


Introduction
The Baja California (“Baja” hereafter) peninsula is rich in biodiversity and endemic species thanks to its unique geological history and physical setting. The peninsula was created when initiation of the San Andreas transform system ∼12 million years ago rifted the sliver of land that was to become Baja California from the Mexican mainland and began transporting it to the northwest (Atwater and Stock, Reference Atwater and Stock2010), eventually leading to the marine flooding of the Gulf of California 6 million years ago (Darin et al., Reference Darin, Dorsey, Niemi, Salgado Munoz, Henry, Pecha, Riggs, Putirka and Wakabayashi2025). Subsequent uplift and block faulting created an irregular spine of mountain ranges that runs down the peninsula (Gastil et al., Reference Gastil, Phillips and Allison1975). Geographically isolated, the narrow peninsula stretches over 1200 km from the Transverse Ranges of California at 32°45′N to south of the Tropic of Cancer at 22°50′N in the Cape Region (Garcillán et al., Reference Garcillán, González-Abraham and Ezcurra2010). Along with its complex topography and latitudinal range, the peninsula’s location between the colder Pacific Ocean and warm Gulf of California results in dramatic northwest–southeast variability in climate, especially the amount and seasonality of precipitation (González-Abraham et al., Reference González-Abraham, Garcillán and Ezcurra2010). Varied substrates, topographies, and climates in turn support a wide array of coastal sage scrub, desertscrub, chaparral, and dry coniferous forest, inspiring competing biogeographic regionalization schemes for Baja California (González-Abraham et al., Reference González-Abraham, Garcillán and Ezcurra2010; Morrone Reference Morrone2021).
In the region around Cataviña in central Baja (29⁰48′N, 114⁰46′W; Fig. 1), vegetation transitions from Sonoran Desert (Chaparral and Lower Colorado River Valley Desert subdivisions) (Brown, Reference Brown1994; González-Abraham et al., Reference González-Abraham, Garcillán and Ezcurra2010; Morrone, Reference Morrone2021) into what has been variously called the Vizcaíno Subdivision of the Sonoran Desert (Shreve, Reference Shreve1951; Brown, Reference Brown1994), Vizcaíno Desert (Zippin and Vanderwier, Reference Zippin and Vanderwier1994), or Central Desert (Wiggins, Reference Wiggins1960; Garcillán and Ezcurra, Reference Garcillán and Ezcurra2003; González-Abraham et al., Reference González-Abraham, Garcillán and Ezcurra2010; Morrone, Reference Morrone2021). We will use Central Desert herein. Importantly, the transitional area between these two divisions has long been recognized for its diverse and endemic flora (Shreve, Reference Shreve1936, Reference Shreve, Shreve and Wiggins1964; Peinado et al., Reference Peinado, Alcaraz, Aguirre, Delgadillo and Aguado1995) and includes ∼1300 species of vascular plants, 900 of which are endemic to the peninsula. Covering only one-fifth of the surface area, the transition between the Lower Colorado Valley Desert and the Central Desert hosts half of the species and three-fourths of the vascular plant genera in Baja California (Peinado et al., Reference Peinado, Alcaraz, Aguirre, Delgadillo and Aguado1995; for an annotated checklist, refer to Rebman et al., Reference Rebman and Gibson2016).
Map of northern and central Baja California, Mexico, showing the current midden study area at Cataviña and previous midden records in Baja California and the Sonoran Desert mainland (SSPM = Sierra San Pedro Mártir). The dashed lines show land that would have been exposed with a 100 m drop in sea level during the last glacial. The map was compiled and drafted by hand from 1:500,000 scale topographic sheets prepared by the United States War Department, Military Intelligence Division, Geographic Branch (1936). Physical maps are archived with the North American Packrat Midden Collection at the University of Arizona’s Desert Laboratory in Tucson, AZ.

During glacial–interglacial cycles, changes in temperature and precipitation resulted in expansion and contraction of species’ ranges and the reshuffling of communities in the North American deserts, including Baja California (Betancourt et al., Reference Betancourt, Van Devender and Martin1990; Becklin et al., Reference Becklin, Betancourt, Braasch, Dézerald, Díaz, González and Harbert2024). In addition, during glacial periods lower global sea levels exposed extensive areas of continental shelf that could have been colonized by terrestrial plant and animal communities. During the last glacial maximum ∼21,000 years ago, sea level was ∼100–120 m lower than today and exposed 20,000–30,000 km2 of land in the Gulf of California (Fig. 1) that is now submerged (Durham and Allison, Reference Durham and Allison1960; Stanford et al., Reference Stanford, Hemingway, Rohling, Challenor, Medina-Elizalde and Lester2011).
Much of what is known about late Quaternary ecological dynamics in arid and semi-arid environments in the Americas comes from fossil rodent middens (Betancourt et al., Reference Betancourt, Van Devender and Martin1990; Betancourt, Reference Betancourt, Lomolino and Heaney2004) and middens hold great potential for addressing emerging ecological and evolutionary questions (Becklin et al., Reference Becklin, Betancourt, Braasch, Dézerald, Díaz, González and Harbert2024). In North America, members of the genus Neotoma (packrats or woodrats) produce middens, deposits containing abundant plant remains, fecal pellets, pollen, insects, and bones. These materials are encased in a rock-hard crystallized urine matrix that can preserve the middens for tens of thousands of years in dry rock shelters and crevices. Middens contain copious material for radiocarbon dating and allow for high taxonomic resolution, with plant identification often possible at the level of species or even subspecies, which is critical for discerning endemic flora. Abundant and well-preserved plant and animal remains in packrat middens lend themselves to a battery of morphological, geochemical, and genetic analyses (e.g., Becklin et al., Reference Becklin, Medeiros, Sale and Ward2014; Holmgren et al., Reference Holmgren, Hunter and Betancourt2019; Smith et al., Reference Smith, Kaminski, Kanda, Sweet, Betancourt, Holmgren, Hempel, Alberti and M2021). Regional arrays of midden series enable detailed studies of plant movements in time and space (Norris et al., Reference Norris, Betancourt and Jackson2016), including trait-based analyses of migration velocities for different species (Butterfield et al., Reference Butterfield, Anderson, Holmgren and Betancourt2019a, Reference Butterfield, Holmgren, Anderson and Betancourt2019b).
Despite being the mainstay of vegetation reconstruction in the North and South American deserts during the late Quaternary (Becklin et al., Reference Becklin, Betancourt, Braasch, Dézerald, Díaz, González and Harbert2024), surprisingly few middens have been studied in Baja California. Most midden studies in the Sonoran Desert (Fig. 1) have been concentrated on the mainland (Cole, Reference Cole1986; Van Devender, Reference Van Devender1987, Reference Van Devender, Burgess and Felger1990; Van Devender et al., Reference Van Devender, Burgess and Felger1990, Reference Van Devender, Burgess, Piper and Turner1994; Anderson and Van Devender, Reference Anderson and Van Devender1995). In Baja, two lengthy and detailed midden series are available from the Sierra San Pedro Mártir (Holmgren et al., Reference Holmgren, Betancourt and Rylander2011) and the Sierra Juárez (Holmgren et al., Reference Holmgren, Betancourt, Peñalba, Delgadillo, Zuravnsky, Hunter, Rylander and Weiss2014) in northeastern Baja (Fig. 1). These studies document the Pleistocene occurrence of chaparral and pinyon–juniper–oak woodland that include some desert elements in what is now desertscrub.
Pleistocene midden assemblages containing chaparral and woodland species with some desert elements at low elevations (150–300 m above sea level [m asl]) along the eastern piedmont of the Sierra Juárez refute the long-held idea that the lowlands in the Lower Colorado River Basin may have served as a glacial-aged refugium harboring an intact assemblage of Sonoran Desert flora (Holmgren et al., Reference Holmgren, Betancourt, Peñalba, Delgadillo, Zuravnsky, Hunter, Rylander and Weiss2014). Less is known about the Lower Colorado–Central Desert transition area in central Baja from which only a few middens had been analyzed prior to this study. These include a single 12,070 cal yr BP midden from the Sierra San Francisco (Rhode, Reference Rhode2002), an 11,740 cal yr BP midden near Misión San Fernando, and a 21,175 cal yr BP (Wells, Reference Wells2000) and 1680 cal yr BP midden from Cataviña (Clark and Sankey, Reference Clark and Sankey1999; Sankey et al., Reference Sankey, Van Devender and Clark2001) (Fig. 1).
The existence of Pleistocene middens in these boulder fields has long been known (Wells, Reference Wells and Everett1986, Reference Wells2000; Van Devender, Reference Van Devender, Betancourt, Van Devender and Martin1990; Peñalba and Van Devender, Reference Peñalba and Van Devender1998; Clark and Sankey, Reference Clark and Sankey1999; Sankey et al., Reference Sankey, Van Devender and Clark2001). Here, we present a new 40,000-year midden series from tonalite boulder fields near Cataviña in central Baja (Fig. 1). This series offers some insights about the biogeographic and climatic history of the Central Desert.
Materials and methods
Study area
The Central Desert, where Cataviña is located, is marked by a sharp gradient in rainfall amount and seasonality, with the strictly winter rains in northwestern Baja contrasting with bimodal precipitation in central Baja. At equivalent elevations, northwestern Baja receives three times the mean annual precipitation of central Baja. This contrast demonstrates the greater orographic precipitation that the northern Peninsular Range batholith intercepts from westerly frontal air masses from the significantly cooler and wetter Pacific compared to the lower, discontinuous volcanic ranges in central Baja (Gastil et al., Reference Gastil, Phillips and Allison1975; Minnich et al., Reference Minnich, Franco-Vizcaíno, Goforth, Wehncke, Ruben Lara-Lara, Alvarez-Borrego and Ezcurra2014). Northeastern Baja is the driest and hottest region of the peninsula as a result of strong air subsidence and the Sierra Juárez and Sierra San Pedro Mártir blocking moisture from the Pacific (Hastings and Turner, Reference Hastings and Turner1965). What little precipitation there is in the northeast is distributed nearly equally in winter and summer. The central Baja peninsula around San Luis, Cataviña, and San Ignacio experiences bimodal precipitation, with rainfall peaks in the late summer/early fall (July–October) and in winter/early spring (November–March), separated by particularly dry months in late spring/early summer (April–June) (Wilson, Reference Wilson2002; Bullock, Reference Bullock2003). Warm season precipitation is derived from the North American monsoon, while much of the fall precipitation comes from occasional tropical storms or “chubascos” that either make landfall or interact with low-pressure troughs or cutoff lows to steer moisture inland (Adams and Comrie, Reference Adams and Comrie1997; Bullock, Reference Bullock2003; Corbosiero et al., Reference Corbosiero, Dickinson and Bosart2009). Further to the south, Baja California’s Cape Region has low winter precipitation and a summer rainfall peak caused partly by the summer monsoon and tropical storm activity (Adams and Comrie, Reference Adams and Comrie1997; Stensrud et al., Reference Stensrud, Gall and Nordquist1997). Winter precipitation in northern and central Baja increases during El Niño events (Minnich et al., Reference Minnich, Franco-Vizcaíno and Dezzani2000; Wilson Reference Wilson2002).
At our Cataviña study area, mean annual temperature is ∼ 19°C with occasionally freezing temperatures, such as in the winter of 1989–1990 (Clark and Sankey, Reference Clark and Sankey1999). Mean annual precipitation at Cataviña averages ∼120–130 mm, with ∼50% falling November–February and ∼ 40% in July–October. Wet winters happen during El Niño events, as experienced during our midden collection trip to Cataviña in February 1998. At Laguna Seca Chapala, 60 km south of Cataviña, copious winter rains filled the usually dry playa during the El Niño events of 1965–1966, 1968–1969, 1972–1973, 1976–1977, 1982–1983, 1991–1993, and 1997–1998 (Eugenio Grosso, Rancho Chapala, oral communication, 1998).
The extensive tonalite boulder fields around Cataviña are a well-known and scenic landmark along Route 1, the main peninsular highway. The tonalite is part of a Cretaceous pluton in the Peninsular Ranges Batholith at the northern end of the Jaraguay Block in central Baja and forms topographic lows with surrounding mesas capped by Pliocene basalts (Gastil et al., Reference Gastil, Phillips and Allison1975; Conca and Rossman, Reference Conca and Rossman1985). In the Cataviña boulder fields, cavernous weathering forms numerous caves, crevices, and rock shelters that harbor and preserve middens. Modern vegetation in these low-elevation boulder piles is characterized by the shrubs Larrea tridentata, Ambrosia dumosa, Ambrosia chenopodiifolia, the cacti Cylindropuntia cholla, Cylindropuntia ganderi, Cylindropuntia molesta, Pachycereus pringlei, Lophocereus schottii, Ferocactus gracilis, and the trees Fouquieria columnaris, Pachycormus discolor, and Neltuma odorata [Prosopis glandulosa var. torreyana] (Fig. 2). A list of all plant taxa mentioned in the text and figures, along with their common names, can be found in Table 1. Plant taxonomic names follow the Jepson Flora Project (2025). For species not contained within Jepson, names follow Plants of the World Online (2025).
Example of endemic flora in close proximity to Cataviña middens, featuring Boojum tree (Fouquiera columnaris), Baja elephant tree (Pachycormus discolor), and Cardón (Pachycereus pringlei). Photograph courtesy of Treve Johnson Photography.

Scientific and common names of plant taxa mentioned in the text and figures.

Midden collection and analysis
In 1989, 1990, and 1998, 30 separate packrat middens were collected from 640 m to 680 m elevation within a 5 km2 area of the Cataviña boulder fields. Plants within 50 m of the midden sites were identified and herbarium specimens collected. Herbarium specimens were deposited into the herbaria at the University of Arizona, Arizona State University, New York Botanical Garden, University of Texas, Universidad de Sonora, and San Diego Natural History Museum.
Middens were processed according to well-established methods (Spaulding et al., Reference Spaulding, Betancourt, Croft, Cole, Betancourt, Van Devender and Martin1990; Holmgren et al., Reference Holmgren, Hornsby, Becklin, Betancourt, Díaz, González and Latorre2023). Hammers and rock chisels were used to free cemented middens in the field, remove the outer weathering rind of modern contaminants, and separate any layers along bedding planes into discrete units to be processed and analyzed separately. Indurated midden sample weights, prior to soaking in a closed plastic bucket, ranged from 360 g to 2400 g.
In the laboratory, a small subsample for pollen analysis was removed before the bulk of the sample was placed in a bucket of water to dissolve the crystallized urine matrix. After wet screening and drying in a low temperature oven, plant macrofossils were identified using a binocular stereomicroscope and an extensive reference collection, both at the University of Arizona’s Desert Laboratory and Herbarium. A random 50 g subsample was selected from each sample and sorted for macrofloral, bone, and insect remains using a 35× zoom binocular stereoscope. In total, 100 plant taxa were identified from plant macrofossils in the middens. Plant macrofossils were quantified using a relative abundance scale of 0 to 5, where 0 = 0 fragments; 1 = 1 fragment; 2 = 2–25 fragments; 2.5 = 26–50 fragments; 3 = 51–75 fragments; 3.5 = 76–100 fragments; 4 = 101–150 fragments; 4.5 = 151–199 fragments; and 5 = >200 fragments.
Pollen samples were analyzed for 22 of the middens (Table 1) at the Universidad de Sonora in Hermosillo, Sonora, Mexico following standard methodology (Fægri and Iversen, Reference Faegri and Iversen1989; Brown, Reference Brown2008). A small, indurated subsample (∼10–20 g) was removed from the original indurated sample for pollen analysis, and 10 g were later separated and soaked in distilled water. Pollen slides were observed under 400× magnification. At least 300 grains per sample were counted, and the rest of the slide was scanned for poorly represented types. Pollen determinations were made based on published works on pollen morphology, atlases, and an extensive Sonoran Desert pollen reference collection. A total of 92 different pollen types, mostly including family and genus level, were identified in midden pollen assemblages. All metadata and plant macrofossil (https://data.neotomadb.org/datasets/66731) and fossil pollen (https://data.neotomadb.org/datasets/66274) identifications, abundances, and percentages are archived in the Neotoma Paleoecology Database (Williams et al., Reference Williams, Grimm, Blois, Charles, Davis, Goring and Graham2018).
Middens were radiocarbon dated and treated as discontinuous snapshots of vegetation from the site to form a time series (Table 2). Dates were calibrated using the CALIB 8.2 program IntCal20 curve (Stuiver and Reimer, Reference Stuiver and Reimer1993). The oldest midden (>42,460) returned an infinite date, indicating it was beyond the limit of radiocarbon dating. All other dates are plotted and reported herein as the midpoint of calibrated 2-sigma age ranges. Midpoints were rounded to the nearest year when the standard error was <50 yr and to the nearest 10 yr when the standard error was between 50 yr and 1000 yr.
Packrat midden site data and radiocarbon dates from Cataviña, central Baja California, Mexico. The A-prefix dates were submitted and reported by the University of Arizona Radiocarbon Lab (Tucson, Arizona) in 1990; GX-prefix dates were reported by Geochronology Laboratories in 1998; the Beta-prefix date was submitted and reported by Beta Analytic (Miami, Florida) in 1989.

Results
Radiocarbon dating
Middens from Cataviña range in age from >42,460 to 140 cal yr BP (Table 2). Three middens (>42,460, 28,110, and 25,900 cal yr BP) date from the Pleistocene. Because the >42,460 BP date was infinite, it should be considered a minimum age only. There is a 14,000 yr gap with no dated middens between the Pleistocene and Holocene middens. Eight middens date from the early Holocene (11,660–8490 cal yr BP), documenting changes associated with the glacial–interglacial transition. Another 19 middens span the middle to late Holocene. The Cataviña record provides unusually even coverage from the end of the Pleistocene through the late Holocene, with only three gaps longer than ∼660 years in the Holocene.
Vegetation record
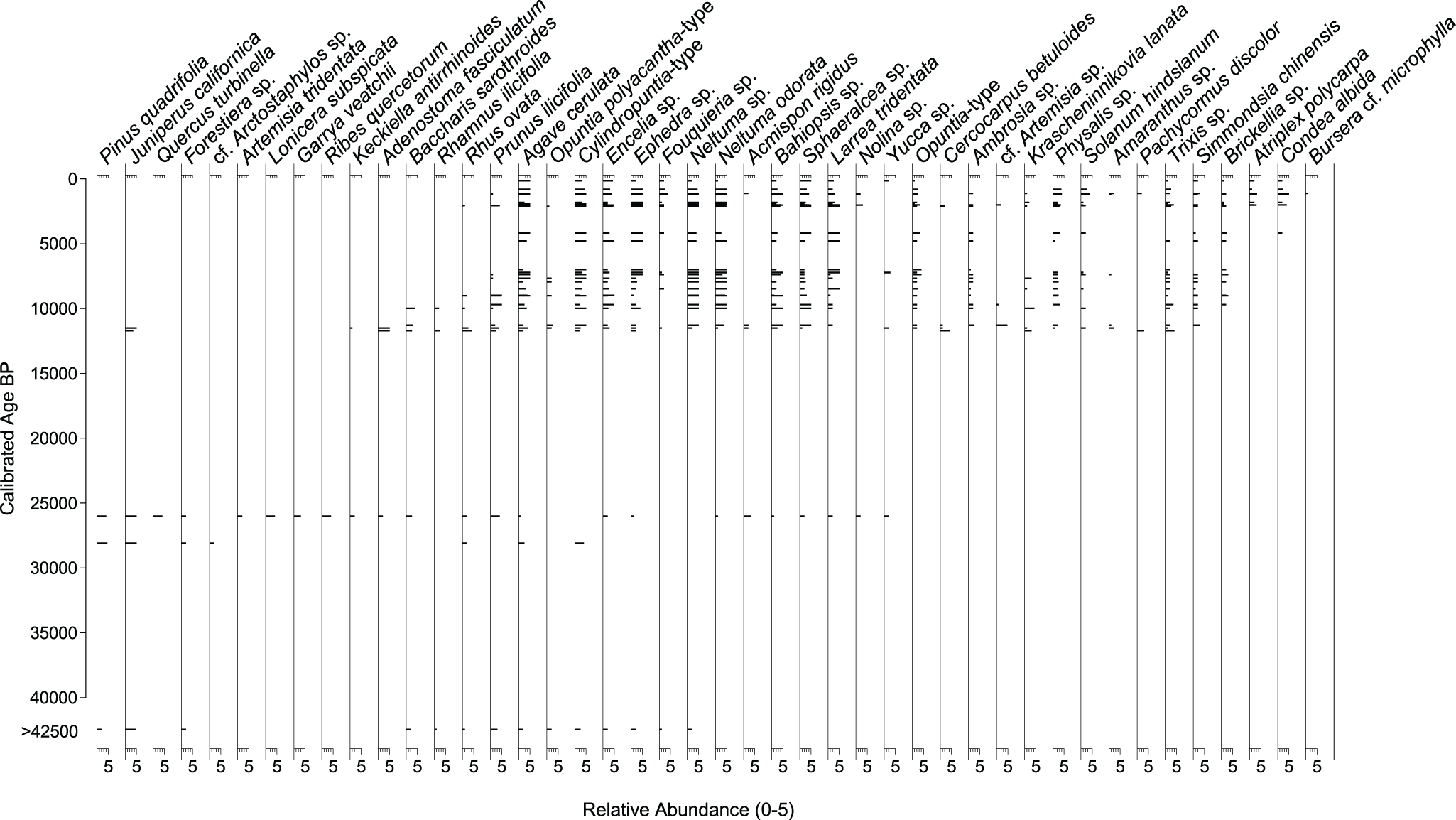
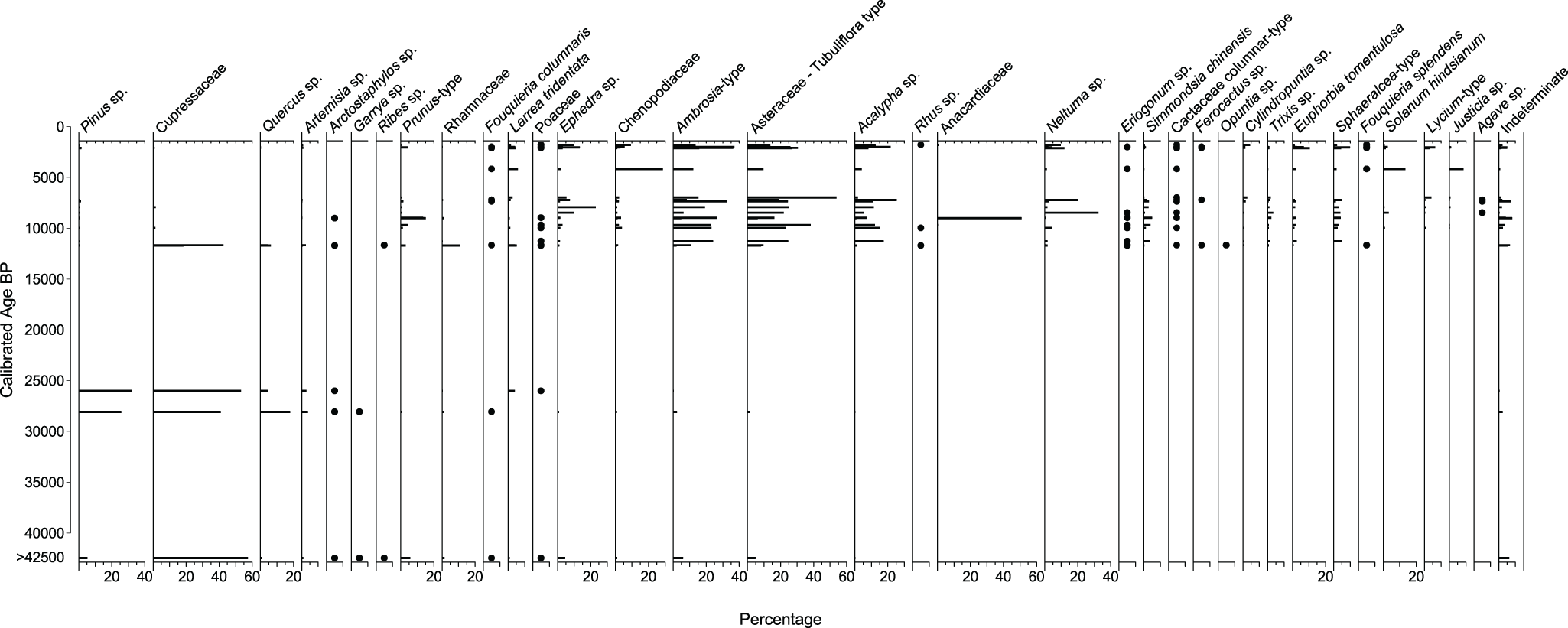
The oldest midden (>42,460 cal yr BP) may be distinguished from full glacial middens (28,110, and 25,900 cal yr BP) by its lower abundance of Pinus quadrifolia and absence of Quercus turbinella and other chaparral shrubs in the macrofossil assemblage, along with the high Juniperus percentage (60%), and moderated frequencies of Pinus, Prunus, Ephedra, and Asteraceae in the pollen assemblage (Figs. 3 and 4). The full glacial middens indicate a rich chaparral/woodland assemblage marked by abundant macrofossils of P. quadrifolia, Juniperus sp. and several chaparral shrubs (cf. Arctostaphylos, Lonicera, Garrya, Ribes, Adenostoma, Keckiella, Rhamnus), and by the co-dominance of Juniperus, Pinus, and Quercus in the pollen assemblages.
Plant macrofossil abundance through time for selected taxa from packrat middens from Cataviña, central Baja California, Mexico.

Pollen percentages through time showing the relative frequencies for selected taxa from packrat middens from Cataviña, central Baja California, Mexico.

Pinus quadrifolia is a sister species to P monophylla due to shared ancestral polymorphism or introgression (Buck et al., Reference Buck, Hyasat, Hossfeld and Flores-Rentería2020; Montes et al., Reference Montes, Peláez, Moreno‐Letelier and Gernandt2022). P. quadrifolia occurs between 1100 and 2000 m asl from the San Jacinto Mountains in southern California to the Sierra San Pedro Mártir in Baja California Norte, where it is the dominant pinyon on the western slope; a southern outlier population also occurs in the Sierra San Francisco in Baja California Sur. At its highest elevations in the Sierra San Pedro Mártir, P. quadrifolia occurs with P jeffreyi. At its mid elevations (∼1500–1600 m asl), P. quadrifolia grows in dense stands. At its lower elevations, P. quadrifolia occurs with another pinyon, P. californiarum, which replaces it at the lowest and driest elevations (Earle, Reference Earle2025a). P. californiarum occurs both in the Sierra Juárez and Sierra de San Pedro Mártir, mostly along their dry eastern escarpments. The nearest P. californiarum stands to Cataviña occur about 75 km northeast near the southern end of the Sierra San Pedro Mártir and 100 km southeast in the Sierra La Asamblea (Earle, Reference Earle2025b).
The Juniperus macrofossils in the full glacial middens are probably J. californica because this is the only juniper species currently found on the Baja peninsula. Although not present at the midden site today, J. californica is distributed from the USA–Mexico border to south of Sierra San Borja (Zanoni and Adams, Reference Zanoni and Adams1973) and isolated patches are known from the Central Desert at an elevation of 1000–1200 m (Minnich et al., Reference Minnich, Franco-Vizcaíno, Goforth, Wehncke, Ruben Lara-Lara, Alvarez-Borrego and Ezcurra2014). Wells (Reference Wells2000) notes a single individual growing at 610 m, 200 m from where he collected an early Holocene midden containing J. californica macrofossils, in a volcanic outcrop northwest of Mission San Fernando. J. californica was also found in glacial-age middens to the north (Holmgren et al., Reference Holmgren, Betancourt and Rylander2011, Reference Holmgren, Betancourt, Peñalba, Delgadillo, Zuravnsky, Hunter, Rylander and Weiss2014) and further to the south in the Sierra San Francisco (Rhode, Reference Rhode2002). Similarly, the Cupressaceae pollen almost certainly represents Juniperus given the copious macrofossils, although transport of Cupressus or Libocedrus pollen (taxa found today in the Sierra de San Pedro Mártir region) cannot be ruled out.
Macrofossils of Q. turbinella, another woodland tree species, are found only in the midden dating to 25,900 cal yr BP. Quercus pollen, on the other hand, is present in all full glacial middens, although it is represented by only two grains in the oldest midden. The presence of macrofossils provides clear evidence that a species was growing locally at a site because woodrats only forage within ∼30–50 m of their dens (Finley, Reference Finley, Betancourt, Van Devender and Martin1990; Vaughan, Reference Vaughan, Betancourt, Van Devender and Martin1990), whereas pollen may become incorporated into middens via the regional pollen rain. Taken together, the macrofossils and pollen indicate the presence of chaparral and woodland with at least the occasional Q. turbinella growing in the vicinity.
In addition to these now extralocal tree species (i.e., species that no longer occur at the site), there are several shrubs found only in the Pleistocene middens. Those represented by macrofossils include the chaparral shrubs Forestiera sp., cf. Arctostaphylos, Artemisia tridentata, Lonicera subspicata, Garrya veatchii, Ribes quercetorum, Keckiella antirrhinoides, Adenostoma fasciculatum, and Rhamnus ilicifolia. Similarly, pollen from Garrya and Ribes are found exclusively in the Pleistocene, while Arctostaphylos pollen is last found in the 9020 cal yr BP midden. Along with the tree species discussed earlier, these extralocal species are characteristic of more mesic pinyon–juniper–shrub oak woodland and/or chaparral and provide additional evidence for cooler, wetter conditions during the Pleistocene.
Some desert species are also first found in the full glacial middens either as macrofossils or pollen and persist into the late Holocene, including Agave cerulata, Cylindropuntia-type and Opuntia polyacantha-type, Encelia sp., Fouquieria columnaris, Larrea tridentata, and Neltuma odorata. Today, the endemic F. columnaris is found between 27.48ºN and 30.43ºN latitude, roughly from San Quintín to Volcán las Tres Vírgenes, with Cataviña in the northern part of its range (Webb et al., Reference Webb, Salazar-Ceseña and Turner2014). A. cerulata is another Baja endemic that has occupied Cataviña near its northern limit (Turner et al., Reference Turner, Bowers and Burgess1995) for at least the last 40,000 years. Other species including Acmispon rigidus, Nolina sp., Sphaeralcea sp., Bahiopsis sp., and Yucca sp. first appear in the Pleistocene middens and tend to increase in abundance during the Holocene.
We acknowledge that significant vegetation shifts probably happened during the 14,000-year gap between our full glacial and early Holocene (11,660–8490 cal yr BP) middens. During the early Holocene, extralocal woodland species decline, while other species first appear or increase in abundance in the midden sequence. After 11,630 cal yr BP, Pinus, Juniperus, and Quercus disappear from the macrofossil record and are present only at very low pollen percentages (<2%). These low percentages of anemophilous (wind-pollinated) species likely indicate longer-distance transport of pollen rather than continued local presence, especially considering the disappearance of these species from the macrofossil record. In the early Holocene, new species begin to appear including Acalypha sp., Amaranthus sp., Ambrosia sp., Artemisia sp., Brickellia sp., Cercocarpus betuloides, Eriogonum sp., Euphorbia tomentulosa, Fouquieria splendens, Krascheninnikovia lanata, Pachycormus discolor, Physalis sp., Simmondsia chinensis, Solanum hindsianum, and Trixis sp. More recent arrivals include Condea albida (4170 cal yr BP), Atriplex polycarpa (1981 cal yr BP), and Bursera cf. microphylla (1110 cal yr BP), all of which are common today on gravelly plains, washes, and rocky desert hillslopes. Right after 11,630 cal yr BP, both plant macrofossil (Fig. 3) and pollen (Fig. 4) assemblages show immediate shifts to highly diverse desert plant communities.
Discussion
Like other packrat midden records from Baja, the record from Cataviña suggests that chaparral and woodland were much more extensive during the last glacial into the early Holocene. At Cataviña, California chaparral and/or pinyon–juniper–oak woodland extended down to at least 600 m (see contour in Fig. 1) and probably 100–200 m lower throughout central Baja during the late Pleistocene (Wells, Reference Wells2000). Today, more extensive California chaparral is found from California, USA, to the southern extremity of the Sierra San Pedro Mártir; it is only found as isolated stands in montane “sky islands” above ∼1500 m or as small outlying populations at lower elevations between 30°N and 27.5°N in central Baja (Wells, Reference Wells2000; Minnich et al., Reference Minnich, Franco-Vizcaíno, Goforth, Wehncke, Ruben Lara-Lara, Alvarez-Borrego and Ezcurra2014). Scattered patches of Juniperus californica have been found today near Cataviña on granitic slopes (Wells Reference Wells2000), as well as on a basalt mesa at ∼1000 m along with the chaparral species Rhus ovata and Hesperoyucca whipplei (Minnich et al., Reference Minnich, Franco-Vizcaíno, Goforth, Wehncke, Ruben Lara-Lara, Alvarez-Borrego and Ezcurra2014). The abundance and much larger suite of species seen in the macrofossil assemblages from 640 m to 680 m elevation, including the more mesic Pinus quadrifolia in the oldest three middens, suggests chaparral/woodland was much more widespread during the late Pleistocene–early Holocene. This supports the inference by Wells (Reference Wells2000) that today’s disjunct islands of chaparral/woodland species in central Baja are remnants of what was formerly a more continuous distribution. Whether this resulted from expansion from the Sierra San Pedro Mártir (Wells, Reference Wells2000; Minnich et al., Reference Minnich, Franco-Vizcaíno, Goforth, Wehncke, Ruben Lara-Lara, Alvarez-Borrego and Ezcurra2014), long-distance colonization (Minnich et al., Reference Minnich, Franco-Vizcaíno, Goforth, Wehncke, Ruben Lara-Lara, Alvarez-Borrego and Ezcurra2014) and subsequent infilling, or some combination of the two is unknown. What is clear is that many chaparral/woodland species contracted to higher latitudes and/or higher elevation sky islands following the last ice age.
The southern extent of the late glacial–early Holocene chaparral/woodland expansion is poorly constrained. The single midden dating to the Younger Dryas from the Sierra San Francisco (Rhode, Reference Rhode2002) provides evidence for expansion at least that far south, but additional midden series from the Sierra San Borja, Sierra San Francisco, and Sierra Guadalupe (south of Santa Rosalía) could greatly improve the geographic coverage. This general area was proposed by Wells (Reference Wells2000) as a possible isolating barrier for pinyons, with P. quadrifolia restricted today to the Sierra San Pedro Mártir and P. californiarum reaching a southern outpost on Cerro San Luis. These species are replaced by P. lagunae in the southern Cape region. Likewise, J californica and Arctostaphylos peninsularis reach their modern southern limit at the Sierra San Borja, with a southern outpost of chaparral as well as Fouquieria columnaris in the Volcán las Tres Vírgenes above Santa Rosalía (Wells, Reference Wells2000). Future midden surveys perhaps could help elucidate the biogeographic history of this important transition zone into modern monsoonal areas to the south.
There is some debate about the role that increased precipitation played in supporting the expansion of chaparral/woodland to lower elevations and latitudes. As a note of caution about precipitation estimates from midden assemblages, Minnich et al. (Reference Minnich, Franco-Vizcaíno, Goforth, Wehncke, Ruben Lara-Lara, Alvarez-Borrego and Ezcurra2014) argue that granitic or volcanic substrates at midden sites result in a cistern effect that produces anomalous moist sites, leading to overestimates of vegetation and climatic change from middens. Granitic substrates where many middens in Baja have been collected tend to have greater infiltration due to boulder piles, coarse soils, and highly fractured bedrock compared to volcanics with similar precipitation amounts (Shreve, Reference Shreve, Shreve and Wiggins1964). However, there is no evidence for anomalous moisture conditions at Cataviña or other midden sites in northern Baja (Holmgren et al., Reference Holmgren, Betancourt and Rylander2011, Reference Holmgren, Betancourt, Peñalba, Delgadillo, Zuravnsky, Hunter, Rylander and Weiss2014). Substrates at midden sites are neither unusual nor localized. Central and northern Baja consist of extensive areas of granitic (especially the Cretaceous granodiorite of the Peninsular Range) and volcanic rocks, as well as Quaternary alluvium partially derived from these (Gastil et al., Reference Gastil, Phillips and Allison1975). Because middens require caves, rock shelters, or boulders for preservation, they indicate vegetation growing on or at the base of rocky hillslopes that receive more runoff than the more xeric vegetation in the valleys (often extensive plains of the extremely drought tolerant Larrea tridentata and halophytic Atriplex). Nevertheless, modern vegetation at Cataviña consists of the desertscrub found extensively throughout the area, not the smaller bands of chaparral or upland fan palm habitats of more mesic microsites (Minnich et al., Reference Minnich, Franco-Vizcaíno, Goforth, Wehncke, Ruben Lara-Lara, Alvarez-Borrego and Ezcurra2014).
Here, we estimate winter precipitation may have been roughly twice (>250 mm) modern to support the ∼300–500 m lowering of vegetation like J. californica seen in the Cataviña middens, although this lowering may have been moderated by cooler temperatures and reduced evapotranspiration. On the east side of the Gulf of California, early Holocene packrat middens collected at 200 m elevation from the Sierra Bacha in Sonora lack woodland species but contain extralocal species that also indicate greater winter rainfall (Van Devender et al., Reference Van Devender, Burgess, Piper and Turner1994; Anderson and Van Devender, Reference Anderson and Van Devender1995).
During the last glacial, the elevational displacement of species from their modern distributions, as observed in the midden record at a handful of sites, indicates regional-scale changes associated with cooler growing seasons and increases in the frequency of frontal storms during the cool season. Especially notable is the presence of J. californica in 38 glacial-aged middens from the Sierra Juárez, all collected from 300 m to 150 m, thus making it a minimum estimate for downslope displacement of chaparral/woodland (Holmgren et al., Reference Holmgren, Betancourt, Peñalba, Delgadillo, Zuravnsky, Hunter, Rylander and Weiss2014). The decline or disappearance of several extralocal chaparral and woodland species by 11,630 cal yr BP suggests a decline in effective moisture in the early Holocene. Further corroboration of a wet glacial in the Central Desert is the presence of a large pluvial lake (∼66 km2) at Laguna Chapala, 60 km south of Cataviña prior to 10,225 cal yr BP, which then desiccated by 7450 cal yr BP, and is now a dry playa (Davis, Reference Davis2003).
The Cataviña middens record several desert taxa mixed in with chaparral and woodland species during the Pleistocene and persisting throughout the Holocene and the quick arrival of other desert species after ∼11,000 cal yr BP. This resembles what Holmgren et al. (Reference Holmgren, Betancourt, Peñalba, Delgadillo, Zuravnsky, Hunter, Rylander and Weiss2014) found in middens from the Sierra Juárez in northern Baja. As noted in that study, the presence of desert elements within chaparral/woodland during the Pleistocene supports the idea that desert species could have survived in microsites and/or under the protection of nurse plants during unfavorable glacial periods (Valiente-Banuet et al., Reference Valiente-Banuet, Rumebe, Verdú and Callaay2006); the Sonoran Desert is a biome that has repeatedly assembled during interglacial periods such as the Holocene from many disparate populations of desert plants that survived the glacial period in dry and protected microsites (Van Devender, Reference Van Devender, Betancourt, Van Devender and Martin1990; Holmgren et al., Reference Holmgren, Betancourt, Peñalba, Delgadillo, Zuravnsky, Hunter, Rylander and Weiss2014). The present data do not allow us to make any conclusions concerning potential desert distribution downhill, although it remains feasible that broader areas with strictly desert plants existed in central Baja at elevations lower than the chaparral/woodland midden assemblages found at Cataviña (640–680 m) and San Fernando (550–595 m; Wells, Reference Wells2000), especially given the much greater areal extent of lowlands exposed by the drop in sea level during the last glacial (Fig. 1). The quick arrival of several desert species shortly after ∼12,000 cal yr BP at Cataviña also suggests dispersal from nearby microsites or lower elevations harboring desert plants. Pollen assemblages from the Sierra Bacha middens also show modern Sonoran Desert elements were established in coastal Sonora by ∼11,465 cal yr BP, with near-modern desertscrub present by the middle Holocene (Van Devender et al., Reference Van Devender, Burgess, Piper and Turner1994; Anderson and Van Devender, Reference Anderson and Van Devender1995). Vegetation during the remainder of the Holocene appears relatively stable at Cataviña.
A weaker monsoon during the late glacial/early Holocene is implied by the lack of C4 grasses and summer-flowering annuals in middens at Cataviña, the Sierra San Francisco midden to the south (Rhode, Reference Rhode2002), or the Sierra Bacha middens on the Mexican mainland (Van Devender et al., Reference Van Devender, Burgess, Piper and Turner1994). This contrasts with midden studies to the north that contain these species (Van Devender, Reference Van Devender, Betancourt, Van Devender and Martin1990; Holmgren et al., Reference Holmgren, Betancourt and Rylander2010, Reference Holmgren, Betancourt and Rylander2011, Reference Holmgren, Betancourt, Peñalba, Delgadillo, Zuravnsky, Hunter, Rylander and Weiss2014). Likewise, leaf wax δD values from cores in the Gulf of California suggest that, although weaker, the summer monsoon still contributed more than 20% of regional rainfall in the Guaymas Basin (near Santa Rosalía) during the last glacial maximum ∼21,000 yr BP (Bhattacharya et al., Reference Bhattacharya, Tierney, Addison and Murray2018). While still present, this weakening of the monsoon may have fallen below the threshold for C4 grasses and summer-flowering annuals in central Baja, especially when combined with the effects of local topography. Around Cataviña, the lower elevations may not provide as much orographic uplift to aid summer convection as areas to the north and south. In any case, we can infer that the late glacial increase in precipitation in central Baja needed to support chaparral and woodland may have resulted from enhanced winter precipitation. Based on their expert knowledge about the peninsula’s modern vegetation and climate, Minnich et al. (Reference Minnich, Franco-Vizcaíno, Goforth, Wehncke, Ruben Lara-Lara, Alvarez-Borrego and Ezcurra2014) reason instead that during the last glacial period both extension of chaparral/woodland elements to lower latitudes and elevations could be explained simply by lower growing season temperatures and evapotranspiration, perhaps supplemented by greater marine layer penetration onto the peninsula. Better Pleistocene midden coverage, and detailed climate modeling focused on the peninsula, could help resolve these differences in interpretation. An important clue may be the widespread presence of endemic desert plants (e.g., Fouquiera columnaris), and their climatic tolerances, in late Pleistocene chaparral/woodland midden assemblages below 1000 m elevation.
In addition to its high plant endemism and diversity, the Central Desert also is defined by numerous and clear genetic breaks in plant and animal populations. Regions with elevated endemism are presumably generated through increased speciation rates or an unusual capacity for certain areas to sustain refugial populations. Examples of such endemics at Cataviña include Fouquieria columnaris, Pachycormus discolor, Pachycereus pringlei, and Agave cerulata. In the last decade, a multidisciplinary working group of earth and life scientists, the Baja GeoGenomics (BGG) consortium, has documented a strong pattern of intraspecific north–south genetic divergence in more than 60 disparate plant and animal species in the Baja peninsula (Dolby et al., Reference Dolby, Bennett, Lira-Noriega, Wilder and Munguía-Vega2015, Reference Dolby, Bennett, Dorsey, Stokes, Riddle, Lira-Noriega, Munguía-Vega and Wilder2022). BGG researchers are evaluating three hypotheses for these genetic breaks: (1) isolation of populations by a mid-peninsular seaway; (2) isolation in glacial refugia; and (3) temporal isolation imposed by a climate dipole in seasonal precipitation. Previously, several biogeographers hypothesized the presence of a cross-peninsular marine seaway during the Pliocene–Pleistocene to explain the genetic divergence (Hafner and Riddle, Reference Hafner, Riddle, Yates, Gannon and Wilson1997, Reference Hafner, Riddle, Upchurch, McGowan and Slater2011; Upton and Murphy, Reference Upton and Murphy1997; Riddle et al., Reference Riddle, Halfner, Alexander and Jaeger2000; Lindell et al., Reference Lindell, Ngo and Murphy2006; Vázquez-Miranda et al., Reference Vázquez-Miranda, Zink and Pinto2022). However, the BGG conducted a stratigraphic and sedimentological study of Pliocene–Pleistocene beds in the low-lying San Ignacio Trough, a northeast–southwest oriented topographic saddle in central Baja, defined by stacked paleosol sequences, cross-bedding, and terrestrial trace fossils, that strongly refutes the seaway hypothesis (Gardner et al., Reference Gardner, Hasiotis, Dorsey, Darin, Hausback, Bennett, Heizler and Dolby2024). This points to significant niche and genetic divergence in populations between northern and southern Baja, most likely indicating the dynamics of an evolving and complex topography, fluctuating air temperature regimes and air mass boundaries, and shifting refugia during both the Neogene and the Quaternary (Dolby et al., Reference Dolby, Munguia-Verga, Dorsey, Bennett, Drain, Gardner and Araya-Donoso2024; Araya-Donoso et al., Reference Araya-Donoso, Dávalos-Dehullu, Lukasik-Drescher, Moore, Wilder, Lira-Noriega, Munguía-Vega, Kusumi and Dolby2025). Currently, the late Quaternary midden record in Baja California remains too sparse to rigorously test either the refugium or climatic (seasonal precipitation) dipole hypotheses. Additional studies would be needed to understand the late Quaternary stability of the north–south, seasonal precipitation dipole on the peninsula. The Cataviña midden record lacks botanical evidence for a significant northward expansion of monsoonal precipitation during the last glacial period, and more complete midden coverage is needed to determine the extension of winter storm tracks south of 28°S.
Conclusions
The long record of macrofossils and pollen in this midden study provides new insights into the biogeographic history of central Baja over the past >40,000 years. The presence of California chaparral and conifer woodland during the Pleistocene provides evidence for southward and downward expansion compared to today. In contrast to midden series to the north, Pleistocene middens from Cataviña lack of species indicative of warm-season precipitatio, bolstering evidence from the Sierra San Francisco that the ∼100–150 mm increase in precipitation needed to support chaparral/woodland species is attributable to enhanced winter rainfall. The southern extent of the late glacial–early Holocene chaparral/woodland expansion and the persistence of Fouquieria columnaris, Agave cerulata, and other endemic desert taxa throughout the late Quaternary remains poorly constrained and would benefit from additional midden series from central and southern Baja. Although midden preservation may be limited by greater precipitation, a systematic search for middens at higher elevations could reveal a Pleistocene expansion of diverse conifer forest into the middle elevations of northern and central Baja.
Acknowledgments
JLB acknowledges funding from the US Geological Survey. We thank Tony Burgess, John Wiens, Rene Rondeau, Bill Clark, and Steve Bullock for assistance in the field, Lizbeth Rivero and Lizeth Larios for pollen extractions, and Chuck Sternberg for drafting Figure 1. We also thank J. Sankey for contributing midden data and Stephen Jackson and two anonymous reviewers for thoughtful suggestions that improved this article. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.





 Open access
Open access