


Introduction
In temperate climates, many plants exhibit physiological dormancy that is released by cold winter conditions (CC Baskin and JM Baskin Reference Baskin and Baskin2014; JM Baskin and CC Baskin Reference Baskin and Baskin2004; Finch-Savage and Leubner-Metzger Reference Finch-Savage and Leubner-Metzger2006). This stratification requirement is common among temperate weed species (Batlla et al. Reference Batlla, Ghersa and Benech-Arnold2020; Duke Reference Duke2018; Gioria and Pyšek Reference Gioria and Pyšek2017). For such weeds, physiological dormancy and dormancy polymorphism serve multiple purposes, for example, avoiding winter germination and temporal bet-hedging. Dormancy dynamics influence the likelihood of weed population expansion in different environments. For instance, weedy or invasive species requiring shorter stratification periods could outperform species requiring longer stratification periods in regions with short winters, or if winters are shortened by climate change (Gioria and Pyšek Reference Gioria and Pyšek2017). More broadly, species with shorter stratification requirements may be better able to germinate early in the growing season, potentially gaining a competitive advantage over other plants (Gioria et al. Reference Gioria, Pyšek and Osborne2018). A better understanding of weed seed dormancy can help improve invasion risk assessments and models of weed seedling emergence to enable time-specific weed management (Batlla and Luis Benech-Arnold Reference Batlla and Benech-Arnold2007; Grundy Reference Grundy2003; Marschner et al. Reference Marschner, Colucci, Stup, Westbrook, Brunharo, DiTommaso and Mesgaran2024).
This study focuses on four species that can behave as weeds in the context of tallgrass prairie management for livestock production. The four species are Baldwin’s ironweed (Vernonia baldwinii Torr.), common evening primrose (Oenothera biennis L.), hollow Joe-pye weed [Eutrochium fistulosum (Barratt) E.E. Lamont], and tall goldenrod (Solidago altissima L.). Oenothera biennis is usually biennial, and the others are perennial. Vernonia baldwinii is unpalatable to cattle and increases with overgrazing (Missouri Department of Conservation 2025b). This species is associated with a long history of chemical and mechanical control efforts (Klingman and McCarty Reference Klingman and McCarty1958; McCarty and Linscott Reference McCarty and Linscott1962; McCarty and Scifres Reference McCarty and Scifres1969). Oenothera biennis exhibits opportunistic behavior, especially in disturbed habitats (Hall et al. Reference Hall, Steiner, Threadgill and Jones1988; Steckel et al. Reference Steckel, Sosnoskie and Steckel2019). Its impressive seed longevity (80 yr) and tolerance to herbicides such as glyphosate and dicamba contribute to difficulties with control (Steckel et al. Reference Steckel, Sosnoskie and Steckel2019). Although less frequently considered a problematic weed, E. fistulosum, like other Eutrochium species, is mostly not palatable to grazing animals (Missouri Department of Conservation 2025a). Solidago altissima has a clonal growth habit, enabling it to aggressively outcompete other plants (Stoll et al. Reference Stoll, Egli and Schmid1998; Szymura et al. Reference Szymura, Szymura, Wolski and Świerszcz2018; Werner et al. Reference Werner, Gross and Bradbury1980). This species has also been reported to exhibit allelopathic effects against forage crops (Gam et al. Reference Gam, Adhikari, Kang, Injamum-Ul-Hoque, Shaffique, Woo, Jeon, An, Back, Kim, Kang and Lee2024).
In general, many North American prairie forbs in the family Asteraceae, which would include V. baldwinii, E. fistulosum, and S. altissima, require 4 to 6 wk of moist-cold stratification to break dormancy (Hind Reference Hind2023). Recommendations for propagation of Vernonia and Eutrochium species have suggested 30 d of stratification, although some sources suggest up to 90 d (Belt Reference Belt2012; Diboll Reference Diboll2008; Hind and Langhorne Reference Hind and Langhorne2023). Baskin et al. (Reference Baskin, Baskin and Leck1993) stratified E. fistulosum seeds from Ohio, USA, and S. altissima seeds from Kentucky, USA, at 5 C for 0, 4, 6, 8, 10, or 12 wk. In both species, percentage germination increased with increasing stratification duration. Stratification also reduced the minimum temperature at which seeds would germinate (a Type 2 response; Baskin et al. Reference Baskin, Baskin and Leck1993). In O. biennis, seeds from Ontario, Canada, and Ohio and Kentucky were subjected to 0, 4, 8, or 12 wk of stratification at 4 C, then germinated at temperatures of 5/5, 20/10, 25/15, 30/20, or 35/25 C (Susko et al. Reference Susko, Ismail and Rahman2025). There was a three-way interaction between population (state of origin), stratification duration, and germination temperature. The seeds from Kentucky did not respond to stratification. For the other seeds, increasing stratification duration helped rescue germination at low temperatures, but stratification was not necessary to enable germination at higher temperatures (Susko et al. Reference Susko, Ismail and Rahman2025).
Our study tested the relationship between stratification duration and germination in rangeland biotypes of these four weed species from Kansas, USA. For the Asteraceae species, our study is among the first formal analyses of stratification response. For O. biennis, given that geographic location is known to impact response to stratification, our study is novel in that it introduces a more western population. As Kansas has a more arid climate than the states in which most research has been conducted, it is plausible that we might see a different pattern.
For all species, we hypothesized that stratification (2, 4, 6, or 12 wk) would increase total germination and reduce time to germination relative to the unstratified control (0 wk). We also expected that increasing stratification duration would increase these benefits, but that we would start to see diminishing returns at longer durations (i.e., the 6-wk and 12-wk treatments might be similar).
Materials and Methods
Seed Sources
All seeds used in the study (V. baldwinii, O. biennis, E. fistulosum, and S. altissima) were collected near Manhattan, KS (collection coordinates: 39.11°N to 39.14°N, 96.54°W to 96.52°W) on October 15, 2024. Seeds were collected from range habitat (V. baldwinii), two plants in roadside habitat (O. biennis), one plant in range habitat (E. fistulosum), and range habitat (S. altissima). The seeds of each species were mature at the time of collection. All seeds were stored under dry, dark conditions in paper envelopes in a refrigerator at approximately 3 C until the experiment, which was conducted in 2025. For Run 1, data were collected from June 24 to October 7, 2025. For Run 2, data were collected from September 6 to December 17, 2025.
Experimental Procedure
The experimental treatments were factorial combinations of species (V. baldwinii, O. biennis, E. fistulosum, and S. altissima) by stratification treatment (0-wk control, 2 wk, 4 wk, 6 wk, or 12 wk). For each combination, we tested five replicate petri dishes, each containing 15 seeds. The experiment was repeated in time (two runs), for a total of 10 petri dishes tested per combination of species and stratification treatment.
Dry seeds were rehydrated by soaking them in water in a sealed container for 24 h. They were sanitized by being soaked in 1% sodium hypochlorite solution for 10 min, then thoroughly rinsed with distilled water for 10 min. After sanitization, batches of 15 seeds were placed in a single layer in a moistened paper towel within a sealed plastic bag. The plastic bags were refrigerated at approximately 3 C under dark conditions for the designated stratification period. The seeds were monitored for mold growth during stratification and remoistened by lightly spraying them with distilled water as necessary.
At the end of the stratification period, seeds were removed and placed into petri dishes (15 seeds per dish). The petri dishes contained two sheets of filter paper, moistened with 6 to 10 ml of distilled water. Petri dishes were placed into a growth chamber (Conviron ATC60 Multi-Tier Reach-In, Winnipeg, MB, Canada). The growth chamber settings maintained throughout both runs of the experiment were 26/22 C (light/dark) and 75% relative humidity at 12-h photoperiod. These conditions were selected to mimic spring conditions in Kansas and because a temperature regime of 26/22 C is suitable for germination of the study species or closely related species (Baskin et al. Reference Baskin, Baskin and Leck1993; Hind and Langhorne Reference Hind and Langhorne2023; Meyer and Schmid Reference Meyer and Schmid1999; Susko et al. Reference Susko, Ismail and Rahman2025). In the chamber, petri dishes were checked daily and moistened with distilled water as needed to keep filter paper moist.
The number of germinated seeds was determined at 7-d intervals for 28 d. Germination was defined as visible radicle emergence. To prevent overcrowding and mold, seedlings were removed when the radicle reached 2-mm long. At the end of the 28-d period, potential viability of ungerminated seeds was assessed by hand dissection and visual assessment. The visual assessment criteria were that seeds that were visibly moldy, easily crushed, or otherwise abnormal were recorded as not viable. Visually normal seeds were recorded as potentially viable. We note that a tetrazolium test was not performed, which represents a limitation of the experiment. For this reason, it is possible that some seeds categorized as potentially viable were actually not viable. However, qualitative procedures involving crush tests and visual inspection often give similar results to tetrazolium tests as assessments of weed seed viability (Sawma and Mohler Reference Sawma and Mohler2002).
Statistical Analysis
We excluded two petri dishes (both S. altissima) in which the recorded number of seeds in a dish did not sum to 15. Thus, the number of data points in the analysis was 198 (4 species by 5 treatments by 5 replicates by 2 runs, minus 2).
Data analyses were performed in R v. 4.4.1 (R Core Team 2024). The probability of germination was analyzed using a generalized linear mixed model with a logit link and beta-binomial family (package glmmTMB). The beta-binomial was selected rather than binomial due to overdispersion. Species, stratification treatment, and the interaction between species and stratification treatment were included as fixed effects. Run was retained as a random effect, because the model including run fit better than the corresponding model without run, based on a likelihood-ratio test (P = 0.04) and a lower Akaike information criterion. Model fit was assessed using simulated residuals (package DHARMa) to create diagnostic plots and to conduct dispersion and zero-inflation tests. After verifying that assumptions were met, we performed Type 3 ANOVA (package car) and post hoc testing using the Tukey method for P-value adjustment (package emmeans). Post hoc tests compared stratification treatments within species. Tests were performed on the log odds ratio scale at α = 0.05. Estimates and 95% confidence intervals were back-transformed from this scale. For petri dishes that did not reach 100% germination, we used the same modeling approach to evaluate probability of viability (as defined earlier under “Experimental Procedure”) among nongerminated seeds.
Cumulative germination over time was modeled separately for each species. For these models, we used generalized linear mixed models with a logit link and binomial family, as the beta-binomial family did not improve model fit for any of the four species (packages glmmTMB, DHARMa). Fixed effects were stratification treatment, time (Day 7, Day 14, Day 21, or Day 28), and the interaction between stratification treatment and time. Random effects were run and petri dish (accounting for repeated measures). Model diagnostics using simulated residuals were performed as described earlier, except that zero-inflation was not considered, because lower values would be biologically expected at Day 7. For O. biennis, we observed minor departures from assumptions in the quantile–quantile and residual versus predicted plots, but no dispersion issues. The model for this species was therefore analyzed qualitatively. For other species, we used the package emmeans to calculate the instantaneous slope (rate of increase in cumulative germination) at the mean value of time.
Results and Discussion
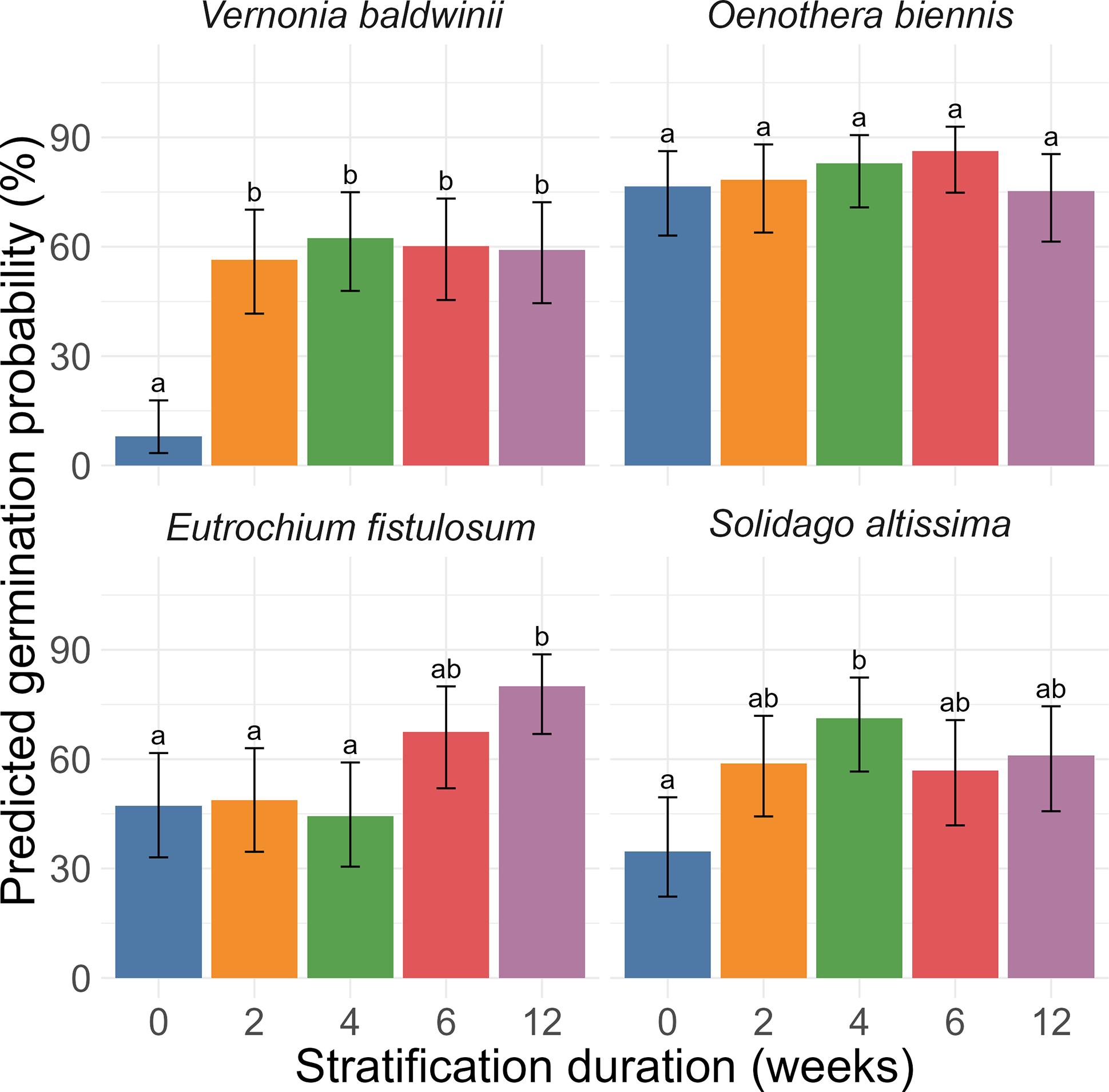
Species (P < 0.001), stratification treatment (P < 0.001), and the interaction between species and stratification treatment (P < 0.001) jointly impacted the probability of germination (Figure 1). For V. baldwinii, germination probability was lower for seeds in the 0-wk treatment (8%) than seeds in all other treatments (56% to 62%). For O. biennis, germination probability was between 75% and 86% and did not differ between stratification treatments. For E. fistulosum, germination in the 12-wk treatment (80%) exceeded germination in the 4-wk, 0-wk, and 2-wk treatments (44% to 49%). The 6-wk treatment was intermediate (68%). For S. altissima, germination in the 4-wk treatment (71%) exceeded germination in the 0-wk treatment (35%). Other treatments were intermediate (57% to 61%).
Effects of species and stratification duration (weeks) on germination probability. Data are back-transformed estimates with 95% confidence intervals. Within species, stratification duration treatments indicated by the same letter are not significantly different at α = 0.05. Each estimate represents 10 petri dishes (5 replicates by 2 runs), each containing 15 seeds.

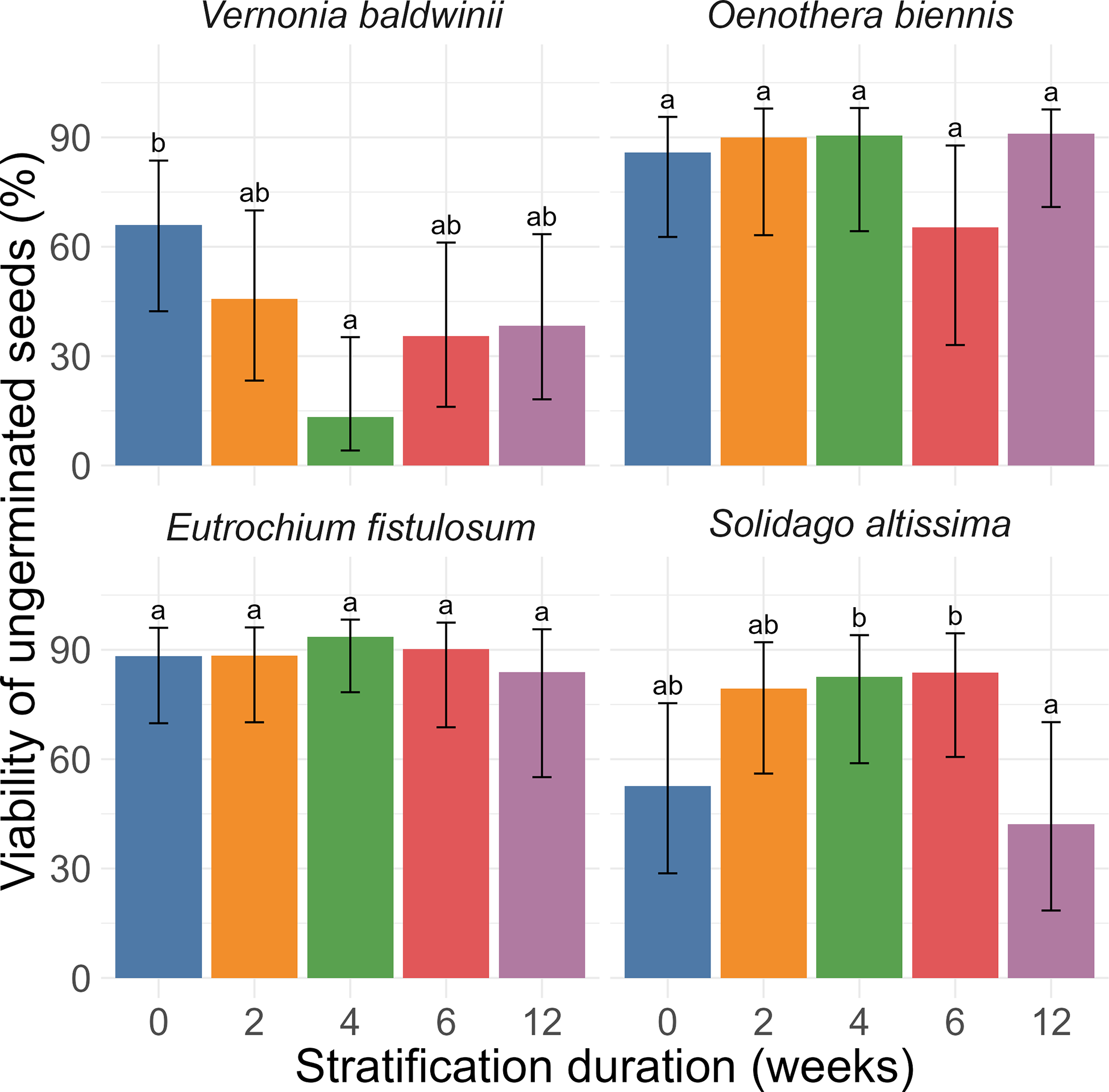
Among seeds that did not germinate, the probability of potential viability depended on species (P < 0.001) and the interaction between species and stratification treatment (P < 0.001) (Figure 2). For V. baldwinii, the probability of potential viability was higher in the 0-wk treatment (66%) than the 4-wk treatment (13%). Other treatments were intermediate (35% to 46%). For O. biennis, the probability of potential viability did not differ among treatments (65% to 91%). Likewise, for E. fistulosum, the probability of potential viability did not differ among treatments (84% to 93%). Finally, for S. altissima, the probability of potential viability was higher in the 4-wk (83%) and 6-wk (84%) treatments than the 12-wk treatment (42%). Other treatments were intermediate (53% to 79%).
Effects of species and stratification duration (weeks) on the probability of viability among seeds that did not germinate. Data are back-transformed estimates with 95% confidence intervals. Within species, stratification duration treatments indicated by the same letter are not significantly different at α = 0.05. Each estimate represents up to 10 petri dishes (5 replicates by 2 runs), each containing 15 seeds. Petri dishes in which all 15 seeds germinated were not included in this analysis.

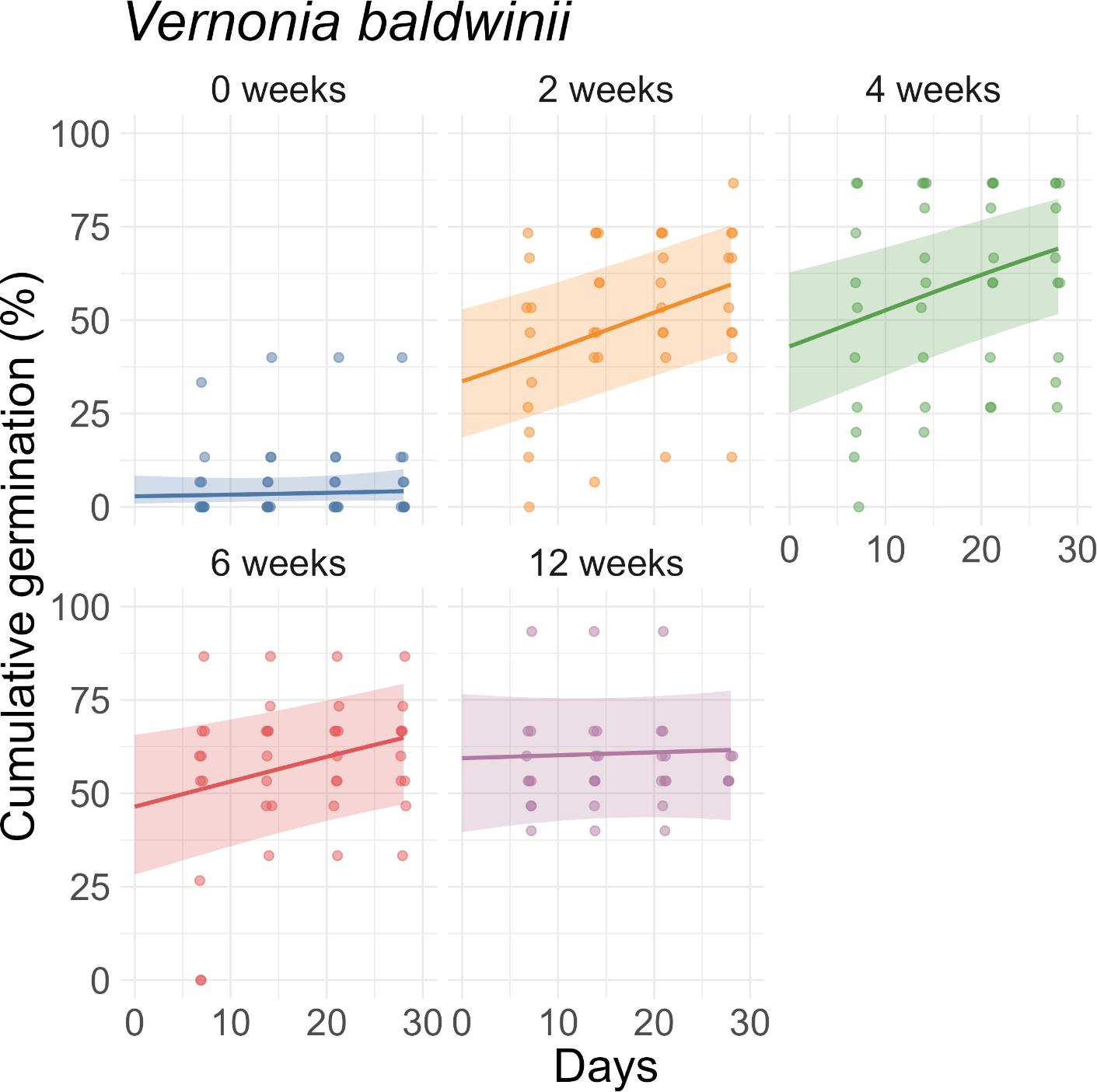
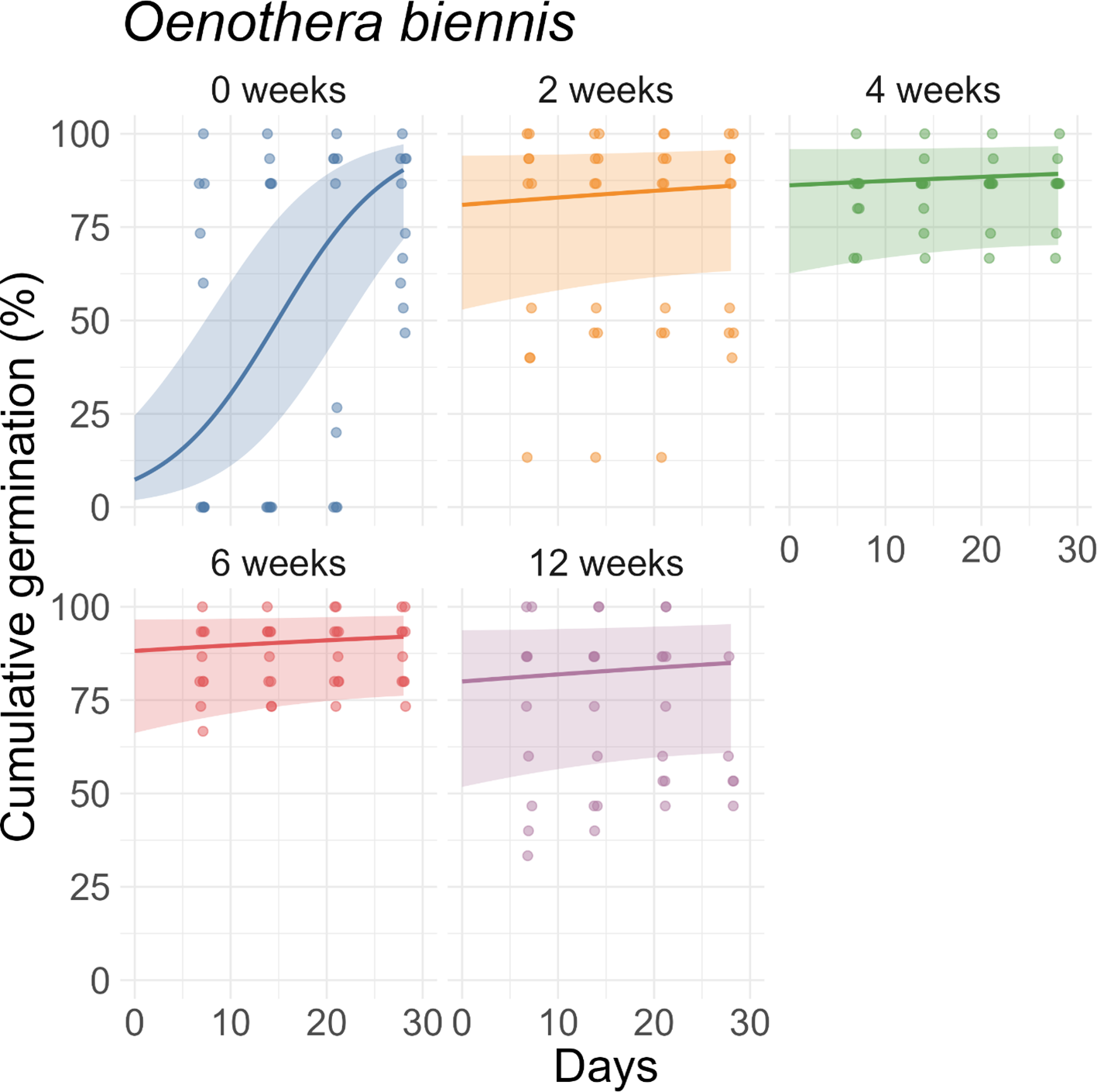
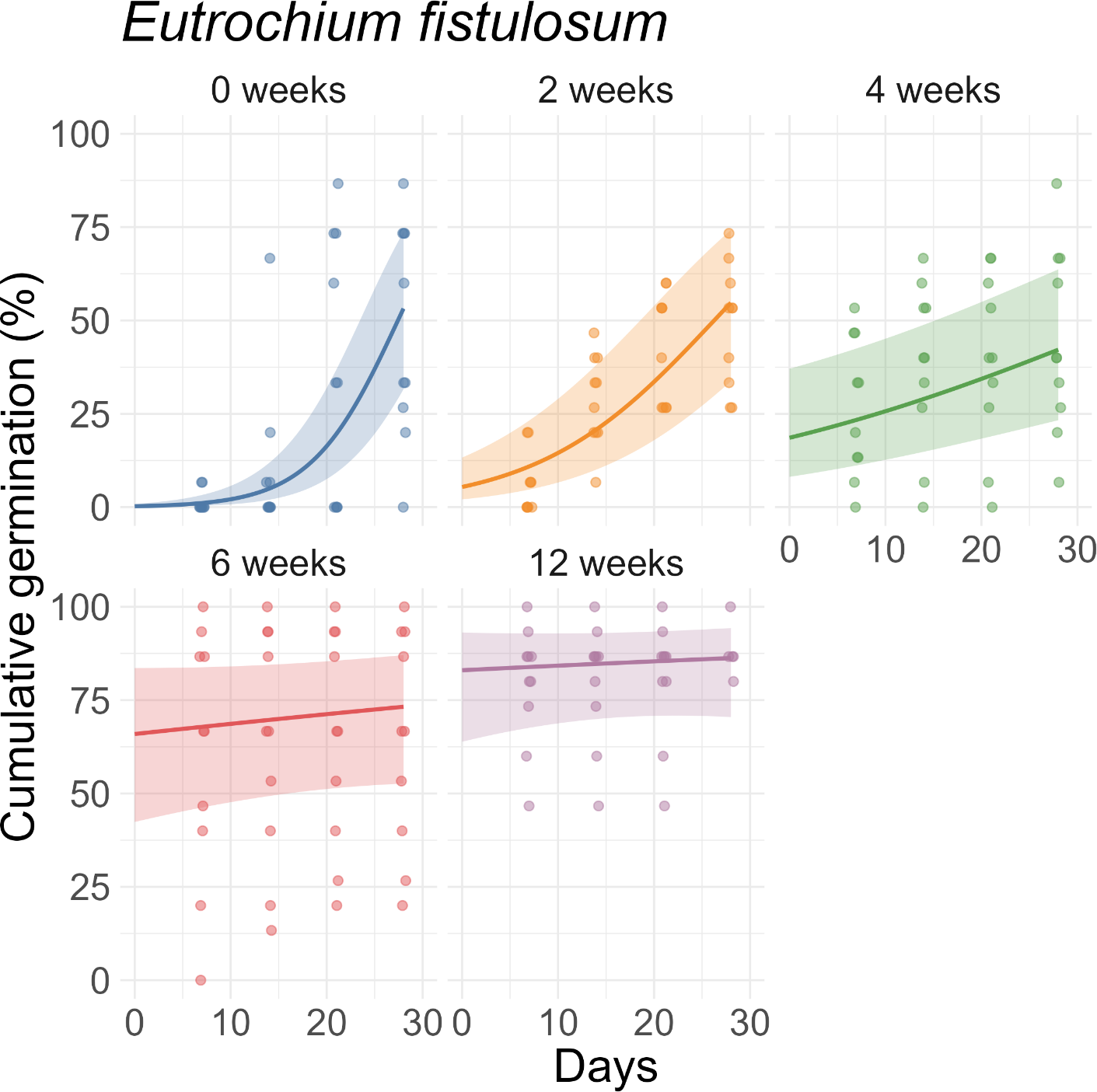
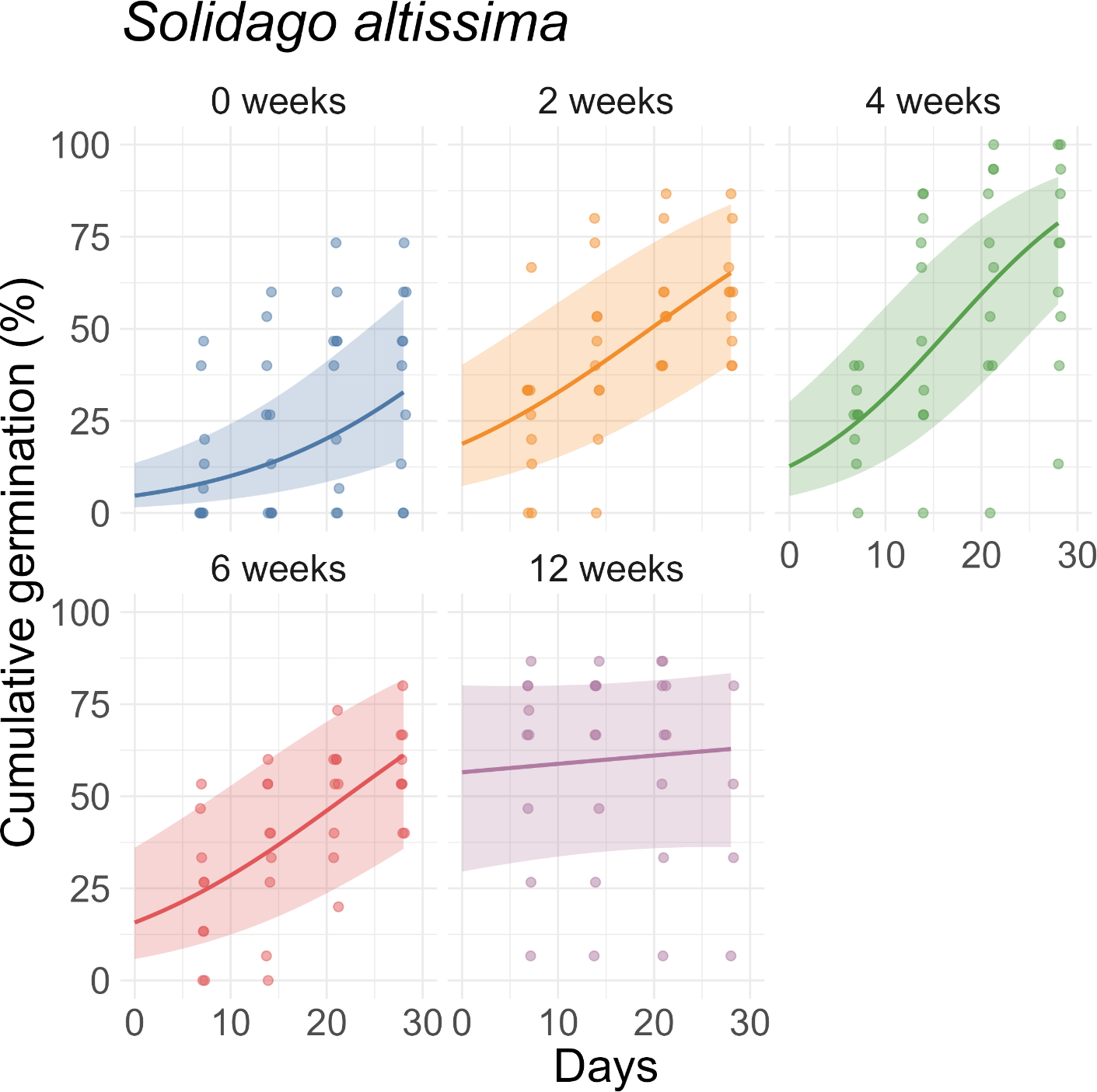
For V. baldwinii, the rate of increase in cumulative germination reached a few percent per day in the 2-wk, 4-wk, and 6-wk stratification treatments (Figure 3). However, for the 12-wk treatment, the rate of increase in cumulative germination was approximately an order of magnitude slower. This slow rate of change indicates that, for the 12-wk stratification treatment, nearly all the germination that would occur had already happened by the 7-d mark (Figure 3). For O. biennis, the rate of increase in cumulative germination (assessed halfway through the germination period) was much larger in the 0-wk treatment than in the other treatments, indicating a longer average time to germination (Figure 4). For E. fistulosum, the rate of increase in cumulative germination decreased with increasing stratification duration (Figure 5). For S. altissima, the rate of increase in cumulative germination was again lowest in the 12-wk stratification treatment (Figure 6). Full results from these models (fixed-effect estimates, standard errors, and P-values) are reported in Supplementary Tables S1–S4.
Effects of stratification duration (0, 2, 4, 6, or 12 wk) on cumulative germination of Vernonia baldwinii. Points are observed percentages of seeds germinated per dish. Lines and shaded ribbons represent predictions from a binomial generalized linear mixed model with 95% confidence intervals.

Effects of stratification duration (0, 2, 4, 6, or 12 wk) on cumulative germination of Oenothera biennis. Points are observed percentages of seeds germinated per dish. Lines and shaded ribbons represent predictions from a binomial generalized linear mixed model with 95% confidence intervals.

Effects of stratification duration (0, 2, 4, 6, or 12 wk) on cumulative germination of Eutrochium fistulosum. Points are observed percentages of seeds germinated per dish. Lines and shaded ribbons represent predictions from a binomial generalized linear mixed model with 95% confidence intervals.

Effects of stratification duration (0, 2, 4, 6, or 12 wk) on cumulative germination of Solidago altissima. Points are observed percentages of seeds germinated per dish. Lines and shaded ribbons represent predictions from a binomial generalized linear mixed model with 95% confidence intervals.

The finding that O. biennis did not respond strongly to stratification (Figure 1) is largely consistent with previous research. Although O. biennis is known to exhibit some physiological dormancy (Baskin and Baskin Reference Baskin and Baskin1994; Nightingale and Baker Reference Nightingale and Baker1995; Steckel et al. Reference Steckel, Sosnoskie and Steckel2019), a previous study reported that the positive impact of stratification was strongest when subsequent germination temperatures were too low (Susko et al. Reference Susko, Ismail and Rahman2025). Because our seeds germinated under favorable conditions (26/22 C), prior stratification may not have been crucial. Previous research also reported no stratification requirement for seeds from the warmest region tested (Kentucky; Susko et al. Reference Susko, Ismail and Rahman2025), and our seeds were collected from a relatively warm climate as well (Kansas). Third, our seeds had been stored under cold conditions (approximately 4 C), although not in an imbibed state, before the experiment. The cold storage period might have helped alleviate primary dormancy even without moist-cold stratification.
All three Asteraceae species were significantly impacted by stratification. For V. baldwinii, 2 wk of stratification was sufficient to maximize total germination (Figure 1). Most seeds that failed to germinate after stratification appeared not to be viable (Figure 2). This finding suggests that, although 1 to 2 mo of stratification are typically recommended by those seeking to propagate V. baldwinii (Diboll Reference Diboll2008; Prairie Moon Nursery 2025), a winter with only brief periods of cold weather should not prevent recruitment in the subsequent season. However, a longer stratification period might accelerate germination once temperatures are suitable, potentially increasing the weed’s competitive potential.
Our findings support the idea that germination of E. fistulosum and S. altissima may be maximized with longer stratification periods (12 wk for E. fistulosum and 4 wk for S. altissima). These periods are broadly consistent with the 4+ wk often used as a rule of thumb for Asteraceae species (Hind Reference Hind2023). In E. fistulosum, previous research found that non-stratified seeds could germinate at 35/20 or 30/15 C (Baskin et al. Reference Baskin, Baskin and Leck1993). However, stratified seeds could germinate at 35/20, 30/15, 25/15, 20/10, or 15/6 C. In most germination-temperature environments, percent germination increased with increasing stratification duration up to 6 or 8 wk of stratification. However, in the coldest germination-temperature environment (15/6 C), percent germination increased with increasing stratification duration up to 12 wk of stratification (Baskin et al. Reference Baskin, Baskin and Leck1993). Similar results were observed for S. altissima, except that increasing stratification duration continued to enhance percent germination up to 10 or 12 wk of stratification in most germination-temperature environments (Baskin et al. Reference Baskin, Baskin and Leck1993). Although our results are slightly different (suggesting that the optimal stratification duration is longer in E. fistulosum than S. altissima), we also found a positive relationship between stratification duration and percent germination.
Overall, our research demonstrates that, although stratification can help release dormancy and enhance germination in weedy rangeland forbs, a long period of cold weather is not essential. Variable winter and spring weather conditions are more likely to cause erratic germination patterns than to prevent germination altogether. Our study adds to the growing body of literature that can be used to develop more mechanistic weed emergence models for decision support. As reported for agronomic situations (Marschner et al. Reference Marschner, Colucci, Stup, Westbrook, Brunharo, DiTommaso and Mesgaran2024), these models will help improve weed management efficacy. For example, they can help land managers identify the optimal time to spray herbicides and gauge how the timing of mowing, spraying, or prescribed fire operations would influence weed recruitment. For land managers faced with weed recruitment from a persistent seedbank, these advances will lead to more cost-effective and time-efficient weed management strategies.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/wsc.2026.10118
Acknowledgments
We thank Grant Snider and Lisa Auen for collecting the seeds used in this study.
Funding statement
This research was supported in part by the Weed Science Society of America Innovative Grants Program (2024 to 2025).
Competing interests
The authors declare no conflicts of interest.



 Open access
Open access