

1. Introduction
Syntactic processing, a core cognitive component of language comprehension, exhibits age-related decline that serves as a sensitive marker of cognitive ageing. Some studies suggest that older adults’ difficulties with complex syntactic structures are closely associated with reduced working memory capacity and diminished cognitive control (Campbell et al., Reference Campbell, Samu, Davis, Geerligs, Mustafa and Tyler2016; Shafto & Tyler, Reference Shafto and Tyler2014). Notably, pauses, as typical disfluency features in speech production, may offer unique insights into the cognitive mechanisms underlying age-related changes in syntactic processing. Acoustically, pauses are defined as the absence of speech signals (Lickley, Reference Lickley2015), while prosodically, they are defined as prosodic boundary markers (Männel & Friederici, Reference Männel and Friederici2016). Emerging evidence indicates that pause frequency and distribution can reliably predict the progression of neurodegenerative diseases (Yuan et al., Reference Yuan, Bian, Cai, Huang, Ye and Church2020, Reference Yuan, Cai, Bian, Ye and Church2021). However, while existing research has primarily focused on the clinical diagnostic value of pauses in neurodegenerative disorders (e.g., Alzheimer’s disease [AD]), their potential compensatory role in syntactic processing during normal ageing remains underexplored.
The current research debate centres on whether pauses encountered by older adults during complex syntactic processing reflect cognitive overload in online syntactic construction (where pauses may amplify the difficulty of disambiguating garden-path [GP] sentences) or represent an active cognitive regulation strategy employed by older adults (utilizing pauses to integrate cognitive resources and maintain processing efficiency). Resolving this issue would further elucidate how pauses modulate older adults’ syntactic processing and contribute to a deeper understanding of the neural mechanisms underlying late-life language processing in cognitive ageing.
1.1. Syntactic processing ability in older adults
In terms of sentence production, older adults tend to reduce their use of complex syntax, such as non-garden-path (NGP) sentences, showing significantly decreased syntactic complexity in oral narratives (Agmon et al., Reference Agmon, Pradhan, Ash, Nevler, Liberman, Grossman and Cho2024; Kemper et al., Reference Kemper, Herman and Lian2003). They are more likely to produce syntactically incomplete sentences in spontaneous speech (Kemper et al., Reference Kemper, Herman and Liu2004), a phenomenon attributed to age-related limitations in syntactic construction abilities (Campbell et al., Reference Campbell, Samu, Davis, Geerligs, Mustafa and Tyler2016; Davis et al., Reference Davis, Zhuang, Wright and Tyler2014; Meunier et al., Reference Meunier, Stamatakis and Tyler2014; Samu et al., Reference Samu, Campbell, Tsvetanov, Shafto and Tyler2017).
Regarding sentence comprehension, whether older adults’ syntactic processing abilities are affected by ageing remains controversial. Some healthy older adults demonstrate performance comparable to younger adults in tasks involving ambiguous sentence resolution (e.g., judging the plausibility of subsequent constituents) and syntactic-semantic integration (Campbell et al., Reference Campbell, Samu, Davis, Geerligs, Mustafa and Tyler2016; Tyler et al., Reference Tyler, Shafto, Randall, Wright, Marslen-Wilson and Stamatakis2010), suggesting relatively preserved syntactic processing abilities. However, when tasks require real-time processing of progressively complex syntactic structures (e.g., hierarchically nested constructions), older adults show significantly prolonged reaction times (RTs) and decreased accuracy (ACC) (Obler et al., Reference Obler, Fein, Nicholas and Albert1991; Waters & Caplan, Reference Waters and Caplan2001) and this performance is not improved by factors such as altered speech rate (Small & Cochrane, Reference Small and Cochrane2024). This discrepancy may stem from methodological differences: offline tasks (e.g., sentence plausibility judgments) allow reliance on semantic compensation strategies, whereas online tasks (e.g., picture-word matching) are more sensitive to age-related declines in syntactic processing efficiency (Antonenko et al., Reference Antonenko, Brauer, Meinzer, Fengler, Kerti, Friederici and Flöel2013; Kemtes & Kemper, Reference Kemtes and Kemper1997).
In studies of syntactic processing in older adults, ambiguous structures serve as sensitive indicators of syntactic processing ability due to their high complexity. Beyond general ambiguous structures, researchers have particularly focused on older adults’ processing of ‘garden-path (GP)’ sentences. The term ‘garden-path’ derives from the experience of parsing such sentences: much like being led astray on a garden path, readers proceed linearly until the end of the sentence, only to realize they must backtrack and reanalyse the structure (Bever, Reference Bever and Hayes1970). The GP effect arises primarily because a potential ambiguous structure in the earlier part of the sentence becomes unequivocally ambiguous as the sentence unfolds, leading the reader or listener to initially build an incorrect syntactic representation. When subsequent words cannot be integrated into the structure, a syntactic mismatch or expectation violation occurs, triggering reanalysis to seek an alternative syntactic structure (Slattery et al., Reference Slattery, Sturt, Christianson, Yoshida and Ferreira2013). Moreover, studies indicate that reanalysis of GP sentences occurs mainly at the disambiguating region. Eye-tracking research has shown that participants exhibit longer reading times and higher regression rates at disambiguation points (Slattery et al., Reference Slattery, Sturt, Christianson, Yoshida and Ferreira2013). ERP studies on GP sentences also reveal that syntactic violations at the disambiguation point elicit a larger P600 component amplitude (O’Rourke & Colflesh, Reference O’Rourke and Colflesh2015; Tanner & Van Hell, Reference Tanner and Van Hell2014). Furthermore, Samu et al. (Reference Samu, Campbell, Tsvetanov, Shafto and Tyler2017) combined behavioural and neuroimaging (magnetic resonance imaging [MRI]) approaches to demonstrate that older adults make significantly more errors than younger adults when process sentences with syntactic ambiguity, accompanied by atypical prefrontal activation patterns, suggesting that declines in inhibitory control may hinder syntactic reanalysis. However, existing research has primarily focused on Indo-European languages, leaving the investigation of Chinese-speaking older adults an area requiring further exploration.
1.2. The impact of pauses on syntactic processing in older adults
In existing research, silent intervals exceeding a specific duration threshold are typically defined as ‘silent pauses’ or ‘unfilled pauses’ (Zellner, Reference Zellner and Keller1994). In this study, we simply refer to them as ‘pauses’. Pauses primarily function to maintain prosodic structure during syntactic processing, indicating word-finding difficulties (Goldman-Eisler, Reference Goldman-Eisler1958a, Reference Goldman-Eisler1958b; Kircher et al., Reference Kircher, Brammer, Levelt, Bartels and McGuire2004; Maclay & Osgood, Reference Maclay and Osgood1959) while delaying the initiation of subsequent lexical processing. This delay may facilitate the identification of content following pause insertion (Howell & Young, Reference Howell and Young1991), thereby potentially enhancing overall syntactic processing.
Research on older adults with neurodegenerative disorders (e.g., mild cognitive impairment [MCI], AD) demonstrates that pauses can sensitively reflect cognitive load during semantic retrieval or syntactic planning (Gayraud et al., Reference Gayraud, Lee and Barkat-Defradas2011). Older patients with MCI or AD exhibit significantly higher pause frequencies than cognitively healthy older adults, particularly when facing semantic processing challenges. Prolonged pauses may compensate for episodic memory impairment by extending information integration time, thereby assisting AD patients in memory retrieval during narrative tasks (Pistono et al., Reference Pistono, Jucla, Barbeau, Saint-Aubert, Lemesle, Calvet, Köpke, Puel and Pariente2016). Pause distribution shows sensitivity to syntactic positions, occurring more frequently at sentence-initial positions and within complex syntactic structures (e.g., before noun phrases). Increased word repetition following pauses suggests that AD patients compensate for language impairment by selecting more common words (Gayraud et al., Reference Gayraud, Lee and Barkat-Defradas2011), a phenomenon interpreted as a simplification strategy resulting from declining syntactic planning abilities (Lofgren & Hinzen, Reference Lofgren and Hinzen2022). Older adults with impaired complex syntactic production ability may produce pauses more frequently at prosodic boundaries, suggesting an association between pauses and syntactic processing (Lee et al., Reference Lee, Huber, Jenkins and Fredrick2019). Neuroimaging studies further reveal that reduced grey matter density in the prefrontal BA10 region correlates with increased pause frequency at syntactically relevant positions, while prefrontal atrophy may require patients to spend more time integrating information when recalling events (Pistono et al., Reference Pistono, Jucla, Barbeau, Saint-Aubert, Lemesle, Calvet, Köpke, Puel and Pariente2016) and decreased grey matter density in the left temporal pole positively correlates with pause frequency during semantic tasks (Pistono et al., Reference Pistono, Pariente, Bézy, Lemesle, Le Men and Jucla2019). Notably, AD patients’ pause patterns dynamically change with disease progression: early-stage patients increase filled pauses (e.g., ‘um’, ‘uh’) to maintain conversational fluency, but lose this regulatory ability as the disease advances (Lofgren & Hinzen, Reference Lofgren and Hinzen2022; Pistono et al., Reference Pistono, Pariente, Bézy, Lemesle, Le Men and Jucla2019). These findings suggest that pauses serve dual functions as both pathological markers and cognitive compensatory mechanisms. A cross-linguistic study highlights the complex relationship between pauses and syntactic processing. Spanish-speaking AD patients show significantly higher standard deviations and pairwise variability indices of syllabic intervals including pauses in spontaneous speech compared to healthy controls, indicating impaired prosodic regularity (Martínez-Sánchez et al., Reference Martínez-Sánchez, Meilán, Vera-Ferrandiz, Carro, Pujante-Valverde, Ivanova and Carcavilla2017). This finding suggests language-specific effects of pauses on syntactic processing.
Although existing evidence demonstrates close associations between pauses and syntactic processing in clinical populations, critical gaps remain regarding healthy ageing. Whether findings from Indo-European languages generalize to morphologically impoverished languages like Chinese remains questionable. Unlike Indo-European languages, which rely on morphological characters, Chinese depends primarily on word order to convey grammatical and semantic information (Du & Chen, Reference Du and Chen2021). Consequently, complex word order may impose greater memory load on older adults to maintain and parse syntactic structures, leading to reduced comprehension of long-distance or nested constructions (Beese et al., Reference Beese, Werkle-Bergner, Lindenberger, Friederici and Meyer2019; Poulisse et al., Reference Poulisse, Wheeldon and Segaert2019). This aligns with the domain-general decline theory, which posits that older adults exhibit constrained processing resources (Cansino et al., Reference Cansino, Torres-Trejo, Estrada-Manilla, Pérez-Loyda, Vargas-Martínez, Tapia-Jaimes and Ruiz-Velasco2020), yet may also develop compensatory strategies to maintain language comprehension. Thus, empirical studies on pause-syntax interactions in healthy Chinese older adults are urgently needed. In particular, whether pauses can facilitate healthy Chinese older adults’ processing of complex syntactic structures (e.g., GP sentence) requires systematic investigation.
1.3. Brain mechanisms associated with pauses in garden-path sentence processing
The P600 component, a well-established marker of syntactic processing, is closely linked to syntactic violation detection, syntactic reanalysis (Friederici, Reference Friederici2002) and syntactic integration (Kaan & Swaab, Reference Kaan and Swaab2003). This component primarily activates in parietal lobe and mid-posterior scalp regions when processing phrase structure errors or lexical category violations (Friederici, Reference Friederici2011; Hao et al., Reference Hao, Duan and Zhang2021), while P600 activity in fronto-central region correlates with the resolution of syntactic complexity and structural ambiguity (Kaan et al., Reference Kaan, Harris, Gibson and Holcomb2000). Research on GP sentence demonstrates P600 activation at points where the parser attempts to revise syntactic ambiguity (Frisch et al., Reference Frisch, Schlesewsky, Saddy and Alpermann2002). However, inserted pauses may reinforce initial misanalysis, requiring additional time for disambiguation and consequently attenuating P600 amplitude (Maxfield et al., Reference Maxfield, Lyon and Silliman2009).
The N400 component, predominantly associated with left fronto-temporal neural networks (Payne & Silcox, Reference Payne and Silcox2019), also shows occasional activation in left posterior middle temporal gyrus (Lau & Namyst, Reference Lau and Namyst2019) and bilateral temporal and partial occipital visual cortex regions (Khateb et al., Reference Khateb, Pegna, Landis, Mouthon and Annoni2010). This component reflects lexical-semantic processing and discourse-level integration (Kutas et al., Reference Kutas, Van Petten and Kluender2006), as well as predictive processing of upcoming input (Lau et al., Reference Lau, Namyst, Fogel and Delgado2016; MacGregor et al., Reference MacGregor, Corley and Donaldson2009). During pause-modulated syntactic processing, the predictability of post-pause words could significantly modulate N400 effects (Corley et al., Reference Corley, MacGregor and Donaldson2007; MacGregor et al., Reference MacGregor, Corley and Donaldson2009). Some studies suggest that pauses delay the appearance of predicted words, consequently postponing N400 peak latency – a phenomenon attributed to missing coarticulation cues that require additional cognitive resources for processing unexpected phonetic information (Besson et al., Reference Besson, Faita, Czternasty and Kutas1997). GP sentence studies reveal N400 activation at points of temporary ambiguity, indicating listeners’ tendency to maintain initial misanalyses while attempting to integrate subsequent disambiguating elements (e.g., disambiguating verbs) into this incorrect interpretation (Maxfield et al., Reference Maxfield, Lyon and Silliman2009). Collectively, these findings establish the N400’s dual role in pause-related GP sentence processing: reflecting both the difficulty induced by pauses and engagement in syntactic disambiguation.
Building on this evidence, the current study employs event-related potential (ERP) techniques with Chinese GP sentences to address two fundamental questions: (1) whether pauses modulate older adults’ syntactic processing by providing additional time for cognitive resource integration and (2) whether pause-related ERP components reveal neural signatures of compensatory resource reallocation in ageing. The findings will advance our understanding of how disfluency features interact with age-related syntactic processing while identifying potential neurocognitive markers for language-based assessment of cognitive ageing.
2. Methods
2.1. Participants
The study included 24 older adults and 24 college students, all from Nantong City, Jiangsu Province of China, with 4 male older adults and 9 male young adults. The older participants were aged 60–80 years (M = 66.08, SD = 6.04), while the younger participants were aged 18–30 years (M = 22.04, SD = 3.22). All older participants passed standardized cognitive screening using the Beijing version of the Montreal cognitive assessment (MoCA) and mini-mental state examination (MMSE) scales. The average score of MoCA is over 26 (MoCA: M = 26.46, SD = 2.25; corrected by education factor) and the average score of MMSE is over 27 (MMSE: M = 27.58, SD = 2.19). Hearing assessment using the HHIE-S showed normal hearing levels, with older participants scoring an average of 2.75 (5th percentile = 0, 95th percentile = 10).
All participants met the following criteria: (a) at least elementary school education (older adults: 6–16 years, M = 10.67, SD = 3.00; young adults: 12–17 years, M = 14.96, SD = 1.85). An independent samples t-test was conducted on the years of education (YoE) between the older and younger groups, with the following results: the junior group had significantly higher YoE than the senior group, with a mean difference of 4.42, t (37.69) = −6.17, p < 0.001, Hedges’ g = −1.75. Although a significant difference was observed, this primarily reflects an objective reality of China’s social history: the older group has fewer YoE, largely because they grew up in an era with relatively limited educational resources, especially in rural areas (Pei et al., Reference Pei, Cong and Wu2020; Ross & Zhang, Reference Ross and Zhang2008), while the younger group has benefited from the expansion of higher education in recent decades. Therefore, this intergroup difference in YoE is systemic and generational, rather than a random error in our research design. Secondly, the study on language ageing indicates that the native language syntactic knowledge system of older adults remains relatively stable and is primarily influenced by age-related general cognitive abilities (e.g., working memory) rather than by YoE itself (Bertola et al., Reference Bertola, Ávila, Bicalho and Malloy-Diniz2019). Thus, differences in YoE may not affect the findings of this study regarding syntactic processing across different age groups; (b) right-handedness; (c) normal or corrected-to-normal vision; (d) no history of cognitive training, brain trauma, neurological disorders, or genetic diseases and (e) fluent Mandarin proficiency. All participants signed informed consent forms, and their personal information was kept strictly confidential. Participants could terminate the experiment at any time and received appropriate compensation after completing the study.
2.2. Experiment design and materials
The experiment employed a 2 × 2 within-subjects design, with sentence type (GP sentences versus ‘Non-garden-path’ sentences) and pause existence (pause-absent [PA] versus pause-present [PP]) as independent variables. Each condition consisted of 30 trials, totalling 120 trials per participant. Trials were presented in a pseudo-randomized order, with unique sequences for each participant.
All auditory stimuli were professionally recorded using a female announcer certified at Level 1-B (the second-highest tier) in Mandarin proficiency. The recordings were edited using Adobe Audition software and digitized at 16-bit resolution with a 44.1 kHz sampling rate (mono format).
Linguistic Materials
We use two types of sentences as materials: (1) GP sentences: These contained the structurally ambiguous ‘NP1 + VP + NP2 + 的 (Pinyin: de, structural particle) + NP3’ construction (e.g., ‘医生嘱托了病人的家属’, Pinyin: yī shēng zhǔ tuō le bìng rén de jiā shǔ, The doctor instructed the patient’s family member), where NP2 and NP3 had a possessive relationship (e.g., ‘病人的家属’, Pinyin: bìng rén de jiā shǔ, the patient’s family member). (2) NGP sentences: These followed the unambiguous ‘NP1 + VP + AP + 的 (structural particle) + NP2’ structure (e.g., ‘医生嘱托了那位住院的病人’, Pinyin: yī shēng zhǔ tuō le nà wèi zhù yuàn de bìng rén, The doctor instructed that hospitalized patient), where AP modified NP2 (e.g., ‘住院的病人’, Pinyin: zhù yuàn de bìng rén, hospitalized patient).
Each sentence was followed by a yes/no comprehension question (‘NP1 + VP + 的 (Pinyin: de, structural particle) + 是(Pinyin: shì, linking verb) + NP2 + 吗(Pinyin: ma, interrogative particle)?’; e.g., ‘医生嘱托的是病人吗?’, Pinyin: yī shēng zhǔ tuō de shì bìng rén ma? Was it the patient whom the doctor instructed?). Pauses (1000 ms) were inserted immediately before ‘的’ to induce ‘garden-path’ effects.
Prior researches on pause thresholds for prosodic boundaries in Mandarin indicate that silent segments longer than 200 ms can generally be effectively perceived as pauses and retain independent prosodic functions to adequately mark boundary strength (Wang et al., Reference Wang, Xu and Ding2017; Wu et al., Reference Wu, Adda-Decker and Lamel2020). In contrast, generally, silent segments exceeding 1000 ms in duration are less likely to be perceived as natural prosodic pauses (Campione & Véronis, Reference Campione and Véronis2002; Šturm & Volín, Reference Šturm and Volín2023). Therefore, a 1000 ms pause was inserted immediately before ‘的’ to induce ‘garden-path’ effect in this study.
2.2.1. Lexical controls
All content words were disyllabic, with matched durations across conditions. For NP1, we employed common human referents (e.g., ‘医生’ Pinyin: yī sheng, doctor; ‘教授’ Pinyin: jiào shòu, professor; ‘小红’ Xiaohong [Name]). VPs consisted of high-frequency verbs (e.g., ‘嘱托’ Pinyin: zhǔ tuō, instruct; ‘忘记’ Pinyin: wàng jì, forget; ‘赞美’ Pinyin: zàn měi, praise) suffixed with ‘了 (Pinyin: le, perfective aspect marker)’. In GP sentences, NPs (NP2 and NP3) were semantically related (e.g., ‘病人的家属’ Pinyin: bìng rén de jiā shǔ, patient’s family member; ‘作家的文章’ Pinyin: zuò jiā de wén zhāng, writer’s article). For NGP sentences, adjectives/adjectival phrases (e.g., ‘住院(的)’ Pinyin: zhù yuàn (de), hospitalized; ‘优秀’ Pinyin: yōu xiù, excellent; ‘老旧’ Pinyin: lǎo jiù, old-fashioned) modified NP2.
All lexical items were selected from the Modern Chinese Dictionary (7th edition). All materials can be found on the Open Science Framework at https://doi.org/10.17605/OSF.IO/C2FNB.
To mitigate the potential influence of stimulus familiarity on experimental results, a rigorous material screening procedure was implemented prior to the formal experiment. Through an online survey platform, 140 participants who did not participate in the subsequent main experiment were recruited. They systematically evaluated the familiarity of the experimental materials using a standardized 5-point Likert scale (1 = very unfamiliar, 5 = very familiar). Statistical analysis confirmed that the familiarity ratings of all selected materials met the predetermined criteria, effectively ensuring balanced familiarity across all experimental conditions. Since the four sentence types shared identical syntactic structures and morphemic composition, only the familiarity ratings for the GP sentences without pauses and the NGP sentences without pauses were analysed. The familiarity ratings for both sentence types were above 4 (GP sentences without pauses: M = 4.19, SD = 0.11; NGP sentences without pauses: M = 4.04, SD = 0.11), with no significant difference in familiarity between them (p > 0.05). Seventy-three additional participants (not in main experiment/familiarity evaluation) rated experimental materials’ semantic plausibility via a standardized 5-point Likert scale (1 = very implausible, 5 = very plausible). Statistical analysis showed no significant semantic plausibility difference across conditions (GP sentences without pauses: M = 4.27, SD = 0.31; NGP sentences without pauses: M = 4.39, SD = 0.21; p > 0.05).
Example stimuli are provided in Table 1.
Examples of experimental stimuli

Table 1. Long description
The table has two columns labeled Stimuli type and Example sentences. From top to bottom, the first row is GP-PA with the Chinese sentence Xiaohong checked the goods’ colours, Pinyin transcription, and English translation. The second row is GP-PP with the same sentence but a marked pause before the structural particle, shown in both Pinyin and English as [pause]. The third row is NGP-PA with the Chinese sentence Xiaohong checked the recycled goods, Pinyin, and English translation. The fourth row is NGP-PP with the same sentence but a pause before the structural particle, indicated in both Pinyin and English as [pause]. The table footnote explains GP as garden-path sentences with ambiguous NP1 plus VP plus NP2 plus de plus NP3 structure, NGP as unambiguous NP1 plus VP plus AP plus de plus NP2 structure, PA as pause-absent, and PP as pause-present with a 1000 millisecond pause before de. It notes all content words are disyllabic and that NP2-NP3 pairs in GP share possessive relationships.
Note: GP, Garden-path sentences with temporarily ambiguous ‘NP1 + VP + NP2 + 的 (Pinyin: de, structural particle) + NP3’ structure (e.g., ‘货物的颜色’ interpreted as the goods’ colours before disambiguation). NGP with unambiguous ‘NP1 + VP + AP + 的 (Pinyin: de, structural particle) + NP2’ structure (e.g., ‘回收的货物’ parsed as the recycled goods). PA, Pause-absent; PP, Pause-present (1000 ms pause inserted before ‘的 [structural particle]’). All content words were disyllabic, and NP2-NP3 pairs in GP shared possessive relationships (e.g., 货物-颜色 goods-colours).
2.3. Experimental apparatus
The experiment was programmed using E-Prime 3.0. Electroencephalography (EEG) data were recorded via a 64-channel Ag/AgCl electrode cap (following the international 10–20 system) using a Neuroscan acquisition system. Auditory stimuli were delivered through computer speakers, and behavioural responses (ACC and RT) were collected via an external keyboard. The experimenter monitored participants’ task comprehension and response correctness.
2.4. Procedure
The experiment was conducted in a sound-attenuated booth, administered by two trained graduate research assistants. Prior to the formal task, participants received detailed instructions and completed a practice session with distinct materials (excluded from analysis).
Each trial began with an 800 ms fixation cross (‘+’) displayed at the centre of the screen. Following this, participants heard an auditory sentence stimulus, with durations varying by condition: PA sentences lasted 8200 ms, while PP sentences lasted 9200 ms due to the insertion of a 1000 ms pause before the critical word de. After stimulus presentation, participants judged the correctness of a follow-up question by keypress. Individual trial durations averaged approximately 10s (PA) and 11 s (PP), and the entire experimental session lasted around 25 minutes.
See Figure 1 for the schematic workflow.
Schematic workflow of the experiment.

2.5. EEG recording and analysis
2.5.1. Data acquisition & preprocessing
EEG data were recorded using a 64-channel Neuroscan system with electrodes placed according to the international 10–20 system. Electrode impedances were maintained below 10 kΩ, and signals were sampled at 500 Hz with a bandpass filter (0.01–100 Hz). Online recording used the right mastoid (M2) as the reference, while vertical (VEOG) and horizontal (HEOG) electrooculograms were monitored. Offline preprocessing was performed in MATLAB R2023a with EEGLAB 2024.2 (Delorme & Makeig, Reference Delorme and Makeig2004). The EEG data were re-referenced to the average reference following the detection of anomalies in the left mastoid (M1) channel for some participants. A bandpass filter (0.1–40 Hz) and a notch filter (48–52 Hz) were applied to remove line noise and high-frequency artefacts. Bad channels were corrected via spherical interpolation, and artefactual segments were manually rejected. Ocular and muscular artefacts were further removed using independent component analysis (ICA). Finally, EEG epochs corresponding to behavioural trials with 0% ACC or RTs exceeding ±3 standard deviations from the mean were excluded from analysis.
2.5.2. Time-domain analysis
For time-domain analysis, epochs were segmented from −1000 to 2000 ms time-locked to the onset of the critical word ‘的de’, with a baseline correction window of −1000 to 0 ms. Regions of interest (ROIs) were selected for signal averaging across the following electrode clusters: frontal (F3, F1, F4, F2, FZ), fronto-central (FC3, FC1, FC4, FC2, FCZ), central (C3, C1, C4, C2, CZ), centro-parietal (CP3, CP1, CP4, CP2, CPZ) and parietal (P3, P1, P4, P2, PZ) regions. Through the observation of participants’ EEG waveforms, we found the widespread presence of the P200 component across all conditions. Therefore, it was also included in the analysis. Based on grand average waveforms of older adult participants, the P200 component exhibited a concentrated peak, while the N400 and P600 components showed greater variability across conditions. Therefore, based on the fronto-central peak amplitudes observed in older adults and the specific requirements of this study, the analysis windows were determined as follows: 170–240 ms for P200 (centred on the fronto-central peak ±20 ms), 340–440 ms for N400 (100 ms window anchored to the fronto-central peak) and 600–700 ms for P600 (100 ms window anchored to the fronto-central peak).
2.5.3. Time-frequency analysis
Time-frequency analysis was conducted using the Letswave 7 toolbox (https://github.com/NOCIONS/letswave7) with MATLAB custom scripts (https://doi.org/10.17605/OSF.IO/C2FNB) to estimate event-related spectral perturbation (ERSP). The short-time Fourier transform (STFT) was applied with a frequency resolution of 1 Hz (1–30 Hz range) using a 100 ms Hanning window. Following preprocessing, a new baseline correction window (−800 to −200 ms) was applied, and ERSP data were baseline-normalized using the subtractive method. ROIs matched those used in the time-domain analysis.
2.5.4. Frequency bands of interest
The power in the θ-band demonstrates increased activity in response to syntactic violations, reflecting the cognitive load of syntactic integration (Pérez et al., Reference Pérez, Molinaro, Mancini, Barraza and Carreiras2012) while also being closely associated with online processing of prosodic rhythm (Yan & Zhang, Reference Yan and Zhang2022). Significant oscillations in the α-band may indicate predictive processing and inferential reasoning of syntactic structures, actively participating in real-time syntactic computation (Zioga et al., Reference Zioga, Weissbart, Lewis, Haegens and Martin2023). Accordingly, based on the statistical patterns observed in ERSP, this study focused its analysis on mean power within the θ-band (4–8 Hz) and α-band (10–14 Hz), using a consistent time window of 200–800 ms for both bands.
3. Results
3.1. Behavioural data measurements
Trials with ACC rates of 0%, RTs shorter than 100 ms or longer than 2000 ms, or RTs exceeding ±3 standard deviations were excluded from analysis. Statistical analyses were performed on the mean RTs and ACC rates under each condition for both the older and younger adult groups. Detailed results are presented in Tables 2 and 3.
The result of older and younger adults’ RT

Table 2. Long description
The table has five columns: Age, Sentence type, Pause existence, RT mean (M), and RT standard deviation (SD), with values in milliseconds. For older adults: GP, PA has M 987, SD 354; GP, PP has M 882, SD 285; NGP, PA has M 750, SD 184; NGP, PP has M 764, SD 186. For younger adults: GP, PA has M 681, SD 94; GP, PP has M 690, SD 97; NGP, PA has M 658, SD 94; NGP, PP has M 666, SD 99. GP stands for garden-path sentences, NGP for non-garden-path sentences, PA for pause-absent, PP for pause-present. RT is reported in milliseconds as whole numbers.
Note: RT data are reported in milliseconds (ms) as whole numbers, reflecting the intrinsic precision of the measurement system.
Abbreviations: GP, garden-path sentences; NGP, non-garden-path sentences; PA, pause-absent; PP, pause-present; RT, RT (in milliseconds); M, mean; SD, standard deviation.
The result of older and younger adults’ ACC

Table 3. Long description
Starting from the top, the table is divided into two main Age groups: Older and Younger. For each group, data are presented for two Sentence types: GP (garden-path) and NGP (non-garden-path). Each Sentence type is further split into Pause existence: PA (pause-absent) and PP (pause-present). For Older adults: GP with PA has mean accuracy 82.1 percent, SD 0.17; GP with PP has mean 79.2 percent, SD 0.18; NGP with PA has mean 96.5 percent, SD 0.08; NGP with PP has mean 92.8 percent, SD 0.10. For Younger adults: GP with PA has mean 82.3 percent, SD 0.15; GP with PP has mean 84.7 percent, SD 0.15; NGP with PA has mean 93.6 percent, SD 0.09; NGP with PP has mean 92.7 percent, SD 0.10. Abbreviations are defined as follows: GP is garden-path sentences, NGP is non-garden-path sentences, PA is pause-absent, PP is pause-present, ACC is accuracy in percent, M is mean, SD is standard deviation.
Abbreviations: GP, garden-path sentences; NGP, non-garden-path sentences; PA, pause-absent; PP, pause-present; ACC, accuracy (in percentage); M, mean; SD, standard deviation.
A 2 (Age group: older, younger) × 2 (Sentence type: GP versus NGP) × 2 (Pause existence: PA versus PP) three-way repeated-measures ANOVA was conducted for both ACC and RT data. Bonferroni correction was applied to all post hoc pairwise comparisons.
For RT analysis: A significant main effect of Age group was observed (F (1, 46) = 11.77, p = 0.001,
$ {\eta}_p^2= $
 0.20) with older adults showing significantly longer RTs than younger adults. Additionally, a significant main effect of Sentence type was found (F (1, 46) = 26.41, p < 0.001,
$ {\eta}_p^2= $
0.20) with older adults showing significantly longer RTs than younger adults. Additionally, a significant main effect of Sentence type was found (F (1, 46) = 26.41, p < 0.001,
$ {\eta}_p^2= $
 0.37), indicating that GP eliciting longer RTs than NGP. Finally, a significant main effect of Pause existence emerged (F (1, 46) = 4.51, p = 0.039,
$ {\eta}_p^2= $
0.37), indicating that GP eliciting longer RTs than NGP. Finally, a significant main effect of Pause existence emerged (F (1, 46) = 4.51, p = 0.039,
$ {\eta}_p^2= $
 0.09), showing shorter RTs in PP conditions. Significant two-way interactions were found. First, a significant Age group × Sentence type interaction emerged (F (1, 46) = 15.56, p < 0.001,
$ {\eta}_p^2= $
0.09), showing shorter RTs in PP conditions. Significant two-way interactions were found. First, a significant Age group × Sentence type interaction emerged (F (1, 46) = 15.56, p < 0.001,
$ {\eta}_p^2= $
 0.25). Second, there was a significant Age group× Pause existence interaction (F (1, 46) = 6.58, p = 0.014,
$ {\eta}_p^2= $
0.25). Second, there was a significant Age group× Pause existence interaction (F (1, 46) = 6.58, p = 0.014,
$ {\eta}_p^2= $
 0.13). Finally, a significant Sentence type × Pause existence interaction was observed (F (1, 46) = 13.77, p < 0.001,
$ {\eta}_p^2= $
0.13). Finally, a significant Sentence type × Pause existence interaction was observed (F (1, 46) = 13.77, p < 0.001,
$ {\eta}_p^2= $
 0.23). In addition, a significant three-way interaction was also observed: Age group × Sentence type × Pause existence (F (1, 46) = 12.42, p < 0.001,
$ {\eta}_p^2= $
0.23). In addition, a significant three-way interaction was also observed: Age group × Sentence type × Pause existence (F (1, 46) = 12.42, p < 0.001,
$ {\eta}_p^2= $
 0.21).
0.21).
Simple effects analyses revealed that, for both GP and NGP, older adults showed significantly longer RTs than younger adults (GP: F (1, 46) = 14.73, p < 0.001,
$ {\eta}_p^2= $
 0.24; NGP: F (1, 46) = 5.92, p = 0.019,
$ {\eta}_p^2= $
0.24; NGP: F (1, 46) = 5.92, p = 0.019,
$ {\eta}_p^2= $
 0.11). Older adults had significantly longer RTs for GP than NGP condition (F (1, 46) = 41.26, p < 0.001,
$ {\eta}_p^2= $
0.11). Older adults had significantly longer RTs for GP than NGP condition (F (1, 46) = 41.26, p < 0.001,
$ {\eta}_p^2= $
 0.47). In both PA and PP conditions, older adults showed significantly longer RTs than younger adults (PA: F (1, 46) = 13.22, p < 0.001,
$ {\eta}_p^2= $
0.47). In both PA and PP conditions, older adults showed significantly longer RTs than younger adults (PA: F (1, 46) = 13.22, p < 0.001,
$ {\eta}_p^2= $
 0.22; PP: F (1, 46) = 9.54, p = 0.003,
$ {\eta}_p^2= $
0.22; PP: F (1, 46) = 9.54, p = 0.003,
$ {\eta}_p^2= $
 0.17). Older adults responded significantly faster in PP than PA conditions (F (1, 46) = 10.99, p = 0.002,
$ {\eta}_p^2= $
0.17). Older adults responded significantly faster in PP than PA conditions (F (1, 46) = 10.99, p = 0.002,
$ {\eta}_p^2= $
 0.19). In both pause conditions, GP elicited significantly longer RTs than NGP (PA: F (1, 46) = 30.38, p < 0.001,
$ {\eta}_p^2= $
0.19). In both pause conditions, GP elicited significantly longer RTs than NGP (PA: F (1, 46) = 30.38, p < 0.001,
$ {\eta}_p^2= $
 0.40; PA: F (1, 46) = 14.54, p < 0.001,
$ {\eta}_p^2= $
0.40; PA: F (1, 46) = 14.54, p < 0.001,
$ {\eta}_p^2= $
 0.24). For GP, PP conditions showed significantly shorter RTs than PA conditions (F (1, 46) = 10.69, p = 0.002,
$ {\eta}_p^2= $
0.24). For GP, PP conditions showed significantly shorter RTs than PA conditions (F (1, 46) = 10.69, p = 0.002,
$ {\eta}_p^2= $
 0.19).
0.19).
Simple-simple effects analysis of the significant three-way interaction showed that, across all four conditions (GP/PA, GP/PP, NGP/PA, NGP/PP), older adults had significantly longer RTs than younger adults (Fs > 5.32, ps < 0.05,
$ {\eta}_p^2 $
 s > 0.10). For older adults, GP showed significantly longer RTs than NGP in both pause conditions (PA: F (1, 46) = 49.77, p < 0.001,
$ {\eta}_p^2= $
s > 0.10). For older adults, GP showed significantly longer RTs than NGP in both pause conditions (PA: F (1, 46) = 49.77, p < 0.001,
$ {\eta}_p^2= $
 0.52;PP: F (1, 46) = 20.71, p < 0.001,
$ {\eta}_p^2= $
0.52;PP: F (1, 46) = 20.71, p < 0.001,
$ {\eta}_p^2= $
 0.31).Older adults showed significantly shorter RTs for GP in PP versus PA conditions (F (1, 46) = 22.56, p < 0.001,
$ {\eta}_p^2= $
0.31).Older adults showed significantly shorter RTs for GP in PP versus PA conditions (F (1, 46) = 22.56, p < 0.001,
$ {\eta}_p^2= $
 033). No significant Sentence type × Pause existence interaction was found for younger adults.
033). No significant Sentence type × Pause existence interaction was found for younger adults.
For ACC analysis, while no significant main effect of Age group (F (1, 46) = 2.44, p = 0.16,
$ {\eta}_p^2= $
 0.05) or Pause existence (F (1, 46) = 1.50, p = 0.23,
$ {\eta}_p^2= $
0.05) or Pause existence (F (1, 46) = 1.50, p = 0.23,
$ {\eta}_p^2= $
 0.03) were observed, a significant main effect of Sentence type emerged (F (1, 46) = 25.15, p < 0.001,
$ {\eta}_p^2= $
0.03) were observed, a significant main effect of Sentence type emerged (F (1, 46) = 25.15, p < 0.001,
$ {\eta}_p^2= $
 0.35) with lower ACC for GP than NGP. Furthermore, A significant Sentence type × Pause existence interaction was found (F (1, 46) = 14.00, p < 0.001,
$ {\eta}_p^2 $
0.35) with lower ACC for GP than NGP. Furthermore, A significant Sentence type × Pause existence interaction was found (F (1, 46) = 14.00, p < 0.001,
$ {\eta}_p^2 $
 = 0.23) with no other interactions reached significance. Simple effects analyses showed that, in both pause conditions, GP had significantly lower ACC than NGP (PA: F (1, 46) = 34.25, p < 0.001,
$ {\eta}_p^2= $
= 0.23) with no other interactions reached significance. Simple effects analyses showed that, in both pause conditions, GP had significantly lower ACC than NGP (PA: F (1, 46) = 34.25, p < 0.001,
$ {\eta}_p^2= $
 0.43; PP: F (1, 46) = 14.17, p < 0.001,
$ {\eta}_p^2= $
0.43; PP: F (1, 46) = 14.17, p < 0.001,
$ {\eta}_p^2= $
 0.26). For GP, PP conditions also showed significantly higher ACC than PA conditions (F (1, 46) = 6.88, p = 0.012,
$ {\eta}_p^2= $
0.26). For GP, PP conditions also showed significantly higher ACC than PA conditions (F (1, 46) = 6.88, p = 0.012,
$ {\eta}_p^2= $
 0.13).
0.13).
To account for the YoE’s confounding effect, an ANCOVA was conducted. For RT, Age group × YoE interaction was non-significant (F (1, 44) = 0.42, p = 0.52,
$ {\eta}_p^2= $
 0.01), supporting a standard ANCOVA. Adjusted for YoE, Age group main effect (F (1, 45) = 3.39, p = 0.072,
$ {\eta}_p^2 $
0.01), supporting a standard ANCOVA. Adjusted for YoE, Age group main effect (F (1, 45) = 3.39, p = 0.072,
$ {\eta}_p^2 $
 = 0.07) and YoE main effect ((F (1, 45) = 1.27, p = 0.27,
$ {\eta}_p^2= $
= 0.07) and YoE main effect ((F (1, 45) = 1.27, p = 0.27,
$ {\eta}_p^2= $
 0.02) were non-significant. For ACC: the Age group × YoE interaction was significant (F (1, 44) =9.55, p = 0.03,
$ {\eta}_p^2= $
0.02) were non-significant. For ACC: the Age group × YoE interaction was significant (F (1, 44) =9.55, p = 0.03,
$ {\eta}_p^2= $
 0.18), prompting moderation analysis. This interaction improved the model for GP/PA (ΔR2 = 0.112, F (1, 44) = 6.38, p = 0.015; B = −0.048, SE = 0.019, t (44) = −2.53, p = 0.015) and NGP/PA (ΔR2 = 0.159, F (1, 44) = 8.87, p = 0.005; B = −0.030, SE = 0.010, t (44) = −2.98, p = 0.005), but not for GP/PP (ΔR2 = 0.051, p = 0.11) or NGP/PP (ΔR2 = 0.062, p = 0.084).
0.18), prompting moderation analysis. This interaction improved the model for GP/PA (ΔR2 = 0.112, F (1, 44) = 6.38, p = 0.015; B = −0.048, SE = 0.019, t (44) = −2.53, p = 0.015) and NGP/PA (ΔR2 = 0.159, F (1, 44) = 8.87, p = 0.005; B = −0.030, SE = 0.010, t (44) = −2.98, p = 0.005), but not for GP/PP (ΔR2 = 0.051, p = 0.11) or NGP/PP (ΔR2 = 0.062, p = 0.084).
3.2. Time-domain analysis results
A four-way repeated-measures analysis of variance (ANOVA) with factors of 2 (Age group: older, younger) × 2 (Sentence type: GP, NGP) × 2 (Pause existence: PA, PP) × 5 (ROI: frontal, fronto-central, central, centro-parietal, parietal) was conducted on mean amplitudes for P200, N400 and P600 amplitudes. All post hoc pairwise comparisons were adjusted using the Bonferroni correction.
For the P200 component, no significant main effects were observed for any factors: Age group (F (1, 46) = 0.16, p = 0.90,
$ {\eta}_p^2= $
 0.001), Sentence type (F (1, 46) = 0.17, p = 0.68,
$ {\eta}_p^2= $
0.001), Sentence type (F (1, 46) = 0.17, p = 0.68,
$ {\eta}_p^2= $
 0.004), Pause existence (F (1, 46) = 2.23, p = 0.14,
$ {\eta}_p^2 $
0.004), Pause existence (F (1, 46) = 2.23, p = 0.14,
$ {\eta}_p^2 $
 = 0.05), or ROI (F (1.27, 58.44) = 1.08, p = 0.32,
$ {\eta}_p^2= $
= 0.05), or ROI (F (1.27, 58.44) = 1.08, p = 0.32,
$ {\eta}_p^2= $
 0.02). A significant Sentence type × ROI interaction was observed (F (1.39, 63.82) = 3.75, p = 0.044,
$ {\eta}_p^2= $
0.02). A significant Sentence type × ROI interaction was observed (F (1.39, 63.82) = 3.75, p = 0.044,
$ {\eta}_p^2= $
 0.075). At the three-way level, a significant Age group × Sentence type × ROI interaction also was observed (F (1.39, 63.82) = 7.46, p = 0.004,
$ {\eta}_p^2= $
0.075). At the three-way level, a significant Age group × Sentence type × ROI interaction also was observed (F (1.39, 63.82) = 7.46, p = 0.004,
$ {\eta}_p^2= $
 0.14). Simple effect analysis showed that GP elicited significantly smaller P200 mean amplitudes than NGP in the frontal region (F (1, 46) = 4.37, p = 0.042,
$ {\eta}_p^2= $
0.14). Simple effect analysis showed that GP elicited significantly smaller P200 mean amplitudes than NGP in the frontal region (F (1, 46) = 4.37, p = 0.042,
$ {\eta}_p^2= $
 0.09). Under the NGP condition, the mean amplitude of the P200 component elicited in older adults was significantly larger than that in younger adults over the frontal region (F(1, 46) = 6.61, p = 0.013,
$ {\eta}_p^2= $
0.09). Under the NGP condition, the mean amplitude of the P200 component elicited in older adults was significantly larger than that in younger adults over the frontal region (F(1, 46) = 6.61, p = 0.013,
$ {\eta}_p^2= $
 0.13); whereas it was significantly smaller than that in younger adults over the centro-parietal region (F (1, 46) = 6.35, p = 0.015,
$ {\eta}_p^2 $
0.13); whereas it was significantly smaller than that in younger adults over the centro-parietal region (F (1, 46) = 6.35, p = 0.015,
$ {\eta}_p^2 $
 = 0.12) and the parietal region (F (1, 46) = 11.10, p = 0.002,
$ {\eta}_p^2= $
= 0.12) and the parietal region (F (1, 46) = 11.10, p = 0.002,
$ {\eta}_p^2= $
 0.19). Within the older adult group, the mean P200 amplitude elicited by the NGP condition was significantly larger than that by the GP condition over the frontal region (F (1, 46) =10.82, p = 0.002,
$ {\eta}_p^2= $
0.19). Within the older adult group, the mean P200 amplitude elicited by the NGP condition was significantly larger than that by the GP condition over the frontal region (F (1, 46) =10.82, p = 0.002,
$ {\eta}_p^2= $
 0.19). Conversely, it was significantly smaller than that by the GP condition over the centro-parietal region (F (1, 46) = 6.62, p = 0.013,
$ {\eta}_p^2= $
0.19). Conversely, it was significantly smaller than that by the GP condition over the centro-parietal region (F (1, 46) = 6.62, p = 0.013,
$ {\eta}_p^2= $
 0.13) and the parietal region (F (1, 46) = 12.01, p = 0.001,
$ {\eta}_p^2= $
0.13) and the parietal region (F (1, 46) = 12.01, p = 0.001,
$ {\eta}_p^2= $
 0.21). No other interactions reached statistical significance.
0.21). No other interactions reached statistical significance.
For the N400 component, no significant main effect of Age group was observed (F (1, 46) = 0.44, p = 0.51,
$ {\eta}_p^2= $
 0.01). While a marginally non-significant main effect of Sentence type merged (F (1, 46) = 3.59, p = 0.065,
$ {\eta}_p^2= $
0.01). While a marginally non-significant main effect of Sentence type merged (F (1, 46) = 3.59, p = 0.065,
$ {\eta}_p^2= $
 0.07), a significant main effect of Pause existence was found (F (1, 46) = 9.00, p = 0.004,
$ {\eta}_p^2= $
0.07), a significant main effect of Pause existence was found (F (1, 46) = 9.00, p = 0.004,
$ {\eta}_p^2= $
 0.16), with post-hoc tests revealing larger N400 mean amplitudes in PP conditions compared to PA conditions. No significant main effect was observed for ROI (F (1.26, 57.73) = 0.32, p = 0.63,
$ {\eta}_p^2= $
0.16), with post-hoc tests revealing larger N400 mean amplitudes in PP conditions compared to PA conditions. No significant main effect was observed for ROI (F (1.26, 57.73) = 0.32, p = 0.63,
$ {\eta}_p^2= $
 0.01), and none of the interactions between factors reached statistical significance (all ps > 0.05).
0.01), and none of the interactions between factors reached statistical significance (all ps > 0.05).
Analysis of the P600 component revealed multiple significant main effects. First, older adults demonstrated significantly larger P600 amplitudes than younger adults (Age group: F (1, 46) = 5.33, p = 0.025,
$ {\eta}_p^2 $
 = 0.10). Additionally, GP sentences elicited stronger P600 responses compared to NGP (Sentence type: F (1, 46) = 4.15, p = 0.048,
$ {\eta}_p^2= $
= 0.10). Additionally, GP sentences elicited stronger P600 responses compared to NGP (Sentence type: F (1, 46) = 4.15, p = 0.048,
$ {\eta}_p^2= $
 0.08). Notably, pause conditions significantly modulated the component, with PP trials showing larger amplitudes than PA trials (Pause existence: F (1, 46) = 13.30, p < 0.001,
$ {\eta}_p^2 $
0.08). Notably, pause conditions significantly modulated the component, with PP trials showing larger amplitudes than PA trials (Pause existence: F (1, 46) = 13.30, p < 0.001,
$ {\eta}_p^2 $
 = 0.22). Finally, regional variations were observed, as indicated by a significant ROI effect F (1.36, 62.35) = 3.71, p = 0.046,
$ {\eta}_p^2= $
= 0.22). Finally, regional variations were observed, as indicated by a significant ROI effect F (1.36, 62.35) = 3.71, p = 0.046,
$ {\eta}_p^2= $
 0.08). The presence of these main effects, particularly those with modest effect sizes (
$ {\eta}_p^2 $
0.08). The presence of these main effects, particularly those with modest effect sizes (
$ {\eta}_p^2 $
 < 0.14, according to Cohen [Reference Cohen1992]), suggests that the contributions of these factors are not independent. This logically leads to the investigation of how these factors interact. Therefore, we next turn to the critical interaction effects, which provide a more nuanced account of the data and clarify the specific conditions under which the P600 modulation occurs. Significant interaction effects were also observed across multiple levels. For two-way interactions, ROI showed significant interactions with both Age group (F (1.36, 62.35) = 11.18, p < 0.001,
$ {\eta}_p^2= $
< 0.14, according to Cohen [Reference Cohen1992]), suggests that the contributions of these factors are not independent. This logically leads to the investigation of how these factors interact. Therefore, we next turn to the critical interaction effects, which provide a more nuanced account of the data and clarify the specific conditions under which the P600 modulation occurs. Significant interaction effects were also observed across multiple levels. For two-way interactions, ROI showed significant interactions with both Age group (F (1.36, 62.35) = 11.18, p < 0.001,
$ {\eta}_p^2= $
 0.20) and Pause existence (F (1.65, 75.78) = 10.19, p < 0.001,
$ {\eta}_p^2= $
0.20) and Pause existence (F (1.65, 75.78) = 10.19, p < 0.001,
$ {\eta}_p^2= $
 0.18), while Sentence type and Pause existence also interacted significantly (F (1, 46) = 9.28, p = 0.004,
$ {\eta}_p^2= $
0.18), while Sentence type and Pause existence also interacted significantly (F (1, 46) = 9.28, p = 0.004,
$ {\eta}_p^2= $
 0.17). At the three-way level, significant interactions emerged between Age group × ROI × Sentence type (F (1.63, 75.04) = 3.63, p = 0.04,
$ {\eta}_p^2= $
0.17). At the three-way level, significant interactions emerged between Age group × ROI × Sentence type (F (1.63, 75.04) = 3.63, p = 0.04,
$ {\eta}_p^2= $
 0.07) and ROI × Sentence type × Pause existence (F (1.60, 73.42) = 4.24, p = 0.026,
$ {\eta}_p^2= $
0.07) and ROI × Sentence type × Pause existence (F (1.60, 73.42) = 4.24, p = 0.026,
$ {\eta}_p^2= $
 0.08). Furthermore, a significant four-way interaction was detected involving all factors: Age group × ROI × Sentence type × Pause existence (Age group × ROI × Sentence type × Pause existence: F (1.60, 73.42) = 6.07, p = 0.006,
$ {\eta}_p^2= $
0.08). Furthermore, a significant four-way interaction was detected involving all factors: Age group × ROI × Sentence type × Pause existence (Age group × ROI × Sentence type × Pause existence: F (1.60, 73.42) = 6.07, p = 0.006,
$ {\eta}_p^2= $
 0.12). Although statistically significant, the main effects and higher-order interactions all yielded small effect sizes (as per conventional benchmarks), with the interactions mirroring the pattern seen in the main effects. Consequently, the presentation emphasizes the two-way interactions and the corresponding post-hoc tests.
0.12). Although statistically significant, the main effects and higher-order interactions all yielded small effect sizes (as per conventional benchmarks), with the interactions mirroring the pattern seen in the main effects. Consequently, the presentation emphasizes the two-way interactions and the corresponding post-hoc tests.
Simple effects analyses revealed that, significant age differences were observed in P600 amplitudes across prefrontal (PF), fronto-central (FC) and parietal (P) regions. Older adults exhibited significantly larger amplitudes than younger adults in both PF (F (1, 46) = 12.09, p = 0.001,
$ {\eta}_p^2= $
 0.21) and FC regions (F (1, 46) = 12.55, p < 0.001,
$ {\eta}_p^2= $
0.21) and FC regions (F (1, 46) = 12.55, p < 0.001,
$ {\eta}_p^2= $
 0.21), but showed significantly smaller amplitudes in the P region (F (1, 46) = 9.15, p = 0.004,
$ {\eta}_p^2= $
0.21), but showed significantly smaller amplitudes in the P region (F (1, 46) = 9.15, p = 0.004,
$ {\eta}_p^2= $
 0.17). Within the older adult group, significant ROI differences were found (F (4, 43) = 4.39, p = 0.005,
$ {\eta}_p^2= $
0.17). Within the older adult group, significant ROI differences were found (F (4, 43) = 4.39, p = 0.005,
$ {\eta}_p^2= $
 0.29). Specifically, centro-parietal (CP) amplitudes were significantly smaller than those in PF (p = 0.008), FC (p = 0.002) and central (C) regions (p = 0.003). Similarly, P amplitudes were significantly smaller than those in PF (p = 0.003), FC (p < 0.001), C (p = 0.002) and CP regions (p = 0.037). No such regional differences were observed in younger adults. Pause existence also significantly modulated P600 amplitudes across ROI. PP conditions elicited larger amplitudes than PA in PF (F (1, 46) = 12.18, p = 0.001,
$ {\eta}_p^2= $
0.29). Specifically, centro-parietal (CP) amplitudes were significantly smaller than those in PF (p = 0.008), FC (p = 0.002) and central (C) regions (p = 0.003). Similarly, P amplitudes were significantly smaller than those in PF (p = 0.003), FC (p < 0.001), C (p = 0.002) and CP regions (p = 0.037). No such regional differences were observed in younger adults. Pause existence also significantly modulated P600 amplitudes across ROI. PP conditions elicited larger amplitudes than PA in PF (F (1, 46) = 12.18, p = 0.001,
$ {\eta}_p^2= $
 0.21), FC (F (1, 46) = 22.97, p < 0.001,
$ {\eta}_p^2 $
0.21), FC (F (1, 46) = 22.97, p < 0.001,
$ {\eta}_p^2 $
 = 0.33) and C regions (F (1, 46) = 9.14, p = 0.004,
$ {\eta}_p^2= $
= 0.33) and C regions (F (1, 46) = 9.14, p = 0.004,
$ {\eta}_p^2= $
 0.17). Under PP conditions, significant regional differences emerged (F (4, 43) = 5.88, p < 0.001,
$ {\eta}_p^2= $
0.17). Under PP conditions, significant regional differences emerged (F (4, 43) = 5.88, p < 0.001,
$ {\eta}_p^2= $
 0.35). C amplitudes were significantly smaller than FC (p = 0.014), while CP amplitudes were smaller than PF (p = 0.014), FC (p < 0.001) and C (p = 0.002). P amplitudes were also smaller than PF (p = 0.002), FC (p < 0.001) and C (p < 0.001). Sentence-type effects were observed specifically in PP conditions, where GP elicited significantly larger P600 amplitudes than NGP (F (1, 46) = 13.82, p < 0.001,
$ {\eta}_p^2= $
0.35). C amplitudes were significantly smaller than FC (p = 0.014), while CP amplitudes were smaller than PF (p = 0.014), FC (p < 0.001) and C (p = 0.002). P amplitudes were also smaller than PF (p = 0.002), FC (p < 0.001) and C (p < 0.001). Sentence-type effects were observed specifically in PP conditions, where GP elicited significantly larger P600 amplitudes than NGP (F (1, 46) = 13.82, p < 0.001,
$ {\eta}_p^2= $
 0.23). Additionally, within GP sentences, PP conditions produced significantly larger amplitudes than PA (F (1, 46) = 24.80, p < 0.001,
$ {\eta}_p^2= $
0.23). Additionally, within GP sentences, PP conditions produced significantly larger amplitudes than PA (F (1, 46) = 24.80, p < 0.001,
$ {\eta}_p^2= $
 0.35).
0.35).
Follow-up analyses revealed complex interaction patterns through hierarchical simple effect analysis of the significant three-way (Age group × ROI × Sentence type; ROI × Sentence type × Pause existence) and four-way (Age group × ROI × Sentence type × Pause existence) interactions. The key findings demonstrated that age effects emerged in specific combinations of brain regions, sentence types and pause conditions.
Sentence-type effects varied significantly between age groups. Older adults showed larger P600 amplitudes for GP versus NGP in the PF (F (1, 46) = 19.72, p < 0.001,
$ {\eta}_p^2= $
 0.30) and FC (F (1, 46) = 12.25, p = 0.001,
$ {\eta}_p^2= $
0.30) and FC (F (1, 46) = 12.25, p = 0.001,
$ {\eta}_p^2= $
 0.21) regions during PP conditions. Pause effects also demonstrated age-specific patterns. For older adults, pause existence significantly enhanced amplitudes in the PF (F (1, 46) = 11.90, p = 0.001,
$ {\eta}_p^2= $
0.21) regions during PP conditions. Pause effects also demonstrated age-specific patterns. For older adults, pause existence significantly enhanced amplitudes in the PF (F (1, 46) = 11.90, p = 0.001,
$ {\eta}_p^2= $
 0.21) and FC (F (1, 46) = 10.95, p = 0.001,
$ {\eta}_p^2= $
0.21) and FC (F (1, 46) = 10.95, p = 0.001,
$ {\eta}_p^2= $
 0.19) regions during GP processing. Younger adults consistently demonstrated pause-related amplitude enhancements across multiple regions: for GP in prefrontal (F (1, 46) = 10.47, p = 0.002,
$ {\eta}_p^2 $
0.19) regions during GP processing. Younger adults consistently demonstrated pause-related amplitude enhancements across multiple regions: for GP in prefrontal (F (1, 46) = 10.47, p = 0.002,
$ {\eta}_p^2 $
 = 0.19), FC (F (1, 46) = 15.62, p < 0.001,
$ {\eta}_p^2= $
= 0.19), FC (F (1, 46) = 15.62, p < 0.001,
$ {\eta}_p^2= $
 0.25) and C (F (1, 46) = 9.85, p = 0.003,
$ {\eta}_p^2= $
0.25) and C (F (1, 46) = 9.85, p = 0.003,
$ {\eta}_p^2= $
 0.18) regions. The P region in younger adults showed enhanced amplitudes in GP-PP condition (F (1, 46) = 10.66, p = 0.002,
$ {\eta}_p^2= $
0.18) regions. The P region in younger adults showed enhanced amplitudes in GP-PP condition (F (1, 46) = 10.66, p = 0.002,
$ {\eta}_p^2= $
 0.19).
0.19).
Finally, ROI differences emerged in specific Age groups, Sentence types and Pause existence combinations. For older adults processing GP with PP, significant ROI variations were observed (F (4, 43) = 7.23, p < 0.001,
$ {\eta}_p^2 $
 = 0.40), with the PF region showing smaller amplitudes than central (p = 0.009), CP (p < 0.001) and P (p < 0.001) regions. The FC region was smaller than C (p = 0.003), while the C region was smaller than CP (p = 0.001), which in turn was smaller than P (p = 0.033). During NGP-PA condition processing, older adults showed smaller amplitudes in FC compared to PF regions (F (4, 43) = 2.70, p = 0.043,
$ {\eta}_p^2= $
= 0.40), with the PF region showing smaller amplitudes than central (p = 0.009), CP (p < 0.001) and P (p < 0.001) regions. The FC region was smaller than C (p = 0.003), while the C region was smaller than CP (p = 0.001), which in turn was smaller than P (p = 0.033). During NGP-PA condition processing, older adults showed smaller amplitudes in FC compared to PF regions (F (4, 43) = 2.70, p = 0.043,
$ {\eta}_p^2= $
 0.20). Younger adults exhibited no significant ROI differences across conditions.
0.20). Younger adults exhibited no significant ROI differences across conditions.
Similarly, ANCOVAs adjusting for YoE were conducted for the ERP components P200, N400 and P600. The YoE × Age group interaction was not significant for all components (P200: F (1, 44) = 0.06, p = 0.81,
$ {\eta}_p^2= $
 0.001; N400: F (1, 44) = 0.75, p = 0.39,
$ {\eta}_p^2= $
0.001; N400: F (1, 44) = 0.75, p = 0.39,
$ {\eta}_p^2= $
 0.02; P600: F (1, 44) = 0.30, p = 0.59,
$ {\eta}_p^2= $
0.02; P600: F (1, 44) = 0.30, p = 0.59,
$ {\eta}_p^2= $
 0.01), justifying standard ANCOVAs without the interaction term. For the P200, the main effect of YoE (F (1, 45) = 0.001, p = 0.97,
$ {\eta}_p^2 $
0.01), justifying standard ANCOVAs without the interaction term. For the P200, the main effect of YoE (F (1, 45) = 0.001, p = 0.97,
$ {\eta}_p^2 $
 < 0.001) and Age group (F (1, 45) = 0.01, p = 0.91,
$ {\eta}_p^2 $
< 0.001) and Age group (F (1, 45) = 0.01, p = 0.91,
$ {\eta}_p^2 $
 < 0.001) were not significant. For the N400, the Age group main effect was not significant (F (1, 45) = 0.87, p = 0.36,
$ {\eta}_p^2 $
< 0.001) were not significant. For the N400, the Age group main effect was not significant (F (1, 45) = 0.87, p = 0.36,
$ {\eta}_p^2 $
 = 0.02), but a significant main effect of YoE was found (F (1, 45) = 4.83, p = 0.033,
$ {\eta}_p^2 $
= 0.02), but a significant main effect of YoE was found (F (1, 45) = 4.83, p = 0.033,
$ {\eta}_p^2 $
 = 0.10). For the P600, main effects were not significant (YoE: F (1, 45) = 1.33, p = 0.26,
$ {\eta}_p^2 $
= 0.10). For the P600, main effects were not significant (YoE: F (1, 45) = 1.33, p = 0.26,
$ {\eta}_p^2 $
 = 0.03; Age group: F (1, 45) = 0.96, p = 0.33,
$ {\eta}_p^2 $
= 0.03; Age group: F (1, 45) = 0.96, p = 0.33,
$ {\eta}_p^2 $
 = 0.02). However, regression analysis revealed that the regression coefficients for YoE were not significant across conditions for the N400 (all ps > 0.05). Pearson correlation analysis revealed a significant positive correlation between YoE and age (r = 0.66, p < 0.001) (Figures 2–5).
= 0.02). However, regression analysis revealed that the regression coefficients for YoE were not significant across conditions for the N400 (all ps > 0.05). Pearson correlation analysis revealed a significant positive correlation between YoE and age (r = 0.66, p < 0.001) (Figures 2–5).
ERP waveforms and topographic maps elicited by garden-path sentences in older adults.
Note: GP, garden-path sentences; PA, pause-absent; PP, pause-present.

Figure 2. Long description
From top left, five line graphs show E R P amplitude in microvolts versus latency in milliseconds for Frontal, Fronto-central, Central, Centro-parietal, and Parietal regions. Each graph has two lines: blue for G P dash P A and orange for G P dash P P. Shaded vertical bands mark P200, N400, and P600 time windows. In all regions, G P dash P P shows larger negative deflections in the N400 window and reduced positive deflections in the P600 window compared to G P dash P A. The three rightmost panels are topographic scalp maps for P200, N400, and P600, with color bars indicating amplitude from negative (blue) to positive (red) microvolts. P200 shows a strong positive maximum at the vertex, N400 shows a central negativity, and P600 shows a parietal positivity.
ERP waveforms and topographic maps elicited by NGP sentences in older adults.
Note: NGP, non-garden-path sentences; PA, pause-absent; PP, pause-present.

Figure 3. Long description
From top left, the first five panels are line graphs labeled Frontal, Fronto-central, Central, Centro-parietal, and Parietal. Each graph has amplitude in microvolts on the y-axis and latency in milliseconds on the x-axis, with vertical colored bands marking P200 (orange, 200 ms), N400 (green, 400 ms), and P600 (purple, 600 ms) time windows. Two lines are plotted: blue for N G P dash P A and red for N G P dash P P, as indicated in the legends. In all regions, N G P dash P P shows larger negative deflections at N400 and more positive deflections at P600 compared to N G P dash P A. The bottom right contains three circular scalp maps labeled P200, N400, and P600, each with a color bar from blue (negative) to red (positive) microvolts. The P200 map shows a central maximum, N400 shows a centro-parietal negativity, and P600 shows a parietal positivity. These maps illustrate the spatial distribution of E R P effects across the scalp for each time window.
ERP waveforms and topographic maps elicited by GP sentences in younger adults.
Note: GP, garden-path sentences; PA, pause-absent; PP, pause-present.

Figure 4. Long description
Starting at the top left, the Frontal panel shows a line graph with amplitude in microvolts on the y-axis and latency in milliseconds on the x-axis, comparing G P dash P A and G P dash P P conditions. Colored vertical bands highlight P200, N400, and P600 intervals. The Fronto-central and Central panels to the right follow the same format, each showing distinct waveform differences between the two conditions, especially in the P600 window where G P dash P P shows a higher positive peak. The bottom row begins with the Centro-parietal and Parietal panels, also plotting amplitude versus latency for both conditions, with similar interval highlights. To the right, three circular topographic scalp maps are stacked vertically, labeled P200, N400, and P600. Each map uses a color scale from blue (low amplitude) to red (high amplitude) to show the spatial distribution of E R P amplitudes across the scalp, with P200 showing a central maximum, N400 a more diffuse pattern, and P600 a posterior maximum. Legends and axis labels are consistent across all panels.
ERP waveforms and topographic maps elicited by non-garden-path sentences in younger adults.
Note: NGP, non-garden-path sentences; PA, pause-absent; PP, pause-present.

Figure 5. Long description
Starting at the top left, the first five panels are line graphs labeled Frontal, Fronto-central, Central, Centro-parietal, and Parietal. Each graph has latency in milliseconds on the x-axis and amplitude in microvolts on the y-axis. Two lines are plotted: N G P dash P A (blue) and N G P dash P P (orange). Vertical shaded bands mark P200, N400, and P600 time windows. In all regions, the N G P dash P P condition shows higher positive amplitudes, especially during the P600 window. The bottom right contains three topographic scalp maps for P200, N400, and P600, with color bars ranging from negative (blue) to positive (red) amplitudes. The P600 map shows the strongest positive activity in parietal regions.
ERSP at FZ channel in older adults and younger adults.
Note: GP, garden-path sentences; NGP, non-garden-path sentences; PA, pause-absent; PP, pause-present. θ-band, 4–8 Hz; α-band, 10–14 Hz.

Figure 6. Long description
The grid contains eight panels. The top row shows older adults, the bottom row shows younger adults. Each column is labeled at the top: G P dash P A, G P dash P P, N G P dash P A, N G P dash P P. Each panel is a heatmap with x axis labeled Times in seconds from 0 to 0.8, y axis labeled Frequency in hertz from 0 to 30. A vertical color bar at the right of each panel ranges from negative 0.5 to 1.5, with blue for low values and red for high values. In all panels, red bands appear between 4 and 14 hertz, corresponding to theta and alpha bands, with the most intense red in G P dash P P and N G P dash P P for both age groups. Blue regions dominate higher frequencies. The patterns are similar across age groups, but younger adults show slightly more pronounced red bands in the pause-present conditions. The spatial distribution of color indicates stronger E R S P responses in the theta and alpha bands during pause-present conditions, especially for garden-path sentences.
ERSP at FCZ channel in older adults and younger adults.
Note: GP, garden-path sentences; NGP, non-garden-path sentences; PA, pause-absent; PP, pause-present. θ-band, 4–8 Hz; α-band,10–14 Hz.

Figure 7. Long description
There are eight panels arranged in two rows and four columns. The top row shows older adults, the bottom row shows younger adults. Each column represents a condition: GP-PA, GP-PP, NGP-PA, NGP-PP. X-axes are labeled Times (s) from 0 to 0.8, y-axes are labeled Frequency (Hz) from 5 to 30. Each panel uses a color scale from -0.5 (blue) to 1.5 (red) to indicate ERSP values. In both age groups, PP conditions (second and fourth columns) show strong red bands between 4 and 14 Hz, especially in the theta and alpha bands, while PA conditions (first and third columns) show weaker or more diffuse activity. The effect is more pronounced in younger adults, with higher intensity and broader frequency range in the PP panels. All panels show time-locked activity starting near 0.1 seconds, with the strongest responses in the GP-PP and NGP-PP conditions.
3.3. Time-frequency analysis results
For θ-band and α-band mean power, separate 2 (Age group: older, younger) × 2 (Sentence type: GP, NGP) × 2 (Pause existence: PA, PP) × 5 (ROI: prefrontal, fronto-central, central, centro-parietal, parietal) four-way repeated-measures ANOVAs were conducted. Bonferroni correction was used for all post hoc pairwise comparisons.
Analysis of θ-band activity revealed a significant main effect of Age group (F (1, 46) = 5.28, p = 0.026,
$ {\eta}_p^2 $
 = 0.10), indicating that older adults exhibited significantly lower θ-band power compared to younger adults. However, no other significant main effects were observed, with non-significant results for Sentence type (F (1, 46) = 0.03, p = 0.87,
$ {\eta}_p^2 $
= 0.10), indicating that older adults exhibited significantly lower θ-band power compared to younger adults. However, no other significant main effects were observed, with non-significant results for Sentence type (F (1, 46) = 0.03, p = 0.87,
$ {\eta}_p^2 $
 = 0.001), Pause existence (F (1, 46) = 1.16, p = 0.29,
$ {\eta}_p^2 $
= 0.001), Pause existence (F (1, 46) = 1.16, p = 0.29,
$ {\eta}_p^2 $
 = 0.03) and ROI (F (1.20, 54.95) = 2.86, p = 0.09,
$ {\eta}_p^2 $
= 0.03) and ROI (F (1.20, 54.95) = 2.86, p = 0.09,
$ {\eta}_p^2 $
 = 0.06). A significant Age group × Sentence type interaction emerged (F (1, 46) = 4.08, p = 0.049,
$ {\eta}_p^2 $
= 0.06). A significant Age group × Sentence type interaction emerged (F (1, 46) = 4.08, p = 0.049,
$ {\eta}_p^2 $
 = 0.08). Simple effects analysis showed that for GP sentences, older adults had significantly lower θ-band power than younger adults (F (1, 46) = 10.85, p = 0.002,
$ {\eta}_p^2 $
= 0.08). Simple effects analysis showed that for GP sentences, older adults had significantly lower θ-band power than younger adults (F (1, 46) = 10.85, p = 0.002,
$ {\eta}_p^2 $
 = 0.19). No other significant interactions were observed.
= 0.19). No other significant interactions were observed.
The α-band analysis demonstrated distinct age-related differences, with older adults displaying significantly reduced power relative to younger adults (Age group: F (1, 46) = 6.00, p = 0.018,
$ {\eta}_p^2 $
 = 0.12). The remaining factors failed to reach statistical significance, as evidenced by Sentence type (F (1, 46) = 0.10, p = 0.81,
$ {\eta}_p^2 $
= 0.12). The remaining factors failed to reach statistical significance, as evidenced by Sentence type (F (1, 46) = 0.10, p = 0.81,
$ {\eta}_p^2 $
 = 0.001), Pause existence (F (1, 46) = 1.89, p = 0.18,
$ {\eta}_p^2 $
= 0.001), Pause existence (F (1, 46) = 1.89, p = 0.18,
$ {\eta}_p^2 $
 = 0.04) and ROI (F (1.53, 70.18) = 2.88, p = 0.077,
$ {\eta}_p^2 $
= 0.04) and ROI (F (1.53, 70.18) = 2.88, p = 0.077,
$ {\eta}_p^2 $
 = 0.06) all showing non-significant effects. A significant Age group × Sentence type interaction was obviously found (F (1, 46) = 4.78, p = 0.034,
$ {\eta}_p^2 $
= 0.06) all showing non-significant effects. A significant Age group × Sentence type interaction was obviously found (F (1, 46) = 4.78, p = 0.034,
$ {\eta}_p^2 $
 = 0.09). Simple effects analysis showed that for GP sentences, older adults had significantly lower α-band power than younger adults (F (1, 46) = 10.04, p = 0.003,
$ {\eta}_p^2 $
= 0.09). Simple effects analysis showed that for GP sentences, older adults had significantly lower α-band power than younger adults (F (1, 46) = 10.04, p = 0.003,
$ {\eta}_p^2 $
 = 0.18). No other significant interactions were observed.
= 0.18). No other significant interactions were observed.
To account for YoE’s confounding effect on θ/α-band group differences, an ANCOVA was conducted. The YoE × Age group interaction was not significant for both bands and removed from the final model. In θ-band, neither YoE(F (1, 45) = 0.53, p = 0.47,
$ {\eta}_p^2 $
 = 0.01) nor Age group (F (1, 45) = 1.53, p = 0.22,
$ {\eta}_p^2 $
= 0.01) nor Age group (F (1, 45) = 1.53, p = 0.22,
$ {\eta}_p^2 $
 = 0.03) had significant main effects. In α-band, YoE (F (1, 45) = 0.02, p = 0.90,
$ {\eta}_p^2 $
= 0.03) had significant main effects. In α-band, YoE (F (1, 45) = 0.02, p = 0.90,
$ {\eta}_p^2 $
 < 0.001) and Age group (F (1, 45) = 3.02, p = 0.09,
$ {\eta}_p^2 $
< 0.001) and Age group (F (1, 45) = 3.02, p = 0.09,
$ {\eta}_p^2 $
 = 0.06) main effects were also non-significant (Figures 6 and 7).
= 0.06) main effects were also non-significant (Figures 6 and 7).
4. Discussion
This study systematically investigated the age-specific regulatory effects of pauses on syntactic processing of Chinese GP sentences in older adults by comparing behavioural and neurophysiological indicators across age groups. Behavioural results revealed significant age effects in sentence processing among older adults, with pause-induced facilitation of syntactic processing demonstrating age specificity. Pauses significantly reduced RTs in older adults during GP sentence processing, while no similar effect was observed in younger adults. The improvement in ACC represented a cross-age commonality, as pauses significantly enhanced judgment ACC for GP sentences without being modulated by age factors.
At the neural mechanism level, time-domain analysis demonstrated that P600 component activation in older adults was more strongly modulated by syntactic structure and pause conditions, exhibiting significant regional specificity. Older adults showed significantly larger P600 amplitudes than younger adults in prefrontal and fronto-central regions under PP conditions. In contrast, N400 component modulation by pauses displayed age-independent and syntax-general characteristics, with pauses significantly enhancing N400 amplitudes across different syntactic complexity conditions for both groups. No significant modulation of P200 components by experimental factors was observed, though GP sentences elicited significantly reduced P200 amplitudes in prefrontal regions. Time-frequency analysis indicated that older adults exhibited significantly lower θ-band and α-band power than younger adults during GP sentence processing, suggesting age-related alterations in neural oscillatory patterns. More specifically, between-group comparisons revealed significant correlations between α-band power and syntactic complexity, with GP sentences attenuating θ-band and α-band ERSP responses in older adults; however, no direct neural oscillatory evidence was found to indicate that pauses influence syntactic processing in older adults. Additionally, ANCOVA incorporating YoE revealed that YoE only affected partial behavioural ACC (GP/PP and NGP/PP conditions for non-pause sentences) and the N400 component. Collectively, based on behavioural results and time-domain ERP analysis, these findings demonstrate that Chinese older adults effectively utilize pauses to regulate ambiguous complex syntactic structures, with pauses accelerating their syntactic processing while eliciting a prefrontal-dominant neural activation pattern.
4.1. Age differences in syntax processing under speech pause modulation
This comparative study between older and younger adults reveals the dynamic characteristics of ageing mechanisms in syntactic processing. The results demonstrate significant differences between older and younger adults in both behavioural and electrophysiological measures, suggesting that ageing effects influence language abilities related to syntactic processing in older adults. While older adults showed significantly longer RTs overall compared to younger adults, no significant age-related differences emerged in ACC rates. This pattern indicates that older adults compensate for age-related declines by extending processing time to maintain syntactic judgment ACC, consistent with the explanation of the processing speed theory (PST) (Salthouse, Reference Salthouse1996). According to PST, the slowing of cognitive operations with age affects the efficiency of syntactic information processing (Albinet et al., Reference Albinet, Boucard, Bouquet and Audiffren2012). We posit that this behavioural pattern suggests that older adults employ temporal compensatory mechanisms to maintain syntactic processing ACC. This aligns with the core premise of the PST, which posits that a generalized slowing of central processing speed means older adults require more time to execute each cognitive operation – such as lexical retrieval, syntactic structure building and ambiguity resolution (Salthouse, Reference Salthouse1996). Moreover, the ACC data indicate that this increased processing time, or reduced speed, does not necessarily lead to a decline in syntactic processing ability. Instead, by prolonging processing time – whether proactively or reactively – older adults effectively compensate for their decreased processing efficiency per unit time, thereby striving to achieve final performance levels comparable to those of younger adults. Consequently, these findings not only reflect the principles of PST but also provide direct behavioural evidence for cognitive compensation.
Furthermore, this processing strategy shows a close association with speech pauses, as evidenced by a significant interaction between pause and age. The facilitative effect of pauses on RTs demonstrates age-specific characteristics, particularly manifested in older adults showing significantly reduced RTs for GP sentences under PP conditions, while younger adults exhibited no such effect. The cognitive reserve theory suggests that older adults rely more on extra cues to optimize cognitive function (Stern, Reference Stern2009). As working memory and processing speed decline with age in older adults (De Bot et al., Reference De Bot, Plejert, Simonsen, Fyndanis, Hansen, Norvik, Svendsen and Svennevig2020), they tend to rely more heavily on contextual information and semantic cues as external supports during syntactic processing (De Bot et al., Reference De Bot, Plejert, Simonsen, Fyndanis, Hansen, Norvik, Svendsen and Svennevig2020; Ryskin & Nieuwland, Reference Ryskin and Nieuwland2023). We propose that pauses facilitate older adults’ processing of complex syntactic structures (e.g., GP sentences) by providing additional processing time (Brennan & Schober, Reference Brennan and Schober2001) or by marking syntactic boundaries where ambiguity arises (Bailey & Ferreira, Reference Bailey and Ferreira2003). This offers older adults either extended processing duration or clearer positional cues for syntactic reanalysis. Furthermore, older adults demonstrate heightened sensitivity to external cues, enabling them to actively utilize prosodic information to compensate for age-related declines in cognitive resources. In contrast, since younger adults do not yet require external cues to maintain normal syntactic processing abilities, no significant regulatory effect of pauses on GP sentence processing was observed in this group.
Neurophysiological evidence reveals distinct compensatory mechanisms in older adults’ pause-modulated syntactic processing. The significant age-related main effect observed in the P600 component, with older adults exhibiting notably stronger activation in the prefrontal region under PP condition during GP sentence processing compared to younger adults, suggests two plausible explanations: pauses may either facilitate initial syntactic processing by eliciting P600 activity in older adults, or they could potentially disrupt their syntactic processing. However, considering the behavioural results demonstrating that pauses significantly improved both ACC rates and RTs in older adults, the enhanced P600 activity is more likely to be triggered by the facilitative effect of pauses on syntactic processing in this population. This suggests that older adults require greater mobilization of prefrontal resources for reanalysis of complex syntactic structures. In contrast, older adults showed significantly smaller P600 amplitudes in parietal regions compared to younger adults, along with marked suppression of θ-band and α-band neural oscillations. Considering the late activation pattern of P600, these findings indicate that older adults need more time for neural synchronization to maintain syntactic integration capacity. This ‘prefrontal enhancement-parietal suppression’ activation pattern aligns with the posterior–anterior shift in ageing (PASA) phenomenon (Davis et al., Reference Davis, Dennis, Daselaar, Fleck and Cabeza2008). This phenomenon reflects a functional reallocation of cognitive resources to address processing inefficiencies in posterior brain regions (e.g., parietal lobe): by enhancing recruitment of prefrontal executive control resources, the brain compensates for declines in the posterior sensory integration network, thereby maintaining behavioural performance. This indicates that older adults undergo functional reorganization when processing conditions of high syntactic complexity, utilizing alternative neural resources for compensation (Cabeza & Dennis, Reference Cabeza and Dennis2012). The observed P600 activation in prefrontal regions in the study may be associated with the prefrontal cognitive control system enhancing goal-directed syntactic reanalysis and integration of syntactic ambiguity (Alekseeva et al., Reference Alekseeva, Myachykov, Bermudez-Margaretto and Shtyrov2022). Conversely, the reduced P600 amplitude in parietal regions, combined with the generalized attenuation of θ and α band power in time-frequency characteristics, suggests age-related deficits in the neuroelectrophysiological activity of syntactic integration within the parietal lobe and associated networks. Together, these findings constitute multimodal evidence for cognitive ageing in older adults. Although no direct pause-related effects on θ and α band power were observed, we speculate that the enhanced P600 in the prefrontal region of the elderly group may be attributed not only to the high syntactic complexity consuming greater cognitive resources but also to potential influences of prosodic features – such as pauses – on resource allocation during syntactic processing. This may reflect a distinct or more ‘economical’ neural processing strategy. Future research should further investigate the electrophysiological relationship between the presence of pauses and cognitive ageing to explore whether older adults effectively utilize prosodic cues to facilitate syntactic processing as a cognitive strategy. Additionally, we adjusted for YoE to account for its potential confounding effect among older adults. The results showed that YoE significantly affected only behavioural ACC (non-pause sentences: GP/PP, NGP/PP conditions) and the N400, demonstrating semantic processing differences to education and cognitive reserve (Milburn et al., Reference Milburn, Dickey, Warren and Hayes2023; Montemurro et al., Reference Montemurro, Jarema and Mondini2021; Xu et al., Reference Xu, Chen, Yang and Zhu2020). The regression and correlation analyses of N400 further indicated that YoE and age exhibit collinearity. In contrast, YoE had no significant effect on the P600 or pause facilitation, demonstrating robust syntactic processing in older adults (Charles et al., Reference Charles, Stephen, Robert, Noam and Johan2017) and that education modulates ageing semantic, not syntactic, language processing.
4.2. The facilitation effect of pauses on syntactic processing in older adults
Within-group data from older adults demonstrated that pauses significantly facilitate the processing of Chinese GP sentences in healthy older adults. In PA conditions, older adults showed markedly prolonged RTs for GP sentences compared to NGP sentences. However, when pause segments were inserted, the RT difference between these two sentence types was significantly reduced while maintaining comparable ACC levels.
This finding aligns with Howell and Young (Reference Howell and Young1991), suggesting that pauses may improve older adults’ syntactic processing performance by delaying the processing of subsequent content and providing additional time for syntactic integration, thereby alleviating working memory constraints on syntactic reanalysis (Samu et al., Reference Samu, Campbell, Tsvetanov, Shafto and Tyler2017). This mechanism shows functional similarity with how AD patients utilize prolonged pauses to compensate for memory impairments (Pistono et al., Reference Pistono, Jucla, Barbeau, Saint-Aubert, Lemesle, Calvet, Köpke, Puel and Pariente2016), implying that regardless of cognitive status, older adults may preferentially employ strategies that optimize cognitive resource allocation to compensate for syntactic processing difficulties.
Notably, pause conditions did not significantly affect the processing of NGP sentences. This indicates that the regulatory effect of pauses on older adults’ syntactic processing is more sensitive to complex syntactic structures, providing no significant benefit for simpler structures. These findings are consistent with conclusions from Obler et al. (Reference Obler, Fein, Nicholas and Albert1991) and Waters and Caplan (Reference Waters and Caplan2001), showing that older adults’ syntactic processing abilities decline progressively with increasing syntactic complexity. Therefore, for NGP sentences, older adults can demonstrate relatively good processing ability without relying on pause elements. These findings suggest that the facilitation effect of pauses is specifically targeted at complex syntactic structures that challenge older adults’ processing capacity, rather than being a general enhancement of all syntactic processing.
4.3. Specific neural mechanisms of syntactic processing in older adults
The time-domain analysis revealed unique neural mechanisms underlying Chinese older adults’ syntactic processing. Building upon the discussion in Section 4.1 and considering within-group results from older adults, the significantly larger P600 amplitudes elicited by GP sentences compared to NGP sentences – with pauses further enhancing this difference in prefrontal and fronto-central regions – suggests that healthy Chinese older adults can normally activate syntactic reanalysis processes when handling complex syntactic structures and ambiguous content (Friederici, Reference Friederici2011; Frisch et al., Reference Frisch, Schlesewsky, Saddy and Alpermann2002; Hao et al., Reference Hao, Duan and Zhang2021). This finding contrasts with Kutas and Federmeier (Reference Kutas and Federmeier2011) conclusions, indicating that healthy Chinese older adults do not consequently reinforce initial erroneous analyses. While Indo-European language studies suggest prefrontal activation is compromised by ageing (Samu et al., Reference Samu, Campbell, Tsvetanov, Shafto and Tyler2017), the P600 activation observed in our study demonstrates that Chinese older adults can utilize the additional time provided by pauses to more effectively regulate prefrontal resources, highlighting typical language-specific characteristics in how Chinese populations employ prosodic cues compared to Indo-European language speakers.
The N400 amplitude patterns in GP sentences also warrant attention. The lack of significant influence from syntactic complexity on N400 activation contradicts classical findings (Corley et al., Reference Corley, MacGregor and Donaldson2007; MacGregor et al., Reference MacGregor, Corley and Donaldson2009; Maxfield et al., Reference Maxfield, Lyon and Silliman2009). In our study, the N400 component showed no enhanced activation at temporary ambiguity positions in GP sentences, suggesting that at the semantic level, Chinese older adults do not persist in integrating initially incorrect semantic relationships into subsequent sentence processing. However, pauses elicited larger N400 amplitudes, indicating their regulatory role in facilitating proper syntactic processing and semantic comprehension in Chinese older adults. Speech pauses enhanced processing capabilities at both semantic and syntactic levels. Since pauses can maintain prosodic structure during syntactic processing (Kircher et al., Reference Kircher, Brammer, Levelt, Bartels and McGuire2004), Chinese older adults appear to preferentially use prosodic cues for early syntactic prediction, thereby somewhat mitigating semantic conflicts in later reanalysis stages. We speculate that the significantly reduced power in the θ and α bands for GP sentences observed in the time-frequency results may be explained by the possibility that older adults rely more on prosodic structure and pauses to adjust processing. This would involve increasing the weight of syntactic prediction to compensate for the decline in online syntactic integration ability (Zioga et al., Reference Zioga, Weissbart, Lewis, Haegens and Martin2023), while leveraging the facilitative effect of pauses on syntactic processing, thereby mitigating the cognitive load induced by syntactic violations (Pérez et al., Reference Pérez, Molinaro, Mancini, Barraza and Carreiras2012).
The P200 component is closely associated with phonological information extraction (de Diego Balaguer et al., Reference de Diego Balaguer, Toro, Rodriguez-Fornells and Bachoud-Levi2007) and primarily distributed in fronto-central and parieto-occipital regions (Kong et al., Reference Kong, Zhang, Zhang and Kang2012; Landi & Perfetti, Reference Landi and Perfetti2007; Liu et al., Reference Liu, Perfetti and Hart2003). Although P200 showed no statistically significant relationships with age, syntactic complexity, or pauses in its activation patterns, the observed significant reduction in average amplitude for GP sentences in prefrontal regions suggests that regardless of age, increased syntactic complexity can inhibit auditory processing (Shin et al., Reference Shin, Noh, Park and Sung2023) and affect higher-order cognitive resource regulation processes. Furthermore, this indicates that older adults can effectively maintain late-stage information integration in higher-order cognitive processes, specifically manifested in correctly extracting prosodic-acoustic cues provided by pauses and optimizing attention allocation in fronto-central and parietal regions (Novak et al., Reference Novak, Ritter and Vaughan1992), thereby assisting cognitive resource compensation strategies for late syntactic processing.
Additionally, this study provides some cross-linguistic perspective on the syntactic regulatory mechanisms of prosodic boundary markers (pauses). By inserting pauses before disambiguating elements in Chinese GP sentences, we artificially segmented the prosodic structure at the prosodic level to strengthen the boundary-marking function of pauses. Comparing with the related study in Spanish (Martínez-Sánchez et al., Reference Martínez-Sánchez, Meilán, Vera-Ferrandiz, Carro, Pujante-Valverde, Ivanova and Carcavilla2017), we found that pauses in Chinese simultaneously serve dual functions of marking both syntactic and prosodic boundaries, with artificial prosodic segmentation not affecting pauses’ regulatory role in syntactic processing. The synchronized enhancement of P600 components in older adults’ prefrontal and fronto-central regions demonstrates that Chinese pauses can improve syntactic reanalysis by redistributing cognitive resources. In contrast, the significant enhancement of pause-induced N400 components indicates that older adults can reduce reliance on initial erroneous analyses, thereby overcoming interference from temporary ambiguous structures at both sentential and semantic levels. Pauses in Chinese play a more active role in syntactic analysis and regulatory mechanisms.
5. Conclusion
Older adults demonstrated significant age-related effects in linguistic abilities, yet they could compensate for natural cognitive ageing through pauses and cognitive resource reallocation. Building upon ageing effects, within-group analyses revealed that GP sentences showed marked facilitation from pauses, while NGP sentences remained unaffected by pause manipulation. YoE was included as a covariate, and results showed it influenced age-group differences in semantic measures but not others. This demonstrates that pauses exhibit distinct sensitivity to syntactic complexity, enhancing older adults’ complex syntactic processing by providing additional time for cognitive resource allocation.
Pauses facilitated increased P600 activation in prefrontal and fronto-central regions, indicating that Chinese older adults could effectively suppress initial erroneous analyses while strengthening late-stage correct syntactic processing. The elicitation of larger N400 mean amplitudes by pauses suggests Chinese older adults preferentially utilize such prosodic cues for early syntactic prediction, thereby mitigating semantic conflicts during later processing stages. These regulatory effects of pauses on syntactic processing in older adults demonstrate unique cross-linguistic characteristics. Unlike findings in Indo-European languages or Arabic, pauses in Chinese do not inhibit proper syntactic reanalysis but instead play a more active role in syntactic analysis and regulation.
This study has several limitations: First, it primarily examined pause effects on offline syntactic processing without extending to natural speech production contexts. Future research should investigate multimodal regulatory mechanisms in naturalistic settings. Second, the sensitivity of pauses to syntactic complexity gradients requires further validation, as the current study only employed two sentence types. Subsequent studies could incorporate functional magnetic resonance imaging (fMRI) to precisely localize key brain regions involved in pause-modulated syntactic processing, enabling comprehensive exploration of temporal–spatial neural mechanisms underlying older adults’ use of pauses for syntactic regulation and deeper investigation of syntax-pause interactions. Third, demographic characteristics should be further controlled to avoid confounding effects of education attainment, such as vocational skills training, educational quality, or lifelong learning activities.
Data availability statement
The data that support the findings are available at https://doi.org/10.17605/OSF.IO/C2FNB.
Acknowledgments
This work was supported by grants from the China National Social Sciences Funding (No. 23AYY013), Jiangsu Major Research Projects in Philosophy and Social Sciences at Higher Education Institutions (No. 2023SJZD110).
Author contribution
JC: Data collection, Software, Methodology, Writing-original draft, Writing-review & editing. XC: Data collection, Software, Methodology. YYM: Data collection, Software, Methodology. YJM: Data collection, Software. XP: Data collection, Software. SF: Funding acquisition, Formal Analysis, Methodology, Resources, Supervision, Writing-original draft, Writing-review & editing.
Competing interests
The authors declare no competing interests.
Ethical approval
This study received ethical approval from the Special Committee on Research Ethics of the Academic Committee at Jiangsu Normal University (Approval No.: JSNU-H2025XM-001) in January 2025. It was conducted in strict compliance with international ethical guidelines, including the Declaration of Helsinki. This ensured the standards of participant protection and research integrity.
Informed consent
Prior to the experiment, the principal investigator provided all participants (24 elderly individuals aged over 60 and 24 undergraduate students) with written informed consent.

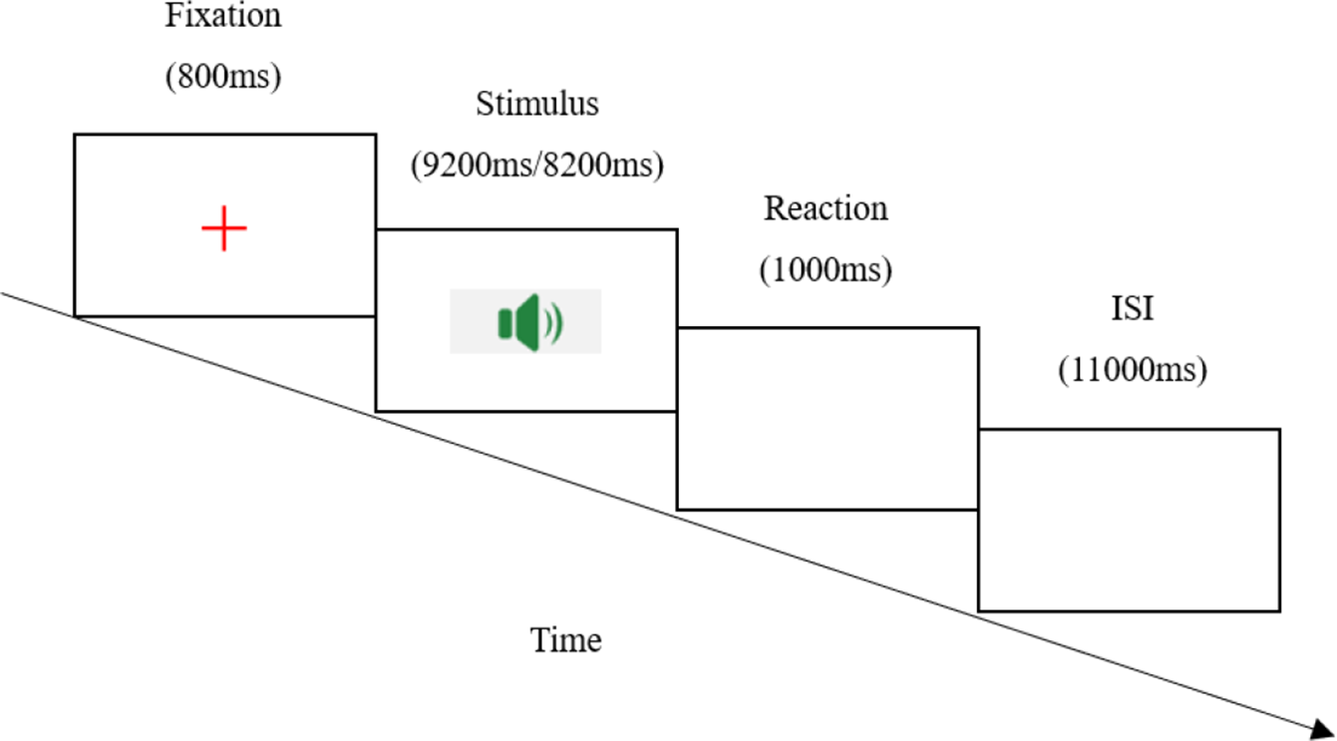
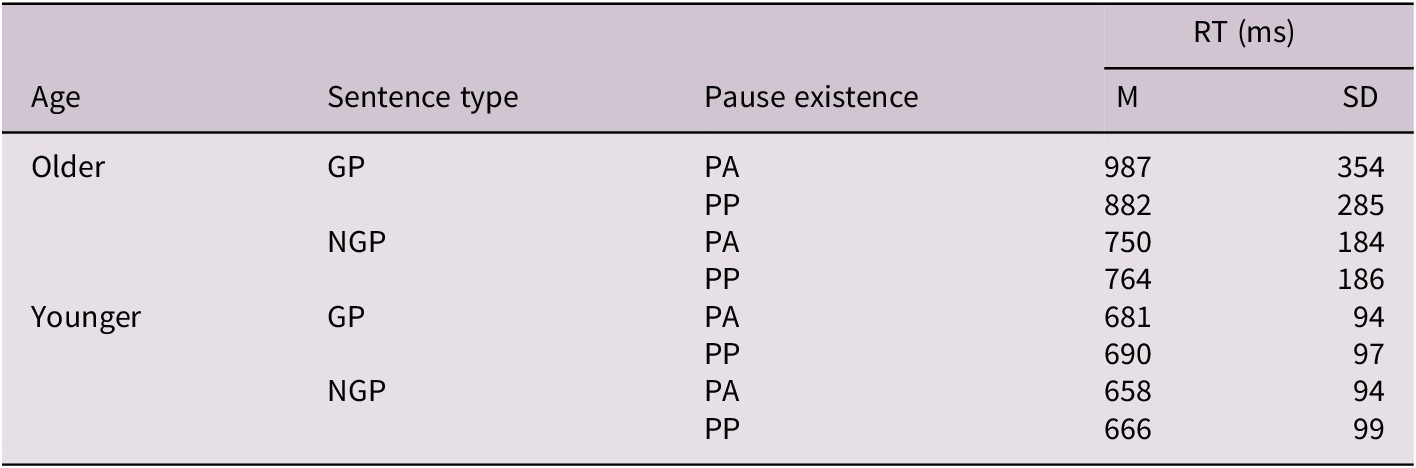
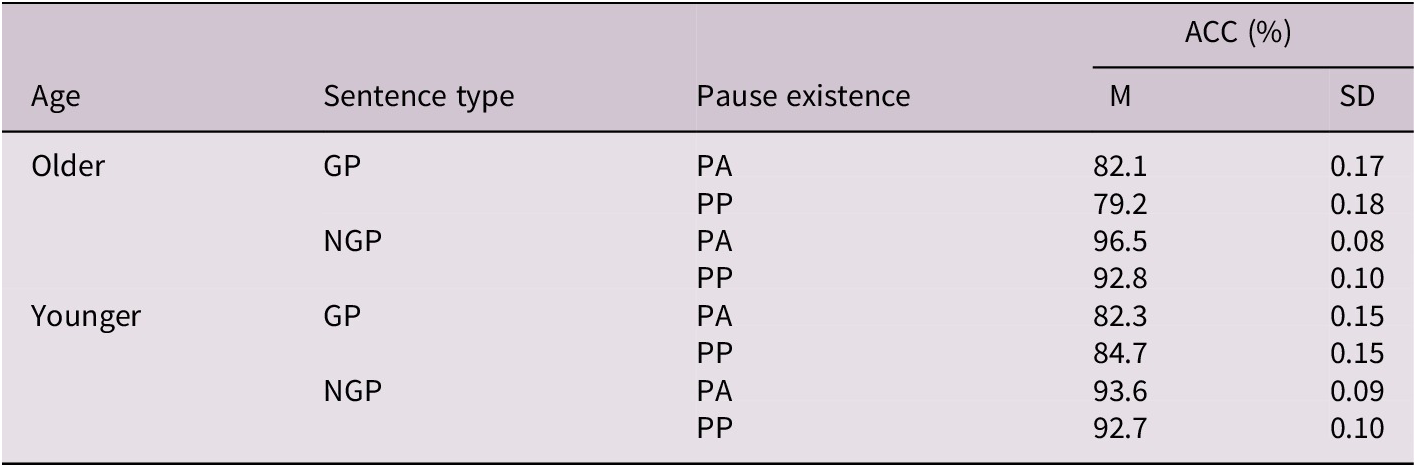







 Open access
Open access