Introduction
The Mediterranean Sea is among the regions most heavily affected by Non-Indigenous Species (NIS) worldwide (Costello et al., Reference Costello, Dekeyzer, Galil, Hutchings, Katsanevakis, Pagad, Robinson, Turon, Vandepitte, Vanhoorne, Verfaille, Willan and Rius2021). More than 1,000 NIS have been recorded to date, with approximately half likely introduced through the Suez Canal, as the primary pathway (Galanidi et al., Reference Galanidi, Aissi, Ali, Bakalem, Bariche, Bartolo, Bazairi, Beqiraj, Bilecenoglu, Bitar and Zenetos2023). Many of these Canal-introduced species – referred to as Lessepsian species – have established thriving populations in the Levant Basin, while others have expanded into the central and western Mediterranean (Galil et al., Reference Galil, Boero, Campbell, Carlton, Cook, Fraschetti, Gollasch, Hewitt, Jelmert, Macpherson and Ruiz2015; Öztürk, Reference Öztürk2021). Once established, some NIS can rapidly spread and become invasive alien species (IAS), exerting significant ecological, economic, and human-health impacts (Pyšek et al., Reference Pyšek, Hulme, Simberloff, Bacher, Blackburn, Carlton, Dawson, Essl, Foxcroft, Genovesi and Richardson2020). Notable examples include the herbivorous rabbitfishes Siganus luridus and Siganus rivulatus, which have transformed formerly diverse algal assemblages into extensive ‘barrens’ in the Levantine infralittoral zone, reducing habitat complexity, biodiversity, and biomass (Sala et al., Reference Sala, Kizilkaya, Yildirim and Ballesteros2011). Similarly, the toxic silver-cheeked toadfish Lagocephalus sceleratus is considered one of the most harmful IAS in the region due to its rapid spread, high abundance, and wide-ranging impacts (Streftaris and Zenetos, Reference Streftaris and Zenetos2006; Galil, Reference Galil, Mazza and Tricarico2018). The proliferation of Lessepsian species has been associated with marked declines in native fauna, including local extirpations. For instance, the Indo-Pacific goldband goatfish Upeneus moluccensis has replaced the native red mullet Mullus barbatus in Levantine fisheries (Ulman et al., Reference Ulman, Abd Rabou, Al Mabruk, Bariche, Bilecenŏglu, Demirel, Galil, Hüseyinoğlu, Jimenez and Hadjioannou2024). Interactions and potential competition with native mullids may also involve other Lessepsian goatfishes, such as Por’s goatfish Upeneus pori and the Red Sea goatfish Parupeneus forsskali (Fourmanoir & Guézé, 1976) (Otero et al., Reference Otero, Cebrian, Francour, Galil and Savini2013; Evagelopoulos et al., Reference Evagelopoulos, Nikolaou, Michailidis, Kampouris and Batjakas2020). The latter species is native to the Red Sea and Gulf of Aden, where it is the dominant mullid in the northern Red Sea (Ben-Tuvia and Kissil, Reference Ben-Tuvia and Kissil1988; Sabrah, Reference Sabrah2015).
Parupeneus forsskali (Fourmanoir & Guézé, 1976) was first recorded in the Mediterranean Sea in Türkiye in 2004 (Çinar et al., Reference Çinar, Bilecenoglu, Öztürk and Can2006) and has since been reported throughout the Levant and southern Mediterranean (Bariche et al., Reference Bariche, Bilecenoglu and Azzurro2013, Reference Bariche, Al-Mabruk, Ateş, Büyük, Crocetta, Dritsas, Edde, Fortič, Gavriil, Gerovasileiou, Gökoğlu, Huseyinoglu, Karachle, Kleitou, Terbiyik Kurt, Langeneck, Lardicci, Lipej, Pavloudi, Pinna, Rizgalla, Rüştü Özen, Sedano, Taşkin, Yildiz and Zangaro2020; Sonin et al., Reference Sonin, Salameh, Edelist and Golani2013; Ali et al., Reference Ali, Diatta, Alkusairy, Saad and Capapé2016; Chartosia and Michailidis, Reference Chartosia and Michailidis2016; Mehanna et al., Reference Mehanna, Mahmoud and Hassanien2016), reaching Tunisian waters in the central basin in 2016 (Capapé et al., Reference Capapé, Zaouali, Amor and Amor2018). Here, we document the presence of P. forsskali in the waters of Lampedusa Island, representing the first confirmed record of the species in Italian waters, supported by both morphological and molecular evidence. We further reconstruct the relationships of the present record within its conspecific and within the genus Parupeneus based on publicly available COI barcode sequences. Finally, we highlight the essential role of fishers in the detection and collection of NIS, underscoring their contribution to early monitoring efforts.
Materials and methods
On 19 November 2024, a goatfish displaying unusual coloration was captured by trawl approximately 4 nautical miles south of Lampedusa Island at a depth of 60 m. The specimen was preserved frozen by the fisherman and subsequently delivered to the ISPRA Marine Ecology Laboratory (Palermo) for identification. In the laboratory, the specimen was photographed after thawing (Figure 1a), and morphometric and meristic characters were recorded following Ben-Tuvia and Kissil (Reference Ben-Tuvia and Kissil1988) and Randall and Heemstra (Reference Randall and Heemstra2009). Measurements were taken to the nearest 1 mm using an ichthyometer or to the nearest 0.1 mm using a caliper, and the specimen was weighed. A muscle tissue sample was collected and preserved in 90% ethanol for molecular analyses. The specimen was deposited in the ichthyological collection of the Museum of Zoology ‘P. Doderlein’ of Palermo under accession code PL 392. One year later, seven additional specimens (Figure 1b–g) were captured in the same area and photographed by local fishers, who forwarded the images to ISPRA researchers for identification. All individuals were caught together with species of the genus Mullus. Details of all captures are provided in Table 1.
Specimens of Parupeneus forsskali caught near Lampedusa Island on: (A) 19 November 2024; (B) 2 November 2025; (C) 16 November 2025; (D-E) 21 November 2025; (F) 21 December 2025; (G) 22 December 2025.

Figure 1 Long description
Seven close-up photographs show goatfish specimens in side view. A) One goatfish lies on a measuring board with evenly spaced vertical lines and numbered markings. B) One goatfish lies on a flat surface. A scale bar reads 2 cm. C) One goatfish lies on a flat surface. D) One goatfish lies on a flat surface. E) Two goatfish lie side by side on a flat surface, one larger and one smaller. F) One goatfish lies on a slotted crate or grate surface. G) Two goatfish lie on netting with a mesh pattern.
Records of Parupeneus forsskali from Lampedusa Island, including date of capture, number of specimens (N), total length (TL), fishing zone, geographical coordinates, bottom depth (m), fishing gear, and corresponding image in Figure 1

Table 1 Long description
The table lists seven fishing events with date, number of individuals, total length when recorded, fishing zone, coordinates, depth, gear type, and the related figure panel. Five events are offshore and all use trawl gear at depths from 60 to 75 m, each reporting one individual. Two events are in coastal waters and use gill nets at shallower depths, one at 7 to 8 m with one individual and one at 12 to 15 m with two individuals. Total length is reported only for the first two offshore records, at 187 mm on 19 November 2024 and 210 mm on 2 November 2025; all later entries have length missing. Offshore coordinates cluster around 35.38 to 35.47 degrees north and 12.60 to 12.97 degrees east, while coastal coordinates are near 35.50 degrees north and 12.63 degrees east. Because most length values are absent and sample sizes per event are very small, comparisons of size or abundance across dates and zones are limited.
Molecular analysis
Genomic DNA was extracted using the PureLink Genomic DNA Kit (Invitrogen). PCR amplification of the COI barcode region was performed using 50 ng of purified DNA, the FishF1/FishR1 primer pair (Ward et al., Reference Ward, Zemlak, Innes, Last and Hebert2005) at 1 µM, 1 × reaction buffer, 0.2 mM dNTPs (Euroclone), BSA (0.5 µg/µl; New England BioLabs), and Platinum Taq DNA Polymerase (Thermo Fisher Scientific Inc.) in a final volume of 25 µl. PCR cycling conditions consisted of an initial denaturation at 94 °C for 2 min, followed by 30 cycles of 94 °C for 30 s, 50 °C for 30 s, and 72 °C for 45 s, with a final extension at 72 °C for 7 min. Amplification products were visualized on agarose gel, purified, and sequenced using an ABI Prism 373 automated sequencer (Macrogen Europe). Sequence identification was performed using BLASTn (https://www.ncbi.nlm.nih.gov). The obtained sequence was compared with Parupeneus spp. sequences available in GenBank (https://www.ncbi.nlm.nih.gov/genbank/) (Supplementary Table S1). Pairwise genetic distance among Parupeneus species was estimated using the Kimura 2-parameter model (Kimura, Reference Kimura1980). Their relationships were reconstructed using the Maximum Likelihood (ML) method, and Kimura 2-parameter model of nucleotide substitutions to construct a tree with the highest log likelihood as implemented in MEGA 6.0 (Tamura et al., Reference Tamura, Stecher, Peterson, Filipski and Kumar2013).
Results
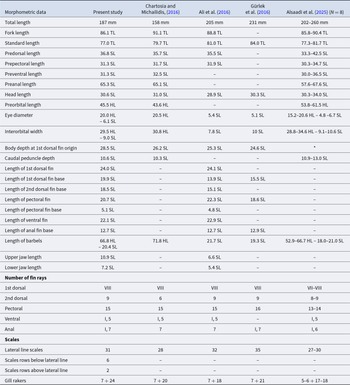
The first specimen exhibited an elongated body with large scales and two long chin barbels (Figure 1a). Each jaw bore a row of well-spaced teeth, whereas teeth were absent on the vomer and palatines – diagnostic features of the genus Parupeneus (Uiblein, Reference Uiblein2021). The coloration of the fresh specimen was reddish dorsally, with a longitudinal dark stripe extending to the posterior end of the second dorsal-fin base and a distinct black spot on the upper caudal peduncle. The flanks were pinkish and marbled with yellow, including the head, with scattered reddish spots, while the ventral surface was whitish. The first dorsal, pectoral, and pelvic fins were pinkish; the second dorsal fin displayed three yellow longitudinal stripes; and the anal and caudal fins were yellowish. This colour pattern closely matched descriptions of P. forsskali previously reported from the Mediterranean Sea (Randall and Myers, Reference Randall and Myers2002; Bariche et al., Reference Bariche, Bilecenoglu and Azzurro2013; Sonin et al., Reference Sonin, Salameh, Edelist and Golani2013; Gürlek et al., Reference Gürlek, Gündüz, Uyan, Doğdu, Karan, Gürlek, Ergüden and Turan2016). The specimen measured 187 mm total length (TL) and weighed 77.7 g. Morphometric and meristic characters were consistent with published accounts of Lessepsian P. forsskali (Table 2) (Ali et al., Reference Ali, Diatta, Alkusairy, Saad and Capapé2016; Chartosia and Michailidis, Reference Chartosia and Michailidis2016; Gürlek et al., Reference Gürlek, Gündüz, Uyan, Doğdu, Karan, Gürlek, Ergüden and Turan2016; Alsaadi et al., Reference Alsaadi, Elbaraasi, Shoeib, Alkklosh, Almabrouk, Elsalini, Tiralongo and Azzurro2025). The seven additional specimens photographed by local fishers exhibited the same diagnostic colour pattern and were likewise identified as P. forsskali (Figure 1b–g).
Morphometric and meristic characters of Parupeneus forsskali from Lampedusa Island (Italian waters), compared with published data for other Mediterranean specimens. Measurements are expressed as percentages of total length (TL), standard length (SL) and/or head length (HL). *The Authors reported Maximum body depth, resulting in 25.0-30.3 %SL, instead of Body depth at 1st dorsal fin origin

Table 2 Long description
The table compares morphometric measurements and meristic counts for the same species across five sources, reporting total length in millimeters and many other dimensions as proportions of total length, standard length, or head length. Total length differs most among studies, ranging from 158 mm (Chartosia and Michailidis, 2016) to 231 mm (Gürlek et al., 2016), with Alsaadi et al., 2025 reporting 202 to 260 mm and the present study at 187 mm. Fork length and standard length are broadly consistent as proportions of total length, with fork length around the mid to high 80s percent and standard length around the high 70s to mid 80s percent where reported. Several pre-length measures (predorsal, prepectoral, preventral, preanal) cluster in similar ranges across studies but include missing entries in some columns. Head length is close across sources at roughly three tenths of standard length, while eye diameter and interorbital width are given either as head-length proportions, standard-length proportions, or both, and generally fall within the ranges reported by Alsaadi et al., 2025. Fin measurements (for example dorsal, pectoral, ventral, and anal fin bases) are provided for some studies and absent for others, limiting direct comparisons for those traits. Meristic data show small differences: first dorsal fin rays are typically eight, second dorsal rays range from six to nine, pectoral rays are mostly fifteen to sixteen but Alsaadi et al., 2025 reports thirteen to fourteen, and anal rays are usually seven with one study reporting six. Scale counts vary modestly, with lateral line scales spanning 27 to 35, and gill rakers differ across studies, with the present study reporting 7 plus 24 and other sources reporting lower totals. Because several cells are missing and some measures are expressed using different reference lengths, comparisons should be interpreted as approximate rather than exact.
Molecular analysis
The COI barcode region was successfully amplified (609 bp before trimming; GenBank accession number PX619320). BLAST analysis showed a 98.9% identity match with P. forsskali from the Red Sea (Suez Gulf; accession numbers LC543901, LC543902, and LC543903; sequence query cover of 100%), and 99.0–99.2% identity with other P. forsskali sequences from the Red Sea and Mediterranean (MH331820, KR861545, MN560944, and MF123975, with a lower sequence query cover 95–98%). Alignment with GenBank sequences yielded 570 bp, of which 231 were polymorphic and 199 informative. The relationships among Parupeneus species assessed by using ML method (Figure 2) recovered a Red Sea/Mediterranean clade that grouped with P. margaritus and was clearly separated from P. forsskali specimens originating from the Indian Ocean (FJ347965 from India) and South China Sea (MK777803 from Vietnam). Conversely, Indian Ocean and South China Sea P. forsskali clustered with the P. fraserorum clade. Kimura 2-parameter distances among Parupeneus species ranged from 0.3% (P. crassilabris vs P. insularis) to 25.5% (P. barberinoides vs P. fraserorum; Supplementary Table S2).
COI-based phylogenetic tree of the genus Parupeneus constructed with the Maximum Likelihood method and Kimura (1980) 2-parameter model of nucleotide substitutions. Tree rooted at its mid-point. The specimen from Lampedusa Island (GenBank accession Number PX619320) is bordered in red as the two clades of Parupeneus forsskali.

Figure 2 Long description
The diagram is a vertical phylogenetic tree illustrating the evolutionary relationships among Parupeneus species using the Maximum Likelihood method. At the top, the tree begins with several branches leading to species such as Parupeneus crassilabris and Parupeneus insularis, with GenBank accession numbers like MN560944 and MF123975. As the tree progresses downward, branches split and merge, showing connections between species like Parupeneus macronemus and Parupeneus indicus. Midway, the tree features a clade grouping Parupeneus margaritus with sequences from the Red Sea and Mediterranean, clearly separated from Parupeneus forsskali specimens from the Indian Ocean and South China Sea. This separation is highlighted with red boxes around the labels for P. forsskali from the Red Sea and Mediterranean and P. forsskali from the Indian Ocean and South China Sea. Further down, the tree includes species such as Parupeneus fraserorum, with branches labeled with accession numbers like FJ347965 and MK777803. The tree concludes with species like Parupeneus barberinoides, showing the diverse evolutionary paths within the genus.
Discussion
Marine biological invasions are among the most significant drivers of ecological change in the Mediterranean Sea (Tsirintanis et al., Reference Tsirintanis, Azzurro, Crocetta, Dimiza, Froglia, Gerovasileiou and Katsanevakis2022). Accelerating climate change, rising sea temperatures, increased maritime traffic, and aquaculture activities, together with the continued influx of Lessepsian migrants through the Suez Canal, favour the establishment and spread of non-indigenous species (Öztürk, Reference Öztürk2021; Vagenas et al., Reference Vagenas, Karachle, Oikonomou, Stoumboudi and Zenetos2024). The faunistic identity of the Mediterranean basin is undergoing a gradual transformation as species enter through multiple pathways, resulting in a progressive ‘demediterraneization’ of its biota (Azzurro et al., Reference Azzurro, Smeraldo and D’Amen2022). Warming trends have progressively increased the suitability of Mediterranean habitats for thermophilic fauna, thereby facilitating the eastward expansion of Atlantic species as well as the westward advance of numerous Red Sea and Indo-Pacific species (Albano et al., Reference Albano, Schultz, Wessely, Taviani, Dullinger and Danise2024). In this context, the Strait of Sicily – strategically located at the centre of the Mediterranean basin – represents a crossroads for major dispersal routes and a key observation point for monitoring the range expansion of species entering the Mediterranean through the Strait of Gibraltar, as well as of Lessepsian species (Azzurro et al., Reference Azzurro, Ben Soussi, Boughedir, Castriota, Deidun, Falautano, Ghanem, Zammit-Mangion and Andaloro2014; Badalucco et al., Reference Badalucco, Tiralongo, Crocetta and Lo Brutto2024). Among the latter is the recent detection of the Red Sea goatfish P. forsskali in the Strait of Sicily. The specimen collected from Lampedusa Island displayed morphometric and meristic characters that fall within the range reported for P. forsskali in Cyprus, Syria, Türkiye, and Libya (Ali et al., Reference Ali, Diatta, Alkusairy, Saad and Capapé2016; Chartosia and Michailidis, Reference Chartosia and Michailidis2016; Gürlek et al., Reference Gürlek, Gündüz, Uyan, Doğdu, Karan, Gürlek, Ergüden and Turan2016; Alsaadi et al., Reference Alsaadi, Elbaraasi, Shoeib, Alkklosh, Almabrouk, Elsalini, Tiralongo and Azzurro2025).
Molecular analysis confirmed its identity as P. forsskali, representing the first verified record of the Red Sea goatfish in Italian waters. Furthermore, the Italian specimen clusters tightly with Red Sea and Mediterranean P. forsskali, forming a distinct lineage in the ML tree. The presence of a distinct haplotype in the Mediterranean Sea may suggest multiple, potentially ongoing, incursions from the Red Sea. This Red Sea/Mediterranean lineage is clearly separated from the conspecifics of the Indian Ocean and the South China Sea (Indo-Pacific) and is positioned close to the P. margaritus clade. The high mean genetic distance (20%) between Red Sea/Mediterranean P. forsskali and putative conspecifics from the Indian Ocean and South China Sea is comparable to interspecific distances within the genus, raising the possibility of misidentified sequences in GenBank. Such errors have occurred in the past, particularly among sympatric species that are difficult to distinguish using morphology-based identification keys, and because many species lack species-specific diagnostic markers that can be verified in museum-preserved specimens, which are essential for accurate identification (Randall and Heemstra, Reference Randall and Heemstra2009). Alternatively, the observed pattern may reflect genuine evolutionary divergence between the Red Sea and the Indo-Pacific. The Red Sea is a semi-enclosed basin with high endemism, harbouring 14.7% of the world’s endemic marine fishes (Bogorodsky and Randall, Reference Bogorodsky, Randall, Rasul and Stewart2019), and has been proposed as an ‘evolutionary incubator’ generating distinct genetic lineages. Restricted species dispersal through the Bab al-Mandab Strait (connecting the Gulf of Aden with the Red Sea) and historical fluctuations in salinity and sea level may have contributed to the partial isolation of the Red Sea from the Indo-Pacific populations (Di Battista et al., Reference Di Battista, Choat, Gaither, Hobbs, Lozano-Cortés, Myers, Paulay, Rocha, Toonen, Westneat and Berumen2016). Furthermore, the low genetic distance between Mediterranean and Red Sea P. forsskali supports the hypothesis of ongoing gene flow between the two basins. Broader sampling and additional molecular markers will be necessary to clarify population structure and evolutionary relationships. The difficulty in resolving interspecific boundaries within Parupeneus is consistent with previous studies reporting comb-like phylogenetic branching patterns (Kim, Reference Kim2002) and suggesting rapid diversification within the genus, requiring additional investigations (Longo et al., Reference Longo, Faircloth, Meyer, Westneat, Alfaro and Wainwright2017; Nash et al., Reference Nash, Lungstrom, Hughes and Westneat2022). Parupeneus forsskali is native to the Red Sea and Gulf of Aden, where it is abundant in shallow waters (<30 m) (Ben-Tuvia and Kissil, Reference Ben-Tuvia and Kissil1988). Its introduction into the Mediterranean Sea likely occurred via the Suez Canal, as documented for other Eritrean mullids such as Upeneus moluccensis and U. pori. Since its first Mediterranean record in Türkiye in 2004 (Çinar et al., Reference Çinar, Bilecenoglu, Öztürk and Can2006), the species has expanded throughout the Levantine Basin, including Lebanon, Israel, Cyprus, Syria, and Egypt (Bariche et al., Reference Bariche, Bilecenoglu and Azzurro2013; Sonin et al., Reference Sonin, Salameh, Edelist and Golani2013; Ali et al., Reference Ali, Diatta, Alkusairy, Saad and Capapé2016; Chartosia and Michailidis, Reference Chartosia and Michailidis2016; Mehanna et al., Reference Mehanna, Mahmoud and Hassanien2016). It is now established in Cyprus, where it supports a commercial fishery, and has recently been reported from the Dodecanese Islands and Greece (Evagelopoulos et al., Reference Evagelopoulos, Nikolaou, Michailidis, Kampouris and Batjakas2020). Its presence along the Turkish coast suggests a well-established population in the eastern Mediterranean (Gürlek et al., Reference Gürlek, Gündüz, Uyan, Doğdu, Karan, Gürlek, Ergüden and Turan2016). The westward expansion of the species was reported in Libyan (Bariche et al., Reference Bariche, Al-Mabruk, Ateş, Büyük, Crocetta, Dritsas, Edde, Fortič, Gavriil, Gerovasileiou, Gökoğlu, Huseyinoglu, Karachle, Kleitou, Terbiyik Kurt, Langeneck, Lardicci, Lipej, Pavloudi, Pinna, Rizgalla, Rüştü Özen, Sedano, Taşkin, Yildiz and Zangaro2020; Alsaadi et al., Reference Alsaadi, Elbaraasi, Shoeib, Alkklosh, Almabrouk, Elsalini, Tiralongo and Azzurro2025) and Tunisian waters (Capapé et al., Reference Capapé, Zaouali, Amor and Amor2018), with the latter being the westernmost confirmed record at that time. Other central Mediterranean records remain uncertain due to a lack of documentation (Sciberras and Schembri, Reference Sciberras and Schembri2007; Perzia et al., Reference Perzia, Cillari, Crociata, Deidun, Falautano, Franzitta, Galdies, Maggio, Vivona and Castriota2023). The first concerns a record from Gozo (Maltese Islands), described by Cilia (Reference Cilia1979) as Pseudupeneus barberinus (Lacepède, 1801) (= Parupeneus barberinus). According to Golani et al. (Reference Golani, Orsi-Relini, Massuti and Quignard2004), P. barberinus should be excluded from the Mediterranean list of exotic fishes. The markings of P. barberinus and P. forsskali exhibit a high degree of similarity, raising the possibility that the specimen reported by Cilia (Reference Cilia1979) as P. barberinus may have been misidentified and could, in fact, correspond to P. forsskali. However, in the absence of further information, this record cannot be reliably assigned to the species and has therefore been listed as Parupeneus sp. (Sciberras and Schembri, Reference Sciberras and Schembri2007). Additional records of P. forsskali reported in 2017 and 2018 from Maltese waters during citizen science activities cannot be validated, as no photos or videos are available (Perzia et al., Reference Perzia, Cillari, Crociata, Deidun, Falautano, Franzitta, Galdies, Maggio, Vivona and Castriota2023). In its native range, P. forsskali inhabits coral reefs, sandy bottoms adjacent to reefs, and artificial structures (Hobson, Reference Hobson1974; Ben-Tuvia and Kissil, Reference Ben-Tuvia and Kissil1988; Golani and Diamant, Reference Golani and Diamant1999), while juveniles occur along sandy shores (Golani, Reference Golani1993). In the eastern Mediterranean, the species has been observed on rocky and soft bottoms, often near seagrass meadows (Posidonia oceanica and Cymodocea nodosa), typically shallower than 40 m, although occurrences down to 110 m have been reported (Evagelopoulos et al., Reference Evagelopoulos, Nikolaou, Michailidis, Kampouris and Batjakas2020). It reaches 28.5 cm TL (Sabrah, Reference Sabrah2015), with first maturity around 16 cm (Farrag et al., Reference Farrag, Osman, Mehanna and Osman2018; Evagelopoulos et al., Reference Evagelopoulos, Nikolaou, Michailidis, Kampouris and Batjakas2020). P. forsskali is a benthic predator feeding mainly on invertebrates associated with sandy substrates (Hobson, Reference Hobson1974). In the eastern Mediterranean, its diet is dominated by crustaceans, followed by polychaetes and, to a lesser extent, foraminiferans, fishes, molluscs, echinoderms, sipunculans, and plant material (Evagelopoulos et al., Reference Evagelopoulos, Nikolaou, Michailidis, Kampouris and Batjakas2020; Vagenas et al., Reference Vagenas, Dogrammatzi, Kondylatos and Karachle2023; Michail et al, Reference Michail, Pyle and Chartosia2024). Its trophic level is comparable to that of other mullids (Mullus spp. and Upeneus spp.), suggesting potential competition for resources (Michail et al., Reference Michail, Pyle and Chartosia2024). Recent studies indicate that P. forsskali is an opportunistic predator capable of exploiting diverse prey in its new environment, with potential impacts on local food webs and commercially important species (Michail et al., Reference Michail, Pyle and Chartosia2024). The apparent replacement of native mullids in Cypriot artisanal fisheries may reflect competitive interactions for space and resources (Evagelopoulos et al., Reference Evagelopoulos, Nikolaou, Michailidis, Kampouris and Batjakas2020). Given these dynamics, the ecological consequences of its westward expansion warrant close monitoring, particularly in areas of high conservation value. The present records from Lampedusa Island highlight once again the crucial role of fishers in detecting and reporting NIS. The capture of multiple specimens over 2 consecutive years suggests that P. forsskali may be in an early stage of establishment in the Strait of Sicily. Strengthening collaborations with fishers is therefore essential for improving NIS surveillance, as demonstrated in previous initiatives (Mannino and Balistreri, Reference Mannino and Balistreri2018; Tiralongo et al., Reference Tiralongo, Crocetta, Riginella, Lillo, Tondo, Macali, Mancini, Russo, Coco, Paolillo and Azzurro2020; Perzia et al., Reference Perzia, Cillari, Crociata, Deidun, Falautano, Franzitta, Galdies, Maggio, Vivona and Castriota2023). Their involvement should be considered a key component of the early detection system (Maggio et al., Reference Maggio, Perzia, Falautano, Visconti and Castriota2022), which is most effective when invasions are still in their initial phases (Simberloff et al., Reference Simberloff, Martin, Genovesi, Maris, Wardle, Aronson, Courchamp, Galil, García-Berthou, Pascal, Pyšek, Sousa, Tabacchi and Vilà2013). Integrating fishers’ observations into structured citizen-science frameworks could provide an efficient early-warning mechanism, enabling timely detection and rapid response to biological invasions in the Mediterranean Sea.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0025315426101349.
Acknowledgements
The authors are very grateful to Mr. Piero Billeci, president of the local fishermen’s cooperative and to the fishermen of Lampedusa who provided us the specimens and related documentation of Parupeneus forsskali: Vincenzo Di Maggio, Pietro Famularo, Antonino Prestipino, Salvatore Prestipino, Salvatore Riso, Daniele Riso, Domenico Palmisano, Rosario Palmisano, and Pietro Palmisano. Our thanks also go to Enrico Bellia, Curator of the Museum of Zoology ‘P. Doderlein’ of Palermo, for cataloguing the first specimen. During the preparation and revision of this manuscript, the authors used Microsoft 365 Copilot for the purpose of English language revision. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Author contributions
M.F., L.C., and T.M. provided the same contribution to conceptualization, methodology, validation, formal analysis, investigation, resources, data curation, writing – original draft preparation, writing – review & editing, visualization, and supervision. All authors have read and agreed to the published version of the manuscript.
Funding
The present research received no external funding.
Competing interests
The authors declare none.
Data availability statement
The sequence of Parupeneus forsskali obtained in this study is publicly available on GenBank with the accession number PX619320. The specimen analysed in this study was deposited in the ichthyological collection of the Museum of Zoology ‘P. Doderlein’ of Palermo under accession code PL 392.
Ethical standards
Ethical review and approval were not required for this study because this work does not contain any experimental studies with live animals.