


Spanish colonialism introduced new animals, plants, production systems, and foodways to the Americas. Religious missions were one of the main vehicles by which this transmission occurred. Archaeological investigations into Spanish missions have changed over time (Funari and Senatore Reference Funari and Senatore2015; Graham Reference Graham1998; Panich and Schneider Reference Panich and Schneider2014; Van Buren Reference Van Buren2010). Although initial research focused on architectural structures and mission conservation, these interpretations failed to address dynamic social and cultural interactions taking place in these landscapes. Since the 1990s, archaeologists have approached missions as sites of interaction among European, Indigenous, and African and Afrodescendant peoples. Increasingly, researchers highlight the agency with which Indigenous people negotiated their interactions with the Spanish, including through their knowledge of local foodways and their labor as related to plant and animal procurement and production (Panich and Schneider Reference Panich and Schneider2014). Recent research addresses, for example, transformations and persistence in identity and culture; the biological impacts of plant, animal, and pathogen translocations; and the ways these topics relate to wider global integration (Van Buren Reference Van Buren2010). The archaeology of Spanish missions today is centered on investigating the multidirectional negotiation of power, land, and lifeways in a multiethnic and quickly changing landscape. A key lens through which to do this is the archaeology of foodways.
Foodways capture not only what people eat but also what methods of resource management and procurement they use; the activities involved in and rules that govern how foods are prepared, consumed, discarded, and given meaning; and the development of regional or cultural cuisines (Peres Reference Peres2017). Studies of foodways are relevant to wider discussions of trade, labor, and gender, race, ethnicity, and class identities. The growing field of historical zooarchaeology addresses these themes (Landon Reference Landon2005; Landon and Opishinski Reference Landon, Opishinski, Orser, Zarankin, Funari and Lawrence2020; Tourigny and Gordon Reference Tourigny and Gordon2023). In North America, this research largely focuses on European colonization, attempts to replicate aspects of Eurasian food systems, and the ways Indigenous people chose to respond, leading to the forging of new foodways in areas that were being colonized.
Zooarchaeologists have demonstrated considerable variation across the missions, presidios, ranches, mining towns, and other settlements of northern New Spain in the degrees to which Eurasian domestic and native wild fauna were exploited. In the southeastern United States, most Spanish colonial sites incorporated terrestrial and aquatic food sources that would not have been part of traditional Iberian diets (Reitz and Waselkov Reference Reitz and Waselkov2015), whereas in the Southwest and California, cattle- and sheep-based ranching systems developed early, supporting burgeoning industries that demanded not only food but hides, wool, or tallow (Pavão-Zuckerman Reference Pavão-Zuckerman2011).
Lying between these two regions is Tejas: present-day central and southeastern Texas and western Louisiana. Tejas was sufficiently distant from Mexico City to be neglected by colonial administrators, creating a borderlands milieu in which Spanish, French, and Indigenous peoples interacted with relative autonomy, potentially shaping foodways. Ranching, a key element of Texas foodways today, is rooted in Spanish colonialism. Spanish missionaries and settlers introduced livestock and management practices, and ranching depended on Indigenous labor. Feral livestock populations would later form the foundations of modern ranching (Jackson Reference Jackson1986). Despite this well-documented history, direct archaeofaunal evidence for Spanish ranching is thin, and the relationship between sites of production and consumption is unclear. Five of the six San Antonio missions, for example, were each attached to a rancho that provisioned them. But only one of these—Rancho de las Cabras—has been excavated and analyzed (Taylor and Fox Reference Taylor and Fox1985). Most of our knowledge comes from missions and presidios. Although there is a robust archaeological record for Spanish missions in Texas (Fox Reference Fox1991; Hester Reference Hester and Thomas1989), most faunal data are hidden in gray literature, and only two presidios are well documented (Fradkin and Walter Reference Fradkin and Walter2018; Pavão-Zuckerman and Loren Reference Pavão-Zuckerman and Loren2012). Here, we contextualize new zooarchaeological research at Mission Dolores within the first review of Tejas mission and presidio faunal data.
Mission Dolores was founded at the far eastern reaches of Spanish control, along the main artery to Mexico City, and near the New France border. In this milieu, where missionaries would be in regular contact with the Ais and other Caddo-speaking Indigenous groups, and with their French neighbors, we might expect a transformation of European foodways into something new. To our knowledge, the only rancho that may have been associated with Mission Dolores is that of Antonio Gil Y’Barbo, but there is no archaeological or documentary evidence to indicate its exact location or its relationship with the mission (Iruegas and Iruegas Reference Iruegas and Iruegas2018). Written records, described further below, suggested that Mission Dolores residents were failing to thrive and failing in their mission to attract converts. Our research questions for zooarchaeological analysis therefore centered on provisioning strategies, livestock management, and culinary practices. They include the following:
• Research Question 1 is foundational: What were the provisioning strategies at Mission Dolores? What animals were people eating, and what proportion of the animal-based diet was obtained from domesticates compared to wild fowl, game, or fish? What do these proportions suggest about subsistence activities and potential relationships with other groups?
• Research Question 2 centers on the domestic proportion of the fauna. Does livestock species composition reflect the preferences of the missionaries, local ecology, or the impacts of raiding? Is it possible to make inferences from mortality profiles about livestock management for meat, milk, wool, traction, or other animal-derived products and labor?
• Research Question 3 investigates culinary practices. To do so, we first determine which taphonomic processes are observable, which could bias our interpretation of the faunal remains. Next, we ask what carcass parts are represented, and whether this evidence indicates butchery on-site, obtaining animal parts through trade, or other processes. Does evidence for food processing reveal a regional or cultural pattern of culinary practices?
These questions were guided by what is known of other Spanish mission foodways, summarized below. Our findings contribute to a growing understanding of the variation in animal-based foodways across the Spanish borderlands of North America.
Spanish Colonial Foodways and Zooarchaeology
Animal remains are a partial window on foodways. Unfortunately, plant-based foods are rarely preserved, recovered, or studied at the sites considered here. But historical zooarchaeology has great potential for comparison of written, faunal, and material records of foodways (Landon and Opishinski Reference Landon, Opishinski, Orser, Zarankin, Funari and Lawrence2020). Spanish colonists introduced a new suite of animals, including horses, donkeys, cattle, sheep, goat, pigs, and chickens. The on-the-ground reality of this aspect of the Columbian Exchange (Crosby Reference Crosby1972) is best understood through direct archaeological evidence (Pavão-Zuckerman and Reitz Reference Pavão-Zuckerman, Reitz and Smith2011). An emerging theme from zooarchaeology of the Spanish Americas is regional and site-specific variability in animal-based diets, reflecting geography, ecology, political control, and relationships with Indigenous groups.
Variability in Foodways across Spanish Americas
Much early work on Spanish colonial faunal assemblages was conducted in the southeastern United States (Peres Reference Peres2017, Reference Peres, Peres and Marrinan2021, Reference Peres2023; Reitz Reference Reitz1992a, Reference Reitz1992b; Reitz et al. Reference Reitz, Pavão-Zuckerman, Weinand, Duncan and Thomas2010). This work demonstrated how the Spanish—and their livestock—struggled to adapt to humid environments, often turning to local resources likely obtained through Indigenous labor. For example, at St. Augustine in La Florida, residents consumed diverse wild terrestrial and aquatic animals—as well as pig and chicken, which were better adapted to the humid climate (Reitz Reference Reitz1992b). Reitz and Waselkov (Reference Reitz and Waselkov2015) demonstrate that such experimentation and reliance on wild fauna was the norm in the earliest phases of colonization but that southeastern foodways eventually consolidated around cattle ranching. Elsewhere, even the earliest settlements were focused on cattle; for example, at Puerto Real, Haiti (Delsol Reference Delsol2024a; Reitz Reference Reitz1986).
In the Southwest and California, Spanish colonists were generally more successful in introducing domesticates (Pavao-Zuckerman and Reitz Reference Pavão-Zuckerman, Reitz and Ubelaker2006, Reference Pavão-Zuckerman, Reitz and Smith2011). This may be partially due to the adaptability of caprines and cattle to arid and semiarid lands, where their ability to convert grass into protein is especially valuable. The success of ranching and burgeoning industries reliant on animal products in the Southwest and California depended heavily on Indigenous labor (Lightfoot Reference Lightfoot2005; Noe Reference Noe2022; Pavão-Zuckerman Reference Pavão-Zuckerman2011; Spielmann et al. Reference Spielmann, Clark, Hawkey, Rainey and Fish2009). Whereas southeastern Indigenous groups generally resisted adoption of livestock until the nineteenth century (Pavão-Zuckerman and Reitz Reference Pavão-Zuckerman, Reitz and Ubelaker2006), southwestern Indigenous peoples often rapidly incorporated these animals into their lives, with studies examining not only the impacts of livestock on Indigenous political and economic systems but also the forging of novel foodways and crafts (Campbell Reference Campbell2021; Gifford-Gonzalez and Sunseri Reference Gifford-Gonzalez, Sunseri and Twiss2007; Noe Reference Noe2022, Reference Noe2023; Sunseri Reference Sunseri2017).
The Pimería Alta region on the Arizona-Mexico border has been especially well studied (Mathwich and Pavão-Zuckerman Reference Mathwich and Pavão-Zuckerman2018; Pavão-Zuckerman Reference Pavão-Zuckerman2011, Reference Pavão-Zuckerman, Douglass and Graves2017; Pavão-Zuckerman and LaMotta Reference Pavão-Zuckerman and LaMotta2007; Pavão-Zuckerman and Martínez-Ramírez Reference Pavão-Zuckerman and Martínez-Ramírez2020). Variation in caprine versus cattle ranching strategies—and in degrees of reliance on wild resources—can be linked to local ecologies, connections to nearby markets, and the threat of raiding (Pavão-Zuckerman Reference Pavão-Zuckerman, Douglass and Graves2017; Pavão-Zuckerman and Martínez-Ramírez Reference Pavão-Zuckerman and Martínez-Ramírez2020; Spielmann et al. Reference Spielmann, Clark, Hawkey, Rainey and Fish2009). For example, at Mission Cocóspera, there was rarely a resident priest, giving O’odham residents greater freedom (Pavão-Zuckerman Reference Pavão-Zuckerman, Douglass and Graves2017). This mission sat at high elevation and suffered occasional raids; faunal analysis revealed relatively greater reliance on wild game, perhaps as a coping strategy (Pavão-Zuckerman and Martínez-Ramírez Reference Pavão-Zuckerman and Martínez-Ramírez2020). In contrast, nearby Mission San Agustín sat at a lower elevation, with hot, dry conditions. Insulation from other communities meant fewer raids, and there was a more regular presence of Spanish priests; the dominance of cattle is linked to both ecological factors and greater colonial supervision (Pavão-Zuckerman Reference Pavão-Zuckerman, Douglass and Graves2017). At both missions, cattle carcasses were being processed for tallow. The Pimería Alta shows that patterns of animal resource use in this area have unifying themes in ranching, raiding, and rendering, with site-specific differences in reliance on wild game and domesticates (Pavão-Zuckerman Reference Pavão-Zuckerman, Douglass and Graves2017).
Zooarchaeological research in Latin American colonial contexts makes for informative comparisons to the southern United States (Delsol Reference Delsol2024a), but a detailed review is beyond our scope. Although many studies in Central and South America demonstrate the impact of Eurasian domesticates on local foodways (deFrance and Hanson Reference deFrance and Hanson2008; Delsol Reference Delsol2024b; Kennedy et al. Reference Kennedy, Chiou and VanValkenburgh2019; Kennedy and VanValkenburgh Reference Kennedy and VanValkenburgh2016), other sites show continuity of Indigenous foodways with minimal integration of introduced livestock (e.g., deFrance et al. Reference deFrance, Wernke and Sharpe2016). Recent work has also focused on carcass processing and Indigenous labor (e.g., Delsol Reference Delsol2020). There is potential here to draw broader conclusions about transformation of both Indigenous and colonizer foodways, as well as impacts on animals themselves (deFrance Reference deFrance2010; Delsol et al. Reference Delsol, Stucky, Oswald, Cobb, Emery and Guralnick2023).
Historical Zooarchaeology in Spanish Tejas
Contrasting with the above discussion, the zooarchaeology of Spanish colonialism in Tejas is poorly known, with most relevant data found in report appendices, student work, or other gray literature. Here, we provide an initial scholarly review of this corpus (Figure 1; Supplementary Material 1). Although there were nearly three dozen missions in Texas, only eight (including Mission Dolores) have archaeofaunal data. Faunal data are reported from three of the San Antonio Missions in central Texas—San José, San Juan, and Concepción (Clark Reference Clark1978; Hard et al. Reference Hard, Fox, Cox, Gross, Meissner, Mendez, Tennis and Zapata1995; Henderson and Clark Reference Henderson and Clark1984; Schuetz Reference Schuetz1969; Scurlock and Fox Reference Scurlock and Fox1977); from two of the three locations of Mission Espíritu Santo, one of the two in Victoria County (Mission Valley, 41VT11; 41VT10 has no faunal data) and one in Goliad County (41GD1); and from Missions Rosario and Refugio, also at Goliad, in the south Texas coastal plains (deFrance Reference deFrance1999; Gilmore Reference Gilmore1974; Tennis Reference Tennis2002).
Map of eastern Texas and Louisiana—showing mission and presidio sites with faunal data—and the Camino Real de los Tejas. Basemap from Natural Earth (naturalearthdata.com); Camino Real data from National Parks Service (https://irma.nps.gov/DataStore/Reference/Profile/2238909).

These missions reveal diverse procurement strategies alongside livestock management, including hunting deer, trapping small game such as rabbits and turtles, catching fish, and collecting shellfish. The kinds of wild animals pursued, and the relative contributions of different domesticates, vary across sites in ways that we discuss along with the Mission Dolores fauna below. Although several mission reports provide little more than faunal lists, others consider (1) spatial variation in faunal distribution, as at Mission San Juan (Schuetz Reference Schuetz1969), or (2) diachronic trends, as at Mission San José, where a shift from cattle to caprines is seen as a response to cattle raiding (Clark Reference Clark1978; Hard et al. Reference Hard, Fox, Cox, Gross, Meissner, Mendez, Tennis and Zapata1995), and at Mission Refugio, where there is an increase in fishing over time (Tennis Reference Tennis2002). Finally, DeFrance (Reference deFrance1999) attributed variation in faunal remains across Missions Espíritu Santo (two locations) and Rosario to the cultural traditions of Indigenous groups living at the sites.
More recently, zooarchaeologists have studied two Tejas presidios: Presidio San Sabá, around 200 km northwest of San Antonio (Fradkin and Walter Reference Fradkin and Walter2018), and Presidio Los Adaes in present-day Louisiana, around 50 km east of the Texas border (Pavão-Zuckerman and Loren Reference Pavão-Zuckerman and Loren2012). Because these analyses are recent and in-depth, they are useful here, although we recognize that military and religious sites will differ considerably—most notably in that Indigenous peoples were encouraged to live at the missions, whereas presidios were fortified against some Indigenous groups. But even more so than the missions, presidios were expected to be self-sufficient (as was also the case for Mission Dolores), which makes them appropriate comparative datasets.
Presidio San Sabá was one of the largest Spanish settlements in the northern borderlands, with a peak resident population of nearly 400 soldiers and families, and it also served as a base for settlers expanding northward (Chipman and Joseph Reference Chipman and Joseph2010; Fradkin and Walter Reference Fradkin and Walter2018). Written records attest that the delivery of food and goods was unreliable, that the presidio faced regular threats from Comanche groups, and that presidio residents coped with malnutrition and disease. This informed resource availability. Fradkin and Walter (Reference Fradkin and Walter2018) show that the residents relied on livestock, particularly caprines, which were well adapted to the hilly Edwards Plateau; cattle were also a key part of the diet, but pig and chicken were less common. Wild fauna—absent from written records—were few but present; these included deer, fish, ducks, and turtles. They may have been procured by the Spanish or obtained via trade with Lipan Apache (Walter Reference Walter2021).
Presidio Los Adaes is particularly relevant as the site closest to Mission Dolores, approximately 80 km to its west. It served as the capital of Tejas from 1729 to 1772 (Chipman and Joseph Reference Chipman and Joseph2010; Gregory et al. Reference Gregory, Avery, Aubra and Jay2004; Marceaux Reference Marceaux2011) and was established as part of a broader effort to construct six East Texas missions (including Mission Dolores) to protect against French encroachment from the east at Natchitoches and to maintain good relationships with Caddoan groups. The Caddo Nation had local power and influence, and Spanish administrators wanted to dissuade their alliances with the French (Gregory et al. Reference Gregory, Avery, Aubra and Jay2004). But in reality, the presidio functioned as a center of exchange among Spanish, French, and Caddo. Los Adaes and East Texas missions were closer to their French neighbors than to any major Spanish settlements, and these neighbors relied on each other despite being enemies on paper (Galán Reference Galán2020).
In this unique context, it is no surprise that the faunal remains recovered from Los Adaes are distinct from those at San Sabá, even though both presidios were started with substantial livestock herds and an expectation of self-sufficiency. Written records suggest that although the large numbers of livestock at Los Adaes were thriving, crop failures led to food shortages (Pavão-Zuckerman and Loren Reference Pavão-Zuckerman and Loren2012). Pavão-Zuckerman and Loren (Reference Pavão-Zuckerman and Loren2012) note that caprines were relatively uncommon at Los Adaes and would have had trouble adapting to the humid climate, whereas cattle—although not in an ideal climate—would have had sufficient grasslands for grazing, and indeed these are the most common domesticate. But the presidio residents also ate a substantial amount of wild fauna, including deer and small mammals, birds, fish, turtles, and alligator—several of which were considered unsavory by Spanish cultural standards.
Pavão-Zuckerman and Loren (Reference Pavão-Zuckerman and Loren2012) coanalyzed fauna and ceramics from the governor’s house, the house of a low-middle-class European settler (potentially a French trader), and a likely mixed-ethnicity lower-class household. The governor’s household had ceramics that were expected of Spanish elites, and preserved skeletal elements that suggested tastier cuts of meat. However, every household—even the governor’s—was eating similar amounts of wild fauna overall. This suggests the difficulty of obtaining foods that matched cultural expectations (Pavão-Zuckerman and Loren Reference Pavão-Zuckerman and Loren2012). These results reflect the subsistence challenges and possibly creation of new identities even amid differences in class and ethnicity.
With these frameworks in mind, we initially approached the Mission Dolores assemblage seeking to understand foodways particular to this site and to compare our results to Presidio Los Adaes. We expected that we might find a similar dependence on wild resources, working on the assumption that humid eastern Texas would be less conducive to ranching than central Texas or the coastal plains. In addition to characterizing the assemblage in basic terms of taxonomy, we also conducted an analysis of butchery, initially seeking culturally specific culinary practices.
Butchery Analysis and Culinary Practice at Spanish Colonial Sites
Butchering an animal is a culturally informed process (Seetah Reference Seetah2019). The analysis of cut marks to investigate butchery practices has long been established, especially in prehistoric archaeology (Domínguez-Rodrigo and Pickering Reference Domínguez-Rodrigo and Pickering2017). Although taphonomic concerns came later to historical zooarchaeology, bone surface modifications, bone breakage, and implications for carcass processing have recently been centered in many studies (e.g., Heinrich Reference Heinrich2014; Pavão-Zuckerman Reference Pavão-Zuckerman2011; Pavão-Zuckerman and LaMotta Reference Pavão-Zuckerman and LaMotta2007; Seetah Reference Seetah2019; Sunseri Reference Sunseri2017).
Much of this research aims to understand desired end products (meat, tallow, etc.) through taphonomic analysis. Scholars also examine cultural patterns of butchery. For example, Richard Gilmore (Reference Gilmore1999) compared butchery patterns from Spanish and British colonial sites with the goal of characterizing cultural butchery practices and making inferences about culturally specific foodways and cuisine. Gilmore found key differences in the butchery process for caprines between Spanish and English colonial sites and related this to cuisine. Although it is tempting to determine a culturally Spanish pattern of butchery, there are some drawbacks to this study. Gilmore's (Reference Gilmore1999) data came from a nonrandom sample of remains from a range of sites, each with its own formation processes—including degrees of carnivore involvement—which bias the relative abundances of skeletal elements, specific portions, and therefore cut-mark frequencies. This study did not include limb shafts, which in the decades since Gilmore’s work have been increasingly recognized as fundamental (e.g., Pickering et al. Reference Pickering, Marean and Domı́nguez-Rodrigo2003). In our work, we found it difficult to replicate Gilmore's (Reference Gilmore1999) descriptive taxonomy of cut, slice, chop, and shear marks.
In paleoanthropological sites, research focuses on confirming the presence of cut marks, eliminating other confounding variables, and interpreting the results in terms of human agency. This often involves statistical analyses of the spatial distribution of cut marks across elements, most recently by plotting them in three-dimensional (3D) space (Domínguez-Rodrigo, Baquedano, et al. Reference Domínguez-Rodrigo, Baquedano, Organista, Cobo-Sánchez, Mabulla, Maskara and Gidna2021; Domínguez-Rodrigo, Gidna, et al. Reference Domínguez-Rodrigo, Gidna, Baquedano, Cobo-Sánchez, Mora, Courtenay, Gonzalez-Aguilera, Mate-Gonzalez and Prieto-Herráez2021). Here, we show that methods used in paleoanthropology can be easily transferred to historical assemblages, which are often well preserved, making cut-mark analysis straightforward. The Mission Dolores assemblage provides a test case for this approach.
The Mission Dolores Site
History of Mission Dolores
Mission Dolores (41SA25) is a State Historic Site in San Augustine County managed by the Texas Historical Commission (THC). It was part of a group of missions and presidios founded to convert the Indigenous population to sedentary and devout Catholic lives and defend against French encroachment (Figure 1). In the early 1700s, the French had established a stable and thriving settlement at Natchitoches, maintaining amicable relationships with the powerful Caddo Nation via trade and intermarriage. Administrators in Mexico City saw this as a threat to Spain’s claims (Chipman and Joseph Reference Chipman and Joseph2010; Gregory et al. Reference Gregory, Avery, Aubra and Jay2004). In 1717, six missions and two presidios were established to secure the eastern border. They were poorly provisioned and were abandoned in 1719 following French raids on Presidio Los Adaes, and they were reestablished in 1721 (Chipman and Joseph Reference Chipman and Joseph2010). However, half of the missions closed by 1730, leaving only Mission Dolores, Mission Nuestra Señora de Guadalupe de los Nacogdoches, and Mission San Miguel de Linares de los Adaes. The missions were located along the Camino Real de los Tejas, a central artery of economic and cultural exchange stretching from Natchitoches via San Antonio to Mexico City.
Mission Dolores was reestablished with about 10–20 Spanish inhabitants, approximately 1.2 km from its AD 1717 location, in a small open prairie, surrounded by forest, along a creek, and overlooking the Ayish Bayou—a freshwater tributary of the Angelina River (Chipman and Joseph Reference Chipman and Joseph2010; Corbin et al. Reference Corbin, Brown, Canavan and Toups1990). Because of the steep topography between the mission and the creek, irrigation of agricultural fields was not possible. It is unclear what pasture was available for livestock.
Although the Hasinai Caddo were the most prominent Indigenous group in the region, the Ais were the focus of conversion efforts. The Ais numbered around 70 families and roughly 300 individuals living within 10 km of Mission Dolores. Apart from occasional baptisms, multiple accounts indicate there were no Ais living at the mission (Carlson and Quinn Reference Carlson and Quinn1996; Corbin et al. Reference Corbin, Brown, Canavan and Toups1990; Kress et al. Reference Kress, Fray Gaspar José and Mattie Austin1931; Magnaghi Reference Magnaghi1984). One account states that the Ais did congregate briefly at Mission Dolores, until the lack of resources prompted them to leave (Nunley Reference Nunley1975).
The conversion efforts at Mission Dolores—as elsewhere in East Texas—were therefore a failure. Although relations between Caddo Nation peoples and Europeans were generally peaceful at this time, the Caddo saw no need to settle at Spanish missions, which, according to written records, failed to raise the most basic crops to support themselves. In contrast, the Caddo Nation held considerable political power, with a large population, strong alliances with other tribes, a diversified food system, and well-developed exchange networks (Marceaux and Wade Reference Marceaux, Wade, Panich and Schneider2014).
When the French ceded their claim on Louisiana to the Spanish, the East Texas missions lost their primary justification. In 1767, the Marqués de Rubí completed an inspection of Spanish settlements across northern New Spain and noted inadequate military defenses at the presidios and the failure of the missions to attract converts, recommending their closure. In 1773, all East Texas missions and presidios were formally abandoned. At Mission Dolores, the architecture fell into disrepair, its location forgotten until the twentieth century.
Archaeology of Mission Dolores
Kathleen Gilmore led the first systematic attempt to relocate Mission Dolores in 1972–1973, following a 1962 survey by Edward B. Jelks, Dee Ann Suhm, and Lathel F. Duffield (Corbin et al. Reference Corbin, Alex and Kalina1980). Excavations focused on Mission Hill—long thought to be the mission’s location—north of State Highway 147, but these were not fruitful, and Gilmore indicated that future work should focus south of the highway. James Corbin directed such excavations with Stephen F. Austin State University volunteers and field schools in 1976, 1977, and 1978 (Corbin Reference Corbin1977; Corbin et al. Reference Corbin, Alex and Kalina1980). Most of Mission Dolores’s main features were identified in these seasons, including the southeast perimeter wall, three structures, four trash pits, and a well. Excavators recovered thousands of artifacts, including European and Indigenous ceramics, metals, glass, stone, and faunal remains. Animal bones were especially abundant in Features 8 and 18, which Corbin (Reference Corbin1977:3) noted were similar to refuse pits at Presidio Los Adaes. Corbin led additional excavations in 1984, expanding on some previously documented features; artifacts were similar to those previously reported (Corbin et al. Reference Corbin, Brown, Canavan and Toups1990). This work showed that most of the mission was destroyed by highway construction and previous logging activities, and that the remaining mission deposits were actively eroding.
Further archaeology in 1995–1996 was part of impact assessments when building a museum complex and parking lot (Carlson and Quinn Reference Carlson and Quinn1996). In these seasons, 360 shovel test pits and seven 1 m2 units were excavated. Only one unit produced many artifacts, of which only one bead was definitively associated with the mission period. Finally, in 2008, geophysical surveys of perimeter walls and shovel test pits were employed to calculate the mission’s dimensions in anticipation of a reconstruction and replication project (Avery Reference Avery2016).
Evidence for Foodways at Mission Dolores
Written records suggest that East Texas missions and presidios leaned heavily on trading relationships (Galán Reference Galán2008, Reference Galán2020). Mission Dolores occupants traded with the Spanish at Presidio Los Adaes and the French at Natchitoches (Chipman and Joseph Reference Chipman and Joseph2010; Marceaux Reference Marceaux2011). The French would often attend religious services at the Spanish missions, and relations were maintained through intermarriage and godparent relationships. The eighteenth-century French–Spanish border was a hub of cross-cultural contact, especially at Presidio Los Adaes, where half the soldiers were of mixed Spanish, Indigenous, and African heritage (Gregory et al. Reference Gregory, Avery, Aubra and Jay2004). Such cultural diversity makes East Texas mission and presidio foodways an intriguing subject of inquiry.
The 1721 expedition to reestablish the six East Texas missions brought 2,800 horses, 4,800 cattle, and 6,400 sheep and goats to distribute among the settlements along their journey (Chipman and Joseph Reference Chipman and Joseph2010). However, primary accounts specific to Mission Dolores are scant, and since we have no records of any associated ranchos, we assume that livestock rearing occurred at or near the site. Fray Gaspar José de Solís inspected Mission Dolores in 1768, noting the residents would “plant onions, garlic, cabbage, lettuce, and green vegetables of all kinds” and gather peaches, figs, and native fruits and nuts from a nearby orchard (Kress et al. Reference Kress, Fray Gaspar José and Mattie Austin1931:67). He noted “a few horses, about fifteen or twenty mules, about ten or twelve cows, and as many bulls, about sixteen or twenty oxen for farming” (Kress et al. Reference Kress, Fray Gaspar José and Mattie Austin1931). He also mentioned raiding: “The Indians steal as many as they can, the bulls, cows, and calves for eating, the horses, mules, and mares (of which there is a drove) for their commerce and traffic with the French” (Kress et al. Reference Kress, Fray Gaspar José and Mattie Austin1931:67). No sheep, goat, pigs, or chickens were mentioned by de Solís. Another firsthand account indicates that during the brief period during which some of the Ais may have lived on the mission, men worked as herdsmen and women would “use the metates” to perform culinary labor, although it is unclear what animals were herded, what was cooked, or how this labor was negotiated (Nunley Reference Nunley1975). Most other sources emphasize that besides this outlier period of residence, the Ais neither converted nor congregated at the mission, and they were generally disagreeable to interactions with the padres, suggesting there may be limited food exchange or sharing (Kress et al. Reference Kress, Fray Gaspar José and Mattie Austin1931:67–68; Magnaghi Reference Magnaghi1984). Documentary records do not mention either game procurement or trading.
Prior to this study, direct zooarchaeological evidence has been limited. Faunal remains were recovered from all of the above-mentioned campaigns, but analyses have been mainly conducted as student projects buried in archaeological reports. For the largest assemblage, from the 1976–1978 seasons, only a short qualitative report is provided, which noted abundant large bovid remains and possible remains of deer, caprines, and birds (Corbin et al. Reference Corbin, Alex and Kalina1980). A report from the 1984 excavation includes quantitative data on taxonomic and skeletal-part representation as well as notes on butchery marks (Corbin et al. Reference Corbin, Brown, Canavan and Toups1990). Among the 344 identified specimens listed in that report’s Table 11 (which shows discrepancies with Appendix VI in the same report), there are many cattle (Bos taurus) remains (more than half of identified specimens) and an important contribution of white-tailed deer (Odocoileus virginianus) remains (one-quarter of identified specimens), with few remains of domestic goat (Capra hircus), pig (Sus scrofa), and felid or canid (Felis and Canis sp.), which were listed under domestic animals. Wild fauna included turtles or tortoises, birds, fish, and shellfish. In this analysis, most fragments were grouped into either the large and robust “Bos taurus” category or the medium-sized and gracile “Odocoileus virginianus” category, even if those fragments were not identifiable to element.
Faunal remains from the impact assessment were analyzed, but none could be definitively related to the mission occupation (Carlson and Quinn Reference Carlson and Quinn1996). In total 1,951 specimens were identified, more than 97% of which were only identifiable to class and/or size (e.g., “large mammal”). Large bovids, a medium ungulate (deer or pronghorn), and small fauna (hare/rabbit, turtle, fish) were reported. Avery’s (Reference Avery2016) work identified a trash pit outside the perimeter wall that was linked to a soldier or priests’ house. Among the 48 identifiable specimens were cattle, fish, chicken, turtles, rabbit, and pig, and the more than 1,400 unidentified remains were mainly of large mammals.
Given the limits of the faunal assemblages described above, the best opportunity to understand foodways at Mission Dolores comes from the extensive 1976–1978 excavations with their abundant well-preserved faunal remains. These are the focus of our analysis—one part of a broader effort by the THC to rehabilitate the Mission Dolores collection. Additionally, we reanalyze the identified specimens from the 1984 assemblage. In parallel, archaeobotanical analysis of the 1976–1978 plant remains was completed by Leslie Bush (Reference Bush2023). When taken together with the faunal remains, we hope that these two lines of evidence will provide a far richer assessment of Texas mission foodways than has been possible until now.
Materials and Methods
We analyzed the vertebrate remains from the 1976–1978 excavations and reanalyzed the 1984 assemblage. There is a small invertebrate shell assemblage—appearing to be mostly oyster or other bivalve—that awaits identification. Prior to our analysis, the 1977–1978 vertebrate remains had been roughly organized in THC storage by degree of preservation (that is, the best-preserved context bags were boxed together). Because of limitations on our time and experience, and the limits of the available comparative collection, we chose to leverage this existing organizational system to prioritize the most complete and well-preserved material as well as bags from trash-pit features. Roughly 50% of the entire assemblage (1977–1978 and 1984 seasons combined), by weight, was evaluated in this project. By evaluate, we mean that all remains in each bag were examined and sorted into more and less diagnostic specimens, with the highly diagnostic specimens identified and catalogued. The other 50% of the assemblage consists of mostly fragmentary and less identifiable bone. These remains were only counted and weighed.
Because these are legacy collections, there are important limitations for interpretation. It was not always possible to link the accession numbers on faunal bags to excavated contexts. For the 1977–1978 excavations, nearly all of the remains could be linked to specific squares and/or features, although depths were inconsistently recorded. Most remains came from one of the trash pits (Feature 8) excavated in 1977, where fauna were well preserved (Figure 2). However, we had more difficulties determining the contexts of fauna excavated during the 1984 season, although the excavation report suggests that the majority of animal bone again came from trash pits akin (and in some cases adjacent) to those excavated in 1977 (Corbin et al. Reference Corbin, Brown, Canavan and Toups1990). In the 1984 assemblage, individual bones had been inked with lot numbers that are relatable to specific contexts, but the bones were then bagged by taxon and element, thereby mixing contexts. Today, the inked lot numbers are often illegible; we recorded them to the best of our ability in our reanalysis. Given these challenges, the simplest approach was to interpret the identified specimens from the two excavation seasons jointly, with all contexts as a single analytical unit.
Plan of Feature 8, interpreted as a trash pit. Redrawn after Corbin et alia (Reference Corbin, Alex and Kalina1980:Figure 35).

To investigate provisioning strategies at Mission Dolores, we recorded variables including taxon, element, portion, and symmetry, and we evaluated taphonomic marks for each identified specimen. Identification followed standard zooarchaeological methods, using the William F. McClure Faunal Collection in the Rice University Archaeology Lab and osteological guides (Balkwill and Cumbaa Reference Balkwill and Cumbaa1992; Barone Reference Barone1976; Gilbert Reference Gilbert1990; Hillson Reference Hillson2005; Pales et al. Reference Pales, Lambert and Garcia1971; Schmid Reference Schmid1972). Preliminary identifications by Orta were confirmed by Prendergast. For specimens initially identified as large mammal or large bovid (bison/cattle), the criteria of Balkwill and Cumbaa (Reference Balkwill and Cumbaa1992) were applied to provide, in many cases, more precise identifications.
In this analysis, fragments classified as not identified (NID) included those that could, with additional time and effort, be identified to element group, such as rib, vertebral, and limb shaft fragments; future inclusion of such portions would increase the robusticity of taphonomic analysis, particularly as limb shafts preserve highly informative butchery marks. Additionally, teeth that were less than half complete, bone fragments under approximately 7.6 cm (3 inches) in size (unless highly diagnostic), or fragments that lacked any distinguishing features were generally considered NID. Taxonomic identifications frequently made use of size classes. In this analysis, large mammals are bison, cattle, and horse sized; medium mammals are deer, caprine, and pig sized; and small mammals are dogs, cats, and everything smaller (except micromammals, such as rodents).
From the identified specimens, the minimum number of elements (MNE) and individuals (MNI) was estimated for each taxonomic group by laying all identified specimens out on a table by element group and comparing bone portions, symmetry, individual size, and fusion to obtain the most accurate estimates. Both epiphyseal fusion and tooth eruption/wear were used to estimate age at death, although only the latter was found useful for calculating mortality profiles due to its greater precision. For cattle, we coded dental-eruption/wear stages following Grant (Reference Grant, Wilson, Grigson and Payne1982) and used age-at-death correlations published by Janzen (Reference Janzen2015) and Jones and Sadler (Reference Jones and Sadler2012). For caprines, we coded dental-eruption/wear stages following Payne (Reference Payne1987) and Zeder (Reference Zeder1991) and used age-at-death correlations published by Zeder (Reference Zeder and Ruscillo2006). In practice, the number of caprines was so few that no mortality profiles could be constructed. Skeletal-part representation of the major mammal size classes was evaluated using the metric of survivorship for body portions as defined by Brain (Reference Brain1981), where survivorship (%MNI) is the MNE of a given skeletal element, relative to the expected number of elements in the assemblage based on the MNI. In our survivorship analysis, we grouped elements into body portions following Clark (Reference Clark2011).
To investigate butchery practices, we examined each identified specimen under oblique light with 10×–30× hand lenses, and we recorded cut marks as well as other visible marks. We followed criteria outlined by Domínguez-Rodrigo et alia (Reference Domínguez-Rodrigo, Barba Egido and Egeland2007) for cut, percussion, and abrasion marks; Domínguez-Rodrigo and Barba (Reference Domínguez-Rodrigo and Barba2006) for biochemical pits and etching (including root marks); and Backwell et alia (Reference Backwell, Parkinson, Roberts, d’Errico and Huchet2012) for insect marks. For cut marks, we followed the criteria of Greenfield (Reference Greenfield1999) and Olsen (Reference Olsen and Olsen1988) to distinguish marks made by metal tools as opposed to stone tools.
For cattle or large bovid remains with cut marks, we performed additional analyses. To record the precise position of cut marks on a bone in three dimensions, we employed Ikhnos open-source software (Mora et al. Reference Mora, Aramendi, Courtenay, González-Aguilera, Yravedra, Ángel Maté-González, Prieto-Herráez, Vázquez-Rodríguez and Barja2022). This is the first application of this software to a historic site, and there are some features that we found limiting. Models are only provided for the six long limbs that dominate many Plio-Pleistocene sites (humerus, radio-ulna, metacarpal, femur, tibia, metatarsal), but historic assemblages often include many axial elements not accounted for in Ikhnos. Furthermore, we could only use those fragments identifiable to the left or right sides for the analysis. For the fraction of cut-marked specimens that fit these criteria, the spatial distribution of cut marks on each specimen was recorded with a singular point representing the midpoint of each mark. Spatial data for each element/symmetry pair (e.g., all left femora) were then visualized on a single 3D model using R.
Results
Assemblage Overview
We examined 3,154 bone and tooth specimens, of which 81.5% (n = 2,572) were classified as nonidentified (NID) and 19% (n = 582) were identified. After accounting for modern breaks and refitting, we had a total number of identified specimens (NISP) of 485, representing a minimum number of individuals (MNI) of 19. Most identified fauna (60% of NISP) came from Feature 8, which reflects our prioritization of this feature and the best-preserved remains. Bones from Feature 8 were so well preserved that we could occasionally link specimens to excavation drawings and photographs. Corbin et alia (Reference Corbin, Alex and Kalina1980:414–415) noted that outside of pit features, preservation was poor. Indeed, only 11% of NISP came from other defined contexts (including other pits) in the 1977–1978 excavations, and another 3.5% came from undefined contexts. The remaining 25% of NISP came from our reanalysis of the 1984 assemblage.
Our selected subsample was well preserved, with good cortical preservation, little to no weathering damage, and a low degree of fragmentation. This enabled the identification of taphonomic signatures such as butchery marks. Biogenic marks—including root etching, other biochemical erosion, and insect marks likely made by termites—were extensive and easy to distinguish from anthropogenic marks (Supplementary Material 2). We noted qualitatively that most breakage planes were smooth and oblique, indicating breakage when bones were fresh or “green,” as opposed to postdepositional breakage. A detailed taphonomic analysis follows below.
Cattle-Dominant Taxonomic Representation at Mission Dolores
Cattle were the dominant meat source for Mission Dolores residents. We carefully reviewed criteria for Bison/Bos discrimination using postcranial elements, recording all visible morphological differences, as well as the success rates for each skeletal element according to Balkwill and Cumbaa (Reference Balkwill and Cumbaa1992). These results (Supplementary Material 3) show that of 40 bovine postcranial specimens with sufficient preservation for differentiation, 36 were identified as Bos taurus, most at high confidence. The remaining four specimens were more tentatively identified as possible Bison bison, but two of these had only one observable morphological feature, one of them had a mixture of typical Bos and Bison features, and the last specimen had bison-like features that Balkwill and Cumbaa (Reference Balkwill and Cumbaa1992) document as less reliable. We note that the morphologically ambiguous specimens are generally larger than other bovine remains. It is possible that there is at least one bison represented at Mission Dolores, but the preponderance of evidence suggests that most or all bovine specimens are cattle, and they are treated as such here.
Bos taurus is the taxonomic group with the highest NISP (39%) in the assemblage, and if these are combined with specimens identified as “Large mammal cf. Bos taurus” group, they represent 78% of the total NISP (Table 1; Figure 3). Given that NISP is highly sensitive to both fragmentation and identifiability, we also consider MNI estimates. Cattle give the highest MNI of any group (MNI = 7, or 37% of total), based on dentition. We believe that this is a conservative (low) estimate, and we note that cattle-sized remains dominate the unidentified fauna.
Relative abundance of mammalian taxa at Mission Dolores as a percentage of the total Number of Identified Specimens (%NISP) and Minimum Number of Individuals (%MNI), with domestic and wild fauna represented as separate color groups. Note that for this generalized representation, more cautious attributions (e.g., “cf. Felis catus”) are treated as definitive, and categories such as “cattle/likely cattle” and “deer/likely deer” are combined. (Color online)

Identified Taxa at Mission Dolores by Number of Identified Specimens (NISP) and Minimum Number of Individuals (MNI).

There are minor contributions from other domesticates at Mission Dolores. Caprines—either sheep (Ovis aries) or goat (Capra hircus), which could not be distinguished here—represent 6% of NISP and an MNI of 2, when including specimens more conservatively identified as likely caprines. Other probable domesticates include a canid and felid that are very likely Canis familiaris (NISP = 2) and Felis catus (NISP = 2), respectively; these are not thought to form part of the diet. It is notable that no domestic pigs or chicken were identified in the assemblage, nor was (wild or domestic) turkey identified, all of which are commonly reported at Spanish missions.
Hunting at Mission Dolores centered on deer, including both white-tailed deer (Odocoileus virginianus) and at least one specimen which, based on size and morphology (following Jacobson Reference Jacobson2003), is likely mule deer (Odocoileus hemionus). Cervids represent 6% of the NISP and an MNI of 4 when including specimens identified as likely cervid. There are scant small animal remains, including one hare/rabbit specimen, two catfish specimens, and some fragments of turtle/tortoise carapace. One indeterminate bird specimen was identified.
Macromammalian Skeletal-Part Representation
To analyze the survivorship of specific body parts, macromammalian specimens were grouped by carcass size into large (cattle), medium (caprines, cervids), and small (felid, canid, lagomorph) classes. For each size class, skeletal elements were grouped into body portions (axial, forelimb, etc.) following Clark (Reference Clark2011). Large mammals had higher overall survivorship of body portions compared to medium and small mammals (Table 2). For large mammals, portions with the highest survivorship are the skull (86%), forelimb (50%), and hindlimb (36%), which are robust parts of the skeleton that tend to preserve well. However, survivorship for large mammal feet is only 3%, even though those elements typically preserve well, leading to a head-and-foot pattern at many sites. The paucity of foot bones suggests that they may have remained attached to the hide during processing and were subsequently discarded. Medium mammals had overall lower survivorship than large mammals, and small mammals had the lowest survivorship overall, with each of the three individuals (cat, dog, hare/rabbit) being represented by scant limb bones (Figure 4).
Survivorship (%MNI) by carcass portion for large, medium, and small mammals. Survivorship is calculated as the MNE present, relative to the expected number of elements given the MNI for this size class. Here, survivorship is based on an MNI of seven large mammals (cattle), six medium mammals (cervids/caprines), and three small mammals (dog, cat, hare). (Color online)

Skeletal Part Representation and Survivorship at Mission Dolores.

Notes: Skeletal part representation is shown by Number of Identified Specimens (NISP) and Minimum Number of Elements (MNE), with elements grouped into body portions following Clark (Reference Clark2011). Survivorship (%MNI) is calculated as the MNE present, relative to the expected number of elements given the MNI for this size class.
Cattle Mortality Profile
Because cattle had the highest MNI, greatest survivorship, and the most complete dentition of the species represented in this assemblage, mortality profiles based on dental eruption and wear could be constructed; this was impossible for caprines due to small samples. Epiphyseal fusion patterns were also analyzed, but this did not result in greater resolution on age at death than dentition, nor did it indicate that young individuals were being overlooked by using dentition alone. For each of the seven cattle individuals, dental eruption and wear were used to conservatively determine a timespan during which the animal was slaughtered (Figure 5).
Mission Dolores cattle mortality profile, showing estimated age at death of each of seven individuals identified and aged based on dentition.

A single infantile individual could not have been more than six months old, and likely it was stillborn, or it died from illness or injury—assuming that veal would have been a luxury the missionaries could not afford. Four individuals were slaughtered between two and four years old, which falls within the prime age window during which cattle were slaughtered for beef in other historic North American contexts (Bowen Reference Bowen1998). One individual was slaughtered between six and eight years of age, and another was more than eight years old. This indicates that some cattle were kept alive longer than is typical for meat production, potentially for breeding, milk production, or traction. Unfortunately, no features diagnostic of sex were present to help address this question.
Taphonomy and Cut-Mark Frequency
Bone-surface modifications consisted of both anthropogenic and other biogenic marks (Supplementary Material 4). Due to a database error, both insect marking and percussion marks had their data deleted, but qualitatively, we noted that there was a very high prevalence of insect marks, likely from termites. In contrast, there were only a few instances of percussion marking, likely from occasionally smashing bones open for boiling; we did not note the high degrees of limb-bone fragmentation that have been associated elsewhere in the Spanish borderlands with tallow production. Root etching was present on 68% of NISP, and biochemical erosion (e.g., from fungi or bacteria in the burial environment) was present on 93% of NISP (these figures exclude teeth). Carnivore tooth marks were rare, observed on only 0.5% of NISP, although this may be a conservative underestimate due to the possibility of confusing marks with biochemical erosion. Rodent gnawing was present on 0.9% of NISP. Less than 1% of identified specimens (NISP), but 9% of unidentified (NID) specimens, were burnt, showing that although burnt bone was uncommon, most of it consisted of small, fragmented pieces that were not identifiable.
Cut marks were recorded on 28% of NISP across the assemblage as a whole. When focusing only on cattle, however, this frequency rises to 38% of NISP, which makes sense given their large body size and the extent of butchery required. In contrast, 29% of caprine remains and 19% of deer remains had cut marks, frequencies sensitive to the small sample sizes for these taxa (Table 3). The nonmammalian and small-mammal remains bore no observed cut marks.
Cut-Mark Frequency by NISP and Element for Large Bovids, Cervids, and Caprines.

Cattle elements with the highest percentage of cut marks include the lumbar vertebra (69% NISP), mandible (67% NISP), and innominate (64% NISP). For caprine and cervid elements, sample sizes are small, but they indicate that the tibia and metatarsal were often cut marked.
Butchery marks at Mission Dolores bear traits associated with metal tools, following Greenfield (Reference Greenfield1999) and Olsen (Reference Olsen and Olsen1988). Mission Dolores cut marks were regular in shape, sharply defined with a V-shaped cross section, and narrow and repetitive at the same location; these are features associated with metal tools. Most cross sections were symmetrical, with an incision axis largely perpendicular to the bone surface, which is also consistent with metal-tool marks. Some chop marks in the assemblage bear a more oblique angle. The Mission Dolores marks had no shoulder effects—that is, shallower striae parallel to the main groove. They also did not have cortical flaking visible at the surface adjacent to the mark. These traits are typical of stone-tool marks. We also observed few or no microstriations on the walls of the grooves, which are often associated with stone-tool marks. The few microstriations documented occur at the base of the grooves and indicate repetitive slicing, as is expected with use of metal tools. Traits associated with metal tools can be seen on both appendicular and axial bones, and on remains of cattle, caprines, and deer. Some magnified cut and chop marks are illustrated in Supplementary Material 5.
Three-Dimensional Visualization of Butchery Marks
For selected cattle specimens, the exact placement of the cut marks was recorded using Ikhnos to observe patterns of butchery. Due to limitations with this software, only a subsample of the cut-marked cattle specimens could be recorded: the six long limb bones, and only fragments identified to left or right sides (Supplementary Material 6). This excludes the aforementioned axial elements that have the highest occurrence of cut marks in the assemblage.
3D models for each element/symmetry combination of limb bones enabled us to visualize the distribution of all cut marks along the element for all specimens combined. For example, there were three right cattle humeri with cut marks identified in this analysis, which together had a total of 13 cut marks present. These were then recorded in 3D space on one model (Figure 6). To view all 10 models, see Supplementary Material 7.
Example of 3D representation of cut-mark distribution—in this case, on the right humerus of cattle. For additional models representing other elements and sides, see Supplementary Material 7.

Based on visual inspection of these models, most cut marks on limbs occur across the shaft, with the exception of the distal femur. The presence of cut marks on the midshaft implies removal of the meat from the bone directly from the carcass, whereas cut marks on the epiphyses implies dismemberment into more transportable sections. Based on these visualizations, it is apparent that there is less evidence for dismemberment in the cattle butchery patterns, suggesting that whole carcasses were processed for meat at the mission—an interpretation further explored below. This interpretation is underscored by the fact that our method was biased toward epiphyseal specimens because they were more likely to be attributed to a specific element and sided; midshafts are underrepresented in our analysis, yet they bear relatively more cut marks. If all limb bone fragments were analyzed and visualized here, we would expect to see an even more extreme version of the observed pattern—that is, greater evidence for defleshing.
Summary of Results
Our first research question focused on characterizing the meat-based portion of the diet at Mission Dolores. Our results show that occupants relied heavily on cattle, with their diets supplemented by caprines and cervids. Fish, reptiles, birds, and small mammals may be related to the diet, but they bore no cut marks that would allow us to confirm this, and in any case, they are so rare as to be quite marginal. We conclude that ranching was the major subsistence activity for the mission. Our second research question centered on livestock management. The higher proportion of cattle relative to caprines in our sample, and the absence of pigs or chicken, is notable compared to other Spanish colonial settlements. Mission Dolores cattle herds were likely managed primarily for beef production, given that most individuals were slaughtered at prime adult age. Finally, our third research question investigated culinary practices. Cut marks were made by metal tools and were mostly found on cattle remains. Analysis of cut-mark placement and skeletal-part representation suggests that cattle were butchered whole on-site rather than being transported in pieces. It remains unclear if this means that cattle were being raised on the site or were raised elsewhere and transported to Mission Dolores for slaughter.
Discussion
Provisioning Strategies at Mission Dolores: Comparing Archaeological and Written Records
The Mission Dolores assemblage is dominated by mammals, especially domesticates, and particularly cattle. Mission residents supplemented their largely beef-based diet by hunting deer and raising caprines. Occasionally, they appear to have consumed small game (such as rabbit or hare) and aquatic fauna, including turtle, catfish, and (as-yet-unidentified) freshwater shellfish. Although such diverse wild fauna are frequently interpreted in mission contexts as signaling the presence of Indigenous groups, written records suggest that the Ais rarely congregated at the mission. The absence of cut marks or other human-imparted marks on small-animal remains means that we cannot be certain that they were part of the mission diet and that they could alternatively have been brought to the site by dogs or cats, both of which are tentatively identified in the faunal remains.
The paucity of small animals at Mission Dolores does not appear to be an artifact of research bias—at least at the level of faunal analysis. Given our interest in wild fauna exploitation, we specifically sought out remains of small animals in each bag, and we spent considerable effort identifying those few remains present. It is likely that small animals were either uncommon at the site (or in our subsampled contexts) or were not recovered during excavation. Corbin et alia (Reference Corbin, Alex and Kalina1980:51, 123) state that unspecified “fine screen” samples were collected and that additionally, samples from features were water-screened; both kinds of screened samples produced glass beads, so we have no reason to think small-faunal remains would not be retained, although we do not know what proportion of excavated sediments were dry- or wet-screened. The vast majority of remains that we left unidentified appear to be fragmentary remains of medium and especially large (cattle-sized) mammals.
Our zooarchaeological findings generally support the documentary evidence while providing new information about mission foodways. As noted earlier, Kress et alia (Reference Kress, Fray Gaspar José and Mattie Austin1931) observed a small herd of cows, bulls, and oxen, as well as horses and mules at Mission Dolores. We identified no equid remains (they were not likely eaten), but the abundant cattle remains agree with the documentary evidence. No caprines were mentioned by de Solís, and our results suggest they were a minor part of the diet. It is possible that although Mission Dolores was founded with caprines, the herd was diminished or absent by the time de Solís evaluated the mission, or was not seen as worthy of mention. It is also possible that caprines were difficult to herd in the low-lying, humid environment. Neither pigs or chickens were identified in the fauna, nor were they mentioned in any primary sources about Mission Dolores. None of the written sources remark on hunting wild game, and this is an area where faunal analysis provides new insights.
Cattle Ranching and Herd Management at Mission Dolores
With cattle being so important at Mission Dolores, we consider how these animals were managed. To our knowledge, there is no archival or archaeological evidence directly associating any rancho with Mission Dolores, so we began with questions as to whether the mission was a site of both cattle production and consumption or if whole animals or carcass parts were being traded from other settlements.
Based on age-at-death estimates, most of the individuals represented in the assemblage were slaughtered at a prime age for beef consumption, which suggests that the herd was being managed with meat production in mind (Bowen Reference Bowen1998). A few individuals were kept alive for breeding, labor, or milking. Primary and secondary historical sources describe the subsistence struggles of Mission Dolores residents and the failure to attract congregants because of resource scarcity. However, our results suggest that the missionaries focused on beef consumption rather than broadening their diet to include diverse wild fauna, as has been documented at other missions and presidios where resource stress is hypothesized.
It is entirely possible, however, that Mission Dolores residents were struggling to provide themselves with fundamental plant-based nutrition. Records indicate that people at both Presidio los Adaes and Mission Dolores had failed to raise basic staples such as corn and beans, and they would instead trade for them with the French and the Ais or other Caddo speakers (Corbin et al. Reference Corbin, Brown, Canavan and Toups1990; Galán Reference Galán2020). We investigated the Mission Dolores faunal assemblage expecting that we might find evidence for trade of cattle as well; however, there is no skeletal-part survivorship pattern that might indicate this was the case, and future research might be able to better address this question through strontium isotope analysis or other approaches.
Other questions remain about cattle herd management practices at this site, including how and where these cattle were being grazed or foddered, and who was doing the herding at the mission. In a 1754 letter pleading for the mission to remain open amid talk of the closing of East Texas missions, Mission Dolores resident Fray Vallejo claims that the Ais had briefly congregated at the mission before and that the men would serve as laborers and herdsmen while women would perform kitchen-related labor (Nunley Reference Nunley1975). However, Fray de Solís notes in a 1768 diary that the Ais refused to congregate at Mission Dolores and that livestock at the site were targeted in Ais raids, with cattle being eaten and horses used for trade with the French (Kress et al. Reference Kress, Fray Gaspar José and Mattie Austin1931). In his 1744 report detailing his inspection of the East Texas settlements—particularly Presidio Los Adaes—Governor Winthuysen notes that there were no Indigenous residents at the Mission Dolores and that it seemed there never were (Magnaghi Reference Magnaghi1984). At the same time, archaeological work revealed material culture (mainly ceramics) typically associated with Indigenous groups (Corbin et al. Reference Corbin, Brown, Canavan and Toups1990). It remains challenging based on current evidence to assess the extent to which Indigenous peoples lived and labored at the site, and the question of ranching labor, therefore, remains unanswered.
Carcass Processing and Culinary Practices at Mission Dolores
Our cut-mark analysis supports an interpretation that cattle were slaughtered and butchered on site rather than obtained as parts through trade. Cut marks were common on cattle remains: 38% of cattle NISP exhibited at least one cut mark, and many specimens bore multiple marks. The most frequently cut-marked cattle elements were the lumbar vertebrae, innominates, and the mandible. For long-limb bones, we observed via Ikhnos that there were more cut marks on the midshafts than the epiphyses of the specimens (Figure 6; Supplementary Material 7). This indicates that people were removing the meat directly from the dead animal and were not regularly sectioning carcasses into parts for transport. It is noteworthy that there was much lower survivorship for feet than other skeletal parts. Potentially, the foot bones remained attached to hides during skinning and were processed or discarded in a different, unexcavated part of the site. However, crania and mandibles are often also discarded along with foot bones, and we did not find an unexpectedly low number of these elements. This might suggest that the head and first cervical vertebrae were removed first and discarded at the butchery site before the remainder of the carcass was butchered, and that skins (with feet attached) were then brought elsewhere for separate hide processing.
It is possible that carcasses were suspended from a wooden rack with hooks during butchery, with the majority of remaining bones discarded in this single area. This might explain the concentration of faunal remains in specific pit features, particularly Feature 8. Unfortunately, there is no documentary evidence to support this scenario, nor are there many artifacts reported that could be indicative of butchery processes. Small ferrous knife fragments are illustrated by Corbin et alia (Reference Corbin, Alex and Kalina1980, Reference Corbin, Brown, Canavan and Toups1990), but none are complete, and no hooks were depicted that could be unambiguously related to butchery. Further research might use a combination of skeletal-part representation, spatial location of faunal remains across the mission, and bone-surface modifications to better understand where and how butchery, cooking, and discard occurred.
There are limits to the extent to which we can interpret these data in terms of the social and cultural dimensions of culinary practice at Mission Dolores. As noted above, for ranching labor, it is challenging to make inferences as to who was involved in meat processing, from slaughter through butchery to cooking, serving, and discard. It is also especially challenging to assess whether the Mission Dolores fauna represents a “Spanish” pattern of butchery distinct from that of “English” patterns, as described by Gilmore (Reference Gilmore1999). We recommend that future researchers revisit this work, define patterns in terms of skeletal-part representation and cut-mark placement (including all limb shafts), and apply this approach more widely to colonial sites.
The 3D modeling and visualizing of cut-mark distribution using software such as Ikhnos, although well established in paleoanthropology, is new to historical archaeology in the United States. There are limits to our application of this method at Mission Dolores. First, our sample size, although it approximates the median NISP for Spanish colonial sites in Texas (see below), could be more robust. Second, we did not identify all limb-bone shafts, which would have enlarged our cut-marked sample. Finally, the Ikhnos program is limited to limb bones, whereas our work shows the importance of considering whole animals in carcass processing and culinary practices. Taking into account these limits, we see an expanded version of our approach as offering a new way of analyzing culinary practices. This research presents a first introduction of this powerful method to a historical collection, and there is future potential to use this method to analyze other collections to learn more about historic meat consumption, butchery practices, and cuisines.
Mission Dolores in Context: A Zooarchaeological Review of Spanish Texas
Here, we provide the first synthesis of zooarchaeological data from presidios and missions in Spanish Tejas (Supplementary Materials 1, 8, and 9). This analysis shows that domestic mammals, and especially cattle, played an exceptionally prominent role at Mission Dolores. Our review relies primarily on assemblages for which NISP data are included; we also consider three assemblages for which only MNI values are available, but we do not illustrate them given that NISP and MNI data are not directly comparable. Sample sizes vary widely, from 49 to 9,790 NISP for 17 assemblages excluding Mission Dolores (some assemblages represent different seasons at the same site). The median NISP is 458, similar to that of Mission Dolores. Many excavations occurred around the same time as those of Mission Dolores in the 1970s and 1980s, and we infer that they likely employed comparable faunal recovery and recording methods.
When comparing general taxonomic categories, Mission Dolores has the highest proportion of mammals, and especially of ungulates (Figure 7). Other Spanish colonial sites in Texas had a much greater faunal diversity, with proportionally more reliance on fish, reptiles, and birds. There are notably high numbers of fish remains at Presidio Los Adaes, Presidio San Sabá, Mission Refugio, and Mission Espíritu Santo at Goliad. In contrast, apart from a few catfish remains, these are virtually absent at Mission Dolores. Reptiles—turtles, terrapins, and tortoises in particular—are common at many sites and especially at Mission Espíritu Santo (at Mission Valley) and Mission Rosario, where softshell turtles and freshwater sliders are well represented.
Major faunal groups represented at Texas missions and presidios, using NISP (Number of Identified Specimens) and excluding remains identified as indeterminate vertebrate or mammal. We aggregated data for different seasons and features at Mission San José and at Mission Refugio. For details, see Supplementary Materials 8–9. (Color online)

Avian fauna are well represented at many sites in our review, but they are nearly absent at Mission Dolores. There are exceptionally diverse wild avifauna—mainly waterfowl—at Mission Refugio, which is unsurprising given its proximity to Mission Bay near the Gulf of Mexico. Presidio Los Adaes also has abundant waterfowl. Many sites, however, have mainly ground-dwelling fowl, such as domestic chicken and either wild or domestic turkeys. Chicken or turkey are especially well documented at Mission Refugio, Presidio Los Adaes, and Presidio San Sabá, but they are entirely absent at Mission Dolores.
Rodents are rarely reported at sites in our review, and they are absent at Mission Dolores. One exception is Mission Concepción. The analysts interpreted this as resulting from owl predation (Scurlock and Fox Reference Scurlock and Fox1977). It is significant that this is the only site in our review to report Rattus rattus—a nonnative species introduced from Europe that authors suggest was transported to Texas via grain from Mexico. R. rattus is documented on a seventeenth-century French ship that sank in Matagorda Bay but is otherwise not documented in early Texas (Guiry et al. Reference Guiry, Kennedy, Orton, Armitage, Bratten, Dagneau and Dawdy2024).
For sites where only MNI was provided, the relative abundance of ungulates is generally lower: approximately 60% of MNI for Mission San Juan and two Mission Rosario assemblages. Mission San Juan has an exceptionally high number of rodents (15% of MNI). Nonmammals are relatively abundant in these assemblages, with fish, reptiles, and birds jointly comprising 13% of MNI at Mission San Juan and 32% of MNI in the Mission Rosario assemblages. However, MNI tends to (over)emphasize the importance of taxa represented by few specimens.
When examining only mammals in the comparative dataset, Mission Dolores has greater relative abundance of cattle remains than any other site in our review, and the greatest relative abundance of domestic mammal remains overall (Figure 8). Mission Dolores is the most extreme case of a pattern identified at the majority of the reviewed sites: reliance on domesticates and especially bovines. Given that bovines were identified at differing levels of precision, we treated all remains identified as “cattle” as one group, and all those identified as “cow/bison,” “bovine,” or “bison” as another group, noting that remains identified specifically as bison are exceedingly rare across all sites in our review. Taking these two groups together, bovines comprise greater than three-quarters of mammalian NISP (excluding indeterminate mammals) at Missions Dolores, Refugio, Rosario, Espíritu Santo at Mission Valley, and Espíritu Santo at Goliad, as well as at Presidio Los Adaes. For sites where only MNI was available, the pattern is similar, with bovines forming 50% of MNI at Mission San Juan and approximately 60% of MNI at Mission Rosario. The only sites that do not follow this pattern are Missions San José and Concepción, where cattle or combined bovine remains are less than half of mammalian NISP, and where caprines and pigs were relatively important; and Presidio San Sabá, which has an unusually high frequency of caprines (71% of mammalian NISP). This presidio is located on the Edwards Plateau, which has a cooler, hilly environment advantageous for raising caprines (Walter Reference Walter2021).
Major mammalian groups represented at Texas missions and presidios, using NISP (Number of Identified Specimens), where blue tones are domesticated and orange tones are wild. Here, we excluded rodents, and remains identified as indeterminate mammal, indeterminate ungulate, and carnivore remains that similarly could not be defined as wild or domestic (e.g., those identified as Canis sp.). We grouped remains identified as “cow/bison,” “bovine,” or “bison” into a “cattle or bison” group while keeping those specified as cattle separate. “Other (wild)” includes peccary, wild carnivores (e.g., raccoon, fox, skunk), opossum, and armadillo. “Other (domestic)” includes equids, dogs, and cats. We aggregated data for different seasons and features at Mission San José and at Mission Refugio. For details, see Supplementary Materials 8–9. (Color online)

Wild mammals (excluding rodents) never comprise more than 20% of mammalian NISP in this analysis. At nearly all sites, most wild mammal remains are those of deer, with rabbit or hare also being relatively important at Missions Refugio, San José, and Concepción and at Presidio San Sabá. For sites where only MNI data were available, wild game appears relatively more abundant—21%–34% of MNI at Missions San Juan and Rosario; however, we urge caution given that MNI values may inflate the importance of animals represented by isolated specimens. We also note that the “other wild” category here includes several small carnivores that were unlikely food sources—such as skunk and raccoon—and it would be challenging to discern food taxa from background taxa in the absence of clear signs of human intervention; similarly, “other domestic” includes unlikely food animals, such as cat, dog, and horse.
We began our analysis aiming to compare Presidio Los Adaes and Mission Dolores. The unexpected contrasts between these assemblages are therefore worth further exploration, particularly given the sites’ geographic proximity and similar settings in low-lying ridges or hills with a humid climate. Zooarchaeological evidence from Presidio Los Adaes indicates that although cattle were important along with domesticates such as caprines, pig, chicken, and either wild or domestic turkey, the occupants also relied on a strikingly diverse range of wild fauna. This contrast is most evident in Figure 7 (see also Supplementary Materials 8 and 9), which shows that Adaesaños were exploiting fish (especially catfish) and turtles, and occasionally consuming fauna such as alligator (which would have been especially unfamiliar to Spanish palates). Among mammals, the Adaesaños exploited a moderately greater number of deer and rabbits or hares. The diverse fauna at Los Adaes was interpreted by Pavão-Zuckerman and Loren (Reference Pavão-Zuckerman and Loren2012) as evidence that even elite residents of the presidio—including the governor—were struggling to put food on the table that aligned with their background. Instead, they resorted to eating catfish and alligator, even as tablewares suggested that they maintained norms of presentation. Based on documentary evidence of the subsistence struggles at Mission Dolores, we expected to see a similar reliance on culturally unfamiliar foods. To our surprise, Mission Dolores residents instead consumed beef from prime-age cattle, suggesting more food security—at least in terms of the animal-based part of the diet—than is indicated in written records.
Conclusions and Future Directions
This research represents the first in-depth zooarchaeological analyses of an East Texas Spanish mission, which shows that cattle ranching, supplemented by sheep/goat herding and deer hunting, was the central subsistence activity at the short-lived Mission Dolores. In contrast with the neighboring and environmentally similar Presidio Los Adaes, the Mission Dolores residents did not hunt, fish, or capture a wide range of wild fauna, nor did they typically consume animals foreign to the Spanish palate. They also slaughtered their cattle at a prime age for meat consumption, suggesting that the herd was not under stress. These findings contrast with documentary evidence describing the struggles of Mission Dolores residents to feed themselves; instead, they suggest a successful cattle-based subsistence strategy. We also found no evidence to suggest that cattle were being obtained through trade, despite written accounts attesting to trade in agricultural products. Although Mission Dolores is located in a borderlands zone with abundant opportunities for French, Spanish, Indigenous, and African cultural contact to forge new food traditions, we did not find evidence of this in the faunal assemblage; instead, we saw evidence of adherence to a European diet. This new evidence is important for learning more about the effects of Spanish colonization on the cultural history and ecology of this region and its people and for providing some of the earliest evidence of the ranching practices central to Texas heritage today.
This article also represents an initial synthesis of faunal data for Spanish settlements in Texas, which were mostly hidden in gray literature. In comparing faunal records at Mission Dolores to other sites, we demonstrate that although Mission Dolores falls within a pattern of cattle-dominated ranching at Texas missions and presidios, it is an extreme version of this pattern, with fewer nonmammalian fauna and fewer nonbovine domesticates than any other site in our review. A more in-depth comparative analysis of Texas faunal assemblages is beyond the scope of this article, but it would be useful in the future for examining variation in both herd management and culinary practices across Texas during Spanish occupation. Such comparison could incorporate the few Spanish sites in Texas that are neither missions nor presidios (Taylor and Fox Reference Taylor and Fox1985; Tennis Reference Tennis2001) and could integrate the understudied zooarchaeology of Texas sites with the abundant and diverse datasets from across New Spain’s northern borderlands.
Although traditional zooarchaeological analysis provides a robust starting point for understanding Spanish colonial foodways in Texas, revisiting these legacy collections using additional methods could reveal a more detailed picture of the cattle-ranching activities and herd-management strategies at Mission Dolores and at the other Spanish Tejas sites reviewed in this article. Osteometry could offer insights into the average size (and therefore meat yields) of animals when slaughtered. If osteometric approaches were taken across multiple Texas assemblages to increase sample size and spatial and chronological transects, they could potentially shed light on the breeds utilized by the Spanish and the origins of North American breeds such as the Texas longhorn, which originate from feral populations of Spanish cattle (McTavish et al. Reference McTavish, Decker, Schnabel, Taylor and Hillis2013).
Ancient DNA analysis of cattle remains could also contribute to reconstructing population history. Many heirloom Spanish breeds were largely lost during the nineteenth century (Specht Reference Specht2016), and using present-day cattle genetic data alone will be limited in terms of capturing past diversity. In other colonial contexts in the Americas, ancient DNA is revealing diverse origins and pathways for cattle, including previously underappreciated contributions of African cattle breeds to populations introduced by the Spanish (Delsol et al. Reference Delsol, Stucky, Oswald, Cobb, Emery and Guralnick2023). Such work might also reveal biological adaptations to new environments (Ramírez-Ayala et al. Reference Ramírez-Ayala, Rocha, Ramos-Onsins, Leno-Colorado, Charles, Bouchez, Rodríguez-Valera, Pérez-Enciso and Ramayo-Caldas2021). This would be particularly relevant in Texas, where longhorns are especially adapted to drought.
Stable isotope analysis could also shed light on herd mobility and management strategies, and help address questions about not only where and how animals were being grazed but whether large and small stock were managed together or separately. Such analyses have been limited—but informative—in Spanish and French colonial contexts in North America, where they have revealed faunal procurement strategies (Colclasure et al. Reference Colclasure, Andrus and Blair2025) and livestock management practices (Freiwald and Pugh Reference Freiwald and Pugh2018; Guiry et al. Reference Guiry, Jones, deFrance, Bruseth, Durst and Richards2018; Mathwich Reference Mathwich2022).
This investigation of Mission Dolores foodways contributes to increasingly complex, archaeologically derived narratives about Spanish colonialism and the extent to which missions and presidios were sites of cultural exchange and innovation. Whereas foodways across the northern borderlands of New Spain are relatively well documented in the southeastern and southwestern United States, the vast area comprising Spanish Tejas remains poorly known. Our review suggests that in contrast with documentary evidence stressing the risks and failures of Texas missions and presidios, most had thriving cattle-based subsistence systems. There is great potential for future research in this area to further elucidate foodways and culinary practices in Spanish Texas and how this influenced the historical development of ranching and new cuisines.
Acknowledgments
We thank the Friends of the Texas Historical Commission and Rice University Department of Anthropology for their support. We gratefully acknowledge Manuel Domínguez-Rodrigo for guidance on bone surface modifications, statistical analyses, and visualization in Ikhnos.
Funding Statement
This project was initiated as part of the Texas Historical Commission’s Mission Dolores Archeological Collections Rehabilitation Project, funded by the National Parks Service Save America’s Treasures program, via a grant to the Friends of the Texas Historical Commission (Award ST-251087-OMS-21).
Data Availability Statement
All data used in this article are provided in the online supplementary information. A copy of the original faunal database has been deposited with the Texas Historical Commission and is available by requesting it from that organization.
Competing Interests
The authors declare no competing interests.
Supplementary Material
The supplementary material for this article can be found at https://doi.org/10.1017/aaq.2025.10160.
Supplementary Material 1. Archaeofaunal assemblages used for comparison to Mission Dolores in this study (table).
Supplementary Material 2. Examples of bone surface modifications observed at Mission Dolores (figure).
Supplementary Material 3. Differentiation of Bos/Bison at Mission Dolores, following Balkwill and Cumbaa (Reference Balkwill and Cumbaa1992) (table).
Supplementary Material 4. Bone surface modifications observed at Mission Dolores (table).
Supplementary Material 5. Cut marks on cattle remains at Mission Dolores (figure).
Supplementary Material 6. Cut marks observed on cattle long-limb bones at Mission Dolores and used in Ikhnos spatial analysis (table).
Supplementary Material 7. 3D model diagrams of cut-mark distribution on cattle specimens (figure).
Supplementary Material 8. Taxonomic representation at Mission Dolores compared with regional mission and presidio faunal assemblages (table).
Supplementary Material 9. Taxonomic attributions of faunal remains from Texas mission and presidio sites (table).


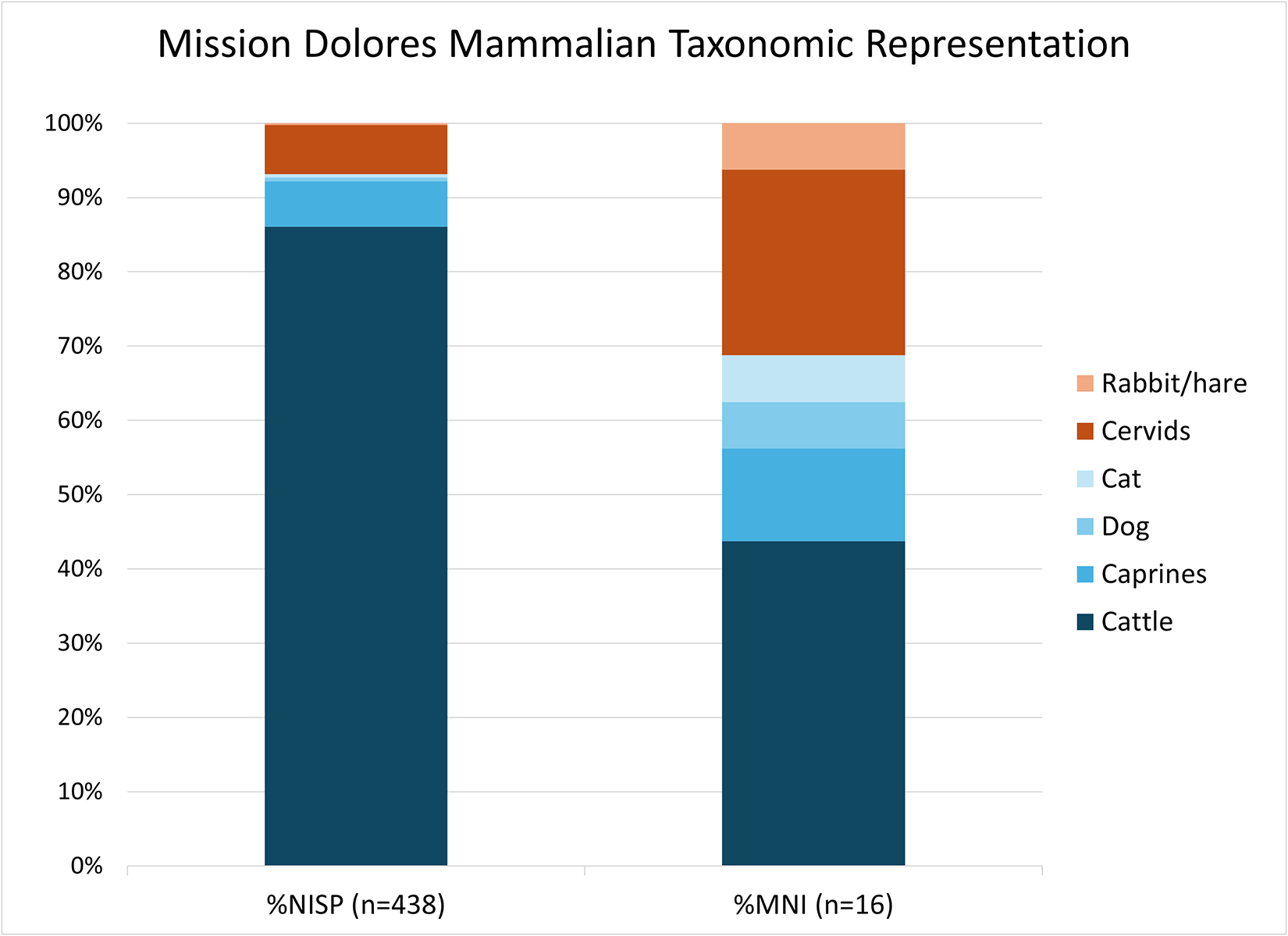



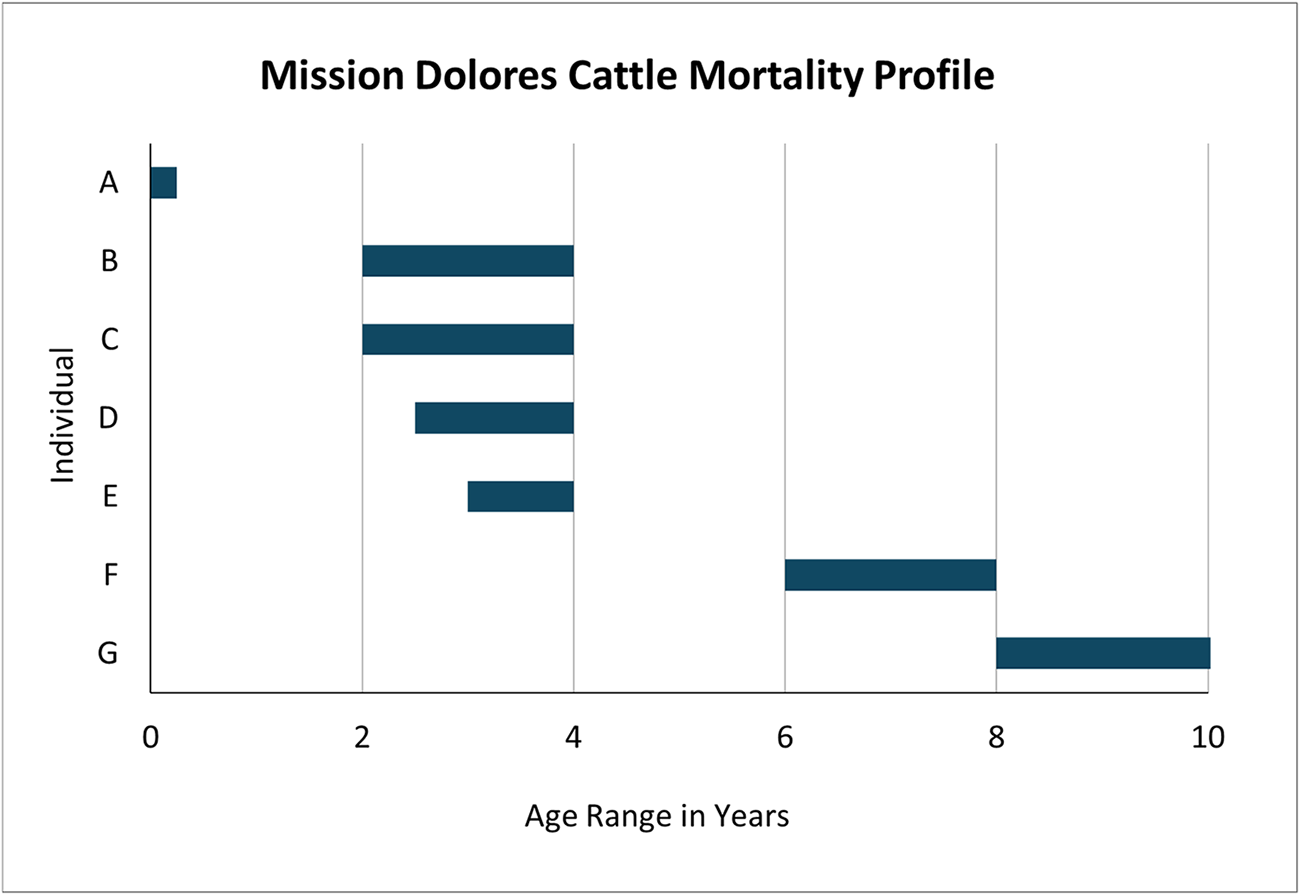







 Open access
Open access