


Non-technical Summary
A new species of extinct shark, Pseudocorax heteroserratus n. sp., has been identified through isolated teeth from fossil-rich Cretaceous rock in Morocco. Unlike previously known Pseudocorax species, this new species is distinguishable through its broad crown, elongated tooth base, and unusually variable serrations. These serrations range from unserrated to coarsely serrated and provide new insights into Pseudocorax evolutionary history. It raises the question of whether serrations in this lineage evolved gradually in a continuous process or from multiple independent origins. The findings of this study emphasize how large sample sizes of shark teeth are critical in palaeoichthyology, especially when species are known only from their teeth.
Introduction
The Pseudocoracidae is an extinct lamniform chondrichthyan family found in Late Cretaceous marine strata. This family is composed of two genera, Pseudocorax Priem (Reference Priem1897) and Galeocorax Cappetta (Reference Cappetta2012), both of which are believed to have been restricted to the Northern Hemisphere (Cappetta, Reference Cappetta2012). While the latter genus is monotypic, the former is currently composed of five species, ranging from the Cenomanian to the Maastrichtian (Cappetta, Reference Cappetta2012; Jambura et al., Reference Jambura, Stumpf and Kriwet2021). These five species are, from oldest to youngest, Pseudocorax kindlimanni Jambura et al., Reference Jambura, Stumpf and Kriwet2021; Pseudocorax duchaussoisi Guinot et al., Reference Guinot, Underwood, Cappetta and Ward2013; Pseudocorax laevis (Leriche, Reference Leriche1906); Pseudocorax granti Cappetta and Case, Reference Cappetta and Case1975; and Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843). The taxonomic validity of P. granti has come under scrutiny, with Hamm and Shimada (Reference Hamm and Shimada2007) synonymizing the taxon with P. laevis. While some have agreed with the synonymy (Shimada, Reference Shimada2009; Ciampaglio et al., Reference Ciampaglio, Cicimurri, Ebersole and Runyon2013), others have refuted it (Guinot et al., Reference Guinot, Underwood, Cappetta and Ward2013).
Maastrichtian Pseudocorax
Two Pseudocorax species have been described from Maastrichtian strata: Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843) and Pseudocorax granti Cappetta and Case, Reference Cappetta and Case1975. The isolated occurrence of P. granti in the Maastrichtian is noted from the Kemp Clay, Texas, USA (Case and Cappetta, Reference Case and Cappetta1995); however, the present study is scrutinous on whether the teeth truly originated from the unit, or if they instead were reworked from older strata. Consequently, the present study will not recognize the occurrence until it is further reaffirmed.
The most abundant Maastrichtian species is the nominal species Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843). Historically, P. affinis has been the only described serrated Pseudocorax species and has been reported numerous times, particularly from European localities (Herman, Reference Herman1977; Cappetta and Odin, Reference Cappetta and Odin2001; Adolfssen and Ward, Reference Adolfssen and Ward2014). Furthermore, P. affinis has historically been reported from both North Africa and the Middle East (Arambourg, Reference Arambourg1952; Lewy and Cappetta, Reference Lewy and Cappetta1989; Cappetta et al., Reference Cappetta, Bardet, Pereda Suberbiola, Adnet, Akkrim, Amalik and Benabdallah2014). More recent reports have noted P. affinis from various North American localities in the Atlantic Coastal Plain (Case et al., Reference Case, Cook, Sadorf and Shannon2017; Boles et al., Reference Boles, Ullmann, Putman, Ford and Deckhut2024). In this study, a sample of Pseudocorax from Khouribga Province, Morocco, is analyzed, and a new Pseudocorax species is described.
Geologic setting
Situated approximately 150 km south of Rabat, the Oulad Abdoun Basin is the largest and northernmost of the Moroccan phosphate basins (Fig. 1) (Bardet et al., Reference Bardet, Pereda Suberbiola, Jouve, Bourdon, Vincent, Houssaye, Rage, Jalil, Bouya and Amaghzaz2010). It is an economically important area for commercial phosphate quarrying, with numerous open-pit quarries. The phosphate-bearing strata within the Oulad Abdoun Basin are a part of the Mediterranean Tethyan phosphogenic province, a large belt of sedimentary deposits that extends around the southern border of the Mediterranean Sea (Lucas and Prévôt-Lucas, Reference Lucas, Prévôt-Lucas, Narin, Ricou, Vrielynck and Dercourt1996). Many stratigraphic units within this basin are divided into several informal layers classically called “Couches” and “Sillons,” ranging from the Late Cretaceous (Maastrichtian) to the Eocene (Lutetian) in age (Duffin and Ward, Reference Duffin and Ward2025).
(1) Map of northern Morocco showing the location of the Moroccan phosphates. (2) Map of the Oulad Abdoun Basin in Khouribga Province.

The majority of Maastrichtian units at the Sidi Chennane phosphate quarry are predominantly contained within Couche III (see Duffin and Ward, Reference Duffin and Ward2025, fig. 1). Couche III, which is often subdivided into upper and lower groupings, consists broadly of limestones, phosphorite sands, and both phosphatic and oxidized marls (Kocsis et al., Reference Kocsis, Gheerbrant, Mouflih, Cappetta, Yans and Amaghzaz2014; Duffin and Ward, Reference Duffin and Ward2025). The upper bone bed of Couche III, from which the sample described in the present study originates, occurs within a phosphatic marl that directly overlies the limestone unit separating the upper and lower Couche III units. Couche III has been dated to the latest Maastrichtian on the basis of both selachian biostratigraphy and carbon and oxygen isotope chemostratigraphy (Cappetta et al., Reference Cappetta, Bardet, Pereda Suberbiola, Adnet, Akkrim, Amalik and Benabdallah2014; Kocsis et al., Reference Kocsis, Gheerbrant, Mouflih, Cappetta, Yans and Amaghzaz2014; Duffin and Ward, Reference Duffin and Ward2025). Besides neoselachians, the diverse vertebrate remains of the upper Couche III include mosasaurs, actinopterygians, crocodilians, plesiosaurs, marine turtles, pterosaurs, and rare dinosaurs (Suberbiola et al., Reference Suberbiola, Bardet, Jouve, Iarochène, Bouya and Amaghzaz2003; Vincent et al., Reference Vincent, Bardet, Suberbiola, Bouya, Amaghzaz and Meslouh2011; de Lapparent de Broin et al., Reference de Lapparent de Broin, Bardet, Amaghzaz and Meslouh2014; Longrich et al., Reference Longrich, Pereda-Suberbiola, Jalil, Khaldoune and Jourani2017, Reference Longrich, Martill and Andres2018; Vullo et al., Reference Vullo, Cavin, Khalloufi, Amaghzaz, Bardet, Jalil, Jourani, Khaldoune and Gheerbrant2017; Rempert et al., Reference Rempert, Martens and Vinkeles Melchers2022). These units were deposited in a warm and shallow marine environment at an approximate paleolatitude of 25°S (Bardet et al., Reference Bardet, Pereda Suberbiola, Jouve, Bourdon, Vincent, Houssaye, Rage, Jalil, Bouya and Amaghzaz2010).
Materials and methods
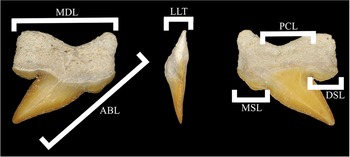
Specimens were sieved out of freshly excavated in situ Couche III sediments from the upper bone bed at the Sidi Chennane quarry and its immediate surroundings. This sieving was conducted partially by state-licensed fossil dealer Boubker Chaibi and his daughter Ikhlass Chaibi, while other specimens were purchased by Boubker from other local collectors employing the same field methodologies on site. These teeth were purchased directly from Boubker and subsequently were accessioned within the Mississippi Museum of Natural Science (MMNS) in Jackson, Mississippi, USA. References for specimen measurements in Table 1 are shown in Figure 2.
Measurements of Pseudocorax heteroserratus n. sp. figured specimens from the Sidi Chennane quarry, Morocco. JP, jaw position; RP, row position within jaw; ABL, apicobasal length along the mesial crown (and tooth base edge); MDL, largest mesiodistal length (laterally, edge to edge); PCL, principal cusp mesiodistal length at the widest point; MSL, mesial shoulder length mesiodistally; DSL, distal shoulder length mesiodistally; LLT, labiolingual thickness of crown at the widest point. *measurement incomplete due to incomplete specimen.

Pseudocorax heteroserratus n. sp. measurements indexed in Table 1. ABL, apicobasal length along the mesial crown (and tooth base edge); MDL, largest mesiodistal length (laterally, edge to edge); PCL, principal cusp mesiodistal length at the widest point; MSL, mesial shoulder length mesiodistally; DSL, distal shoulder length mesiodistally; LLT, labiolingual thickness of crown at the widest point.

Specimens were photographed using the Keyence VHX-7000 digital microscope owned by the Alabama Materials Institute at The University of Alabama in Tuscaloosa, Alabama, USA. Specimen angles (e.g., profile, labial, apical, and basal views) that could not be sufficiently captured by placing the specimen on the microscope stage were taken with the assistance of Geo-Tac Mineral and Fossil Mounting Display Putty. Figures were subsequently edited and created using Adobe Photoshop 2024 and Adobe Illustrator 2024. Morphological nomenclature follows the convention of Cappetta (Reference Cappetta2012), outside of the use of carinae for the tooth cutting edge. Further, the present study uses “tooth base” in place of root in similar convention to both modern ichthyology (i.e., Ritter and Dellios, Reference Ritter and Dellios2018) and Paleozoic palaeoichthyology (i.e., Ginter et al., Reference Ginter, Hampe, Duffin and Schultze2010) as these teeth affix to the jaws via connective tissues rather than being rooted into the jaw cartilage itself.
Repository and institutional abbreviation
MMNS VP-: Mississippi Museum of Natural Science, Jackson, Mississippi, USA.
Systematic paleontology
Class Chondrichthyes Huxley, Reference Huxley1880
Subclass Elasmobranchii Bonaparte, Reference Bonaparte1838
Cohort Euselachii Hay, Reference Hay1902
Subcohort Neoselachii Compagno, Reference Compagno1977
Order Lamniformes Berg, Reference Berg1958
Family Pseudocoracidae Cappetta, Reference Cappetta2012
Genus Pseudocorax Priem, Reference Priem1897
Type species
Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843), from the upper Maastrichtian, Netherlands.
Pseudocorax heteroserratus new species
Reference Gemmellaro1920 Corax affinis; Gemmellaro, p. 152, 156, 186, Tavola II, figs. 20–23.
Reference Arambourg1952 Pseudocorax affinis; Arambourg, p. 14, 117, 367, Tableau I, II, VI, Annexe I, Planche XXI, figs. 4–6.
Reference Cappetta, Bardet, Pereda Suberbiola, Adnet, Akkrim, Amalik and Benabdallah2014 Pseudocorax affinis; Cappetta et al., p. 221, 236, table 1.
Holotype
MMNS VP-12820, isolated upper left anterior tooth.
Paratypes
MMNS VP-12821, isolated lower left anterior tooth; MMNS VP-12822, isolated lower right anterior tooth; MMNS VP-12823, isolated upper left lateroposterior tooth; MMNS VP-12824, isolated lower right lateroposterior tooth; MMNS VP-12825, isolated upper right lateroposterior tooth; MMNS VP-12826, isolated lower left lateroposterior tooth.
Referred specimens
MMNS VP-12827, 20 isolated complete to near-complete teeth; MMNS VP-12828, 38 isolated complete to near-complete teeth.
Diagnosis
Crown and tooth base mesiodistally expanded, especially along upper and lower jaw anterior teeth. Mesial shoulder generally more prominent along basal crown, extending further apically. Variable serrations, from absent to full and coarse to fine, regardless of jaw position or potential ontogenetic stage. Deep basal margin on tooth base with variable tooth base lobe morphology.
Occurrence
Upper Couche III layer of the Oulad Abdoun Basin, Sidi Chennane quarry, near Oued Zem, Khouribga Province, Morocco.
Description
Tooth measurements can be found in Table 1 (Fig. 2).
Upper anterior files
Upper anterior crowns are broad, short, and distally inclined with minor mesial recurve apically (Fig. 3). Marked shoulders hang apically from the basalmost lateral portions of the crown. There is a distinct notch separating the apical shoulder from the principal cusp, although the shoulders are still well connected to the basal principal cusp. Shoulders apexes can either be blunt or come to a well-defined point. Apically from the shoulders, carinae are convex, with the distal carina being more so than the mesial. The mesial carina then becomes weakly concave toward the crown apex. Serrations are often present but variable along the crown. Shoulders are weakly serrated to unserrated. The crown exhibits strong labiolingual compression, with the labial face being more or less flat and the lingual face convex. The crown is labiolingually thickest toward its base and tapers down apically. A protuberant radicular “waist” is present along the labial crown at the crown–tooth base junction, labially overhanging the tooth base. A weak dental band is present at the crown–tooth base junction.
Upper anterior Pseudocorax heteroserratus n. sp. teeth. (1–5) MMNS VP-12820 (holotype), upper left anterior tooth in (1) lingual, (2) basal, (3) apical, (4) profile, and (5) labial views. (6–10) MMNS VP-12827.01, upper left anterior tooth in (6) lingual, (7) basal, (8) apical, (9) profile, and (10) labial views. Scale bars = 5 mm.

The tooth base is markedly bifurcated, with the mesial lobe being strongly mesiodistally compressed compared with the highly expanded distal lobe. Tooth base lobes are ovate. In profile view, the apicalmost tooth base is thickest and projects lingually; the tooth base then tapers basally. The tooth base margins pinch near the apicalmost tooth base, creating a marked concavity before flaring convexly toward the crown–tooth base junction. A marked axial furrow is present.
Lower anterior files
Broad, elongated lower anterior crowns are distally inclined to varying degrees (Fig. 4). In anteriormost files, carinae are straight to weakly convex; there is more carinae variance in laterally situated anterior files. In these laterally situated anterior files, the convexities are more prominent; however, toward the apex, the mesial carina straightens. Ovaloid shoulders are present along the basalmost crown, directly lateral to the principal cusp. A marked notch is present separating the apical shoulder and the principal cusp, although the shoulders are still well connected to the basal principal cusp. The shoulder apexes are predominantly blunt, especially when compared with upper anterior files. Serrations are often present but variable along the crown, from fine to coarse. The crown exhibits strong labiolingual compression, with the labial face being more or less flat and the lingual face convex. The crown is labiolingually thickest toward its base and tapers down apically. A protuberant radicular “waist” is present along the labial crown at the crown–tooth base junction, labially overhanging the tooth base. A weak dental band is present along the lingual crown–tooth base margin.
Lower anterior Pseudocorax heteroserratus n. sp. teeth. (1–5) MMNS VP-12821 (paratype), lower left anterior tooth in (1) lingual, (2) basal, (3) apical, (4) profile, and (5) labial views. (6–10) MMNS VP-12827.02, lower left anterior tooth in (6) lingual, (7) basal, (8) apical, (9) profile, and (10) labial views. (11–15) MMNS VP-12827.03, lower left anterolateral tooth in (11) lingual, (12) basal, (13) apical, (14) profile, and (15) labial views. (16–20) MMNS VP-12822 (paratype), lower right anterolateral tooth in (16) lingual, (17) basal, (18) apical, (19) profile, and (20) labial views. (21–25) MMNS VP-12827.04, lower right anterolateral tooth in (21) lingual, (22) basal, (23) apical, (24) profile, and (25) labial views. Scale bars = 5 mm.

The tooth base is strongly bifurcated, with the distal tooth base lobe being marginally mesiodistally wider than the mesial. Tooth base lobes are ovaloid. In profile view, the apicalmost tooth base is thickest and projects lingually; the tooth base then tapers basally. The tooth base margins pinch near the apicalmost tooth base, creating a marked concavity before flaring convexly toward the crown–tooth base junction. A distinct axial furrow is present.
Upper lateroposterior files
Broad, distally inclined upper lateral crowns sometimes exhibit a mesial recurve in the apical crown (Fig. 5). More posteriorly, crowns exhibit more apicobasal compression (Fig. 7). Ovaloid or rare pointed shoulders are present along the basalmost portions of the crown, laterally from the principal cusp. The mesial shoulder is less defined compared with the distal; in some more posteriorly positioned files, the mesial shoulder may be nearly to completely absent. The distal shoulder remains markedly defined but still well connected to the basal principal cusp. Apically from the shoulders, distal carinae vary from straight to weakly convex, becoming uniformly convex approaching the apex. The mesial carinae are convex, with recurving crowns becoming concave apically whereas only distally inclined crowns become straight apically. Serrations are often present along the crown but are highly variable. Serrations vary, ranging from fully serrated to completely unserrated. Serrations also vary from coarse to fine. The crown is strongly labiolingually compressed, with the labial face being more or less flat and the lingual face convex. The crown is labiolingually thickest toward its base and tapers down apically. A protuberant radicular “waist” is present along the labial crown at the crown–tooth base junction, labially overhanging the tooth base. A weak dental band is present along the lingual crown–tooth base margin.
Upper lateroposterior Pseudocorax heteroserratus n. sp. teeth. (1–5) MMNS VP-12823 (paratype), upper left lateroposterior tooth in (1) lingual, (2) basal, (3) apical, (4) profile, and (5) labial views. (6–10) MMNS VP-12827.05, (?) upper right lateroposterior tooth in (6) lingual, (7) basal, (8) apical, (9) profile, and (10) labial views. (11–15) MMNS VP- 12827,06, (?) upper left lateroposterior tooth in (11) lingual, (12) basal, (13) apical, (14) profile, and (15) labial views. (16–20) MMNS VP-12827.07, upper left lateroposterior tooth in (16) lingual, (17) basal, (18) apical, (19) profile, and (20) labial views. (21–25) MMNS VP-12827.08, upper right lateroposterior tooth in (21) lingual, (22) basal, (23) apical, (24) profile, and (25) labial views. (26–30) MMNS VP-12827.09, upper left lateroposterior tooth in (26) lingual, (27) basal, (28) apical, (29) profile, and (30) labial. Scale bars = 5 mm.

The tooth base is markedly bifurcated, with the distal tooth base lobe being variably wider mesiodistally than the medial lobe. The distal tooth base lobe varies from being only slightly to strongly more mesiodistally expanded than the mesial tooth base lobe. Tooth base lobes are ovaloid. In profile view, the apicalmost tooth base is thickest and projects lingually; the tooth base then tapers basally. The tooth base margins pinch near the apicalmost tooth base, creating a marked concavity before flaring convexly toward the crown–tooth base junction. A distinct axial furrow is present.
Lower lateroposterior files
Lateroposterior crowns are distally inclined and range from broad to somewhat gracile (Fig. 6). More posteriorly positioned files are more distally inclined (Fig. 8). Ovaloid shoulders are present along the basalmost portions of the crown, laterally from the principal cusp. The mesial shoulder is less defined compared with the distal. Most shoulders have a notch separating them from the principal cusp; however, the shoulders are still well connected to the basal principal cusp. Some mesial shoulders are unnotched. Apically from the shoulders, distal carinae are straight to slightly convex. The mesial carinae are convex basally and often become straight apically. Mesial carinae, near the crown apex, are sometimes weakly concave. Serrations are often present but generally expressed weaker and finer than on other files. The crown is strongly compressed labiolingually, with the labial face being more or less flat and the lingual face convex. The crown is labiolingually thickest toward its base and tapers down apically. A protuberant radicular “waist” is present along the labial crown at the crown–tooth base junction, labially overhanging the tooth base. A weak dental band is present along the lingual crown–tooth base margin.
Lower lateroposterior Pseudocorax heteroserratus n. sp. teeth. (1–5) MMNS VP-12827.10, lower right lateroposterior tooth in (1) lingual, (2) basal, (3) apical, (4) profile, and (5) labial views. (6–10) MMNS VP-12827.11, lower right lateroposterior tooth in (6) lingual, (7) basal, (8) apical, (9) profile, and (10) labial views. (11–15) MMNS VP-12827.12, lower right lateroposterior tooth in (11) lingual, (12) basal, (13) apical, (14) profile, and (15) labial views. (16–20) MMNS VP-12824 (paratype), lower left lateroposterior tooth in (16) lingual, (17) basal, (18) apical, (19) profile, and (20) labial views. Scale bars = 5 mm.

The tooth base is markedly bifurcated, with the distal tooth base lobe being slightly mesiodistally wider than the mesial. Tooth base lobes are ovaloid. In profile view, the apicalmost tooth base is thickest and projects lingually; the tooth base then tapers basally. The tooth base margins pinch near the apical-most tooth base, creating a marked concavity before flaring convexly toward the crown–tooth base junction. A distinct axial furrow is present.
Etymology
“Hetero-”, Latin for “different”, and “serratus” for “serrations,” highlighting the variability in the presence and size of serrations along the crown.
Comparison and remarks.
Pseudocorax comparisons
Pseudocorax heteroserratus n. sp. can be differentiated from Pseudocorax laevis (Leriche, Reference Leriche1906), Pseudocorax granti Cappetta and Case, Reference Cappetta and Case1975, Pseudocorax duchaussoisi Guinot et al., Reference Guinot, Underwood, Cappetta and Ward2013), and Pseudocorax kindlimanni Jambura et al., Reference Jambura, Stumpf and Kriwet2021 by the presence of serrations along the carinae. While P. laevis (sensu stricto Leriche, Reference Leriche1906) exhibits the broadest crown of unserrated Pseudocorax, P. heteroserratus possesses a substantially broader crown than any aforementioned taxa. Similarly, P. heteroserratus has much greater mesiodistal expansion of the tooth base lobes than the preceding taxa (Case, Reference Case1991, plate 2, fig. 1; Guinot et al., Reference Guinot, Underwood, Cappetta and Ward2013, fig. 14I; Jambura et al., Reference Jambura, Stumpf and Kriwet2021, fig. 3).
Pseudocorax heteroserratus n. sp. and Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843) both exhibit serrated crowns, necessitating more subtle morphological distinctions. P. heteroserratus crowns are generally broader than those of P. affinis, with notably more convex carinae. The difference in crown breadth is best seen in anterior and upper lateroposterior comparisons between the two taxa (Figs. 3, 4, 5, 7) (Case and Cappetta, Reference Case and Cappetta2004, plate 3, figs. 3a, b; Adolfssen and Ward, Reference Adolfssen and Ward2014, fig. 8G–J; Case et al., Reference Case, Cook, Sadorf and Shannon2017, fig. 3H). One of the more striking differentiating morphology between P. heteroserratus and P. affinis crowns is the predominance of the mesial shoulder. P. heteroserratus exhibits a more predominant mesial shoulder than P. affinis, especially in upper anterior and some lateroposterior files (Figs. 3, 5, 7) (Cappetta and Odin, Reference Cappetta and Odin2001, plate 1, figs. 2, 3; Boles et al., Reference Boles, Ullmann, Putman, Ford and Deckhut2024, fig. 2H1, H2). Finally, P. heteroserratus serrations are often much more variable and, on occasion, completely absent, whereas P. affinis serrations are often finer and more regular (Fig. 8; Table 1) (Case and Cappetta, Reference Case and Cappetta2004, plate 3, fig. 3a, b; Adolfssen and Ward, Reference Adolfssen and Ward2014, fig. 8H–J; Boles et al., Reference Boles, Ullmann, Putman, Ford and Deckhut2024, fig. 2H1, H2). The tooth base of Pseudocorax heteroserratus n. sp. differs from that of Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843) by its greater mesiodistal expansion, thus increasing the axial asymmetry between the mesial and distal tooth base lobes (Boles et al., Reference Boles, Ullmann, Putman, Ford and Deckhut2024, fig. 2H1, H2). This is especially prevalent within upper anterior files. Further, the flare of the P. heteroserratus lateral tooth base edges is substantially more prominent than on P. affinis.
Upper lateroposterior Pseudocorax heteroserratus n. sp. teeth. (1–5) MMNS VP-12825 (paratype), upper right lateroposterior tooth in (1) lingual, (2) basal, (3) apical, (4) profile, and (5) labial views. (6–10) MMNS VP-12827.13, upper right lateroposterior tooth in (6) lingual, (7) basal, (8) apical, (9) profile, and (10) labial views. (11–15) MMNS VP-12827.14, upper right lateroposterior tooth in (11) lingual, (12) basal, (13) apical, (14) profile, and (15) labial views. (16–20) MMNS VP-12827.15, upper right lateroposterior tooth in (16) lingual, (17) basal, (18) apical, (19) profile, and (20) labial views. (21–25) MMNS VP-12827.16, upper right lateroposterior tooth in (21) lingual, (22) basal, (23) apical, (24) profile, and (25) labial views. (26–30) MMNS VP-12827.17, upper left lateroposterior tooth in (26) lingual, (27) basal, (28) apical, (29) profile, and (30) labial views. Scale bars = 5 mm.

Lower lateroposterior Pseudocorax heteroserratus n. sp. teeth. (1–5) MMNS VP-12827.18, lower right lateroposterior tooth in (1) lingual, (2) basal, (3) apical, (4) profile, and (5) labial views. (6–10) MMNS VP-12827.19, lower right lateroposterior tooth in (6) lingual, (7) basal, (8) apical, (9) profile, and (10) labial views. (11–15) MMNS VP- 12827.20, lower right lateroposterior tooth in (11) lingual, (12) basal, (13) apical, (14) profile, and (15) labial views. (16–20) MMNS VP-12826 (paratype), lower left lateroposterior tooth in (16) lingual, (17) basal, (18) apical, (19) profile, and (20) labial views. Scale bars = 5 mm.

Serration variability along the Pseudocorax heteroserratus n. sp. crown. (1–3) MMNS VP-12828.01, upper right lateroposterior tooth showing (1) smooth distal carina, (2) entire tooth, and (3) smooth mesial carina. (4–6) MMNS VP-12828.02, upper left lateroposterior tooth showing (4) irregularly serrated mesial carina, (5) entire tooth, and (6) irregularly serrated mesial carina. (7–9) MMNS VP-12828.03, upper left lateroposterior tooth showing (7) finely serrated mesial carina, (8) entire tooth, and (9) irregularly serrated distal carina. (10–12) MMNS VP-12828.04, lower right lateroposterior tooth showing (10) finely serrated mesial carina, (11) entire tooth, and (12) irregularly serrated distal carina. (13–15) MMNS VP-12828.05, upper left lateroposterior tooth showing (13) finely serrated mesial carina, (14) entire tooth, and (15) finely serrated distal carina. (16–18) MMNS VP-12828.06, upper right lateroposterior tooth showing (16) coarsely serrated distal carina, (17) entire tooth, and (18) coarsely serrated mesial carina. All magnified figures are 3× magnification. Scale bar = 5 mm.

Concerns about tooth size bias can be raised on the basis of the sifting methods used within the Moroccan phosphates as all Pseudocorax heteroserratus n. sp. teeth within the present sample size reside within a roughly 0.66 cm apicobasal range (see Table 1). The present Moroccan sample size, when compared with a 17-tooth-type stratum sample of Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843) from the MMNS collections, falls within the range of the largest teeth in that type stratum sample. The potential of the present Moroccan sample being a hypothetical ontogenetic stage of mature adult P. affinis is highly unlikely due not only to the size overlap with the MMNS P. affinis sample, but also to its size overlap with figured P. affinis apicobasal lengths (Case and Cappetta, Reference Case and Cappetta2004; Case et al., Reference Case, Cook, Sadorf and Shannon2017). Further, there has yet to be an extant or extinct selachian known that has lost serrations as the animal matures. There is, however, ample evidence of the opposite within extant and extinct elasmobranchs wherein younger animals lack or have coarse, irregular serrations while adult animals have finer and more regular serrations (Chandler et al., Reference Chandler, Chiswell and Faulkner2006; Ehret et al., Reference Ehret, Hubbell and Macfadden2009; Türtscher et al., Reference Türtscher, Jambura, López-Romero, Kindlimann, Sato, Tomita and Kriwet2022; Ebersole et al., Reference Ebersole, Kelosky, Huerta-Beltrán, Cicimurri and Drymon2023).
The most diagnostic contrasting tooth positions between Pseudocorax heteroserratus n. sp. and Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843) are the upper anterior files (Fig. 3) (Herman, Reference Herman1977, plate 4, fig. 5b); some lateroposterior files may lack the prominent mesial shoulder or exhibit tooth base structures similar to P. affinis. The crowns of upper anterior P. heteroserratus are squatter than those of P. affinis and feature a unique weak mesial recurve of the apex. P. affinis upper anterior crowns are more straight, gracile, and apicobasally elongated by comparison. The mesial shoulder of P. heteroserratus is far more prominent, forming a pointed, superficially cusp-like structure, whereas P. affinis shoulders are considerably weaker and blunter. The tooth bases of P. heteroserratus upper anterior files can rival and even supersede the apicobasal length of the crown, causing them to feature greater mesiodistal elongation than in the tooth bases of P. affinis. Further, the P. heteroserratus tooth base lobes possess greater asymmetry than P. affinis, with the mesial lobes being somewhat compressed and the distal lobes notably expanded mesiodistally.
Pseudocorax heteroserratus n. sp. lateroposterior upper and lower jaw teeth are sometimes difficult to distinguish between one another. This leads some lateral file placements of isolated teeth in the present study to be somewhat tentative. These ambiguous placements are noted in figure captions with “(?).” Gradient monognathic heterodonty is present within both palatoquadrates and Meckel’s cartilage. Within both jaws, the crowns become more distally inclined posteriorly. This distal inclination is more gradual within the palatoquadrate as, contrarily, lower anterior teeth are much more erect than more distal lower files. Dignathic heterodonty is also present, but not as starkly as in other lamniform taxa (e.g., Siversson et al., Reference Siversson, Lindgren, Newbrey, Cederström and Cook2015; Shimada and Everhart, Reference Shimada and Everhart2019). While both the palatoquadrates and Meckel’s cartilage have distally inclined teeth, teeth of the latter have more gracile crowns with less distal inclination. Both jaws having notable distal inclination diverts P. heteroserratus from the classic “lamnoid” tooth plan as described by Shimada (Reference Shimada2002). Further, there are no notable reduced UA3 teeth within the P. heteroserratus sample. There are similar dignathic patterns within Haimirichia amonensis (Cappetta and Case, Reference Cappetta and Case1975). Associated dentitions from this taxon favor the gnathic trends of P. heteroserratus in having notable distal inclination throughout both palatoquadrates and Meckel’s cartilage (Vullo et al., Reference Vullo, Guinot and Barbe2016). No study of articulated Pseudocorax remains has differentiated upper and lower jaw teeth, nor have similar inferences been made with large samples of isolated teeth.
Taxonomic history
Pseudocorax heteroserratus n. sp. has historically been described under Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843) from multiple phosphate quarries in the Mediterranean Tethys Margin region. The oldest description of the taxon occurs in Gemmellaro (Reference Gemmellaro1920) as “Corax affinis” from Sciarauna-el-Ghibli, Sibaiya (Egypt). Arambourg (Reference Arambourg1952) similarly describes occurrences of P. heteroserratus under P. affinis; however, the study aligns the species correctly with the genus Pseudocorax. These Arambourg (Reference Arambourg1952) occurrences originate from Moroccan phosphates in quarries near where the type and referred specimens herein were found.
Pseudocorax heteroserratus n. sp. occurrences are possibly described but not figured as “Pseudocorax aff. affinis” by Lewy and Cappetta (Reference Lewy and Cappetta1989) from three informal uppermost Campanian to Maastrichtian “beds” within the Mishash and Menuha formations, Israel. The study notes that, in the Campanian–Maastrichtian boundary beds of “bed 4,” the Pseudocorax often exhibit a partially serrated crown suggested to be an intermediate morphology within the Pseudocorax laevis (Leriche, Reference Leriche1906)—Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843) lineage (Lewy and Cappetta, Reference Lewy and Cappetta1989). The synonymy herein does not include these and the younger, more serrated occurrences labeled P. affinis by Lewy and Cappetta (Reference Lewy and Cappetta1989) as the Israeli specimens are unfigured and no other serrated Pseudocorax occurrences have been described from the country.
By contrast, the present study does include the unfigured occurrences listed by Cappetta et al. (Reference Cappetta, Bardet, Pereda Suberbiola, Adnet, Akkrim, Amalik and Benabdallah2014) within the synonymy of Pseudocorax heteroserratus n. sp. on the basis of their geographic and stratigraphic proximity to the type specimens described herein.
General remarks
Pseudocorax heteroserratus n. sp. marks the second Pseudocorax to have a serrated crown; however, these serrations are highly variable and range from completely unserrated to fully finely to coarsely serrated (Fig. 8; Table 1). There are no notable positional trends with the presence or absence of serrations as many different teeth of similar jaw positions have high variance. Broader crown morphologies also do not positively correlate with serration variability. Sexual dimorphism cannot be sufficiently ruled out with a sample of isolated teeth; however, there are no two distinct morphologies but rather a serration gradation. This makes the case for sexual dimorphism of the carinae an improbable one.
Discussion
Pseudocorax serration evolution
Pseudocorax heteroserratus n. sp. marks the second unequivocal Maastrichtian Pseudocorax species and is the second congeneric species to feature a serrated crown. Serrated crowns present within two distinct Pseudocorax taxa calls into question the evolution of serrations within the genus. Possible scenarios include: (1) evolution happened progressively as a singular event throughout a widely spread population, or (2) two separate Pseudocorax lineages evolved serrations independently. Only Lewy and Cappetta (Reference Lewy and Cappetta1989) discuss the evolution of serrations within Pseudocorax, highlighting an intermediate form present in a potential Pseudocorax laevis (Leriche, Reference Leriche1906)—Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843) chronospecies lineage. While the validity of the latter species being P. affinis needs verification as they are unfigured in Lewy and Cappetta (Reference Lewy and Cappetta1989), the presence of intermediate, partially serrated Pseudocorax in the Campanian–Maastrichtian boundary beds of Israel suggests a genesis of serrations within the genus occurred soundly in the Campanian.
The presence of irregular serrations, and even a lack thereof, among some Pseudocorax heteroserratus n. sp. carinae has not been observed in teeth at the Pseudocorax affinis type locality (Münster in Agassiz, Reference Agassiz1843) (Table 1; Fig. 8). This difference between contemporaneous-yet-geographically separate populations of Pseudocorax could give credence to the hypothesis that serrations evolved independently at least twice within the genus and that the ecology within the Tethyan region did not drive the evolution of serrations to fixation in P. heteroserratus before the Cretaceous–Paleogene (K-Pg) extinction event. Alternatively, this may suggest that during a broad, progressive evolutionary transition from smooth to serrated carinae, populations in Morocco and likely the broader Tethyan region diverged into a distinct taxon before the complete development of serrated crowns, potentially due to the absence of ecological pressures necessary to drive serration fixation. Perhaps P. heteroserratus became extinct (likely via the K-Pg extinction event) before serration fixation could take hold within its population.
If serrations evolved two separate times within Pseudocorax, then two potential phylogenetic scenarios are proposed: (1) there are two distinct, broad-crowned, unserrated Pseudocorax species existing in geographic isolation from one another that evolved serrations separately; or (2) the Pseudocorax laevis (Leriche, Reference Leriche1906) lineage had two populations independently serrate, resulting in homoplastic serration genesis in Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843) and Pseudocorax heteroserratus n. sp. The current paleobiogeographic range of P. laevis is restricted to the eastern Atlantic Ocean as per Guinot et al. (Reference Guinot, Underwood, Cappetta and Ward2013), suggesting the former scenario is less likely. However, reanalysis of the unfigured P. laevis discussed by Lewy and Cappetta (Reference Lewy and Cappetta1989) may further elucidate the relationship.
If serrations evolved only a single time within Pseudocorax, the most feasible process by which that would occur is for the western Tethys Pseudocorax population to diverge at a point between initial serration genesis in Pseudocorax laevis (Leriche, Reference Leriche1906) and serration fixation in Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843). It can be implied that an equally broad variation in carinae was present in a transitional Pseudocorax within the P. laevis–P. affinis lineage as it progressively came into serration fixation.
Conclusion
Reexamination of Pseudocorax teeth from the upper Maastrichtian Moroccan phosphates has yielded a new species, Pseudocorax heteroserratus n. sp., which is characterized most prominently by teeth with highly variable serrations, regardless of jaw position. Other diagnostic characters include a strongly pronounced mesial shoulder on some jaw positions and an overall broader tooth morphology. Upper anterior files are particularly diagnostic for the taxon. The variability in P. heteroserratus serration breadth and size leads to differing hypotheses as to how two distinct serrated Pseudocorax species were present within the Maastrichtian. The first hypothesis suggests that serrations evolved two separate times in the genus, whereas the second hypothesis suggests that serrations evolved once, with P. heteroserratus diverging from an intermediate Pseudocorax species between Pseudocorax laevis (Leriche, Reference Leriche1906) and Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843). This evolution into a distinct Tethyan taxon likely corresponded with the loss of ecological factors that were actively driving serrations into fixation within P. affinis.
Further study is necessary to provide better insights into Pseudocorax heteroserratus n. sp. and, more broadly, generic phylogeny. Continued examination of late Campanian through early Maastrichtian Pseudocorax, in particular, may yield more information on serration evolution within the genus. Expectations of a transitional taxon bridging the temporal gap between unserrated Pseudocorax laevis (Leriche, Reference Leriche1906) and serrated Pseudocorax affinis (Münster in Agassiz, Reference Agassiz1843) seem reasonable given the case of Carcharodon hubbelli Ehret et al., Reference Ehret, Macfadden, Jones, Devries, Foster and Salas-Gismondi2012 as a similar intermediate in the evolution of serrations in the Cenozoic Carcharodon lineage.
In a broader sense, Pseudocorax heteroserratus n. sp. highlights the importance of using large sample sizes to determine intraspecific variability, even within a singular tooth position. The new species remained undescribed up to the present study possibly due to insufficient sample size collection, as the taxon is an uncommon occurrence within Moroccan phosphates. Selachian odontology, especially when describing novel taxa using isolated teeth, benefits from as large a sample as possible.
Acknowledgments
We are immensely grateful for A. Stubbers (Alabama Materials Institute) and the Alabama Materials Institute for generously allowing the lead author to use their Keyence VHX-7000 digital microscope to create Pseudocorax figures. We also thank the father and daughter fossil hunters, B. and I. Chaibi, for collecting the sample used herein. G. Phillips, J.C. Sloan, C. Kroll, and anonymous reviewers provided insightful revisions to this manuscript.
Competing interests
The authors declare there are no competing interests.









 Open access
Open access