


Nutrition is increasingly recognised as a key modifiable lifestyle factor influencing both physical and mental health, whereby inadequate nutrient intake is one of the main causes for poor quality of life, and contributes to long-term illnesses(Reference Gheonea, Oancea and Mititelu1). Growing evidence links specific nutrients and dietary patterns to improvements in (or modulation of) cognitive function, mood, sleep quality and metabolic regulation in both healthy and clinical populations(Reference Gheonea, Oancea and Mititelu1–Reference Zhong, Han and Li4). These effects occur through multiple interacting biological pathways, including, but not limited to, the modulation of inflammation, improved mitochondrial function, reduction of oxidative stress and regulation of gut microbiota and the gut–brain axis(Reference Li, Pan and Shen5–Reference Del Portillo, Clemente-Suárez and Ruisoto7).
Of specific importance, asparagus (Asparagus officinalis), a nutrient-dense vegetable containing a variety of bioactive compounds, including polyphenols (such as flavonoids), steroidal saponins, essential oils and dietary fibre, has gained attention(Reference Guo, Wang and Liu8,Reference Nadeem, Khan and Ahmad9) . These compounds are known to exhibit antioxidant activity by reduction of reactive oxygen species and anti-inflammatory effects through inhibition of nuclear factor kappa B – a key regulator of inflammation that drives the release of molecules such as IL-6 and tumour necrosis factor-α, and by reducing pro-inflammatory cytokines(Reference Yahfoufi, Alsadi and Jambi10). Such actions may affect various health outcomes, including cognitive function, depression, anxiety and sleep disorders, in both animal models and humans(Reference Naomi, Yazid and Teoh11,Reference Pereira, da Silva and Hermsdorff12) . Asparagus also provides key micronutrients, including potassium, calcium, magnesium and selenium, which support neurological function and mental well-being(Reference Guo, Wang and Liu8). In addition, better overall well-being, including improvements in sleep, mood and reduced inflammation, is supported by diets rich in these compounds and minerals, with effects thought to be mediated in part through pathways involving the gut microbiota(Reference Sejbuk, Siebieszuk and Witkowska13,Reference Zheng, Qin and Wen14) .
Furthermore, asparagus constituents enhance levels of Heat Shock Protein 70 (HSP70), a protective molecule that helps cells cope with stress by preventing protein damage and reducing the harmful effects of inflammation and oxidative stress(Reference Inoue, Takanari and Abe15). In vitro studies showed asparagus extract up-regulated HSP70 messenger ribonucleic acid (mRNA) expression in human HL-60 cells (a human promyelocytic leukaemia cell line)(Reference Inoue, Takanari and Abe15); additionally, HSP70 injections in sleep-deprived mice led to alleviated sleep-deprivation-induced anxiety and cognitive impairment, restored brain-derived neurotropic factor levels and reduced microglia-induced neuroinflammation in the hippocampus(Reference Kang, Lin and Su16).
Despite these promising mechanistic insights, evidence from human studies remains limited. Only a small number of clinical trials have investigated the effects of asparagus or asparagus-based products on cognitive function, sleep, mood and relevant biomarkers, showing reduction in mental fatigue(Reference Takanari, Nakahigashi and Sato17–Reference Sakai, Nagata and Nagata19), depressive symptoms(Reference Takanari, Nakahigashi and Sato17,Reference Zenimoto and Takahashi20,Reference Mikhailichenko, Lai and Wei21) and sleep disturbances(Reference Huang, Zhou and An22). It is essential to highlight that while initial human studies provide insight into the potential effects of asparagus, considerable heterogeneity exists across studies in terms of participants, interventions and outcome measures, and no systematic review has yet synthesised these findings. Conducting a systematic review, therefore, allows for a structured evaluation of the available evidence, identification of consistent trends and potential mechanisms, and clarification of knowledge gaps, providing a foundation for future research and guiding the design of more rigorous trials. Hence, this review aims to evaluate and synthesise existing evidence from randomised controlled and quasi-experimental trials examining the impact of orally consumed asparagus or asparagus-based products on various health outcomes, including cognition, mood, sleep and relevant biomarkers in human adults.
Methods
The protocol for this systematic review was registered in the International Prospective Register of Systematic Reviews (PROSPERO; Registration ID: CRD42025636834), and the articles were selected following the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines(Reference Page, McKenzie and Bossuyt23).
Data sources and search strategy
Five electronic databases (PubMed/MEDLINE, Web of Science Core Collection, PsycINFO, Scopus and Google Scholar) were searched up until the end of April 2025.
The searches were conducted with the following strings, adjusted to be compatible which each search engine based on title and abstract: (((asparagus*) OR (enzyme treated asparagus stem)) OR (ETAS®)) AND ((cogn*) OR (memory) OR (attention*) OR (Executive Function) OR (Mood) OR (Anxi*) OR (Depressi*) OR (Sleep*) OR (Stress) OR (Inflammatory) OR (Brain) OR (Blood) OR (IFN-γ) OR (IL-1β) OR (IL-2) OR (IL-4) OR (IL-6) OR (IL-8) OR (IL-10) OR (IL-12p70) OR (IL-13) OR (TNF-α) OR (CRP) OR (Tau) OR (Metabolites)). The searches were restricted to the Title/Abstract fields.
Scientific publication from the Amino Up Ltd webpage has also been searched.
Inclusion and exclusion criteria
The inclusion criteria were as follows:
-
• Treatment: Dietary intervention of asparagus or an asparagus-based product (no dose or type limit)
-
• Outcome measures: cognitive, affective (defined following Gross et al. (Reference Gross, Uusberg and Uusberg24)) as encompassing emotions, stress responses, impulses (e.g. appetite, and mood), sleep outcomes or relevant biomarkers
-
• Participants: No restriction to age, sex or health status
-
• Design: Randomised controlled or quasi-experiment trials (including pre-post-designs with non-equivalent control groups(Reference Miller, Smith and Pugatch25)
-
• Language: Any
Exclusion criteria were as follows:
-
• Design: Case report, letter to editor, conference paper, thesis, personal opinion or commentary
-
• Animal studies, in vitro and ex vivo studies
Study selection and data extraction
All search results were organised using the EndNote program (Clarivate, Philadelphia, PA, USA). One reviewer (AD) independently assessed the selected records for eligibility, with any disagreements regarding study inclusion resolved through discussion with a second (PH) and third (DL) reviewer. Data extraction was conducted using a standardised form, capturing details including:
-
– Publication details: authors, year, journal
-
– Participant characteristics: number of participants recruited, number of participants included in the study, number of participants (intervention), number of participants (control), number of participants (other intervention), health status, gender and age range
-
– Study design: design and blinding
-
– Intervention characteristics: intervention duration, washout period, asparagus supplement type, asparagus supplement dose
-
– Control characteristics: presence/absence of control/placebo, control/placebo doses
-
– Outcome measures: cognitive, affective, sleep outcomes or related markers
-
– Remarks: notes on the factors that might affect results/data quality
The study quality and potential risk of bias of the selected studies were evaluated using the Cochrane Risk of Bias tool(Reference Higgins and Green26).
Results
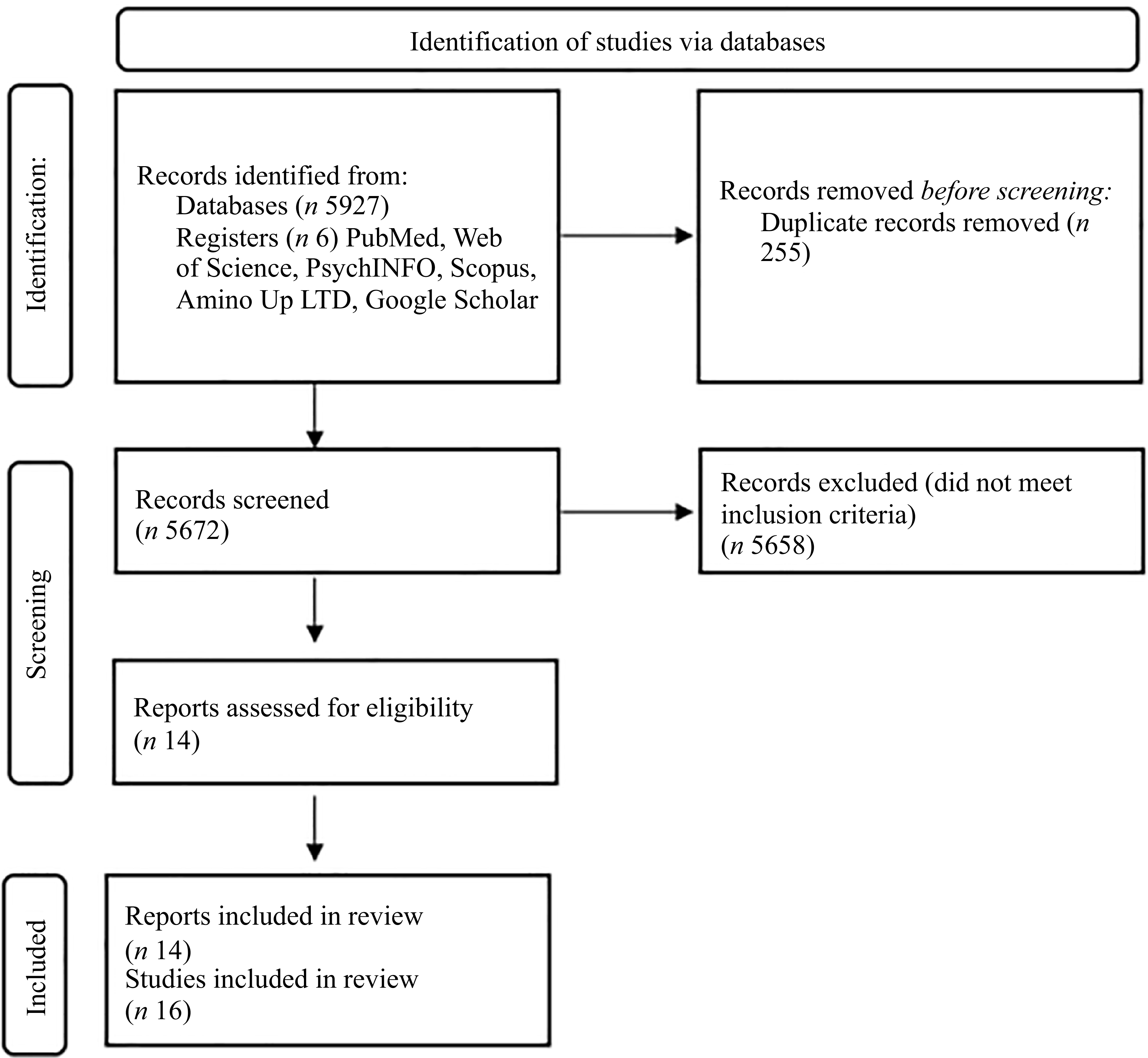
The literature search identified 5927 publications. Following the removal of duplicates and papers that failed to meet the inclusion and exclusion criteria, fourteen papers were included in this review. However, Ito et al. (Reference Ito, Goto and Takanari27) and Cheng et al. (Reference Cheng, Pan and Wang28) reported on two studies within the same paper. Therefore, sixteen studies were included in the systematic review (Figure 1). Thirteen were randomised control trials, and three were pre/post-designs(Reference Sakai, Nagata and Nagata19,Reference Huang, Zhou and An22,Reference Cheng, Pan and Wang28) . There were n 503 participants in total, with mean age ranging from 19·5(Reference Cheng, Pan and Wang28) to 87·4(Reference Zenimoto and Takahashi20). The most frequently measured outcome was affective (n 12 studies), followed by sleep (n 10), and cognition (n 7).
PRISMA flow diagram.

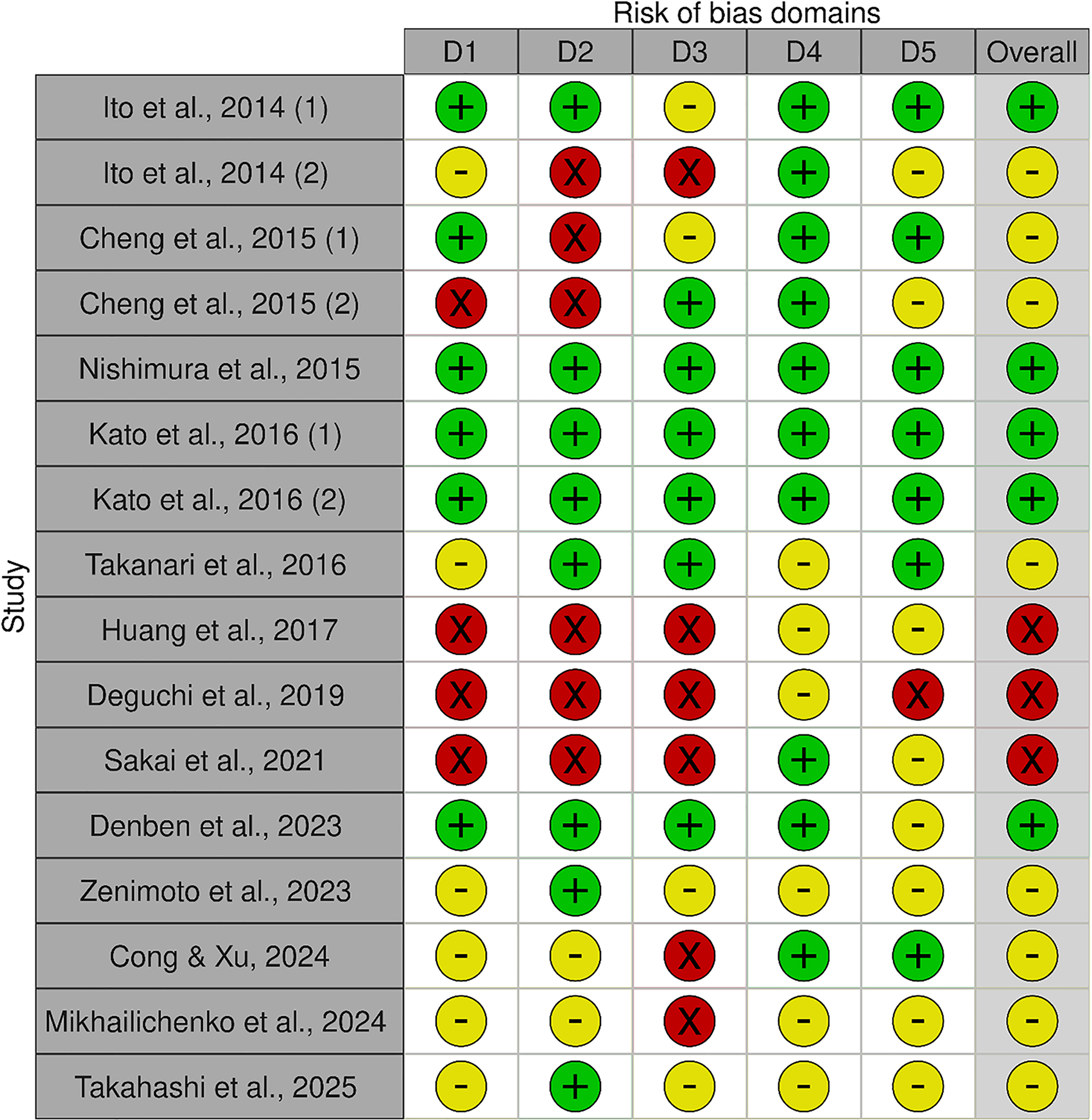
One reviewer (AD) evaluated the quality of the studies included in this review (Figure 2). Most studies demonstrated a low or unclear risk of bias across domains, with only a minimal proportion classified as high risk. Specifically, most studies were judged to have a low-to-moderate risk of selection bias, attrition bias and reporting bias. However, there was greater uncertainty regarding performance and detection biases. Generally, quasi-experiments showed higher bias(Reference Sakai, Nagata and Nagata19,Reference Huang, Zhou and An22,Reference Cheng, Pan and Wang28) .
Cochrane risk of bias.

Summaries of all the studies are presented in Table 1 (Methods) and Table 2 (Outcomes). Green indicates low risk of bias, yellow indicates some concerns, and red indicates high risk of bias. D1 = selection bias; D2 = performance bias; D3 = detection bias; D4 = attrition bias; D5 = reporting bias.
Characteristics of included studies

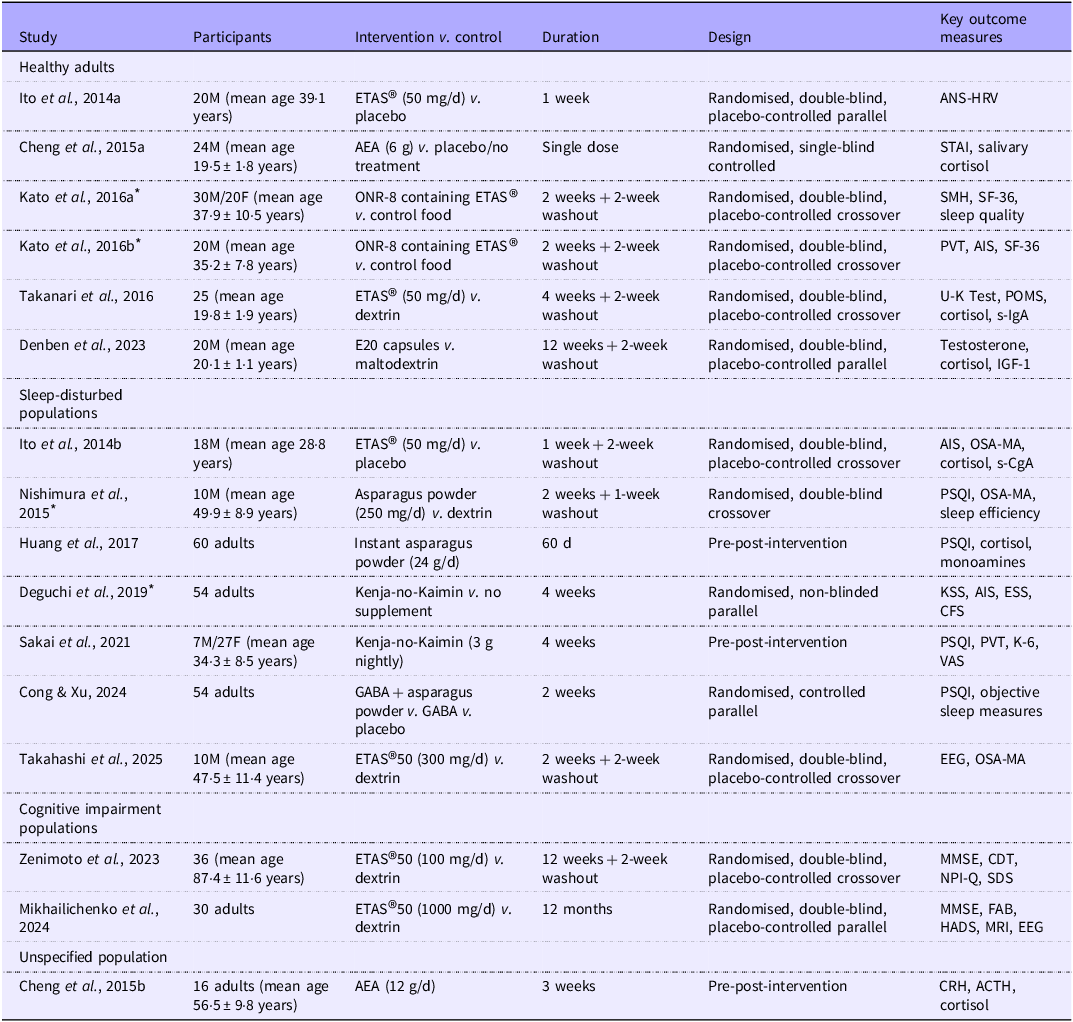
Table 1. Long description
The table presents characteristics of included studies, organized into categories: Healthy adults, Sleep-disturbed populations, Cognitive impairment populations, and Unspecified population. It has six columns: Study, Participants, Intervention v. control, Duration, Design, and Key outcome measures. The table includes 18 rows with detailed information for each study. Each row lists the study reference, participant details, intervention and control conditions, study duration, study design, and key outcome measures. The studies vary in participant numbers, ages, interventions, durations, designs, and outcome measures.
* Study translated from the original language into English for data extraction and review purposes. Abbreviations: ANS-HRV, ANS, Autonomic Nervous System-Heartrate Variability; ETAS®, enzyme-treated asparagus stem; AEA, aqueous extract of asparagus; M, male; F, female; AIS, Athens Insomnia Scale; OSA-MA, Oguri-Shirakawa-Azumi Sleep Inventory; PVT, Psychomotor Vigilance Test; PSQI, Pittsburgh Sleep Quality Index; POMS, Profile of Mood States; KSS, Karolinska Sleepiness Scale; ESS, Epworth Sleepiness Scale; CFS, Chalder Fatigue Scale; MMSE, Mini-Mental State Examination; FAB, Frontal Assessment Battery; HADS, Hospital Anxiety and Depression Scale; STAI, State-Trait Anxiety Inventory; SMH, St. Mary’s Hospital; SF-36, 36-Item Short Form Health Survey; VAS, Visual Analogue Scale; EEG, Electroencephalography; CDT, Clock Drawing Test; NPI-Q, Neuropsychiatric Inventory Questionnaire; CRH, corticotropin-releasing hormone; ACTH, adrenocorticotropic hormone; GABA, Gamma-aminobutyric acid; IGF, Insulin-like growth factor; DBP, Diastolic Blood Pressure; JUMACL, Japanese UWIST Mood Adjective Checklist.
Summary of outcomes of included studies

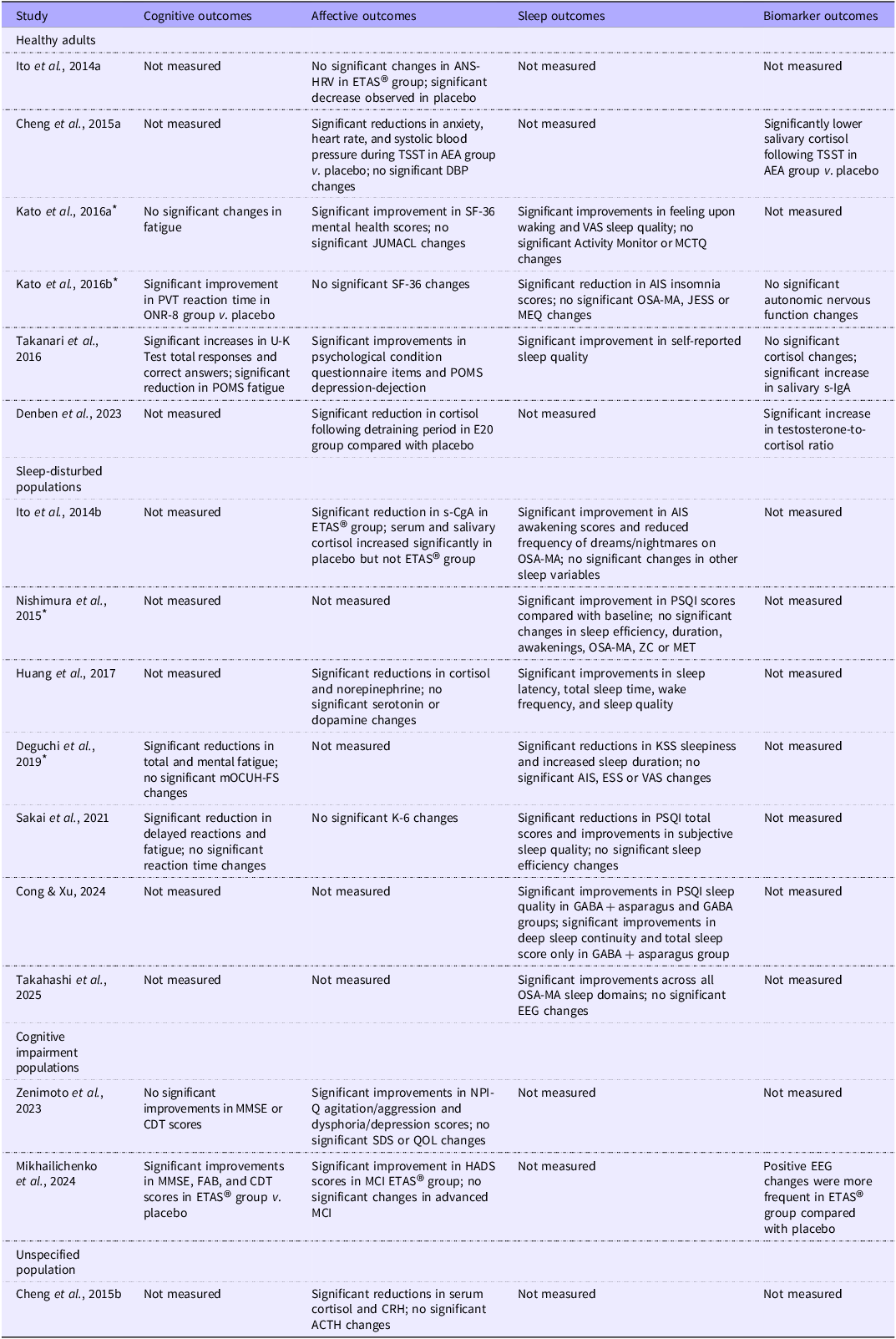
Table 2. Long description
The table presents a summary of outcomes from various studies, focusing on cognitive, affective, sleep, and biomarker outcomes across different populations. It includes columns for study names, cognitive outcomes, affective outcomes, sleep outcomes, and biomarker outcomes. Each row corresponds to a specific study and its findings. The table has 18 rows and 5 columns. Row 1: Ito et al., 2014a, Not measured, No significant changes in ANS-HRV in ETAS group; significant decrease observed in placebo, Not measured, Not measured. Row 2: Cheng et al., 2015a, Not measured, Significant reductions in anxiety, heart rate, and systolic blood pressure during TSST in AEA group v. placebo; no significant DBP changes, Not measured, Significantly lower salivary cortisol following TSST in AEA group v. placebo. Row 3: Kato et al., 2016a, No significant changes in fatigue, Significant improvement in SF-36 mental health scores; no significant JUMACL changes, Significant improvements in feeling upon waking and VAS sleep quality; no significant Activity Monitor or MCTQ changes, Not measured. Row 4: Kato et al., 2016b, Significant improvement in PVT reaction time in ONR-8 group v. placebo, No significant SF-36 changes, Significant reduction in AIS insomnia scores; no significant OSA-MA, JESS or MEQ changes, No significant autonomic nervous function changes. Row 5: Takanari et al., 2016, Significant increases in U-K Text total responses and correct answers; significant reduction in POMS fatigue, Significant improvements in psychological condition questionnaire items and POMS depression rejection, Significant improvement in self-reported sleep quality, No significant cortisol changes; significant increase in salivary s-IgA. Row 6: Denben et al., 2023, Not measured, Significant reduction in cortisol following detraining period in E20 group compared with placebo, Not measured, Significant increase in testosterone-to-cortisol ratio. Row 7: Ito et al., 2014b, Not measured, Significant reduction in s-CgA in ETAS group; serum and salivary cortisol increased significantly in placebo but not ETAS group, Significant improvement in AIS awakening scores and reduced frequency of dreams/nightmares on OSA-MA; no significant changes in other sleep variables, Not measured. Row 8: Nishimura et al., 2015, Not measured, Not measured, Significant improvement in PSQI scores compared with baseline; no significant changes in sleep efficiency, duration, awakenings, OSA-MA, ZC or MET, Not measured. Row 9: Huang et al., 2017, Not measured, Significant reductions in cortisol and norepinephrine; no significant serotonin or dopamine changes, Significant improvements in sleep latency, total sleep time, wake frequency, and sleep quality, Not measured. Row 10: Deguchi et al., 2019, Significant reductions in total and mental fatigue; no significant mOCUH-FS changes, Not measured, Significant reductions in KSS sleepiness and increased sleep duration; no significant AIS, ESS or VAS changes, Not measured. Row 11: Sakai et al., 2021, Significant reduction in delayed reactions and fatigue; no significant reaction time changes, No significant K-6 changes, Significant reductions in PSQI total scores and improvements in subjective sleep quality; no significant sleep efficiency changes, Not measured. Row 12: Cong & Xu, 2024, Not measured, Not measured, Significant improvements in PSQI sleep quality in GABA + asparagus and GABA groups; significant improvements in deep sleep continuity and total sleep score only in GABA + asparagus group, Not measured. Row 13: Takahashi et al., 2025, Not measured, Not measured, Significant improvements across all OSA-MA sleep domains; no significant EEG changes, Not measured. Row 14: Zenimoto et al., 2023, No significant improvements in MMSE or CDT scores, Significant improvements in NPI-Q agitation/aggression and dysphoria/depression scores; no significant SDS or QOL changes, Not measured, Not measured. Row 15: Mikhailichenko et al., 2024, Significant improvements in MMSE, FAB, and CDT scores in ETAS group v. placebo, Significant improvement in HADS scores in MCI ETAS group; no significant changes in advanced MCI, Not measured, Positive EEG changes were more frequent in ETAS group compared with placebo. Row 16: Cheng et al., 2015b, Not measured, Significant reductions in serum cortisol and CRH; no significant ACTH changes, Not measured, Not measured.
* Study translated from the original language into English for data extraction and review purposes. Abbreviations: ANS-HRV, ANS, Autonomic Nervous System-Heartrate Variability; ETAS®, enzyme-treated asparagus stem; AEA, aqueous extract of asparagus; TSST, Trier Social Stress Test; PVT, Psychomotor Vigilance Test; AIS, Athens Insomnia Scale; OSA-MA, Oguri-Shirakawa-Azumi Sleep Inventory; JESS, Japanese Epworth Sleepiness Scale; MEQ, Morningness-Eveningness Questionnaire; U-K Test, Uchida-Kraepelin Test; POMS, Profile of Mood States; s-IgA, salivary immunoglobulin A; s-CgA, salivary chromogranin A; PSQI, Pittsburgh Sleep Quality Index; KSS, Karolinska Sleepiness Scale; ESS, Epworth Sleepiness Scale; mOCUH-FS, City University Hospital Fatigue Scale; VAS, Visual Analogue Scale; EEG, electroencephalography; MMSE, Mini-Mental State Examination; FAB, Frontal Assessment Battery; CDT, Clock Drawing Test; HADS, Hospital Anxiety and Depression Scale; MCI, mild cognitive impairment; CRH, corticotropin-releasing hormone; ACTH, adrenocorticotropic hormone; SF-36, 36-Item Short Form Health Survey; MCTQ, Munich ChronoType Questionnaire; NPI-Q, Neuropsychiatric Inventory Questionnaire; SDS, Self-Rating Depression Scale; QOL, Quality of Life; ZC, Zeitgeber Cycle; MET, Metabolic Equivalent of Task.
Cognitive outcomes
Seven studies assessed the effects of ETAS® or related formulations (e.g. Kenja-no-Kaimin) on cognitive outcomes. Participant health status varied, including healthy adults(Reference Takanari, Nakahigashi and Sato17,Reference Kato, Shoichiro and Sato29,Reference Kato, Inoue and Sato30) , individuals with sleep problems(Reference Deguchi, Mizuno and Tanaka18,Reference Sakai, Nagata and Nagata19) and those with cognitive impairment ranging from mild cognitive impairment (MCI) to dementia(Reference Zenimoto and Takahashi20,Reference Mikhailichenko, Lai and Wei21) . Intervention durations ranged from 2 weeks to 12 months, with doses ranging from 50 mg to 1000 mg/d.
In healthy participants, ONR-8, a food containing asparagus extract, significantly improved reaction times in a Psychomotor Vigilance Test compared with controls after 2 weeks(Reference Kato, Inoue and Sato30). Additionally, Takanari et al. (Reference Takanari, Nakahigashi and Sato17) reported that 4 weeks of ETAS® supplementation (50 mg ETAS® + 200 mg dextrin/d) improved accuracy on the Uchida-Kraepelin test, a serial addition task measuring attention and cognitive flexibility compared with baseline and placebo groups (dextrin, 250 mg/d).
Several studies reported reductions in mental fatigue. Takanari et al. (Reference Takanari, Nakahigashi and Sato17) observed significantly lower Profile of Mood States fatigue scores in the ETAS® group compared with placebo, whereas Kato et al. (Reference Kato, Shoichiro and Sato29) did not detect a significant change in fatigue after 2 weeks of ONR-8. Among participants with sleep problems, in a pre-post-design study, 4 weeks of Kenja-no-Kaimin (3 g/d before sleep) was found to significantly reduce mental fatigue scores compared with baseline(Reference Sakai, Nagata and Nagata19). Deguchi et al. (Reference Deguchi, Mizuno and Tanaka18) similarly found that 4 weeks of Kenja-no-Kaimin (1 sachet/d) reduced total subjective fatigue and mental fatigue scores on the Chalder Fatigue Scale compared with a control group.
In participants with cognitive impairment, Mikhailichenko et al. (Reference Mikhailichenko, Lai and Wei21) reported that 12 months of ETAS® supplementation (1000 mg ETAS®50/d) significantly improved Mini-Mental State Examination scores, assessing general cognitive function including orientation, attention, language, memory and visuospatial skills, as well as Frontal Assessment Battery scores, assessing frontal lobe functions such as abstract thinking, verbal fluency, motor control and inhibitory regulation, compared with placebo in individuals with mild and advanced MCI. Additionally, Zenimoto et al. (Reference Zenimoto and Takahashi20) found that 12 weeks of ETAS® (100 mg ETAS®50/d) supplementation significantly improved Neuropsychiatric Inventory–Questionnaire scores (assessing the dementia symptom severity) compared with placebo (dextrin, dose not specified). Improvements in Mini-Mental State Examination and Clock Drawing Test scores (useful for measuring moderate to severe cognitive impairment and measuring visuospatial ability, executive function and semantic processing)(Reference De Paula, De Miranda and De Moraes31) were also observed within the ETAS® group. Still, they were not statistically significant compared with the placebo.
Affective outcomes
Twelve studies examined affective outcomes (as categorised by Gross et al. (Reference Gross, Uusberg and Uusberg24)), encompassing emotions, stress responses and mood, in healthy adults(Reference Takanari, Nakahigashi and Sato17,Reference Ito, Goto and Takanari27–Reference Kato, Inoue and Sato30,Reference Denben, Sripinyowanich and Ruangthai32) , individuals with sleep problems(Reference Sakai, Nagata and Nagata19,Reference Huang, Zhou and An22,Reference Ito, Goto and Takanari27) , participants with cognitive decline(Reference Zenimoto and Takahashi20,Reference Mikhailichenko, Lai and Wei21) and unspecified health status(Reference Cheng, Pan and Wang28). Interventions included ETAS® (50–1000 mg/d), asparagus powder (24 g/d), E20 (32·2 mg/d), Aqueous Extract of Asparagus (AEA, 6 g/d) and Kenja-no-Kaimin (1 sachet/d), with durations ranging from 1 day to 12 months.
In healthy participants, Takanari et al. (Reference Takanari, Nakahigashi and Sato17) reported that 4 weeks of ETAS® (50 mg + 200 mg dextrin/d) significantly reduced Profile of Mood States depression scores compared with placebo. However, no significant changes were observed in serum or salivary cortisol. Kato et al. (Reference Kato, Shoichiro and Sato29) observed improvements in the mental health subscale of 36-Item Short Form Health Survey following 2 weeks of ONR-8 (3 g/d) supplementation, whereas Kato et al. (Reference Kato, Inoue and Sato30) found no significant changes after 2 weeks of ONR-8 (2 capsules/d). Cheng et al. (Reference Cheng, Pan and Wang28) reported a significant reduction in trait anxiety following a single dose (6 g) of AEA in a pre-post-test design study.
Among participants with sleep problems, in a pre-post-test design study, Sakai et al. (Reference Sakai, Nagata and Nagata19) found no significant changes in the K-6 psychological distress scale after 4 weeks of Kenja-no-Kaimin supplementation. Ito et al. (Reference Ito, Goto and Takanari27) reported that 1 week of ETAS® (50 mg + 200 mg pinedex/d) intake had no significant effect on serum or salivary cortisol measured between 18.00–20.00, whereas the placebo group showed a significant increase, suggesting a potential protective effect of ETAS® on evening cortisol levels. ETAS® supplementation also significantly reduced salivary chromogranin A (s-CgA), particularly in participants with above-average sleep duration.
Among participants with cognitive decline, Mikhailichenko et al. (Reference Mikhailichenko, Lai and Wei21) found that 12 months of ETAS® (1000 mg/d) significantly reduced Hospital Anxiety and Depression Scale depression and anxiety scores in the MCI group, while no changes were observed in the advanced MCI group. Zenimoto et al. (Reference Zenimoto and Takahashi20) reported improvements in Neuropsychiatric Inventory–Questionnaire agitation/aggression and dysphoria/depression scores following 12 weeks of ETAS® (100 mg/d).
Sleep outcomes
Ten studies examined the effects of ETAS®, Kenja-no-Kaimin, ONR-8 or asparagus powder on sleep-related outcomes. Populations included healthy adults(Reference Takanari, Nakahigashi and Sato17,Reference Ito, Goto and Takanari27,Reference Kato, Shoichiro and Sato29,Reference Kato, Inoue and Sato30) and individuals with sleep problems(Reference Deguchi, Mizuno and Tanaka18,Reference Sakai, Nagata and Nagata19,Reference Huang, Zhou and An22,Reference Nishimura, Takahashi and Kohzai33–Reference Takahashi, Seol and Kokubo35) . Intervention durations ranged from 1 week to 60 d, with doses spanning from 50 mg to 24 g/d.
Ito et al. (Reference Ito, Goto and Takanari27) found that 1 week of ETAS® supplementation (50 mg ETAS® + 200 mg pinedex/d) significantly improved the Athens Insomnia Scale item ‘awakening earlier than desired’ and significantly reduced sleep time when sleep efficiency was over 90 % indicating participants woke closer to their desired time. They also reported significantly reduced frequency of nightmares and dreams based on the Oguri-Shirakawa-Azumi Sleep Inventory (OSA-MA). Similarly, Kato et al. (Reference Kato, Inoue and Sato30) reported significantly lower overall Athens Insomnia Scale scores following 1 week of ONR consumption compared with placebo. Takanari et al. (Reference Takanari, Nakahigashi and Sato17) observed improved self-reported sleep quality after 4 weeks of ETAS® (50 mg ETAS® + 200 mg dextrin/d) supplementation. Compared with controls, Kato et al. (Reference Kato, Shoichiro and Sato29) found that 2 weeks of ONR-8 consumption improved ‘feeling asleep after waking’ scores on the St Mary’s Hospital Sleep Questionnaire and increased Visual Analogue Scale-rated sleep quality. Deguchi et al. (Reference Deguchi, Mizuno and Tanaka18) also demonstrated that 4 weeks of Kenja-no-Kaimin (1 sachet/d) reduced Karolinska Sleepiness Scale-measured sleepiness and increased sleep duration compared with controls. Cong & Xu(Reference Cong and Xu34) reported significant improvements in Pittsburgh Sleep Quality Index (PSQI) sleep quality scores following 4 weeks of GABA and asparagus extract supplementation, with more minor improvements in the GABA-only group and no significant changes in the placebo group. Consistently, objective sleep outcomes measured by the Huawei Sport Band 7 showed that the combined intervention significantly improved deep sleep continuity and total sleep scores, whereas changes in the GABA-only and placebo groups were not statistically significant.
Nishimura et al. (Reference Nishimura, Takahashi and Kohzai33) reported that 2 weeks of asparagus capsule supplementation (250 mg/d) reduced sleep disturbances compared with placebo (dextrin, 235 mg/d). Preliminary findings from the uncontrolled pre-post-study by Huang et al. (Reference Huang, Zhou and An22) suggested that 60 d of asparagus powder (24 g/d) decreased sleep onset latency and increased total sleep time and quality compared with baseline. Takahashi et al. (Reference Takahashi, Seol and Kokubo35) further showed that 2 weeks of ETAS® (300 mg/d) improved scores across all five OSA-MA factors including sleepiness on rising, initiation and maintenance of sleep, frequent dreaming/nightmares, restorative sleep and daytime alertness – relative to baseline, with no comparable improvements observed in the placebo group (dextrin, 600 mg/d). Finally, Sakai et al. (Reference Sakai, Nagata and Nagata19) reported that 4 weeks of Kenja-no-Kaimin (3 g/d) significantly reduced total PSQI scores and improved Visual Analogue Scale scores compared with baseline; however, results from the accelerometer-based activity metre were not consistent with subjective findings, and the study lacked a control group.
Discussion
The current review examined the effects of asparagus-derived supplementation consumption on cognition, mood, sleep and related physiological markers in adults, drawing on evidence from randomised controlled and quasi-experimental studies. Across populations, asparagus supplementation appears to influence multiple interrelated domains, including cognitive performance, stress regulation, sleep quality and inflammation. However, the magnitude and consistency of effects vary by population, dose and intervention duration.
Cognition
Regarding cognitive outcomes, in young healthy adults, short-term ETAS® supplementation improved reaction time(Reference Kato, Inoue and Sato30) and accuracy of attention(Reference Takanari, Nakahigashi and Sato17), indicating potential enhancements in attentional processes. Fatigue results were mixed such that while Kato et al. (Reference Kato, Inoue and Sato30) showed no significant improvements, Sakai et al. (Reference Sakai, Nagata and Nagata19) identified significant improvements in fatigue utilising Visual Analogue Scale. However, the absence of a control group limits the inference that the changes are due to the intervention(Reference Paulus, Dahabreh and Balk36). Further evidence supporting improvements in fatigue comes from Takanari et al. (Reference Takanari, Nakahigashi and Sato17) who reported improvements on the Profile of Mood States fatigue subscale and Deguchi et al. (Reference Deguchi, Mizuno and Tanaka18) who observed improvements on the Chalder Fatigue Scale. As two of the four included studies recruited sleep-disturbed individuals(Reference Deguchi, Mizuno and Tanaka18,Reference Sakai, Nagata and Nagata19) and 3 out of 4 studies utilised intervention durations of 4 weeks, improvements in fatigue may have been more readily detectable in sleep-impaired populations or with longer intervention durations. In contrast, recruitment of healthy participants in for a 2-week long intervention in Kato et al. (Reference Kato, Shoichiro and Sato29) may have afforded little or no room for improvement, thereby limiting the likelihood of observing a measurable change.
In older adults with cognitive impairments, improvements were strongly dose- and duration-dependent. High-dose ETAS® (1000 mg/d) over 12 months improved global and executive functioning (Mini-Mental State Examination, Frontal Assessment Battery, Clock Drawing Test)(Reference Mikhailichenko, Lai and Wei21), whereas a shorter 12-week, lower-dose intervention (100 mg/d) produced only subtle within-group changes(Reference Zenimoto and Takahashi20). Cellular findings mirrored this pattern; a 2-week ETAS® 50 (300 mg/d) intervention in sleep-disturbed adults did not elicit significant changes in HSP70(Reference Takahashi, Seol and Kokubo35), whereas long-term, high-dose ETAS® robustly increased HSP70 in adults with MCI and advanced MCI(Reference Mikhailichenko, Lai and Wei21). Dose-dependent HSP70 induction has also been demonstrated at 100–150 mg/d of ETAS®(Reference Ito, Maeda and Goto37), while 50 mg/d over 1 week was insufficient to produce measurable effects(Reference Ito, Goto and Takanari27). These findings support the notion that long-term, high-dose ETAS® administration is required to achieve both cognitive and cellular effects.
Age-related increases in inflammation and oxidative stress may also influence responsiveness to interventions(Reference Marzola, Melzer and Pavesi38). Despite the older sample (mean age 87·4 years) in Zenimoto et al. (Reference Zenimoto and Takahashi20) having presumably higher stress and inflammatory burden, they showed greater cognitive improvements than the younger cohort in Mikhailichenko et al. (Reference Mikhailichenko, Lai and Wei21) (mean age 66 years), suggesting that higher doses and longer intervention durations can mitigate age-related constraints on cognitive responsiveness. Importantly, the capacity to induce HSP70 in response to stress declines with age(Reference Heydari, Wu and Takahashi39), but ETAS® may help restore HSP70 inducibility in adults ≥ 80 years(Reference Mikhailichenko, Lai and Wei21), potentially supporting cognitive resilience in the face of age-related physiological stress. Effects may be less pronounced in younger older adults (60–70 years) with lower stress burden. Together, these findings highlight the potential importance of early intervention and appropriate dosing to maximise cognitive outcomes in older adults.
Affect and relevant biomarkers
Regarding affective outcomes, anxiety was shown to reduce following a single dose of AEA(Reference Cheng, Pan and Wang28), while Takanari et al. (Reference Takanari, Nakahigashi and Sato17) observed reductions in depression-dejection following 4 weeks of supplementation, supporting short-term mood benefits. Similarly, Kato et al. (Reference Kato, Shoichiro and Sato29) found improved mental health scores. However, it is important to highlight that a similar study involving only males did not observe these effects(Reference Kato, Inoue and Sato30), suggesting that there may be sex differences in affective outcomes. More research is required to elucidate these outcomes. It is important to highlight that, although Kato et al. (Reference Kato, Shoichiro and Sato29) observed improved mental health with the 36-Item Short Form Health Survey there were no corresponding improvements in the UWIST Mood Checklist (Japanese version) checklist highlighting the need for consistent measurement tools to compare mental health outcomes within and between studies accurately(Reference Breedvelt, Zamperoni and South40). In unhealthy cohorts, similarly, Mikhailichenko et al. (Reference Mikhailichenko, Lai and Wei21) reported reductions in anxiety and depression in participants with MCI; however, this was not seen in advanced MCI. Furthermore, improvements were found in measures reflecting behavioural disturbances associated with dementia, e.g. hallucinations and agitation(Reference Zenimoto and Takahashi20). This suggests that the severity of cognitive impairment may limit benefits, with some symptoms more responsive than others. For example, behavioural and neuropsychiatric symptoms (e.g. agitation) are prominent across dementia severity but remain responsive to targeted non-pharmacological interventions even after a dementia diagnosis(Reference Wiels, Wittens and Zeeuws41). In contrast, affective symptoms such as depression are more commonly observed in earlier stages of cognitive impairment, where interventions, including natural supplements, can lead to improvements in cognitive performance and depressive symptoms in MCI and early stages, with less consistent effects as dementia severity increases(Reference Businaro, Vauzour and Sarris42).
Asparagus supplementation also appears to modulate stress-related biomarkers via multiple physiological pathways, with variability attributable to methodological differences across studies. Several studies support improvements in cortisol levels. In a pre-post-test study without a control group, Cheng et al. (Reference Cheng, Pan and Wang28) found that AEA dampened the salivary cortisol response and decreased serum corticotropin-releasing hormone and serum cortisol; however, there were no significant differences in adrenocorticotropic hormone levels(Reference Cheng, Pan and Wang28). As the stress response is mediated by the hypothalamic–pituitary–adrenal axis, starting with hypothalamic corticotropin-releasing hormone release, followed by pituitary adrenocorticotropic hormone secretion, and culminating in cortisol production(Reference Lee, Meyer and Nenke43), these results indicate that supplementation of AEA may modulate the stress response specifically at the corticotropin-releasing hormone and cortisol stages. However, the reduction of cortisol in the absence of changes in adrenocorticotropic hormone also suggests possible peripheral regulation at the level of the adrenal cortex, such as autonomic innervation of the adrenal cortex or adrenal medullary–derived signals influencing cortisol secretion, rather than exclusive hypothalamic corticotropin-releasing hormone modulation(Reference Tsigos and Chrousos44).
Furthermore, in individuals with sleep disturbances, in a pre-post-test design study, Huang et al. (Reference Huang, Zhou and An22) reported significant reductions in both blood cortisol and norepinephrine. However, they did not specify whether serum, plasma or whole blood was considered. While Ito et al. (Reference Ito, Goto and Takanari27) did not specifically find reduced cortisol in their experimental group, their control group showed a significant increase in salivary and serum cortisol, indicating that ETAS® supplementation may have a stabilising effect on hypothalamic–pituitary–adrenal axis activity. They also found that the ETAS® group had lower s-CgA levels, suggesting reduced sympathetic arousal(Reference Gallina, Di Mauro and D’Amico45). Notably, salivary cortisol levels are generally lower than serum cortisol(Reference Ljubijankić, Popović-Javorić and Šćeta46), highlighting that salivary cortisol measurements may be less sensitive or reliable as biomarkers of asparagus-related stress modulation. Decreased serum cortisol and elevated testosterone-to-cortisol ratios (associated with greater anabolic potential and lower stress burden) were observed following 12 weeks of E20 supplementation(Reference Denben, Sripinyowanich and Ruangthai32), whereas weeks of ETAS® supplementation had no measurable effect on cortisol(Reference Takanari, Nakahigashi and Sato17). The exclusively male sample in Denben et al. (Reference Denben, Sripinyowanich and Ruangthai32) may partly account for the larger cortisol reduction, as higher baseline cortisol in the male sample could allow for larger measurable decreases following intervention(Reference Reschke-Hernández, Okerstrom and Edwards47). Differences in intervention duration (12 v. 4 weeks) and formulation (E20 v. ETAS®) likely also contributed. Although Takanari et al. (Reference Takanari, Nakahigashi and Sato17) did not find significant improvements in serum or salivary cortisol in healthy populations, they reported an increase in salivary immunoglobulin A. Given that lower levels of salivary immunoglobulin A have been associated with perceived stress and depressive symptoms(Reference Engeland, Hugo and Hilgert48), this may reflect improvements in mood, consistent with previously observed reductions in depression–dejection. In addition to reductions in cortisol, in a pre-post-test design study, Huang et al. (Reference Huang, Zhou and An22) reported decreases in circulating norepinephrine alongside trends toward increased serotonin and dopamine, suggesting modulation of stress-related neuroendocrine responses. These biomarkers are closely linked to affective regulation and stress responsiveness and may contribute to downstream effects on mood and cognitive function(Reference Arnsten49). Collectively, these findings indicate that asparagus supplementation may influence peripheral stress-related biological systems with relevance to affective and cognitive outcomes.
Sleep
In relation to sleep outcomes, across two studies by Kato et al. (Reference Kato, Shoichiro and Sato29,Reference Kato, Inoue and Sato30) , participants reported more rested feelings and reduced insomnia symptoms. However, daytime sleepiness (measured by Epworth Sleepiness Scale) did not improve, suggesting asparagus interventions may primarily improve nocturnal sleep quality rather than daytime alertness(Reference Kato, Inoue and Sato30). This may partly reflect the distinct constructs captured by sleep measures, as PSQI and Athens Insomnia Scale assess subjective nocturnal sleep quality and insomnia symptoms, whereas Epworth Sleepiness Scale reflects daytime alertness(Reference Kawaratani, Miyaaki and Hiraoka50). Daytime alertness is influenced by multiple factors beyond sleep quality, particularly dietary composition and meal timing, which can affect sleep architecture and next-day alertness, as dietary patterns have been shown to influence both night-time sleep and related daytime functioning(Reference St-Onge, Mikic and Pietrolungo51). As dietary intake was not measured or controlled in most included studies, its potential influence on daytime sleepiness could not be accounted for, which may help explain why asparagus interventions improved nocturnal sleep quality without corresponding reductions in daytime alertness. Emotional stress and stressful life events have also been shown to influence daytime functioning independently of nocturnal sleep quality, indicating that stress-related factors may persist even when night-time sleep improves(Reference Salahuddin, Barroso and Leserman52). Taken together, these findings suggest that improvements in nocturnal sleep following asparagus supplementation may not translate directly into enhanced daytime alertness due to the persistent influence of diet, circadian rhythms and stress. Converging evidence comes from Takanari et al. (Reference Takanari, Nakahigashi and Sato17) who also reported improved self-reported sleep quality through the use of an unvalidated questionnaire limit comparability. In addition, Cong and Xu(Reference Cong and Xu34) observed significant improvements in self-reported sleep quality and objectively measured deep sleep continuity (Huawei Sport Band 7) only in the combined GABA and asparagus extract group, whereas the GABA-only group showed no measurable benefits. Although GABA was included in both formulations, the superior outcomes in the combined group suggest that asparagus extract itself likely plays a key contributory role in producing these sleep improvements.
While improvements are evident in healthy adults, sleep-disturbed populations demonstrated more pronounced and consistent benefits. Ito et al. (Reference Ito, Goto and Takanari27) observed reduced insomnia scores (as measured by the Athens Insomnia Scale) and fewer dreams/nightmares (as measured by OSA-MA). These effects were particularly evident in participants who slept longer than average. This may reflect a normalising effect, aligning sleep duration with healthier ranges, as both insufficient and excessive sleep are associated with adverse outcomes(Reference Chattu, Manzar and Kumary53,Reference Ohayon, Reynolds and Dauvilliers54) . Nishimura et al. (Reference Nishimura, Takahashi and Kohzai33) reported improved sleep quality (as measured by the PSQI) after 2 weeks of dried asparagus powder intake, although the small male sample size limits inferences. In pre-post-test studies without control groups, Sakai et al. (Reference Sakai, Nagata and Nagata19) found improved sleep quality (as measured by Visual Analogue Scale), but not efficiency, following 4 weeks of Kenja-no-Kaimin, while Huang et al. (Reference Huang, Zhou and An22) observed improvements across multiple PSQI domains, including sleep latency, total sleep time, awakenings and subjective sleep quality following high-dose supplementation (24 g/d). Takahashi et al. (Reference Takahashi, Seol and Kokubo35) reported improvements across all OSA-MA domains after 2 weeks of ETAS®, although electroencephalography outcomes did not change significantly. By contrast, the larger sample in Deguchi et al. (2019) showed lower Karolinska Sleepiness Scale scores and longer total sleep time. Karolinska Sleepiness Scale is generally highly correlated with electroencephalography(Reference Shahid, Wilkinson and Marcu55), suggesting that Takahashi et al. (Reference Takahashi, Seol and Kokubo35) may have been underpowered. Overall, these studies suggest that asparagus-related interventions can improve sleep quality, particularly in sleep-disturbed populations, though methodological heterogeneity and small sample sizes in some studies warrant cautious interpretation.
Mechanistically, several pathways may contribute to sleep-related effects observed with asparagus supplementation. One known mechanism is ETAS’s® modulation of neurons in the suprachiasmatic nucleus and melatonin receptors, with evidence that ETAS® may increase suprachiasmatic nucleus neuron number and normalises melatonin receptor 1 and 2 expression(Reference Chan, Wu and Wu56). ETAS has also been shown to increase HSP70 expression, potentially enhancing cellular stress resilience and supporting sleep regulation(Reference Ito, Maeda and Goto37). Complementing this cellular stress pathway, inhibitory neurotransmission via GABA plays a central role in sleep regulation, as GABAergic signalling in key brain regions suppresses wake-promoting neuronal activity and facilitates sleep initiation and maintenance(Reference Gottesmann57). The study by Cong and Xu(Reference Cong and Xu34) further suggests that asparagus extract, when combined with GABA, improved deep sleep continuity more than GABA alone, raising the possibility that asparagus may modulate GABAergic signalling. Together, these mechanisms highlight how asparagus extract may influence multiple neurophysiological targets that support sleep quality.
Strengths, limitations and gaps
Most studies included in this review were randomised controlled trials, which provide relatively strong internal validity by reducing bias and controlling for confounding variables(Reference Hariton and Locascio58). In contrast, quasi-experimental and uncontrolled pre-post-studies(Reference Sakai, Nagata and Nagata19,Reference Huang, Zhou and An22,Reference Cheng, Pan and Wang28) offered preliminary mechanistic and exploratory insights despite inherent limitations, including greater susceptibility to selection bias, regression to the mean, placebo effects, confounding variables and poorer internal validity(Reference Flannelly, Flannelly and Jankowski59). Therefore, findings derived from these lower-quality designs should be interpreted cautiously.
Across studies, common limitations included varied (and generally small) sample sizes (10–60), male-dominant cohorts and short intervention periods. In addition, supplementation formulations and doses varied widely (ETAS®, Kenja-no-Kaimin, ONR-8, asparagus power, E20), limiting cross-study comparability. Moreover, bioactive components were often not consistently reported, and in some cases insufficiently characterised, reducing reproducibility and making meaningful comparisons across trials challenging(Reference Coskun, Wise and Kuszak60). Most studies also did not take participants’ diets into account, which may have influenced the observed effects, suggesting that future studies should monitor or control dietary intake(Reference St-Onge, Mikic and Pietrolungo51).
A notable gap concerns individuals with cognitive impairment, as only two studies to date have examined this population(Reference Zenimoto and Takahashi20,Reference Mikhailichenko, Lai and Wei21) . Cognitive decline is commonly accompanied by hypothalamic–pituitary–adrenal axis dysregulation(Reference Joshi and Praticò61), inflammation(Reference Leonardo and Fregni62), oxidative stress(Reference Papathanasiou, Fradelos and Malli63) and sleep disruption(Reference Yaffe, Falvey and Hoang64); These are pathways asparagus interventions appear to influence, indicating a clear potential for further investigation.
Future well-powered research employing standardised dosing, consistent supplement formulations, balanced sex representation and validated outcome measures across the domains of cognition, affect, sleep and biomarkers is warranted. Mechanistic assessments, particularly cortisol, inflammatory markers and HSP70, should be integrated with clinical designs. Longer-term interventions are needed to assess the durability of effects, and studies should prioritise populations most likely to benefit, including those with sleep disruption and cognitive decline.
Conclusion
Asparagus-based supplementation appears to provide modest benefits across multiple domains, including cognition (reaction time, attention and fatigue), mood (anxiety, depression and reduced agitation) and stress-related physiology (reduced cortisol), and particularly quality and duration of sleep. Given the overlap between cognitive function, affective states, stress biomarkers and sleep, improvements in one domain may influence others, making it difficult to isolate a single therapeutic target. The observed effects are likely mediated by anti-inflammatory and stress-protective mechanisms, potentially involving hypothalamic–pituitary–adrenal axis regulation, reduced oxidative stress and modulation of HSP70. Notably, HSP70 induction may represent a distinctive mechanism of asparagus, as most other vegetables only exhibit HSP70 activity as part of their own stress response(Reference Usman, Rafii and Martini65), whereas asparagus supplementation has been shown to increase HSP70 expression in humans. Current evidence suggests that long-term, higher-dose interventions may be required to elicit meaningful cognitive, affective and cellular benefits, particularly in older adults or those with cognitive impairment or sleep disturbances, while lower doses and shorter interventions may suffice in healthy populations. Although findings from randomised controlled trials are promising, limitations such as small sample sizes, short intervention durations, heterogeneous supplement types, inconsistent reporting of bioactive components and failure to account for participants’ diet highlight the need for more rigorous, long-term studies with standardised protocols. Future research should focus on clarifying underlying mechanisms, optimising dosage and formulation, and evaluating efficacy in clinically relevant populations, especially those with cognitive impairment or sleep disruption.
Acknowledgements
None.
This research was conducted as part of a PhD studentship funded by Amino Up Co., Ltd and the University of Reading. Amino Up Co., Ltd had no role in the design, data collection, analysis, interpretation, or writing of the manuscript.
Abigail Davies: Data curation – Equal, Formal analysis-Equal, Methodology – Equal, Writing – original draft – Lead; Daniel Lamport: Conceptualisation – Equal, Supervision – Equal, Validation – Equal, Writing – review & editing-Equal; Kohei Homma: Writing – review & editing – Equal; Jun Takanari: Writing – review & editing – Equal; Piril Hepsomali: Conceptualisation – Equal, Funding acquisition – Lead, Validation – Equal, Writing – review & editing – Equal.
A. D., D. L. and P. H. have received research funding, consultancy, travel support and/or speaking fees from various industrial companies. K. H. and J. T. are employees of Amino Up Co., Ltd.
All relevant data are within the paper.

 Open access
Open access