurus proriger, Eonatator sternbergi, Dallasaurus turneri, Clidastes sp., Eremiasaurus heterodontus, Mosasaurus sp., and Plotosaurus bennisoni) were selected for predictive analysis (see Osborn Reference Osborn1899: fig. 13; Bardet and Pereda Suberbiola Reference Bardet and Suberbiola2001: fig. 1A; Lindgren Reference Lindgren2007: fig. 3A; Lindgren et al. Reference Lindgren, Jagt and Caldwell2007: fig. 3A, 2008: fig. 4A, 2010: fig. 8A, 2011: fig. 12; LeBlanc et al. Reference LeBlanc, Caldwell and Bardet2012: fig. 10C; Makádi et al. Reference Makádi, Caldwell and Ősi2012: fig. 4B; Carpenter Reference Carpenter2017: fig. 9). The caudal vertebrae of Mosasaurus sp. were adjusted according to the articulated tail of MDC 20269994 (Mosasaurus beaugei), an undescribed specimen from Morocco (N. Longrich personal communication 2024) and IRSNB R29 (Mosasaurus lemonnieri). The mosasaurians were divided into two distinct morphotypes based on limb and pelvic morphology: (1) plesiopedal forms characterized by limbs and a pelvis retaining terrestrial-like features (Pontosaurus, Vallecillosaurus, Pannoniasaurus, and Dallasaurus); and (2) hydropedal forms exhibiting flipper-like extremities and loss of sacro-iliac contact (Platecarpus, Tylosaurus, Eonatator, Clidastes, Eremiasaurus, Mosasaurus, and Plotosaurus) (Bell and Polcyn Reference Bell and Polcyn2005).
Anatomical studies of various sharks and reptiles have demonstrated that within the tail fin region, the shape of the caudal musculature broadly corresponds to the outline of the caudal vertebrae when observed in lateral view (Flammang Reference Flammang2010; Young et al. Reference Young, Dumais, John, Lyons, Macduff, Most, Reiser and Reiser2016; LeBlanc et al. Reference LeBlanc, MacDougall, Haridy, Scott and Reisz2018; Díez Díaz et al. Reference Díez Díaz, Demuth, Schwarz and Mallison2020). Consequently, this study considers the skeletal outline of the caudal vertebrae of all fossil species to broadly represent the outline of the associated musculature (Fig. 1).
Based on osteological studies of the tail anatomy in derived ichthyosaurs and ERMNH HFV 197 (Prognathodon sp.), we infer the origin of the tail fin region from the vertebral centra exhibiting the lowest centrum length/centrum height ratio in the posterior portion of the intermediate caudal series (Buchholtz Reference Buchholtz2001; Lindgren et al. Reference Lindgren, Kaddumi and Polcyn2013). As in the CFS/M subset of the training (shark) set, 3 fixed landmarks and 21 semi-landmarks were defined and digitized from skeletal reconstructions of the nine selected mosasaur species (Fig. 1B).
Because the predictive model is built mainly from extant shark data, all of the reconstructed mosasaur tail fins in this study have a subterminal lobe (Fig. 1A). However, this soft-tissue structure is regarded as unique to sharks (Shadwick and Goldbogen Reference Shadwick and Goldbogen2012) and is accordingly not known from any Mesozoic marine reptile (Maisch Reference Maisch2008; Spindler et al. Reference Spindler, Lauer, Tischlinger and Mäuser2021; Delsett et al. Reference Delsett, Friis, Kölbl-Ebert and Hurum2022; Eriksson et al. Reference Eriksson, De La Garza, Horn and Lindgren2022). Consequently, we manually adjusted the terminal part of the predicted ventral lobe to better align with the documented tail fin outlines of marine reptiles, such as ichthyosaurs and metriorhynchid crocodylimorphs.
The predicted TFEST coordinates reflect covariance patterns captured by the 2B-PLS method rather than the correspondence with the original CFS/M. To reconcile these geometric disparities, we performed thin-plate spline deformation to align the base of the TFEST with the original CFS/M (3 fixed, 21 semi-landmarks) (Fig. 1A). This process anchored the reconstructed soft-tissue data to the skeleton/musculature framework and preserved the spatial relationship between the base of the TFEST and TFO.
Morphometric Analyses
To infer lifestyle and swimming mode of the species with predicted tail fin outlines, and to quantitatively assess morphological differences among the various mosasaurians, we employed geometric morphometrics (Mitteroecker and Gunz Reference Mitteroecker and Gunz2009) to analyze the morphospace occupied by sharks and marine reptiles in two dimensions. We applied the same method used when constructing the aforementioned predictive model to digitize the TFO of 89 specimens, including 70 extant sharks, 7 ichthyosaurs, and 12 mosasaurians (9 of the last group were reconstructed using the predictive model introduced in this study; see Supplementary Table 1).
We conducted a principal component analysis (PCA) of Procrustes-transformed morphometric data using the geomorph v. 4.0.3 package (Baken et al. Reference Baken, Collyer, Kaliontzopoulou and Adams2021) in R Studio to identify the positions of the fossil marine reptiles in morphospace relative to those of living sharks. Most sharks were classified into three broad groups based on their swimming mode: anguilliform, carangiform, and thunniform (Sternes and Shimada Reference Sternes and Shimada2020). However, because alopiid sharks exhibit specialized tail fins compared to other carangiforms, we classified them separately as “carangiform-alopiids” (Oliver et al. Reference Oliver, Turner, Gann, Silvosa and D’Urban Jackson2013).
Limitations of the Study
Our predictive approach relies heavily on distinct similarities in both morphology and arrangement of the neural spines and haemal arches in the caudal region of mosasaurians and extant sharks (Lindgren et al. Reference Lindgren, Jagt and Caldwell2007, Reference Lindgren, Polcyn and Young2011, Reference Lindgren, Kaddumi and Polcyn2013; Carpenter Reference Carpenter2017). Consequently, its applicability is limited to predictions of the tail fin of mosasaurians, aquatic varanids, some basal ichthyosaurs (e.g., Mixosaurus), and most sharks. It is not suitable for taxa that lack distinctly elongate and slender neural spines and haemal arches in the caudal region, such as thalattosuchian crocodylomorphs, squaliform sharks, and derived ichthyosaurs (Fig. 2). Moreover, because our predictive model is based on the overall shape of the caudal fin skeleton/musculature, the caudal vertebral segment (including the haemal arches and neural spines) needs to be nearly complete and additionally pristinely preserved (i.e., undeformed) for accurate predictions of the target specimens.
Examples of the caudal vertebral morphology in applicable and non-applicable species (pertaining to the predictive model). A, Plotosaurus bennisoni (modified from Lindgren et al. Reference Lindgren, Jagt and Caldwell2007: fig. 5A). B, Scalloped hammerhead shark, Sphyrna lewini (modified from Lindgren et al. Reference Lindgren, Jagt and Caldwell2007: fig. 5B). C, Mixosaurus cornalianus (redrawn from Klug et al. Reference Klug, Sivgin, Miedema, Scheffold, Reisdorf, Stössel, Maxwell and Scheyer2024: fig. 6). D, Cricosaurus suevicus (redrawn from Williston and Gregory Reference Williston and Gregory1925: fig. 85). E, Cookiecutter shark, Isistius brasiliensis (redrawn from de Figueiredo Petean and de Carvalho Reference de Figueiredo Petean and de Carvalho2018: fig. 1B). F, Ophthalmosaurus icenicus (redrawn from Cleary et al. Reference Cleary, Moon, Dunhill and Benton2015: fig. 1B).

It is also worth pointing out that tail fin morphology alone does not fully reflect the swimming mode of any animal. Rather, it needs to be integrated with studies dealing with the overall body shape, biomechanics, and hydrodynamics to more accurately reflect the ecology and swimming capabilities of mosasaurians. Nevertheless, soft-tissue reconstructions still remain crucial indicators of paleoecology (Ferrón et al. Reference Ferrón, Martínez-Pérez and Botella2017).
Institutional Abbreviations.
AMNH: American Museum of Natural History, New York, New York, USA; DMNH: Perot Museum of Nature and Science, Dallas, Texas, USA; ERMNH: Eternal River Museum of Natural History, Amman, Jordan; GSM: Georgia Southern Museum, Georgia Southern University, Statesboro, Georgia, USA; IRSNB: Institut Royal des Sciences Naturelles de Belgique, Brussels, Belgium; KUVP: Natural History Museum and Biodiversity Research Center, Lawrence, Kansas, USA; LACM: Natural History Museum of Los Angeles County, Los Angeles, California, USA; MDC: Musée des Confluences, Lyon, France; MTM: Hungarian Natural History Museum, Budapest, Hungary; PMU: Museum of Evolution, Paleontology Section, Uppsala University, Uppsala, Sweden; TMM: Texas Memorial Museum, Dallas, Texas, USA; UALVP: University of Alberta Laboratory for Vertebrate Paleontology, Edmonton, Alberta, Canada; UCMP: University of California Museum of Paleontology, Berkeley, California, USA.
Results
Relationship between CFS/M and Tail Fin Morphology
The results of our LOOCV analysis revealed a significant increase in prediction error when the number of LVs exceeded three (Fig. 3), which indicates that the optimal prediction for the tail fin morphology is achieved with the first three LVs. Collectively, the selected LVs explain more than 95% of the total covariance between the CFS/M and TFEST in the 2B-PLS analysis, with each LV accounting for 73.88%, 21.99%, and 1.91% of the total variance, respectively. Accordingly, these LVs are sufficient to effectively describe the covariation between the two structures.
Error plot for the selection of latent variables (LVs) in the predictive model. The intersection marked by a dashed line indicates the optimum number of LVs (= 3), which corresponds to the minimum mean square error (MSE).

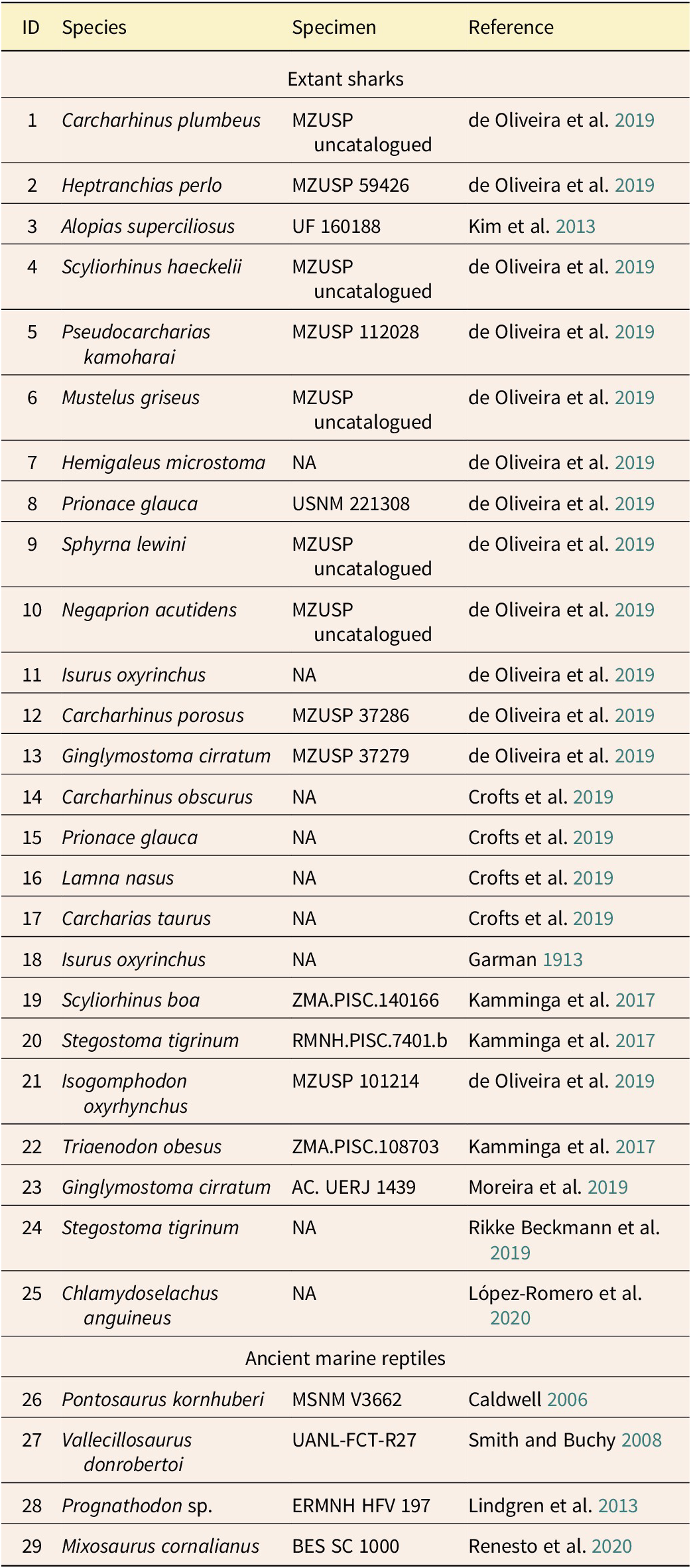
The 2B-PLS analysis demonstrated strong morphological integration between CFS/M and TFEST, showing high correlations (r > 0.9, p < 0.05) and statistically significant singular values (p < 0.05) across the selected LVs. This covariation pattern remained consistent in both sharks and marine reptiles (Fig. 4), including the only known derived mosasaurian with a preserved tail fin outline (ERMNH HFV 197). These results thus validate the predictive model’s capability to accurately reconstruct mosasaurian tail fin shapes through CFS/M–TFEST covariation.
Two-block partial least squares (2B-PLS) analysis of caudal fin skeleton/musculature (CFS/M) and tail fin expanded soft tissue (TFEST) covariation for each of the three latent variables (LVs) selected in the predictive model. The dots represent differences in landmark placement of the two structures along each axis. In the first (A), second (B), and third (C) LVs, variations in CFS/M and TFEST show significant covariation. Note placement of ERMNH HFV 197 (Prognathodon sp.), which indicates that the covariation pattern between the two structures in derived mosasaurians is consistent with that of the predictive model.

Mosasaurian Tail Fin Predictions
Figure 5 presents the tail fin morphologies of nine mosasaurian species based on the predictive model. The two plesiopedal forms (i.e., Pannoniasaurus inexpectatus and Dallasaurus turneri) possess broadly paddle-like tails, and the predicted morphologies for all hydropedal species include a prominent dorsal lobe, particularly in derived mosasaurines (i.e., Clidastes, Eremiasaurus, Mosasaurus, and Plotosaurus). Among the hydropedal mosasaurines, Plotosaurus exhibits the most symmetrical tail fin (Fig. 5L). Conversely, Platecarpus is reconstructed with an elongate ventral fin lobe and a relatively small dorsal lobe (Fig. 5E).
Mean tail fin outline shapes of sharks (turned upside down) and ichthyosaurs used in our principal component analysis (PCA), together with the predicted fluke shape of select mosasaurians. A, Mean caudal fin shape in sharks employing anguilliform swimming modes. B, Mean caudal fin shape in sharks (excluding members of the Alopiidae) employing carangiform swimming modes. C, Mean caudal fin shape in sharks and ichthyosaurs employing thunniform swimming modes. Reconstructed tail fin outline in D, Pannoniasaurus inexpectatus (MTM 2011.43.1); E, Platecarpus tympaniticus (LACM 128319); F, Tylosaurus proriger (composite of AMNH FR 221 and GSM-1); G, Eonatator sternbergii (PMU R163); H, Dallasaurus turneri (TMM 43209-1); I, Clidastes sp. (KUVP 14596); J, Eremiasaurus heterodontus (UALVP 51744); K, Mosasaurus sp. (UCMP 61221); and L, Plotosaurus bennisoni (composite of UCMP 33913, CIT 2750, CIT 2755, and CIT 2804).

Tail Fin Shape Analysis
The first two principal components (PCs) collectively explain 73.68% of the total variance, and largely distinguish different swimming modes in morphospace occupation (Fig. 6, Supplementary Fig. 1). PC 1 (56.1%) describes the development of an upper (dorsal) lobe in marine reptiles (lower/ventral lobe in sharks). Shape changes from the sample mean in the negative direction along PC 1 are associated with a more eel-like (single-lobed) tail fin, whereas in the positive direction, the tail fin becomes more lunate.
Two-dimensional morphospace of tail fin shape variation based on the first two principal components (PCs). Deformation grids represent the extreme shape of each axis.

PC 2 (17.6%) represents the sharpness of the tail fin lobes: in the negative direction, the shape transitions into a fin with short and blunt lobes, whereas in the positive direction, it evolves into a fin with elongate and sharp lobes.
Seven hydropedal mosasaurians (Eonatator, Clidastes, Tylosaurus, Prognathodon, Eremiasaurus, Mosasaurus, and Plotosaurus) fall exclusively within the “carangiform” morphospace, with Plotosaurus exhibiting the highest PC 1 scores. Platecarpus occupies an intermediate position between the carangiform and anguilliform sharks. The two plesiopedal mosasaurians Pannoniasaurus and Dallasaurus, along with the two non-mosasaurid mosasaurians Vallecillosaurus and Pontosaurus are positioned at the extreme negative end of PC 1, exhibiting lower scores than the included anguilliform sharks.
Discussion
In previous studies, the presence of a dorsal fin lobe in marine reptiles, such as derived mosasaurians, could be qualitatively inferred based on morphometrics and osteological characteristics of the underlying caudal vertebrae (e.g., Wiman Reference Wiman1920; Lindgren et al. Reference Lindgren, Polcyn and Young2011; Konishi et al. Reference Konishi, Lindgren, Caldwell and Chiappe2012). However, for complex structures, such as tail fins, performing direct quantitative analyses and reconstructions only from traditional morphometric methods remain challenging, because it is difficult to quantify and analyze morphological shape. The findings of the present study indicate that modern ecomorphological analogues can serve as appropriate comparative samples for extinct species reconstructions through morphometric analysis. Moreover, significant potential exists for quantitative morphological reconstructions in paleontology through integration of 2B-PLS and geometric morphometrics.
The results of our 2B-PLS analysis demonstrate that in both sharks and mosasaurians, the CFS/M is strongly correlated with the soft-tissue outline of the tail fin, and thus can serve as a reliable predictor for life reconstructions (Fig. 4). In sharks and derived mosasaurians, a blunt, short CFS/M indicates well-developed chevrons/haemal arches and neural spines, which provided support for the fleshy dorsal fin lobe (ventral fin lobe in sharks) through connective tissues (Lindgren et al. Reference Lindgren, Polcyn and Young2011, Reference Lindgren, Kaddumi and Polcyn2013; Crofts et al. Reference Crofts, Shehata and Flammang2019).
Ecological Inferences
The pattern of morphospace occupation indicates high disparity of tail fin morphologies among mosasaurians, from Pontosaurus to Plotosaurus (Fig. 6, Supplementary Fig. 1). For axial-undulatory swimmers, a more symmetrical tail fin (characterized by well-developed dorsal and ventral lobes and a high aspect ratio) indicates greater cruising efficiency and thereby better adaptations to open-water (pelagic) environments (Lindgren et al. Reference Lindgren, Polcyn and Young2011; Matta et al. Reference Matta, Pendar, Battaglia and Bayandor2020; Sternes and Shimada Reference Sternes and Shimada2020). The two non-mosasaurid mosasaurians Pontosaurus and Vallecillosaurus, along with the basally branching russellosaurine Pannoniasaurus, lack a bilobed tail fin (Figs. 5, 6). Instead, these species exhibit an even more elongate and slender fluke than those of anguilliform sharks, morphologically resembling the tail of semiaquatic varanids, such as Mertin’s water monitor (Varanus mertensi). However, unlike the other plesiopedal mosasaurians, Dallasaurus may have possessed an incipient dorsal lobe (Fig. 5H), allowing its tail fin to superficially resemble the composite structure of the anal and tail fins in the living epaulette shark (Hemiscyllium hallstromi)—a typical anguilliform species adapted to life in complex reef environments (Ebert et al. Reference Ebert, Dando and Fowler2021). These results are consistent with the primitive (terrestrial-type) limb morphology of these plesiopedal mosasaurians, suggesting that they represent semiaquatic forms (Bell and Polcyn Reference Bell and Polcyn2005; Caldwell Reference Caldwell2006; Smith and Buchy Reference Smith and Buchy2008; Makádi et al. Reference Makádi, Caldwell and Ősi2012; Cross et al. Reference Cross, Moon, Stubbs, Rayfield and Benton2022).
All hydropedal mosasaurians exhibit a bilobed tail fin with a distinct dorsal lobe, yet their flukes show considerable variation (Figs. 5, 6). According to our analyses, Platecarpus occupies a position somewhere between anguilliform and carangiform sharks, being characterized by a highly asymmetrical hypocercal tail fin (Figs. 5E, 6). In morphospace, Platecarpus is most closely associated with active predatory anguilliform sharks (such as the broadnose sevengill shark, Notorynchus cepedianus), rather than with slower benthic forms, such as the zebra shark (Stegostoma varium) and nurse shark (Ginglymostoma spp.) (Fig. 7A). Furthermore, evidence from limb morphology (Cross et al. Reference Cross, Moon, Stubbs, Rayfield and Benton2022), bone microstructure (Sheldon Reference Sheldon, Callaway and Nicholls1997), and biogeochemistry (Harrell and Pérez-Huerta Reference Harrell and Pérez-Huerta2015) all support the hypothesis that Platecarpus exhibited an active littoral predatory lifestyle with occasional bursts of speed, similar to N. cepedianus in shallow seas today (Ebert et al. Reference Ebert, Dando and Fowler2021).
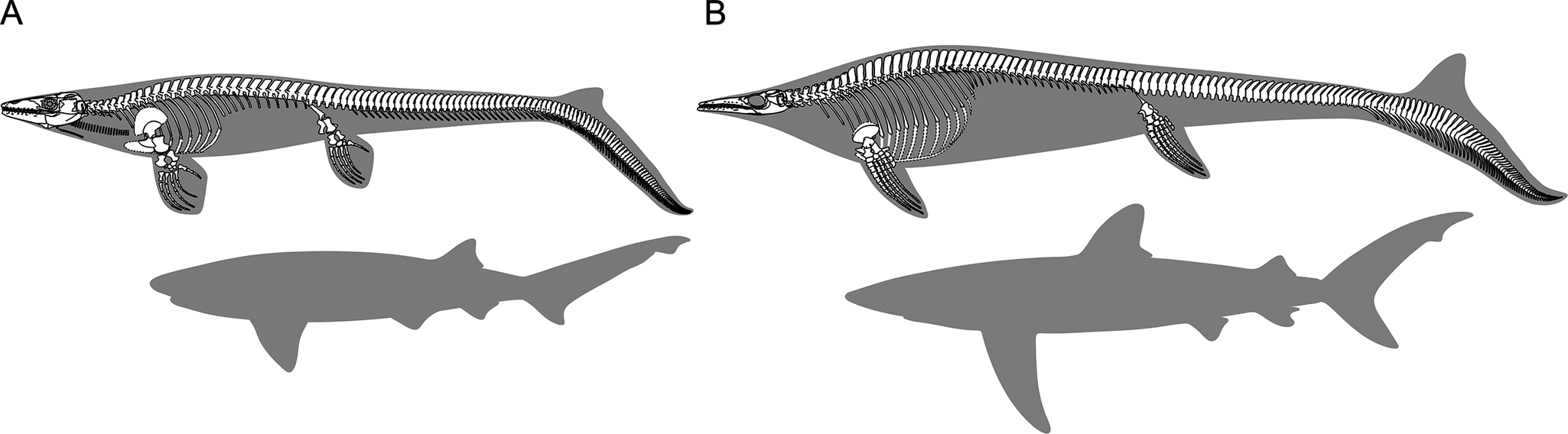
Reconstructed mosasaurians (top) compared against the body outline of extant sharks with similar tail fin morphologies (bottom). Specimens are not drawn to scale; instead, the size of the sharks has been adjusted for correspondence of tail fin length. A, Platecarpus tympaniticus (modified from Lindgren et al. Reference Lindgren, Caldwell, Konishi and Chiappe2010: fig. 8B) and the broadnose sevengill shark (Notorynchus cepedianus). B, Plotosaurus bennisoni (modified from Lindgren et al. Reference Lindgren, Jagt and Caldwell2007: fig. 3B) versus the blue shark (Prionace glauca).

The tail fin morphology of the small-sized hydropedal mosasaurian Eonatator is similar to smaller carangiform sharks, such as the demersal sailback houndshark (Gogolia filewoodi). It presumably represents an early fully aquatic form, characterized by a relatively primitive morphology suited for shallow-water environments (Bardet and Pereda Suberbiola 2001; Cross et al. Reference Cross, Moon, Stubbs, Rayfield and Benton2022).
The predicted tail fin shapes of the larger mosasaurians are similar to those of typical carangiform sharks, with Mosasaurus being closer to large sharks inhabiting the open seas (e.g., the bigeye sand tiger shark, Odontaspis noronhai) than Clidastes, Eremiasaurus, and Tylosaurus. Morphospace distribution indicates that the tail fin morphology of Plotosaurus is close to that of sharks with more oceanic habits, such as the blue shark (Prionace glauca) (Fig. 7B). Our new reconstruction further offers indirect evidence that Plotosaurus may have been a pelagic cruiser, highly adapted to open-marine environments (Camp Reference Camp1942; Lindgren et al. Reference Lindgren, Jagt and Caldwell2007, Reference Lindgren, Caldwell and Jagt2008, Reference Lindgren, Polcyn and Young2011; Ebert et al. Reference Ebert, Dando and Fowler2021; Cross et al. Reference Cross, Moon, Stubbs, Rayfield and Benton2022).
It is noteworthy that ERMNH HFV 197 (Prognathodon sp.)—the only known fossil of a derived mosasaurian with a preserved tail fin outline—was inferred to represent a juvenile individual by Lindgren et al. (Reference Lindgren, Kaddumi and Polcyn2013). In similarity with the ontogenetic development of the caudal fin in some sharks and ichthyosaurs, Prognathodon may have exhibited positive allometric tail fin growth, to suggest more efficient locomotor performance in adult specimens (Lindgren et al. Reference Lindgren, Kaddumi and Polcyn2013; Fu et al. Reference Fu, Hammerschlag, Lauder, Wilga, Kuo and Irschick2016; Seamone et al. Reference Seamone, Sternes, McCaffrey, Tsao and Syme2024). However, it has conversely been proposed that juvenile mosasaurians could achieve faster and more effective swimming than adults (Carpenter Reference Carpenter2017). These conflicting views necessitate further investigations into the ontogenetic growth of the mosasaurian tail fin.
Swimming Modes of Mosasaurians
It should be noted that although derived mosasaurians likely had tail fins that were superficially similar to those of pelagic sharks, they nonetheless exhibit notable differences in, for example, overall body proportions (Fig. 7). Specifically, mosasaurians differ from sharks in several key anatomical features, including: (1) a mobile cervical region; (2) a longer pre-fluke caudal region; (3) absence of an anal fin; and (4) lack of a dorsal fin in most, if not all, species (but see also Konishi et al. Reference Konishi, Ohara, Misaki, Matsuoka, Street and Caldwell2023). These dissimilarities indicate that although mosasaurians were comparable to sharks in certain aspects, differences in, for example, locomotor performance still could have existed.
Complexity of Tail Fin Evolution in Mosasauria
Figure 8 illustrates evolutionary changes occurring in the tail fin of mosasaurians. The integration of fluke reconstructions and phylogenetic studies in mosasaurine mosasaurians is consistent with the findings of Lindgren et al. (Reference Lindgren, Polcyn and Young2011), yet reveals a complex evolutionary pattern within Mosasauria: a bilobed fluke evolved independently at least four times (in mosasaurines, plioplatecarpines, tylosaurines, and halisaurines). Similar instances of multiple origins of specific aquatic adaptations are observed, for example, in extant sea snakes (Hydrophiinae and Laticaudinae) (Sanders et al. Reference Sanders, Rasmussen and Elmberg2012). This pattern, supported by the evidence of convergent evolution in mosasaurian limbs, corroborates the hypothesis of multiple transitions from semiaquatic to fully pelagic lifestyles in derived mosasaurians (Makádi et al. Reference Makádi, Caldwell and Ősi2012; Mekarski Reference Mekarski2017).
Evolution of tail fin shape among various mosasaurians. The simplified phylogenetic tree is based on data in Augusta et al. (Reference Augusta, Zaher, Polcyn, Fiorillo, Jacobs, Gower and Zaher2022), Polcyn et al. (Reference Polcyn, Bardet, Albright and Titus2023), and Zietlow et al. (Reference Zietlow, Boyd and Van Vranken2023). Images of the tail fin in Pontosaurus, Vallecillosaurus, and Prognathodon are modified from Caldwell (Reference Caldwell2006: fig. 2A), Smith and Buchy (Reference Smith and Buchy2008: fig. 2B, 3A), and Lindgren et al. (Reference Lindgren, Kaddumi and Polcyn2013: fig. 3E), respectively.

Acknowledgments
We thank A. Folie and J. Hagström at Institut Royal Sciences Naturelles de Belgique (Brussels, Belgium) for scanning Mosasaurus lemonnieri specimen IRSNB R29. We also thank D. Berthet at Musée des Confluences and N. Longrich at University of Bath for providing photos and additional information about Mosasaurus beaugei. Finally, we thank A. Zietlow and an anonymous reviewer for providing helpful feedback that improved this article.
Competing Interests
The authors declare no competing interests.
Data Availability Statement
Supplementary Material is available from the Zenodo Digital Repository: https://doi.org/10.5281/zenodo.17154012.
Author Contribution
Y.S.: Conceptualization; methodology; investigation; formal analysis; software; writing—original draft; writing—review and editing. J.L.: Conceptualization; supervision; writing—review and editing. Both authors approved the submitted version.







 Open access
Open access