


Introduction
Cambrian trilobites of Laurentia have been studied extensively for over a century (Walcott, Reference Walcott1886, Reference Walcott1890, Reference Walcott1916a, Reference Walcottb, Reference Walcott1917). However, there are many gaps in the geographic coverage, especially in northern Canada. Due to the remoteness of this region, with a few exceptions (Fritz, Reference Fritz1972, Reference Fritz1973, Reference Fritz1991; Pratt, Reference Pratt1992), most biostratigraphic information is at a reconnaissance level (e.g., Fritz, Reference Fritz1968, Reference Fritz1979; Dewing and Nowlan, Reference Dewing and Nowlan2012; Pyle, Reference Pyle2012) and organized using the traditional, Laurentian, genus-based biozonation. Now that more-refined, species-based zones are being erected in a number of localities (e.g., Sundberg, Reference Sundberg1994; McCollum and Sundberg, Reference McCollum and Sundberg2007), a higher resolution biostratigraphy is needed in support of local basin stratigraphy and long-distance correlation. The well-exposed, fossiliferous, lower and middle Cambrian formations of the eastern Mackenzie Mountains provide an untapped opportunity to develop a biozonation in this region (MacNaughton et al., Reference MacNaughton, Pratt and Fallas2013).
The Mount Clark and Mount Cap formations of the northern Canadian Cordillera record sedimentation in a shallow, epicontinental sea—one of several that rimmed Laurentia during the early and middle Cambrian (Aitken et al., Reference Aitken, Macqueen and Usher1973; Pyle, Reference Pyle2012). This basin is thought to have been partially isolated from the open ocean by a series of uplifted, Precambrian-cored arches (Dixon and Stasiuk, Reference Dixon and Stasiuk1998; MacLean, Reference MacLean2011). Regional stratigraphic studies suggest that the contact between the Mount Clark and Mount Cap formations is diachronous (MacNaughton et al., Reference MacNaughton, Pratt and Fallas2013; Serié et al., Reference Serié, Bergquist and Pyle2013). Early reports on reconnaissance collections of trilobites (Kobayashi, Reference Kobayashi1936; Fritz, Reference Fritz1969; Aitken et al., Reference Aitken, Macqueen and Usher1973) identified taxa indicating an age range from the Bonnia-Olenellus through Glossopleura zones (Cambrian Stage 4 and Wuliuan). Preliminary biostratigraphy by Pratt (in MacNaughton et al., Reference MacNaughton, Pratt and Fallas2013) documented that trilobites occurred sporadically in offshore facies that herein are assigned to the Mount Clark Formation, and more or less throughout the Mount Cap Formation. The purpose of this study is to provide a detailed taxonomic, biostratigraphic, and paleoecological account of the trilobite faunas from a series of sections cropping out in the eastern Mackenzie Mountains (Fig. 1).
Locality of study area in the eastern Mackenzie Mountains, Northwest Territories. (1) National map; (2) location in the Northwest Territories; (3) location in the eastern Mackenzie Mountains. Outcrop belts of the Mount Clark, Mount Cap, and Saline River formations are shaded in gray. Localities studied are indicated by black dots, which include Dodo Canyon (64.937525°N, 127.265209°W), Little Bear River (64.479199°N, 126.794262°W), Inlin Brook (64.282846°N, 16543348°W), Carcajou Falls (64.670639°N, 127.161682°W), and Grafe River (64.988407°N, 127.649351°W). The dashed line in (3) roughly delineates the edge of the Mackenzie Arch. Major Laramide anticlines indicated by crosses with arrows.
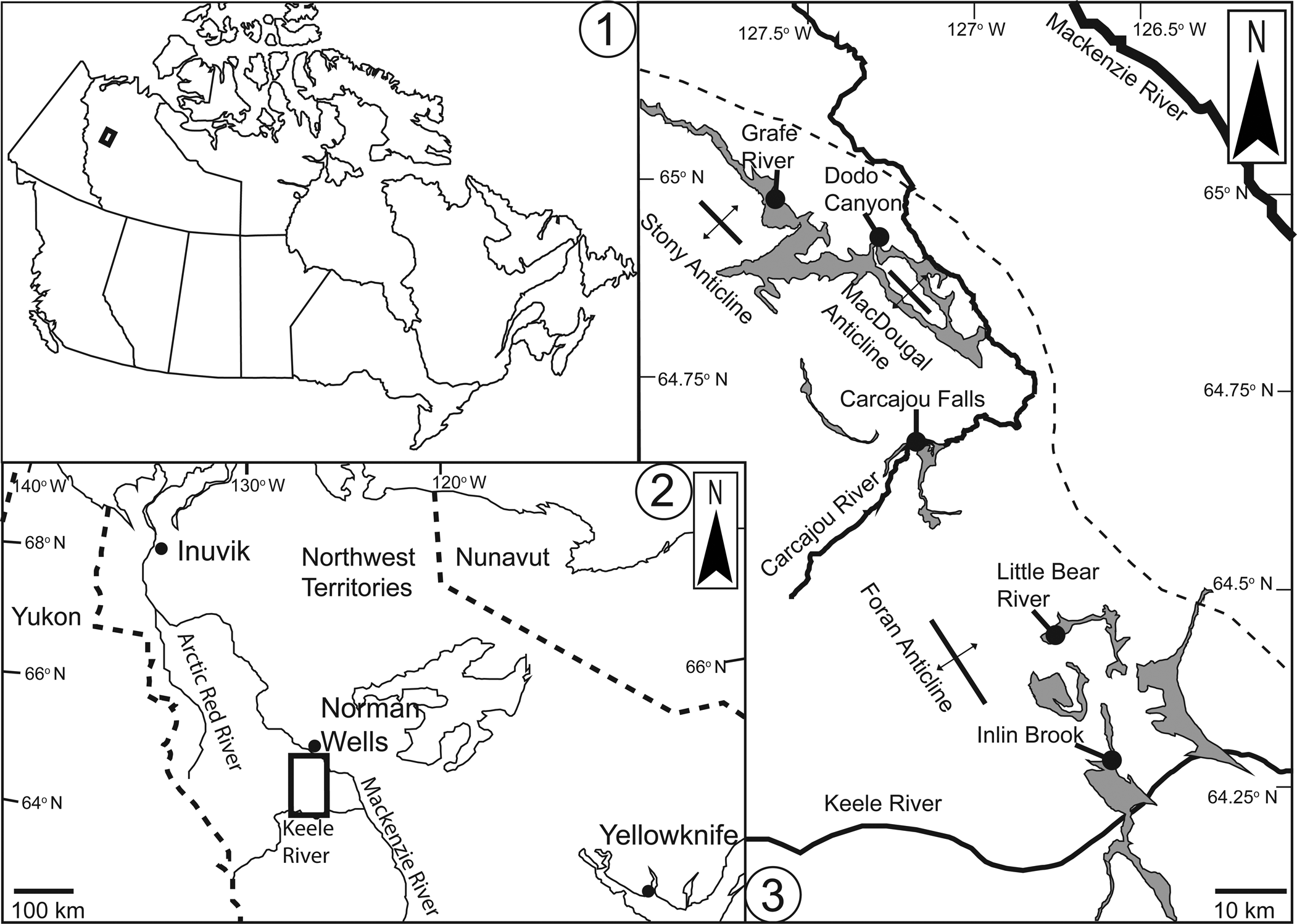
Geological setting
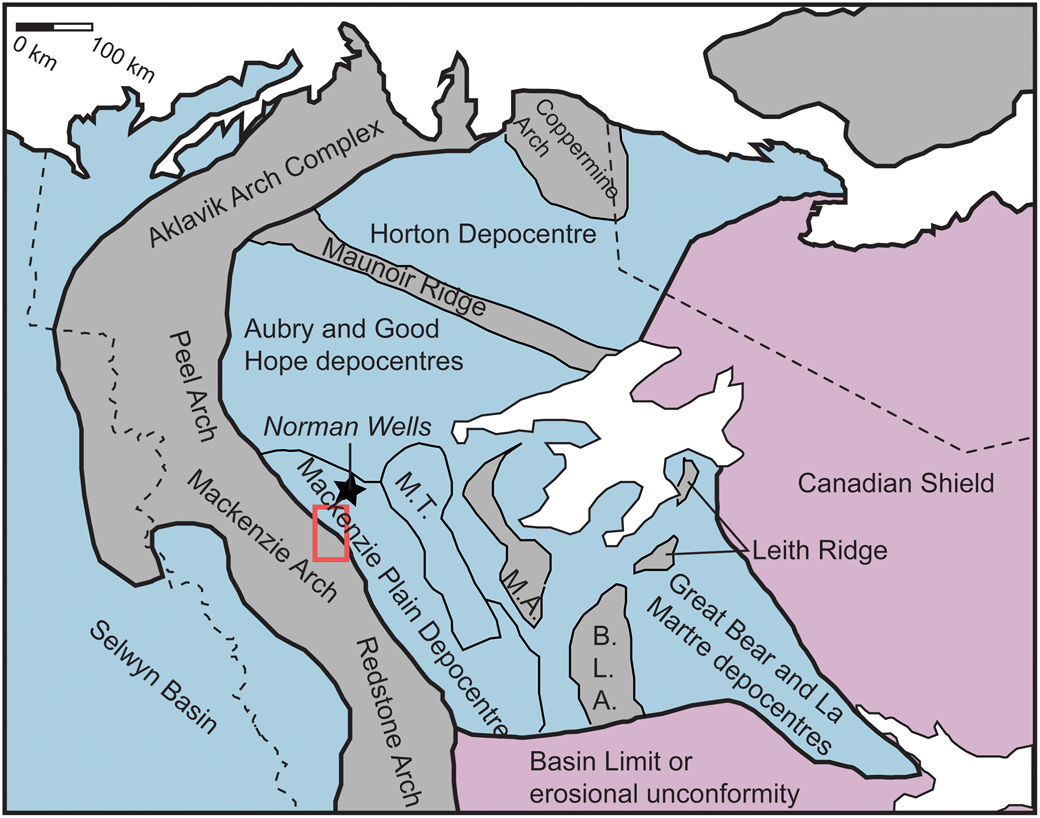
Strata of the lower and middle Cambrian basin are present in the subsurface of the Mackenzie Plain and northern Interior Plains (Dixon and Stasiuk, Reference Dixon and Stasiuk1998; MacLean, Reference MacLean2011; Herbers et al., Reference Herbers, MacNaughton, Timmer and Gingras2016; Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020), and are uplifted and exposed in the Franklin Mountains and the Canyon Ranges of the eastern Mackenzie Mountains (Aitken et al., Reference Aitken, Macqueen and Usher1973; Aitken and Cook, Reference Aitken and Cook1974). The basin was flanked to the modern-day west by the Mackenzie, Redstone, and Peel arches, to the north by the Aklavik Arch Complex and Coppermine Arch, and to the east by the paleoshoreline on the Canadian Shield (Aitken et al., Reference Aitken, Macqueen and Usher1973; Dixon and Stasiuk, Reference Dixon and Stasiuk1998) (Fig. 2). Convergent orientations of the Aklavik Arch Complex and Coppermine Arch indicate that they may have merged, which isolated the Cambrian basin to the north (Dixon and Stasiuk, Reference Dixon and Stasiuk1998), although similar stratigraphy on Victoria Island suggests a connection with basins in the Arctic Islands (Durbano et al., Reference Durbano, Pratt, Hadlari and Dewing2015; Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020). The southern extent of the Cambrian basin is poorly understood (Aitken and Cook, Reference Aitken and Cook1974; Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020). Subsurface data indicate that this basin was subdivided into depocenters by paleotopographic highs composed of Precambrian rocks (Dixon and Stasiuk, Reference Dixon and Stasiuk1998, MacLean, Reference MacLean2011; Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020) (Fig. 2).
Depositional limit of the inland Cambrian basin and other adjacent basins. Blue shading indicates areas where deposition occurred during the early to middle Cambrian. Positive tectonic elements during the Cambrian are shaded in gray. Precambrian basement is shaded in pink. M.T. = Mackenzie Trough, B.L.A. = Bulmer Lake Arch, M.A. = Mahony Arch. Study area within the red box. Modified from Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020.
Outcrops in the eastern Mackenzie Mountains represent the westernmost occurrences of the Mount Clark and Mount Cap formations. Strata onlap the eastern flank of the Mackenzie Arch, which is composed of deformed Neoproterozoic strata of the Mackenzie Mountains Supergroup (Aitken et al., Reference Aitken, Macqueen and Usher1973; Aitken and Cook, Reference Aitken and Cook1974; Turner and Long, Reference Turner and Long2012; MacNaughton et al., Reference MacNaughton, Pratt and Fallas2013; Long and Turner, Reference Long and Turner2014). The basal Cambrian formation in the basin is the Mount Clark Formation. At proximal sections in the Stony and Foran anticlines (Fig. 1.3), the Mount Clark Formation is composed of variably burrowed quartz arenite and argillaceous sandstone, whereas at distal sections in the MacDougal Anticline (Fig. 1.3) the formation is composed of burrowed sandstone and siltstone, silty mudstone, and limestone (Williams, Reference Williams1922, Reference Williams1923; Aitken et al., Reference Aitken, Macqueen and Usher1973; Aitken and Cook, Reference Aitken and Cook1974; Fallas and MacNaughton, Reference Fallas and MacNaughton2012; MacNaughton et al., Reference MacNaughton, Pratt and Fallas2013; Serié et al., Reference Serié, Bergquist and Pyle2013). These more distal strata have been treated by previous researchers as a basal, sandy unit of the Mount Cap Formation (e.g., Aitken et al., Reference Aitken, Macqueen and Usher1973; MacNaughton et al., Reference MacNaughton, Pratt and Fallas2013), but herein are assigned to the Mount Clark Formation on the basis of their stratigraphic position and overall sand-rich lithology. Although typical outcrop exposures of the Mount Clark Formation are dominated by sandstone (Aitken et al., Reference Aitken, Macqueen and Usher1973), strata assigned to the Mount Clark Formation in the subsurface also can contain a significant proportion of mudstone (Dixon and Stasiuk, Reference Dixon and Stasiuk1998; Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020).
The Mount Clark Formation is conformably overlain by the Mount Cap Formation, which consists of mudstone and lime mudstone with minor siltstone (Williams, Reference Williams1923; Aitken et al., Reference Aitken, Macqueen and Usher1973; Aitken and Cook, Reference Aitken and Cook1974; MacNaughton et al., Reference MacNaughton, Pratt and Fallas2013). Reconnaissance biostratigraphy of the Mount Clark and Mount Cap formations identified the Olenellus, Albertella, and Glossopleura zones. The apparent absence of the Plagiura-Poliella Zone was regarded as evidence for an unconformity (Fritz, Reference Fritz1969; Aitken and Cook, Reference Aitken and Cook1974), although this interpretation has been disputed (MacNaughton et al., Reference MacNaughton, Pratt and Fallas2013). Unconformably overlying the Mount Cap Formation in outcrop is the Saline River Formation, which is composed of mudstone, dolostone, and gypsum (Aitken et al., Reference Aitken, Macqueen and Usher1973; Aitken and Cook, Reference Aitken and Cook1974; Dixon and Stasiuk, Reference Dixon and Stasiuk1998; Serié et al., Reference Serié, Bergquist and Pyle2013; Bouchard and Turner, Reference Bouchard and Turner2017). The westward truncation of the Mount Cap Formation beneath the Saline River Formation indicates that the Mackenzie Arch was variably tectonically active during this time (Aitken et al., Reference Aitken, Macqueen and Usher1973; Aitken and Cook, Reference Aitken and Cook1974).
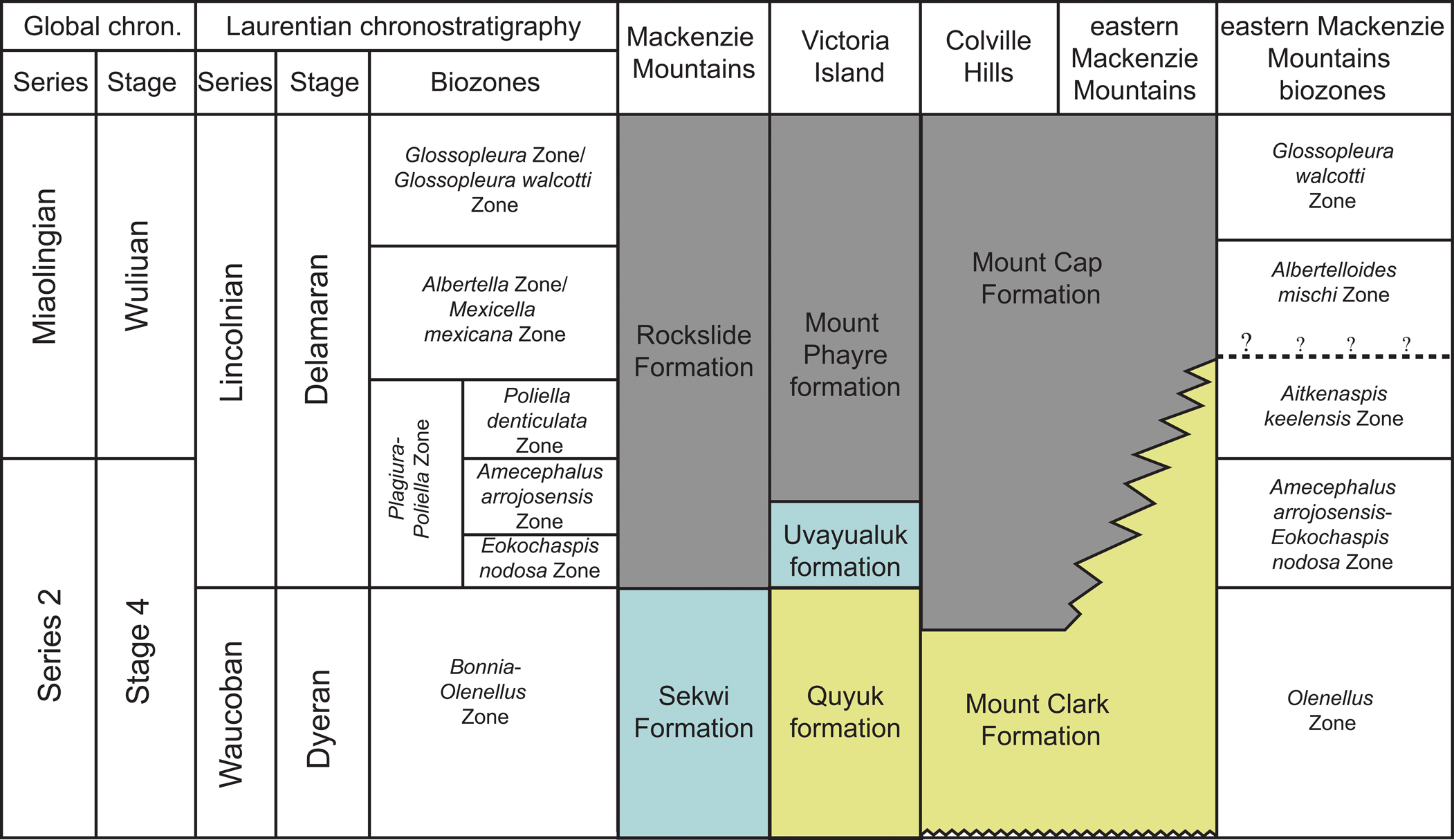
In the Mackenzie Plain to the northeast, the Mount Clark and Mount Cap formations are thicker, and the contact between the two formations is in the Olenellus Zone (Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020) (Fig. 3). On the western flank of the Mackenzie Arch, lower and middle Cambrian formations of the Selwyn Basin record a carbonate platform to slope setting on the passive continental margin (Pratt, Reference Pratt1989; Dilliard et al., Reference Dilliard, Pope, Coniglio, Hasiotis and Lieberman2010; Kimmig and Pratt, Reference Kimmig and Pratt2016). Limestones of the Sekwi Formation contain trilobites belonging to the Fallotaspis through Olenellus zones (Cambrian stages 3 and 4; Fritz, Reference Fritz1972), suggesting that the initial Cambrian transgression had flooded the western continental margin prior to the inland Cambrian basin. Conformably overlying the Sekwi Formation is the Rockslide Formation, with trilobites indicative of the Plagiura-Poliella through Bolaspidella zones (Fritz, Reference Fritz1979).
Lithostratigraphic correlation chart and biostratigraphic ages of Cambrian formations of the Mackenzie Mountains, Arctic Islands, Colville Hills, and eastern Mackenzie Mountains. Color of formations indicates the dominant lithology: yellow = dominantly sandstone, gray = dominantly mudstone, and blue = dominantly carbonate. Biostratigraphic and lithostratigraphic data from Fritz, Reference Fritz1972, Reference Fritz1979; Pratt, Reference Pratt1989; Palmer, Reference Palmer1998a; Durbano et al., Reference Durbano, Pratt, Hadlari and Dewing2015; Kimmig and Pratt; Reference Kimmig and Pratt2016; Sundberg et al., Reference Sundberg, Geyer, Kruse, McCollum, Pegel, Zylinska and Zhuravlev2016, Reference Sundberg, Karlstrom, Geyer, Foster, Hagadorn, Mohr, Schmitz, Dehler and Crossey2020; and Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020. Biostratigraphic framework of the eastern Mackenzie Mountains based on Fritz (Reference Fritz1969), Aitken et al. (Reference Aitken, Macqueen and Usher1973), and this work. Biostratigraphy of Victoria Island is currently under investigation by Pratt.
Materials and methods
Fieldwork was carried out at five sites in the Canyon Ranges: Dodo Canyon, Little Bear River, Inlin Brook, Carcajou Falls, and Grafe River (Fig. 4; see Handkamer, Reference Handkamer2020, for measured section data), ~50 km southwest of Norman Wells (NTS 96D) (Fig. 1.3). Material was also collected by Pratt in 2011 from Dodo Canyon and Carcajou Falls, and by MacNaughton in 2012 from three additional measured sections: MWB-12-s04 and MWB-12-s05 located northwest of Dodo Canyon, and 12-MWB-s06 located west of Little Bear River (see MacNaughton, Reference MacNaughton2021, for stratigraphic horizons of collections). All specimens were coated ammonium chloride sublimate and photographed, mainly with a macro lens, and in the case of small specimens, with a bellows attachment. Figures were assembled using Adobe Photoshop and Adobe Illustrator, and brightness and contrast were adjusted as needed to increase image quality. Simple rarefaction plots were generated in Past4.0 and modified visually using Illustrator, and population counts were tabulated in Microsoft Excel.
Outcrop views of the Mount Clark, Mount Cap, and Neoproterozoic formations of the Canyon Ranges, Northwest Territories. Solid lines indicate lithostratigraphic boundaries and dashed lines indicate biostratigraphic boundaries. (1) Section at Dodo Canyon. Stratal thickness = 73.9 m. The Mount Cap Formation-Saline River Formation contact is exposed farther downriver (in the distance). Central talus cone covers normal faults. (2) Section at Inlin Brook with the units exposed in an overturned anticline. Stratal thickness of Cambrian formations = 18.3 m. The top of the section is marked where debris covers the Mount Cap Formation. (3) Upper part of the section exposed at Carcajou Falls. Stratal thickness = 28.2 m. The section was measured upriver from this cliff (behind the observer).

Repositories and institutional abbreviations
All figured specimens are deposited in the type collection of the Geological Survey of Canada, Ottawa. Specimens not illustrated are housed at Geological Survey of Canada, Calgary. Institutional abbreviations are: GSC = Geological Survey of Canada; USNM = U.S. National Museum of Natural History, Smithsonian Institution, Washington DC; DMNH = Denver Museum of Natural History; and MGUH = Natural History Museum, Copenhagen, Denmark.
Sedimentology
Several reports have included measured sections through the Mount Clark and Mount Cap formations in the eastern Mackenzie Mountains (Aitken et al., Reference Aitken, Macqueen and Usher1973; Pyle and Gal, Reference Pyle and Gal2009; Serié et al., Reference Serié, Bergquist and Pyle2013; Bouchard and Turner, Reference Bouchard and Turner2017; Handkamer, Reference Handkamer2020; MacNaughton, Reference MacNaughton2021). Regionally, facies of the Mount Clark and Mount Cap formations record the evolution from a nearshore to deeper water environment (MacNaughton et al., Reference MacNaughton, Pratt and Fallas2013; Herbers et al., Reference Herbers, MacNaughton, Timmer and Gingras2016; Handkamer, Reference Handkamer2020; Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020). In the present study area, low-angle cross-bedded, wavy bedded, and wave-rippled sandstone with minor glauconite of the Mount Clark Formation indicate a nearshore setting (Handkamer, Reference Handkamer2020). The dominance of storm-induced features, with rare sand bars, suggest a relatively low-energy storm-dominated shoreline with weak tidal influence. In distal sections of the Mount Clark Formation, interbedded sandstone and siltstone with wave ripples, gutter casts, local hummocky cross-stratification, and minor unidirectional ripples (Handkamer, Reference Handkamer2020) record fluctuating conditions produced by storms in the offshore setting. Interbedded, rippled sandy grainstone and argillaceous lime mudstone (Handkamer, Reference Handkamer2020) represent similar conditions during episodes of localized carbonate production. Strata of the Mount Clark Formation are dominated by a deposit-feeding ichnofauna (Handkamer, Reference Handkamer2020). The scarcity of burrows cast by tempestites in the offshore setting is attributed to scour during deposition. A vertical, suspension-feeding ichnofauna is dominant in strata representing upper-shoreface deposits (Handkamer, Reference Handkamer2020), due to organic productivity in the water column.
The contact between the Mount Clark and Mount Cap formations is interpreted to be a flooding surface. Variably calcareous and fossiliferous silty mudstone, clay-rich mudstone, and minor siltstone of the Mount Cap Formation (Handkamer, Reference Handkamer2020) are interpreted as deeper water hemipelagites. Carbonate strata are common especially at Dodo Canyon and Carcajou Falls. Carbonate lithologies include: (1) thin-bedded, rippled sandy grainstone; (2) wavy bedded and rippled, argillaceous or sandy dolostone; (3) thin- to medium-bedded, bioclastic and peloidal wackestone; (4) variably burrowed, wavy bedded lime mudstone; and (5) interbedded, burrowed lime mudstone and siliciclastic mudstone with minor bioclastic grainstone lenses and bioclastic wackestone (Handkamer, Reference Handkamer2020). The first two are interpreted to be higher energy, shallow-subtidal ramp sediments, the third and fourth lower energy, deeper subtidal ramp sediments, and the fifth intercalated, deeper subtidal lime mud and siliciclastic mud, punctuated by minor tempestites. The contrast between the thick carbonate accumulations at Dodo Canyon and Carcajou Falls versus the minor proportions at Little Bear River, Inlin Brook, and Grafe River, is considered to record localized carbonate production in a well-oxygenated setting. In the Mount Cap Formation, the greater proportion of carbonate strata in the eastern Mackenzie Mountains compared to the Colville Hills (Handkamer, Reference Handkamer2020; Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020) indicates that there was a more productive, but geographically localized, carbonate factory in this region. The Colville Hills were closer to the basin's eastern paleoshoreline and may have been supplied with a more continuous amount of siliciclastic mud compared to the intermittently tectonically active Mackenzie Arch. In outcrops studied for this report, horizontal, deposit-feeding burrows are rare, either preserved by pyrite or on the surfaces of carbonate concretions in silty mudstone. These surfaces indicate well-oxygenated surface sediments, whereas the apparent absence of burrows in mudstone is regarded as taphonomic. Variably numerous trilobites, hyoliths, and linguliformean brachiopods comprise the benthic, shelly biota (Handkamer, Reference Handkamer2020) that developed during times of well-oxygenated conditions.
Biostratigraphy
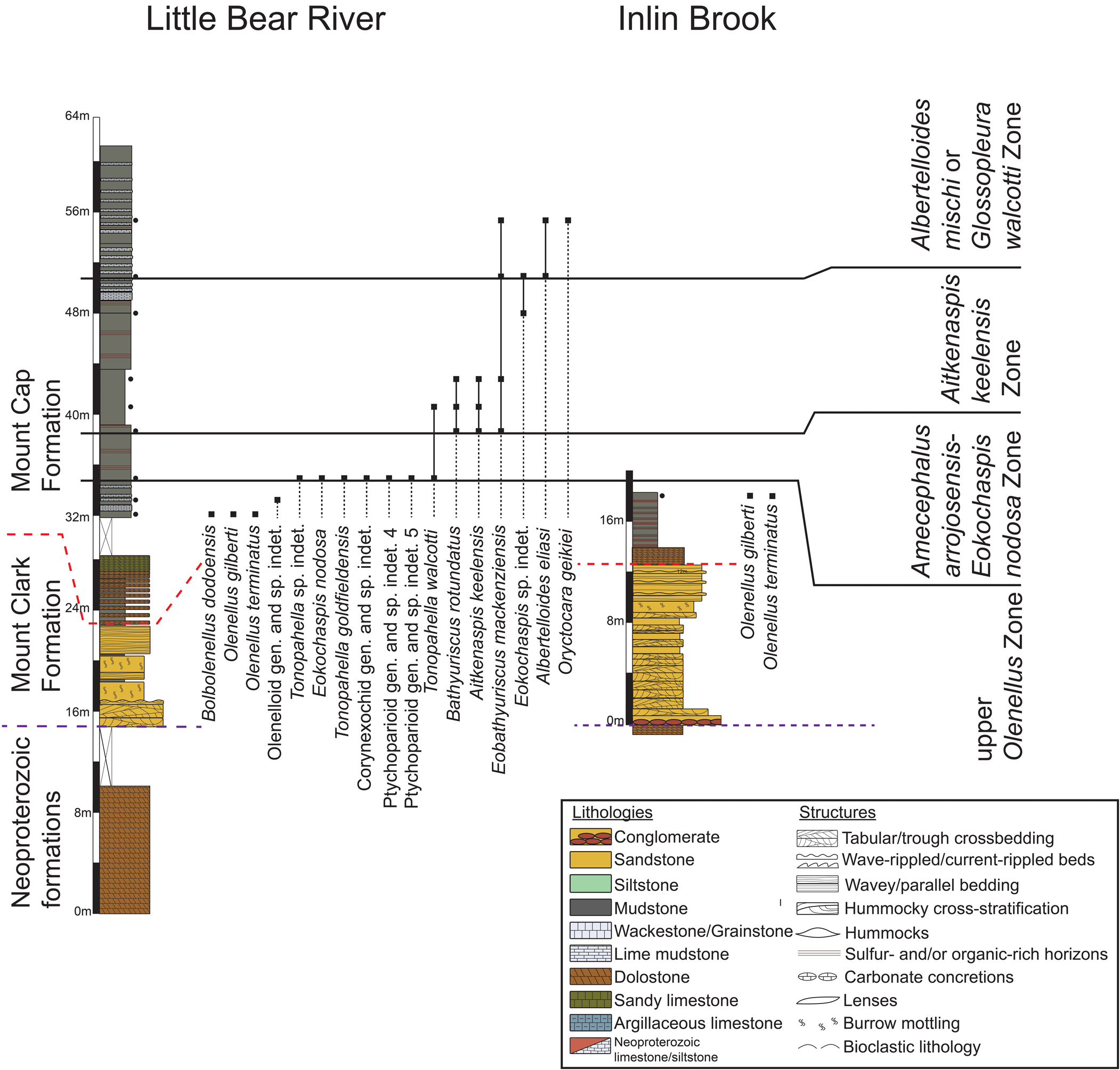
Trilobites from the Mount Clark and Mount Cap formations indicate an age spanning the Laurentian upper Dyeran and Delamaran stages of the upper Waucoban and Lincolnian series, respectively, and the global upper Stage 4 and Wuliuan stages of the upper Series 2 and lower Miaolingian series (Fig. 3). Some of the traditional Laurentian zones are recognized (Rasetti, Reference Rasetti1951; Lochman-Balk and Wilson, Reference Lochman-Balk and Wilson1958; Robison, Reference Robison1976; Palmer and Halley, Reference Palmer and Halley1979; Palmer, Reference Palmer1998a, Reference Palmerb; Sundberg and McCollum, Reference Sundberg and McCollum2000, Reference Sundberg and McCollum2003a, Reference Sundberg and McCollumb; McCollum and Sundberg, Reference McCollum and Sundberg2007; Webster, Reference Webster2011b; Pratt and Bordonaro, Reference Pratt and Bordonaro2014; Sundberg et al., Reference Sundberg, Karlstrom, Geyer, Foster, Hagadorn, Mohr, Schmitz, Dehler and Crossey2020). However, two new zones (the Aitkenaspis keelensis Zone and the Albertelloides mischi Zone) are established due to a lack of typical Laurentian taxa in part of the Mount Cap Formation. These zones are, in ascending order, the upper Olenellus, Amecephalus arrojosensis-Eokochaspis nodosa, Aitkenaspis keelensis, Albertelloides mischi, and Glossopleura walcotti zones (Figs. 5, 6).
Measured sections of the Mount Clark and Mount Cap formations exposed at Dodo Canyon, Carcajou Falls, and Grafe River. Black dots beside stratigraphic sections denote sampling horizons. The legend for the stratigraphic columns is in Figure 6. The purple dashed line denotes the lithostratigraphic contact between the Mount Clark Formation and underlying Neoproterozoic formations (Handkamer, Reference Handkamer2020). The red dashed line denotes the Mount Clark Formation-Mount Cap Formation contact.
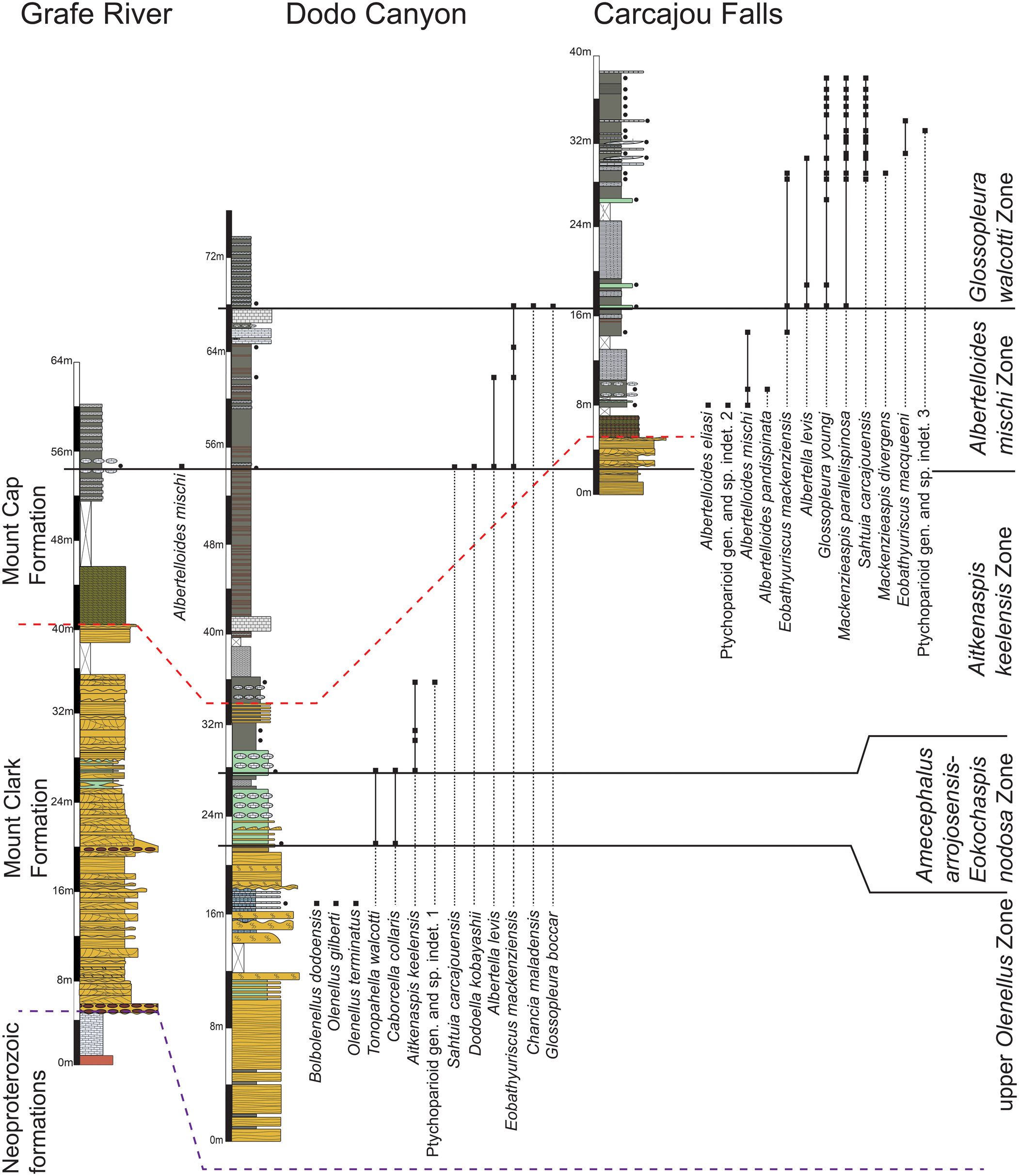
Measured sections of the Mount Clark and Mount Cap formations exposed at Little Bear River and Inlin Brook. Black dots beside stratigraphic sections denotes sampling horizons. The purple dashed line denotes the lithostratigraphic contact between the Mount Clark Formation and underlying Neoproterozoic formations (Handkamer, Reference Handkamer2020). The red dashed line denotes the Mount Clark Formation-Mount Cap Formation contact.
The base of the Olenellus Zone of the Dyeran (Palmer, Reference Palmer1998a) is recorded by the first appearance datum (FAD) of an olenellid, indicating the traditional marker for the uppermost early Cambrian of Laurentia. Olenellus gilberti Meek in White, Reference White1874, and O. terminatus Palmer, Reference Palmer1998b, which occur in the Mount Clark and Mount Cap formations, span the two uppermost zones of the species-based biozonation in the Great Basin (Webster, Reference Webster2011b), the Bolbolenellus euryparia Zone and overlying Nephrolenellus multinodus Zone. This prevents the assignment to either of these zones, but the lack of an unconformity above the fossil-bearing horizons possibly indicates the N. multinodus Zone.
The overlying zone is the undifferentiated Amecephalus arrojosensis-Eokochaspis nodosa Zone. The Eokochaspis nodosa Zone and the younger Amecephalus arrojosensis Zone were established as separate zones in the Pioche Formation (Sundberg and McCollum, Reference Sundberg and McCollum2000). However, in the La Laja Formation of the Precordillera of western Argentina, A. arrojosensis and E. nodosa are present in reverse order, suggesting that these two taxa co-existed and are not temporally exclusive (Pratt and Bordonaro, Reference Pratt and Bordonaro2014). Furthermore, E. nodosa and Tonopahella walcotti (Sundberg and McCollum, Reference Sundberg and McCollum2000), a species previously recorded from the Amecephalus arrojosensis Zone, occur in the same bed at Little Bear River. The base of the A. arrojosensis-E. nodosa Zone in the Mount Cap Formation is probably synchronous with the base of the E. nodosa Zone in the Pioche, Emigrant, and Monola formations (Sundberg and McCollum, Reference Sundberg and McCollum2000, Reference Sundberg and McCollum2003b), and the Plagiura-Poliella Zone elsewhere (Lochman-Balk and Wilson, Reference Lochman-Balk and Wilson1958; Robison, Reference Robison1976). It is at this time when simple ptychoparioids (referable as ‘kochaspids’) diversified (Sundberg and McCollum, Reference Sundberg and McCollum2000).
The succeeding zone in the Mount Clark and Mount Cap formations is the Aitkenaspis keelensis Zone, established herein. The base of this zone is inferred to be equivalent to the base of the Poliella denticulata Zone in restricted-shelf settings (Sundberg and McCollum, Reference Sundberg and McCollum2003a) and the Oryctocephalus indicus Zone in open-shelf settings in the Great Basin (Sundberg and McCollum, Reference Sundberg and McCollum2003b), due to its position above the A. arrojosensis-E. nodosa Zone and lack of any erosional unconformity at the base. The temporal relationships of the A. keelensis Zone relative to the Wenkchemnia-Stephenaspis and Plagiura-Kochaspis zones of the Mount Whyte Formation of Alberta and British Columbia (Rasetti, Reference Rasetti1951), or the upper Amecephalus arrojosensis-Eokochaspis nodosa and lower Mexicella mexicana zones of La Laja Formation, are not well constrained. The A. keelensis and equivalent zones record when corynexochids became significant components of Laurentian faunas.
The new Albertelloides mischi Zone overlies the Aitkenaspis keelensis Zone in the Mount Cap Formation. This zone is recognized by the FAD of Albertelloides mischi Fritz, Reference Fritz1968, which appears to be synchronous with the FADs of A. pandispinata Fritz, Reference Fritz1968, and A. eliasi Handkamer and Pratt, n. sp. In the Great Basin, A. pandispinata is present close to the base of the Albertellina aspinosa Subzone of the Mexicella mexicana Zone, whereas Albertelloides mischi is likely present higher up in the Albertella highlandensis Subzone (Fritz, Reference Fritz1968; Eddy and McCollum, Reference Eddy and McCollum1998; McCollum and Sundberg, Reference McCollum and Sundberg2007). These two species co-occur in the Mount Cap Formation, so the age relationship of the base of the Albertelloides mischi Zone with respect to that of the Mexicella mexicana Zone is uncertain. Alternatively, the co-occurrence of Oryctocara geikiei Walcott, Reference Walcott1908b, and Albertelloides eliasi Handkamer and Pratt, n. sp., within the uppermost strata at Little Bear River may suggest that part of this zone is equivalent to the lower Glossopleura walcotti and lower Ptychagnostus praecurrens zones (Sundberg and McCollum, Reference Sundberg and McCollum1997, Reference Sundberg and McCollum2003a, Reference Sundberg and McCollumb). Two biofacies are represented in the Albertelloides mischi Zone: the higher diversity and abundance Albertelloides biofacies is present at Carcajou Falls, Little Bear River, and Grafe River in variably calcareous, silty mudstone and lime mudstone, whereas the sparse biofacies is present at Dodo Canyon in organic-rich mudstone. The contrast between the two biofacies correlates with shoreline trajectory, in that diversity and abundance decrease farther offshore.
The youngest zone recognized in the Mount Cap Formation is the Glossopleura walcotti Zone, the base of which is indicated by the FADs of G. boccar (Walcott, Reference Walcott1916b) and G. youngi Handkamer and Pratt, n. sp. In the southern Canadian Rocky Mountains, the oldest species of Glossopleura is G. skokiensis (? = G. walcotti; McCollum and Sundberg, Reference McCollum and Sundberg2007), indicating that the base of the G. walcotti Zone is near the top of the Cathedral Formation (Rasetti, Reference Rasetti1951; McCollum and Sundberg, Reference McCollum and Sundberg2007). The basal beds of the overlying Stephen Formation contain G. boccar, suggesting that this species occurs slightly above the base of the G. walcotti Zone. Therefore, the base of the G. walcotti Zone in the Mount Cap Formation is considered slightly younger than that in the Carrara Formation, Lyndon Limestone, Chisholm Formation, upper Emigrant Formation, and possibly the Bright Angel Shale of the Great Basin (Palmer and Halley, Reference Palmer and Halley1979; Eddy and McCollum, Reference Eddy and McCollum1998; Sundberg, Reference Sundberg2005, Reference Sundberg2018; Foster, Reference Foster2011). It is also considered younger than the base of the Glossopleura Zone of the Cathedral Formation (Rasetti, Reference Rasetti1951), although equivalent to the Glossopleura Zone of the Stephen and Snake Indian formations of the Canadian Rocky Mountains (Rasetti, Reference Rasetti1951; Mountjoy and Aitken, Reference Mountjoy and Aitken1978; Fletcher and Collins, Reference Fletcher and Collins1998). The base of the G. walcotti Zone in the Mount Cap Formation is also probably younger than the G. walcotti Zone of the Cape Wood Formation of Greenland (Poulsen, Reference Poulsen1927) and the G. walcotti Zone of the upper La Laja Formation of the Precordillera (Foglia and Vaccari, Reference Foglia and Vaccari2010).
Biostratigraphic zonation of the Mount Clark and Mount Cap formations indicates that: (1) the formations are partly time equivalent; (2) the contact between the Mount Clark Formation and the Mount Cap Formation, interpreted as a flooding surface, is diachronous; and (3) there is no biostratigraphic evidence for an unconformity within the Mount Cap Formation as was postulated by Fritz (Reference Fritz1969, also in Aitken et al., Reference Aitken, Macqueen and Usher1973), which is consistent with the lack of any physical evidence of an unconformity. Flooding began during the upper Olenellus Zone in the southeastern part of the study area and continued into the Albertelloides mischi Zone in the northwestern part of the study area.
Paleoecology
Trilobites co-occur with linguliformean brachiopods in siltstone, limestone, and mudstone, and co-occur with hyoliths in mudstone. Soft-bodied faunal elements are present at Little Bear River, which include anomalocarids, bivalved arthropods, chancelloriids, and small carbonaceous fossils of planktonic acritarchs, crustaceans, and lophotrochozoans (Butterfield and Nicholas, Reference Butterfield and Nicholas1996; Harvey and Butterfield, Reference Harvey and Butterfield2011). The rare occurrence of protaspids and meraspids of Sahtuia carcajouensis Handkamer and Pratt, n. gen. n. sp., and Eobathyuriscus mackenziensis Handkamer and Pratt, n. gen. n. sp., in mudstone and lime mudstone, respectively, indicates that these species were able to reproduce in the environments in which those facies were deposited.
Trilobite faunas
In Laurentia, trilobite faunas of the inner detrital belt are composed of endemic taxa, while those of the outer detrital belt contain globally distributed genera (Robison, Reference Robison1976; see also Pegel, Reference Pegel2000). In the Mount Cap Formation, the faunas are overall less diverse than those in other middle Cambrian formations of Laurentia, particularly at higher taxonomic levels in the Albertelloides mischi and Glossopleura walcotti zones. Genera and species present are typical of a restricted-shelf setting (Robison, Reference Robison1976; Palmer and Halley, Reference Palmer and Halley1979; Eddy and McCollum, Reference Eddy and McCollum1998), and the closest faunal analogs are those present in the inner detrital belt facies of the Great Basin. Faunas of the Albertella/Mexicella mexicana and Glossopleura walcotti zones of the Pioche, Chisholm, and inner-shelf Carrara formations are comparable (Fritz, Reference Fritz1968; Palmer and Halley, Reference Palmer and Halley1979; Eddy and McCollum, Reference Eddy and McCollum1998; Sundberg, Reference Sundberg2005). Rarefaction of trilobite collection data from Mount Cap Formation and these three formations indicate that they represent a reasonable approximation of the original trilobite biotas (Fig. 7.1).
Quantitative paleoecology of the trilobite faunas from the Albertelloides mischi/Albertella/Mexicella mexicana and Glossopleura walcotti/Glossopleura zones of the Mount Cap, Pioche, Chisholm, and Carrara formations. (1) Simple rarefaction curves of collections. Dashed lines denote 95% confidence intervals. (2) Abundance of trilobite groups. Individual cranidia and pygidia, partially complete specimens, and complete specimens were all given equal value in the abundance count. Counts from the Great Basin are compiled from (a) Fritz, Reference Fritz1968; (b) Palmer and Halley, Reference Palmer and Halley1979; (c) Eddy and McCollum, Reference Eddy and McCollum1998; and (d) Sundberg, Reference Sundberg2005.
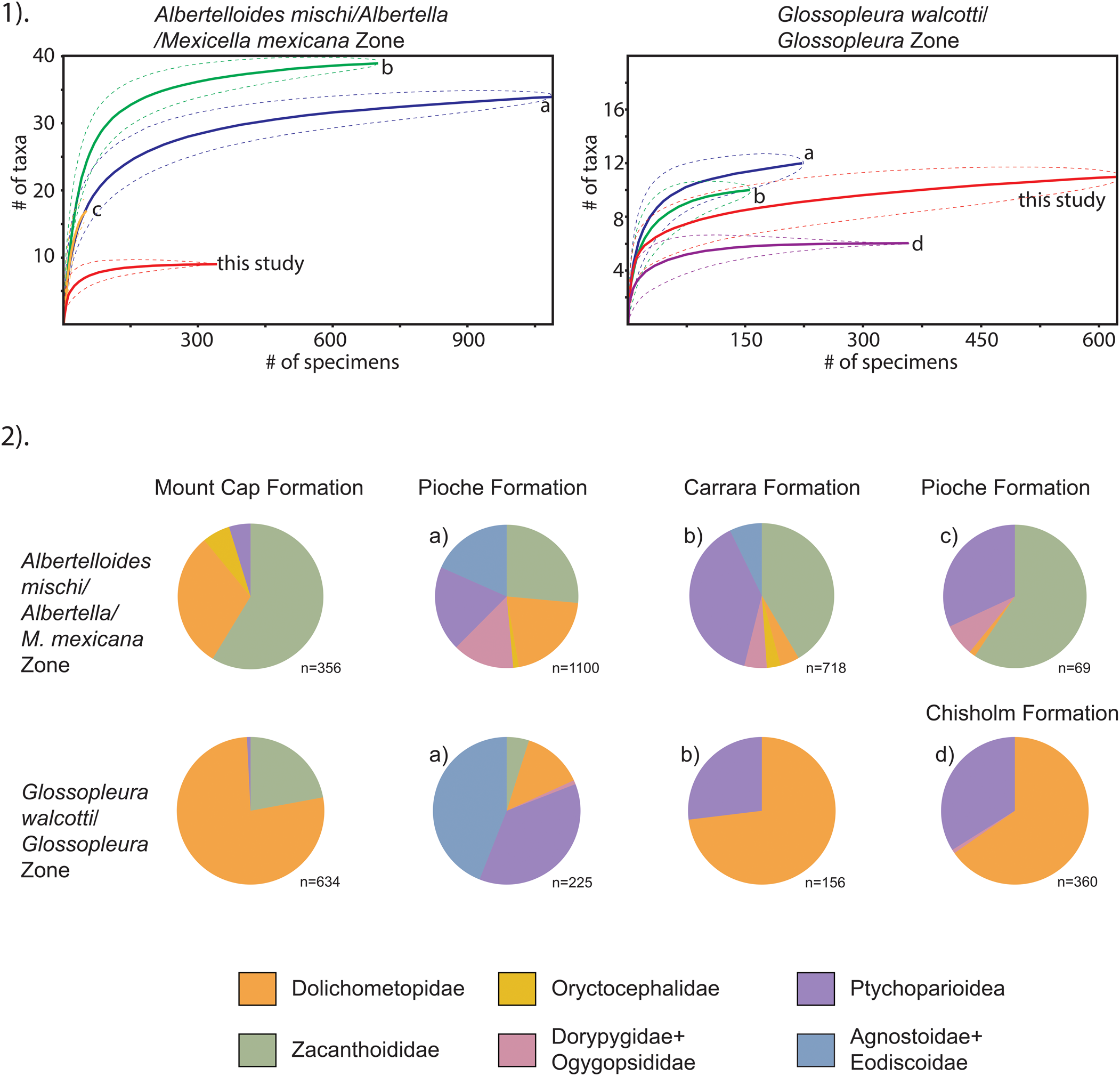
Dolichometopids and zacanthoidids are relatively abundant in the Mount Cap Formation, whereas ptychoparioids and oryctocephalids are rare, and agnostoids, eodiscoids, dorypygids and ogygopsidids are absent (Fig. 7.2). The dominance of dolichometopids and zacanthoidids increases in younger zones, and ptychoparioids virtually disappear. The absence of agnostoids and eodiscoids and paucity of oryctocephalids are to be expected within a semi-enclosed basin far from deep-water settings where these taxa tend to be common (Robison, Reference Robison1976; Whittington, Reference Whittington1995; Sundberg and McCollum, Reference Sundberg and McCollum1997). The lack of dorypygids and ogygopsidids contrasts with other areas where these families are present, such as in the southern Canadian Rocky Mountains (Rasetti, Reference Rasetti1951), eastern Alaska (Palmer, Reference Palmer1968), the Great Basin (Fritz, Reference Fritz1968; Robison, Reference Robison1976; Palmer and Halley, Reference Palmer and Halley1979; Eddy and McCollum, Reference Eddy and McCollum1998; Sundberg, Reference Sundberg2005, Reference Sundberg2018; Foster, Reference Foster2011; Robison and Babcock, Reference Robison and Babcock2011), the Precordillera (Bordonaro et al., Reference Bordonaro, Banchig, Pratt and Raviolo2008), and Greenland (Poulsen, Reference Poulsen1927; Babcock, Reference Babcock1994). Also, these two groups are present in both inner and outer detrital belt faunas. Environmental conditions in the Mount Cap Formation were favorable, or tolerable, to zacanthoidids and dolichometopids, but less so to ptychoparioids, and not so for dorypygids and ogygopsidids. Facies in the Pioche (Merriam and Palmer, Reference Merriam and Palmer1964; Eddy and McCollum, Reference Eddy and McCollum1998) and Chisholm formations (Sundberg, Reference Sundberg2005), broadly similar to those in the Mount Cap Formation, suggest environmental preference was independent of substrate type or physical sedimentary processes, such as bottom turbulence. Three possible paleoecological stressors include variation in water temperature, salinity, or bottom-water oxygenation. Variations in any of these could be expected within a semi-enclosed basin, but to single them out specifically has not yet been achieved.
Paedomorphic trilobites
Several taxa unique to the Mount Cap Formation of the eastern Mackenzie Mountains exhibit paedomorphic anatomical features, interpreted as arising due to heterochronic changes during ontogeny (McNamara, Reference McNamara1986a, Reference McNamarab). These species are Dodoella kobayashii Handkamer and Pratt, n. gen. n. sp., and Sahtuia carcajouensis Handkamer and Pratt, n. gen. n. sp., in the Albertelloides mischi Zone at Dodo Canyon, and S. carcajouensis n. gen. n. sp., Mackenzieaspis parallelispinosa Handkamer and Pratt, n. gen., n. sp., and M. divergens Handkamer and Pratt, n. gen., n. sp., in the Glossopleura walcotti Zone at Carcajou Falls. Non-paedomorphic relatives of all four species were collected from the Mount Cap Formation as well: Albertella levis, Eobathyuriscus mackenziensis Handkamer and Pratt, n. gen., n. sp., and Albertelloides eliasi Handkamer and Pratt, n. sp., or A. mischi, respectively, which are regarded as ancestral due to their similarity to typical zacanthoidids and dolichometopids of Laurentia (see McNamara, Reference McNamara1982). These ancestor-descendant relationships are supported by many subtle morphological similarities (see remarks on new genera). Identification of the ancestral form is a criterion for recognizing the occurrence of paedomorphosis (McNamara, Reference McNamara1986a). Compared to the ancestral apaedomorphs, paedomorphic species have: (1) fewer thoracic segments; (2) a larger pygidium exhibiting more furrows; and (3) shorter thoracic spines. Exoskeletons of the holaspids of both paedomorphic and apaedomorphic species are broadly similar in their sagittal length. Morphological differences indicate that only the development of the thorax was retarded, interpreted as arising from post-displacement. Paedomorphosis is demonstrated in three separate lineages in the Mount Cap Formation due to the presence of three separate paedomorph-apaedomorph pairs. This indicates that the development was likely environmentally provoked, rather than from random mutation or genetic drift. Almost all the paedomorphic specimens were collected from 20 meters of abundantly fossiliferous mudstone at Carcajou Falls, but the reason for this distribution is unclear.
Systematic paleontology
New taxonomic names are authored by Handkamer and Pratt. The descriptors “long” and “short” refer to sagittal and exsagittal measurements, while “wide” and “narrow” refer to transverse measurements. In the materials sections, a “nearly complete exoskeleton” refers to specimens lacking either sclerites of the cephalon, thorax, or pygidium.
Class Trilobita Walch, Reference Walch1771
Order Redlichiida Richter, Reference Richter, Dittler, Joos, Korschelt, Linek, Oltmanns and Schaum1932
Suborder Olenellina Walcott, Reference Walcott1890
Superfamily Olenellioidae Walcott, Reference Walcott1890
Family Olenellidae Walcott, Reference Walcott1890
Subfamily Olenellinae Walcott, Reference Walcott1890
Genus Olenellus Hall, Reference Hall1861
Type species
Olenus thompsoni Hall, Reference Hall1859, from the Olenellus Zone of the Parker Formation (Slate), Vermont.
Remarks
Palmer and Repina's (Reference Palmer and Repina1993) and Palmer's (Reference Palmer1998b) generic classification is followed. Observations on the ontogeny and intraspecific variation of Olenellus gilberti Meek in White, Reference White1874, have raised doubt about this classification (Webster, Reference Webster2015). Because of this uncertainty, subgenera proposed by Palmer (Reference Palmer1998b) are not adopted here.
Olenellus gilberti Meek in White, Reference White1874
Figure 8.1–8.4, 8.6, 8.10
- Reference White1874
Olenellus gilberti Meek in White, p. 7.
- Reference White1877
Olenellus gilberti; White, p. 44, pl. 2, fig. 3a–c.
- ?Reference Kobayashi1936
Olenellus mackenziensis Kobayashi, p. 162, pl. 21, fig. 32.
- Reference Webster2015
Olenellus gilberti; Webster, p. 11, figs. 6–14, 19–21, 25–29, 31–33, 36A–W. [see for synonymy]
- Reference Boyce2021
Olenellus cf. gilberti; Boyce, p. 67, pl. 1.
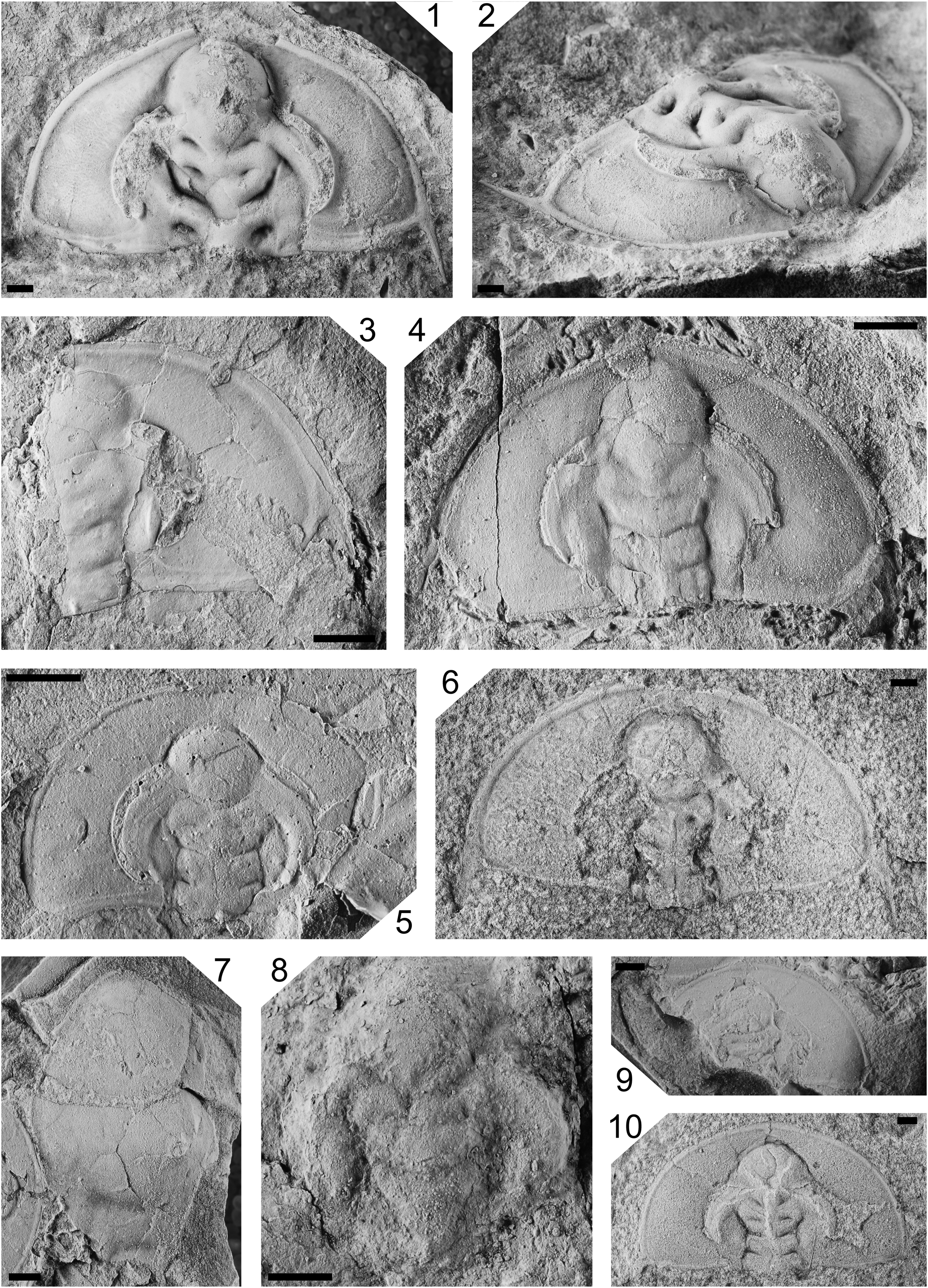
Species of Olenellus Hall, Reference Hall1861, from the upper Olenellus Zone, Northwest Territories: (1–4, 6, 10) Olenellus gilberti Meek in White, Reference White1874; (1, 2) cephalon (dorsal, oblique) GSC 142281, sandy limestone, Mount Clark Formation, Dodo Canyon; (3) partially complete cephalon (dorsal) GSC 142282, silty mudstone, Mount Cap Formation, Inlin Brook; (4) fractured cephalon (dorsal) GSC 142283, silty mudstone, Mount Cap Formation, Inlin Brook; (6) cephalon (dorsal) GSC 142284, sandstone, Mount Clark Formation, measured section 12-MWB-05; (10) cephalon (dorsal) GSC 142285, carbonate, Mount Clark Formation, measured section 12-MWB-06. (5, 7, 9) Olenellus terminatus Palmer, Reference Palmer1998b; (5) cephalon (dorsal, latex mold) GSC 142286, silty mudstone, Mount Cap Formation, Little Bear River; (7) possible fractured hypostome (dorsal) GSC 142287, silty mudstone, Mount Cap Formation, Inlin Brook; (9) partially complete cephalon (dorsal) GSC 142287, silty mudstone, Mount Cap Formation, Inlin Brook. (8) Olenellid gen. and sp. indet., partially complete cephalon (dorsal) GSC 142288, lime mudstone, Mount Cap Formation, Little Bear River. All scale bars = 2 mm.
Holotype
Cephalon (USNM 15411a) from the Olenellus Zone of the Delamar Member, Pioche Formation, Nevada (White, Reference White1877, pl. 2, fig. 3a; Walcott, Reference Walcott1910, pl. 36, fig. 3).
Occurrence
Sandstone, Mount Clark Formation, measured section 12-MWB-05; carbonate, Mount Cap Formation, measured section 12-MWB-06; sandy limestone, Mount Clark Formation, Dodo Canyon; silty mudstone, Mount Cap Formation, Little Bear River and Inlin Brook, Northwest Territories, upper Olenellus Zone; Buelna Formation, Sonora, Mexico (Cooper et al., Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952); Carrara Formation, Nevada, Olenellus Zone (Palmer and Halley, Reference Palmer and Halley1979); Delamar Member, Pioche Formation, Utah and Nevada, Bolbolenellus euryparia and Nephrolenellus multinodus zones, upper Olenellus Zone (Palmer, Reference Palmer1998b; Webster, Reference Webster2011b, Reference Websterc); Cadiz Formation, California, Olenellus Zone (Webster, Reference Webster2015); middle shale, Forteau Formation, Newfoundland, Bonnia-Olenellus Zone (Boyce, Reference Boyce2021).
Material
Twenty cephala and one hypostome. Figured material: GSC 142281–142285.
Remarks
Olenellus gilberti is characterized by a long cephalic border that is the same length or slightly shorter than the preglabellar field, a moderately long ocular lobe that has the posterior tip opposite the occipital furrow, and an advanced genal spine. Specimens from the Mount Cap Formation are nearly identical to the holotype from the Pioche Formation. One specimen (Fig. 8.10) from the Mount Clark Formation appears to have a shorter ocular ridge, although this is attributed to compaction.
Olenellus terminatus Palmer, Reference Palmer1998
Figure 8.5, 8.7, 8.9
- Reference Palmer and Halley1979
Olenellus clarki (Resser); Palmer and Halley, p. 68, pl. 3, figs. 1, 2, 5.
- Reference Palmer1998b
Olenellus terminatus Palmer, p. 670, fig. 12.1–12.8.
- Reference Webster2011c
Olenellus terminatus; Webster, fig. 4.2.
Holotype
Cephalon (DMNH 16085) from the Olenellus Zone of the Delamar Member, Pioche Formation, Nevada (Palmer, Reference Palmer1998b, fig. 12.3).
Occurrence
Sandy limestone, Mount Clark Formation, Dodo Canyon; silty mudstone, Mount Cap, Little Bear River, and Inlin Brook, Northwest Territories, upper Olenellus Zone; Carrara Formation, Nevada and California, Olenellus Zone (Palmer and Halley, Reference Palmer and Halley1979); Delamar Member, Pioche Formation, Nevada, Bolbolenellus euryparia and Nephrolenellus multinodus zones, upper Olenellus Zone (Palmer, Reference Palmer1998b; Webster, Reference Webster2011c).
Material
Twenty-seven cephala, two thoracic segments, and one possible hypostome. Figured material: GSC 142286, 142287.
Remarks
Olenellus terminatus differs from O. gilberti by the shorter cephalic border and less-advanced genal spine. Olenellus terminatus differs from O. clarki by having a longer ocular lobe with the posterior tip opposite the anterior half of the occipital ring.
Genus Bolbolenellus Palmer and Repina, Reference Palmer and Repina1993
Type species
Olenellus euryparia Palmer in Palmer and Halley, Reference Palmer and Halley1979, from the Olenellus Zone of the Carrara Formation, Nevada.
Remarks
This genus was originally classified in subfamily Bristoliinae by Palmer and Repina (Reference Palmer and Repina1993), but was later reclassified within Olenellinae due to the presence of a macropleural thoracic segment (Palmer, Reference Palmer1998b). The most distinctive feature is the absence of a preglabellar field, where the anterior glabellar lobe abuts or overlaps the cephalic border (Palmer and Repina, Reference Palmer and Repina1993).
Bolbolenellus dodoensis Handkamer and Pratt, new species
Figure 9
Holotype
Cephalon (GSC 142289) from the upper Olenellus Zone of the Mount Clark Formation, Dodo Canyon (17.3 m below the base of the Mount Cap Formation), Northwest Territories (Fig. 9.1, 9.2).
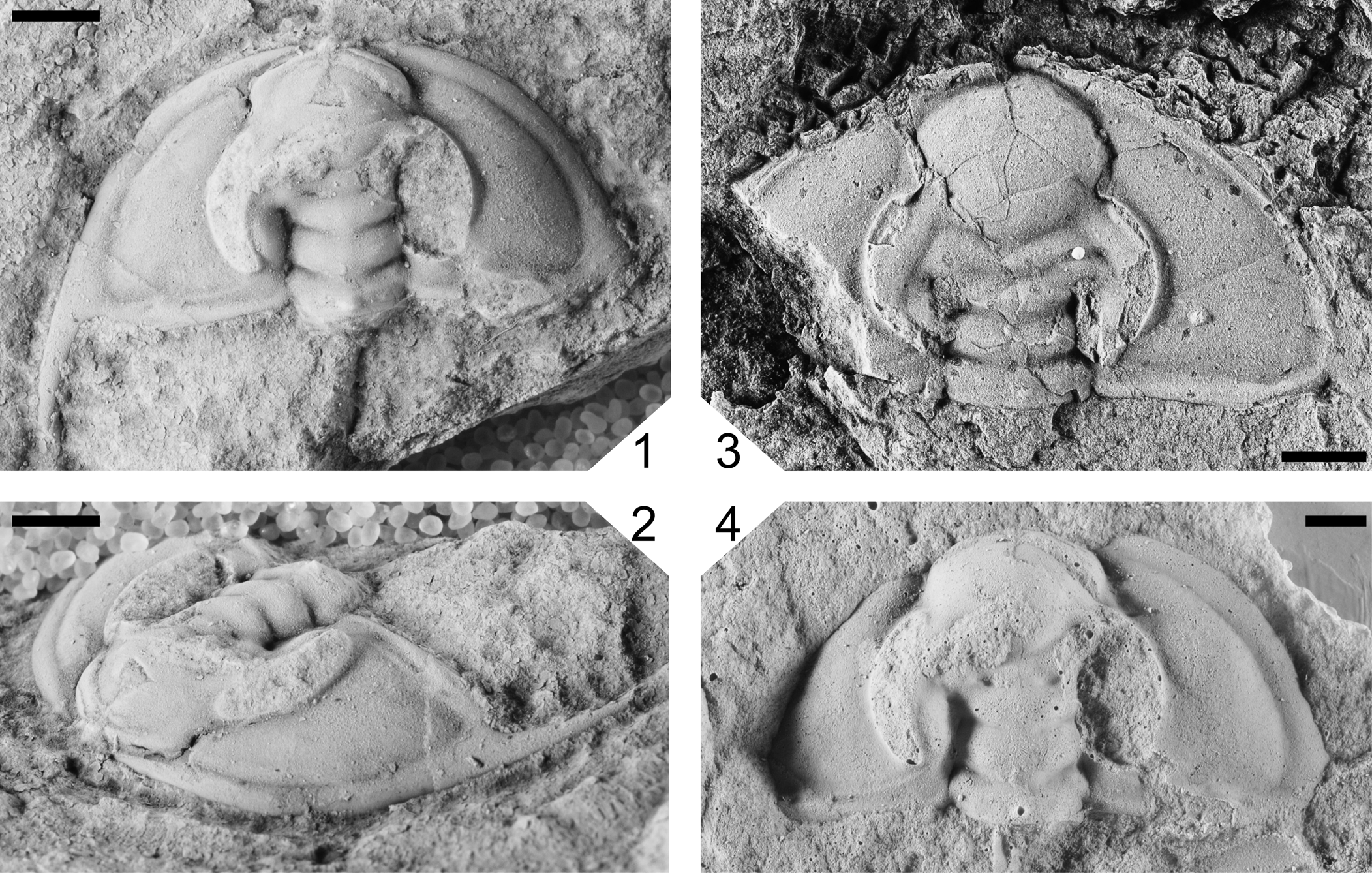
Bolbolenellus dodoensis Handkamer and Pratt, n. sp., from the upper Olenellus Zone, Northwest Territories. (1, 2) Holotype cephalon (dorsal, oblique) GSC 142289, sandy limestone, Mount Clark Formation, Dodo Canyon; (3) partially complete paratype cephalon (dorsal) GSC 142290, silty mudstone, Mount Cap Formation, Little Bear River; (4) paratype cephalon (dorsal, latex mold) GSC 142291, sandy limestone, Mount Clark Formation, Dodo Canyon. All scale bars = 2 mm.
Diagnosis
A species of Bolbolenellus with the anterior glabellar lobe intersecting or overlapping the anterior cephalic border furrow, ocular lobe long, posterior cephalic border nearly transverse, genal spine slightly advanced and moderately long.
Occurrence
Sandy limestone, Mount Clark Formation, Dodo Canyon; silty mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, upper Olenellus Zone.
Description
Cephalon is semicircular in outline, the length equal to half the width. Anterior cephalic border ranges in length from half to slightly less than that of the occipital ring. Cephalic border furrow is moderately to well defined. Preglabellar field is absent, with the anterior edge of the glabella either intersecting or overlapping the anterior border furrow. Strongly convex glabella with a well-defined axial furrow, narrowing from the occipital ring to L2, and expanding from L2 to the rounded anterior lobe. S1 and S2 are well defined and oriented obliquely backward, S2 does not intersect the axial furrow, and S3 is moderately defined and oriented transversely. L3 expands posterolaterally to envelope the lateral side of L2. Occipital ring and furrow are well defined, with a medial occipital tubercle. Strongly convex ocular lobe is crescent-shaped, narrows posteriorly, with the anterior tip intersecting the posterior part of the anterior glabellar lobe, and the posterior tip opposite the occipital furrow. Length of the ocular lobe is half that of the glabella. Interocular cheek at the widest point in width is one-third that of the occipital ring. Width of the moderately convex extraocular area at the widest point one-and-one-third times that of the occipital ring. Posterior border oriented nearly transverse and in width is one-and-one-half times that of the occipital ring. Genal spine is slightly advanced, the length is five-sixths that of the glabella.
Hypostome, thorax, and pygidium are unknown.
Etymology
Named after Dodo Canyon, Canyon Ranges, eastern Mackenzie Mountains.
Material
Eleven cephala. Type material: holotype, GSC 142289; paratypes, GSC 142290, GSC 142291.
Remarks
Specimens of Bolbolenellus dodoensis Handkamer and Pratt, n. sp., from the Mount Cap Formation show variation in the length of the anterior cephalic border and position of the anterior lobe of the glabella with respect to the cephalic border. The border length and position of the anterior glabellar lobe with respect to the anterior cephalic border varies in Olenellus gilberti (Webster, Reference Webster2015, figs. 4F, 5A). These features are considered to represent intraspecific variation in B. dodoensis n. sp. as well.
Olenellid genus and species indet.
Figure 8.8
Occurrence
Dolomitic limestone of the Mount Cap Formation, Little Bear River, Northwest Territories, upper Olenellus Zone.
Material
One partial cephalon. Figured material: GSC 142288.
Remarks
The fragmentary cephalon shows the ocular lobe intersecting the posterior edge of L4, and L3 expanding posterolaterally to envelope the side of L2. This separates S2 from the axial furrow. Olenellid gen. and sp. indet. may be a poorly preserved specimen of Olenellus gilberti or O. terminatus. The presence of a possible preglabellar field suggests this specimen is not Bolbolenellus dodoensis Handkamer and Pratt, n. sp.
Order Corynexochida Kobayashi, Reference Kobayashi1935
Family Dolichometopidae Walcott, Reference Walcott1916
Genus Bathyuriscus Meek, Reference Meek1873
Type species
Bathyurus (?) haydeni Meek, Reference Meek1873, from the Meagher Formation (Limestone), Montana (Diess, Reference Diess1936).
Remarks
The generic diagnosis by Robison (Reference Robison1964) is followed here. See Young and Ludvigsen (Reference Young and Ludvigsen1989) for the history of the discussion on the generic name.
Bathyuriscus rotundatus (Rominger, Reference Rominger1887)
Figures 10, 11
- Reference Rominger1887
Embolimus rotundata Rominger, p. 16, pl. 1, figs. 4, 5.
- Reference Walcott1908a
Bathyuriscus rotundatus; Walcott, pl. 4, fig. 2.
- Reference Walcott1916b
Bathyuriscus rotundatus; Walcott, p. 346, pl. 47, figs. 2, 2a, b. [see for synonymy]
- Reference Rasetti1951
Bathyuriscus rotundatus; Rasetti, p. 158, pl. 28, figs. 2, 3.
- Reference Hu1985
Bathyuriscus rotundatus; Hu, pl. 6, figs. 18, 21, 25.
Bathyuriscus rotundatus (Rominger, Reference Rominger1887) from the Aitkenaspis keelensis Zone, mudstone, Mount Cap Formation, Little Bear River, Northwest Territories. (1–3) Nearly complete exoskeleton (dorsal, lateral, oblique) GSC 142292; (4, 5) cranidium (dorsal, oblique) GSC 142293; (6) free cheek, (dorsal) GSC 142294; (7, 8) pygidium (dorsal, oblique) GSC 142295; (9, 10) nearly complete exoskeleton, (dorsal, oblique) GSC 142296. All scale bars = 2 mm.

Bathyuriscus rotundatus (Rominger, Reference Rominger1887) from the Aitkenaspis keelensis Zone, mudstone, Mount Cap Formation, Little Bear River, Northwest Territories. (1, 2) Early holaspid exoskeleton lacking free cheeks (dorsal, oblique) GSC 142297; (3) cranidium with one free cheek (dorsal) GSC 142298; (4) exoskeleton lacking free cheeks (dorsal) GSC 142299; (5) exoskeleton lacking free cheeks (dorsal) GSC 142300; (6) articulated thorax and pygidium (dorsal) GSC 142294. All scale bars = 2 mm.

Holotype
Embolimus rotundata (specimen appears to be lost) from the Stephen Formation, British Columbia (Rominger, Reference Rominger1887, pl. 1, fig. 4).
Diagnosis
A species of Bathyuriscus with the anterior facial suture divergent, interocular area narrow, posterior border moderately wide; pygidium subisopygous, five to six axial rings, border spines absent.
Occurrence
Mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, Aitkenaspis keelensis Zone; Stephen Formation, British Columbia, Bathyuriscus-Elrathina Zone (Rominger, Reference Rominger1887; Walcott, Reference Walcott1908a, Reference Walcott1916b; Rasetti, Reference Rasetti1951; Hu, Reference Hu1985).
Material
Six complete exoskeletons, 23 incomplete exoskeletons or nearly complete exoskeletons, two early holaspid exoskeletons, 10 cranidia, 15 free cheeks, three thoracic segments, and 27 pygidia. Figured material: GSC 142292–142300.
Remarks
Pygidia from the Mount Cap Formation vary in the length to width ratio (Figs. 10.1, 11.4, 11.6), similar to Walcott's material (Reference Walcott1916b, pl. 47, figs. 2, 2a) from the Stephen Formation. Because the ratio does not correlate to the overall size or number of pygidial segments, this is interpreted as intraspecific variation. The length of the terminal spines of the thoracic segments seems to vary slightly in B. rotundatus. Also, the free cheeks from the Mount Cap Formation appear to have a slightly longer genal spine than those in the Stephen Formation. This occurrence of B. rotundatus is older than previously reported collections.
Two early holaspids of Bathyuriscus rotundatus were collected (Fig. 11.1, 11.2). These differ from late holaspid exoskeletons by the narrower posterior cranidial border; narrower thoracic pleura; smaller, triangular-shaped pygidium with an axis that is nearer the border furrow, which is composed of one less axial ring; and a faintly denticulated pygidial margin. Except for the denticulated margin, these differences are similar to those between early and late holaspids of B. fimbriatus Robison, Reference Robison1964 (Robison, Reference Robison1967).
Genus Eobathyuriscus Handkamer and Pratt, new genus
Type species
Eobathyuriscus mackenziensis Handkamer and Pratt, n. sp. from the Albertelloides mischi and Glossopleura walcotti zones of the Mount Cap Formation, Northwest Territories.
Diagnosis
Dolichometopid with glabella expanding anteriorly, anterior border short to absent, palpebral lobe short, interocular area wide to moderately wide; free cheek moderately wide, genal spine long; eight or nine thoracic segments; subisopygous, four or five axial rings, pleural field moderately narrow, pleural furrows poorly to moderately defined, border furrow well defined, border short.
Etymology
Occurs in strata older than those typically containing species of Bathyuriscus.
Remarks
Eobathyuriscus Handkamer and Pratt, n. gen., is similar to Bathyuriscus, Poliella Walcott, Reference Walcott1916b, and Wenkchemnia Rasetti, Reference Rasetti1951. Eobathyuriscus n. gen., differs from Bathyuriscus by having a generally wider interocular area, smaller pygidium composed of fewer axial rings, and less well-defined pygidial pleural furrows. It differs from Poliella in having a wider anterior glabellar lobe, generally shorter palpebral lobe, larger pygidium composed of more axial rings, and narrower pygidial border. Eobathyuriscus n. gen., differs from Wenkchemnia in having a wider interocular area and larger pygidium with more axial rings.
Eobathyuriscus Handkamer and Pratt, n. gen., is intermediate in form between Wenkchemnia and Bathyuriscus. The length of the palpebral lobe (40–48% of the cranidial length) is like that of Wenkchemnia (35–41% of the cranidial length) and some species of Bathyuriscus, such as B. terranovensis Young and Ludvigsen, Reference Young and Ludvigsen1989 (40–50% of the cranidial length). The size of the pygidium in Eobathyuriscus n. gen., (60–65% of the cranidial length) and number of axial rings (four or five) is intermediate between Wenkchemnia (45–57% of the cranidial length, two or three rings) and Bathyuriscus (75–120% of the cranidial length, five to eight axial rings). Some specimens of B. rotundatus have the same number of axial rings, although the pygidium is proportionally larger and has better defined pleural furrows than Eobathyuriscus n. gen., (Fig. 11.6; Walcott, Reference Walcott1916b, pl. 47, figs. 2, 2a).
Eobathyuriscus mackenziensis Handkamer and Pratt, new species
Figures 12, 13
Holotype
Eobathyuriscus mackenziensis (GSC 142301) from the Glossopleura walcotti Zone of the Mount Cap Formation, Carcajou Falls (24.4 m above the base of the Mount Cap Formation), Northwest Territories (Fig. 12.1).
Eobathyuriscus mackenziensis Handkamer and Pratt, n. gen., n. sp., from the Mount Cap Formation, Northwest Territories. (1) Holotype exoskeleton lacking free cheeks (dorsal) GSC 142301, lime mudstone, Carcajou Falls, Glossopleura walcotti Zone; (2) paratype complete exoskeleton (dorsal) GSC 142302, mudstone, Carcajou Falls, Glossopleura walcotti Zone; (3) paratype exoskeleton lacking free cheeks (dorsal) GSC 142303, wackestone, Carcajou Falls, Glossopleura walcotti Zone; (4) paratype exoskeleton lacking a free cheek (dorsal) GSC 142304, mudstone, Little Bear River, Albertelloides mischi or Glossopleura walcotti Zone; (5) paratype exoskeleton lacking free cheeks (dorsal) GSC 142305, lime mudstone, Carcajou Falls, Glossopleura walcotti Zone; (6) paratype exoskeleton (dorsal) GSC 142306, lime mudstone, Carcajou Falls, Glossopleura walcotti Zone; (7) paratype exoskeleton (dorsal) GSC 142307, lime mudstone, Carcajou Falls, Glossopleura walcotti Zone; (8) paratype exoskeleton (dorsal) GSC 142308, mudstone, Carcajou Falls, Glossopleura walcotti Zone; (9) disarticulated exoskeleton (dorsal, latex mold) GSC 142309, mudstone, Carcajou Falls, Glossopleura walcotti Zone. All scale bars = 2 mm.

Eobathyuriscus mackenziensis Handkamer and Pratt, n. gen n. sp. from the Mount Cap Formation, Northwest Territories. (1) Paratype exoskeleton lacking free cheeks (dorsal) GSC 142310, mudstone, Carcajou Falls, Glossopleura walcotti Zone; (2) paratype exoskeleton lacking free cheeks (dorsal) GSC 142311, mudstone, Carcajou Falls, Glossopleura walcotti Zone; (3) paratype disarticulated thorax and pygidium (dorsal) GSC 142312, siltstone, Carcajou Falls, Glossopleura walcotti Zone; (4) paratype cranidium (dorsal) GSC 142313, siltstone, Carcajou Falls, Glossopleura walcotti Zone; (5) paratype nearly complete exoskeleton (dorsal) GSC 142314, siltstone, Carcajou Falls, Glossopleura walcotti Zone; (6) paratype free cheek (dorsal) GSC 142315, mudstone, Little Bear River, Albertelloides mischi Zone; (7) paratype pygidium (dorsal) GSC 142316, siltstone, Carcajou Falls, Glossopleura walcotti Zone; (8) paratype pygidium (dorsal) GSC 142306, lime mudstone, Carcajou Falls, Glossopleura walcotti Zone; (9) paratype meraspid cranidium (dorsal) GSC 142317, lime mudstone, Carcajou Falls, Glossopleura walcotti Zone, scale bar = 500 μm; (10) paratype hypostome (dorsal) GSC 142318, lime mudstone, Carcajou Falls, Glossopleura walcotti Zone, (11) paratype cranidium (dorsal) GSC 142319, mudstone, Carcajou Falls, Glossopleura walcotti Zone. All scale bars = 2 mm unless stated otherwise.

Diagnosis
Eobathyuriscus Handkamer and Pratt, n. gen., with interocular area wide, slightly variable, posterior border wide; free cheek narrow, genal spine short; eight or nine thoracic segments, thoracic spines short; four or five pygidial axial rings, pleural field narrow, border short, variably with shallow medial indentation and very short border spine.
Occurrence
Mudstone, siltstone, lime mudstone, and wackestone, Mount Cap Formation, Little Bear River, Carcajou Falls, and Dodo Canyon, Northwest Territories, Aitkenaspis keelensis, Albertelloides mischi, and Glossopleura walcotti zones.
Description
Exoskeleton is subelliptical in outline. Exoskeleton length 14–25 mm.
Cranidium is subtrapezoidal in outline. Axial furrow is well defined. Glabella extends to anterior border furrow or margin if anterior border is absent. Four pairs of lateral glabellar furrows are present. Glabella narrows gently from the occipital ring to S1 and widens from S1 to the anterior lobe. S1 is well defined and oriented obliquely backwards; S2 is moderately defined and oriented obliquely backwards; and S3 and S4 are poorly defined and oriented obliquely forward. Occipital furrow is well defined, the occipital ring variably bearing a median tubercle. Anterior course of the facial suture is slightly divergent. Interocular area at the widest point is two-thirds to half the width of the occipital ring. Palpebral lobe is strongly arched, with the anterior tip intersecting the axial furrow opposite S4, and the posterior tip opposite S1. Length of the palpebral lobe is half that of the glabella. Posterior course of the facial suture is oriented obliquely backwards. Posterior border is equal in width to the occipital ring, with a moderately defined border furrow.
Free cheek is equal in width to the occipital ring; flat lateral border comprises one-fifth the width. Genal spine is non-advanced and comprises one-third the length of the free cheek.
Hypostome is conterminant, widening anteriorly and narrowing posteriorly. Length of the posterior lobe is equal to one-fifth of the total length.
Thorax consists of eight, rarely nine segments. Axial furrow is well defined; each axial ring has a medial tubercle. Pleura is slightly wider than the axis, with well-defined pleural and interpleural furrows. Pleural spines moderately short to moderately long and narrow.
Pygidium subisopygous, semicircular in outline, and the length is equal to two-thirds the width. Pygidial axis tapers gently posteriorly; the posterior tip is slightly anterior of the border furrow, and is composed of four or five rings and a terminal piece. Pleural field is equal in width to the axis, with four or five pairs of pleural furrows. The two or three anterior furrows are moderately to poorly defined and intersect the border furrow, and the two posterior furrows are poorly defined or effaced and do not intersect the border furrow. The border comprises one-tenth or less the length of the pygidium. One pair of short border spines is variably present, opposite the anteriormost pleural furrow. The posterior edge of the border may have a medial indentation.
Etymology
Named after the Mackenzie Mountains.
Material
Twenty-nine complete and nearly complete exoskeletons, 93 exoskeletons lacking the free cheeks, 78 holaspid cranidia, three meraspid cranidia, four isolated thoracic segments, and 87 pygidia. Type material: holotype, GSC 142301; paratypes GSC 142302–142319.
Remarks
Specimens of Eobathyuriscus mackenziensis Handkamer and Pratt, n. gen. n. sp., show variation in the width of the interocular area, number of thoracic segments, length and width of the border of the free cheek and pygidium, number of pygidial axial rings and furrows, presence of a medial indentation in the pygidial margin, and presence of border spines. The width of the interocular area correlates to the size of the cranidium in that it is narrower in larger specimens (Figs. 12.1, 12.7, 13.8). Larger pygidia have five rather than four axial rings and pleural furrows, and border spines are present only on the smaller pygidia (Figs. 12.4–12.6, 13.2). These features are interpreted as growth related. Variation in border length, number of thoracic segments, and presence of a medial indentation do not correlate with exoskeleton size and are regarded as intraspecific variation.
Eobathyuriscus macqueeni Handkamer and Pratt, new species
Figure 14
Holotype
Eobathyuriscus macqueeni (GSC 142320) from the Glossopleura walcotti Zone of the Mount Cap Formation, Carcajou Falls (27.2 m above the base of the Mount Cap Formation), Northwest Territories (Fig. 14.1–14.3).
Eobathyuriscus macqueeni Handkamer and Pratt, n. gen. n. sp. from the Glossopleura walcotti Zone, Mount Cap Formation, Carcajou Falls, Northwest Territories. (1–3) Holotype complete exoskeleton (dorsal, oblique, lateral) GSC 142320, lime mudstone; (4) paratype nearly complete exoskeleton (dorsal, latex mold) GSC 142321, lime mudstone; (5, 6) paratype cranidium (dorsal, oblique) GSC 142322, lime mudstone; (7, 8) paratype cranidium (dorsal, oblique) GSC 142323, wackestone; (9, 10) paratype pygidium (dorsal, oblique) GSC 142324, lime mudstone; (11, 12) paratype pygidium (dorsal, oblique) GSC 142323; wackestone. All scale bars = 2 mm.

Diagnosis
Eobathyuriscus Handkamer and Pratt, n. gen., with an interocular area narrow, posterior border narrow; free cheek wide, genal spine long; nine thoracic segments, thoracic spine long; five or six pygidial axial rings, pleural field wide, border long, medial indentation and border spines absent.
Occurrence
Lime mudstone and rarely wackestone, Mount Cap Formation, Carcajou Falls, Northwest Territories, Glossopleura walcotti Zone.
Description
Exoskeleton is subquadrate in outline. Exoskeleton length 40 mm.
Cranidium is subtrapezoidal in outline. Axial furrow is well defined. Glabella extends to anterior border furrow or margin if anterior border is absent. Four pairs of lateral glabellar furrows are present. Glabella narrows gently from the occipital ring to S1 and widens from S1 to the anterior lobe. S1 is well defined and oriented obliquely backwards; S2 is poorly defined or effaced and oriented obliquely backwards; and S3 and S4 are poorly defined or effaced and oriented obliquely forward. Occipital furrow is well defined, the occipital ring variably bearing a medial tubercle. Anterior course of the facial suture is divergent. Moderately convex interocular area at the widest point is equal to half the width of the occipital ring. Palpebral lobe is strongly arched, with the anterior tip intersecting the axial furrow opposite S4, and the posterior tip opposite the middle of L1. Length of the palpebral lobe is equal to half that of the glabella. Posterior course of the facial suture is oriented obliquely backwards. Weakly convex posterior border is slightly narrower than the occipital ring, with a well-defined border furrow.
Free cheek is slightly narrower than the occipital ring, moderately convex, and the border comprises one-third of the width. Genal spine is non-advanced and comprises one-third to two-thirds the length of the free cheek.
Hypostome is conterminant.
Thorax consists of nine segments. Moderately convex axis with a well-defined axial furrow; each axial ring has a medial tubercle. Gently convex pleura is slightly wider than the axis, with well-defined pleural and interpleural furrows. Pleural spines are moderately long and narrow.
Pygidium subisopygous, semicircular in outline, and the length is equal to half to two-thirds the width. Moderately convex axis narrows posteriorly, the posterior tip is slightly anterior of the border furrow, and is composed of five or six rings and a terminal piece. Axial ring furrows are well defined. Gently convex pleural field is slightly wider than the axis, with five pairs of pleural furrows that intersect the border furrow. The three anterior furrows are well defined, and the two posterior furrows are poorly defined. The flat border comprises one-eighth the length of the pygidium. Border spines are absent.
Etymology
Named after Roger W. Macqueen, who was the first GSC researcher to study the lower Paleozoic stratigraphy of the eastern Mackenzie Mountains in detail.
Material
One complete exoskeleton, one nearly complete exoskeleton, six cranidia, and seven pygidia. Type material: holotype, GSC 142320; paratypes, GSC 14321–142324.
Remarks
Eobathyuriscus macqueeni Handkamer and Pratt, n. gen. n. sp. differs from E. mackenziensis n. gen. n. sp. in its larger size, narrower interocular area, wider free cheek, longer genal spine, slightly longer thoracic spines, occasional additional axial ring, wider pleural field, better defined pleural furrows, slightly longer border, and lack of short border spines. Eobathyuriscus macqueeni n. gen. n. sp. was only collected in limestone, whereas E. mackenziensis n. gen. n. sp. was collected in mudstones as well.
Genus Glossopleura Poulsen, Reference Poulsen1927
Type species
Dolichometopus boccar Walcott, Reference Walcott1916b, from the Glossopleura Zone of the Stephen Formation, British Columbia.
Remarks
Glossopleura Poulsen, Reference Poulsen1927, has >50 species and is in need of review because most species are likely synonymous (Sundberg, Reference Sundberg2005; Robison and Babcock, Reference Robison and Babcock2011) and features used to discern species of this genus can overlap significantly (Sundberg, Reference Sundberg2005). Hitherto, Glossopleura species for which the thorax is known had seven or eight segments. Glossopleura youngi Handkamer and Pratt, n. sp., bears six thoracic segments, which is not regarded as taxonomically important at the generic level.
Glossopleura boccar (Walcott, Reference Walcott1916)
Figure 15.1–15.6
- Reference Walcott1916b
Dolichometopus boccar Walcott, p. 363, pl. 52, fig. 1, 1a–f.
- Reference Sundberg2005
Glossopleura boccar; Sundberg, p. 65, fig. 6.10–6.14. [see for synonymy]
- Reference Foster2011
Glossopleura boccar; Foster, p. 105, fig. 4.1–4.3.
Dolichometopids from the Glossopleura walcotti Zone, Mount Cap Formation, Northwest Territories. (1–6) Glossopleura boccar (Walcott, Reference Walcott1916b), Dodo Canyon; (1) cranidium (dorsal) GSC 142448, mudstone; (2) cranidium (dorsal) GSC 142325, mudstone; (3) cranidium (dorsal) GSC 142326, lime mudstone; (4) partially complete exoskeleton lacking the cephalon (dorsal) GSC 143327, mudstone; (5) pygidium (dorsal) GSC 142328, mudstone; (6) pygidium (dorsal) GSC 142329, mudstone. (7, 8) Polypleuraspis solitaria Poulsen, Reference Poulsen1927, lime mudstone, measured section 12-MWB-04; (7) pygidium (dorsal) GSC 142330; (8) pygidium (dorsal) GSC 142331. All scale bars = 2 mm.

Holotype
Dolichometopus boccar (USNM 62702) from the Glossopleura Zone of the Stephen Formation, British Columbia (Walcott, Reference Walcott1916b, pl. 52, fig. 1).
Diagnosis
Glossopleura with anterior glabellar lobe moderately wide to wide, S1 well defined, S2, S3, and S4 poorly defined or effaced, palpebral lobe long and narrow; posterior border wide; thorax with seven segments; pygidial length two-thirds the width, axis long, intersecting the border furrow, ring furrows moderately defined, pleural furrows and border furrow moderately to poorly defined, border moderately long (modified from Sundberg, Reference Sundberg2005).
Occurrence
Mudstone, lime mudstone, and wackestone, Mount Cap Formation, Dodo Canyon and measured section 12-MWB-04, Northwest Territories, Glossopleura walcotti Zone; Stephen Formation, Alberta and British Columbia, Glossopleura Zone (Walcott, Reference Walcott1916b; Rasetti, Reference Rasetti1951); Chisholm Formation, Utah, Glossopleura walcotti Zone (Sundberg, Reference Sundberg2005; McCollum and Sundberg, Reference McCollum and Sundberg2007); Bright Angel Shale, Arizona, Glossopleura walcotti Zone (Foster, Reference Foster2011).
Material
Eleven nearly complete exoskeletons, 28 cranidia, eight free cheeks, 13 thoracic segments, and 52 pygidia. Figured material: GSC 142325–142329, 142448.
Remarks
Glossopleura boccar from the Mount Cap Formation varies in the width of the anterior glabellar lobe (Fig. 15.1 vs. 15.2, 15.3) and the depth of the pygidial border and pleural furrows (Fig. 15.4–15.6), similar to G. boccar in the Chisholm Formation (Sundberg, Reference Sundberg2005, fig 6.10–6.14). This species has been tentatively reported from the Mount Cap Formation in the subsurface of the Colville Hills as well (Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020).
Glossopleura youngi Handkamer and Pratt, new species
Figure 16
Holotype
Glossopleura youngi (GSC 142332) from the Glossopleura walcotti Zone of the Mount Cap Formation, Carcajou Falls (28.5 m above the base of the Mount Cap Formation), Northwest Territories (Fig. 16.1).
Glossopleura youngi Handkamer and Pratt, n. sp., from the Glossopleura walcotti Zone, Mount Cap Formation, Carcajou Falls, Northwest Territories. (1) Holotype exoskeleton lacking free cheeks (dorsal, latex mold) GSC 142332, mudstone; (2) paratype nearly complete exoskeleton (dorsal) GSC 142333, mudstone; (3) paratype partially complete exoskeleton (dorsal) GSC 142334, mudstone; (4) paratype complete exoskeleton (dorsal) GSC 142335, mudstone; (5) paratype cranidium (dorsal) GSC 142336, mudstone; (6) paratype pygidium (dorsal) GSC 142337, mudstone; (7) paratype cranidium (dorsal) GSC 142338, mudstone; (8) paratype pygidium (dorsal) GSC 142339, mudstone; (9, 10) paratype pygidium (dorsal, oblique) GSC 142340, grainstone; (11) paratype articulated pygidium and thorax (dorsal) GSC 142341, mudstone. All scale bars = 2 mm.

Diagnosis
Glossopleura with anterior glabellar lobe moderately wide, lateral glabellar furrows poorly defined to effaced, palpebral lobe long and wide, posterior border narrow; thorax with six segments; pygidial length half to three-fifths the width, axis short, anterior of border furrow, ring furrows poorly defined or effaced, pleural furrows poorly defined or effaced, border furrow well defined, border long.
Occurrence
Mudstone and grainstone, Mount Cap Formation, Carcajou Falls, Northwest Territories, Glossopleura walcotti Zone.
Description
Exoskeleton is subelliptical in outline. Exoskeleton length 18–28 mm.
Cranidium is subquadrate in outline. Axial furrow is moderately defined. Glabella is subrectangular in shape and extends to anterior border. Four pairs of lateral glabellar furrows are present. Glabella widens gently from S1 to the anterior lobe. S1 is poorly defined and oriented obliquely backwards; S2 is poorly defined or effaced and oriented obliquely backwards; and S3 and S4 are poorly defined or effaced and oriented obliquely forward. Occipital furrow is poorly to moderately defined. Length of the anterior border is less than one-tenth that of the occipital ring. Anterior course of the facial suture is slightly divergent. Width of the interocular area at the widest point is one-third that of the occipital ring. Palpebral lobe is strongly arched, with the anterior tip intersecting the axial furrow slightly posterior of S4, and the posterior tip opposite the occipital ring. Length of the palpebral lobe is three-fifths that of the glabella, and the width is slightly narrower than that of the interocular area. Posterior course of the facial suture is transverse. Width of the posterior border is two-thirds that of the occipital ring, with a moderately defined border furrow.
Free cheek width is unknown. Free cheek border comprises one-sixth the width of the free cheek. Genal spine is short.
Hypostome is unknown.
Thorax consists of six segments. Axial furrow is well defined. Pleural width is equal to or slightly wider than that of the axis, pleura with moderately defined pleural and interpleural furrows. Pleural spines are very short.
Pygidium isopygous, semicircular in outline, and the length is half to three-fifths the width. Moderately convex axis narrows gently posteriorly, with the posterior tip slightly anterior of the border furrow, and is composed of at least three rings. Axial ring furrows are poorly defined or effaced. Gently convex pleural field is slightly narrower than the axis, with two poorly defined or effaced pleural furrows. Flat border comprises one-quarter to one-fifth the length of the pygidium. Border is covered in terrace lines. Border spines are absent.
Etymology
Named after Graham A. Young, Manitoba Museum, Winnipeg, Manitoba.
Material
One complete exoskeleton, 21 nearly complete exoskeletons or complete exoskeletons lacking free cheeks, 10 cranidia, and 19 pygidia. Type material: holotype, GSC 142332; paratypes, GSC 142333–142341.
Remarks
Glossopleura youngi Handkamer and Pratt, n. sp., has one thoracic segment fewer than in other species of Glossopleura for which the thorax is known. In those species, Glossopleura varies in having either seven or eight thoracic segments, but the number is stable within individual species. Apart from the number of thoracic segments, Glossopleura youngi n. sp. is similar to G. boccar and G. producta (Walcott, Reference Walcott1916b), differing from the former by having a more effaced glabella, a slightly longer and wider palpebral lobe, a narrower posterior border, a slightly shorter pygidial axis, a less well-defined axial furrow, a near absence of pleural furrows, and a longer border; and differing from the latter by having an more effaced glabella, a slightly shorter pygidial axis, and a longer pygidial border.
Genus Polypleuraspis Poulsen, Reference Poulsen1927
Type species
Polypleuraspis solitaria Poulsen, Reference Poulsen1927, from the Glossopleura walcotti Zone of the lower Cape Wood Formation, Greenland.
Remarks
Peel's (Reference Peel2020) generic diagnosis is followed herein. Polypleuraspis currently contains three species: P. solitaria, P. insignis Rasetti, Reference Rasetti1951, and P. glacialis Peel, Reference Peel2020, all from the Glossopleura Zone. Peel (Reference Peel2020) allowed for some intraspecific variation in the pygidium of P. glacialis, in outline, convexity, and shape of the median indentation of the posterior margin.
Polypleuraspis solitaria Poulsen, Reference Poulsen1927
Figure 15.7, 15.8
- Reference Poulsen1927
Polypleuraspis solitaria Poulsen, p. 270, pl. 16, figs. 37, 38.
- Reference Peel2020
Polypleuraspis solitaria; Peel, p. 20, figs. 2D, K, L, N, P, Q, S, T, 4A–U. [see for synonymy]
Holotype
Pygidium (MGUH 2292) from the Glossopleura Zone of the lower Cape Wood Formation, northwestern Greenland (Poulsen, Reference Poulsen1927, pl. 16, figs. 37, 38; Peel, Reference Peel2020, fig. 2I, K, N).
Occurrence
Lime mudstone, Mount Cap Formation, measured section 12-MWB-04, Northwest Territories, Glossopleura walcotti Zone; Cap Wood Formation, Kap Kent, Greenland, Glossopleura Zone (Poulsen, Reference Poulsen1927; Peel, Reference Peel2020).
Material
Two pygidia. Figured material: GSC 142330, 142331.
Remarks
The maximum pygidial width of these specimens is at the midlength, which is consistent with P. solitaria. The two pygidia in the Mount Cap Formation are wider than most specimens from northwestern Greenland. The axial nodes are preserved only on one specimen.
Genus Sahtuia Handkamer and Pratt, new genus
Type species
Sahtuia carcajouensis Handkamer and Pratt, n. sp., from the Glossopleura walcotti Zone of the Mount Cap Formation, Northwest Territories.
Diagnosis
As for species.
Etymology
Named after the Sahtu Region of the Northwest Territories, within which the study area lies.
Remarks
The glabellar shape and size, orientation of the palpebral lobe, size and shape of the postocular fixed cheek, lack of a fixigenal spine, narrow pygidial border, and lack of border spines indicate that Sahtuia Handkamer and Pratt, n. gen. belongs to Dolichometopidae and is closely related to Bathyuriscus and Eobathyuriscus Handkamer and Pratt, n. gen. Sahtuia n. gen., compared to other dolichometopids, has fewer thoracic segments and a larger pygidium. At present the genus is monospecific.
Sahtuia carcajouensis Handkamer and Pratt, new species
Figures 17, 18
Holotype
Sahtuia carcajouensis (GSC 142342) from the Glossopleura walcotti Zone of the Mount Cap Formation, Carcajou Falls (26.3 m above the base of the Mount Cap Formation), Northwest Territories (Fig. 17.1).
Sahtuia carcajouensis Handkamer and Pratt, n. gen. n. sp., from the Glossopleura walcotti Zone, mudstone, Mount Cap Formation, Carcajou Falls, Northwest Territories. (1) Holotype exoskeleton lacking a free cheek (dorsal) GSC 142342; (2) paratype complete exoskeleton (dorsal, latex mold) GSC 142343; (3) paratype exoskeleton lacking free cheeks (dorsal, latex mold) GSC 142344; (4) paratype exoskeleton lacking free cheeks (dorsal, latex mold) GSC 142345; (5) paratype nearly complete exoskeleton (dorsal) GSC 142346; (6) paratype complete exoskeleton (dorsal) GSC 142347; (7) paratype exoskeleton lacking free cheeks (dorsal) GSC 142348; (8) paratype exoskeleton lacking free cheeks (dorsal) GSC 142349; (9) paratype complete exoskeleton (dorsal, latex mold) GSC 142350. All scale bars = 2 mm.

Sahtuia carcajouensis Handkamer and Pratt, n. gen. n. sp., from the Glossopleura walcotti Zone, Mount Cap Formation, Carcajou Falls, Northwest Territories. (1, 2) Paratype cranidium (dorsal, oblique) GSC 142351, grainstone; (3) paratype cranidium (dorsal) GSC 142352, mudstone; (4, 5) paratype pygidium (dorsal, oblique) GSC 142353, grainstone; (6) paratype pygidium (dorsal) GSC 142354; mudstone; (7) paratype free cheek (dorsal) GSC 142355, mudstone; (8) paratype cranidium (dorsal) GSC 142356, mudstone; (9) paratype pygidium (dorsal) GSC 142357, mudstone; (10) paratype early meraspid cranidium (dorsal) GSC 142358, mudstone, scale bar = 125 μm; (11) paratype protaspid (dorsal) GSC 142359, mudstone, scale bar = 125 μm; (12) paratype late meraspid cranidium (dorsal) GSC 142360, mudstone, scale bar = 500 μm. All scale bars = 2 mm unless stated otherwise.

Diagnosis
Dolichometopid with glabella anteriorly wide, palpebral lobe short, postocular fixed cheek long and wide; four thoracic segments; macropygous, eight or nine axial rings, eight or nine pleural furrows and seven interpleural furrows well defined, border short, pygidial spines absent.
Occurrence
Mudstone, lime mudstone, wackestone, and grainstone, Mount Cap Formation, Dodo Canyon, upper Albertelloides mischi Zone, Carcajou Falls, Glossopleura walcotti Zone, Northwest Territories.
Description
Exoskeleton is subelliptical in outline. Exoskeleton length 8–33 mm.
Cranidium is subtrapezoidal in outline. Axial furrow is well defined. Strongly convex glabella is subrectangular in outline, extending anteriorly to the border. Glabella narrows gently from the occipital ring to S1 and widens from S1 to the anterior lobe. Four pairs of lateral glabellar furrows are present. S1 is well defined and oriented obliquely backwards, S2 is moderately defined and oriented obliquely backwards, and S3 and S4 are poorly defined or effaced and oriented obliquely forward. Occipital furrow is well defined, the occipital ring bearing a medial tubercle. Length of the anterior border is one-sixth that of the occipital ring. Anterior course of the facial suture is parallel or slightly divergent. Moderately convex interocular area at the widest point is three-fifths the width of the occipital ring. Palpebral lobe is strongly arched, with the anterior tip intersecting the axial furrow opposite S4, and the posterior tip opposite the anterior-half to middle of L1. Length of the palpebral lobe is equal to half that of the glabella. Posterior course of the facial suture is oriented obliquely backwards. Posterior border is equal in width to that of the occipital ring, with a well-defined posterior border furrow.
Free cheek is slightly wider than the occipital ring and the border comprises one-fifth to one-sixth the width. Genal spine is slightly advanced and comprises slightly less than half the length of the free cheek. Free cheek is variably caecate. Terrace lines are present on the border.
Hypostome is conterminant, widening anteriorly, and narrowing gently posteriorly. Medial lobe is covered in terrace lines. The posterior lobe comprises one-quarter of the length of the hypostome.
Thorax consists of four segments. Axial furrow is well defined; each ring bearing a medial tubercle. Pleural width is one-and-one-half times that of the axis, pleura with well-defined pleural and interpleural furrows. Pleural spine is short.
Pygidium is macropygous, subelliptical in outline, and the length is equal to four-fifths the width. Strongly convex axis narrows gently posteriorly, the posterior tip intersecting the border furrow, and is composed of eight or nine rings, each bearing a medial tubercle, and a terminal piece. Axial ring furrows are well to moderately defined. Moderately convex pleural field is slightly wider than the axis, with eight or nine pleural and seven interpleural furrows. The seven anterior pleural and interpleural furrows are well defined and intersect the border furrow. The one or two posterior pleural furrows are poorly defined and do not intersect the border furrow. Flat border comprises one-tenth to slightly less than one-tenth the length of the pygidium. Border spines are absent. Interpleural areas are caecate.
Etymology
Named after the Carcajou River.
Material
Fourteen complete exoskeletons, 52 exoskeletons lacking free cheeks, 17 holaspid cranidia, two meraspid cranidia, one protaspid, three free cheeks, and 30 pygidia. Type material: holotype, GSC 142342; paratypes, GSC 142343–142360.
Remarks
Sahtuia carcajouensis Handkamer and Pratt, n. gen. n. sp., is distinguished by the large size of the pygidium, yet reduced number of thoracic segments in holaspids. The unrelated zacanthoidids Mackenzieaspis parallelispinosa Handkamer and Pratt, n. gen. n. sp., and Dodoella kobayashii Handkamer and Pratt, n. gen. n. sp., also both have four thoracic segments and a large pygidium.
Family Oryctocephalidae Beecher, Reference Beecher1897
Subfamily Oryctocarinae Hupé, Reference Hupé and Piveteau1953
Genus Oryctocara Walcott, Reference Walcott1908
Type species
Oryctocara geikiei Walcott, Reference Walcott1908b, from the Spence Shale, Idaho
Remarks
The diagnosis by Whittington (Reference Whittington1995) is followed herein.
Oryctocara geikiei Walcott, Reference Walcott1908
Figure 20.7–20.10
- Reference Walcott1908b
Oryctocara geikiei Walcott, p. 23, pl. 1, figs. 9, 10.
- Reference Sundberg2020
Oryctocara geikiei; Sundberg, p. 16, fig. 15.1–15.29. [see for synonymy]
Lectotype
Oryctocara geikiei (USNM 53426, 53428) from the Spence Shale, Idaho (Walcott, Reference Walcott1908b, pl. 1, fig. 9; Whittington, Reference Whittington1995, pl. 4, figs. 5, 6).
Occurrence
Mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, Albertelloides mischi Zone or Glossopleura walcotti Zone; Spence Shale, Idaho and Utah, Glossopleura walcotti Zone (Walcott, Reference Walcott1908b; Campbell, Reference Campbell1974; Whittington, Reference Whittington1995); Lakeview Limestone, Idaho, Ptychagnostus praecurrens Zone (Resser, Reference Resser1938, Reference Resser1939a; Sundberg, Reference Sundberg2020); Stephen Formation, British Columbia, Glossopleura walcotti Zone (Whittington, Reference Whittington1995).
Material
Ten cranidia and nine pygidia. Figured material: GSC 142377–142379.
Remarks
Specimens of Oryctocara geikiei Walcott, Reference Walcott1908b, from the Mount Cap Formation resemble the meraspid specimens collected by Sundberg (Reference Sundberg2020, fig. 15.6, 15.8, 15.11, 15.18–15.20) from the Lakeview Limestone. The presence of this taxon in the uppermost strata of Little Bear River may indicate the Glossopleura walcotti Zone (Whittington, Reference Whittington1995; Sundberg, Reference Sundberg2020). However, its co-occurrence with Albertelloides eliasi Handkamer and Pratt, n. sp., as well as the lack of any other taxa indicative of the G. walcotti Zone makes the assignment of either zone uncertain.
Family Zacanthoididae Swinnerton, Reference Swinnerton1915
Remarks
Review of the literature has revealed that Mexicaspis has already been assigned as a genus name. Mexicaspis Lochman, Reference Lochman1948 (type species Mexicaspis stenopyge Lochman, Reference Lochman1948) is preoccupied by Mexicaspis Spaeth, Reference Spaeth1936 (Coptocycla azteca, Champion, Reference Champion1894), a tortoise beetle. The replacement name Mexicaspidella Handkamer and Pratt is proposed herein.
Genus Aitkenaspis Handkamer and Pratt, new genus
Type species
Aitkenaspis keelensis Handkamer and Pratt, n. gen. n. sp., from the Aitkenaspis keelensis Zone of the Mount Cap Formation, Little Bear River, Northwest Territories.
Diagnosis
As for species.
Etymology
Named after James D. Aitken of the GSC who was one of the first researchers to investigate the stratigraphy of the eastern Mackenzie Mountains in detail, and who also carried out ground-breaking studies of the Cambrian of the Canadian Rocky Mountains.
Remarks
Aitkenaspis Handkamer and Pratt, n. gen., resembles Fieldaspis Rasetti, Reference Rasetti1951, and Stephenaspis Rasetti, Reference Rasetti1951. Cranidia of Fieldaspis and Stephenaspis differ by the distance from the posterior tips of the palpebral lobe to the axial furrow (Rasetti, Reference Rasetti1951). This distance in the type species of Stephenaspis, S. bispinosa Rasetti, Reference Rasetti1951, is one-quarter of the width of the occipital ring, whereas in the type species of Fieldaspis, F. furcata Rasetti, Reference Rasetti1951, the distance is one-sixth of the width of the occipital ring. In Aitkenaspis keelensis Handkamer and Pratt, n. gen. n. sp., this distance is the same as in F. furcata. The pygidium of Aitkenaspis n. gen. lacks the border spines or median indentation in the posterior margin present in species of Fieldaspis and Stephenaspis, although the well-defined border furrow of Aitkenaspis n. gen. is within the interspecific variation of Stephenaspis. Thus, Aitkenaspis n. gen. is intermediary between Fieldaspis and Stephenaspis. At present, the genus is monospecific.
Aitkenaspis keelensis Handkamer and Pratt, new species
Figures 19, 20.1–20.6
- ?Reference Butterfield and Nicholas1996
Zacanthoides sp. Butterfield and Nicholas, fig. 2.5.
Aitkenaspis keelensis Handkamer and Pratt, n. gen. n. sp., from the Aitkenaspis keelensis Zone, mudstone, Mount Cap Formation, Little Bear River, Northwest Territories. (1) Holotype exoskeleton lacking free cheeks (dorsal) GSC 142361; (2) paratype exoskeleton lacking free cheeks and the pygidium (dorsal) GSC 142362; (3) paratype exoskeleton lacking free cheeks (dorsal, latex mold) GSC 142363; (4) paratype nearly complete exoskeleton (dorsal) GSC 142364; (5) paratype exoskeleton lacking the cephalon (dorsal, latex mold) GSC 142365; (6) paratype exoskeleton lacking the cephalon (dorsal) GSC 142366; (7) paratype cranidium (dorsal) GSC 142367; (8) paratype exoskeleton lacking the cephalon (dorsal) GSC 142368; (9) paratype free cheek (dorsal) GSC 142369; (10) paratype pygidium (dorsal) GSC 142370. All scale bars = 2 mm.

Corynexochids from the Mount Cap Formation, Northwest Territories. (1–6) Aitkenaspis keelensis Handkamer and Pratt, n. gen. n. sp., from the Aitkenaspis keelensis Zone; (1) paratype hypostome (dorsal) GSC 142371, mudstone, Mount Cap Formation, Little Bear River; (2) paratype cranidium (dorsal, latex mold) GSC 142372, silty mudstone, Mount Clark Formation, Dodo Canyon; (3) paratype pygidium (dorsal) GSC 142373, siltstone, Mount Clark Formation, Dodo Canyon; (4) paratype cranidium (dorsal) GSC 142374, mudstone, Mount Cap Formation, Little Bear River, scale bar = 500 μm; (5) paratype cranidium (dorsal) GSC 142375, silty mudstone, Mount Cap Formation, Little Bear River, scale bar = 1 mm; (6) paratype pygidium (dorsal) GSC 142376, siltstone, Mount Clark Formation, Dodo Canyon. (7–10) Oryctocara geikiei Walcott, Reference Walcott1908b, mudstone, Mount Cap Formation, Little Bear River, Albertelloides mischi or Glossopleura walcotti Zone; (7) cranidium (dorsal) GSC 142377, scale bar = 350 μm; (8) cranidium (dorsal) GSC 142378, scale bar = 400 μm; (9) pygidium (dorsal) GSC 142379, scale bar = 500 μm; (10) pygidium (dorsal) GSC 142377, scale bar = 750 μm. All scale bars = 2 mm unless stated otherwise.

Holotype
Aitkenaspis keelensis (GSC 142361) from the Aitkenaspis keelensis Zone of the Mount Cap Formation, Little Bear River, (17.7 m above the base of the Mount Cap Formation), Northwest Territories (Fig. 19.1).
Diagnosis
Zacanthoidid with glabella anteriorly wide, palpebral lobe long; genal spine non-advanced; nine thoracic segments, pleural spines long, macropleural spines on the fifth and ninth segments; oval-shaped pygidium subisopygous, four axial rings, four pleural furrows, border furrow well defined, border spines absent.
Occurrence
Silty mudstone and siltstone, Mount Clark Formation, Dodo Canyon, mudstone and silty mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, Aitkenaspis keelensis Zone.
Description
Exoskeleton is subelliptical in outline. Exoskeleton length 10–20 mm.
Cranidium is subquadrate in outline. Axial furrow is well defined. Glabella is subquadrate in outline, extending anteriorly to the border. Glabella narrows gently from the occipital ring to S1 and widens from S1 to the anterior lobe. Four pairs of lateral glabellar furrows are present. S1 is well defined and oriented obliquely backwards; S2 is poorly defined and nearly transverse; and S3 and S4 are poorly defined and oriented obliquely forward. Occipital furrow is well defined, the occipital ring bearing a medial tubercle. Length of the anterior border is one-sixth that of the occipital ring. Anterior course of the facial suture is divergent. Interocular area at the widest point is one-third to half the width of the occipital ring. Palpebral lobe is strongly arched, with the anterior tip intersecting the axial furrow opposite S3, and the posterior tip opposite the occipital furrow. Length of the palpebral lobe is two-thirds that of the glabella. Posterior course of the facial suture is transverse. Posterior border is equal in width to the occipital ring, with a poorly defined posterior border furrow. Intergenal spines possibly present, but poorly preserved if so.
Free cheek is slightly wider than the occipital ring and the border comprises one-fifth the width. Genal spine is non-advanced and comprises half of the length of the free cheek.
Hypostome is conterminant, widening anteriorly and narrowing gently posteriorly. Anterior lobe is covered in terrace lines. Posterior lobe comprises one-sixth the length.
Thorax consists of nine segments. Axial furrow is well defined; each axial ring bearing a medial tubercle. Pleura width is one-and-one-half to twice that of the axis, with well-defined pleural and interpleural furrows. Thoracic spines are moderately long; posteriorly directed macropleural spines are present on segments five and nine, with the latter extending just past to well beyond the posterior edge of the pygidium.
Pygidium subisopygous, oval in outline, and the length is equal to two-thirds the width. Axis narrows posteriorly, with the posterior tip anterior of the border furrow, and is composed of four rings, each bearing a medial tubercle, and a terminal piece. Ring furrows are moderately defined. Pleural field width is two-thirds that of the axis and has four pleural furrows, which are poorly to well defined and do not intersect the well-defined border furrow. Border comprises one-eighth the length of the pygidium. Border spines are absent. Terrace lines are variably present on the border.
Etymology
Named after the Keele River in the eastern Mackenzie Mountains.
Material
Two complete exoskeletons, 13 exoskeletons lacking free cheeks, 46 cranidia, 12 free cheeks, 13 thoracic segments, 27 pygidia, and one hypostome. Type material: holotype, GSC 142361; paratypes, GSC 142362–142376.
Remarks
Zacanthoides sp. in Butterfield and Nicholas (Reference Butterfield and Nicholas1996) has a nearly identical cephalon and thorax to that of Aitkenaspis keelensis Handkamer and Pratt, n. gen. n. sp. The pygidium of that specimen is partially covered by matrix and thus cannot be compared to the pygidium of A. keelensis n. gen. n. sp.
Genus Albertella Walcott, Reference Walcott1908
Type species
Albertella helena Walcott, Reference Walcott1908b, from the Albertella Zone of the Gordon Shale, Montana.
Remarks
Albertella Walcott, Reference Walcott1908b, has a subrectangular glabella, a moderately long cranidial border, a moderately short palpebral lobe with the posterior tip opposite S1, a wide interocular area, a narrow free cheek, a macropleural third thoracic segment, a pygidium that is wider than long, a pygidial axis composed of four rings and a terminal piece, and a posterolaterally oriented pygidial border spine (Rasetti, Reference Rasetti1951; Palmer and Halley, Reference Palmer and Halley1979). This genus has been recognized in the Mount Cap Formation of the Colville Hills (Sommers et al., Reference Sommers, Gingras, MacNaughton, Fallas and Morgan2020).
Albertella levis Walcott, Reference Walcott1917
Figure 21.1–21.6
- Reference Walcott1917
Albertella levis Walcott, p. 39, pl. 7, fig. 1, 1a.
Zacanthoidids from the Mount Cap Formation, Carcajou Falls, Northwest Territories. (1–6) Albertella levis Walcott, Reference Walcott1917, Glossopleura walcotti Zone; (1) cranidium (dorsal) GSC 142380, silty mudstone; (2, 3) pygidium (dorsal, oblique) GSC 142381, grainstone; (4) cranidium (dorsal) GSC 142382, silty mudstone; (5) pygidium (dorsal) GSC 142383, silty mudstone; (6) pygidium (dorsal, latex mold) GSC 142384, silty mudstone. (7–11) Albertelloides mischi Fritz, Reference Fritz1968, silty mudstone, Albertelloides mischi Zone; (7) cranidium (dorsal) GSC 142385; (8) cranidium (dorsal) GSC 142386; (9) pygidium (dorsal) GSC 142387; (10) cranidium (dorsal) GSC 142388; (11) pygidium (dorsal) GSC 142389. (12) Albertelloides pandispinata Fritz, Reference Fritz1968, pygidium (dorsal) GSC 142390, silty mudstone, Albertelloides mischi Zone. All scale bars = 2 mm.

Holotype
Cranidium (USNM 63758) from the Albertella Zone of the Chetang Formation, Alberta (Walcott, Reference Walcott1917, pl. 7, fig. 1).
Occurrence
Silty mudstone and mudstone, Mount Cap Formation, Dodo Canyon, Albertelloides mischi Zone, Carcajou Falls, Glossopleura walcotti Zone, Northwest Territories; Chetang Formation, Alberta, Albertella Zone (Walcott, Reference Walcott1917).
Material
Twelve cranidia and 14 pygidia. Figured material: GSC 142380–142384.
Remarks
Albertella levis is intermediate between Albertella and Paralbertella Palmer in Palmer and Halley, Reference Palmer and Halley1979. Features shared with Albertella include a moderately short palpebral lobe, a pygidium that is wider than long, poorly defined pleural furrows, and a pair of posterolaterally oriented border spines. Features shared with Paralbertella include an anteriorly wide glabella and a pygidial axis composed of five rings. The pygidium of Albertelloides eliasi Handkamer and Pratt, n. sp., is broadly similar to Albertella levis, but the exoskeleton of A. eliasi n. sp. is distinguished by having a longer palpebral lobe, well-defined pygidial furrows, and a shorter pygidial axis. Pygidia from the Mount Cap Formation differ from those from the Chetang Formation by having a slightly shorter pygidial axis, which is slightly anterior to the border furrow.
Genus Albertelloides Fritz, Reference Fritz1968
Type species
Albertelloides mischi Fritz, Reference Fritz1968, from the Albertella Zone of the Pioche Formation, Nevada.
Remarks
Albertelloides Fritz, Reference Fritz1968, differs from Albertella by having a wider anterior glabellar lobe, a longer palpebral lobe, a shorter postocular fixed cheek, a narrower, less-convex interocular fixed cheek, a wider free cheek, a non-advanced genal spine, wider thoracic pleura, and a more posteriorly located pygidial border spine. Albertelloides differs from Paralbertella by having a wider anterior glabellar lobe, a non-advanced genal spine, a thorax composed of eight instead of seven segments, four to five instead of five to seven pygidial axial rings, less well-defined pleural furrows, a shallower border furrow, and a more posteriorly located border spine.
Fritz (Reference Fritz1968) described three species, Albertelloides mischi, A. pandispinata, and A. sp. indet. Campbell (Reference Campbell1974) recognized an additional species. Albertelloides rectimarginatus Palmer in Palmer and Halley, Reference Palmer and Halley1979, and more complete material belonging to A. mischi were described from the Carrara Formation (Palmer and Halley, Reference Palmer and Halley1979), and A. kitai Eddy and McCollum, Reference Eddy and McCollum1998, was described from the Pioche Formation, all from Nevada. Fritz (Reference Fritz1968) proposed that the pygidia of Kochaspis maladensis Resser, Reference Resser1939b, and K. dispar Resser, Reference Resser1939b (Resser, Reference Resser1939b, pl. 13, figs. 9–12, 13–15) belong to Albertelloides. Sundberg and McCollum (Reference Sundberg and McCollum2003a) argued that these pygidia belong to a corynexochid, but not Albertelloides due to the fewer axial rings. The species described by Campbell (Reference Campbell1974, pl. 15, figs. 1, 2, 6, 7) probably also does not belong to Albertelloides for the same reason.
Albertelloides mischi, A. kitai, A. pandispinata, and A. rectimarginatus all have four or five pygidial axial rings. These species are distinguished by the variable length, width, and orientation of the pygidial border spine. Albertelloides mischi and A. pandispinata have long and narrow border spines, but differ in the orientation: subparallel in A. mischi and divergent in A. pandispinata. Albertelloides rectimarginatus and A. kitai, by contrast, have short and wide border spines: parallel and sharp-tipped in A. rectimarginatus, whereas they are blunt-tipped in A. kitai (Eddy and McCollum, Reference Eddy and McCollum1998). The interocular area is wider in A. rectimarginatus and A. kitai and narrower in A. mischi. The cranidium of A. pandispinata is unknown.
Albertelloides eliasi Handkamer and Pratt, new species
Figure 22.4–22.13
Holotype
Pygidium (GSC 142396) from the Albertelloides mischi Zone of the Mount Cap Formation, Carcajou Falls (4.4 m above the base of the Mount Cap Formation), Northwest Territories (Fig. 22.6).
Species of Albertelloides Fritz, Reference Fritz1968, from the Albertelloides mischi Zone, silty mudstone, Mount Cap Formation, Carcajou Falls, Northwest Territories. (1–3) Albertelloides mischi Fritz, Reference Fritz1968; (1) cranidium (dorsal, latex mold) GSC 142391; (2) cranidium (dorsal) GSC 142392; (3) pygidium (dorsal) GSC 142393. (4–13) Albertelloides eliasi Handkamer and Pratt, n. sp.; (4) paratype cranidium (dorsal) GSC 142394; (5) paratype cranidium (dorsal) GSC 142395; (6) holotype pygidium (dorsal) GSC 142396; (7) paratype cranidium (dorsal) GSC 142397; (8) paratype pygidium (dorsal) GSC 142398; (9) paratype free cheek (dorsal) GSC 142399; (10) paratype partially articulated exoskeleton (dorsal) GSC 142400; (11) paratype pygidium (dorsal) GSC 142401; (12) paratype partially articulated exoskeleton (dorsal, latex mold) GSC 142402; (13) paratype free cheek (dorsal) GSC 142403. All scale bars = 2 mm.

Diagnosis
Albertelloides with cranidial border short, posterior border oriented gently posterolaterally; free cheek wide, border of free cheek narrow; thoracic segments with moderately long pleural spines; pygidial pleural field wide, three pleural furrows well defined, border short, bearing one pair of moderately short, narrow, and subparallel-oriented spines.
Occurrence
Calcareous siltstone and silty mudstone, Mount Cap Formation, Carcajou Falls and Little Bear River, Northwest Territories, Albertelloides mischi Zone.
Description
Cranidium is subtriangular in outline. Axial furrow is well defined. Glabella is subparallel, extending anteriorly to the border. Glabella narrows from the occipital ring to S1 and widens from S1 to the anterior lobe. Four pairs of lateral glabellar furrows are present. S1 is well defined and oriented obliquely backwards; S2 is poorly defined and transverse; and S3 and S4 are poorly defined and oriented obliquely forward. Occipital furrow is well defined. Length of the anterior border is one-quarter that of the occipital ring. Anterior course of the facial suture is parallel to slightly divergent. Interocular area at the widest point is half to two-thirds the width of the occipital ring. Palpebral lobe is strongly arched, with the anterior tip intersecting the axial furrow opposite L4, and the posterior tip opposite the occipital furrow. Length of the palpebral lobe is two-thirds that of the glabella. Posterior course of the facial suture is oriented gently posterolaterally. Posterior border width is equal to the occipital ring, with a moderately defined border furrow.
Border of the free cheek comprises one-fifth of the width, bearing terrace lines. Genal spine is non-advanced and comprises one-third the length.
Hypostome is unknown.
Thorax consists of at least five segments. Axial furrow is well defined. Pleura width is one-and-one-half times that of the axis, with well-defined pleural and interpleural furrows. Thoracic spines are moderately long.
Pygidium is semicircular in outline and in length equal to two-thirds the width. Axis is parallel-sided, with the posterior end anterior to or intersecting the border furrow, and is composed of four to five rings and a terminal piece. Axial ring furrows are moderately to poorly defined. Pleural field width is equal to that of the axis, with four pleural furrows and three interpleural furrows. The three anterior pleural and interpleural furrows are well defined and intersect the border furrow. The posterior pleural furrow is poorly defined and does not intersect the border furrow. Border comprises one-tenth the length of the pygidium. Border spine is opposite the fourth pygidial pleural furrow, oriented subparallel, and is slightly shorter than half the length of the pygidial axis.
Etymology
Named after Robert J. Elias, Professor, University of Manitoba, Winnipeg, Manitoba.
Material
Three incomplete exoskeletons, 28 cranidia, 10 free cheeks, and 13 pygidia. Type material: holotype, GSC 142396; paratypes, GSC 142394, 142395, 142397–142403.
Remarks
Albertelloides eliasi Handkamer and Pratt, n. sp., differs from other species of Albertelloides by having shorter cranidial and pygidial borders, a posterior cranidial border oriented slightly obliquely backwards, a wider pygidial pleural field, and a short and narrow border spine. Variation in the number of pygidial axial rings and orientation of the border spine are interpreted as intraspecific variation (Fig. 22.6, 22.7, 22.10).
Albertelloides mischi Fritz, Reference Fritz1968
Figures 21.7–21.11, 22.1–22.3
- Reference Fritz1968
Albertelloides mischi Fritz, p. 215, pl. 38, figs. 1–7.
- Reference Palmer and Halley1979
Albertelloides mischi; Palmer and Halley, p. 90, pl. 10, figs. 7–13.
Holotype
Pygidium (USNM 153568) from the Albertella Zone of the Pioche Formation, Nevada (Fritz, Reference Fritz1968, pl. 38, figs. 5, 7).
Occurrence
Calcareous siltstone and silty mudstone, Mount Cap Formation, Carcajou Falls and Grafe River, Northwest Territories, Albertelloides mischi Zone; upper member, Pioche Formation, Nevada, Albertella Zone (Fritz, Reference Fritz1968); Carrara Formation, Nevada, Zacanthoidid Zonule, Albertella Zone (Palmer and Halley, Reference Palmer and Halley1979).
Material
Thirty-five cranidia, six free cheeks, and 21 pygidia. Figured material: GSC 142385–142389, 142391–142393.
Remarks
Specimens from the Mount Cap Formation differ from those of the Pioche and Carrara formations by having a slightly shorter anterior cranidial border, which is interpreted as intraspecific variation (Fritz, Reference Fritz1968, pl. 38, figs. 1–3, 6; Palmer and Halley, Reference Palmer and Halley1979, pl. 10, figs. 8, 9, 13). Albertelloides rectimarginatus also exhibits minor variation in the length of the anterior border (Palmer and Halley, Reference Palmer and Halley1979, pl. 10, figs. 16, 17). The less well-defined pygidial border furrow and presence of a longitudinal groove on the border spines are attributed to compaction.
Albertelloides pandispinata Fritz, Reference Fritz1968
Figure 21.12
- Reference Fritz1968
Albertelloides pandispinata Fritz, p. 216, pl. 39, figs. 31, 32.
Holotype
Pygidium (USNM 153597) from the Albertella Zone of the Pioche Formation, Nevada (Fritz, Reference Fritz1968, pl. 39, figs. 31, 32).
Occurrence
Calcareous siltstone and silty mudstone, Mount Cap Formation, Carcajou Falls, Northwest Territories, Albertelloides mischi Zone; upper member, Pioche Formation, Nevada, Albertella Zone (Fritz, Reference Fritz1968).
Material
Nine pygidia. Figured material: GSC 142390.
Remarks
Albertelloides pandispinata from the Mount Cap Formation is nearly identical to the pygidium illustrated by Fritz (Reference Fritz1968), although some pygidia differ by having five axial rings as opposed to four. This is interpreted as intraspecific variation by analogy with A. mischi, because this species, which is from both the Pioche and Mount Cap formations, ranges from five to six axial rings (Figs. 20.9, 21.3). Because the cranidium of A. pandispinata is unknown, it is possible that some cranidia assigned here to A. mischi belong instead to A. pandispinata.
Genus Dodoella Handkamer and Pratt, new genus
Type species
Dodoella kobayashii Handkamer and Pratt, n. sp., from the Albertelloides mischi Zone of the Mount Cap Formation, Dodo Canyon (20.4 m above the base of the Mount Cap Formation), Northwest Territories.
Diagnosis
As for species.
Etymology
Named after Dodo Canyon, Canyon Ranges, eastern Mackenzie Mountains.
Remarks
Dodoella Handkamer and Pratt, n. gen., is currently monospecific, and the most diagnostic feature is the thorax consisting of four segments. The presence of a short palpebral lobe, anteriorly widened and effaced glabella, effaced pygidial axis and pleural field, and long pygidial border spines indicate that this species belongs to Zacanthoididae and is closely related to Mexicaspidella Handkamer and Pratt, n. gen. (replacement name for preoccupied Mexicaspis) and less so to Albertella. Mexicaspidella radiatus (Palmer in Palmer and Halley, Reference Palmer and Halley1979) n. comb. has twice as many segments as the type species, Dodoella kobayashii Handkamer and Pratt, n. gen. n. sp.
Dodoella kobayashii Handkamer and Pratt, new species
Figure 24.2, 24.5
Holotype
Dodoella kobayashii (GSC 142414) from the Albertelloides mischi Zone of the Mount Cap Formation, Northwest Territories (Fig. 24.2).
Diagnosis
Zacanthoidid with axial furrow moderately defined, two pairs of lateral glabellar furrows poorly defined, palpebral lobe short, posterior border long and narrow; genal spine oriented obliquely backwards; four thoracic segments; macropygous, ring furrows effaced, pleural field narrow, pleural furrows poorly defined or effaced, one pair of border spines oriented obliquely backward or subparallel.
Occurrence
Mudstone, Mount Cap Formation, Dodo Canyon, Northwest Territories, Albertelloides mischi Zone.
Description
Exoskeleton is subelliptical in outline. Exoskeleton length 16 mm.
Cranidium is subtrapezoidal in outline. Axial furrow is moderately defined. Glabella is subrectangular in outline, extending anteriorly to the border. The glabella widens gently from S1 to the anterior lobe. One pair of lateral glabellar furrows is present. S1 is poorly defined and oriented obliquely backwards. Occipital furrow is moderately defined. Anterior border and anterior course of the facial suture are unknown. Interocular area at the widest point in width is three-quarters that of the occipital ring. Palpebral lobe is strongly arched, with the anterior tip intersecting the axial furrow one-third of the glabellar length from the anterior end, and the posterior tip opposite L1. Length of the palpebral lobe is half that of the glabella. Posterior course of the facial suture is oriented obliquely backwards. Posterior border is slightly narrower than the occipital ring, with a well-defined border furrow.
Free cheek is slightly narrower than the occipital ring; the border comprises one-fourth of the width. Genal spine is slightly advanced, oriented obliquely backwards, and comprises three-fifths the length of the free cheek.
Hypostome is unknown.
Thorax consists of four segments. Axial furrow is moderately defined. Pleura is equal in width to the axis, with a moderately defined pleural furrow. Thoracic segments terminate in a short spine.
Pygidium macropygous, semicircular in outline, and the length is equal to five-sevenths the width. Moderately convex axis narrows gently posteriorly, with the posterior tip slightly anterior to or intersecting the border furrow. Ring furrows are effaced. Gently convex pleural field is slightly narrower than the axis, with one or two pairs of poorly defined pleural furrows that intersect the border furrow. Flat border comprises less than one-tenth the length of the pygidium. Flat, narrow border spine is oriented obliquely backwards or subparallel, curving medially, and in length is slightly shorter than the pygidial axis.
Etymology
Named after Teiichi Kobayashi, who first described the Cambrian and Lower Ordovician trilobites of the Mackenzie River valley region.
Material
One nearly complete exoskeleton lacking a free cheek; one pygidium. Type material: holotype, GSC 142414; paratype, GSC 142417.
Remarks
One isolated pygidium (GSC 142417) is assigned to this species, along with the holotype (GSC 142414). GSC 142417 has a slightly shorter axis that does not intersect the border furrow, and wider, more strongly curved, and subparallel rather than divergent, border spines. This is interpreted as intraspecific variation, because similar variation is observed in other species such as Paralbertella bosworthi (Walcott, Reference Walcott1908b) (Walcott, Reference Walcott1917, pl. 7, fig. 3, 3a; Rasetti, Reference Rasetti1951, pl. 17, figs. 1–4, 9) and Albertella microps Rasetti, Reference Rasetti1951 (Rasetti, Reference Rasetti1951, pl. 19, figs. 1, 3, 7).
Genus Mackenzieaspis Handkamer and Pratt, new genus
Type species
Mackenzieaspis parallelispinosa Handkamer and Pratt, n. sp. from the Glossopleura walcotti Zone of the Mount Cap Formation, Northwest Territories.
Diagnosis
Zacanthoidid with a glabella subrectangular, palpebral lobe long; advanced genal spines long; two or four thoracic segments, axis and pleura equal in width; macropygous, six or eight axial rings, three to five pleural furrows well defined, one pair of posteriorly located border spines.
Etymology
Named after the Mackenzie Mountains.
Remarks
Mackenzieaspis Handkamer and Pratt, n. gen. is regarded as a zacanthoidid because it has a long, subrectangular glabella, a long palpebral lobe, a short intergenal spine, and an advanced and long genal spine. This is supported by the presence of moderately long thoracic and pygidial border spines, in common with many other zacanthoidids. This genus includes two new species that have relatively few thoracic segments and larger pygidia compared to other zacanthoidids. The cephalon of Mackenzieaspis n. gen. is most-similar to that of Paralbertella and Albertelloides.
Mackenzieaspis parallelispinosa Handkamer and Pratt, new species
Figures 23, 24.3, 24.4, 24.6–24.11
Mackenzieaspis parallelispinosa Handkamer and Pratt, n. gen. n. sp., from the Glossopleura walcotti Zone, mudstone, Mount Cap Formation, Carcajou Falls, Northwest Territories. (1) Holotype complete exoskeleton (dorsal) GSC 142404; (2) paratype fractured exoskeleton (dorsal) GSC 142405; (3) paratype complete exoskeleton (dorsal, latex mold) GSC 142406; (4) paratype complete exoskeleton (dorsal, latex mold) GSC 142407; (5) paratype complete exoskeleton (dorsal) GSC 142408; (6) paratype exoskeleton lacking free cheeks (dorsal) GSC 142409; (7) paratype fractured exoskeleton (dorsal) GSC 142410; (8) paratype exoskeleton lacking free cheeks (dorsal, latex mold) GSC 142411; (9) paratype complete exoskeleton (dorsal) GSC 142412. All scale bars = 2 mm.

Zacanthoidids from the Mount Cap Formation, Northwest Territories. (1) Mackenzieaspis divergens Handkamer and Pratt, n. gen. n. sp., holotype disarticulated exoskeleton (dorsal, latex mold) GSC 142413, mudstone, Carcajou Falls, Glossopleura walcotti Zone. (2, 5) Dodoella kobayashii Handkamer and Pratt, n. gen. n. sp., mudstone, Dodo Canyon, Albertelloides mischi Zone; (2) holotype exoskeleton lacking a free cheek (dorsal, latex mold) GSC 142414; (5) paratype pygidium (dorsal) GSC 142417. (3, 4, 6–11) Mackenzieaspis parallelispinosa Handkamer and Pratt, n. gen., n. sp., Carcajou Falls, Glossopleura walcotti Zone; (3) paratype exoskeleton lacking free cheeks (dorsal) GSC 142415, mudstone; (4) paratype pygidium (dorsal) GSC 142416, mudstone; (6) paratype pygidium (dorsal) GSC 142418, mudstone; (7, 8) paratype cranidium (dorsal, oblique) GSC 142353, grainstone; (9) paratype cranidium (dorsal) GSC 142419, mudstone; (10) paratype cranidium (dorsal) GSC 142420, mudstone; (11) paratype free cheek (dorsal) GSC 142349, mudstone. All scale bars = 2 mm.

Holotype
Mackenzieaspis parallelispinosa (GSC 142404) from the Glossopleura walcotti Zone of the Mount Cap Formation, Carcajou Falls (26.1 m above the base of the Mount Cap Formation), Northwest Territories (Fig. 23.1).
Diagnosis
Mackenzieaspis Handkamer and Pratt, n. gen. with pits along occipital furrow, palpebral lobe long; free cheek border narrow, genal spine short to moderately long, advanced, and directed posteriorly; four thoracic segments; macropygous, pygidium semicircular in outline, eight axial rings, pleural field wide, four or five pleural furrows, parallel border spines moderately long.
Occurrence
Mudstone, lime mudstone, and grainstone, Mount Cap Formation, Carcajou Falls, Northwest Territories, Glossopleura walcotti Zone.
Description
Exoskeleton is subelliptical in outline. Exoskeleton length 10–31 mm.
Cranidium is subquadrate in outline. Axial furrow is well defined. Strongly convex glabella is subrectangular in outline, extending anteriorly to the border. Four pairs of lateral glabellar furrows are present. S1 and S2 are moderately defined and oriented obliquely backward; S3 is poorly defined or effaced and transverse; and S4 is poorly defined or effaced and oriented obliquely forwards. The occipital furrow is moderately defined with one pair of pits present at the intersection with the axial furrow; the occipital ring bears a medial tubercle. Length of the anterior border is equal to one-quarter that of the occipital ring. Anterior course of the facial suture is parallel. Moderately convex interocular area at the widest point is slightly narrower than the occipital ring. Palpebral lobe is strongly arched, with the anterior tip intersecting the axial furrow at S4, and the posterior tip opposite the occipital furrow. Length of the palpebral lobe is two-thirds that of the glabella. Posterior course of the facial suture is transverse. Posterior border width is four-fifths that of the occipital ring, with a moderately defined border furrow and a short intergenal spine.
Free cheek is slightly wider than the occipital ring and the border comprises one-seventh the width. Genal spine is advanced, oriented posteriorly, and comprises half to three-fifths the length of the free cheek.
Hypostome is conterminant, widening anteriorly, and narrowing gently posteriorly. Anterior and medial lobes bear terrace lines. Posterior lobe comprises one-seventh the length.
Thorax consists of four segments. Axial furrow is well defined; each ring bearing a medial tubercle. Pleura width is equal to the axis, with well-defined pleural and interpleural furrows. Thoracic spines are moderately long.
Pygidium macropygous, semicircular in outline, and in length is equal to two-thirds the width. Axis narrows gently posteriorly, the posterior end intersecting the border furrow, and is composed of eight rings, each with a medial tubercle, and a terminal piece. Axial ring furrows are well to moderately defined. Pleural field is slightly wider than the axis, with four or five pleural furrows and three interpleural furrows. The three anterior pleural and interpleural furrows are well defined and intersect the border furrow. The one or two posterior pleural furrows are moderately to poorly defined and do not intersect the border furrow. Border comprises one-tenth the length of the pygidium. One pair of border spines is present opposite the fourth pleural furrow. Border spine is subparallel and half to one-and-one-half times the length of the pygidial axis.
Etymology
Named after the subparallel pygidial border spines.
Material
Twelve complete exoskeletons, 25 exoskeletons lacking free cheeks, 26 cranidia, eight free cheeks, and 44 pygidia. Type material: holotype, GSC 142404; paratypes GSC 142349, 142353, 142405–142412, 142415, 142416, 142418–142420.
Remarks
Mackenzieaspis parallelispinosa Handkamer and Pratt, n. gen. n. sp., differs from M. divergens Handkamer and Pratt, n. gen. n. sp., by having a slightly longer palpebral lobe, advanced and parallel genal spines, and a semicircular pygidium with more axial rings, a wider pleural field, more pleural furrows, and parallel border spines.
Mackenzieaspis divergens Handkamer and Pratt, new species
Figure 24.1
Holotype
Mackenzieaspis divergens (GSC 142413) from the Glossopleura walcotti Zone of the Mount Cap Formation, Carcajou Falls (24.4 m above the base of the Mount Cap Formation), Northwest Territories (Fig. 24.1).
Diagnosis
Mackenzieaspis Handkamer and Pratt, n. gen., with palpebral lobe short; free cheek border wide, genal spine long, less advanced, and oriented obliquely backwards; at least two thoracic segments; isopygous, pygidium subtriangular in outline, six axial rings, pleural field narrow, three pleural furrows, border spines extending from pleural field, oriented posterolaterally.
Occurrence
Mudstone, Mount Cap Formation, Carcajou Falls, Northwest Territories, Glossopleura walcotti Zone.
Description
Complete exoskeleton is unknown.
Cranidium is subquadrate in outline. Axial furrow is well defined. Glabella is subrectangular in outline, extending anteriorly to the border. At least one pair of lateral glabellar furrows is present. S1 is poorly defined and oriented obliquely backwards. S2, S3, and S4 are not preserved. Occipital furrow is well defined. Anterior border is in length one-quarter that of the occipital ring. Anterior course of the facial suture is parallel. Interocular area at the widest point is in width half that of the occipital ring. Palpebral lobe is strongly arched, with the anterior tip intersecting the axial furrow one-third the glabellar length from the anterior end, and the posterior tip opposite the middle of L1. Length of the palpebral lobe is half that of the glabella. Posterior course of the facial suture is transverse. Posterior border is slightly narrower than the occipital ring, with a well-defined border furrow.
Free cheek width is equal to that of the occipital ring and the border comprises one-fifth of the width. Genal spine is slightly advanced, oriented posterolaterally, and comprises two-thirds of the length of the free cheek.
Hypostome is unknown.
Thorax consists of at least two segments. Axial furrow is well defined. Pleura width is equal to the axis, with a well-defined pleural furrow. Tips of segments are unknown.
Pygidium isopygous, subtriangular in outline, and in length is equal to four-fifths the width. Axis narrows gently posteriorly; the posterior end intersecting the border furrow, and is composed of at least six axial rings. Ring furrows are poorly defined. Pleural field is equal in width to the axis, with three pleural furrows. The two anterior pleural furrows are well defined and intersect the border furrow. The posterior pleural furrow is moderately defined and does not intersect the border furrow. Length of the border is unknown. One pair of spines extends from the pleural field posterior of the third pleural furrow; the spines are oriented posterolaterally and are twice the length of the axis.
Etymology
Named after the strongly divergent pygidial spines.
Material
One nearly complete exoskeleton. Type material: holotype, GSC 142413.
Remarks
Aside from the distinctively low number of thoracic segments, the pair of strongly divergent pygidial spines that extends from the pleural field of M. divergens Handkamer and Pratt, n. gen., n. sp., is unusual, but similar to spines present in Albertella longwelli Palmer in Palmer and Halley, Reference Palmer and Halley1979, and A. spectrensis Palmer in Palmer and Halley, Reference Palmer and Halley1979.
Corynexochid genus and species indet.
Figure 25.13, 25.14
Occurrence
Silty mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, Amecephalus arrojosensis-Eokochaspis nodosa Zone.
Trilobites of the Mount Cap Formation, Northwest Territories. (1) Chancia maladensis (Resser, Reference Resser1939b), exoskeleton lacking free cheeks (dorsal) GSC 142436, mudstone, Mount Cap Formation, Dodo Canyon, Glossopleura walcotti Zone; (2, 3) Caborcella collaris (Rasetti, Reference Rasetti1951), cranidium (dorsal, oblique) GSC 142437, siltstone, Mount Clark Formation, Dodo Canyon, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (4, 7) Eokochaspis sp. indet., mudstone, Mount Cap Formation, Little Bear River, Aitkenaspis keelensis Zone; (4) exoskeleton lacking the cephalon (dorsal, latex mold) GSC 142438; (7) cranidium (dorsal, latex mold) GSC 142441; (5, 9, 10) ptychoparioid gen. and sp. indet. 2, silty mudstone, Mount Cap Formation, Carcajou Falls, Albertelloides mischi Zone; (5) cranidium (dorsal) GSC 142439; (9) cephalon (dorsal) GSC 142443; (10) cranidium (dorsal) GSC 142444; (6, 8) Tonopahella goldfieldensis Sundberg and McCollum, Reference Sundberg and McCollum2003b, silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (6) cranidium (dorsal) GSC 142440; (8) cranidium (dorsal) GSC 142442; (11, 12) ptychoparioid gen. and sp. indet. 1, cranidium (dorsal, frontal) GSC 142445; silty mudstone, Mount Clark Formation, Dodo Canyon, Aitkenaspis keelensis Zone; (13, 14) corynexochid gen. and sp. indet., silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (13) cranidium (dorsal) GSC 142446; (14) free cheek (dorsal) GSC 142447. All scale bars = 2 mm.

Material
Nine cranidia, three free cheeks, and one partial thorax consisting of five segments. Figured material: GSC 142446, 142447.
Remarks
The lack of a pygidium limits identification because early middle Cambrian corynexochids have morphologically conservative cranidia, such as in species of Fieldaspis, Stephenaspis, and Aitkenaspis n. gen.
Order Ptychopariida Swinnerton, Reference Swinnerton1915
Suborder Ptychopariina Richter, Reference Richter, Dittler, Joos, Korschelt, Linek, Oltmanns and Schaum1932
Superfamily Ptychoparioidea Matthew, Reference Matthew1887
Family Ptychopariidae Matthew, Reference Matthew1887
Subfamily uncertain
Genus Caborcella Lochman, Reference Lochman1948
Type species
Caborcella arrojosensis Lochman, Reference Lochman1948, from the Arrojos Formation, Sonora, Mexico.
Remarks
Caborcella Lochman, Reference Lochman1948, is characterized by a concave preglabellar field with a well-defined border furrow (Lochman, Reference Lochman1948; Palmer and Halley, Reference Palmer and Halley1979). Nyella Palmer in Palmer and Halley, Reference Palmer and Halley1979, is similar, but differs by having a convex preglabellar field.
Caborcella collaris (Rasetti, Reference Rasetti1951)
Figures 25.2, 25.3, 26.14, 26.15
- Reference Rasetti1951
Schistometopus collaris Rasetti, p. 239, pl. 14, figs. 1–3.
Ptychoparioids from the Northwest Territories. (1–7) Tonopahella walcotti (Sundberg and McCollum, Reference Sundberg and McCollum2000); (1) exoskeleton lacking free cheeks (dorsal, latex mold) GSC 142421, silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (2) cranidium (dorsal) GSC 142422, silty mudstone, Mount Clark Formation, Dodo Canyon, Aitkenaspis keelensis Zone; (3) cranidium (dorsal) GSC 142423, siltstone, Mount Clark Formation, Dodo Canyon, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (4) partially complete specimen (dorsal, latex mold) GSC 142424, silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (5) free cheek (dorsal) GSC 142425, silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (6) free cheek (dorsal, latex mold) GSC 142426, silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (7) pygidium, (dorsal, latex mold) GSC 142427, silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (8) Eokochaspis nodosa Sundberg and McCollum, Reference Sundberg and McCollum2000, pygidium (dorsal) GSC 142428, silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (9–11) Tonopahella sp. indet., silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (9) cranidium (dorsal) GSC 142429; (10) cranidium (dorsal, latex mold) GSC 142430; (11) cranidium (dorsal) GSC 142431; (12) ptychoparioid gen. and sp. indet. 4, complete exoskeleton (dorsal, latex mold) GSC 142432, silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (13) ptychoparioid gen. and sp. indet. 5, cranidium (dorsal) GSC 142433, silty mudstone, Mount Cap Formation, Little Bear River, Amecephalus arrojosensis-Eokochaspis nodosa Zone; (14, 15) Caborcella collaris (Rasetti, Reference Rasetti1951), cranidium (dorsal, lateral) GSC 142434, silty mudstone, Mount Clark Formation, Dodo Canyon, Aitkenaspis keelensis Zone; (16, 17) ptychoparioid gen. and sp. indet. 3, exoskeleton lacking free cheeks (oblique, dorsal) GSC 142435, mudstone, Mount Cap Formation, Carcajou Falls, Glossopleura walcotti Zone. All scale bars = 2 mm.

Holotype
Cranidium (USNM 116123) from the Plagiura-Kochaspis Zone of the Mount Whyte Formation, Alberta (Rasetti, Reference Rasetti1951, pl. 14, figs. 1–3).
Occurrence
Siltstone and silty mudstone, Mount Clark Formation, Dodo Canyon, Northwest Territories, Amecephalus arrojosensis-Eokochaspis nodosa and Aitkenaspis keelensis zones; Mount Whyte Formation, Alberta and British Columbia, Plagiura-Kochaspis Zone (Rasetti, Reference Rasetti1951).
Material
Five cranidia. Figured material: GSC 142434, 142437.
Remarks
Owing to the poor quality of the holotype of Schistometopus typicalis Resser, Reference Resser1938, which is the type species for the genus Schistometopus, Sundberg (Reference Sundberg2004) advocated for the reassignment of existing species to other genera. Sundberg (Reference Sundberg2004) suggested that S. collaris belongs in Caborcella. Compared to the holotype cranidium, the cranidia from the Mount Cap Formation have a slightly longer anterior border. The presence of an occipital spine is uncertain because of poor preservation.
Genus Chancia Walcott, Reference Walcott1924
Type species
Chancia ebdome Walcott, Reference Walcott1924, from the Glossopleura walcotti Zone of the Spence Shale, Langston Formation, Idaho (Maxey, Reference Maxey1958; Oriel and Armstrong, Reference Oriel and Armstrong1971).
Remarks
The diagnosis of Chancia Walcott, Reference Walcott1924, used in Palmer and Halley (Reference Palmer and Halley1979) is followed here. Walcott (Reference Walcott1924, Reference Walcott1925) assigned two species to Chancia: C. ebdome Walcott, Reference Walcott1924, and C. evax Walcott, Reference Walcott1925. Resser (Reference Resser1939a) added C. angusta Resser. Four new species were included by Rasetti (Reference Rasetti1951): C. latigena, C. bigranulosa, C. odarayensis, and C. stenometopa. The re-assignment to Chancia of Ptychoparia palliseri Walcott, Reference Walcott1908a, by Rasetti (Reference Rasetti1951), Kochina venusta Resser, Reference Resser1939b, by Fritz (Reference Fritz1968), and Ehmaniella maladensis Resser, Reference Resser1939b, by Palmer (in Palmer and Halley, Reference Palmer and Halley1979) brings the number of species to 10. In addition, Fritz (in Oriel and Armstrong, Reference Oriel and Armstrong1971) listed C. coriacea (Resser, Reference Resser1939a). Hu (Reference Hu1985) added C. conica Hu, Reference Hu1985.
Chancia ebdome, C. evax, C. venusta, C. bigranulosa, and C. stenometopa have a frontal area that is four-tenths of the total cranidial length (Walcott, Reference Walcott1925, pl. 17, figs. 26, 27; Resser, Reference Resser1939b, pl. 6, figs. 9, 10; Rasetti, Reference Rasetti1951, pl. 8, fig. 20, pl. 22, figs. 1–3). Chancia evax has a border that is one-fifth the length of the frontal area and 24 thoracic segments. Chancia ebdome has a border that is one-third the length of the frontal area and 20 thoracic segments. Chancia venusta, C. bigranulosa, and C. stenometopa all have a border that is slightly less than half the length of the frontal area, as well as granular or finely pustulose prosopon on the cranidium. The posterior cranidial border is slightly narrower in C. venusta and slightly wider in C. bigranulosa and C. stenometopa. In addition, C. venusta has a palpebral lobe that is one-third the glabellar length, whereas C. bigranulosa and C. stenometopa have a shorter palpebral lobe that is one-fifth the glabellar length. The similarities in cranidial dimensions between C. stenometopa and C. bigranulosa indicate that the former may be synonymous with the later.
Chancia latigena, C. palliseri, and C. angusta have a frontal area that is one-third of the total cranidial length (Walcott, Reference Walcott1908a, pl. 3, fig. 6; Resser, Reference Resser1939a, pl. 5, fig. 14; Rasetti, Reference Rasetti1951, pl. 21, fig. 15). Chancia palliseri has an anterior border that is one-third the length of the frontal area, a palpebral lobe that is one-quarter the glabellar length, and 20–23 thoracic segments (Rasetti, Reference Rasetti1951). Chancia latigena and C. angusta have a border nearly half the length of the frontal area and a palpebral lobe that is one-fifth the glabellar length. Chancia latigena has a narrower posterior border. Chancia angusta has a distinctly tapering glabella, a wider posterior border, and 18 thoracic segments (Resser, Reference Resser1939b). Chancia maladensis has a frontal area that ranges from slightly less to slightly more than one-third of the total cranidial length, as well as an anterior border that is less than half the length of the frontal area, a palpebral lobe that is one-quarter the glabellar length, an interocular area slightly wider than the glabella, and 18 thoracic segments (Resser, Reference Resser1939b, pl. 12, figs, 20, 21; Sundberg, Reference Sundberg2018, figs. 24.1–24.6, 25.1–25.12). Chancia odarayensis has a frontal area that is one-fifth of the total cranidial length, as well as a border that is nearly half the length of the frontal area, a palpebral lobe that is one-quarter the glabellar length, an interocular area that is equal in width to the glabella, and at least 22 thoracic segments (Rasetti, Reference Rasetti1951, pl. 33, fig. 16).
Chancia maladensis (Resser, Reference Resser1939)
Figure 25.1
- Reference Resser1939b
Ehmaniella maladensis Resser, p. 60, pl. 12, figs. 17–23.
- Reference Sundberg2018
Chancia maladensis; Sundberg, p. 31, figs. 24.1–24.6, 25.1–25.12.
Holotype
Cranidium (USNM 98563a) from the Spence Shale, Idaho (Resser, Reference Resser1939b, pl. 12, figs. 20, 21).
Occurrence
Mudstone, Mount Cap Formation, Dodo Canyon, Northwest Territories, Glossopleura walcotti Zone; Spence Shale, Idaho (Resser, Reference Resser1939b); Rachel Limestone and Emigrant Formation, Nevada, Glossopleura walcotti/Ptychagnostus praecurrens Zone (Sundberg, Reference Sundberg2018).
Material
Two partially complete exoskeletons lacking free cheeks and one cranidium. Figured material: GSC 142436.
Remarks
Chancia maladensis from the Mount Cap Formation resembles specimens from the Spence Shale, Emigrant Formation, and Rachel Limestone, but differs in having a slightly shorter anterior cranidial border. It shares a palpebral lobe which is slightly longer than that in other species.
Genus Eokochaspis Sundberg and McCollum, Reference Sundberg and McCollum2000
Type species
Eokochaspis nodosa Sundberg and McCollum, Reference Sundberg and McCollum2000, from the Eokochaspis nodosa Zone of the Comet Shale Member, Pioche Formation, Nevada.
Remarks
The diagnosis by Sundberg and McCollum (Reference Sundberg and McCollum2000) with amendments in Webster (Reference Webster2011a) is followed. Features used to distinguish species of Eokochaspis include the relative lengths of the preglabellar field and anterior border, presence of a plectrum, depth of axial and lateral glabellar furrows, width of the free cheek, length of the genal spine, number of pygidial axial rings, and presence of a medial indentation on the pygidial border. Sundberg (Reference Sundberg2004) suggested that Eokochaspis is polyphyletic.
Eokochaspis nodosa Sundberg and McCollum, Reference Sundberg and McCollum2000
Figure 26.8
- Reference Sundberg and McCollum2000
Eokochaspis nodosa Sundberg and McCollum, p. 611, fig. 7.1–7.20.
- Reference Sundberg and McCollum2003b
Eokochaspis nodosa; Sundberg and McCollum, p. 967, pl. 2, figs. 1, 2, 4.
- Reference Webster2011a
Eokochaspis nodosa; Webster, p. 218, figs. 7.1–7.20, 28.1–28.29, 29.1–29.31.
- Reference Webster and Zelditch2011
Eokochaspis nodosa; Webster and Zelditch, fig. 1C, D.
- Reference Pratt and Bordonaro2014
Eokochaspis nodosa; Pratt and Bordonaro, p. 916, fig. 8.1–8.10.
Holotype
Cranidium (USNM 497818) from the Eokochaspis nodosa Zone of the Comet Shale Member, Pioche Formation, Nevada (Sundberg and McCollum, Reference Sundberg and McCollum2000, fig. 7.1–7.3).
Occurrence
Silty mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, Amecephalus arrojosensis-Eokochaspis nodosa Zone; Comet Shale Member, Pioche Formation, Nevada, Eokochaspis nodosa Zone (Sundberg and McCollum, Reference Sundberg and McCollum2000; Webster, Reference Webster2011a; Webster and Zelditch, Reference Webster and Zelditch2011); Emigrant Formation, Nevada, Eokochaspis nodosa Zone (Sundberg and McCollum, Reference Sundberg and McCollum2003b); Soldano Member, La Laja Formation, San Juan, Argentina, Amecephalus arrojosensis-Eokochaspis nodosa Zone (Pratt and Bordonaro, Reference Pratt and Bordonaro2014).
Material
Two pygidia. Figured material: GSC 142428.
Remarks
These pygidia have the typical wing nut shape of species of Eokochaspis. The presence of two axial rings in each pygidium is consistent with E. nodosa as opposed to other species of Eokochaspis.
Eokochaspis species indet.
Figure 25.4, 25.7
Occurrence
Mudstone and silty mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, Aitkenaspis keelensis Zone.
Material
One nearly complete exoskeleton and three cranidia. Figured material: GSC 142438, 142441.
Remarks
Eokochaspis sp. indet. is most similar to E. piochensis (Palmer in Palmer and Halley, Reference Palmer and Halley1979), but differs by having a shorter glabella, slightly longer palpebral lobe, and less well-defined axial and lateral glabellar furrows. However, the available cranidia are small.
Genus Tonopahella Sundberg and McCollum, Reference Sundberg and McCollum2003
Type species
Tonopahella goldfieldensis Sundberg and McCollum, Reference Sundberg and McCollum2003b, from the Amecephalus arrojosensis Zone of the Emigrant Formation, Nevada.
Remarks
Sundberg and McCollum (Reference Sundberg and McCollum2003b) proposed this genus to account for two species classified in Kochina Resser, Reference Resser1935, which are somewhat similar to Amecephalus. The cranidium of Tonopahella differs from that of Amecephalus by the constriction of the axial furrow at S2, moderately defined S1 and S2, a narrower interocular area, shorter posterior limb, an absence of the interborder furrow in the anterior border. Tonopahella goldfieldensis differs from T. walcotti (Sundberg and McCollum, Reference Sundberg and McCollum2000) by having a transverse anterior border and a less well-defined border furrow.
Tonopahella goldfieldensis Sundberg and McCollum, Reference Sundberg and McCollum2003
Figure 25.6, 25.8
- Reference Sundberg and McCollum2003b
Tonopahella goldfieldensis Sundberg and McCollum, p. 981, pl. 7, figs. 1–6.
Holotype
Tonopahella goldfieldensis (USNM 517665) from the Amecephalus arrojosensis Zone of the Emigrant Formation, Nevada (Sundberg and McCollum, Reference Sundberg and McCollum2003b, pl. 7, figs. 1, 3).
Occurrence
Silty mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, Amecephalus arrojosensis-Eokochaspis nodosa Zone; Emigrant Formation, Nevada, Amecephalus arrojosensis Zone; Monola Formation, California, Amecephalus arrojosensis Zone (Sundberg and McCollum, Reference Sundberg and McCollum2003b).
Material
Three cranidia. Figured material: GSC 142440, 142442.
Remarks
Cranidia from the Mount Cap Formation differ from the holotype in having a slightly curving anterior margin. This is intermediate between T. goldfieldensis and T. walcotti, which may suggest that the two species are synonymous.
Tonopahella walcotti (Sundberg and McCollum, Reference Sundberg and McCollum2000)
Figure 26.1–26.7
- Reference Sundberg and McCollum2000
Kochina? walcotti Sundberg and McCollum, p. 626, figs. 15.1–15.6.
- Reference Sundberg and McCollum2003b
Tonopahella walcotti Sundberg and McCollum, p. 982, pl. 7, fig. 7.
Holotype
Nearly complete exoskeleton (USNM 497954) from the Amecephalus arrojosensis Zone of the Comet Shale Member, Pioche Formation, Nevada (Sundberg and McCollum, Reference Sundberg and McCollum2000, fig. 15.3).
Occurrence
Silty mudstone, Mount Cap Formation, Little Bear River, siltstone and silty mudstone, Mount Clark Formation, Dodo Canyon, Northwest Territories, Amecephalus arrojosensis-Eokochaspis nodosa and Aitkenaspis keelensis zones; Comet Shale Member, Pioche Shale, Nevada, Amecephalus arrojosensis Zone (Sundberg and McCollum, Reference Sundberg and McCollum2000); Monola Formation, California, Amecephalus arrojosensis Zone (Sundberg and McCollum, Reference Sundberg and McCollum2003b).
Material
Two nearly complete exoskeletons, 30 cranidia, three free cheeks, and one pygidium. Figured material: GSC 142421–142427.
Remarks
The cranidia from the Mount Cap Formation are similar to those from the Pioche and Monola formations (Sundberg and McCollum, Reference Sundberg and McCollum2000, Reference Sundberg and McCollum2003b). These specimens have a slightly shorter postocular fixed cheek compared with that of the holotype specimen, although the difference is considered negligible. The cranidium illustrated in Figure 26.2 differs from the other specimens by having a preserved shallow axial furrow and a more convex anterior border with a more well-defined border furrow. Because this specimen is preserved in siltstone instead of mudstone like the other cranidia, these differences are regarded as arising from differential compaction.
Tonopahella species. indet.
Figure 26.9–26.11
Occurrence
Silty mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, Amecephalus arrojosensis-Eokochaspis nodosa Zone.
Material
Thirteen cranidia. Figured material: GSC 142429–142431.
Remarks
The constriction of the glabella opposite S2, narrow interocular area, and shorter postocular fixed cheek of these specimens are consistent with Tonopahella. Tonopahella sp. indet. is distinct by having a unique combination of morphological features among species of Tonopahella. The strongly divergent anterior facial suture and a curved anterior border and border furrow of Tonopahella sp. indet. is similar to T. walcotti, whereas a longer preglabellar field that is two-thirds of the length frontal area is similar to that of T. goldfieldensis. As stated for T. walcotti, the unique combination of these features may indicate that the two species are synonymous.
Ptychoparioid genus and species indet. 1
Figure 25.11, 25.12
Occurrence
Siltstone, Mount Clark Formation, Dodo Canyon, Northwest Territories, Aitkenaspis keelensis Zone.
Material
One cranidium. Figured material: GSC 142445.
Remarks
This cranidium has a tapered glabella with two lateral glabellar furrows that curve obliquely backward, a shallow anterior border furrow, a wide fixed cheek, and an effaced ocular ridge. It bears a resemblance to Solenopleura conifrons Westergård, Reference Westergård1952, although this taxon is from Baltic and Avalonian strata of the younger Solenopleura? brachymetopa and Paradoxides forchhammeri zones (Guzhangian Stage).
Ptychoparioid genus and species indet. 2
Figure 25.5, 25.9, 25.10
Occurrence
Silty mudstone, Mount Cap Formation, Carcajou Falls, Northwest Territories, Albertelloides mischi Zone.
Description
Cranidium is subtriangular in outline. Axial furrow is well defined. Glabella is subrectangular in outline and with the axial furrow outwardly bowed slightly. Three pairs of lateral glabellar furrows are present. S1 and S2 are well defined and oriented obliquely backward and S3 is poorly defined and nearly transverse. Occipital furrow is well defined, occipital ring with a medial tubercle. Anterior border is in length one-quarter that of the preglabellar field. Border furrow is moderately defined and shallows medially. Anterior course of the facial suture is convergent. Interocular area width is three-quarters that of the glabella at the mid-point. Ocular ridge is well defined and is oriented transverse or obliquely backwards. Mid-point of the palpebral lobe opposite S2, with the anterior tip opposite L3, and the posterior tip opposite L1. Length of the palpebral lobe is nearly half that of the glabella. Posterior course of the facial suture is nearly transverse. Posterior border is slightly wider than the occipital ring, with a well-defined border furrow.
Free cheek is equal in width to the occipital ring, with a border that comprises one-fifth of the width. Genal spine is slightly advanced and comprises one-third of the length of the free cheek.
Hypostome, thorax, and pygidium are unknown.
Material
One cephalon and 16 cranidia. Figured material: GSC 142439, 142443, and 142444.
Remarks
This species is distinct among ptychopariids in that it has a unique combination of a short anterior cranidial border, a convergent anterior facial suture, a long palpebral lobe, and a wide fixed cheek. Typically, genera with convergent facial sutures, such as Plagiura Resser, Reference Resser1935, and Onchocephalus Resser, Reference Resser1937, have a short palpebral lobe located anteriorly and a narrow fixed cheek. The lack of any thoracic and pygidial material of this taxon disfavors the erection of a new genus and species.
Ptychoparioid genus and species indet. 3
Figure 26.16, 26.17
Occurrence
Mudstone, Mount Cap Formation, Carcajou Falls, Northwest Territories, Glossopleura walcotti Zone.
Description
Exoskeleton is subelliptical in outline. Exoskeleton length 20 mm.
Cranidium is subtrapezoidal in outline. Axial furrow is moderately defined. Glabella is subtrapezoidal in outline. Three pairs of lateral glabellar furrows are present. S1 and S2 are poorly defined and oriented obliquely backward, and S3 is nearly effaced and transverse. Occipital furrow is moderately defined. Anterior border is slightly longer than the preglabellar field. Border furrow is moderately defined, shallowing at a poorly defined preglabellar swelling that extends into the border. Anterior course of the facial suture is convergent. Interocular area is equal in width to the occipital ring. Ocular ridge is well defined and nearly transverse; slightly bowed anteriorly. Mid-point of palpebral lobe is opposite S2, with the anterior tip opposite L3, and posterior tip opposite L2. Length of the palpebral lobe is one-quarter that of the glabella. Posterior course of the facial suture is oriented obliquely backwards. Posterior border is in width twice that of the occipital ring, with a well-defined border furrow.
Free cheek and hypostome are unknown.
Thorax consists of 18 segments. Axial furrow is moderately defined. Pleura width is one-and-one-third times that of the axis, with well-defined pleural and interpleural furrows. Pleural spines blunt.
Pygidium micropygous, elliptical in outline, and the length is equal to one-third the width. Axis tapers gently posteriorly, with the posterior tip slightly anterior to the posterior edge of the pygidium, and is composed of two axial rings and a terminal piece. Pleural field is in width twice that of the axis, with two pairs of poorly defined pleural furrows that intersect the margin of the pygidium. Border and border spines are absent.
Material
One complete exoskeleton lacking free cheeks. Figured material: GSC 142435.
Remarks
The cranidium of this specimen bears a resemblance to that of species of Amecephalus, such as A. piochensis (Walcott, Reference Walcott1912) and A. agnesensis (Walcott, Reference Walcott1912), although differs by having a parallel as opposed to divergent anterior facial suture. Furthermore, the pygidium of this specimen is wider in proportion to its length, has well-defined pleural furrows, and lacks a border.
Ptychoparioid genus and species indet. 4
Figure 26.12
Occurrence
Silty mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, Amecephalus arrojosensis-Eokochaspis nodosa Zone.
Material
One nearly complete exoskeleton. Figured material: GSC 142432.
Remarks
The shallow border furrow, longitudinal furrow, lateral glabellar furrows, and thoracic pleural furrows as well as the weakly defined ocular ridge of this specimen are distinct. The morphology of the cranidium is similar to that of Eokochaspis nodosa (Sundberg and McCollum, Reference Sundberg and McCollum2000, fig. 7.1–7.4, 7.6, 7.11), but differs by having a shallow ocular ridge, absence of a glabellar constriction at S2, and narrower genal spine.
Ptychoparioid genus and species indet. 5
Figure 26.13
Occurrence
Silty mudstone, Mount Cap Formation, Little Bear River, Northwest Territories, Amecephalus arrojosensis-Eokochaspis nodosa Zone.
Material
One cranidium. Figured material: GSC 142433.
Remarks
The features of this cranidium are general for ptychoparioids of the middle Cambrian. Distinguishing features of Ptychoparioid gen. and sp. indet. 5 are the slightly convergent axial furrow, short border, and well-defined ocular ridge.
Acknowledgments
This project is part of the Geological Survey of Canada's Geo-mapping for Energy and Minerals (GEM) and Geoscience for New Energy Supply (GNES) programs, which provided bursary support for Handkamer through the Research Affiliate Program of Natural Resources Canada. Funding was also provided by Natural Sciences and Engineering Research Council of Canada (Discovery Grant awarded to Pratt) and Polar Knowledge Canada (Northern Scientific Training Program support awarded to Handkamer). Fieldwork was conducted in the Sahtu Land Claim Settlement Region under a permit issued by the Aurora Research Institute. Helicopter transportation during fieldwork was reliably provided by Sahtu Helicopters. We thank I. Pidskalny for assistance and insight with fieldwork in 2019, as well as S.A. Gouwy, F.A. Sundberg, and an anonymous reviewer for comments on the manuscript. NRCan contribution number 20210288.



























 Open access
Open access