


Highlights
-
• L2 processing of derived words engages morpho-orthographic segmentation.
-
• L2 processing of derived words engages morpho-semantic integration.
-
• L2 processing lacks target-like lateral inhibition in orthographic priming.
-
• Weak lexical representations induce less robust (morpho-)semantic priming in L2.
-
• L2 structural parsing reliably emerges for simple structures with sufficient time.
1. Introduction
One fundamental aspect of linguistic knowledge is the ability to create new words from existing ones. Many languages achieve this through derivation, a morphological process whereby affixes are added to base words to form new lexical items. For example, in English, the suffix -er can be added to certain verbs to create nouns that denote the agent performing the action (e.g., teach-er). Learners of languages utilizing derivational morphology are therefore expected to acquire knowledge of how derived words are structured and interpreted for efficient lexical recognition and processing. How this knowledge is represented and processed in the mental lexicon has attracted considerable attention in psycholinguistic research.
To account for morphological processing mechanisms, two broad branches of theoretical models have been proposed: the decomposition model and the distributed connectionist model. The decomposition model posits that the mental lexicon contains morphologically structured representations, which enable parsers to rapidly segment derived words into stems and affixes based on morpho-orthographic structure, independently of semantic information (e.g., Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023; Grainger & Beyersmann, Reference Grainger and Beyersmann2017; Taft & Nguyen-Hoan, Reference Taft and Nguyen-Hoan2010). In contrast, the distributed connectionist model argues that morphological processing reflects emergent patterns arising from mappings between form and meaning without requiring specific morphemic representations or structural decomposition (e.g., Baayen et al., Reference Baayen, Milin, Đurđević, Hendrix and Marelli2011; Marelli et al., Reference Marelli, Amenta and Crepaldi2015; Plaut & Gonnerman, Reference Plaut and Gonnerman2000). From this perspective, morphological processing is primarily mediated by the degree of form–meaning overlap between morphologically related words.
The two morphological processing models have been tested extensively using priming paradigms (e.g., Beyersmann et al., Reference Beyersmann, Iakimova, Ziegler and Colé2014; Domínguez et al., Reference Domínguez, De Vega and Barber2004; Feldman et al., Reference Feldman, O’Connor and Moscoso2009; Jared et al., Reference Jared, Jouravlev and Joanisse2017; Lavric et al., Reference Lavric, Clapp and Rastle2007, Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025; Morris et al., Reference Morris, Frank, Grainger and Holcomb2007; Rastle et al., Reference Rastle, Davis, Marslen-Wilson and Tyler2000; Smolka et al., Reference Smolka, Gondan and Rösler2015), in which morphological processing effects are inferred from facilitated responses to target stems (e.g., kind) preceded by morphologically related primes (e.g., kindness) relative to unrelated baseline primes (e.g., pore). Using this paradigm, some studies have yielded evidence for the distributed connectionist account, demonstrating morphological priming effects that vary with the degree of form–meaning overlap from the early stages of lexical access (e.g., Feldman et al., Reference Feldman, O’Connor and Moscoso2009; Jared et al., Reference Jared, Jouravlev and Joanisse2017; Morris et al., Reference Morris, Frank, Grainger and Holcomb2007). In contrast, a larger body of empirical findings has reported a distinct morphological facilitation that exceeds the additive effects of form and meaning overlap, supporting the presence of explicit morphological representations in the mental lexicon (e.g., Beyersmann et al., Reference Beyersmann, Iakimova, Ziegler and Colé2014, 2016; Domínguez et al., Reference Domínguez, De Vega and Barber2004; Lavric et al., Reference Lavric, Clapp and Rastle2007, Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025; Rastle et al., Reference Rastle, Davis, Marslen-Wilson and Tyler2000; Smolka et al., Reference Smolka, Gondan and Rösler2015).
Specifically, evidence for the decomposition account indicates a biphasic pattern of morphological processing, which begins with morpho-orthographic segmentation followed by morpho-semantic integration (Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023; Grainger & Beyersmann, Reference Grainger and Beyersmann2017; Taft et al., Reference Taft, Li and Beyersmann2019). During the initial stage of lexical access, morpho-orthographic segmentation indiscriminately operates on any orthographic string that can be exhaustively parsed into morphemic representations, such as farmer (farm + er) and corner (corn + er), irrespective of semantic transparency (Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023; Grainger & Beyersmann, Reference Grainger and Beyersmann2017). Following morpho-orthographic segmentation, morpho-semantic integration serves to validate the meanings of decomposed stems via semantic connections between stem and whole word forms (Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023; Lavric et al., Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025; Rastle & Davis, Reference Rastle and Davis2008).
While both morpho-orthographic segmentation and morpho-semantic integration seem to be essential for derived-word processing, a substantial body of L2 research has primarily focused on the former. This line of work typically employs the masked priming paradigm, which captures automatic processes in the initial stages of visual word recognition through the use of brief priming durations (approximately 50 ms or less; Forster & Davis, Reference Forster and Davis1984). Importantly, the short prime–target interval in masked priming minimizes the involvement of semantic processing, making this paradigm particularly well suited to isolating automatic, semantically blind morpho-orthographic segmentation during derived-word processing. However, given that lexical processing unfolds across distinct temporal stages and that morpho-orthographic segmentation constitutes only one component of derived-word processing (Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023; Lee et al., Reference Lee, Maeng, Lack and Song2025). It is important to examine the temporal dynamics of derived-word processing with a focus on the biphasic pattern of morphological decomposition, encompassing both morpho-orthographic segmentation and morpho-semantic integration.
To this end, the overt priming paradigm, when combined with electroencephalography (EEG), provides a particularly effective means of capturing the temporal dynamics of morphological decomposition that characterize this biphasic pattern (Barber et al., Reference Barber, Domínguez and De Vega2002; Domínguez et al., Reference Domínguez, De Vega and Barber2004; Lavric et al., Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025). Furthermore, recent L2 studies have raised concerns about the suitability of the masked priming paradigm for L2 research (Koval, Reference Koval2025; Nakayama & Lupker, Reference Nakayama and Lupker2018), given the inconsistent results reported across previous studies (e.g., Diependaele et al., Reference Diependaele, Duñabeitia, Morris and Keuleers2011; Heyer & Clahsen, Reference Heyer and Clahsen2015). In particular, it has been argued that the brief prime durations characteristic of the masked paradigm may obscure L2 speakers’ ability to engage in morphological processing.
Against this background, the present study revisits the issue of how L2 speakers process derived words by employing event-related potential (ERP) measures in conjunction with the overt priming paradigm. Specifically, it examines whether L2 speakers exhibit the biphasic pattern of morphological decomposition, in which initial morpho-orthographic segmentation is followed by morpho-semantic integration.
1.1. L1 morphological priming in ERP
Previous masked priming studies have yielded mixed results regarding morphological processing mechanisms involved in the early stages of lexical access. Some findings provide evidence for the distributed connectionist account, reporting larger effects for genuine morphological priming (e.g., farm–farm) than for pseudo-morphological priming (e.g., corner–corn) under extremely short priming durations (e.g., Feldman et al., Reference Feldman, O’Connor and Moscoso2009; Jared et al., Reference Jared, Jouravlev and Joanisse2017; Morris et al., Reference Morris, Frank, Grainger and Holcomb2007). Such graded effects of masked morphological priming suggest that morphological relatedness arises from continuous mappings between form and meaning, rather than from discrete morphemic representations. In contrast, another line of masked priming studies suggests that morphological priming effects in L1 cannot be fully explained by form–meaning overlap alone. These studies report comparable effects for genuine morphological and pseudo-morphological priming, a pattern interpreted as evidence for semantically blind morpho-orthographic segmentation and for morphological structure explicitly represented in the mental lexicon (e.g., Beyersmann et al., 2016; Lavric et al., Reference Lavric, Clapp and Rastle2007; Rastle et al., Reference Rastle, Davis, Marslen-Wilson and Tyler2000; Taft & Nguyen-Hoan, Reference Taft and Nguyen-Hoan2010).
While an ongoing debate remains between the decomposition model and the distributed connectionist model, ERP studies using the overt priming paradigm have consistently provided supporting evidence for morphologically specialized mechanisms that cannot be explained by simple form–meaning overlap effects. Specifically, these studies have reported a biphasic pattern reflecting early morpho-orthographic segmentation followed by later morpho-semantic integration (Barber et al., Reference Barber, Domínguez and De Vega2002; Beyersmann et al., Reference Beyersmann, Iakimova, Ziegler and Colé2014;Domínguez et al., Reference Domínguez, De Vega and Barber2004; Lavric et al., Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025). Despite the greater involvement of semantic information with extended priming durations, the overt priming ERP studies suggest that the initial stage of derived-word processing operates through semantically blind morpho-orthographic segmentation. This has been supported by comparable negativity attenuation across genuine morphological (e.g., farmer–farm) and pseudo-morphological (e.g., corner–corn) priming conditions, both of which elicit stronger effects than the purely orthographic condition, in the early N400 window, where semantic influences are minimal (Domínguez et al., Reference Domínguez, De Vega and Barber2004;Lavric et al., Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025). Since genuine and pseudo-morphological pairs share similar morpho-orthographic structure but differ in semantic transparency, their comparable priming effects indicate that this early segmentation process operates independently of meaning. More importantly, the stronger effects observed for both conditions relative to orthographic pairs underscore the unique contribution of morphological structure beyond surface-form overlap.
Moreover, late morpho-semantic integration reflects additional structural mechanisms beyond semantic association, driven by interactive feedback between stem and derived-word representations that amplifies semantic facilitation (Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023). ERP evidence for this process comes from overt priming studies showing neural responses corresponding to morpho-semantic integration in the late N400 window, during which the semantic information of a morpho-orthographically decomposed stem is validated against the whole-word meaning of the morphologically complex word (Lavric et al., Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025). When morphological and purely semantic priming conditions are matched for semantic relatedness, morphological pairs consistently elicit stronger negativity attenuation within 380–500 ms, whereas semantic pairs produce only weak or marginal priming effects (Beyersmann et al., Reference Beyersmann, Iakimova, Ziegler and Colé2014; Domínguez et al., Reference Domínguez, De Vega and Barber2004; Lee et al., Reference Lee, Maeng, Lack and Song2025). These findings indicate that morpho-semantic integration cannot be explained by semantic overlap alone, highlighting the unique contribution of morphological structure even when semantic relatedness is fully controlled between the morphological and purely semantic priming conditions (Beyersmann et al., Reference Beyersmann, Iakimova, Ziegler and Colé2014; Domínguez et al., Reference Domínguez, De Vega and Barber2004; Lee et al., Reference Lee, Maeng, Lack and Song2025).
In brief, numerous L1 ERP studies using the overt priming paradigm support the decomposition account, demonstrating that morphological priming effects in L1 cannot be explained by form–meaning overlap alone but instead reflect morphology-specific mechanisms that operate at both structural and semantic levels. Notably, ERP evidence further shows that derived-word processing unfolds through a biphasic morphological mechanism, beginning with morpho-orthographic segmentation and continuing into morpho-semantic integration.
Beyond the biphasic pattern of morphological decomposition, another notable finding from overt priming ERP research concerns lateral inhibition effects observed for purely orthographic priming conditions in the late negativity (LN) window under overt priming (Barber et al., Reference Barber, Domínguez and De Vega2002; Domínguez et al., Reference Domínguez, De Vega and Barber2004; Lee et al., Reference Lee, Maeng, Lack and Song2025). In this time window, orthographic primes elicit increased negativity, reflecting intensified lateral inhibition that suppresses irrelevant lexical competitors during word recognition (McClelland & Rumelhart, Reference McClelland and Rumelhart1981). Crucially, the emergence of such inhibition after the late N400, the time frame associated with semantic integration, suggests that semantic mismatch amplifies competition, and thereby lateral inhibition, among orthographically co-activated words once primes and targets are semantically integrated. In contrast, inhibitory LN effects do not arise for morphological priming, because morpho-orthographic structure and morpho-semantic connections jointly mitigate lexical competition through shared morphological representations (Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023; Grainger & Beyersmann, Reference Grainger and Beyersmann2017; Lee et al., Reference Lee, Maeng, Lack and Song2025).
1.2. Morphological decomposition in L2
Although L1 research seems to suggest that morphological decomposition proceeds through a biphasic mechanism, in which morpho-orthographic decomposition and morpho-semantic integration operate sequentially, L2 research has largely concentrated on examining the operability of morpho-orthographic segmentation during the earliest stages of lexical access, leaving the morpho-semantic stage relatively underexplored (Ciaccio & Clahsen, Reference Ciaccio and Clahsen2020; Ciaccio & Jacob, Reference Ciaccio and Jacob2019; Diependaele et al., Reference Diependaele, Duñabeitia, Morris and Keuleers2011; Heyer & Clahsen, Reference Heyer and Clahsen2015; Jacob et al., Reference Jacob, Heyer and Veríssimo2018; Kirkici & Clahsen, Reference Kirkici and Clahsen2013; Silva & Clahsen, Reference Silva and Clahsen2008). This limited focus can be largely attributed to the predominant use of the masked priming paradigm in L2 research, which prevents conscious processing of primes and thus minimizes semantic influence. Within this methodological framework, however, findings have been somewhat mixed. Most of the previous studies have shown that native speakers typically exhibit null or even inhibitory effects from orthographic primes alongside robust facilitation from morphological primes, indicating that the facilitation effect in the morphological condition arises from morpho-orthographic decomposition rather than from nonmorphological form similarity (Ciaccio & Clahsen, Reference Ciaccio and Clahsen2020; Ciaccio & Jacob, Reference Ciaccio and Jacob2019; Diependaele et al., Reference Diependaele, Duñabeitia, Morris and Keuleers2011; Heyer & Clahsen, Reference Heyer and Clahsen2015; Jacob et al., Reference Jacob, Heyer and Veríssimo2018; Kirkici & Clahsen, Reference Kirkici and Clahsen2013). In contrast, these studies have yielded inconsistent results in the orthographic condition for L2 speakers, while consistently reporting reliable facilitation in the morphological condition. Although some studies have reported native-like patterns, showing significantly weaker priming effects in the orthographic than in the morphological condition (e.g., Diependaele et al., Reference Diependaele, Duñabeitia, Morris and Keuleers2011; Li et al., Reference Li, Taft and Xu2017), others have found no difference between the two, interpreting this result as evidence of L2 speakers’ oversensitivity to form overlap (e.g., Heyer & Clahsen, Reference Heyer and Clahsen2015).
Some researchers have suggested that the inconsistent findings from these prior L2 studies may reflect differences in the stages of lexical processing captured by the masked priming paradigm L1 and L2 speakers (Koval, Reference Koval2025; Nakayama & Lupker, Reference Nakayama and Lupker2018). As indicated by L1 ERP research on orthographic priming (Lee et al., Reference Lee, Maeng, Lack and Song2025; Massol et al., Reference Massol, Midgley, Holcomb and Grainger2010, Reference Massol, Molinaro and Carreiras2015; Meade et al., Reference Meade, Grainger, Midgley, Emmorey and Holcomb2018), visual word recognition is initially facilitated through the sublexical co-activation of lexical representations that share letter sequences. However, this initial sublexical facilitation diminishes once the corresponding lexical-orthographic representations become activated, as lateral inhibition among these orthographically similar representations begins to operate. Under masked priming, although L1 lexical processing may initially undergo facilitation at the sublexical level, efficient access to lexical-orthographic representations appears to enable the rapid engagement of lateral inhibition during orthographic priming, resulting in null or inhibitory priming effects in this condition. Alongside lateral inhibition, L1 speakers are assumed to reliably activate morpho-orthographic representations even within the brief prime durations typical of masked priming (e.g., Lavric et al., Reference Lavric, Clapp and Rastle2007; Rastle et al., Reference Rastle, Davis, Marslen-Wilson and Tyler2000). Since morpho-orthographic structure in prime words alleviates lexical competition among co-activated lexical-orthographic representations (Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023; Grainger & Beyersmann, Reference Grainger and Beyersmann2017), L1 speakers consistently exhibit facilitatory priming effects in the morphological condition under masked priming.
In contrast, the short prime duration used in masked priming may constrain L2 speakers’ access to morpho-orthographic and lexical-orthographic representations, given their relatively lower proficiency and reduced efficiency in visual word recognition. As a result, even if morphological representations are available in the L2 mental lexicon, L2 speakers may reliably access only sublexical letter information under masked priming. This limited access, in turn, leads to comparable facilitation in both morphological and orthographic conditions, an effect primarily driven by surface-form overlap rather than genuine morphological decomposition. In sum, L1 and L2 systems may share similar mechanisms of visual word recognition across sublexical and lexical levels, but reduced processing efficiency in L2 may hinder the engagement of morpho-orthographic and lateral inhibition processes within the brief priming duration provided by the masked priming paradigm (Koval, Reference Koval2025; Nakayama & Lupker, Reference Nakayama and Lupker2018).
The predominant use of the masked priming paradigm in L2 research has also constrained the scope of L2 morphological investigation by capturing only the earliest, form-based stages of lexical access. Consequently, morpho-semantic processing has remained relatively underexplored, as the masked priming paradigm is ill-suited for examining later morpho-semantic integration that typically emerges when primes are fully visible (Lavric et al., Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025; Rastle et al., Reference Rastle, Davis, Marslen-Wilson and Tyler2000). However, morpho-semantic integration constitutes an essential stage in deriving the meaning of morphologically complex words, involving the validation of the decomposed stem’s meaning against that of the derived word (Lavric et al., Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025). To address this limitation, paradigms that allow full conscious perception of primes, such as overt priming with longer prime exposure, are required to capture how morpho-orthographic analysis transitions into morpho-semantic integration in L2.
To our knowledge, a single behavioral study has employed overt priming with an SOA of 200 ms to examine both morpho-orthographic and morpho-semantic processing of derived words in L2 (Koval, Reference Koval2025). By comparing morphological, pseudo-morphological, orthographic and semantic priming conditions, Koval (Reference Koval2025) reported graded facilitation effects of morphological and pseudo-morphological primes, with the former producing numerically stronger facilitation, while no facilitation was observed for purely orthographic primes. Importantly, the emergence of pseudo-morphological priming in the absence of purely orthographic priming suggests that L2 speakers can effectively engage in morpho-orthographic analysis and lexical-level lateral inhibition when sufficient processing time is available (Nakayama & Lupker, Reference Nakayama and Lupker2018). Moreover, these findings were interpreted as evidence for the parallel activation of morpho-orthographic and morpho-semantic processes in L2, such that morpho-semantic processing operates directly for genuinely morphological relations, whereas pseudo-morphological relations rely primarily on morpho-orthographic segmentation (Diependaele et al., Reference Diependaele, Duñabeitia, Morris and Keuleers2011).
While Koval (Reference Koval2025) provides crucial insight into the operability of morpho-orthographic segmentation in L2, one limitation of this study concerns the use of behavioral measures, which may not have been sensitive enough to dissociate temporally distinct morpho-orthographic and morpho-semantic processes. Because behavioral measures reflect the cumulative outcome of multiple processing stages, the stronger morphological priming relative to pseudo-morphological priming observed in this study may have resulted from the combined influence of morpho-orthographic segmentation and morpho-semantic integration during derived-word processing in L2. Furthermore, as Koval (Reference Koval2025) found that genuine morphological priming effects were only numerically, but not statistically, different from purely semantic priming effects, it remains unclear whether the study truly captured morpho-semantic integration, as claimed by the author.
In summary, previous L2 research on morphological decomposition has primarily focused on early morpho-orthographic segmentation using masked priming, leaving the morpho-semantic stage largely unexplored. Although Koval (Reference Koval2025) provided initial evidence for L2 speakers’ ability to engage in both morpho-orthographic and morpho-semantic processing under fully visible priming, the reliance on behavioral measures limited temporal resolution and interpretability. To obtain a more comprehensive understanding of L2 morphological decomposition, it is therefore essential to employ time-sensitive methods such as ERP in conjunction with overt priming, which can disentangle the sequential contributions of morpho-orthographic and morpho-semantic processes.
1.3. Implications for L2 theoretical models
Nontarget-like morphological processing patterns in L2, characterized by equivalent effects of morphological and orthographic priming observed in some studies, have often been interpreted as evidence for the Shallow Structure Hypothesis (SSH), which posits that the L2 system tends to underuse structural information due to its heavy reliance on non-structural resources (Clahsen & Felser, Reference Clahsen and Felser2006, Reference Clahsen and Felser2018). While the SSH was originally proposed to account for nontarget-like structural parsing patterns in L2 sentence processing, it has been extended to the morphological domain to explain differential patterns observed in the L2 processing of inflected and derived words (Jacob et al., Reference Jacob, Heyer and Veríssimo2018; Kirkici & Clahsen, Reference Kirkici and Clahsen2013; Silva & Clahsen, Reference Silva and Clahsen2008). Specifically, the SSH proposes that the L2 system relies heavily on whole-word representations in lexical storage, thus reducing the involvement of structural parsing mechanisms (Jacob et al., Reference Jacob, Heyer and Veríssimo2018; Kirkici & Clahsen, Reference Kirkici and Clahsen2013). Assuming that inflected words (particularly regular ones) are typically not stored as whole words in the mental lexicon (e.g., Anderson, Reference Anderson1992), the SSH argues that the strong reliance on lexical storage in L2 may hinder morphological decomposition in the processing of inflectional morphology. Furthermore, the SSH interprets robust priming effects observed for derived words as lexically mediated, reflecting facilitation arising from shared lexemes, or form–meaning overlap, between primes and targets whose whole-word representations exist in the lexicon (Jacob et al., Reference Jacob, Heyer and Veríssimo2018; Kirkici & Clahsen, Reference Kirkici and Clahsen2013). This view is further substantiated by findings that L2 speakers exhibit comparable facilitation for morphological and orthographic primes, whereas L1 speakers show facilitation exclusively for morphological primes, suggesting that L2 morphological processing is largely driven by surface-form similarity rather than morphological structure.
Under the SSH framework, it may therefore be generally challenging for L2 speakers to analyze the morphological structure of derived words. Instead, L2 speakers are expected to rely more heavily on non-structural information, such as surface form overlap without engaging the morphological information associated with derived words, as has been reported in several masked priming studies (e.g., Heyer & Clahsen, Reference Heyer and Clahsen2015; Li & Taft, Reference Li and Taft2020; Taft & Li, Reference Taft and Li2021).
However, an alternative possibility is that non-shallow, full parsing may not be particularly difficult in L2 processing of derived words, as derivational affixes are considered semantically far more salient than inflectional ones. Notably, derivational affixes often make a clear contribution to the meaning of the derived word, for instance, the suffix -able, which denotes ‘being able to VERB’ (Jacob et al., Reference Jacob, Heyer and Veríssimo2018). This stands in sharp contrast to inflectional affixes, which primarily mark abstract syntactic features that are generally known to be difficult to acquire in L2 learning (e.g., Lardiere, Reference Lardiere, Liceras, Zobl and Goodluck2008). The semantic salience of derivational affixes may therefore enable L2 speakers to identify these units more easily during derived-word processing, facilitating the establishment of morphological representations in the mental lexicon. This may be particularly true for bimorphemic words, which exhibit a high degree of structural simplicity. As discussed earlier, target-like processing of bimorphemic words has been reported in some studies (Ciaccio & Clahsen, Reference Ciaccio and Clahsen2020; Li et al., Reference Li, Taft and Xu2017), whereas the structural analysis of trimorphemic constructions is known to be more cognitively demanding for L2 speakers (Jafari et al., Reference Jafari, Marefat and Solaimani2025; Song et al., Reference Song, Do, Thompson, Waegemaekers and Lee2020).
Crucially, contrary to the previously held common understanding, assuming full structural parsing in L2 does not contradict the SSH. In its refined formulation, the SSH proposes that the operability of L2 structural processing is not an all-or-nothing phenomenon, but rather depends on multiple factors such as target structure, proficiency, the nature of the task, etc. (Clahsen & Felser, Reference Clahsen and Felser2018). This view suggests that researchers testing the SSH should focus on the conditions under which L2 speakers are more likely to engage in full parsing, rather than generalizing about its presence or absence across the board. For example, although full parsing of phrases that involve complex syntactic configurations is likely to require exceptionally high proficiency and extensive L2 experience, structural analysis of derived words with simple bimorphemic structures may be attainable even for less proficient L2 speakers, given their structural simplicity and semantic transparency. Therefore, for a particular target structure, instead of positing a general inoperability of structural processing in L2 across different scenarios, it would be more appropriate to identify the specific linguistic and experimental conditions under which target-like parsing is more or less likely to occur.
2. Present study
The present study aims to investigate whether L2 processing of derived words involves a target-like biphasic pattern of morphological decomposition, consisting of morpho-orthographic segmentation and morpho-semantic integration, both of which are uniquely morphological in nature, rather than merely orthographic or semantic. Specifically, to examine these morphological processes, this study employs a 300 ms SOA and compares morphological priming (kindness–kind) with orthographic priming (surgeon–surge) and semantic priming (doctor–nurse) across ERP components that index early and late stages of lexical access. Building on a recent L1 overt priming ERP study (Lee et al., Reference Lee, Maeng, Lack and Song2025), the present study examines three key ERP components: (1) the early N400, associated with initial morpho-orthographic segmentation, (2) the late N400, linked to morpho-semantic integration at later stages of lexical access and (3) the LN, reflecting lateral inhibition. ERP data from L2 speakers were compared with the L1 data reported in Lee et al. (Reference Lee, Maeng, Lack and Song2025).
Lee et al. (Reference Lee, Maeng, Lack and Song2025) found distinct neural responses for morpho-orthographic and morpho-semantic priming, both of which elicited unique patterns compared to purely orthographic and semantic priming across the early and late N400 windows. In the early N400 window, morphological priming elicited a more rapid and robust negativity attenuation than orthographic priming, with only minimal semantic facilitation observed, reflecting the early engagement of morpho-orthographic segmentation. Subsequently, in the late N400 window, the morphological priming effect persisted and became more pronounced, indicating morpho-semantic integration via interactive semantic feedback between a derived word and its stem. In addition, orthographic priming produced a biphasic pattern, characterized by an initial attenuation of negativity in the early N400 window, followed by an inhibitory effect in the LN window, reflecting increased lexical competition. Meanwhile, semantic priming, observed in the late N400 window, exhibited a delayed onset and weaker effects compared to morphological and orthographic priming, underscoring the relative difficulty of integrating semantically related but morphologically unrelated words.
Drawing on the main findings of Lee et al. (Reference Lee, Maeng, Lack and Song2025), the present study examines whether similar patterns emerge in L2 processing: (1) whether morpho-orthographic priming elicits greater attenuation of negativity than orthographic priming in the early N400 window, (2) whether morpho-semantic priming produces greater attenuation compared to semantic priming in the late N400 window and (3) whether orthographic priming induces increased negativity in the LN window.
If the processing of derived words in L2 is shallow, due to a heavy reliance on non-structural information such as letter-sequence overlap (e.g., Heyer & Clahsen, Reference Heyer and Clahsen2015), morphological and orthographic primes are expected to elicit comparable negativity attenuations and latency patterns in the early N400 window among L2 speakers. Furthermore, in the late N400 window, morphological and semantic primes may yield comparable reductions in negativity, as facilitation from morphological primes would be mediated through general semantic associations rather than distinct morpho-semantic mechanisms. In contrast, if morphological parsing is available in L2 processing, we would expect to observe, in the early N400, where semantic influences are minimal, greater attenuation of negativity in the morphological condition than in the orthographic condition, reflecting the involvement of morpho-orthographic segmentation. Moreover, in the late N400, L2 speakers are expected to display larger negativity attenuation in the morphological condition than in the semantic condition, reflecting morpho-semantic integration between the decomposed stem and the derived word.
Finally, two predictions can be made regarding lateral inhibition effects for the orthographic condition in the LN. First, L2 speakers may show increased negativity, reflecting the engagement of lateral inhibition under fully visible priming, which provides sufficient processing time (e.g., Nakayama & Lupker, Reference Nakayama and Lupker2018). Alternatively, they may fail to exhibit such increased negativity due to weakly encoded form–meaning mappings in L2 lexical representations (Gollan et al., Reference Gollan, Forster and Frost1997; Jiang, Reference Jiang1999; Kroll et al., Reference Kroll, Van Hell, Tokowicz and Green2010; Xia & Andrews, Reference Xia and Andrews2015). Prior L1 evidence indicates that lateral inhibition tends to intensify once semantic mismatches among orthographically co-activated representations are fully identified (e.g., Barber et al., Reference Barber, Domínguez and De Vega2002; Lee et al., Reference Lee, Maeng, Lack and Song2025). The weaker form–meaning mappings in L2 may reduce the efficiency of detecting prime–target semantic mismatches in the orthographic condition, thereby preventing the amplification of lateral inhibition. Consequently, increased LN negativity may not arise for the orthographic condition in L2.
3. Methods
3.1. Participants
Thirty advanced Cantonese-speaking learners of English (mean age = 19 years, SD = 1.69, range 18–24; 19 females) participated in the experiment. They were compared with 29 native English speakers from Lee et al. (Reference Lee, Maeng, Lack and Song2025) (mean age = 22 years, SD = 4.30, range 18–35; 15 females). The native speakers were expatriates living in Hong Kong, all of whom identified English as their sole dominant language. All participants were recruited from a university in Hong Kong. They all reported being right-handed and provided informed consent before participation. The study’s protocol and consent forms were approved by the university’s Human Research Ethics Committee.
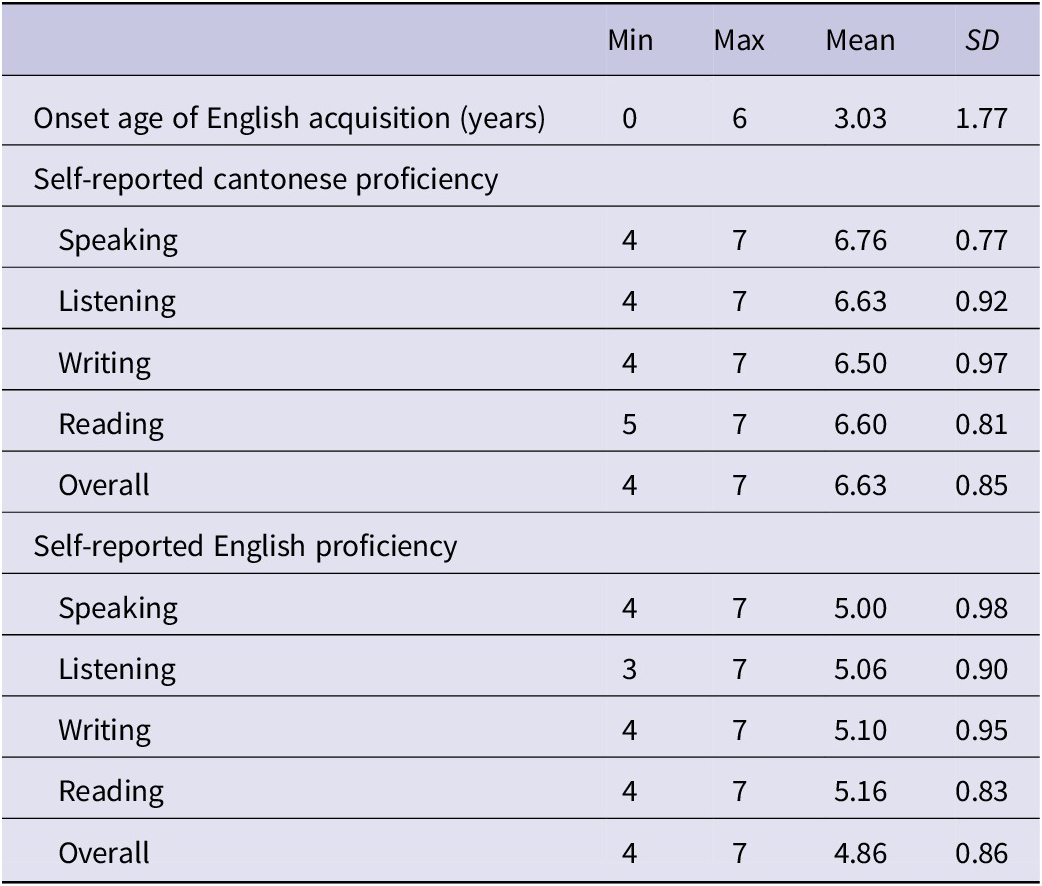
All our L2 speakers reported growing up in Hong Kong with Cantonese as their native and dominant language, as corroborated by their 7-point self-assessment scores for Cantonese (6.6) and English (4.8). Despite the early onset of exposure to English (mean age of acquisition = 3.03, SD = 1.77, range = 0–6 years), as detailed in Table 1, most L2 participants (28 speakers, 93.3%) reported predominantly using Cantonese at home. Moreover, 13 L2 speakers (43.3%) reported using mostly Cantonese at school, with only 7 L2 speakers (23.3%) reporting using equal amounts of Cantonese and English in the school setting. These language background reports indicate that our L2 participants had relatively limited exposure to and use of English compared to Cantonese.
L2 English speakers (n = 30) background information
To control for English proficiency, we strictly recruited L2 speakers who achieved a score of 5 on the Hong Kong Diploma of Secondary Education (HKDSE) English exam. According to the Hong Kong Examinations and Assessment Authority, a score of 5 on the HKDSE (henceforth DSE) is achieved by approximately the top 10% of test-takers annually, which is equivalent to an overall band of 7.55 on the IELTS exam (HKEAA, 2025). We consider these speakers to have a high level of L2 proficiency. The two highest scores, 5* (equivalent to IELTS 7.86) and 5** (equivalent to IELTS 8.22), representing approximately the top 3% and top 1% of scores, respectively, were deliberately excluded, as they might be exceptional language learners and therefore do not represent typical L2 learners.
3.2. Materials
The same stimulus set from Lee et al. (Reference Lee, Maeng, Lack and Song2025) was utilized in this study. The experiment included three prime conditions – morphological, semantic and orthographic – each comprising 80 critical prime–target sets, resulting in a total of 240 critical sets across all conditions. Each set consisted of a target word paired with two types of prime: one related to the target either morphologically, semantically, or orthographically (depending on the condition) and one unrelated prime. All words were selected from the CELEX lexical database (Baayen et al., Reference Baayen, Piepenbrock and Gulikers1995). It is important to note that the present study focuses only on genuine morphological priming, while several studies have included the pseudo-morphological priming condition to investigate morpho-orthographic processing. Prior L1 studies using the overt priming paradigm in combination with ERP measures, which provide high temporal resolution, have effectively distinguished distinct neural signals for initial morpho-orthographic segmentation and subsequent morpho-semantic integration during the processing of semantically transparent morphological primes (Beyersmann et al., Reference Beyersmann, Iakimova, Ziegler and Colé2014; Lee et al., Reference Lee, Maeng, Lack and Song2025; Smolka et al., Reference Smolka, Gondan and Rösler2015). Following the same experimental paradigm, the present study explored whether L2 speakers demonstrate the biphasic pattern of initial morpho-orthographic segmentation followed by morpho-semantic integration during derived-word processing.
In the Morphological condition, all related primes (e.g., swiftly–swift) were derived words constructed with a target stem and a level-two suffix, which is semantically more transparent compared to level-one affixes (e.g., -ity, in-) and phonologically neutral, not causing any phonological change to the base word (Hay & Plag, Reference Hay and Plag2004; Song et al., Reference Song, Do, Thompson, Waegemaekers and Lee2020). Seven different affixes were used to create morphologically related primes: -able, -er, -ful, -ize, -less, -ly and -ness. Each affix occurred 11 or 12 times. The original form of the target stem was retained in the derived word prime, except for five pairs (6.25%) containing base allomorphy, which has minimal impact on morphological processing (Feldman et al., Reference Feldman, Kosti, Basnight-Brown, Urevi and Pastizzo2010; Morris & Stockall, Reference Morris and Stockall2012). In the Semantic condition, 40 related prime–target pairs were synonyms (e.g., explode–burst) and the remaining 40 were semantically associated (e.g., doctor–nurse), following previous studies (e.g., Ciaccio & Jacob, Reference Ciaccio and Jacob2019; Jacob et al., Reference Jacob, Heyer and Veríssimo2018). All related primes were monomorphemic words and were selected from the Oxford English Dictionary and previous morphological priming studies (e.g., Kielar & Joanisse, Reference Kielar and Joanisse2011; Lavric et al., Reference Lavric, Rastle and Clapp2011; Morris et al., Reference Morris, Porter, Grainger and Holcomb2011).
In the Orthographic condition, the related primes were monomorphemic words that orthographically overlap with their targets but without any semantic or morphological relations (e.g., surgeon–surge). In the Orthographic condition, orthographic overlap occurred word-initially in all except five pairs (6.25%) that were word-final/middle overlap pairs. These five pairs were included in our stimuli set due to limited options; however, previous priming research suggests that orthographic overlap positions do not affect either native speakers or L2 learners’ processing (Ciaccio & Clahsen, Reference Ciaccio and Clahsen2020; Ciaccio & Jacob, Reference Ciaccio and Jacob2019; Jiang & Wu, Reference Jiang and Wu2022). Lee et al. (Reference Lee, Maeng, Lack and Song2025) validated that these five pairs do not distort the overall findings by comparing the statistical analysis results, including and excluding them. Additionally, the degree of orthographic overlap was matched between related prime–target pairs in the Morphological condition (62.1%; 4.92 of 7.92 letters) and the Orthographic condition (61.3%; 3.87 of 6.45 letters), t = 0.65, p = .515.
We attempted to match the related primes in the three prime conditions as closely as possible for length and CELEX log frequency, as Table 2 shows. However, pairwise comparisons revealed that related primes in the Orthographic condition were significantly shorter compared to those in the Morphological, t(237) = −6.40, p < .001 and Semantic, t(237) = 4.77, p < .001, conditions. Specifically, the primes in the Morphological condition were on average approximately 1.5 letters longer than those in the Orthographic condition. This length difference was inevitable as some of the attached suffixes in the Morphological condition were three or four letters long (e.g., -ize and -ness). Moreover, the approximately 1.1 letter difference between the Orthographic and Semantic conditions resulted from the selection of relatively long monomorphemic words as our orthographic primes to create a condition where the target is embedded within the prime.
Mean frequencies and lengths of primes and targets

Note: M = Morphological, S = Semantic, O = Orthographic, R = Related, U = Unrelated.
Regarding frequency, pairwise comparisons revealed that primes in the Morphological condition have significantly lower log frequencies than those in the Semantic condition, t(237) = −3.49, p = .002. This difference was unavoidable as all our morphological primes were bimorphemic while all our semantic primes were monomorphemic. Our analysis of the CELEX database confirmed that bi-morphemic words have significantly lower log frequencies than mono-morphemic words, t(43833) = 15.734, p < .001. Furthermore, in each prime condition, unrelated primes were matched to their related prime counterparts in both length and log frequency, as confirmed by pairwise comparisons.
The target words used in the three prime conditions were closely matched for CELEX log frequency and length (i.e., the number of letters). While there was no difference in the log frequency of targets among the three prime conditions, pairwise comparisons revealed that the targets in the Orthographic condition were, on average, approximately one letter shorter than the targets in the Semantic condition, t = −5.70, p < .001, and the Morphological condition, t = −5.36, p < .001. Although the targets in the Orthographic condition were shorter than those in the Morphological condition, the degree of prime–target orthographic overlap was comparable between the two conditions.
The prime–target semantic relatedness in the three prime conditions (see Table 3) was manipulated by using Word2Vec (Mikolov et al., Reference Mikolov, Sutskever, Chen, Corrado and Dean2013), a computational model that predicts the semantic relatedness of a pair of words using a Skip-gram language model. Pairwise comparisons confirmed that the prime–target semantic relatedness of the Orthographic condition was significantly lower than that of the Morphological condition, t(237) = −12.40, p < .001, and the Semantic condition, t(237) = −13.54, p < .001. The Morphological and Semantic condition pairs did not differ significantly in their word2vec scores, t(236) = −1.27, p = .414.
Mean word2vec scores for related and unrelated pairs in all prime conditions

Additionally, the study included 240 filler prime–target pairs. These filler pairs consisted of real-word primes paired with nonword targets (e.g., axis–berm), and the filler targets were matched in length to the critical targets across the three prime conditions. Eight stimulus lists were created, each consisting of 40 related and 40 unrelated prime–target pairs for each of the three prime conditions, in addition to 240 filler pairs. The lists were counterbalanced using a Latin square design to ensure that each participant saw all 80 critical target words in each prime condition, with only half of them occurring with a related prime word.
3.3. Procedure
Upon arrival, participants were asked to fill out a consent form and a language background questionnaire. All participants were seated 1 m away from a 24-inch, 60 Hz LED monitor in a reclining chair, and the experiment was conducted in a dimly lit environment. The experiment was built on Superlab version 6.3, an experiment-building software developed by Cedrus (Haxby et al., Reference Haxby, Parasuraman and Lalonde1993). During the experiment, participants were asked to remain focused and relaxed while minimizing blinking and any other movements. Each participant performed a lexical decision task on visually presented words, responding with two buttons on a game controller assigned to “word” (dominant right hand) and “nonword” (non-dominant left hand) responses. They were asked to respond as quickly and accurately as possible.
The testing session began after participants completed a practice session consisting of 20 prime–target pairs and confirmed their understanding of the procedure. Each trial began with a fixation cross (+) displayed for 1000 ms, followed by a prime word for 200 ms, a blank screen for 100 ms and the target word for 1000 ms (resulting in a stimulus-onset asynchrony of 300 ms). Primes were presented in uppercase using a 22-point Sans Serif font, while targets were presented in lowercase with the same font but at a 26-point size. A total of 480 trials were randomly presented to participants across three blocks, each lasting approximately 15 minutes. Two breaks occurred after completing 1/3 (160 trials) and 2/3 (320 trials) of the total trials, respectively. Overall, each experiment session took approximately 90 minutes, including experimental setup and C-tests.
3.4. EEG recording and preprocessing
EEG data were recorded with 32-channel Neuroscan Quik-Caps Neo Net (AgCl) and a SYNAMPS amplifier, using the Neuroscan software Curry 8. The electrodes on these caps were positioned according to the 10–20 array, and the reference electrode was placed between the Cz and CPz electrodes. Two electrodes placed near the canthi measured horizontal eye movements, and two electrodes above and below the left eye measured vertical eye movements. Impedances were kept below 5 kΩ.
Preprocessing procedures were+ performed using EEGLab version 9.0.0 (Delorme & Makeig, Reference Delorme and Makeig2004) and ERPLAB (Lopez-Calderon & Luck, Reference Lopez-Calderon and Luck2014) according to the preprocessing pipeline from Luck (Reference Luck2022). Continuous EEG data were down-sampled from 1000 Hz to 250 Hz offline, and a bandpass filter between 0.1 and 30 Hz (half-amplitude cut-off, 12 dB/oct roll-off) was applied using the non-causal Butterworth impulse response function. After cleaning line noise at 50 Hz, a bandpass filter between 1 and 30 Hz (half-amplitude cut-off, 48 dB/oct roll-off) was applied again. Breaks and bad electrodes were excluded from the filtered data, and then a threshold criterion of 500 μV was applied to detect and remove eye blinks and other artifacts before running independent component analysis (ICA). ICA components associated with eye blinks and eye movements were then removed, the data were re-referenced to the averaged mastoids (M1/M2), and all electrodes removed before ICA were interpolated.
Epochs were computed from 200 ms before to 1000 ms after the target word onset. Baseline correction was conducted using the −200–0 ms pre-stimulus interval. During ocular artifact rejection, epochs exceeding 50 μV in the vertical eye channels and epochs exceeding 32 μV in the horizontal eye channels were excluded. Then, epochs in which any electrode exceeded the voltage threshold of ±150 μV were excluded using the absolute voltage function and the peak-to-peak function. 17% of trials were removed. Mean amplitudes were calculated by averaging the remaining trials for each participant, electrode and prime condition.
3.5. Data analysis
3.5.1. Behavioral analysis
For behavioral analysis, 0.2% of responses were removed from the data for being longer than 1000 ms. Additionally, outliers exceeding ±3 standard deviations from the group means were further removed, resulting in a total of 1.2% of trials removed. We removed a total of 16 pseudo-morphological and semantically opaque morphological pairs from the orthographic priming analysis,Footnote 1 as a reviewer raised concerns about potential confounding effects arising from pseudo-morphological pairs (e.g., legal–leg) and semantically opaque morphological pairs (e.g., portal–port) in the orthographic priming condition. Although the reviewer initially suggested excluding pairs with primes ending in -le, we decided not to remove these three items (e.g., buckle–buck, nettle–net, nipple–nip) from the analysis because -le no longer functions as a productive suffix for free stems in contemporary English (Booij & Audring, Reference Booij, Audring and Booij2018). The same exclusions were applied to the orthographic priming analysis of the ERP data. For statistical analysis, three-way ANOVAs were conducted using the aov function in R 4.3.0 (R Core Team, 2023). RT was set as the dependent variable, with prime condition (Morphological, Semantic, Orthographic), relatedness (related, unrelated) and participant group (L1, L2) as independent variables. When significant main and interaction effects were identified, pairwise comparisons were conducted using the emmeans function from the emmeans package (Lenth, Reference Lenth2024), with Bonferroni corrections applied. As our primary interest was how priming effects (prime condition × relatedness) were modulated by group effects, our analysis focused on the interpretation of the three-way interaction among prime condition, relatedness and group. While interactions irrelevant to this focus are not reported, their details are available in the Supplementary Materials.
3.5.2. ERP analysis
Preprocessed EEG data were analyzed for the three ERP components representing morpho-orthographic priming, morpho-semantic priming and lateral orthographic inhibition: early N400 (310–410 ms), late N400 (410–500 ms) and late negativity (LN; 500–600 ms). The amplitude of each ERP component was averaged across the fronto-central (FCz and FC3, FC4), central (Cz, C3, and C4), centro-parietal (CPz, CP3, and CP4) and parietal (Pz, P3, and P4) electrodes. The selection of these time windows and electrodes was based on a prior L1 study (Lee et al., Reference Lee, Maeng, Lack and Song2025), relevant findings in the existing literature (Beyersmann et al., Reference Beyersmann, Iakimova, Ziegler and Colé2014; Domínguez et al., Reference Domínguez, De Vega and Barber2004; Grainger & Holcomb, Reference Grainger and Holcomb2009; Kielar & Joanisse, Reference Kielar and Joanisse2011; Lavric et al., Reference Lavric, Rastle and Clapp2011) and visual inspection. These time windows also align with the canonical time windows for the N400 (300–500 ms, Holcomb & Grainger, Reference Holcomb and Grainger2006; Kutas et al., Reference Kutas, Lindamood and Hillyard1984) and LN (450–700 ms, Barber et al., Reference Barber, Domínguez and De Vega2002; Domínguez et al., Reference Domínguez, De Vega and Barber2004). Consistent with the behavioral analysis, three-way ANOVAs and Bonferroni-adjusted pairwise comparisons were conducted, with mean amplitude as the dependent variable, focusing on the interpretation of the three-way interaction among prime condition, relatedness and group. Our ERP analysis is reported in the following format: (1) a descriptive analysis of our ERP waveforms; (2) presentation of statistical findings and validation of observed effects and (3) overall interpretation of our findings.
4. Results
4.1. Behavioral results
Table 4 illustrates the means and standard errors of the reaction times (RTs) for the relatedness conditions (related, unrelated) in each of the three prime conditions (Morphological, Orthographic, Semantic) for both participant groups (L1, L2). The ANOVA results revealed a significant main effect of relatedness, F(1, 13046) = 349.3, p < .001, indicating shorter overall RTs for the related condition compared to the unrelated condition across the three prime conditions (p < .001). A significant main effect of prime condition was also found, F(2, 13046) = 147.1, p < .001, suggesting reliable differences among the three prime conditions (ps < .001). Additionally, a significant main effect of group was observed, F(1, 13046) = 235.9, p < .001, with the post hoc analysis revealing that the L1 group exhibited shorter overall RTs than the L2 group (p < .001).
Mean RTs by prime condition and group

Note: M(SD) in milliseconds. ***p < .001.
More importantly, a significant interaction between prime condition and relatedness was found, F(2, 13046) = 36.7, p < .001. Pairwise comparisons revealed a significant priming effect across all three prime conditions (ps < .001), with differences in the magnitude of the effect. Specifically, the priming effect in the Morphological condition was significantly larger than those in the Orthographic and Semantic conditions (ps < .001), while the priming effects in the Orthographic and Semantic conditions did not differ significantly (p = .538). Most crucially, this interaction between prime condition and relatedness was not significantly modulated by group, F(2, 13046) = 0.1, p = .937, suggesting that the L2 speakers did not significantly differ from the L1 speakers in the relative strengths of the priming effects across the three prime conditions.
4.2. ERP results
4.2.1. Early N400 (310–410 ms)
As Figure 1A illustrates, in the L1 group, the Morphological condition displayed the earliest and least negative peak compared to the other prime conditions within the early N400 time window. This negative peak was immediately followed by a prominent drop in negativity, demonstrating the strongest attenuation of negativity induced by priming. In contrast, the Orthographic condition showed a clear but less attenuated negative peak than the Morphological condition, along with a noticeably slower onset time and a smaller subsequent decline in negativity. Additionally, the Semantic condition exhibited the least attenuated and slowest negative peak, with its amplitude closely resembling the Baseline. The L2 group replicated the overall patterns observed in the L1 group, albeit with relatively less pronounced contrasts among the three prime conditions. Similar to the L1 group, the negative peak of the Morphological condition was the most attenuated among all prime conditions, followed by the greatest drop in negativity. Although the Orthographic condition showed the second most attenuation in the L2 group after the Morphological condition, it lacked the clear negative peak and a subsequent drop in negativity seen in the Morphological condition. As in the L1 group, the Semantic condition in the L2 group exhibited the slowest and most negative peak, exhibiting amplitudes near the Baseline level.
ERP waveforms and topography plots of L1 and L2 for the Morphological (M), Orthographic (O) and Semantic (S) conditions.
Note: (A) ERP time-course of midline electrodes (FCz, Cz, CPz and Pz). The baseline condition was computed by averaging the amplitudes of the targets across the unrelated condition for the three prime conditions. (B) Topographic distribution of the priming effects (unrelated-related). Time windows for each ERP component: Early N400 (310–410 ms), Late N400 (410–500 ms) and LN (500–600 ms).

These observations were consistent with the results of our ANOVA analysis. First, a significant main effect of relatedness was found, F(1, 57) = 3403.9, p < .001, indicating less negative amplitudes for the related condition (−0.91 μV) compared to the unrelated condition (−1.41 μV) across the three prime conditions (p < .001). In addition, there was a main effect of prime condition, F(2, 57) = 984.5, p < .001, reflecting significant amplitude differences among the three prime conditions (ps < .001). The main effect of group was also significant, F(1, 57) = 266.1, p < .001. Pairwise comparisons revealed that the mean amplitude was more negative for the L2 group (−1.20 μV) than the L1 group (−1.11 μV) (p < .001).
In addition, a significant interaction between prime condition and relatedness was found, F(2, 57) = 1854.3, p < .001. Pairwise comparisons indicated a significant attenuation for related pairs in the Morphological (−1.00 μV) and Orthographic (−0.64 μV) conditions (ps < .001), with the magnitude of the priming effect being significantly larger in the Morphological condition (p < .001). In contrast, the Semantic condition exhibited a significant increase in negativity, although the magnitude was relatively small (0.16 μV; p < .001). Crucially, the interaction between prime condition and relatedness was significantly modulated by group, F(2, 57) = 36.9, p < .001. Pairwise comparisons suggested that this group difference was driven by the Semantic condition, where a significant attenuation in negativity (−0.24 μV; p < .001) was found in the L1 group, whereas a significant increase (0.55 μV; p < .001) was observed in the L2 group. Additionally, pairwise comparisons indicated that the group effect was further influenced by more pronounced attenuation in the Morphological and Orthographic conditions in the L1 group (ps < .001), although both L1 and L2 groups showed significant attenuations in these conditions (ps < .001), with greater attenuation in the Morphological condition. For a visualization of these group effects, see the topographic plots in Figure 1B.
In sum, our results demonstrate that each prime condition exhibits a unique temporal profile in the early N400 time window across both language groups. Specifically, the attenuation in the Morphological condition was most distinct and robust, showing the strongest priming effect. While the attenuation in the Orthographic condition was also evident, it was significantly weaker compared to the Morphological condition, and the temporal profiles of attenuation in these two conditions were clearly differentiated. The Semantic condition, on the other hand, was distinct from both the Morphological and the Orthographic conditions in its attenuation time course, resembling the Baseline. However, group effects were observed, as these distinctions among the three prime conditions were more pronounced in the L1 group than the L2 group. In other words, the L1 group exhibited stronger priming effects than the L2 group in the three prime conditions.
4.2.2. Late N400 (410–500 ms)
Figure 1A illustrates an intensified attenuation of the N400 in the Morphological condition for the L1 group, marked by a substantial reduction in negativity, with the peak reaching the least negative amplitude in the entire time course of the condition, followed by an increase in negativity. The Orthographic condition also showed a continued decrease in negativity, but the attenuation was notably less pronounced compared to the Morphological condition. This difference is also revealed in the topographic plots in Figure 1B. Furthermore, Figure 2 illustrates that the attenuation in the Orthographic condition rapidly diminished toward the end of the window for the L1 group, approaching the end of attenuation. In the Semantic condition, the N400 attenuation intensified during this time window, reaching its maximum positive peak and exhibiting its largest divergence from the Baseline. As in the L1 group, in the Morphological condition, the L2 group showed a more pronounced N400 attenuation compared to the previous time window (see Figure 1A). However, the L2 group exhibited notably less attenuation in the Morphological condition compared to the L1 group, as shown in Figure 1. In the Orthographic condition, the L2 group showed a continued decrease in negativity, which transitioned into an increase, similar to the pattern observed in the L1 group, as shown in Figure 1A. However, the figure also shows that the orthographic priming effect persisted longer in the L2 group than in the L1 group, extending beyond this time window. In terms of magnitude, the L2 group displayed noticeably less N400 attenuation in the Orthographic condition compared to the Morphological condition, consistent with the pattern observed in the L1 group, as depicted in Figure 1B. In the Semantic condition, the L2 group was distinguished from the L1 group, displaying much less pronounced N400 attenuation than the L1 group, without a clear peak (see Figure 1A). This group difference is further highlighted in Figure 1B.
Temporal topographic distribution of the priming effects (unrelated–related) within the range of 310–610 ms at 20 ms intervals.
Note: M = Morphological condition; O = Orthographic condition; S = Semantic condition.

These observations align with the findings from the ANOVA analysis. First, there was a significant main effect of relatedness, F(1, 56) = 29479.1, p < .001, revealing a less negative amplitude for the related condition (−0.58 μV) compared to the unrelated condition (−2.51 μV) across the three prime conditions (p < .001). The main effect of prime condition was also significant, F(2, 56) = 6136.9, p < .001, indicating significant amplitude differences among the three prime conditions (ps < .001). Additionally, a significant main effect of group was observed, F(1, 56) = 10543.3, p < .001. Pairwise comparisons revealed that the mean amplitude was more negative in the L2 group (−2.08 μV) than in the L1 group (−1.00 μV; p < .001).
These main effects also significantly interacted with one another. First, the interaction between prime condition and relatedness was significant, F(2, 56) = 3178.1, p < .001. Pairwise comparisons indicated significant attenuation of negativity in the related condition across all prime conditions, each with significantly different magnitudes (ps < .001). The strongest attenuation was observed in the Morphological condition (−3.12 μV), followed by the Orthographic condition (−1.66 μV), with the weakest attenuation in the Semantic condition (−1.02 μV). Crucially, the interaction between group, relatedness and prime condition was also significant, F(2, 56) = 39.4, p < .001. Pairwise comparisons confirmed that both language groups exhibited significant attenuation in all prime conditions (ps < .001), with each effect being significantly stronger in the L1 group than in the L2 group (ps < .001). In both groups, the Morphological condition showed the strongest N400 attenuation, followed by the Orthographic condition, and lastly the Semantic condition (ps < .001).
Taken together, our results in the late N400 window demonstrate that morphological priming further enhanced N400 attenuation, sustaining the strongest attenuation among all prime conditions in both language groups from the early N400 period. While both groups showed the same order of the prime conditions in terms of priming effect magnitude (i.e., Morphological > Orthographic > Semantic), the priming effects observed in the L2 group were significantly smaller than those in the L1 group across all prime conditions. In particular, although semantic priming became most pronounced during this time window for both groups, its emergence was much slower and minimal in the L2 group. On the other hand, the weaker orthographic priming effect lingered longer in the L2 group, whereas it was stronger but diminished more rapidly in the L1 group.
4.2.3. LN (500–600 ms)
Figures 1A and 2 depict that in the L1 group, the gradual decrease in N400 attenuation in the Morphological and Semantic conditions that began in the previous window continued until the waveforms in these conditions returned to the Baseline midway through the window. Notably, in the Orthographic condition, the waveform exhibited greater negativity than the Baseline throughout this time window, forming a distinct negative peak at approximately 520 ms (see Figure 1A). This pronounced negativity is also clearly illustrated in topographic plots in Figure 1B. As discussed earlier, similar instances of this LN, unique to the Orthographic condition, have been reported in previous L1 studies and interpreted as an inhibitory orthographic priming effect that reflects lateral inhibition (e.g., Barber et al., Reference Barber, Domínguez and De Vega2002; Domínguez et al., Reference Domínguez, De Vega and Barber2004). In the L2 group, the Morphological and Semantic conditions exhibited waveform patterns that were overall similar to those of the L1 group. As shown in Figure 1, they demonstrated a continued decline in N400 attenuation, with a slower decline in L2. However, in the Orthographic condition, the L2 group showed a waveform pattern distinct from that of the L1 group. As illustrated in Figure 1A, the L2 waveform did not show clearly greater negativity than the Baseline, instead closely resembling it and lacking the prominent negative peak observed in the L1 group. This group difference is also evident in the topographic plots in Figures 1B and 2.
These observations are supported by the ANOVA results. First, a significant main effect of relatedness was observed, F(1, 56) = 681.9, p < .001, indicating an overall reduction in negativity for the related condition (−0.48 μV) compared to the unrelated condition (−0.66 μV) across the three prime conditions (p < .001). The main effect of prime condition was also significant, F(1, 56) = 6395.2, p < .001, suggesting significant amplitude differences among the three prime conditions (ps < .001). Additionally, the main effect of group was significant, F(1, 56) = 4922.4, p < .001, with pairwise comparisons showing that the mean amplitude was more negative in the L2 group (−0.85 μV) than in the L1 group (−0.29 μV; p < .001).
The analysis further revealed a significant interaction between prime condition and relatedness, F(2, 56) = 570.7, p < .001. Pairwise comparisons showed significantly increased negativity in the related condition within the Orthographic condition, reflecting an inhibitory priming effect (0.18 μV; p < .001). On the other hand, the Morphological (−0.51 μV) and Semantic conditions (−0.19 μV) exhibited significantly attenuated negativity in the related conditions (ps < .01), reflecting facilitatory priming effects. Importantly, the interaction between prime condition and relatedness was significantly modulated by group, F(2, 56) = 297.7, p < .001. Pairwise comparisons revealed that the increased negativity observed in the Orthographic condition was primarily driven by the L1 group (0.47 μV; p < .001), as the L2 group instead showed an attenuation in negativity (−0.11 μV; p < .001). The Morphological and Semantic conditions in both groups exhibited less negativity than the Orthographic condition (ps < .001).
To summarize, in the Morphological and Semantic conditions, the L1 and L2 groups exhibited similar patterns, showing continued reduction in negativity. However, a significant group difference was observed in the Orthographic condition. The L1 group displayed a substantially increased negativity with a clear peak, which is interpreted in the current literature as an orthographic inhibition effect. In contrast, this negativity was absent in the L2 group, indicating that a significant orthographic inhibitory priming effect was found only in the L1 group.
5. Discussion
The present study employed the overt priming paradigm to examine whether L2 speakers can exhibit a target-like biphasic pattern of morphological decomposition, consisting of morpho-orthographic segmentation and morpho-semantic integration, during derived-word processing. Building on previous ERP findings on morphological decomposition in L1 (Lee et al., Reference Lee, Maeng, Lack and Song2025), this study analyzed the early and late N400 components, corresponding to morpho-orthographic and morpho-semantic processes, respectively, to examine whether L2 speakers engage morphologically specialized mechanisms at both structural and semantic levels. Additionally, the study explored the LN, a later time window associated with intensified lexical competition for orthographic priming in L1 (Lee et al., Reference Lee, Maeng, Lack and Song2025), to determine whether L2 speakers also engage target-like lateral inhibition mechanisms.
5.1 Morpho-orthographic segmentation
Our ERP analysis in the early N400 window revealed converging L1–L2 patterns, suggesting that morpho-orthographic segmentation operates similarly across language groups during the initial stages of derived-word processing, where semantic processing is minimally involved (Lavric et al., Reference Lavric, Clapp and Rastle2007, Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025; Rastle et al., Reference Rastle, Davis, Marslen-Wilson and Tyler2000). In morpho-orthographic segmentation, stem recognition is facilitated by morphological structure, through which orthographic input is decomposed into morphemic representations with minimal semantic influence (Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023).
Reflecting this facilitation of stem recognition, the Morphological condition elicited the strongest and earliest attenuation of negativity across L1 and L2 groups, surpassing the effects observed in the Orthographic and Semantic conditions. Importantly, although the Orthographic condition also elicited robust negativity attenuation for both language groups, its magnitude was significantly smaller than that of the Morphological condition, despite both conditions sharing a similar degree of form overlap. Moreover, negativity attenuation in the Semantic condition was minimal and emerged only near the end of the early N400 window across language groups, indicating that semantic processing, if involved at all, exerted only a limited influence on the priming effects in the Morphological condition. Therefore, the additional priming effects observed in the Morphological condition for L1 and L2 speakers, relative to the Orthographic condition, cannot be attributed to the semantic similarity between the morphological prime and target. Taken together, these early N400 findings suggest that L2 speakers, like native speakers, engage in morphological structure analysis during the early stages of derived-word processing.
The morphological priming effects, observed across both L1 and L2 speakers, bear on current models of lexical processing. Specifically, greater effects of morphological priming compared to orthographic priming in the absence of semantic priming highlight that morpho-orthographic segmentation in the initial stage of lexical access is not mediated by form–meaning overlap, as proposed by the distributed connectionist model (e.g., Baayen et al., Reference Baayen, Milin, Đurđević, Hendrix and Marelli2011; Marelli et al., Reference Marelli, Amenta and Crepaldi2015; Plaut & Gonnerman, Reference Plaut and Gonnerman2000). Instead, the clear difference between the Morphological and Orthographic conditions points to an early segmentation mechanism that operates on morphological structure itself, consistent with the decomposition view, which assumes that morphologically complex words are initially parsed into constituent morphemic representations stored in the mental lexicon (Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023; Grainger & Beyersmann, Reference Grainger and Beyersmann2017; Marslen-Wilson et al., 2009; Taft et al., Reference Taft, Li and Beyersmann2019).
Furthermore, our findings in the early N400 time window call for a reinterpretation of previous behavioral studies on L2-derived-word processing, which have reported nontarget-like patterns of morphological decomposition (e.g., Ciaccio & Jacob, Reference Ciaccio and Jacob2019; Heyer & Clahsen, Reference Heyer and Clahsen2015). As discussed earlier, these studies have shown that native speakers typically exhibit null or even inhibitory orthographic priming effects, indicating that surface-form similarity alone does not facilitate lexical access in L1. In contrast, L2 speakers have been found to display comparable facilitation for morphological and orthographic primes. Based on this contrast, prior research advocating nontarget-like L2 morphological processing has underscored that the facilitation observed in the morphological condition merely reflects L2 speakers’ heightened sensitivity to letter-level sequence information rather than genuine morphological parsing, suggesting that such form-based facilitation is a distinctive feature of L2 lexical processing.
However, our ERP findings challenge this interpretation by showing that the influence of sublexical form overlap during early lexical access is not unique to L2 speakers but is also evident in L1 speakers. Both groups demonstrated robust orthographic priming effects in the early N400 window, reflecting initial facilitation through sublexical co-activation of overlapping letter sequences. This pattern aligns with prior ERP research in L1, which has consistently reported a biphasic pattern of form-overlap effects, characterized by early facilitation of target processing through sublexical co-activation of shared letter sequences, followed by later lexical-level lateral inhibition that suppresses competing representations (Lee et al., Reference Lee, Maeng, Lack and Song2025; Massol et al., Reference Massol, Midgley, Holcomb and Grainger2010, Reference Massol, Molinaro and Carreiras2015; Meade et al., Reference Meade, Grainger, Midgley, Emmorey and Holcomb2018). These results suggest that early form-based facilitation is a general characteristic of the lexical processing system, which may be masked in behavioral data by the subsequent inhibitory stage (i.e., the N400 or LN time window) due to the lack of temporal resolution.
Given this, the previous behavioral findings interpreted as evidence of shallow lexical parsing in L2 may instead reflect a methodological artifact of masked priming, which may capture different processing stages in L1 and L2. Under such brief prime exposure, L2 speakers may primarily access sublexical letter information, without fully activating higher-level morpho-orthographic and lexical-orthographic representations, thus resulting in comparable priming effects for morphological and orthographic primes. By contrast, L1 speakers reliably access higher-level representations even under short priming durations, exhibiting distinct patterns for morphological and orthographic priming. Specifically, they show robust morphological priming driven by the facilitative effects of morpho-orthographic structure, whereas orthographic priming elicits null or inhibitory effects, reflecting the suppression of sublexical facilitation through lexical-level inhibition.
5.2. Morpho-semantic integration
Morpho-semantic integration refers to the process of validating the semantic information of a decomposed stem against the whole-word meaning of a derived word (Lavric et al., Reference Lavric, Rastle and Clapp2011; Longtin et al., Reference Longtin, Segui and Hallé2003; Rastle et al., Reference Rastle, Davis, Marslen-Wilson and Tyler2000). During this process, the morpho-semantic connection between a derived word and its decomposed stem generates interactive semantic feedback, which in turn facilitates activation of the stem’s semantic representation (Beyersmann & Grainger, Reference Beyersmann, Grainger and Crepaldi2023; Lee et al., Reference Lee, Maeng, Lack and Song2025). In the present study, this interactive semantic feedback appears to be reflected in the morphological priming effects across both language groups in the late N400, a time window where prime–target semantic integration predominantly occurs (Grainger & Holcomb, Reference Grainger and Holcomb2009; Lavric et al., Reference Lavric, Rastle and Clapp2011; Lee et al., Reference Lee, Maeng, Lack and Song2025). Specifically, morphological priming elicited a markedly greater attenuation of N400 negativity than in the earlier window, significantly exceeding the effects observed for both semantic and orthographic priming. Moreover, although negativity attenuation in the Semantic condition was most pronounced in this time window for both L1 and L2 speakers, the semantic priming effect remained substantially weaker than the morphological priming effect across language groups. This pattern suggests greater cognitive effort during prime–target semantic integration in the absence of morphological overlap (e.g., Brown & Hagoort, Reference Brown and Hagoort1993; Grainger & Holcomb, Reference Grainger and Holcomb2009; Holcomb et al., Reference Holcomb, Reder, Misra and Grainger2005). These findings underscore that the priming effect in the Morphological condition during the late N400 was not merely driven by semantic relatedness but by morpho-semantic connections, which provide stronger interactive semantic feedback via shared morphemes in both L1 and L2 processing.
Additionally, the contribution of orthographic overlap to the priming effect in the Morphological condition appears to be highly limited, as both language groups exhibited distinct trajectories of neural responses for priming in the Morphological and Orthographic conditions. Although both L1 and L2 speakers exhibited sustained negativity attenuation for the Orthographic condition in the late N400 window, its magnitude was considerably smaller than that of the Morphological condition and gradually declined throughout this time frame. In contrast, negativity attenuation for the Morphological condition was amplified through morpho-semantic integration, resulting in the strongest priming effect in the late N400 window.
Taken together, our results from the late N400 revealed converging patterns of morphological priming in L1 and L2 speakers, which cannot be reduced to additive effects of orthographic and semantic overlap, as assumed under the distributed connectionist model (e.g., Baayen et al., Reference Baayen, Milin, Đurđević, Hendrix and Marelli2011; Marelli et al., Reference Marelli, Amenta and Crepaldi2015; Plaut & Gonnerman, Reference Plaut and Gonnerman2000). Notably, this pattern was even more pronounced in L2 speakers than in L1 speakers, as shown in the topographies in Figure 2. Inspection of the topographic distributions revealed that semantic priming effects were minimal or barely visible, while orthographic priming showed only faint attenuation compared to the clear morphological priming effect in the late N400. These findings underscore that the facilitation observed in the Morphological condition among L2 speakers reflects genuine morphological processing rather than an additive combination of form- and meaning-based effects.
L2 speakers, however, diverged from L1 speakers by showing a less robust amplification of morphological priming effects, despite overall similarities in morphological priming patterns across the two groups. This divergence suggests that while L2 speakers can engage in morpho-semantic integration, this process seems to operate with lower efficiency or strength than in L1 speakers. This weaker morphological priming for L2 speakers in the late N400 window may be attributed to relatively weak encoding of form–meaning mappings in L2 lexical representations (Gollan et al., Reference Gollan, Forster and Frost1997; Jiang, Reference Jiang1999; Kroll et al., Reference Kroll, Van Hell, Tokowicz and Green2010). Notably, although L2 speakers exhibited statistically significant semantic priming effects, these effects were numerically small and substantially weaker in magnitude than those observed in L1 processing. Moreover, a visual inspection of our ERP waveforms shows that semantic priming emerged noticeably later in L2 than in L1. Given that morpho-semantic integration entails connecting the meanings of a derived word and its stem, the less efficient semantic processing in L2 may have led to a smaller amount of interactive semantic feedback compared to L1 processing.
5.3. Lateral inhibition
In the LN window, a notable L1–L2 difference emerged in the Orthographic condition: L1 speakers exhibited a pronounced negativity increase with a clear peak, whereas no observable effect was found in L2 speakers. This pattern aligns with prior masked priming findings showing that inhibitory orthographic priming effects emerge in L1 but not in L2 processing (Jiang, Reference Jiang2021; Nakayama & Lupker, Reference Nakayama and Lupker2018; Qiao & Forster, Reference Qiao and Forster2017), suggesting that lateral inhibition is not sufficiently activated in L2. In addition, while some overt priming studies have reported that L2 speakers can engage in lateral inhibition during orthographic priming when given sufficient processing time (Koval, Reference Koval2025; Nakayama & Lupker, Reference Nakayama and Lupker2018), the absence of inhibitory orthographic priming effects in the present study, despite the use of the same paradigm, suggests that lexical representations may be more weakly encoded in L2, leading to reduced lexical competition within the L2 lexicon (e.g., Jiang, Reference Jiang2021). Consistent with this interpretation, the temporal trajectory of orthographic priming further revealed that, although orthographic priming effects completely diminished before the end of the late N400 in L1, they extended into the LN window in L2, reflecting insufficient involvement of lexical inhibition.
The timing of the LN effect observed in L1 also provides important insight into how lateral inhibition operates differently in L1 and L2. Note that in L1 processing, increased negativity in the Orthographic condition emerged in the LN window, a time frame following the late N400, during which prime–target semantic integration primarily occurs (Grainger & Holcomb, Reference Grainger and Holcomb2009). This timing of inhibitory L1 orthographic priming reflects the intensification of lateral inhibition among co-activated orthographic representations as prime–target semantic integration fails at a later stage of lexical access (Barber et al., Reference Barber, Domínguez and De Vega2002; Domínguez et al., Reference Domínguez, De Vega and Barber2004; Lee et al., Reference Lee, Maeng, Lack and Song2025). Given this interplay of orthographic co-activation and semantic integration in the formation of lateral inhibition effects, the absence of increased LN negativity in the L2 orthographic priming condition may reflect weak semantic priming effects across the late N400 and LN windows, which likely stem from weaker form–meaning mappings in L2 lexical representations (Gollan et al., Reference Gollan, Forster and Frost1997; Jiang, Reference Jiang1999; Kroll et al., Reference Kroll, Van Hell, Tokowicz and Green2010). That is, it can be posited that weak form–meaning mappings in L2 may constrain L2 speakers’ ability to detect semantic mismatches between prime and target during orthographic priming, which may in turn limit the reinforcement of lateral inhibition effects.
5.4. Behavioral data
Although our behavioral data generally aligned with ERP responses for both the L1 and L2 groups, a discrepancy between the two measures was observed in the Orthographic condition. While our ERP data revealed a crucial difference between the L1 and L2 groups in the Orthographic condition, this difference was not reflected in the behavioral data. Specifically, in our ERP data, the L1 group exhibited a biphasic pattern, consisting of both an initial facilitative and a later inhibitory priming effect, whereas the L2 group showed only the facilitative effect. In contrast, the behavioral data indicated robust facilitative effects for both groups, with no significant group differences. This discrepancy may reflect the relative insensitivity of behavioral measures to late inhibitory processes under overt priming, resulting in similarly facilitative orthographic priming effects across language groups.
However, our behavioral orthographic priming effects also stand in contrast to an overt priming study employing a similar design to the present study, which found null orthographic priming effects for both L1 and L2 speakers (Koval, Reference Koval2025). One possible explanation for the discrepancies in orthographic priming across studies and measures is that facilitative orthographic priming in the behavioral data of the present study may reflect task effects influenced by stimulus design. In Koval (Reference Koval2025), half of the nonword targets were preceded by orthographically related primes, which may have reduced the reliability of orthographic information for participants, because relying on surface-form overlap does not guarantee correct lexical decisions. As a result, both L1 and L2 parsers may have been prompted to activate lateral inhibition more effectively when making lexical decisions for critical stimuli. In contrast, all nonword targets in the present study were preceded by unrelated primes, which may have induced both L1 and L2 parsers to strategically exploit orthographic information in their behavioral responses, as prime–target form overlap consistently leads to correct lexical decisions for critical stimuli. Further research is needed to clarify how nonword design modulates the engagement of lateral inhibition during visual word recognition.
5.5. Theoretical implications
Our findings on the biphasic pattern of L2 morphological decomposition during derived-word processing make a valuable contribution to our understanding of L2 processing within the framework of the Shallow Structure Hypothesis (SSH). The SSH has often been misunderstood in a large body of L2 research as proposing that full parsing is inoperative in L2 across all grammatical constructions, with target-like processing patterns consequently interpreted as evidence against the hypothesis. However, as clarified in its revised version (Clahsen & Felser, Reference Clahsen and Felser2018), the SSH does not necessarily preclude the engagement of target-like parsing in L2. Rather, it proposes that the operability of detailed structural analysis in L2 depends on multiple factors, including the target structure, proficiency and the nature of the task. Under this formulation of the SSH, structural processing in L2 is not an all-or-nothing, categorical phenomenon but a graded process that can emerge under favorable conditions. Accordingly, we suggest that future L2 research should focus on identifying the specific conditions under which L2 speakers are more likely to engage in target-like structural analysis, rather than merely adjudicating its presence or absence. In this context, our findings on L2 morphological priming effects can be understood in relation to the particular factors that facilitated the structural parsing of derived words.
Crucially, two factors appear to increase the likelihood of morphological parsing in L2: (1) the simplicity of the target structure and (2) the nature of the task. As discussed earlier, the derived word items employed in this study were structurally simple, consisting of only two semantically salient morphemes (Jacob et al., Reference Jacob, Heyer and Veríssimo2018). While the morphological decomposition of inflected words during L2 processing appears to be challenging due to the complexities associated with inflection (e.g., Lardiere, Reference Lardiere, Liceras, Zobl and Goodluck2008), L2 speakers may have fewer difficulties analyzing the structure of derived words whose morphemes contribute substantially to the whole-word meanings, as in the semantically transparent morphological primes (e.g., farm-er) tested in the present study.
Furthermore, even for such structurally simple bimorphemic words, it seems difficult for L2 speakers to consistently capture their morphological structure in the masked priming paradigm, which allows only extremely short processing durations (e.g., Ciaccio & Clahsen, Reference Ciaccio and Clahsen2020; Heyer & Clahsen, Reference Heyer and Clahsen2015; Li et al., Reference Li, Taft and Xu2017; Taft & Li, Reference Taft and Li2021). Yet, our results suggest that when sufficient processing time is available, as in overt priming, L2 speakers who attained advanced-level proficiency in L2 English through typical classroom instructions can reliably analyze the morpho-orthographic structure during early lexical access and subsequently use this structural information to process the semantic information of derived words. Recall that our participants had an average IELTS score of 7.5, corresponding to a proficiency level equivalent to C1 (advanced/effective operational proficiency) in the Common European Framework of Reference for Languages (CEFR), which is distinctly below the C2 level (mastery), representing near-native command of the language.
In brief, our detailed analysis of the temporal dynamics of L2-derived word processing enhances our understanding of L2 lexical processing under the SSH framework by revealing that L2 speakers can successfully engage in structural analysis of simple configurations without achieving native-like proficiency or immersion, provided that sufficient processing time is available.
6. Conclusion
The present overt priming study provides converging ERP evidence for a biphasic mechanism of morphological decomposition in L1 and L2 processing of derived words, encompassing both initial morpho-orthographic segmentation and subsequent morpho-semantic integration. Additionally, the study demonstrates that L1–L2 divergences remain in orthographic and semantic processing, likely reflecting weaker encoding of L2 lexical representations. Overall, our findings suggest that reasonably advanced L2 speakers, even without native-like proficiency, can engage in target-like structural parsing for simple morphological configurations when given sufficient processing time.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S1366728926101199.
Data availability statement
All behavioral and raw EEG data that support the findings of this study are now openly available on the Open Science Framework (OSF) at https://osf.io/ys69q/?view_only=58cf8469352b46aeba89ca3ef02bebb6.
Acknowledgments
This research was supported by the Early Career Scheme of the Research Grants Council of Hong Kong (Project ID: 27608822) for the project “EEG Investigation of Morphologically Complex Word Processing in a Second Language” (Principal Investigator: Yoonsang Song).
Author contribution
Jonah Lack: Conceptualization: J.L., J.M., Y.S.; Formal Analysis: J.L., H.K.L.; Funding Acquisition: Y.S.; Investigation: J.L.; Methodology: Y.S.; Project Administration: Y.S.; Resources: Y.S.; Supervision: Y.S.; Writing – Original Draft: J.L.; Writing – Original Draft (Data Analysis; Results): H.K.L.; Writing – Original Draft (Introduction; Discussion): J.M.; Writing–Review & Editing: Y.S.
Competing interests
The authors declare none.


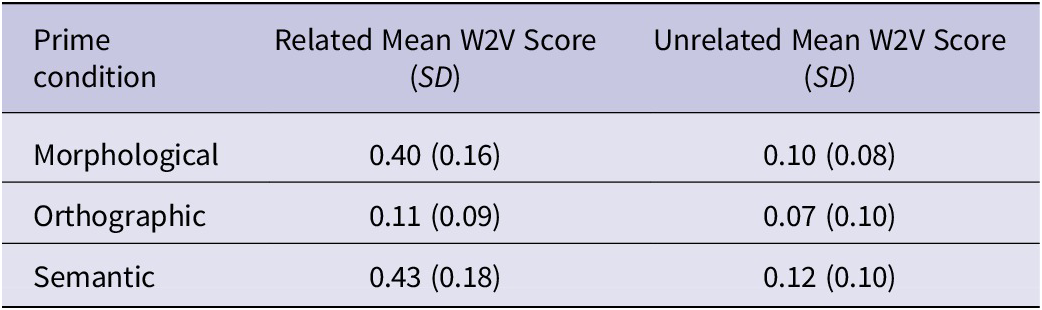






 Open access
Open access