


Introduction
Decreasing the density of weed seeds in the soil seedbank reduces the number of emerged weed seedlings, which can improve overall weed management and decrease the potential for herbicide resistance (Neve et al. Reference Neve, Norsworthy, Smith and Zelaya2011). Due to their chemical and physical defenses, weed seeds in the seedbank can persist in soil for years (Davis et al. Reference Davis, Fu, Schutte, Berhow and Dalling2016; Houlihan et al. Reference Houlihan, Conlin and Chee-Sanford2019), resulting in continued seedling recruitment even when weed control is effective and seed rain is eliminated. Despite this, weed seeds are vulnerable to multiple modes of mortality in the soil, including natural aging, microbial infection, and predation by granivores (Long et al. Reference Long, Gorecki, Renton, Scott, Colville, Goggin, Commander, Westcott, Cherry and Finch-Savage2015). For example, it is estimated that up to 80% of seeds on the soil surface can be lost through seed predation (Bagavathiannan and Norsworthy Reference Bagavathiannan and Norsworthy2013), and pathogenic and/or saprophytic bacteria and fungi can cause up to 50% mortality of weed seeds in the soil (Chee-Sanford Reference Chee-Sanford2008; Davis et al. Reference Davis, Anderson, Hallett and Renner2006; Wagner and Mitschunas Reference Wagner and Mitschunas2008). Consequently, there are multiple incentives to discover agronomic management practices that reduce the number of viable weed seeds in the soil seedbank (Schwartz-Lazaro and Copes Reference Schwartz-Lazaro and Copes2019).
Crop rotations are the foundation of agronomic systems; therefore, understanding whether and to what extent crop species or communities affect rates of weed seed mortality could greatly enhance integrated weed management. For example, plant diversity and community composition may affect weed seed mortality through their effects on the soil microbial community (Wagner and Mitschunas Reference Wagner, Mitschunas, Upadhyaya, Clements and Shrestha2022). A metanalysis found that increasing plant diversity in natural terrestrial ecosystems was associated with greater microbial richness (Liu et al. Reference Liu, Zhu, Wurzburger and Zhang2020), and greater microbial diversity may increase overall microbial activity (Delgado-Baquerizo et al. Reference Delgado-Baquerizo, Maestre, Reich, Jeffries, Gaitan, Encinar, Berdugo, Campbell and Singh2016; Schmid et al. Reference Schmid, van Moorsel, Hahl, De Luca, De Deyn, Wagg, Niklaus and Schmid2021). Functionally distinct plant species may promote distinct soil microbes, through either litter inputs or root exudates, and potentially support greater microbial community diversity when planted in mixtures. For example, when soil microbiomes under mixtures and monocultures of four perennial forages were compared, the soil fungal community from the monoculture of the grass perennial ryegrass (Lolium perenne L.) was distinct from all other monocultures, while the forage mixtures exhibited a blending effect of the microbial communities from the individual monocultures (Fox et al. Reference Fox, Suter, Widmer and Lüscher2020). Greater microbial diversity and activity may allow for greater microbial degradation of weed seeds in the soil seedbank, as some microbes may be able to degrade seed defense chemicals (Davis et al. Reference Davis, Schutte, Iannuzzi and Renner2008; Görge and Werner Reference Görge and Werner1991), while others may be able to degrade compounds like lignin in seed coats (Liu et al. Reference Liu, Luo and Zheng2018; Ruiz-Dueñas and Martinez Reference Ruiz-Dueñas and Martínez2009).
Crop species identity and diversity could potentially affect seed mortality indirectly through effects on soil abiotic conditions. For example, canopy coverage and shading influences soil temperature, moisture, and nutrient availability (Lange et al. Reference Lange, Koller-France, Hildebrandt, Oelmann, Wilcke and Gleixner2019; Lozano-Parra et al. Reference Lozano-Parra, Pulido, Lozano-Fondón and Schnabel2018; Song et al. Reference Song, Zhou, Zhang, Li, Jin and Li2013), which may directly or indirectly affect weed seed mortality. Soil moisture predicted the mortality of velvetleaf (Abutilon theophrasti Medik.) and giant foxtail (Setaria faberi Herrm.) seeds (Schutte et al. Reference Schutte, Davis, Renner and Cardina2008). In warmer, wetter environments, seed physical defenses may be weakened and seed pathogen pressure is higher (Dalling et al. Reference Dalling, Davis, Schutte and Arnold2017), potentially increasing seed mortality. The crop plant community also influences soil nutrients, especially soil nitrogen (N), which may also impact weed seed mortality in the soil. Litter inputs to the soil from plant species varying in carbon:nitrogen (C:N) ratios can alter soil N availability and, potentially, microbial activity (Davis Reference Davis2007; Roumet et al. Reference Roumet, Birouste, Picon-Cochard, Ghestem, Osman, Vrignon-Brenas, Cao and Stokes2016). Davis et al. (Reference Davis, Anderson, Hallett and Renner2006) suggested a negative correlation between soil C:N ratio and weed seed mortality, which is supported by studies that have shown legume cover crops increased weed seed mortality, including for Amaranthus spp. (Mohler et al. Reference Mohler, Taylor, DiTommaso, Hahn and Bellinder2018) and common lambsquarters (Chenopodium album L.) (Hill et al. Reference Hill, Renner, Sprague and Davis2016). Conversely, increased soil N from legume crops may also stimulate weed seed germination, with either fatal or successful outcomes for the individual (Hill et al. Reference Hill, Renner, Sprague and Davis2016).
Perennial crops provide a unique model system for testing the impact of plant community composition and diversity on weed seed mortality, because they are often planted in diverse mixtures. Perennials are frequently used in crop rotations to break up life cycles of common annual weeds (Mohler et al. Reference Mohler, Liebman, Staver, Liebman, Mohler and Staver2001; Teasdale et al. Reference Teasdale, Mangum, Radhakrishnan and Cavigelli2004). The continual and often dense plant cover within perennial forages suppresses annual weed emergence, and the frequent mowing events terminate weeds before seed set, thereby eliminating new seed additions into the seedbank (Meiss et al. Reference Meiss, Médiène, Waldhardt, Caneill and Munier-Jolain2010; Nichols et al. Reference Nichols, Verhulst, Cox and Govaerts2015). Additionally, lack of soil disturbance in perennial systems allows weed seeds in the seedbank to remain undisturbed for years, possibly increasing their susceptibility to mortality if they are buried too deep to successfully germinate. However, it is not yet clear whether functionally distinct perennial forage species or overall forage diversity can affect weed seed mortality in the field, possibly through their effects on soil abiotic conditions or microbial communities infecting weed seeds.
In this study, we examined how monocultures and mixtures of legume, grass, and forb perennial forage crops and associated soil abiotic conditions affect Powell amaranth (Amaranthus powellii S. Watson) and A. theophrasti seed mortality and A. powellii microbial communities. We selected these weed species because A. powellii is a problematic and increasingly herbicide-resistant species (Aicklen et al. Reference Aicklen, Soltani, Tardif, Robinson, Laforest and Sikkema2022), and A. theophrasti is a persistent species in the weed seedbank due to its hard seed coat (Davis et al. Reference Davis, Fu, Schutte, Berhow and Dalling2016). To test whether perennial forage crops can affect seed mortality, we selected the perennials alfalfa (Medicago sativa L.), forage chicory (Cichorium intybus L.), and orchardgrass (Dactylis glomerata L.), because each has unique properties that may influence weed seed mortality and microbial communities (Figure 1). Because more diverse mixtures have diverse litter inputs and exudates entering the soil, which may affect both microbes and soil abiotic conditions, we hypothesized that increasing forage plant richness (number of species) would increase both weed seed mortality and microbial diversity within A. powellii seeds. Alternatively, we hypothesized that weed seed mortality would be greatest in treatments containing a high proportion of the legume (alfalfa), as previous studies have shown a link between legume crops and weed seed mortality (Hill et al. Reference Hill, Renner, Sprague and Davis2016; Mohler et al. Reference Mohler, Taylor, DiTommaso, Hahn and Bellinder2018). We anticipated that perennial forage treatment effects on seed mortality may be mediated through soil abiotic conditions (moisture, nutrients, pH, and temperature). Specifically, we expected that warmer, wetter soils with greater soil N (all of which can increase soil microbial activity) would be positively associated with weed seed mortality.
Photos and information about the three functionally distinct perennial forage species used within our perennial forage diversity gradient (Hall Reference Hall2000; Możdżeń et al. Reference Możdżeń, Barabasz-Krasny, Stachurska-Swakoń, Zandi, Puła, Wang and Turisova2020; Nishimura et al. Reference Nishimura, Kondo, Nagasaka and Satoh2000; Paglialunga et al. Reference Paglialunga, Proietti, Cardarelli, Moscatello, Colla and Battistelli2022; Undersander et al. Reference Undersander, Cosgrove, Cullen, Grau, Rice, Renz, Sheaffer, Shewmaker and Sulc2015; Veronesi et al. Reference Veronesi, Brummer, Huyghe, Boller, Posselt and Veronesi2010; Wang et al. Reference Wang, Xie, Wu, Chen, Wang, Cui, Hu and Wiatrak2012).

Materials and Methods
Experimental Design and Study Site
To evaluate the effect of perennial forage composition and richness on weed seed mortality in the soil, we established seven perennial forage treatments—including monocultures and all possible two- and the three-species mixtures of alfalfa, orchardgrass, and forage chicory—in a randomized complete block design with four blocks. Each plot was 12.2 by 12.2 m. The experiment took place at the Russell E. Larson Agricultural Research Center in Rock Springs, PA (40.717706, −77.946630, 350-m elevation). Soils are predominately comprised of Hagerstown silt loam (fine, mixed, mesic Typic Hapludalfs). Average air temperature at the site is 10.4 C, with average winter temperatures of −1.4 C and average summer temperatures of 21.3 C; the site receives on average 1,050 mm of annual precipitation (NCEI 2024).
Agronomic Management
Before planting, the field was chisel plowed and disked, and then a soil finisher and cultimulcher were used to prepare the seedbed. In September 2021, alfalfa, chicory, and orchardgrass were drill seeded on 19-cm rows (Table 1). Due to high winterkill, the chicory was reseeded in April 2022. We applied an equal low-level amount of N fertilizer across all treatments, despite chicory and orchardgrass not being legumes, to ensure any effects on seed mortality were due solely to the perennial forage treatments. In Spring 2022, we applied 30 kg N ha−1 equally across the entire field, and in Spring 2023, we increased the N amount we applied to 56 kg ha−1. Before planting and again in Spring 2023, we added P and K according to soil test recommendations. We applied Lambda cyhalothrin (Warrior II, Syngenta Crop Protection, Greensboro, NC) once in July 2022 at 0.034 kg ai ha−1 for control of leaf hoppers (Empoasca fabae). No herbicides were used throughout the study duration. The field was mowed four times per year when the alfalfa was at bud to early flowering stage, dried, and baled for hay.
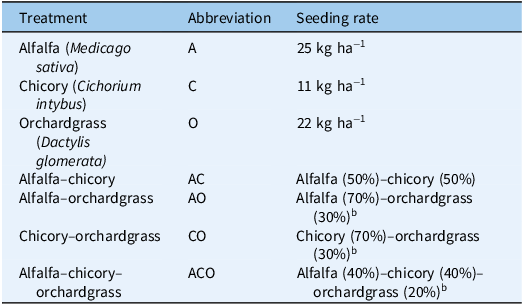
Seeding rates used for each perennial forage monoculture and mixture.a

aMixture percentages are based on the monoculture seeding rates.
bOrchardgrass was used at reduced rates in mixtures because of its highly competitive nature.
Forage Biomass Collection
Forage biomass was collected four times per year (immediately before the field being mowed for harvest) within two 0.25-m2 quadrats per plot, when alfalfa was at bud stage. Biomass was cut approximately 7.5 cm above the soil surface using electric hand shears, sorted to species, dried at 60 C, and weighed.
Weed Seed Mortality Assessment
Weed seeds of A. powellii and A. theophrasti were collected from adjacent fields in October 2021. We separated seed from chaff via an air column and kept the heavier fraction, so only mature seeds were used in the study. We also visually inspected A. theophrasti seeds and removed any seeds with signs of weevil damage. Seeds of each species (100 A. theophrasti and 150 A. powellii, only one species per bag) were combined with 10 g of sieved soil (2-mm sieve) into fine-mesh organza bags. The initial viability of A. theophrasti was approximately 70%, while the starting viability for A. powellii was approximately 97%. Bags were then sewn shut, and a hex nut was attached so bags could be located with a metal detector.
In October 2021, we buried weed seed bags at a 5-cm depth adjacent to the crop row. We buried 10 bags of each species per plot, and 2 bags per species were removed as subsamples for each of four seed extraction time points. We buried two extra bags per plot in case some bags were difficult to locate or in case a bag was ripped during retrieval from the soil. We first retrieved weed seed bags from the field at 1-yr post-burial and then subsequently at 6-mo intervals. These seed extraction time points refer to the length of time the seeds were buried within the soil of our perennial forage treatments and include: (1) 1 yr: Fall 2022; (2) 1.5 yr: Spring 2023; (3) 2 yr: Fall 2023; and (4) 2.5 yr: Spring 2024. At each of these seed extraction time points, we retrieved two seed bags of each species per plot, which were stored on ice and processed within 24 h. First, we pooled the two bags of each species collected from each plot and washed the contents of the seed and soil mixture in EtOH-sanitized sieves to remove soil (500-µM sieves). We counted all intact seeds per bag to obtain the proportion of the original seeds we recovered at each retrieval time point (% Recovery). We then randomly selected 100 A. powellii intact seeds for microbial analyses. These seeds were surface sterilized with 10% bleach for 10 min, followed by three rinses in sterile water (Ercoli et al. Reference Ercoli, Masoni, Pampana and Arduini2007), then frozen at −20 C.
After seeds were separated from soil, the remaining intact seeds not collected for microbial analyses were stored at 4 C for viability testing. In Fall 2022, we initially used the seed crush test (Sawma and Mohler Reference Sawma and Mohler2002) to quantify A. powellii viability, because we were hoping to save the crushed viable and nonviable seeds to evaluate how the seed microbial communities differed between the two. However, we ultimately determined that seed crush test viability estimates were unreliable and not comparable to the more established tetrazolium (TZ) staining method. Therefore, the Fall 2022 A. powellii seed mortality estimates were discarded from our dataset. For all subsequent A. powellii seed viability assessments, and for all A. theophrasti seed viability assessments, we conducted TZ testing according to the AOSA handbook (AOSA/SCST 2010), using 0.1% TZ with minor alterations: (1) A. powellii seeds were pierced at the tapered end of the seed instead of bisected for TZ staining and then later dissected to evaluate whether the embryo stained red; and (2) A. theophrasti seeds were scarified to quicken imbibition.
Total percent weed seed mortality (M total) was calculated as:
 ${M_{\rm total}} = \left[ {1 - \left( {{\rm{recovery\ proportion}} \times {\rm{viable\ proportion}}} \right)} \right]*100$
${M_{\rm total}} = \left[ {1 - \left( {{\rm{recovery\ proportion}} \times {\rm{viable\ proportion}}} \right)} \right]*100$
Microbial DNA Extraction, Library Preparation, and Amplicon Sequencing
Due to funding constraints, we only examined whether perennial forage treatments affect the microbiome of A. powellii seeds. Surface-sterilized A. powellii seeds were freeze-dried in sterilized microcentrifuge tubes covered with breathable sealing film in a freeze dryer for 10 h at −26 C and <500 mTorr.
We then used the Qiagen DNeasy Plant Mini Kit (Germantown, Maryland Qubit) to extract DNA of A. powellii seeds and their associated microbes. The Qiagen kit’s 20-mg restriction of biological tissue greatly limited the number of seeds for which we could extract DNA. Therefore, to obtain a more representative sample, we doubled both the weight of seed and the recommended extraction solution volume in which the seeds are ground. Therefore, we ground 40 mg of weed seeds in 750 μl of extraction solution with a bead mill. We then added the recommended amount of lysate (350 μl) to the Qiagen Shredder Column, and standard protocols were followed thereafter. Additionally, we extracted DNA from the original, unburied seeds collected in Fall 2021 (used to fill the seed bags) to determine a baseline of microbial endophytes. Negative controls to account for contaminants in the processing steps included (1) blank runs on each extraction kit as well as (2) extractions of sterilized metal beads that were subjected to the same protocols as the seeds (surface sterilization, freeze drying, weighing, etc.) and would indicate contaminants in processing steps. Extracted DNA was quantified using a Qubit Fluorometer (Singapore) (broad-spectrum kit), and all the negative controls had undetectable amounts of DNA.
Then, we performed the first of a two-step amplicon PCR process using universal primers with Illumina adapters to amplify the internal transcribed spacer (ITS) (1-2) marker region for fungal taxonomic diversity and the 16S rRNA gene (V4) region for bacterial taxonomic diversity (Supplementary Table 1). For the initial PCR reaction, we used ITS 1-2 primers (Gardes and Bruns Reference Gardes and Bruns1993; Illumina 2019) or 16S rRNA gene V4 515F and 806R (Apprill et al. Reference Apprill, McNally, Parsons and Weber2015; Parada et al. Reference Parada, Needham and Fuhrman2016), with Illumina adapters (Supplementary Table 1). For both ITS and 16S rRNA gene amplification, we used InvitrogenTM 2X PlatinumTM SuperfiTM II Green PCR Master Mix to run 20 μl PCR reactions containing: 10 μl Master Mix (1×), 0.5 μM each of forward and reverse primer, ∼15 ng of DNA, and sterile nuclease-free water to reach 20 μl (concentrations are listed as final concentrations). pPNA and mPNA blockers from PNA Bio were added to 16S rRNA gene reactions for a final concentration of 1.25 μM each to reduce amplification of plastids and mitochondria, respectively. The same PCR was conducted for both 16S rRNA and ITS and run on a thermocycler according to the Superfi II Master Mix protocol: initial denaturing (98 C for 30 s), denature–anneal–extend repeating 30 times (98 C for 10 s, 60 C for 10 s, 72 C for 15 s), final extension (72 C for 5 min), and a hold at 4 C (Invitrogen 2022). PCR blanks (PCR reagents without DNA) were also run to identify any contamination in the PCR reagents. PCR products were checked using a 2% agarose gel, and all controls had no visible amplification. Samples were then submitted to the Penn State University Huck Institute’s Genomics Core Facility, where a second round of PCR was completed to attach index barcodes for sample identification and Illumina sequence adapters, and then amplicons were sequenced (175k pair-end reads) with the Illumina NextSeq 2000 XLEAP P1 600 cycle kit and performing 300 by 300 paired-end sequencing.
Bioinformatic Analysis
We used the DADA2 pipeline (v. 3.18; Callahan et al. Reference Callahan, McMurdie, Rosen, Han, Johnson and Holmes2016) to filter, remove primers, trim reads (according to Phred score), pseudo-pool samples during sample inference, pair reads, and create amplicon sequence variant (ASV) abundance tables. For 16S rRNA gene data, after construction of the sequence table in DADA2, only paired ASVs with a length of 254 bp (the length of the V4 segment) were selected for downstream processing; selection of ASVs of the expected length (based on primers) is intended to remove noise from longer segments likely created from nonspecific priming (Callahan et al. Reference Callahan, McMurdie, Rosen, Han, Johnson and Holmes2016). The majority of the ASVs were exactly 254 bp. We used the GreenGenes2 database (v. 2024.09 genus level) (Callahan et al. Reference Callahan, McMurdie, Rosen, Han, Johnson and Holmes2016; McDonald et al. Reference McDonald, Jiang, Balaban, Cantrell, Zhu, Gonzalez, Morton, Nicolaou, Parks, Karst and Albertsen2024) for 16S rRNA gene taxonomic assignment and the UNITE All-Eukaryotes database (Abarenkov et al. Reference Abarenkov, Zirk, Piirmann, Pöhönen, Ivanov, Nilsson and Kõljalg2024) for ITS taxonomic assignment using pretrained classifiers (ref). Then, in phyloseq (v. 1.46), we removed all 16S rRNA gene ASVs not classified as bacteria, ASVs unclassified at the phylum level, and 16S rRNA gene ASVs associated with mitochondria or chloroplasts.
For ITS, we removed all non-fungal ASVs and all ASVs unclassified at the phylum level. Additionally, we removed poor-quality ITS ASVs containing 20-bp repetitions or longer homopolymers, which are common to ITS (Lindahl et al. Reference Lindahl, Nilsson, Tedersoo, Abarenkov, Carlsen, Kjøller, Kõljalg, Pennanen, Rosendahl, Stenlid and Kauserud2013). To eliminate the influence of rare ASVs in both 16S rRNA gene and ITS, we removed all ASVs that consisted of fewer than 20 total reads or found in less than 5% of samples. Both the 16S rRNA gene and ITS samples were independently rarefied to the minimum number of reads per sample using phyloseq (rarefy_even_depth function), which was 8,284 reads for 16S rRNA gene and 38,980 reads for ITS. We did this because rarefying helps obtain reliable and comparable estimates of alpha diversity across variation in sequencing depth (Weiss et al. Reference Weiss, Xu, Peddada, Amir, Bittinger, Gonzalez, Lozupone, Zaneveld, Vázquez-Baeza, Birmingham, Hyde and Knight2017). Rarefaction curves showed that our sequencing depth adequately captured diversity (Supplementary Figure 1). All downstream analyses occurred on rarefied datasets.
Measurement of Soil Abiotic Conditions
To understand whether the perennial forage treatments could indirectly affect seed mortality through their effects on soil abiotic conditions, we measured soil temperature, moisture, and nutrients in Spring 2023 through Spring 2024. We recorded soil moisture as volumetric water content (three readings per plot, which were averaged) each week during the 2023 growing season (early May to late October) using a HydroSense II (Logan, Utah, USA) soil probe; only three measurement time points were taken in 2024 because the last seed extraction time point was in May 2024. Soil temperature readings were collected hourly via HOBO sensors (Bourne, Massachusetts) (one per plot) that were buried at 5 cm below the soil surface (corresponding with the depth of the seed bags). The HOBO sensors were buried in plots in 2023 and removed in May 2024. During the winter between 2023 and 2024, five HOBO sensors malfunctioned, and therefore temperature data for these plots were removed from the dataset. We then calculated the daily average, maximum, and minimum temperature for each plot across the duration in which the HOBO sensors were deployed in the field. Soil samples were collected twice per year in Spring 2023, Fall 2023, and Spring 2024 for inorganic N analysis, but because other nutrients are less dynamic than N, we sampled soil once per year (Spring 2023 and 2024) for a full-spectrum Mehlich 3 inductively coupled plasma (ICP) analysis. We used a 2.5-cm-diameter corer for soil sampling and collected six soil samples per plot at a depth of 20 cm. Soil samples were then homogenized, and a portion was sent to the Penn State Ag Analytical Services Lab for the Mehlich 3 analysis. To quantify soil inorganic N concentration, we incubated 20 g of fresh soil with 100 ml 2 M KCl for 1 h on a shaker; then filtered extracts were frozen at −20 C until both NH4-N and NO3-N could be quantified based on the Berthelot and Greiss reactions, respectively (Kaye et al. Reference Kaye, Finney, White, Bradley, Schipanski, Alonso-Ayuso, Hunter, Burgess and Mejia2019).
Data Analysis
To examine the effects of perennial forage treatment, richness, or aboveground biomass, as well as soil abiotic conditions on weed seed mortality, we used generalized linear mixed-effects models (GLMM) using the glmmTMB package in R with a beta distribution, which is recommended for proportional data (Douma and Weedon Reference Douma and Weedon2019). Weed species were analyzed separately. Because we had repeated measurements of seed mortality, soil abiotic conditions, and forage biomass over time from each experimental unit of plot, plot was included in each model as a random effect. To determine whether we needed to account for temporal autocorrelation among repeated observations at the plot level, we compared corrected Akaike information criterion scores for models without including any autocorrelation covariance structure and one model in which we modeled the first-order autoregressive (ar1) structure across time at the plot level. However, we including temporal autocorrelation either resulted in failed convergence or did not improve model parsimony. Therefore autocorrelation structures were not retained in any of our final models. We used the emmeans package and applied a Tukey adjustment for pairwise comparisons to determine which perennial forage treatments, if any, differed from one another.
To examine whether forage treatment (treatment is defined as the seven combinations of the three forage species in monocultures or mixtures) affected weed seed mortality, treatment and time till seed extraction were fixed effects in the model. To examine whether forage richness affected weed seed mortality, the number of forage species (one, two, or three) within each treatment and seed extraction time point were fixed effects. To examine whether forage biomass (either total or by individual species) predicted weed seed mortality across our monoculture and mixture species, we used both species (or total) aboveground biomass and time till seed extraction as fixed effects in the model.
To determine how soil abiotic conditions influenced seed mortality, we included time to seed sample extraction and the soil abiotic factor as fixed effects in the model. Because the seeds were all buried at the same time but varied in their seed extraction time points, we averaged all measurements of the soil abiotic conditions within each plot up until a given seed extraction time point. For example, seeds that were extracted at the 2-yr time point also experienced the soil abiotic conditions measured at 1 and 1.5 yr. Therefore, we averaged soil measurements recorded for each plot up until the 2-yr time point, and this was regressed against the 2-yr mortality measurement. We did this for all measured soil abiotic conditions up until the seed extraction time point for each seed sample.
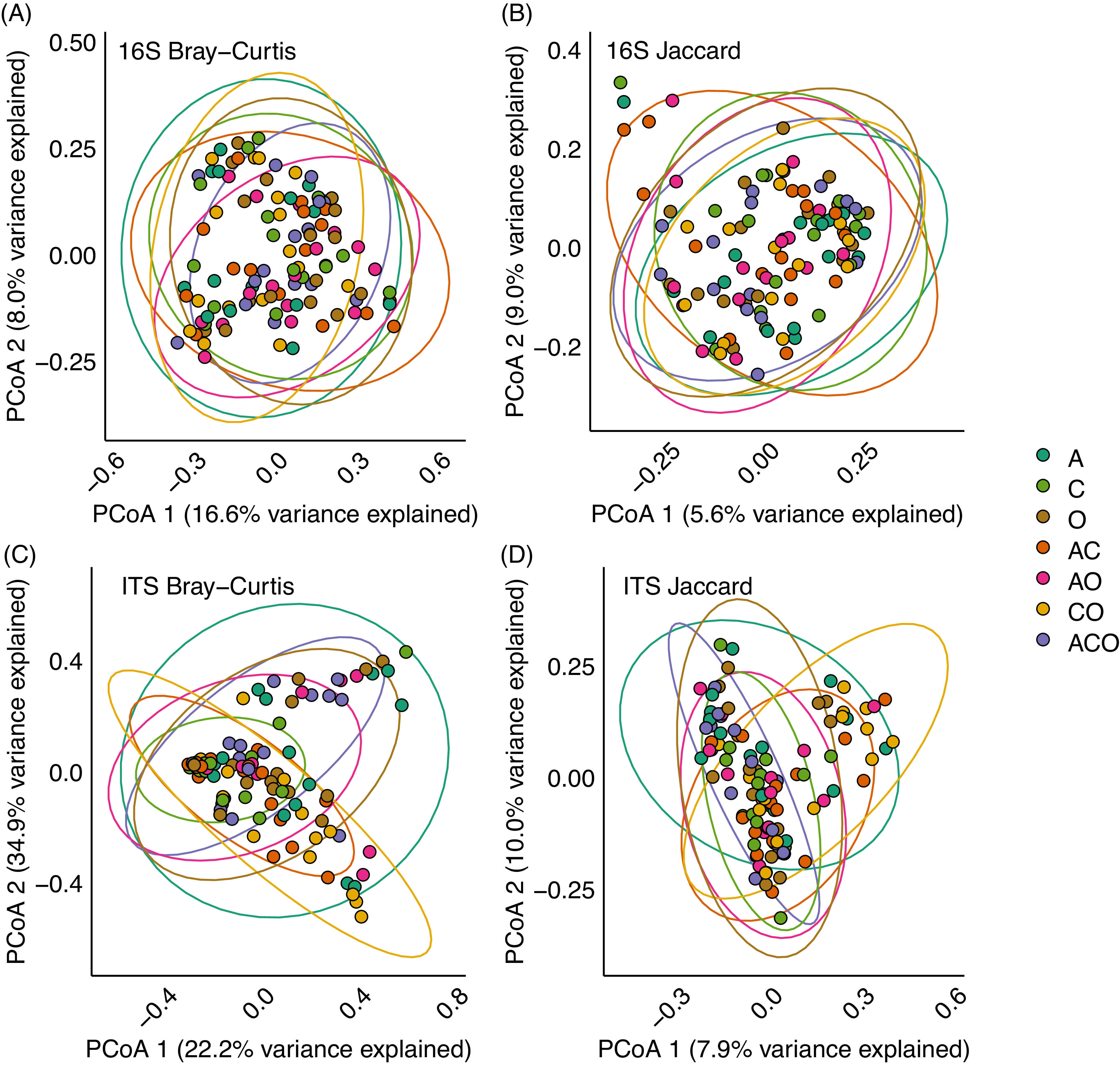
To examine whether the perennial forage treatment affected the A. powellii seed microbiome, we analyzed 16S rRNA gene and ITS data separately and performed a permutational multivariate analysis of variance on the Bray-Curtis dissimilarity matrix using the adonis2 function in the vegan package in R (Oksanen et al. Reference Oksanen, Simpson, Blanchet, Kindt, Legendre, Minchin, O’Hara, Solymos, Stevens, Szoecs, Wagner, Barbour, Bedward, Bolker and Borcard2025), and pairwise comparisons between treatments were performed with the pairwise.adonis function in the pairwiseAdonis package. To visualize A. powellii seed communities, we performed principal coordinates analysis (PCoA) on both the Bray-Curtis dissimilarity matrix and a Jaccard dissimilarity matrix (after converting the community matrix to presence–absence). Finally, we calculated the observed and Shannon diversity using the estimate_richness function in phyloseq, and we examined whether both forage richness and treatment affected both diversity metrics for both weed species using linear mixed-effect models with richness or treatment, retrieval time point, and their interaction as fixed effects and block as the random effect.
Results and Discussion
The mortality of both A. powellii and A. theophrasti increased dramatically between burial and the first seed extraction time point, which is common among weed seeds (Davis et al. Reference Davis, Fu, Schutte, Berhow and Dalling2016). After that, the average mortality remained overall consistent, yet with high variance around the mean (Supplementary Figure 2). For example, the initial mortality in the starting (pre-burial) seed lot of A. theophrasti was approximately 30% and then increased to 47% after 1 yr in the soil, after which it remained relatively constant. The initial A. powellii seed lot had only 3% mortality, which increased to 60% after 1.5 yr buried in the soil, and an average mortality of 70% after 2.5 yr of soil burial. The overall lower mortality in A. theophrasti compared with A. powellii was not surprising, because the hard protective seed coat of A. theophrasti allows the seed to persist longer in the seedbank compared with Amaranthus species (Davis et al. Reference Davis, Fu, Schutte, Berhow and Dalling2016).
The Perennial Forage Community Composition, but Not Diversity, Had Limited Effects on Weed Seed Mortality
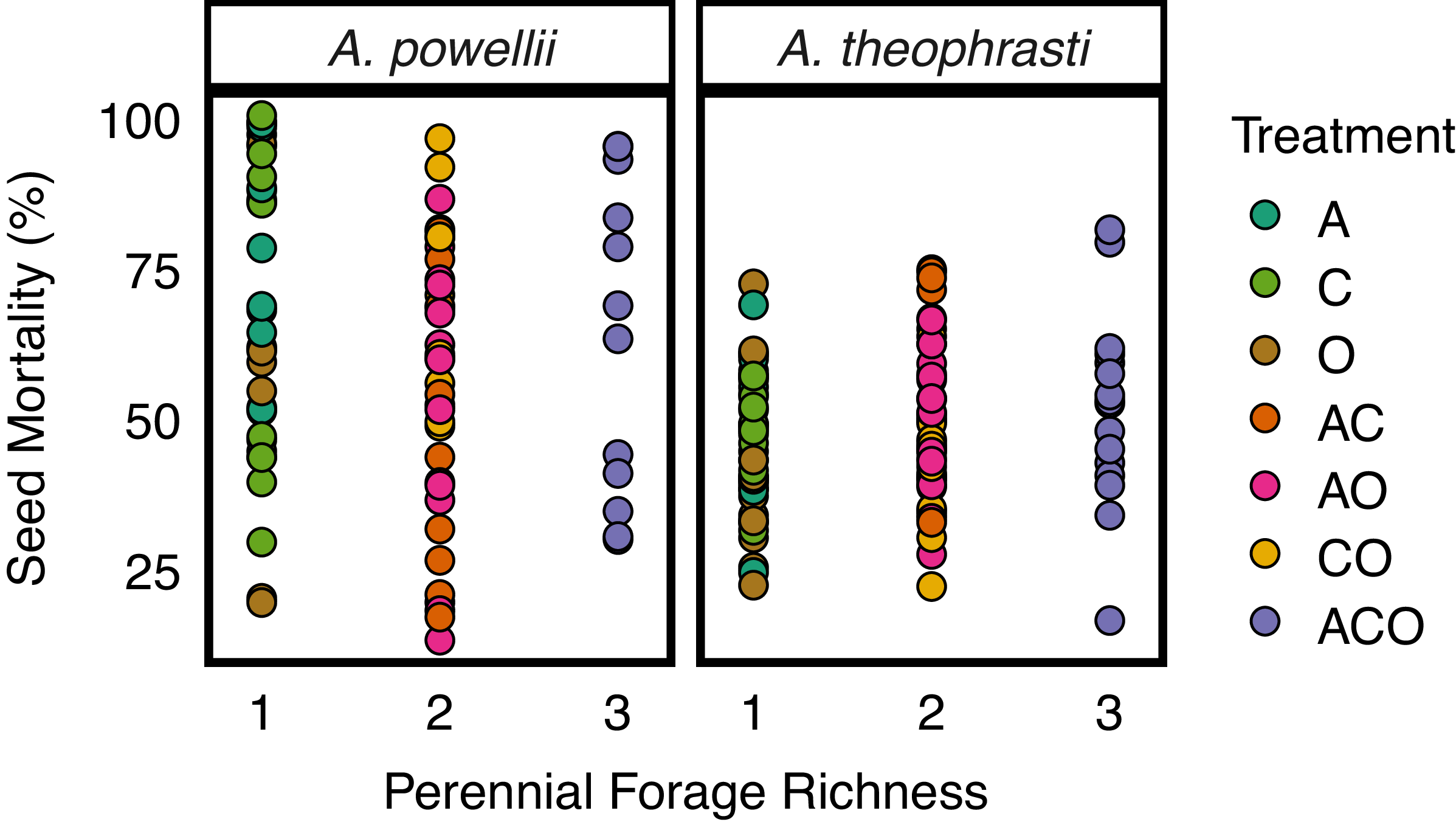
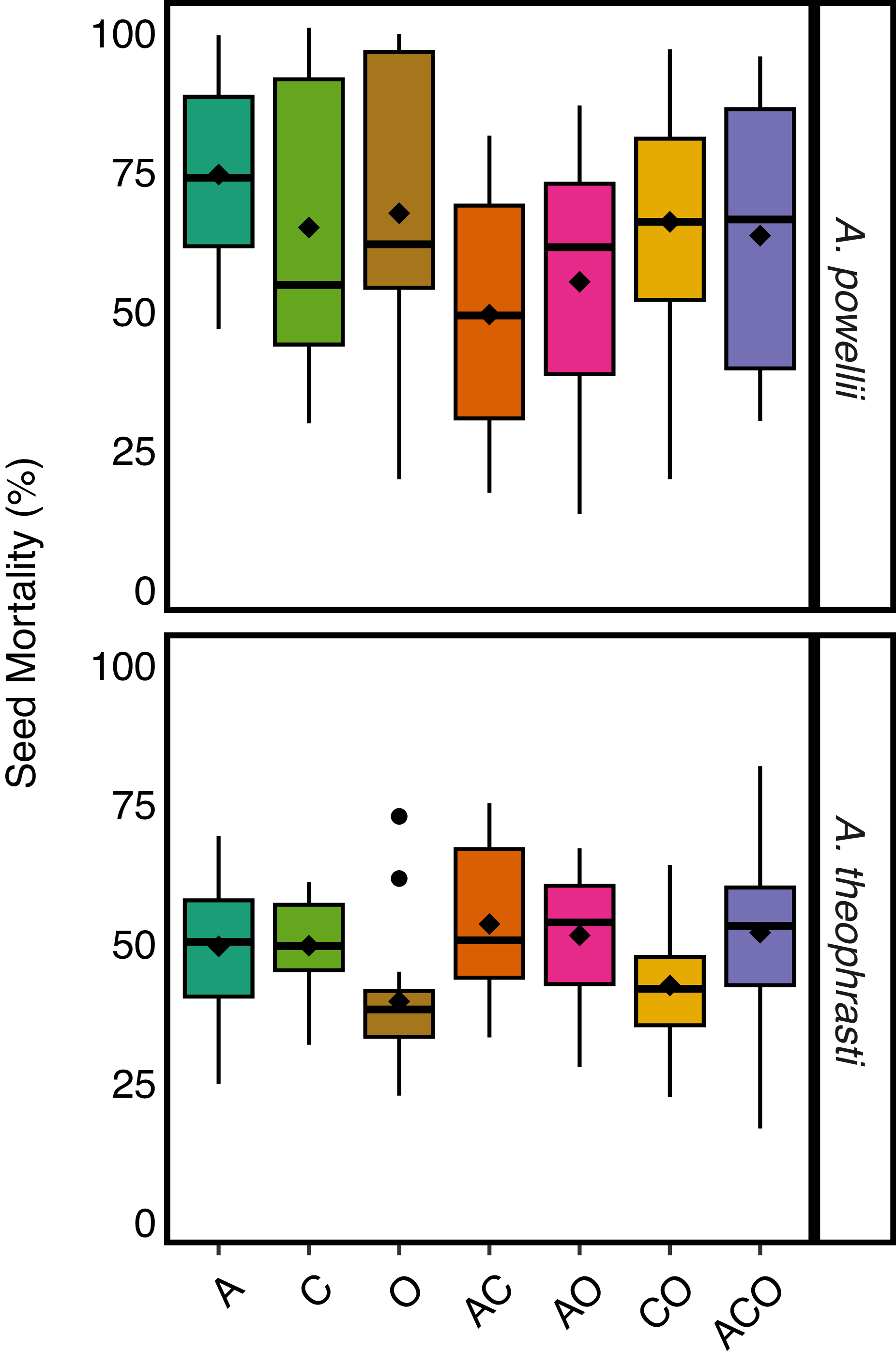
We found no support for our hypothesis that increasing perennial forage richness (number of perennial forage species) increased seed mortality, as we found no effect of perennial forage richness on either A. theophrasti (z-value = 1.5, P-value = 0.124) or A. powellii (z-value = −1.5, P-value = 0.137) seed mortality (Figure 2). However, we did find an effect of perennial forage treatment on A. theophrasti seed mortality (χ2 =17.7, P-value = 0.007; Figure 3), in which the alfalfa–chicory biculture resulted in greater seed mortality compared with the orchardgrass monoculture. Pairwise comparisons revealed no other treatments differed in their effect on A. theophrasti seed mortality. Interestingly, when we did a preplanned contrast comparing all treatments with alfalfa (alfalfa monoculture and the three mixtures with alfalfa) with all treatments without (chicory and orchardgrass monocultures and their biculture), we found higher A. theophrasti mortality in the treatments with alfalfa (z-statistic = −3.13, P = 0.001). We found no effect of treatment on A. powellii seed mortality (χ2 = 8.3, P-value = 0.216).
Effect of perennial forage richness (one, two, or three species) on seed mortality of Amaranthus powellii and Abutilon theophrasti. Treatments include alfalfa (A), chicory (C), orchardgrass (O), and all combinations of the two- and three-species mixtures.

Effect of perennial forage treatments on seed mortality of Amaranthus powellii and Abutilon theophrasti buried in the soil within the perennial forage treatments. Treatments include alfalfa (A), chicory (C), orchardgrass (O), and all combinations of the two- and three-species mixtures.

We also explored whether variation in forage biomass (either total biomass or biomass of an individual species) was associated with weed seed mortality. Consistent with our hypothesis, we found a trend that increasing alfalfa biomass within our monocultures and mixtures was associated with greater A. theophrasti seed mortality (z-statistic = 1.95, P = 0.051, Supplementary Figure 3). Interestingly, we also found that total forage biomass was positively associated with A. theophrasti seed mortality (z-statistic = 2.29, P-value = 0.022). Because we found the lowest A. theophrasti seed mortality in the orchardgrass monoculture, we had expected to find a negative association between orchardgrass biomass and A. theophrasti seed mortality, but this was not the case (z-statistic = −1.33, P-value = 0.184). We also found no association between forage chicory biomass and A. theophrasti seed mortality. Finally, we found no association between A. powellii seed mortality and alfalfa, orchardgrass, chicory, or total forage biomass (Supplementary Figure 4).
Principal coordinate analysis (PCoA) on Bray-Curtis (A and C) or Jaccard (B and D) dissimilarities of bacterial (A and B) and fungal (C and D) community composition in Amaranthus powellii seeds buried in perennial forage monocultures and mixtures. Treatments include alfalfa (A), chicory (C), orchardgrass (O), and all combinations of the two- and three-species mixtures.

Some, but Not All, Soil Abiotic Conditions Were Associated with Seed Mortality
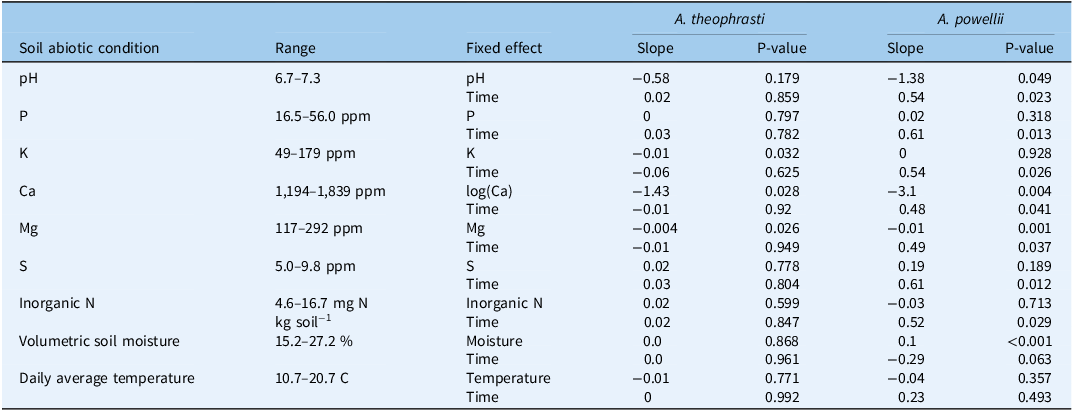
We had expected that soil abiotic conditions would explain the effect of our perennial forage treatments on weed seed mortality. Despite finding limited and inconsistent effects of our perennial forage communities on seed mortality, variation in some, but not all, soil abiotic conditions predicted weed seed mortality. Unexpectedly, soil cations were the strongest soil predictors of seed mortality. Calcium (Ca), potassium (K), and magnesium (Mg) concentrations were negatively associated with A. theophrasti seed mortality (Table 2, P = 0.028, P = 0.032, and P = 0.026, respectively), while Ca and Mg were negatively associated with A. powellii seed mortality (P = 0.004 and P = 0.001, respectively). Interestingly, we found the lowest levels of K, Ca, and Mg in the alfalfa monoculture (Supplementary Table 2), which also consistently had relatively high levels of mortality (Figure 3). Alfalfa is known to heavily utilize and deplete soil K (Kelling Reference Kelling2000), and because the alfalfa monoculture and mixtures were higher yielding, K cations were likely removed from the soil at a higher rate (Arnall Reference Arnall2024). Similarly, we found the lowest A. theophrasti mortality in the orchardgrass monoculture (Figure 3), which had relatively high levels of soil K and Mg (Supplementary Table 2). To the best of our knowledge, no one has directly manipulated soil cation concentrations, such as Ca, Mg, and K, to understand their effects on seed mortality. Previous work found that seed mortality was greatest in soil rich in soil organic matter with higher Ca and Mg concentration compared with a more sandy and red clay soil (Long et al. Reference Long, Steadman, Panetta and Adkins2009). However, they attributed soil type effects on seed mortality to variation in soil moisture and temperature.
Results from generalized linear mixed-effect models testing whether soil abiotic conditions and time till seed extraction predict mortality of Abutilon theophrasti and Amaranthus powellii seeds

We had expected that warmer, wetter soil conditions would favor seed mortality, which has been demonstrated in previous studies (Long et al. Reference Long, Steadman, Panetta and Adkins2009). Soil moisture was positively associated with A. powellii seed mortality (p = 0.001), but not with A. theophrasti seed mortality. A. theophrasti has a hard, persistent seed coat that prevents the seed from imbibing water (Warwick and Black Reference Warwick and Black1988), which may explain a lack of an effect of soil moisture on seed mortality. In contrast, neither soil temperature nor soil inorganic N was associated with A. powellii or A. theophrasti seed mortality. Yet interestingly, soil temperature and moisture were both highest in the orchardgrass monoculture, which had the lowest levels of A. theophrasti mortality.
Multiple previous studies have found greater seed mortality resulting from legumes (Davis et al. Reference Davis, Anderson, Hallett and Renner2006; Hill et al. Reference Hill, Renner, Sprague and Davis2016; Mohler et al. Reference Mohler, Taylor, DiTommaso, Hahn and Bellinder2018), and in soils with lower C:N ratios (Pakeman et al. Reference Pakeman, Small and Torvell2012), presumably due to greater soil N. Therefore, we expected that the perennial forage communities dominated by alfalfa would experience greater seed mortality. We did find a positive association between alfalfa biomass and A. theophrasti mortality. Yet while the alfalfa monoculture consistently had relatively high levels of A. powellii mortality, this trend was not significant, and we did not detect a consistent effect of soil inorganic N on weed seed mortality. However, we found relatively low levels of soil inorganic N concentrations within our system (between 4.6 and 16.7 mg N kg−1 soil; Table 2), which may have limited our ability to detect associations with seed mortality. Most of the previous work examining legume cover crop effects on seed mortality were conducted after the legumes were incorporated into the soil (Hill et al. Reference Hill, Renner, Sprague and Davis2016; Mohler et al. Reference Mohler, Taylor, DiTommaso, Hahn and Bellinder2018). In our perennial forage system, we did not incorporate plant tissue into the soil. Senesced root tissue and fallen leaf litter at harvest were the only legume tissue soil additions, potentially limiting the effect of alfalfa on soil inorganic N concentrations.
Perennial Forage Treatments Had No Effect on Amaranthus powellii Seed Microbiomes
We found no support for our hypothesis that perennial forage community would affect the microbiome within A. powellii seeds, as neither bacterial (F = 1.16, P = 0.06) nor fungal (F = 1.38, P = 0.07) communities varied across the perennial forage monocultures, bicultures, or the three-way mixture (Figure 4). Despite finding a marginal effect of treatment on both bacterial and fungal communities (P-values between 0.05 and 0.1), we found no evidence that any two treatments differed from one another when we conducted pairwise comparisons.
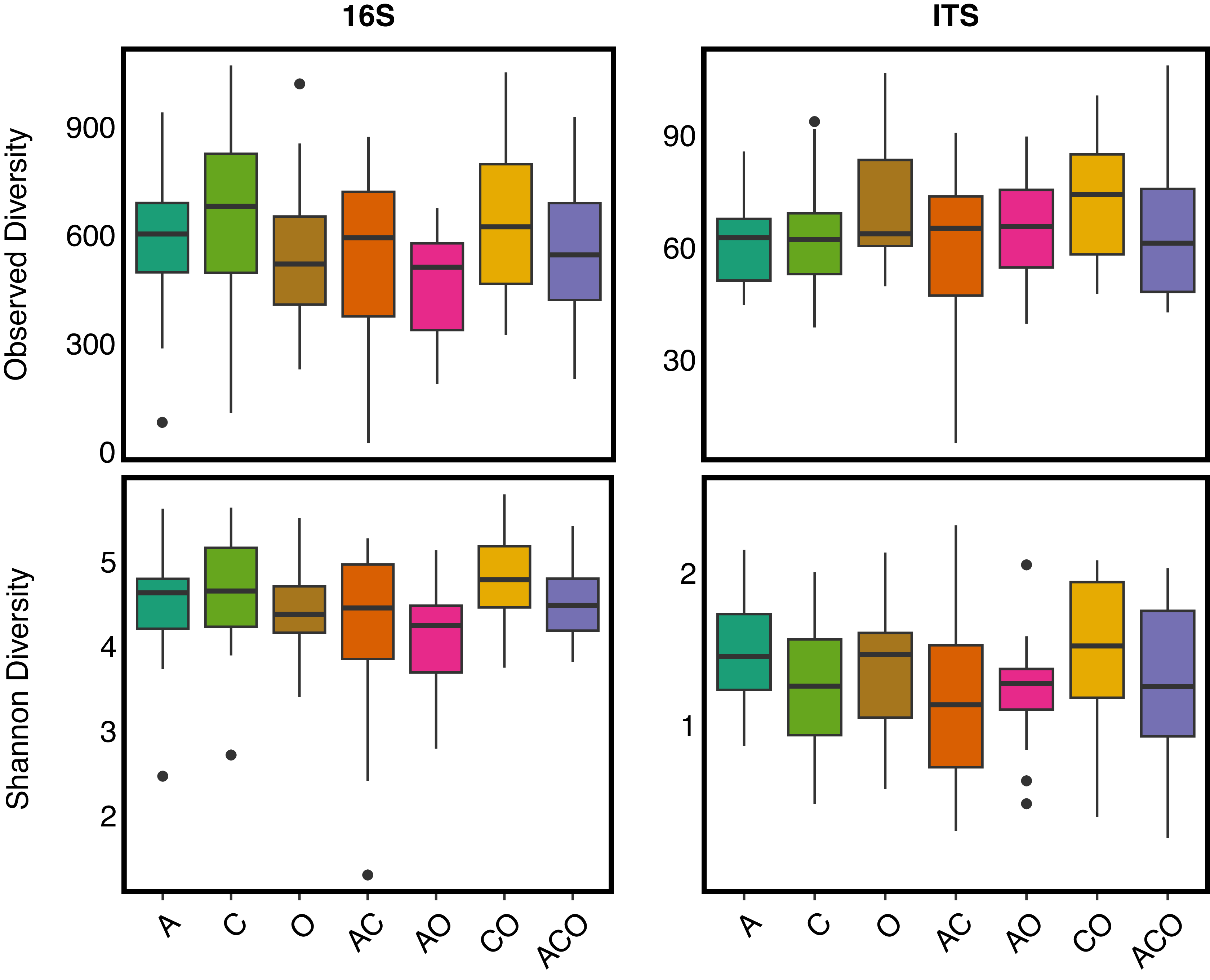
We also found no support for our hypothesis that increased perennial forage richness (i.e., richness) would increase microbial diversity within A. powellii seeds (Figure 5). Instead, we found that perennial forage richness had no effect on Shannon or observed diversity of bacterial or fungal microbial communities within A. powellii seeds (P-value > 0.1). We also found no effect of perennial forage treatment on either bacterial observed or fungal Shannon or observed diversity in A. powellii seeds (Supplementary Table 3).
Observed and Shannon diversity of bacterial (16S) and fungal communities (ITS) in Amaranthus powellii seeds buried in perennial forage monocultures and mixtures. Treatments include alfalfa (A), chicory (C), orchardgrass (O), and all combinations of the two- and three-species mixtures.

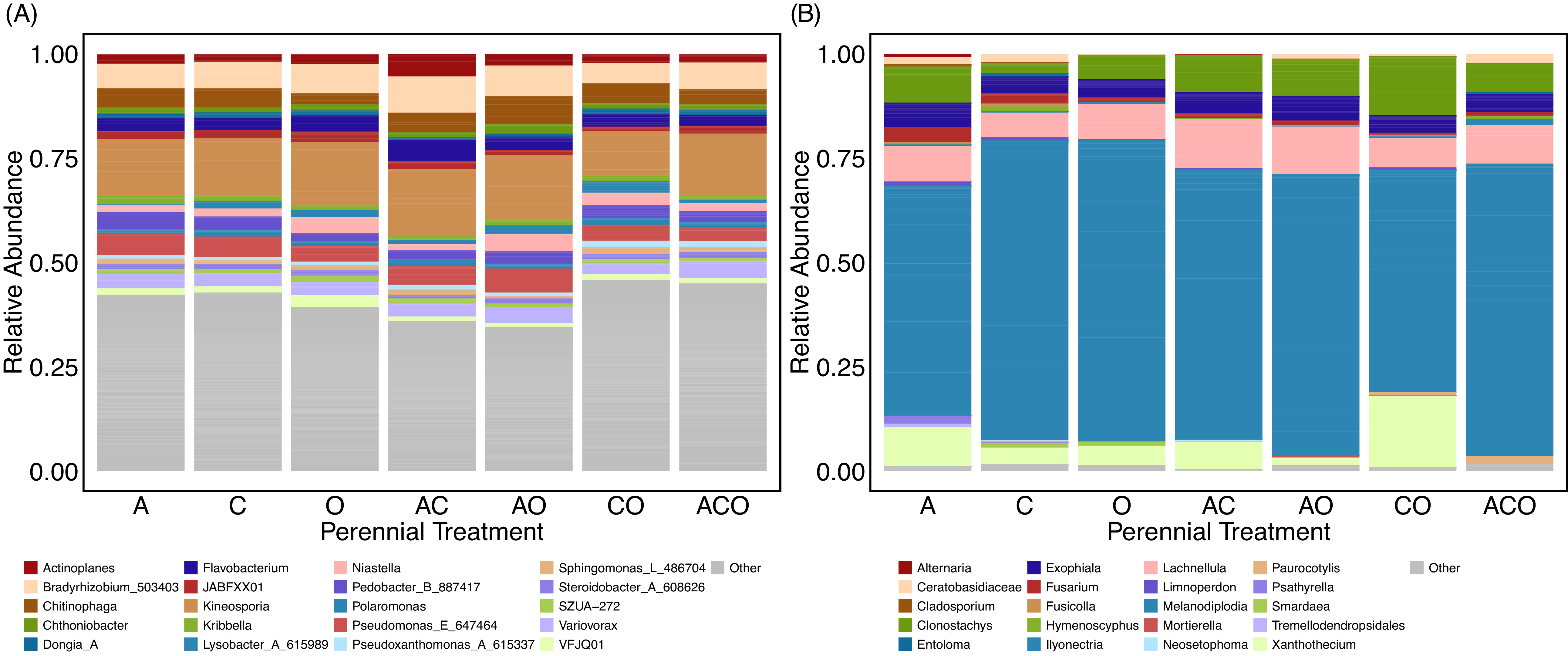
As reported in previous studies, many bacterial genera found in A. powellii seeds were in the Pseudomonadota, Actinomycetota, or Bacteroidota phyla (Acuña et al. Reference Acuña, Hu, Inostroza, Valenzuela, Perez, Epstein, Sessitsch, Qian Zhang and Jorquera2025; Johnston-Monje et al. Reference Johnston-Monje, Gutiérrez and Lopez-Lavalle2021; Figure 6). The most abundant Actinomycetota included the genera Kineosporia, Actinoplanes, and Kribbellam. The Bacteroidota phylum included Chitinophaga, whose species may break down chitin in cell walls (Fernandes et al. Reference Fernandes, Sierra, Brear, Pereira and Lemos2021; Kobayashi and Crouch Reference Kobayashi and Crouch2009), and Flavobacterium, which has been found to have both pathogenic and plant growth–promoting functions (Seo et al. Reference Seo, Kim, Lee and Lee2024). Finally, Pseudomonadota also included genera that could be both pathogenic and plant growth promoters. For example, the known plant pathogen Pseudomonas fluorescens D7 is a soil-borne pathogen and has previously been tested as a biocontrol agent for the invasive plant cheatgrass (Bromus tectorum L.) (Dooley and Beckstead Reference Dooley and Beckstead2010; Kennedy et al. Reference Kennedy, Young, Elliott and Douglas1991). In contrast, Bradyrhizobium is a genus of known plant growth–promoting bacteria often associated with the rhizosphere, but which can also be a seed endophyte, especially in legumes (Mataranyika et al. Reference Mataranyika, Bez, Venturi, Chimwamurombe and Uzabakiriho2024). Variovorax has also been reported to improve plant health, including increasing wheat germination under stress (Acuña et al. Reference Acuña, Rilling, Inostroza, Zhang, Wick, Angela Sessitsch and Jorquera2024). Other Pseudomonadota genera are often characterized as saprotrophs, including Lysobacter, a genus containing species that release lytic enzymes to compete for space with other microbes (Fernandes et al. Reference Fernandes, Sierra, Brear, Pereira and Lemos2021; Lin et al. Reference Lin, Yang, Tao, Shen, Cui, Wang, Wang, Jing, Qian and Shao2023), but can also protect against pathogens (Kobayashi and Crouch Reference Kobayashi and Crouch2009).
Relative abundance of the top 20 most abundant genera of (A) bacterial and (B) fungal within Amaranthus powellii seed buried in the perennial forage treatments, which include alfalfa (A), chicory (C), orchardgrass (O), and all combinations of the two- and three-species mixtures.

The most abundant phyla of fungi within A. powellii seeds were Ascomycota and Basidiomycota (Figure 6). Overwhelmingly, the most abundant genus was Melanodiplodia, followed by Lachnellula (both Ascomycota); these are saprotrophs that degrade wood and other recalcitrant materials (Põlme et al. Reference Põlme, Abarenkov, Henrik Nilsson, Lindahl, Clemmensen, Kauserud, Nguyen, Kjøller, Bates, Baldrian and Frøslev2020). Multiple Ascomycota pathogenic fungi were also found in A. powellii seeds, including Fusarium, Alternaria, Ilyonectria, and Cladosporium, genera that have been associated with weed seed colonization and seed decay (Androsiuk et al. Reference Androsiuk, Okorski, Paukszto, Jastrzębski, Ciesielski and Pszczółkowska2022; Pitty et al. Reference Pitty, Staniforth and Tiffany1987; Põlme et al. Reference Põlme, Abarenkov, Henrik Nilsson, Lindahl, Clemmensen, Kauserud, Nguyen, Kjøller, Bates, Baldrian and Frøslev2020). The most abundant Basidiomycota were not yet classified into genera, but came from the Ceratobasidiaceae family, which includes both plant pathogens and saprotrophs and has been previously found in orchid [Gymnadenia conopsea (L.) R. Br.] seeds during germination (Zhao et al. Reference Zhao, Yang, Wang, Qian, Ding, Jacquemyn and Xing2024).
Thus far, few studies have examined the microbial communities in weeds seeds after being colonized with soil microbes. While it is not yet clear which taxa, if any, may have resulted in or contributed to seed death, results from our study may help develop future hypotheses about which soil microbes can accelerate seed mortality in the soil. Ultimately, understanding which soil microbes result in weed seed mortality and which management practices help build up those microbial populations in the soil would greatly contribute toward regulating seedbank densities of weed populations.
Implications and Future Directions
While perennial phases within crop rotations reduce the density of problematic annual weeds within the soil seedbank (Mohler et al. Reference Mohler, Liebman, Staver, Liebman, Mohler and Staver2001), our study provides some, but limited evidence that distinct perennial forage communities may vary in their ability to promote seed mortality in the soil. We were surprised we did not see more of a perennial forage community effect on seed mortality. However, there are multiple conditions that could have limited our ability to detect any effect of our perennial forage treatments on weed seed mortality or seed microbial communities. First, within a plot, the forage crop stand was quite uneven, which likely contributed to greater soil heterogeneity within a plot, making it harder to detect differences due to our perennial forage treatments. Uneven forage stands also enabled weed infestations that likely altered the effects of our plant community treatments (see Supplementary Figure 5 for forage and weed biomass across time). For example, grass weeds (many Poa spp.) established in the alfalfa plots (especially the alfalfa monocultures), and white clover (Trifolium repens L., a legume) was prevalent in the chicory monocultures. As these weed species represent different plant functional groups, they may have caused confounding effects on soil biotic and abiotic properties, which in turn, may have diluted treatment effects. Finally, we managed all perennial forage treatments in the same way by adding a consistent low level of N fertilizer equally, instead of individually optimizing the management of each individual species or mixture to maximize productivity. Therefore, we may have obtained different results if we had individually managed each forage mixture or monoculture according to management recommendations.
Our study also suggests that the effects of distinct forage communities on seed mortality are likely mediated through effects on soil conditions. For example, we found frequent negative associations between soil cation concentration (K, Ca, and Mg) and weed seed mortality. Few studies have directly manipulated soil abiotic conditions (soil nutrients, soil moisture, and temperature) to examine their effects on seed mortality in soil (Davis Reference Davis2007). However, our work shows the potential for seed mortality to be accelerated through soil fertility management. Understanding whether specific soil amendments accelerate weed seed mortality would improve weed seedbank management, and future work should explicitly test which soil abiotic conditions and amendments influence the mortality of seeds in the soil seedbank.
Additionally, varying plant communities, such as perennial forages, may exhibit more of an effect on seed mortality and seed microbiomes after plant litter is incorporated into the soil, which could be a potentially fruitful avenue for future work. Finally, studies examining the microbiomes of dormant seeds are limited, so future work examining fundamental questions such as how seed mortality and microbiomes vary across soil depths, or soils ranging in texture and structure, could greatly increase our understanding of how to leverage soil microbes to accelerate weed seed mortality in the soil.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/wsc.2026.10116
Acknowledgments
This project would not have been possible without the technical, laboratory, and field assistance from Russell E. Larson farm staff (Corey Dillon, Al Cook, and George Dills), Lowry Lab members (especially David Long, Allie Rice, and Beth Eckert). Thank you to the Spring 2025 Lowry Lab for helpful comments and edits.
Funding statement
This research was financially supported by the Weed Science Society of America (WSSA) through the WSSA Innovative Grants Program and the NE Sustainable Agriculture Research and Education (SARE) Graduate Student Research Grant GNE24-308. This work was also supported by the USDA National Institute of Food and Agriculture and Hatch Appropriations under projects PEN04759 (accession no. 1025327) and PEN04949 (accession no. 7006508).
Competing interests
The authors declare no conflicts of interest.




 Open access
Open access