


Introduction
Italian ryegrass [Lolium perenne L. ssp. multiflorum (Lam.) Husnot], a plant indigenous to the Mediterranean region, is widely utilized as a forage, sod, and cover crop (Kemešytė et al. Reference Kemešytė, Statkevičiūtė, Norkevičienė and Jaškūnė2023; Worku et al. Reference Worku, Tóthi, Orosz, Fébel, Kacsala, Vermeire and Tóth2021). In recent decades, it has become a widespread weed, colonizing roadsides and agricultural lands worldwide. Introduced to China in the 1980s, L. perenne ssp. multiflorum has since evolved into an aggressive weed that competes with wheat (Triticum aestivum L.) for water and nutrients, particularly in provinces such as Jiangsu, Henan, and Anhui (Wu et al. Reference Wu, Song, Zhang, Zhou, Liu, Jin and Zhao2022; Xu et al. Reference Xu, Cheng, Leng, Cao, Su, Sun, Xue, Han and Wu2025; Zhang et al. Reference Zhang, Wu, Xu, Gao, Zhang and Dong2017; Zhu et al. Reference Zhu, Wang, Gao, Liu, Li, Feng and Dong2023). The herbicides commonly used for controlling L. perenne ssp. multiflorum currently include diclofop, pinoxaden, mesosulfuron-methyl, and pyroxsulam (Bararpour et al. Reference Bararpour, Korres, Burgos, Hale and Tseng2018; Jones et al. Reference Jones, Taylor and Everman2021). However, long-term herbicide application has led to the evolution of resistance to multiple herbicides in L. perenne ssp. multiflorum, making it one of the most difficult weeds to control in wheat fields (Nakka et al. Reference Nakka, Jugulam, Peterson and Asif2019). The first case of herbicide resistance in L. perenne ssp. multiflorum was reported in 1987 to the acetyl-CoA carboxylase (ACCase) inhibitor diclofop-methyl in Oregon, USA (Stanger and Appleby Reference Stanger and Appleby1989). Since then, resistance to herbicides in L. perenne ssp. multiflorum has been documented in China, the United Kingdom, France, Chile, and other countries (Heap Reference Heap2025).
Isoproturon, a photosystem II (PSII)-inhibiting herbicide, disrupts photosynthetic electron transport by competitively binding to the D1 protein at the plastoquinone B site, thereby blocking plastoquinone binding (Gardner Reference Gardner1989; Zharmukhamedov and Allakhverdiev Reference Zharmukhamedov and Allakhverdiev2021). This disruption blocks electron transport and suppresses the synthesis of ATP and NADPH+, as well as the Calvin cycle. As a result, normal photosynthetic processes in weeds are inhibited, leading to weed death (Fuerst and Norman Reference Fuerst and Norman1991; Hess Reference Hess2000). However, long-term reliance on a single herbicide can lead to the development of weed resistance. As early as the 1990s, resistance to isoproturon was documented in L. perenne ssp. multiflorum in the United Kingdom and in littleseed canarygrass (Phalaris minor Retz.) in India (Heap Reference Heap2025).
Resistance to herbicides is generally categorized into two main types based on the underlying mechanism: target-site resistance (TSR) and non–target site resistance (NTSR). TSR involves mutations in the genes encoding herbicide target proteins and/or the overexpression of these genes or proteins (Délye et al. Reference Délye, Jasieniuk and Le Corre2013). The psbA gene encodes the D1 protein, the target of isoproturon. To date, eight amino acid substitutions in the psbA gene have been reported to confer resistance to PSII inhibitors in weeds: Leu-218-Val, Val-219-Ile, Ala-251-Val, Phe-255-Ile, Ser-264-Gly, Ser-264-Thr, Asn-266-Thr, and Phe-274-Val (Alfonso et al. Reference Alfonso, Pueyo, Gaddour, Etienne, Kirilovsky and Picorel1996; Bettini et al. Reference Bettini, McNally, Sevignac, Darmency, Gasquez and Dron1987; Johanningmeier et al. Reference Johanningmeier, Bodner and Wildner1987; Lu et al. Reference Lu, Yu, Han, Owen and Powles2018; Masabni and Zandstra Reference Masabni and Zandstra1999; Mengistu et al. Reference Mengistu, Mueller-Warrant, Liston and Barker2000; Ohad and Hirschberg Reference Ohad and Hirschberg1992). In addition to target-site mutations, psbA gene overexpression has also been linked to enhanced resistance. For instance, Yang et al. (Reference Yang, Yu, Cui, Chen and Li2022) demonstrated that overexpression of psbA confers atrazine resistance in Asiatic dayflower (Commelina communis L.). In contrast, NTSR mainly involves enhanced herbicide metabolism and altered absorption and translocation patterns (Gaines et al. Reference Gaines, Duke, Morran, Rigon, Tranel, Kuepper and Dayan2020). Among these mechanisms, elevated metabolic detoxification is the most prevalent, often mediated by key enzymes such as cytochrome P450 monooxygenases, glutathione S-transferases, ABC transporters, and glycosyltransferases (Powles and Yu, Reference Powles and Yu2010 ; Rigon et al. Reference Rigon, Gaines, Küpper and Dayan2020).
While herbicide resistance in L. perenne ssp. multiflorum has become an increasingly serious issue throughout China (Wu et al. Reference Wu, Song, Zhang, Zhou, Liu, Jin and Zhao2022; Xu et al. Reference Xu, Cheng, Leng, Cao, Su, Sun, Xue, Han and Wu2025; Zhang et al. Reference Zhang, Wu, Xu, Gao, Zhang and Dong2017; Zhu et al. Reference Zhu, Wang, Gao, Liu, Li, Feng and Dong2023), the extent and distribution of resistance specifically to isoproturon remain largely unclear. In this study, we measured the isoproturon sensitivity of 87 L. perenne ssp. multiflorum populations collected from Jiangsu and Henan, China, and a highly resistant (HR) population was identified. Therefore, the objectives of this study were to: (1) determine the resistance level, (2) quantify the expression of the psbA gene, (3) measure the activities of antioxidant enzymes and the contents of chlorophyll and carotenoids, and (4) determine cross-resistance and multiple resistance.
Materials and Methods
Plant Materials and Herbicides
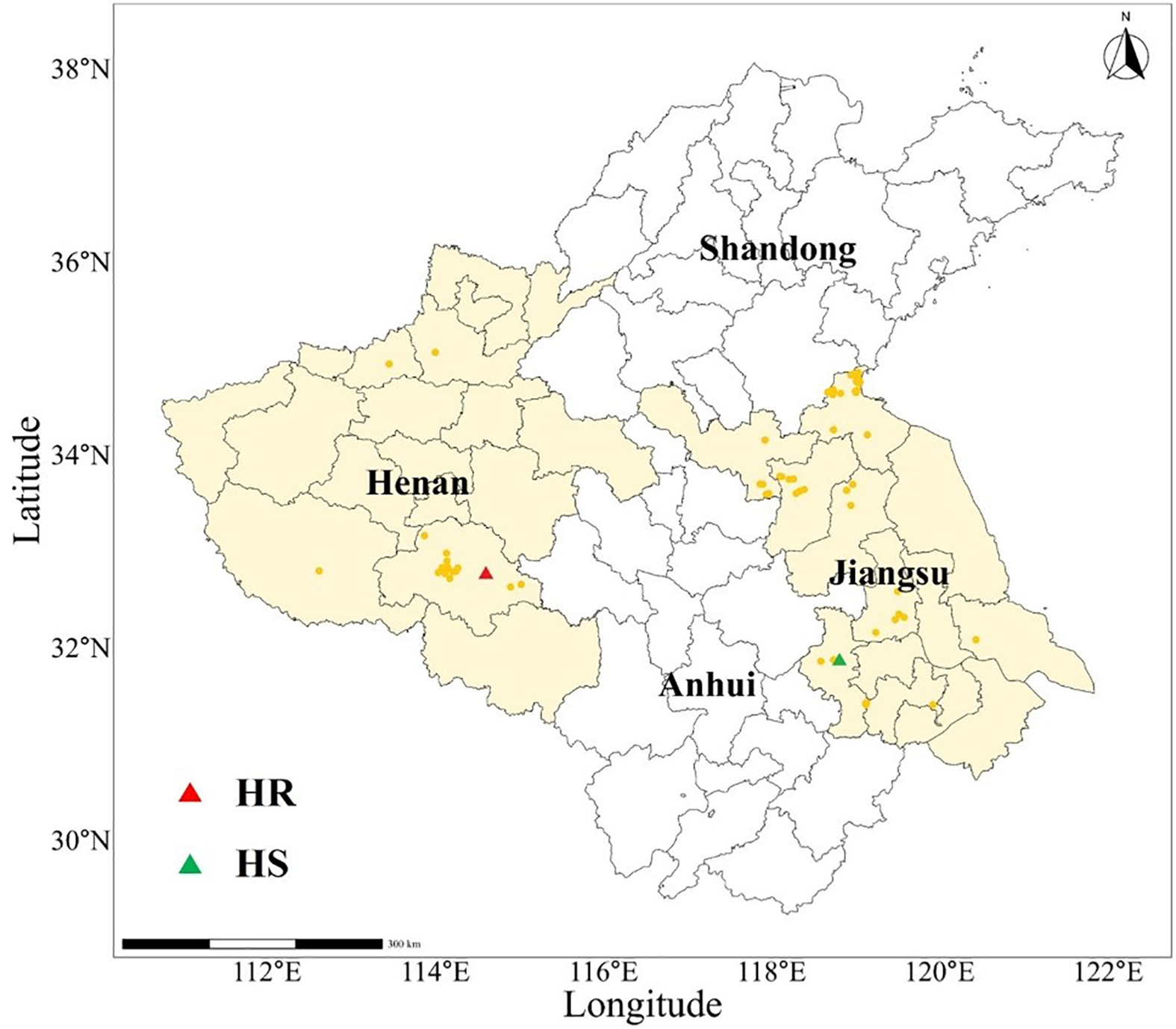
Between 2014 and 2023, we collected L. perenne ssp. multiflorum seeds from winter wheat fields across Henan (24 populations) and Jiangsu (63 populations) provinces, China (Figure 1; Supplementary Tables S1 and S2). All seeds were manually selected, air-dried in the shade, and then stored at 4 C to preserve seed viability.
The geographic distribution of 87 Lolium perenne ssp. multiflorum populations in Henan and Jiangsu provinces. The orange dots indicate the collection sites of the L. perenne ssp. multiflorum populations, the red triangle represents the resistant population (HR), and the green triangle represents the sensitive population (HS).

Isoproturon (50% WP) and chlorotoluron (20% EC) were purchased from Jiangsu Kuaida Agrochemical Company (Jiangsu, China). Pyroxsulam (4% WG) was provided by Corteva Agriscience (Shanghai, China). Pinoxaden (5% EC) was supplied by Syngenta Group China (Shanghai, China).
Plant Single-Dose Assay
Seeds from each of the 87 collected populations (30 seeds per population) were sown in plastic pots (7 by 7 by 7 cm) containing a 2:1 mixture of sandy soil and nutrient matrix (organic matter: 1.1%; pH: 6.2), then lightly covered with the same medium. We then watered the pots until the soil had absorbed enough water. Subsequently, the pots were treated with isoproturon at the field-recommended dose (1,125 g ai ha−1) using a laboratory crawler sprayer (3WP-2000) equipped with a flat-fan nozzle, delivering 280 L ha−1 at 230 kPa. The pots were maintained under moist conditions in a solar greenhouse set at 20/15 C (day/night) with a 12-h photoperiod, and approximately 65% relative humidity. At 21 d after treatment, survival was assessed: deceased plants were classified as susceptible, while survivors were considered resistant. The aboveground biomass of surviving plants was harvested and weighed to calculate the fresh weight inhibition rate.
Whole-Plant Dose–Response Assay
Leveraging data from the single-dose test, we performed whole-plant dose–response assays on suspected sensitive and resistant populations of L. perenne ssp. multiflorum. Plastic pots were filled with the soil described earlier and sown with 20 seeds uniformly. Four replicate pots were used per treatment. The plants were grown in greenhouses under the cultivation conditions described earlier. Seedlings were thinned to 10 per pot at the 2- to 3-leaf stage. At the 3- to 4-leaf stage, herbicides were applied using a laboratory track sprayer (3WP-2000). Isoproturon was applied at the rates of 0, 281.25, 562.5, 1,125, 2,250, and 4,500 g ha−1 for the suspected resistant populations and at 0, 70.31, 140.63, 281.25, 562.5 and 1,125 g ha−1 for the suspected sensitive populations.
Pyroxsulam and pinoxaden were selected for determining multiple resistance of the isoproturon-resistant population of L. perenne ssp. multiflorum. For pyroxsulam, the doses applied to the isoproturon-resistant population were 0, 7.5, 15, 30, 60, and 120 g ha−1, while doses for the isoproturon-sensitive population were 0, 0.12, 0.23, 0.47, 0.94, 1.88 and 3.75 g ha−1. For pinoxaden, the doses were 0, 15, 30, 60, 120, and 240 g ha−1 for the isoproturon-resistant population and 0, 7.5, 15, 30, 60, and 120 g ha−1 for the isoproturon-sensitive population. Aboveground biomass from each pot was harvested 21 d after treatment, and the fresh weight was measured to calculate the inhibition rate.
psbA Gene Cloning and Sequencing
Genomic DNA was extracted from young shoot tissues (0.1 g per sample) of the HZPY-2023-1(HR) and JNXW-2020-2(HS) populations collected at the 3- to 4-leaf stage using a Plant Genomic DNA Kit (Tiangen Biotech, Beijing, China), following the manufacturer’s instructions, with a total of 20 individual plants sampled per population. The full-length psbA gene was amplified and sequenced as described by Zhu et al. (Reference Zhu, Wang, Gao, Liu, Li, Feng and Dong2023). A polymerase chain reaction (PCR) was performed in a 25-μl reaction mixture containing 300 ng of DNA template, 1 μl of each primer (10 μM), 12.5 μl of 2× Phanta Max Master Mix (Vazyme, Nanjing, China), 0.5 μl of dNTPs, 0.5 μl of Phanta Max super-fidelity DNA polymerase, and ddH2O to a final volume of 25 μl. PCR products were purified and sequenced using the Sanger method.
psbA Gene Expression and psbA Gene Promoter Sequencing Analysis
HS and HR populations were grown in a light incubator under controlled conditions (20/15 C, 12/12 h, light intensity 120 μmol m−2 s−1, 65% humidity) until they reached the 3- to 4-leaf stage. The plants were then treated with isoproturon at 1,125 g ha−1, while a water-treated group served as the control. Aboveground tissues (0.1 g) were collected at 6 h, 12 h, 24 h, 3 d, and 5 d after treatment, immediately frozen in liquid nitrogen, and stored at −80 C for subsequent RNA extraction. Each time point included three biological replicates. Total RNA was extracted from 0.1 g of fresh leaf tissue using the RNA-simple Isolation Reagent (Pudi Biotech, Shanghai, China) according to the manufacturer’s protocol. RNA concentration and quality were assessed using an ultra-micro spectrophotometer (ND-100C, MIULAB, Hangzhou, China). First-strand cDNA was synthesized from 1,000 ng of total RNA using the HiScript IV All-in-One Ultra RT Super Mix (Vazyme, Nanjing, China); genomic DNA was eliminated via DNase treatment. Quantitative real-time PCR (RT-qPCR) was performed on a Quant Studio 1 system (Thermo Fisher Scientific, Waltham, MA, USA) using ChamQ SYBR qPCR Master Mix (Vazyme). In accordance with the methodology proposed by Gaines et al. (Reference Gaines, Lorentz, Figge, Herrmann, Maiwald, Ott, Han, Busi, Yu, Powles and Beffa2014), the Ras family GTPase (RGTP) gene was selected as the internal reference gene, while the amplification primer for the psbA gene was derived from the work of Zhu et al. (Reference Zhu, Wang, Gao, Liu, Li, Feng and Dong2023; Supplementary Table S3). The 2−ΔΔCt method was used to calculate the relative expression levels of the psbA gene in the HS and HR populations. The entire experiment was performed twice, each with three biological replicates. The promoter region of the psbA gene was separately amplified in both the HR and HS populations. The PCR reactions were the same as those for amplifying the psbA gene. The PCR products were purified, cloned, and sequenced using the Sanger method.
Antioxidant Enzyme Activity Content Assay
As mentioned earlier, the HS and HR populations were grown to the 3- to 4-leaf stage, after which isoproturon was applied at a rate of 1,125 g ha−1. The aboveground tissues were collected at 0 h (without isoproturon treatment), 12 h, 1 d, 3 d, and 5 d after treatment, with three replicates per treatment. The collected samples were immediately frozen in liquid nitrogen and stored at −80 C. Each treatment was ground in a mortar and then homogenized with 1 ml of 0.05 M sodium phosphate buffer (pH 7.8). The homogenate was centrifuged at 12,000 rpm for 20 min at 4 C, and the resulting supernatant was collected as the crude enzyme extract for the determination of superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT) activities. The specific assay methods for SOD, POD, and CAT activities followed those described by Guan et al. (Reference Guan, Chen, Qiu, Qian, Chen, Shao, Xie, Deng and Peng2020).
Chlorophyll and Carotenoid Determination
As mentioned earlier, the HS and HR populations were cultured to the 3- to 4-leaf stage for isoproturon treatment. Aboveground tissues (0.1 g) were collected at 0 h (without treatment), 1 d, 3 d, 5 d, and 7 d after treatment, with three replicates for each time point. The chlorophyll and carotenoid contents were determined according to the methods described by Lichtenthaler (Reference Lichtenthaler1987) and Liu et al. (Reference Liu, Gao, Wang, Liang, Yu, Feng and Dong2024). For each sample, 0.1 g of fresh leaves was homogenized in a 15-ml centrifuge tube containing 10 ml of acetone. The centrifugal tubes were covered with aluminum foil and kept at room temperature in a dark place for 3 to 4 d. After incubation, the supernatant was filtered with 0.22-μm filter membrane in order to be tested. A spectrophotometer (TECAN, Infinite 200 Pro, Männedorf, Switzerland) was used to measure the absorbance of the solution at 663 nm, 645 nm, and 470 nm, with 80% acetone as a blank. Chlorophyll a, chlorophyll b, total chlorophyll, and carotenoid content were expressed as fresh weight (mg g−1); the relationship is as follows:
 $$\eqalign{ & {\rm{Chlorophyll}}\,a{\mkern 1mu} \left( {{\rm{mg}}\;{{\rm{g}}^{ - 1}}} \right) = 12.71 \times {\rm{O}}{{\rm{D}}_{663}} - 2.59{\rm{O}}{{\rm{D}}_{645}} \cr
& \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \times V/1,000W \cr} $$
$$\eqalign{ & {\rm{Chlorophyll}}\,a{\mkern 1mu} \left( {{\rm{mg}}\;{{\rm{g}}^{ - 1}}} \right) = 12.71 \times {\rm{O}}{{\rm{D}}_{663}} - 2.59{\rm{O}}{{\rm{D}}_{645}} \cr
& \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \times V/1,000W \cr} $$
 $$\eqalign{ & {\rm{Chlorophyll }}\,b{\mkern 1mu} \left( {{\rm{mg}}\;{{\rm{g}}^{{\rm{ - 1}}}}} \right) = 22.88 \times {\rm{O}}{{\rm{D}}_{645}} - 4.67 \times {\rm{O}}{{\rm{D}}_{663}} \cr
& \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \times V/1,000W \cr} $$
$$\eqalign{ & {\rm{Chlorophyll }}\,b{\mkern 1mu} \left( {{\rm{mg}}\;{{\rm{g}}^{{\rm{ - 1}}}}} \right) = 22.88 \times {\rm{O}}{{\rm{D}}_{645}} - 4.67 \times {\rm{O}}{{\rm{D}}_{663}} \cr
& \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \times V/1,000W \cr} $$
 $$\eqalign{ & {\rm{Total}}\;{\rm{chlorophyll}}\;\left( {{\rm{mg}}\;{{\rm{g}}^{ - 1}}} \right) = \left( {8.04 \times {\rm{O}}{{\rm{D}}_{663}} + 20.29 \times {\rm{O}}{{\rm{D}}_{645}}} \right) \cr & \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \times V/1,000W \cr} $$
$$\eqalign{ & {\rm{Total}}\;{\rm{chlorophyll}}\;\left( {{\rm{mg}}\;{{\rm{g}}^{ - 1}}} \right) = \left( {8.04 \times {\rm{O}}{{\rm{D}}_{663}} + 20.29 \times {\rm{O}}{{\rm{D}}_{645}}} \right) \cr & \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \times V/1,000W \cr} $$
 $$\eqalign{ & {\rm{Carotenoid}}\left( {{\rm{mg}}\;{{\rm{g}}^{ - 1}}} \right) = \left( {1,000 \times {\rm{O}}{{\rm{D}}_{470}} - 3.27X - 104Y} \right) \cr
& \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \times V/\left( {229,000 \times W} \right) \cr} $$
$$\eqalign{ & {\rm{Carotenoid}}\left( {{\rm{mg}}\;{{\rm{g}}^{ - 1}}} \right) = \left( {1,000 \times {\rm{O}}{{\rm{D}}_{470}} - 3.27X - 104Y} \right) \cr
& \quad \quad \quad \quad \quad \quad \quad \quad \quad \quad \times V/\left( {229,000 \times W} \right) \cr} $$
where V is the volume of supernatant, W is the fresh weight (g), X is the concentration of chlorophyll a, and Y is the concentration of chlorophyll b.
LmpsbA Transgenic Oryza sativa and Herbicide Sensitivity
The LmpsbA was cloned into the WMV050-HRG-ubi+3xHA vector (provided by Weimi Biotechnology, Hainan, China). The successfully constructed plasmid was used for genetic transformation of japonica rice (Oryza sativa L.), which was performed by Weimi Biotechnology. Overexpression of the LmpsbA gene in transgenic O. sativa lines (T1 line) was confirmed by Western blot analysis. The relative expression level of the LmpsbA gene in the transgenic O. sativa lines (T2 line) was determined by qPCR, using OsActin2 as the internal reference gene (Yang et al. Reference Yang, Nian, Xie, Feng, Zhang, Jing, Zhang, Dong, Peng, Wang, Qian and Zuo2016). For the homozygous transgenic lines (T2 line), herbicide sensitivity tests were conducted. Herbicide treatments were applied at the 3- to 4-leaf stage of O. sativa plants. The application rates of isoproturon were set at 0, 3,750, 7,500, 15,000, and 30,000 g ha−1, while those of chlorotoluron were 0, 1,687.5, 6,750, 13,500, and 27,000 g ha−1. Each treatment included three replicate pots. At 21 d after herbicide application, the aboveground fresh weight of the plants was measured, and the fresh weight inhibition rate was calculated.
Statistical Analysis
All experimental data were separated using Duncan’s multiple range test (P < 0.05) and SPSS 21.0 software (IBM, Chicago, IL, USA). The results showed that there was no significant difference in plant response between the two repetitions (t test, P > 0.05). SigmaPlot v. 15.0 (Systat Software, Chicago, IL, USA) was used to summarize and fit whole-plant dose–response experimental data to a four-parameter nonlinear logistic regression model to calculate herbicide concentrations that lead to 50% growth reduction (GR50):
 $$y = c + \left( {d - c} \right)/\left[ {1 + \left( {{x \over {{g^b}}}} \right)} \right]$$
$$y = c + \left( {d - c} \right)/\left[ {1 + \left( {{x \over {{g^b}}}} \right)} \right]$$
where x is the applied dose of herbicide, y is the percentage of fresh weight of the living aboveground part in the fresh weight of the control aboveground part, c is the lower limit, d is the upper limit, g is the applied dose of herbicide at the intermediate inflection point between the upper limit and the lower limit, and b is the slope of the curve. The resistance index (RI) was calculated by dividing the GR50 of the HR population by the GR50 of the HS population.
Results and Discussion
Sensitivity of Lolium perenne ssp. multiflorum Populations to Isoproturon
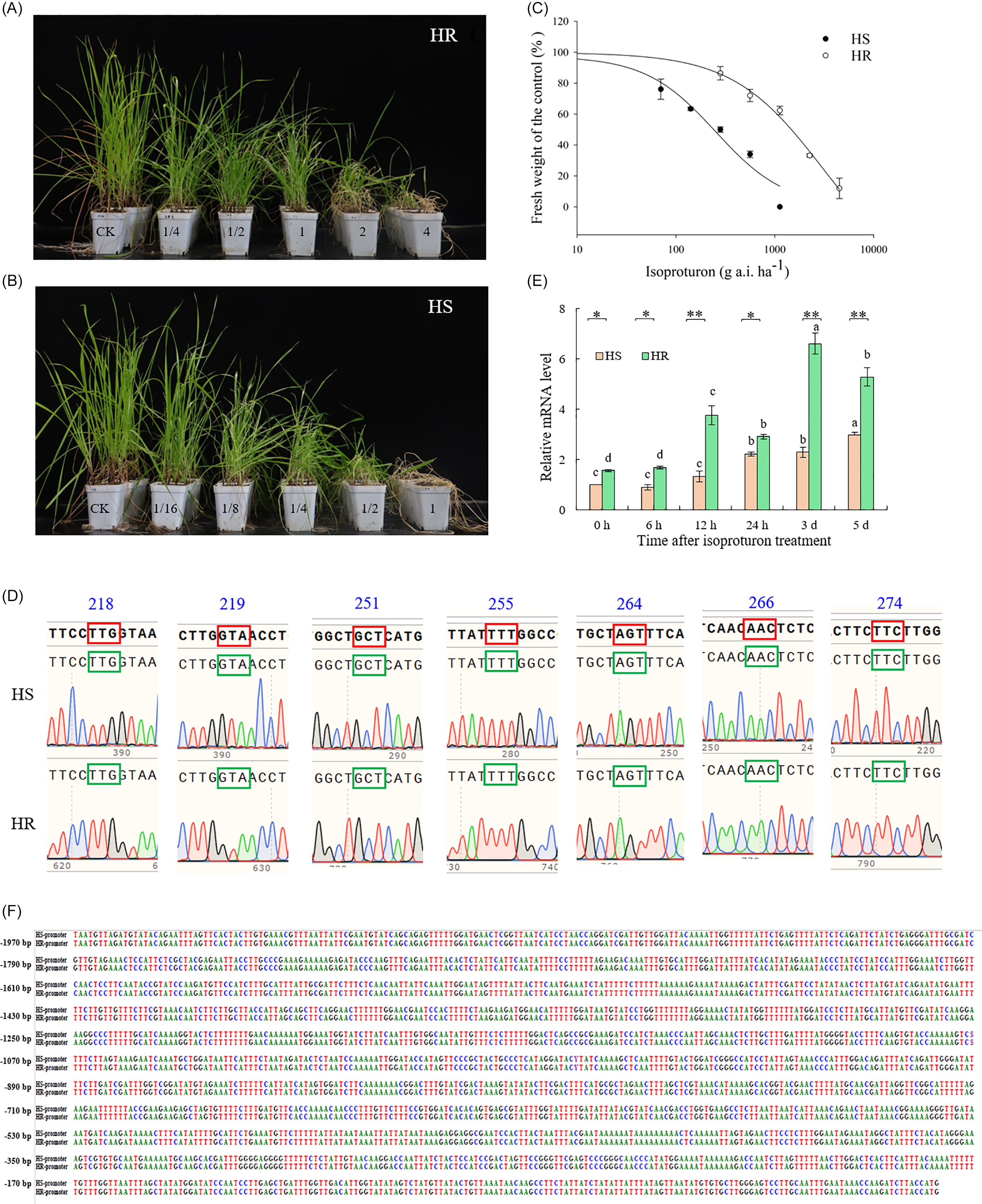
A single-dose assay was conducted to evaluate the sensitivity to isoproturon of 24 L. perenne ssp. multiflorum populations from Henan and 63 from Jiangsu. As shown in Supplementary Tables S1 and S2, under the treatment of isoproturon at the dose of 1,125 g ha−1 (1× field recommended dose), the fresh weight inhibition rates of 6 populations from Henan and 13 populations from Jiangsu were above 95%. We classified these populations as suspected sensitive populations. At the same dose, 4 populations from Henan and 17 from Jiangsu showed fresh weight inhibition rates below 70%. The resistance levels to isoproturon of the suspected sensitive and resistant populations were further determined by whole-plant dose–response assay, and the population with the highest resistance level (HZPY-2023-1, HR) and the population with the lowest resistance level (JNXW-2020-2, HS) to isoproturon were identified for further study (data not shown). The GR50 values of the HR and HS populations were 1,032.5 g ha−1 and 223.5 g ha−1, respectively (Figure 2A–C), and the RI value of the HR population was 4.6. This finding demonstrates that the HR population from Henan province has evolved resistance to isoproturon.
Whole-plant sensitivity bioassays and target-site mechanism of Lolium perenne ssp. multiflorum from China in response to isoproturon. Isoproturon sensitivity of resistant population (HR) (A) and sensitive population (HS). (B) The numbers represent multiples of the recommended field dose. (C) Isoproturon dose–response curves of HS and HR populations. (D) Nucleotide sequence alignment of psbA gene fragments originating from HS and HR populations. (E) Relative mRNA level of psbA gene in HS and HR populations after isoproturon treatment. (F) Sequence alignment of the psbA gene promoter between HS and HR populations. BioEdit 7.7.1 (Borland, Scotts Valley, CA, USA) was used. Lowercase letters indicate comparisons between different time points within the same population; one-way ANOVA was used to analyze the significance of the data: *P < 0.05; ** P < 0.01; a t test was used to analyze the significance between the HS and HR populations. Data are derived from at least three biological replicates.

Lolium perenne ssp. multiflorum has emerged as a predominant malignant weed in wheat fields across China, with its spread continuing to pose significant challenges to crop production, being especially prevalent in Jiangsu, Henan, and Anhui provinces (Xu et al. Reference Xu, Cheng, Leng, Cao, Su, Sun, Xue, Han and Wu2025; Yin et al. Reference Yin, Zhu, Li, Dong and Feng2025; Zhang et al. Reference Zhang, Wu, Xu, Gao, Zhang and Dong2017; Zhu et al. Reference Zhu, Wang, Gao, Liu, Li, Feng and Dong2023). Intensive herbicide use has led to the rapid evolution of resistance in L. perenne ssp. multiflorum, particularly to ACCase and acetolactate synthase (ALS) inhibitors. Although isoproturon has been widely applied for years in China, resistance to this herbicide had not previously been documented. This study reports for the first time the emergence of isoproturon resistance in an L. perenne ssp. multiflorum population from wheat fields. Several studies have documented L. perenne ssp. multiflorum resistance to other herbicide classes. Zhang et al. (Reference Zhang, Wu, Xu, Gao, Zhang and Dong2017) discovered that a population of L. perenne ssp. multiflorum from Henan Province developed severe resistance to fenoxaprop-P-ethyl, haloxyfop-R-methyl, quizalofop-P-ethyl, clodinafop-propargyl, sethoxydim, clethodim, and pinoxaden. Li et al. (Reference Li, Wu, Wang, Jiang, Zhang, Liao, Cao and Zhao2022) reported resistance to the ACCase herbicide clodinafop-propargyl and the ALS herbicide mesosulfuron-methyl, but the population in that study remained susceptible to isoproturon. More recently, Zhu et al. (Reference Zhu, Wang, Gao, Liu, Li, Feng and Dong2023) identified an L. perenne ssp. multiflorum population with resistance to ALS and ACCase herbicides; notably, the isoproturon GR50 value for this population was 307.21 g ha⁻¹, far below the recommended field rate, confirming that the standard dose application will remain effective for control. In contrast, the HR population in the current study was able to survive the recommended field rate of isoproturon, indicating a practical and biologically significant level of resistance. This finding highlights a new and concerning shift in the resistance status of L. perenne ssp. multiflorum in China.
Target Gene Analysis
Sequencing of the psbA Gene
The psbA gene (1,062 bp) was amplified from HS and HR populations of L. perenne ssp. multiflorum using two primer pairs, followed by cloning and sequencing. The obtained psbA sequences were compared through BLAST analysis on NCBI, revealing 99% similarity with the previously reported psbA gene of L. perenne ssp. multiflorum (GenBank accession no. NC_019651), thus confirming the accuracy of the amplified sequence. Sequence alignment of the psbA gene from 20 HS and 20 HR individuals showed complete identity between the two groups, with no mutations detected (Figure 2D). These results suggest that the resistance mechanism in the HR population is not associated with target-site mutation.
TSR to PSII inhibitors in weeds is commonly associated with specific amino acid substitutions in the psbA gene, such as Leu-218-Val, Val-219-Ile, Ala-251-Val, Phe-255-Ile, Ser-264-Gly, Ser-264-Thr, Asn-266-Thr, and Phe-274-Val (Alfonso et al. Reference Alfonso, Pueyo, Gaddour, Etienne, Kirilovsky and Picorel1996; Bettini et al. Reference Bettini, McNally, Sevignac, Darmency, Gasquez and Dron1987; Johanningmeier et al. Reference Johanningmeier, Bodner and Wildner1987; Lu et al. Reference Lu, Yu, Han, Owen and Powles2018; Masabni and Zandstra Reference Masabni and Zandstra1999; Mengistu et al. Reference Mengistu, Mueller-Warrant, Liston and Barker2000; Ohad and Hirschberg Reference Ohad and Hirschberg1992). In this study, there was no difference in psbA gene sequence between HS and HR populations. Therefore, this indicates that the isoproturon resistance of the HR population in this study was not caused by psbA gene mutations.
psbA Gene Expression and Promoter Analysis
The expression levels of the psbA gene in HS and HR populations were determined by qPCR at 0 h, 6 h, 12 h, 24 h, 3 d, and 5 d after isoproturon treatment. As shown in Figure 2E, the relative expression of the psbA gene was upregulated in both populations following herbicide application. In the HS population, expression increased gradually over time, reaching a maximum of 2.97-fold compared with the 0-h control by day 5, although no significant change was observed within the first 12 h. In contrast, the HR population reached peak expression on day 3, at a level 6.61-fold higher than that of the 0-h HS control. The basal expression level (0 h) of the psbA gene in the HR population was 1.56-fold higher than that in the HS population. Throughout the time course, the relative expression of the psbA gene in the HR population remained significantly higher than in the HS population, with the most pronounced difference observed on day 3, when the HR expression level was 2.88-fold greater than that of HS. Therefore, the resistance observed in the HR population may be attributed to psbA overexpression. We sequenced and compared the psbA promoter regions of the HS and HR populations. Sequencing results revealed 100% sequence identity between the two populations, with no mutations detected in this region (Figure 2F). These findings indicate that the overexpression of the psbA gene in the HR population is not driven by promoter region mutations.
Target gene overexpression has been recognized as an important mechanism conferring herbicide resistance in weeds. For instance, Baerson et al. (Reference Baerson, Rodriguez, Biest, Tran, You, Kreuger, Dill, Pratley and Gruys2002) reported a 2.5- to 3-fold increase in EPSPS expression in glyphosate-resistant rigid ryegrass (Lolium rigidum Gaudin), although it remained unclear whether this level of overexpression fully accounted for the resistance phenotype. Similar cases have been frequently documented with glyphosate, where enhanced expression of the target gene contributes to resistance (Chen et al. Reference Chen, Huang, Zhang, Wei, Huang, Chen and Wang2015; Malone et al. Reference Malone, Morran, Shirley, Boutsalis and Preston2016; Salas et al. Reference Salas, Dayan, Pan, Watson, Dickson, Scott and Burgos2012). Likewise, Laforest et al. (Reference Laforest, Soufiane, Simard, Obeid, Page and Nurse2017) found that ACCase overexpression conferred resistance to five distinct ACCase-inhibiting herbicides in large crabgrass [Digitaria sanguinalis (L.) Scop.]. More recently, Yang et al. (Reference Yang, Yu, Cui, Chen and Li2022) reported elevated psbA expression in an atrazine-resistant C. communis population, suggesting that target gene overexpression contributes to resistance to PSII inhibitors. In the present study, the psbA gene was expressed at a significantly higher level in the HR population than in the HS population. Therefore, psbA overexpression may account for the induced resistance of L. perenne ssp. multiflorum to isoproturon.
Antioxidant Enzyme Activity
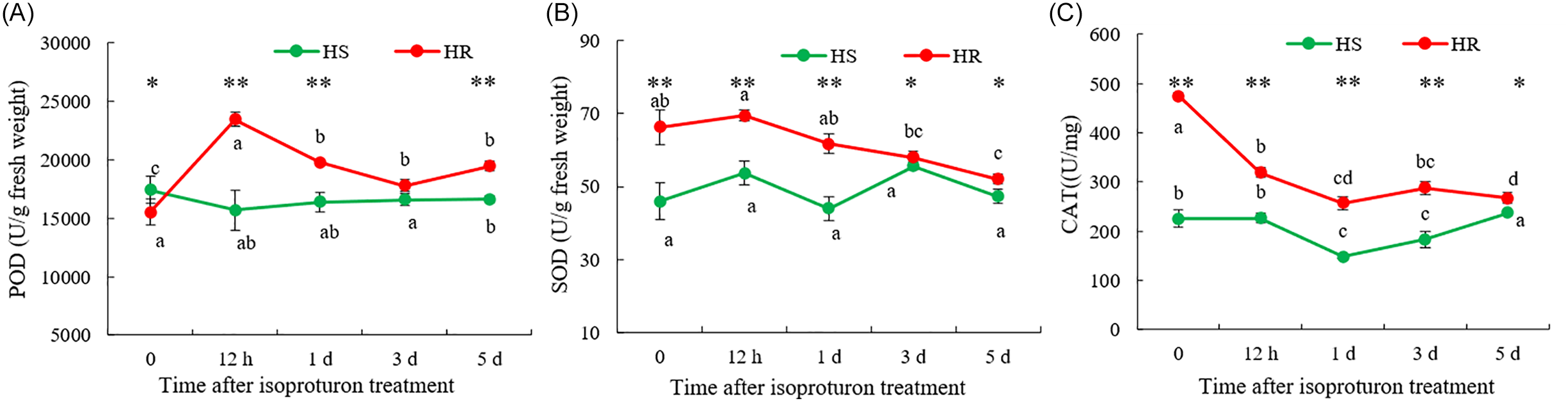
Comparative analysis of antioxidant enzyme activities revealed that the enzyme activities in the HR population were consistently higher than those in the HS population. Specifically, POD activity in the HR population was rapidly and strongly induced by isoproturon treatment, peaking at 12 h at a level 1.5-fold higher than that in the HS population (Figure 3A). Although this activity subsequently declined, it remained consistently elevated above the HS level throughout the experiment. In contrast, the HS population showed no significant response. A similar trend was observed for SOD activity (Figure 3B). The basal SOD level in the untreated HR population was 1.4-fold higher than that in the HS population. Although isoproturon gradually reduced the SOD activity of the HR population, it was still higher than that of the HS population during the experiment. The most significant difference between the HS population and the HR population was CAT activity (Figure 3C). Without treatment, the CAT activity of the HR population was 2.11-fold higher than that of the HS population. After treatment with isoproturon, the CAT activity of the HR population remained significantly higher than that of the HS population. Therefore, the results of antioxidant enzymes indicated that the HR population maintained a highly active antioxidant system, and the activities of POD, SOD, and CAT were significantly higher than in the HS population. This might more effectively eliminate the reactive oxygen species (ROS) induced by herbicides, thereby generating resistance to isoproturon in the HR population.
Antioxidant enzyme activities of Lolium perenne ssp. multiflorum populations from China sensitive (HS) and resistant (HR) to isoproturon. Peroxidase (POD) activity (A), superoxide dismutase (SOD) activity (B), and catalase (CAT) activity (C) of HS and HR populations after isoproturon treatment. Lowercase letters indicate comparisons between different time points within the same population, data are means ± SE (n = 3). * denote comparisons between different populations at the same time. * represents P < 0.05 and ** represents P < 0.01. One-way ANOVA was used to analyze the significance of the data, P < 0.05.

To mitigate oxidative damage induced by environmental stresses, plants have evolved sophisticated defense systems that include antioxidant enzymes such as SOD, CAT, and POD (Garcia-Caparros et al. Reference Garcia-Caparros, De Filippis, Gul, Hasanuzzaman, Ozturk, Altay and Lao2021; Kubis et al. Reference Kubis, Marques, Kitamura, Barroso, Juneau and Gomes2022). Among these, SOD constitutes the first line of defense against oxidative stress by catalyzing the dismutation of superoxide radicals into oxygen and hydrogen peroxide (Alscher et al. Reference Alscher, Erturk and Heath2002). POD and CAT further contribute to ROS detoxification by converting H2O2 into water or other non-toxic molecules and are widely used as biomarkers of oxidative stress in plants. Previous studies have documented that herbicide-resistant weeds often exhibit enhanced antioxidant capacity, including elevated activities of SOD, CAT, and ascorbate peroxidase (APX), which facilitate the maintenance of ROS homeostasis and prevent oxidative damage (Gomes et al. Reference Gomes, Le Manac’h, Moingt, Smedbol, Paquet, Labrecque, Lucotte and Juneau2016; Han et al. Reference Han, Yu, Beffa, Gonzalez, Maiwald, Wang and Powles2021; Kubis et al. Reference Kubis, Marques, Kitamura, Barroso, Juneau and Gomes2022). Consistent with these findings, the HR population showed significantly higher activities of POD, SOD, and CAT compared with the HS population (Figure 3). These results suggest that the resistance of the HR population to isoproturon may be associated with its enhanced antioxidant enzyme activities, which likely improve the ability of L. perenne ssp. multiflorum to cope with herbicide-induced oxidative stress, thereby increasing survival rate under isoproturon treatment.
Chlorophyll and Carotenoid Contents
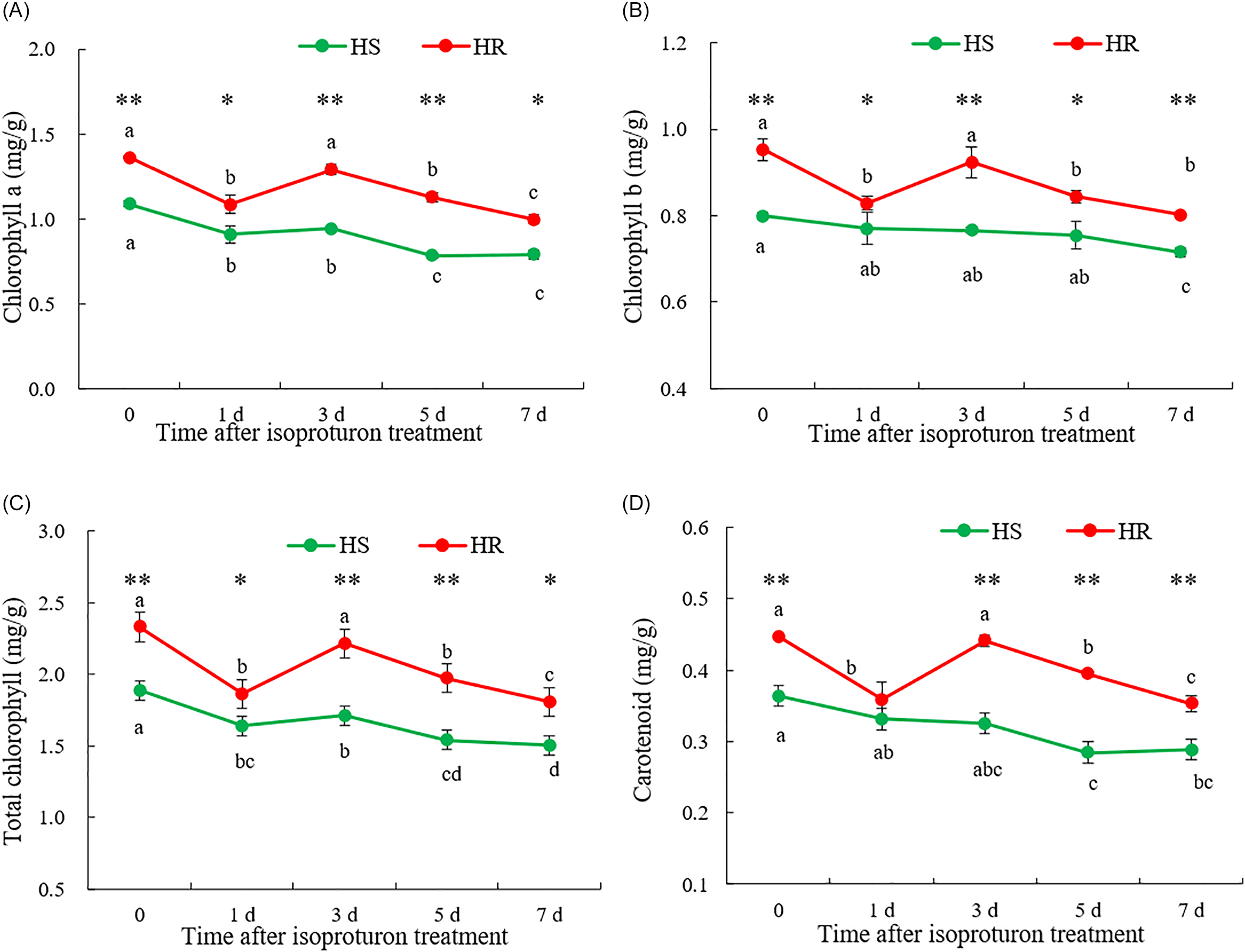
The effects of isoproturon on photosynthetic pigments in the HR and HS populations were tested; isoproturon reduced the chlorophyll and carotenoid contents of L. perenne ssp. multiflorum, but the chlorophyll and carotenoid contents in the HR population were significantly higher than those in the HS population (Figure 4). After isoproturon treatment, the chlorophyll a content in both groups decreased; at 7 d, it had decreased in the HR and HS populations by 26.81% and 27.32%, respectively. However, the chlorophyll a content of the HR population was significantly higher than that of the HS population at all time points. It was 25.11% higher at 0 h and 24.46% higher at 7 d (Figure 4A). During the entire experimental process, the chlorophyll b content of the HR population was still significantly higher than that of the HS population, with the initial value and the final value being 19.24% and 11.81% higher, respectively (Figure 4B). The total chlorophyll content and carotenoid content followed a similar trend (Figure 4C and 4D). Overall, these results indicated that compared with the HS group, the HR group maintained a higher level of photosynthetic pigment content. Although isoproturon caused a reduction in photosynthetic pigments in both populations, the HR population maintained a relatively high pigment content, indicating that the protection or stability of its photosynthetic organs was stronger. This might help it develop herbicide resistance by stabilizing photosynthetic function under herbicide stress.
Chlorophyll and carotenoid contents of Lolium perenne ssp. multiflorum populations from China sensitive (HS) and resistant (HR) to isoproturon. Chlorophyll a (A), chlorophyll b (B), total chlorophyll content (C), and carotenoid content (D) of HS and HR populations after isoproturon treatment. Lowercase letters indicate comparisons between different time points within the same population; data are means ± SE (n = 3). * denote comparisons between different populations at the same time. * represents P < 0.05 and ** represents P < 0.01. One-way ANOVA was used to analyze the significance of the data, P < 0.05.

Photosynthesis serves as the fundamental process supporting essential life activities in green plants, with photosystem I (PSI) and PSII constituting the core photoreaction centers (Nelson and Yocum Reference Nelson and Yocum2006). Isoproturon, a PSII inhibitor, leads to chlorophyll degradation and plant desiccation (Arnaud et al. Reference Arnaud, Taillandier, Kaouadji, Ravanel and Tissut1994). Consistent with this mechanism, the present study observed a decline in both chlorophyll content and photosynthetic rate in L. perenne ssp. multiflorum following isoproturon application. As shown in Figure 4, the HR population maintained higher chlorophyll and carotenoid levels compared with the HS population, indicating a milder physiological impact under herbicide stress. These pigment-related findings are consistent with the results of the whole-plant bioassay. We propose that the observed overexpression of psbA in the HR population may attenuate the effective inhibitory action of isoproturon on the overproduced D1 protein, thereby contributing to the development of the herbicide-resistant phenotype.
Heterogeneous Expression of Lolium perenne ssp. multiflorum psbA in Oryza sativa and Herbicide Sensitivities
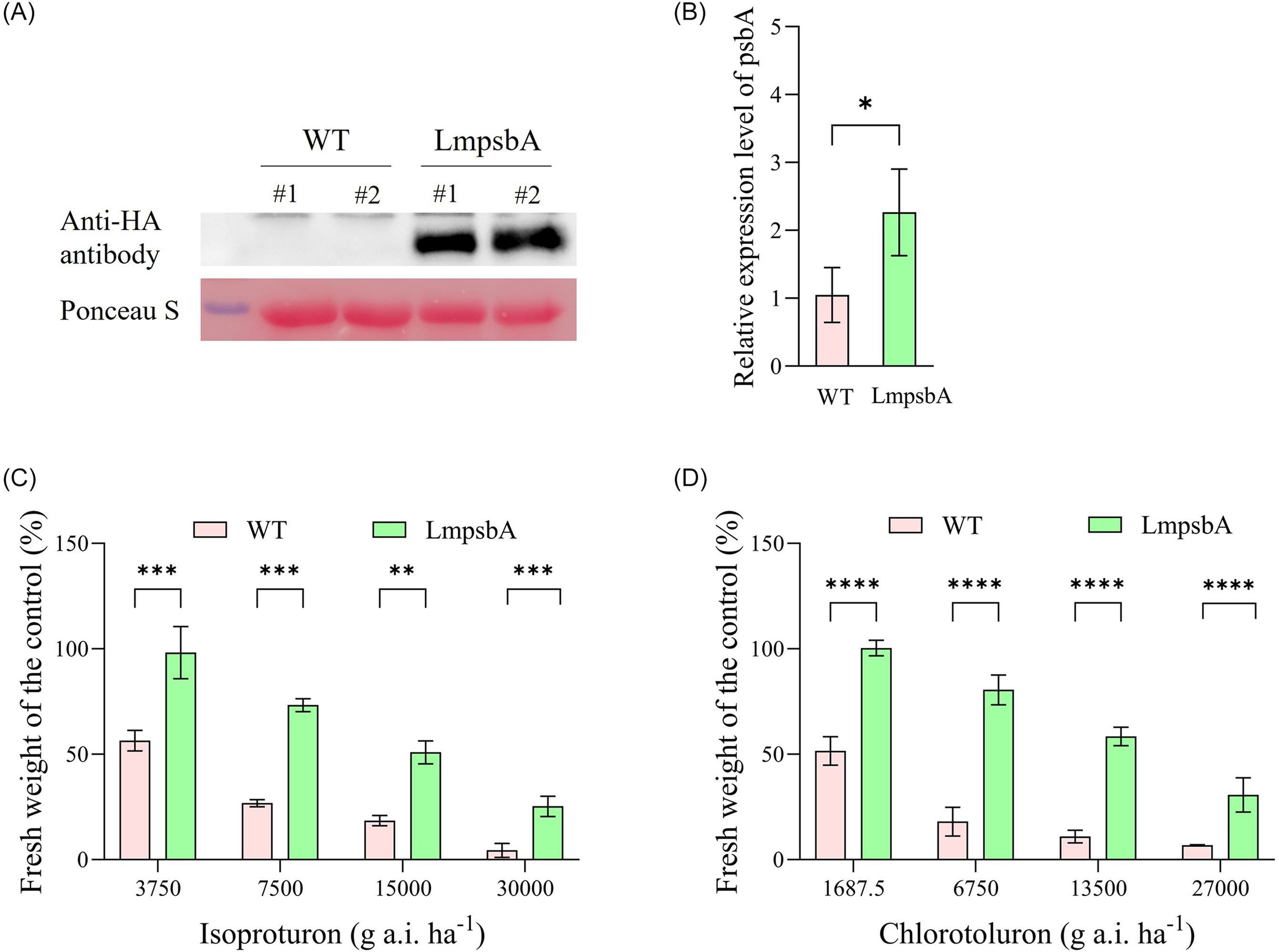
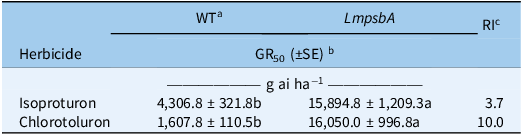
To validate the role of LmpsbA overexpression in isoproturon resistance, we introduced LmpsbA into rice plants for overexpression. The expression level of the LmpsbA gene in transgenic O. sativa is shown in Figure 5A and 5B. The expression of LmpsbA in the transgenic O. sativa (T2 line) was 2.26 times higher than that in the wild type (WT), with the expression level in WT set as 1.0. Due to the high homology between the LmpsbA gene and the rice psbA gene, the rice psbA gene could also be amplified. As shown in Figure 5 and Table 1, the GR50 of isoproturon in the transgenic O. sativa overexpressing LmpsbA was 15,894.8 g ha−1, and the GR50 in the WT was 4,306.8 g ha−1. The RI value of the transgenic O. sativa was 3.7. Similarly, under chlorotoluron treatment, the GR50 value of transgenic O. sativa was 16,050.0 g ha−1, which was significantly higher than that of WT (1,607.8 g ha−1), and the RI value was 10.0 (Figure 5). Therefore, these results confirm that LmpsbA overexpression is a functional resistance mechanism in L. perenne ssp. multiflorum, explaining the isoproturon resistance observed in the HR population.
Herbicide sensitivity of transgenic rice plants expressing LmpsbA. (A) Western blot analysis of LmpsbA protein expression in transgenic rice (T1 line). (B) The relative expression level of the psbA gene in T2 generation transgenic rice. OsActin2 as the internal reference gene. The survival rates of transgenic rice plants expressing LmpsbA after isoproturon treatment (C) and chlorotoluron treatment (D). WT, wild type. *P < 0.05; **P < 0.01; a t-test was used to analyze the significance between the LmpsbA-expressed and WT plants.

Sensitivity of LmpsbA-expressed Oryza sativa plants to different herbicides.

a WT is wild type.
b GR50 is the effective dose of herbicide causing 50% inhibition of fresh weight and is expressed as grams of active ingredient per hectare (g ai ha−1). Different letters indicate significant differences between different treatments (P < 0.05).
c RI is the relative tolerance index; RI = GR50 (LmpsbA)/GR50 (WT).
Multiple Resistance
The results of the whole-plant dose–response assays indicated that the HR population exhibited varying degrees of resistance to ALS-inhibiting and ACCase-inhibiting herbicides (Table 2). The HR population was highly resistant to pyroxsulam with an RI of 204.5, while the GR50 value of the HR population was found to be below the recommended field rate of pyroxsulam (15 g ha−1). The HR population also demonstrated a certain degree of resistance to pinoxaden, with an RI value of 4.4, and exhibited a GR50 value lower than the recommended field rate of pinoxaden (60 g ha−1). It can therefore be concluded that both pyroxsulam and pinoxaden are effective in controlling the HR population and can be employed to manage L. perenne ssp. multiflorum populations exhibiting resistance to isoproturon in wheat fields.
Sensitivity of Lolium perenne ssp. multiflorum populations from China sensitive (HS) and resistant (HR) to pyroxsulam and pinoxaden.

a GR50 is the effective dose of herbicide causing 50% inhibition of fresh weight and is expressed as grams of active ingredient per hectare (g ai ha−1). Different letters indicate significant differences between different treatments (P < 0.05).
b RI is the relative resistance index; RI = GR50 (HR)/ GR50 (HS).
In conclusion, our study supports that LmpsbA gene overexpression can induce the resistance of L. perenne ssp. multiflorum to isoproturon. In addition, the higher activity of antioxidant enzymes and the high levels of pigment content are also the reasons why the L. perenne ssp. multiflorum population developed resistance to isoproturon. To mitigate the proliferation of L. perenne ssp. multiflorum, comprehensive field management strategies, such as herbicide rotation and other nonchemical approaches, are indispensable. Our study provides new insights into isoproturon resistance in L. perenne ssp. multiflorum.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/wsc.2026.10106
Funding statement
This work was supported by the National Natural Science Foundation of China (32372567), the National Key R&D Program of China (2023YFD1400501) and the Student Innovation Research & Entrepreneurship Training of Jiangsu (S202510307071).
Competing interests
The authors declare no conflicts of interest.




 Open access
Open access