


Introduction
Silverleaf nightshade is a perennial broadleaf weed that is recognized as one of the most aggressively invasive plants worldwide (Mekki Reference Mekki2007). The weed has been documented in Greece, India, Portugal, Spain, the United States, Australia, and Northwest Africa (Brunel Reference Brunel2011). Silverleaf nightshade affects a wide range of field and vegetable crops, including corn (Zea mays L.) and processing tomato (Lycopersicon esculentum L.) (Brunel Reference Brunel2011; Qasem et al. Reference Qasem, Al Abdallat and Hasan2019). Silverleaf nightshade propagates both sexually through seeds and asexually through rhizomes. A single plant can produce more than 6,000 seeds that can remain viable for at least 10 yr (Roberts and Florentine Reference Roberts and Florentine2022). In addition, its rhizomes form an extensive underground network, reaching depths of up to 2.8 m (Engelman and Gray Reference Engelman and Gray2007). Furthermore, the weed demonstrates a high tolerance to abiotic stresses, including drought, high temperatures, and high soil salinities (Garrett et al. Reference Garrett, Dendy, Frank, Rouse and Travers2006; Travlos Reference Travlos2013; Uludag et al. Reference Uludag, Gbehounou, Kashefi, Bouhache, Bon, Bell and Lagopodi2016). Mechanical control methods, such as cultivation, and plowing, are largely ineffective due to the weed’s extensive root network (Stanton et al. Reference Stanton, Wu and Lemerle2011). These methods may break the rhizomes, thereby promoting the spread of propagules, which in turn, facilitate the colonization of new areas (Roberts and Florentine Reference Roberts and Florentine2022; Tataridas et al. Reference Tataridas, Kanatas and Travlos2022; Wu et al. Reference Wu, Stanton and Lemerle2016). Consequently, management of silverleaf nightshade often fails to achieve desired levels of control (Kapiluto et al. Reference Kapiluto, Eizenberg and Lati2022; Stanton et al. Reference Stanton, Heap, Carter and Wu2009).
Studies of chemical control against silverleaf nightshade have demonstrated the efficacy of glyphosate and other postemergence herbicides such as glufosinate and various synthetic auxin herbicide mixtures. For example, glyphosate applied at 3,600 g ai ha−1 consistently provided approximately 90% control when applied at early vegetative or flowering stages, from late spring through summer. However, efficacy declined at later growth stages such as post-flowering and seed-set stages (Chalghaf et al. Reference Chalghaf, Aissa, Mellassi and Mekki2007; Gitsopoulos et al. Reference Gitsopoulos, Damalas and Georgoulas2017; Qasem Reference Qasem2014). Conversely, research in Australia, Greece, and Jordan has shown that glufosinate and mixtures of 2,4-D amine + picloram or triclopyr + aminopyralid effectively suppress silverleaf nightshade across vegetative growth stages, from juvenile plants through late vegetative growth prior to flowering (Gitsopoulos et al. Reference Gitsopoulos, Damalas and Georgoulas2017; Qasem Reference Qasem2014; Wu et al. Reference Wu, Stanton and Lemerle2016). These studies underscore the need to expand the toolbox for managing silverleaf nightshade and emphasize the importance of determining how growth stage and propagule types (seed-derived vs. rhizome-derived plants) affect postemergence treatment efficacy. Furthermore, control of silverleaf nightshade using postemergence herbicides is limited by two key factors: inadequate suppression of seed and rhizome emergence in the absence of targeted preemergence options, and the continual evolution of herbicide resistance. Although confirmed resistance has not yet been reported for silverleaf nightshade, several related Solanum species have shown documented resistance to multiple herbicide modes of action, including eastern black nightshade (S. ptycanthum Dunal), which has evolved resistance to herbicides that inhibit acetolactate synthase; and black nightshade (S. nigrum L.) and American black nightshade (S. americanum Mill.), which have shown resistance to photosystem I and II inhibitors (Heap Reference Heap2026; Vencill et al. Reference Vencill, Nichols, Webster, Soteres, Mallory-Smith, Burgos, Johnson and McClelland2012).
To address these challenges, integrating thermal control tools into weed-management practices is increasingly urgent. Thermal methods, such as using the flames from propane torches, kill plants by raising tissue temperatures until cell membranes rupture and the proteins denature (Ascard Reference Ascard1998; Kanellou et al. Reference Kanellou, Economou, Papafotiou, Ntoulas, Lyra, Kartsonas and Knezevic2017; Martelloni et al. Reference Martelloni, Fontanelli, Frasconi, Raffaelli and Peruzzi2016). Previous studies that investigated broadcast flaming of various weed species, such as barnyardgrass [Echinochloa crus-galli (L.) P. Beauv.] and kochia [Kochia scoparia (L.) Schrad], found that flaming efficacy differed among species and growth stages. Generally, broadleaf species and younger weeds were more sensitive than grasses at later stages (Ulloa et al. Reference Ulloa, Datta and Knezevic2010a).
Another emerging thermal control method is electrocution (i.e., the use of electrical energy to kill plants) (Vigneault and Benoît Reference Vigneault, Benoît, Vincent, Panneton and Fleurat-Lessard2001). Electrocution delivers a high-voltage electrical current through plant tissues, causing heating and disruption of cell membranes, which leads to irreversible damage (Bond and Grundy Reference Bond and Grundy2001; Diprose et al. Reference Diprose, Benson and Hackram1980, Reference Diprose, Fletcher, Longden and Champion1985). Recent studies indicate that electrocution control efficacy depends on plant size, morphology, growth stage, and the type of electrical system used (i.e., continuous-contact vs. pulsed systems) (Bloomer et al. Reference Bloomer, Harrington, Ghanizadeh and James2023; Borger and Slaven Reference Borger and Slaven2025; Coe et al. Reference Coe, Bunton, Lee, Tate, Wesslak and Bradley2025).
Several studies have evaluated the use of electrocution treatment for weed control. Vigneault and Benoît (Reference Vigneault, Benoît, Vincent, Panneton and Fleurat-Lessard2001), for example, showed effective control of annual weeds, such as redroot pigweed (Amaranthus retroflexus L.), with factors such as voltage, treatment duration, growth stage, and plant density influencing control performance. However, only a few studies have been conducted to determine the effectiveness of flaming and electrocution on perennial weeds in general and on silverleaf nightshade in particular (Harker and O’Donovan Reference Harker and O’Donovan2013). These knowledge gaps underscore the need to explore new chemical and thermal control options for silverleaf nightshade. Therefore, the overall goal of this study was to evaluate the control efficacy of various treatments against silverleaf nightshade. Specifically, we assessed 1) seven preemergence herbicides in a dose-response experiment targeting seed emergence; 2) seven postemergence herbicides applied at three rates and across three growth stages to plants derived from two propagule types; and 3) two thermal treatments, propane flaming and electrocution, delivered at three rates, using the same three growth stages and propagule types as in the postemergence experiment. We quantified seedling emergence under preemergence treatments and measured biomass reduction under postemergence and thermal treatments to assess overall efficacy and the influence of growth stage. Responses of seed-derived plants for the preemergence and postemergence treatments were evaluated across three populations to assess potential population-level variability in control efficacy. The rhizome-derived plants and thermal treatments were evaluated using a single population.
Materials and Methods
Plant Material
Silverleaf nightshade berries were collected from fields in Kfar Blum (KB) (33.16°N, 35.59°E), Newe Ya’ar (NY) (32.70°N, 35.18°E), and Urim (UR) (31.30°N, 34.52°E) Israel, and treated as detailed previously by Kapiluto et al. (Reference Kapiluto, Eizenberg and Lati2022). These populations were collected from three different agro-ecological regions in Israel to capture potential population level variability in herbicide response. Rhizomes of silverleaf nightshade were collected from a single population located in Nahalal in the Jezreel Valley (32.42°N, 35.12°E), and treated as described by Kapiluto et al. (Reference Kapiluto, Smirnov, Achdary, Eizenberg and Lati2024). This location (near the Newe Ya’ar Research Center) was used to ensure uniformity and sufficient availability of rhizomes for the postemergence and thermal experiments.
Experimental Setup
All experiments were conducted in a net house at the Newe Ya’ar Research Center. Plants were grown in pots, from seeds and rhizomes. The pots were watered by an automated mini-sprinkler irrigation system as needed. Irrigation timing was set to the same hour each day (5:00 AM). Soil moisture was not directly quantified during the treatments. However, the same irrigation timing throughout the experiments aimed to ensure comparable soil moisture among treatments. On days of treatment, the plants were taken out of the net house, and the relevant herbicide was applied using a chain-driven sprayer equipped with a flat-fan 8001E nozzle (TeeJet Technologies, Glendale Heights, IL), delivering 200 L ha−1 at a pressure of 200 kPa, with the nozzle positioned 40 cm above the plant canopy. After the treatment, the plants were returned to the net house until the end of the experiment. Seven preemergence and seven postemergence herbicides were evaluated in this study. These herbicides are representative of eight distinct modes of actions (Table 1). Furthermore, these herbicides are among the most widely used in both annual and perennial cropping systems and have differing regulatory statuses across cropping systems worldwide. Preliminary results revealed lack of control efficacy using all preemergence herbicides on plants grown from rhizomes. Thus, in this part of the study, only seed-derived plants were used. For the postemergence herbicide and thermal control experiments, both seed-derived and rhizome-derived plants were used. Each experiment (preemergence, postemergence, and thermal) was conducted in a completely randomized design with six replicates per treatment and was repeated twice in time (two experimental runs).
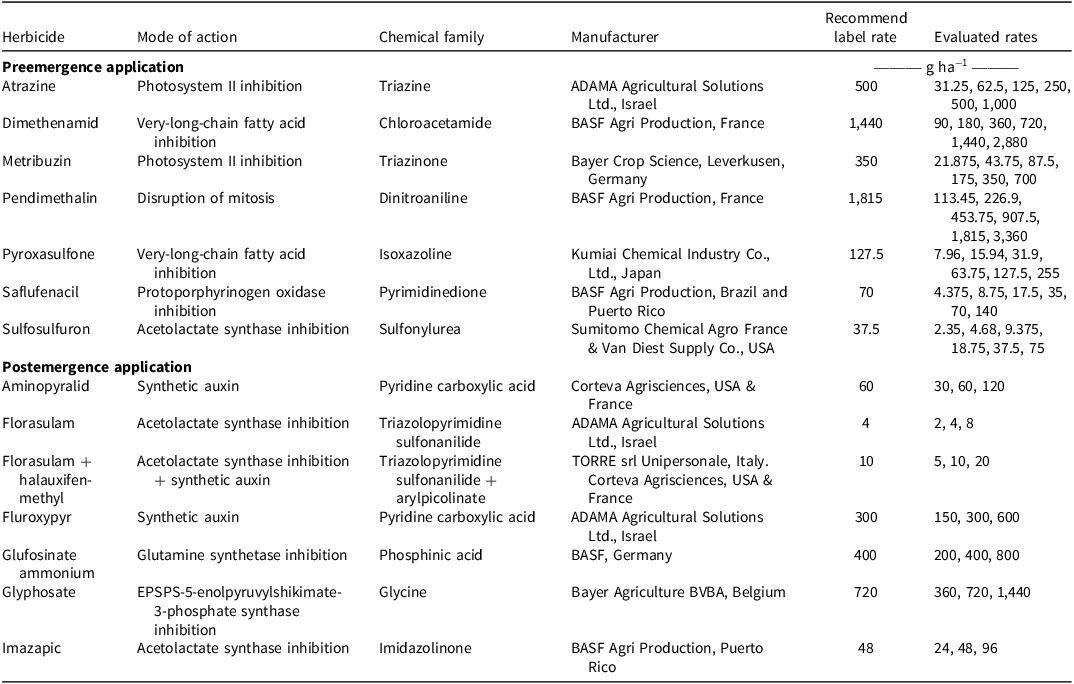
Mode of action, chemical family, manufacturer, recommended label rates and tested rates applied in the preemergence and postemergence herbicide efficacy experiments.

Table 1 Long description
The table compares herbicides, their modes of action, chemical families, manufacturers, and recommended and evaluated application rates for preemergence and postemergence applications. It has 14 rows and 6 columns. The columns are labeled Herbicide, Mode of action, Chemical family, Manufacturer, Recommended label rate, and Evaluated rates. The table is divided into two sections: Preemergence application and Postemergence application. Each row lists a specific herbicide along with its mode of action, chemical family, manufacturer, recommended label rate in grams per hectare, and evaluated rates in grams per hectare. The evaluated rates vary widely for each herbicide.
Preemergence Application of Herbicides to Silverleaf Nightshade Seeds
This experiment was conducted in July and in August 2020 under a 14-h light:10-h dark photoperiod with maximal and minimal temperatures of 35 and 23 C, respectively. A dose-response experiment was conducted for herbicides from five modes of action (Table 1). Twenty silverleaf nightshade seeds were sown per one 0.25-L pots (80 mm in diameter) filled with local soil (chromic haploxerert, fine-clayey, montmorillonitic, thermic) containing 55% clay, 25% silt, and 20% sand. The soil had 2% organic matter, pH 7.2, and a bulk density of 1.37 g cm−3. Herbicides were applied at seven rates (0, 0.0625×, 0.125×, 0.25×, 0.5×, 1×, 2×), as detailed in Table 1. Each population was seeded separately. Seeds were sown at a depth of 10 mm. All herbicides were applied on the same day, directly to the soil, immediately after seeding. Pendimethalin was the only herbicide sprayed onto and incorporated into the soil before the seeds were sown. After herbicide application, pots were irrigated to field capacity and subsequently irrigated as needed. The objective of the preemergence experiment was to quantify inhibition of seedling emergence. Thus, this variable was recorded daily in each pot until 28 d after treatment (DAT). Final emergence counts were expressed as a percentage of the untreated control (mean of six pots) using Equation 1:
 $Seedling{\rm{\;}}emergence{\rm{\;\% }} = \left( {{{{E_{rep}}}}\over{{{{\bar E}_{NTC}}}}} \right) \times 100{\rm{\;\;\;}}$
$Seedling{\rm{\;}}emergence{\rm{\;\% }} = \left( {{{{E_{rep}}}}\over{{{{\bar E}_{NTC}}}}} \right) \times 100{\rm{\;\;\;}}$
where Erep is the seedling count in a given replicate, and Ē NTC represents the mean seedling count in all replicates of the nontreated control (NTC) of the respective population.
Postemergence Applications and Thermal Treatment of Silverleaf Nightshade Grown from Seeds and Rhizomes
The postemergence experiment was conducted in July and in August 2021 under a 14-h light:10-h dark photoperiod with maximal and minimal temperatures of 36 C and 22 C, respectively. The thermal-treatments (flaming and electrocution) were conducted in July and in August 2023 with maximal and minimal temperatures of 36 C and 23 C, respectively, and followed the same photoperiod as described in the preemergence experiment. For both experiments, plants grown from rhizomes and seeds were used. Seeds were initially germinated in seedling trays filled with a commercial potting medium (Tuff; Marom Golan, Israel). Seeds emerged approximately 7 to 10 d after sowing. At the 2-leaf stage, seedlings were transplanted into individual 0.25-L pots (one seedling per pot) filled with the same potting medium. The rhizomes were washed, cut into 5-cm segments, and transplanted at a depth of 1 cm into pots (12.3 cm long, 7.9 cm wide, 5.7 cm high) filled with potting medium. Rhizome sprouts emerged after approximately 4 to 7 d. Treatments were applied after plants reached three growth stages: 2 to 3 true leaves (TL), 4 to 6 TL, and 7 to 10 TL. At the time of treatment, mean plant height was approximately 4 to 5 cm at the 2- to 3-TL stage, 10 to 14 cm at the 4- to 6-TL stage, and 17 to 22 cm at the 7- to 10-TL stage. Herbicides representing four modes of action were applied on the same day using the sprayer described earlier. Each herbicide was applied at three rates: the recommended label rate (1×), and two exploratory rates (0.5× and 2×) (Table 1).
Before flaming treatments were initiated, pots were removed from the net house and arranged in a straight 10-m row, spaced 50 cm apart. The flamethrower was then driven along the row to apply broadcast flaming to each plant. We used liquefied petroleum gas (LPG) in a commercial flamethrower (A 1000 H; Reinert, Germany) fitted with SB 300 burners (30 cm width, 2 bar pressure) mounted on a floating beam (SD30; Garford, England). Dosages were calculated as kilograms of LPG per hectare (kg ha−1) by varying the speed of the tractor (John Deere 6145 with GPS guidance) between 4 and 8 km h−1, yielding three application rates of 33, 50 and 100 kg ha−1, which were selected to represent low-, intermediate-, and high-intensity treatments, respectively. These rates were selected based on previously studies that used similar propane doses and application speeds (Knezevic et al. Reference Knezevic, Stepanovic and Datta2014; Ulloa et al. Reference Ulloa, Datta and Knezevic2010b).
For the electrocution treatments, a high-voltage weeding laboratory prototype was used, as described by Lati et al. (Reference Lati, Rosenfeld, Ben David and Bechar2021). This prototype uses a manual system with two electrodes powered by a 220-V AC input source, operating with an input current of 60 mA, and power factor (cosΦ) of 0.82, resulting in a real input power of 10.8 W (real input power [W] = V ×I × cosΦ). The output current in each electrode is 5 mA and 1,800 V, providing a discharge power of 9 W (discharge power [W] = V × A). The electric current in the two electrodes of the prototype is maintained below the 10-mA threshold that is considered hazardous for AC current. All treatments were held manually as follows: The first electrode was held next to the plants and had continuous contact with the upper leaf, while the second electrode was placed 5 cm below the soil surface for grounding. Treatments were held on the same hour (10:00 AM) to minimize variability in soil moisture. Three application rates were applied, adjusted by varying the duration of the continuous contact of the electrode and the plant upper-leaf (i.e., exposure durations) in seconds: 2 s, 5 s, and 10 s. Since energy equals to the applied power (9 W) multiplied by time (2, 5, and 10 s), the corresponding energy treatments were 18 J, 45 J, and 90 J, respectively (E = P × t). These rates were selected according to established protocols revealed in previous studies (Gafni et al. Reference Gafni, Bechar, Bakshian, Smirnov, Rosenfeld and Lati2025; Kovetz et al. Reference Kovetz, Bechar, Rosenfeld, Smirnov and Lati2025; Lati et al. Reference Lati, Rosenfeld, Ben David and Bechar2021).
In both the postemergence and thermal experiments, the aboveground biomass was harvested at 21 DAT, and oven-dried at 65 C for 72 h to constant weight. The dry biomass data were then converted into percent of biomass reduction (%) relative to the NTC using the Equation 2 (Wortman Reference Wortman2014):
 $Biomass{\rm{\;}}Reduction{\rm{\;\% }} = \left( {1 - \left( {{{{B_{rep}}}}\over{{{{\bar B}_{NTC}}}}} \right)} \right) \times 100{\rm{\;\;\;}}$
$Biomass{\rm{\;}}Reduction{\rm{\;\% }} = \left( {1 - \left( {{{{B_{rep}}}}\over{{{{\bar B}_{NTC}}}}} \right)} \right) \times 100{\rm{\;\;\;}}$
where B
rep
represents the plant biomass in a given replicate, and
 ${\bar B_{NTC\;}}$
represents the mean biomass of plants from the NTC pots, for the respective seed populations or rhizomes.
${\bar B_{NTC\;}}$
represents the mean biomass of plants from the NTC pots, for the respective seed populations or rhizomes.
Statistical and Data Analysis
Statistical analyses were conducted in R (R Core Team 2021) with figures drawn in GGPLOT2 (Wickham Reference Wickham2016). For the preemergence experiment, seedling emergence (%) was modeled with four-parameter log-logistic curves using the drm() function in the drc package as described by Ritz et al. (Reference Ritz, Baty, Streibig and Gerhard2015) and show in Equation 3:
 $Y = c + {{d - c} \over {\left\{ {1 + \exp \left[ {b\left( {log\left( x \right) - \log \left( e \right)} \right)} \right]} \right\}}}$
$Y = c + {{d - c} \over {\left\{ {1 + \exp \left[ {b\left( {log\left( x \right) - \log \left( e \right)} \right)} \right]} \right\}}}$
where Y represents the response variable, seedling emergence (%); d is the upper limit of the response; c is the lower limit of the response; e shows the response halfway between d and c and should not be used as the ED50 (the dose required to result in a 50% reduction in weed biomass) (Keshtkar et al. Reference Keshtkar, Kudsk and Mesgaran2021). To assess whether emergence responses differed among populations or experimental runs, we first fitted a full four-parameter log-logistic model in which each of the three populations (KB, UR, NY) and each run had its own set of b, c, d, and e parameters. We then fitted a reduced model that pooled all populations allowing parameters to vary by run only and compared the two models via F-tests (anova on drm objects; mass package).
No significant differences among populations were found (P ≥ 0.05; data not shown), thus populations were pooled. Since the experimental runs differed significantly (P < 0.05, Table S1), trail data were retained as a curve-ID factor (Table S1 and Figure 1). Effective rates (ED50, ED90) and 95% confidence intervals (CIs) were estimated by the delta method using the ED() function (Keshtkar et al. Reference Keshtkar, Kudsk and Mesgaran2021). To formally test whether ED50 and ED90 differed between runs, we applied EDcomp(), which computes the ratio of ED values and assesses significance via a t-statistic (P < 0.05 denotes a significant run-to-run difference).
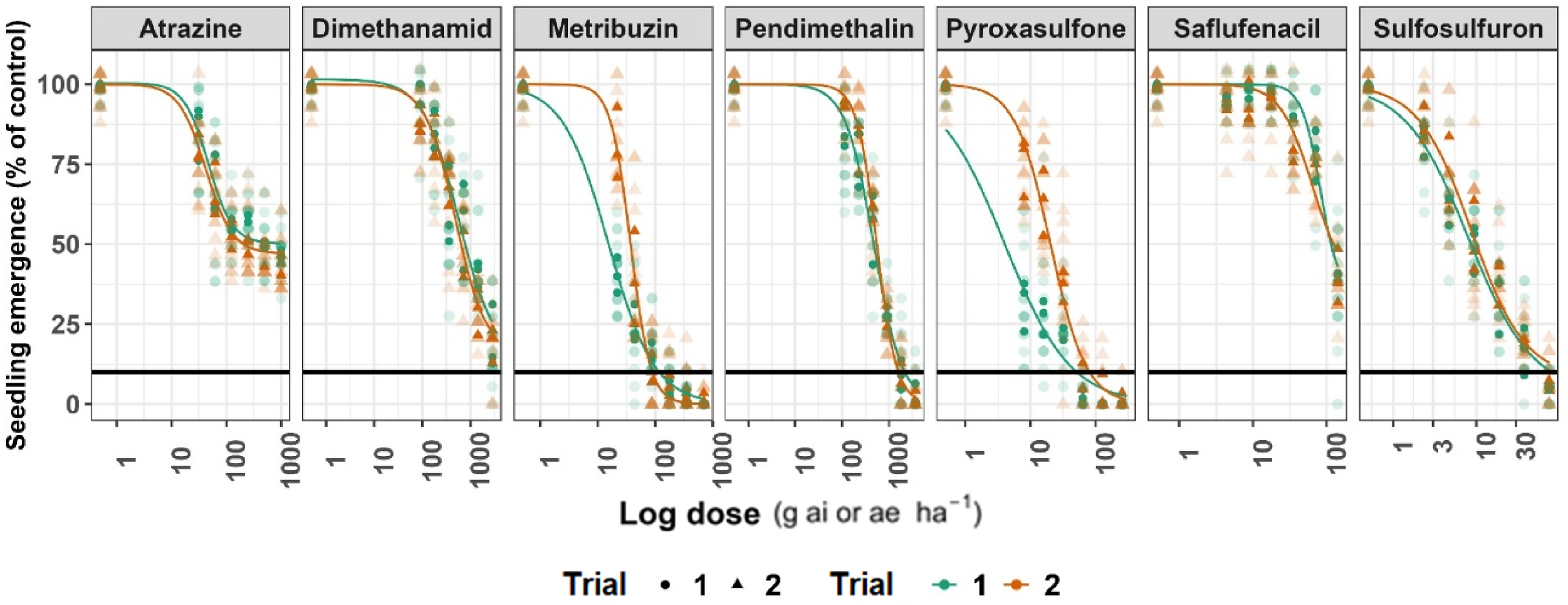
Influence of increasing herbicide rates on seedling emergence (% of untreated control) in silverleaf nightshade, based on pooled data from three populations (Kfar Blum, Newe Ya’ar, and Urim) across two experimental trails. Significant differences were detected between trials (Table S1). Individual observations are shown as circles (Trial 1) and triangles (Trial 2), and fitted log-logistic curves are colored green (Trial 1) and orange (Trial 2). The horizontal black line indicates 90% reduction in emergence. Parameter estimates are presented in Table S1.

In the postemergence and thermal experiments, herbicide or weed control method, application rate, growth stage, propagule type, and population were treated as fixed effects, while experimental run was included as a random effect. In these experiments, biomass reduction (%) was logit-transformed to meet normality assumptions (Warton and Hui Reference Warton and Hui2011) and then analyzed by Gaussian distribution with general linear mixed models (GLMMs) using the glmmTMB package (v.1.0.2.1) (Brooks et al. Reference Brooks, Kristensen, van Benthem, Magnusson, Berg, Nielsen, Skaug, Mächler and Bolker2017). Overall significance was tested via Type II Wald χ2 tests (ANOVA), and Tukey-adjusted pairwise comparisons (α = 0.05) were performed with the emmeans package (v.1.5.4) (Lenth et al. Reference Lenth, Bolker, Buerkner, Giné-Vázquez, Herve, Jung, Love, Miguez, Riebl and Singmann2025). Back-transformed least-square means are reported.
For the postemergence data, we first fitted a GLMM with the full four-way interaction analysis (population × herbicide × rate × growth stage), including all lower-order terms, plus experimental run as a random effect and an untreated control in each series. This model tested three seed populations (KB, UR, NY), seven herbicides (Table 1), three rates (0.5×, 1×, 2×), and three growth stages (2 to 3 TL, 4 to 6 TL, 7 to 10 TL). A Type II Wald ANOVA revealed no population effect (P = 0.91; Table S3), indicating that all seed sources responded similarly. We then analyzed the NY population across the two propagule types, seed-derived and rhizome-derived plants. Using the same GLMM structure, we ran a 2 × 7 × 3 × 3 factorial model (propagule type × herbicide × rate × growth stage), including untreated controls and modeling run as a random effect. All main effects and interactions were assessed by Type II Wald χ2 tests with Tukey-adjusted pairwise comparisons rate × three growth stage. Flaming and electrocution experiments were each modeled separately with a GLMM containing the full three-way interaction. We ran a 2 × 3 × 3 factorial, and fixed effects were propagule type, treatment rate, and growth stage, including untreated controls and experimental run as a random effect.
Results and Discussion
Preemergence Application of Herbicides to Silverleaf Nightshade Seeds
The four-parameter log-logistic model (Equation 3) adequately described the relationship between silverleaf nightshade seed emergence and increasing rates of seven preemergence herbicides, across the two experimental runs (Figure 1 and Table S1). Significant run effects were detected for five of the seven herbicides (atrazine, metribuzin, pendimethalin, pyroxasulfone, and saflufenacil), as indicated by the F- and P-values (Table S1), justifying the presentation of dose-response curves separately for each experimental run. For dimethenamid or sulfosulfuron, no significant effect of the experimental run was observed. The dose response analysis revealed that metribuzin and pyroxasulfone were the most effective herbicides, each reducing the emergence of silverleaf nightshade seedlings by more than 90% (Figure 2 and Table S2), at application rates below their labeled recommendations. In Run 1, the metribuzin ED50 was 15.5 g ha−1 (CI 12.3 to 18.7) and the ED90 was 117.7 g ha−1 (CI 87.9 to 147.3), where the recommended label rate is 350 g ha−1. For pyroxasulfone in Run 1 the ED50 and ED90 values were 4.1 g ha−1 (CI 2.6 to 5.5) and 51.6 g ha−1 (CI 34.5 to 68.8), respectively, where the recommended label rate is 127.5 g ha−1 (Figure 2 and Table S2). Pendimethalin had lower efficacy than the first two herbicides, with an ED90 of 2,183 g ha−1 (CI 1,833 to 2,532) in Run 1, above its 1,815 g ha−1 recommended label rate. However, in Run 2, the ED90 was 1,607 g ha−1 (CI 1,390 to 1,825), suggesting that its efficacy level may be affected by environmental factors.
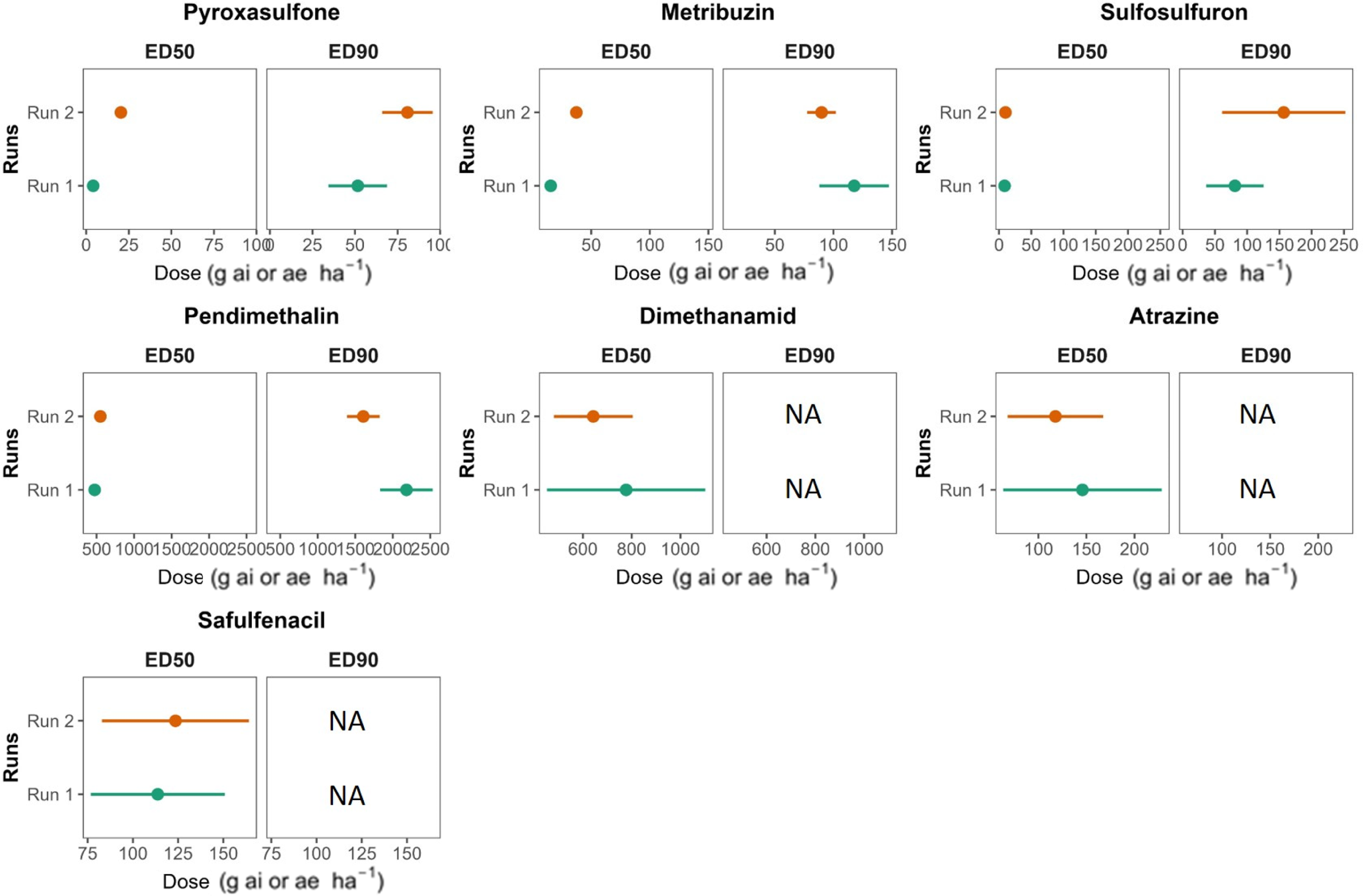
Effective dose estimates (ED50 and ED90; g ai ha−1) with 95% confidence intervals for preemergence treatments (pyroxasulfone, 127.5; pendimethalin, 1,815; saflufenacil, 70; metribuzin, 350; dimethenamid, 1,440; sulfosulfuron, 37.5; and atrazine, 500) on silverleaf nightshade by experimental run. The recommended label rates (g ai ha−1) are shown in parentheses. “NA” indicates that an ED90 could not be estimated. Effective dose estimates are presented in Table S2.

Figure 2 Long description
The image contains multiple scatter plots showing effective dose estimates (ED50 and ED90) for different herbicides on silverleaf nightshade. Each herbicide is represented in two separate plots for ED50 and ED90, with data from two experimental runs. Panel A: Pyroxasulfone. The x-axis represents the dose in grams per hectare, and the y-axis represents the runs. ED50 and ED90 plots show dose values around 25 to 75 and 50 to 100, respectively. Panel B: Metribuzin. The x-axis represents the dose in grams per hectare, and the y-axis represents the runs. ED50 and ED90 plots show dose values around 50 to 150 and 100 to 150, respectively. Panel C: Sulfosulfuron. The x-axis represents the dose in grams per hectare, and the y-axis represents the runs. ED50 and ED90 plots show dose values around 50 to 150 and 100 to 250, respectively. Panel D: Pendimethalin. The x-axis represents the dose in grams per hectare, and the y-axis represents the runs. ED50 and ED90 plots show dose values around 500 to 2500 and 1000 to 2500, respectively. Panel E: Dimethanamid. The x-axis represents the dose in grams per hectare, and the y-axis represents the runs. ED50 plot shows dose values around 600 to 1000, while ED90 is not applicable. Panel F: Atrazine. The x-axis represents the dose in grams per hectare, and the y-axis represents the runs. ED50 plot shows dose values around 100 to 200, while ED90 is not applicable. Panel G: Saflufenacil. The x-axis represents the dose in grams per hectare, and the y-axis represents the runs. ED50 plot shows dose values around 75 to 150, while ED90 is not applicable.
Sulfosulfuron had an ED90 of 81.0 g ha−1 (CI 36.4 to 125.6) and 156.6 g ha−1 (CI 61.1 to 252.1) in Runs 1 and 2, respectively. For this herbicide, the recommended label rate is 37.5 g ha−1, indicating adequate control only at elevated rates (Figure 2). Furthermore, atrazine, saflufenacil, and dimethenamid did not reached 90% emergence reduction at our highest rates, and ED90 values could not be estimate (ED90 “NA”), emphasizing their poor performance. Although ED estimates fluctuated by up to 30% between runs likely reflecting environmental and seed-batch variability the overlapping confidence intervals and the identical herbicide ranking between the runs (metribuzin ≈ pyroxasulfone > pendimethalin > sulfosulfuron ≫ dimethenamid/saflufenacil/atrazine) reinforce the robustness of our results (Table S1; Figures 1 and 2).
Three preemergence herbicides with three distinct modes of action, metribuzin, pyroxasulfone, and sulfosulfuron provided effective (>90%) suppression of seedling emergence at rates below or near their recommended label rates. Among these treatments, metribuzin can be applied to potatoes (Solanum tuberosum L.), tomatoes, and carrots (Daucus carota L.). Pyroxasulfone can be applied to peanut and to legume crops such as chickpea, pea (Pisum sativum L.), lentils (Lens culinaris Medik.), and lupins (Lupinus spp.), and sulfosulfuron can be applied to tomatoes. Notably, none of these three herbicides carry restrictions for use on rotational crops, highlighting their flexibility and suitability for long-term crop rotation strategies. Interestingly, a comparison of metribuzin and atrazine, both PS II inhibitors, showed metribuzin’s superior efficacy, in keeping with previous studies that found metribuzin to be more effective than atrazine in controlling Palmer amaranth (Amaranthus palmeri S. Watson) (Faleco et al. Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022) and waterhemp [Amaranthus tuberculatus (Moq.) J.D. Sauer] (Vennapusa et al. Reference Vennapusa, Faleco, Vieira, Samuelson, Kruger, Werle and Jugulam2018).
Another important, effective preemergence herbicide is pendimethalin, a key weed control option for cotton, tomato, and bean. The use of pendimethalin has increased in recent years following the European Union’s ban on trifluralin under the “Europe-GAP” regulations, making these findings particularly relevant (Achdari et al. Reference Achdari, Hanan, Lati and Matzrafi2022). Our results indicate that achieving 90% control with pendimethalin requires rates above the recommended label rate. However, previous studies have shown that pre-plant incorporated applications can enhance pendimethalin efficacy without reducing crop yield in several rotation crops, including Ethiopian mustard (Brassica carinata A. Braun) (Leon et al. Reference Leon, Ferrell and Mulvaney2017) and peanuts (Arachis hypogaea L.) (Prostko et al. Reference Prostko, Johnson and Mullinix2001). These results highlight the need to optimize application methods and to consider the integration of pendimethalin with other herbicides to enhance its effectiveness without increasing herbicide loads to unsustainable levels.
Postemergence Applications on Silverleaf Nightshade Grown from Seeds and Rhizomes
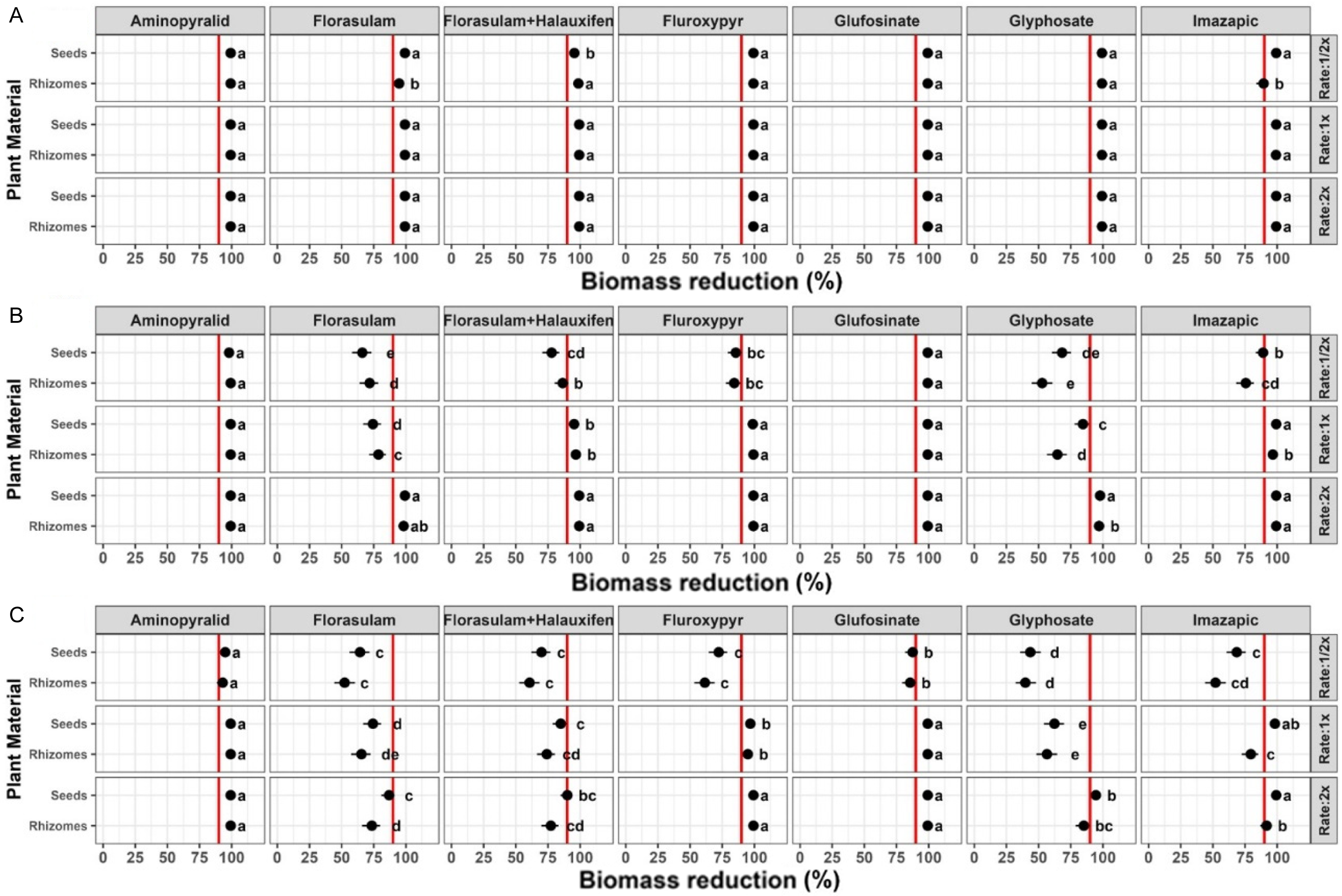
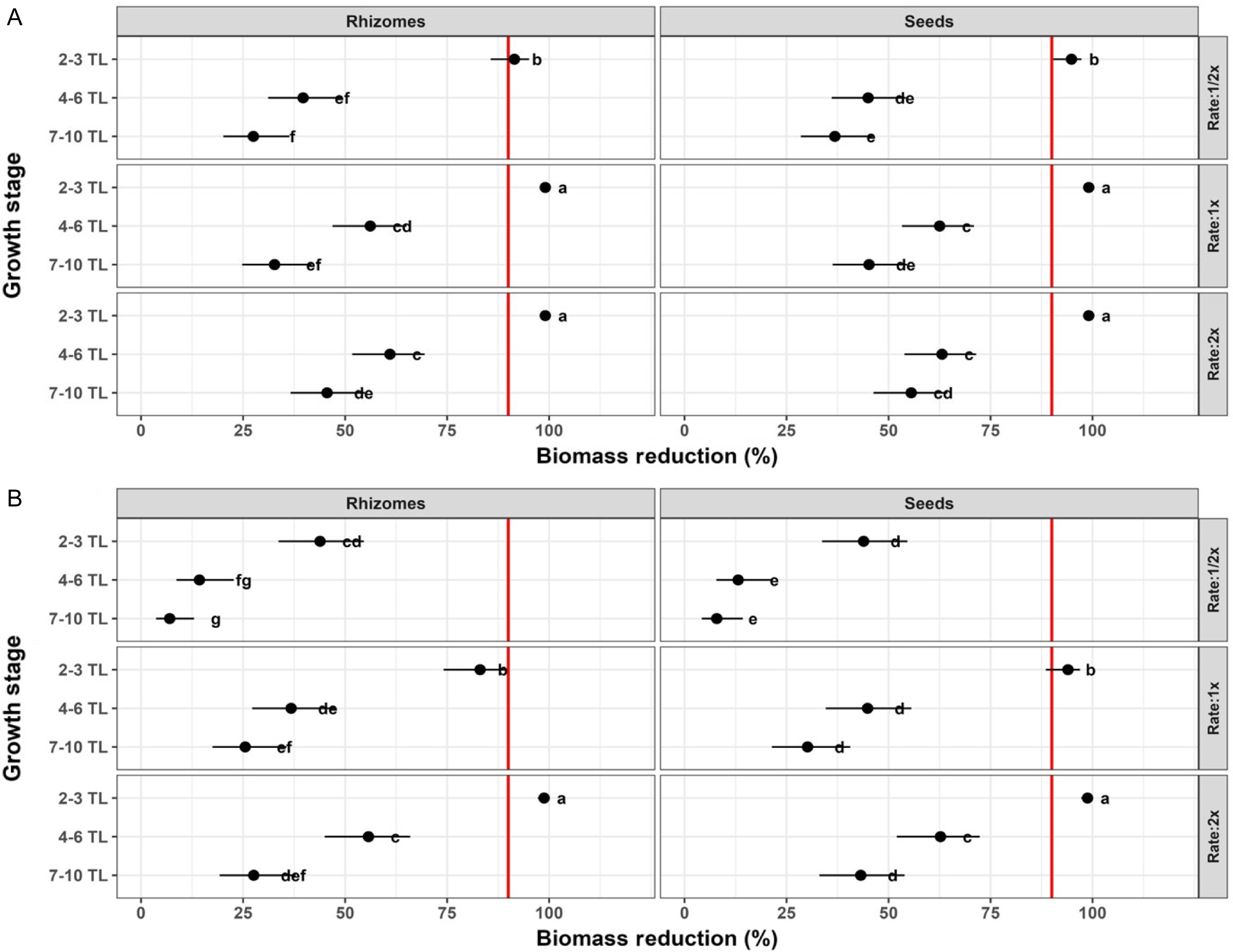
Among all postemergence herbicides, growth stage was the primary driver of biomass reduction, with efficacy consistently declining as plant size increased (Figure 3). At the 2- to 3-TL stage, all treatments achieved high (≥90 %) biomass reduction at all rates, and in both seed-derived and rhizome-derived plants (Figure 3A). This decline in efficacy became marked at the 4- to 6-TL stage (Figure 3B), where biomass reduction below 90% were observed for four treatments at the 0.5× rate (florasulam, florasulam + halauxifen, fluroxypyr and glyphosate) and for two treatments at the 1× rate (florasulam and glyphosate). For example, biomass reduction in plants treated with florasulam that were grown from seeds was approximately 62% and approximately 75% following treatments at 0.5× and 1× rates, respectively. At the later 7- to 10-TL stage, a further decline in biomass reduction was observed (Figure 3C). At that stage, all treatments (except aminopyralid) achieved <90% biomass reductions at the 0.5× rate, and at the 1× rate. For example, at 0.5× rate, the biomass reduction of plants grown from rhizomes was 64% and 54% with applications of fluroxypyr and imazapic, respectively. These results show the importance of early postemergence application for favorable silverleaf nightshade control, and the significant interaction of growth stage and herbicide (Table S4, P < 0.001). However, this study evaluated short-term control efficacy, which was based on biomass reduction at 21 DAT. Further research is needed to determine the regrowth from belowground organs, which is relevant for long-term management of perennial weeds.
Biomass reduction (%) of silverleaf nightshade shoots of plants grown from seeds and rhizomes in response to seven postemergence herbicides (in g ai ha−1: aminopyralid, 60; florasulam, 4; florasulam + halauxifen-methyl, 10; fluroxypyr, 300; glufosinate ammonium, 400; glyphosate, 720; and imazapic, 48) applied at three rate multiples (0.5×, 1×, and 2× of the recommended label rate) across three growth stages: (A) 2 to 3 true leaves (TL), (B) 4 to 6 TL, and (C) 7 to 10 TL. Growth stages are shown in separate panels for graphical clarity. Statistical inference is based on a full factorial model including propagule type (seed-derived vs. rhizome-derived plants), herbicide, application rate, and growth stage. Points represent mean biomass reduction with 95% confidence intervals, and letters denote Tukey HSD groupings (α = 0.05).

The specific herbicide applied and its interactions also had a significant effect on biomass reduction values (Table S4, P < 0.001). In terms of applied herbicide, aminopyralid was the most effective treatment across both propagule types and all three application rates. This herbicide was the only treatment that at the 7- to 10-TL stage showed uniformly high ≥90 % biomass reduction values. In contrast, florasulam provided lower biomass reduction at the 7- to 10-TL stage. At the 0.5× and 2× rates, this treatment reduced biomass of plants grown from rhizomes by 50% and 75%, respectively. Notably, florasulam still provided >90% biomass reduction at the 2- to 3-TL stage, even at the 0.5× rate, further emphasizing the herbicide by growth-stage interaction revealed in this study.
Lastly, propagule type and its interaction with growth stage had a significant effect on biomass reduction, mostly at the 4- to 6-TL and 7- to 10-TL stages (Table S4, P < 0.001). For example, glyphosate at the 7- to 10-TL stage showed approximately10% differences between the biomass reduction values of plants grown from seed and rhizome-derived plants. At the 0.5× rate, biomass reduction of plants grown from seeds and rhizomes was 40% and 50%, respectively. In contrast, at the 2× rate the biomass reduction of plants grown from seeds and rhizomes increased to 90% and 80%, respectively. These results underscore a pronounced interaction between propagule type and growth stage (Table S4, P < 0.001). Nonetheless, the approximate 10% difference in biomass reduction values between the two propagule types has marginal impact under actual real-world field conditions.
Aminopyralid and fluroxypyr provided >90 % biomass reduction at the 2- to 3-TL stage even when applied at 0.5× rate, indicating strong early-stage control. Similar control results were also reported in Australia by Wu et al. (Reference Wu, Stanton and Lemerle2016). Likewise, glufosinate achieved ≥95 % biomass reduction at the 2- to 3-TL state, which is consistent results reported by Gitsopoulos et al. (Reference Gitsopoulos, Damalas and Georgoulas2017), although its use is now restricted under European Union regulations. Among the herbicides that inhibit acetolactate synthase, imazapic was effective at the 2- to 3-TL stage but biomass reduction dropped below 80% by the 7- to 10-TL stage. Our results highlight the often-overlooked temporal aspect of silverleaf nightshade control and clearly show that early application is preferable for achieving optimal efficacy. Previous studies recommended treatment at mature stages, such as flowering and fruiting, when tolerance is higher, thus requiring higher rates for effective control (Gitsopoulos et al. Reference Gitsopoulos, Damalas and Georgoulas2017; Tataridas et al. Reference Tataridas, Kanatas and Travlos2022; Wu et al. Reference Wu, Stanton and Lemerle2016).
On a practical level, fluroxypyr can be applied to maize, orchards, and during seedbed preparation for wheat; imazapic is suitable for peanut; and glufosinate can be applied to orchards. Although aminopyralid was the most effective postemergence treatment, its use is limited to nonarable land. Lastly, propagule type showed a statistically significant effect (Table S4, P < 0.001). Nonetheless, the pairwise comparisons revealed differences of less than 10% biomass reduction for nearly all treatments. These results further indicate that timing and choice of herbicide, rather than propagule type, drive practical efficacy.
Thermal Treatments on Silverleaf Nightshade Grown from Seeds and Rhizomes
With both thermal control methods, propagule type, application rate, and growth stage were highly significant (P < 0.001) drivers of biomass reduction (Table S5). In contrast, any interactions involving propagule type were negligible. For flaming at the 2- to 3-TL growth stage, even the lowest LPG rate (33 kg ha−1) reduced biomass by ≥85 % on both propagule types, while the 1× (50 kg ha−1) and 2× (100 kg ha−1) rates exceeded 90% biomass reduction (Figure 4A). When treatments were applied to plants at the 4- to 6-TL stage, only the 50 and 100 kg ha−1 treatments resulted in ≥90 % biomass reduction. By the later growth stage (7- to 10-TL), flaming at 100 kg ha−1 still achieved 85% biomass reduction (Figure 4A). However, at both lower rates biomass reduction dropped below 70%, confirming a strong rate × growth stage interaction (P < 0.001, Table S5).
Biomass reduction (%) of silverleaf nightshade shoots of plants grown from seed and rhizome in response to thermal weeding by (A) flaming and (B) electrocution applied at three dose intensities (0.5×, 1×, and 2×) across three growth stages (2 to 3 true leaves [TL], 4 to 6 TL, and 7 to 10 TL). The 0.5×, 1×, and 2× doses correspond to 33, 50, and 100 kg LPG ha−1 for flaming, and 18, 45, and 90 J for electrocution. Points represent mean biomass reduction with 95% confidence intervals. Letters denote Tukey HSD groupings (α = 0.05) based on factorial models including propagule type (seed-derived vs. rhizome-derived plants), treatment rate (0.5×, 1×, 2×), and growth stage, and are shown separately for each thermal method (flaming or electrocution).

Electrocution (Figure 4B; Table S5) followed a similar trend of results, though here a marginal propagule type × rate interaction (P = 0.042) was observed. At the 2- to 3-TL stage, even the lowest energy dose (18 J) resulted in ≥80% biomass reduction on both seeds and rhizomes, and the 1× (45 J) and 2× (90 J) rates provided 90% biomass reduction. At the 4- to 6-TL stage, only the 2× rate achieved ≥90 % biomass reduction. The 1× and 0.5× rates resulted in 75% to 85 % and 60% biomass reduction, respectively. By the 7- to 10-TL stage, electrocution at 2× achieved a biomass reduction of approximately 80%, while 1× and 0.5× reduction values dropped to ≤60 %. This highlights a strong rate × growth stage interaction (P < 0.001) and shows that plants grown from seeds and rhizomes responded similarly (propagule type × growth stage P = 0.48).
Similar to the postemergence treatments, rhizome-derived plants exhibited slightly greater resilience at the 4- to 6-TL stage following the thermal treatments (Figure 4, A and B), consistent with their larger carbohydrate reserves and higher stress tolerance (Kapiluto et al. Reference Kapiluto, Smirnov, Achdary, Eizenberg and Lati2024). Under field conditions, however, plants that emerge from rhizomes are expected to be more difficult to control due to their extensive belowground organs, as reported for other perennial weeds (Ascard Reference Ascard1998). Accordingly, our results reflect short-term aboveground responses under controlled conditions and should be interpreted in this context. Furthermore, Feys et al. (Reference Feys, De Cauwer, Reheul, Sciffer, Clercx and Palmans2023) showed that electrocution alone often fails to eliminate the development of yellow nutsedge (Cyperus esculentus L.) underground tubers, underscoring the need to combine thermal weeding with other integrated weed management practices. However, early intervention at the 2- to 3-TL stage not only maximizes aboveground control with lower energy inputs (ED90 ≈ 31 to 34 kg ha−1 for flaming) but also minimizes shoot regrowth and seed production, reducing the need for repeated application.
Conducting the study in field soil might better mimic actual conductivity potentials, nonetheless, our results from a controlled environment allowed us to establish a fundamental proof of concept. Future research should validate these findings in the field to account for soil-specific variables that may affect the overall efficacy of these thermal treatments. Furthermore, it is essential to test the integration of thermal treatments with chemical and cultural tactics to ensure the sustainable suppression of both aboveground shoots and belowground propagules.
Practical Implications
Our experiment results demonstrate that silverleaf nightshade control is most effective when control activities are applied early and diversified across chemical and thermal tactics that include preemergence applications of metribuzin, pyroxasulfone, sulfosulfuron, and pendimethalin, which are readily integrated into existing rotations of potato, tomato, carrot, peanut, corn, and chickpea crops. Pendimethalin only reaches the ED90 at or above their recommended rates and may require pre-plant incorporation or tank mixes on cotton and bean crops. For postemergence control, aminopyralid, glufosinate, and imazapic stand out. Although effective silverleaf nightshade control was achieved with rates lower than those specified on the label for some preemergence and postemergence treatments, reducing application rates is not recommended due to the risk of selecting herbicide-resistant populations of other weed species. This concern is particularly relevant for preemergence treatments for which the full diversity of the weed community may not yet be known. Thermal methods likewise achieve >90% control at the 2- to 3-TL stage but decline sharply by the 7- to 10-TL stage. Under field conditions, plants emerging from rhizomes may exhibit greater tolerance due to their larger belowground reserves, and long-term control will require evaluation of regrowth potential. However, effective weed suppression early in the growing season may facilitate crop establishment and rapid crop canopy development, thereby reducing weed recovery later in the season. Integrating preemergence and postemergence herbicides with thermal tools, together with rotation of modes of action, can suppress both seedlings and rhizomes, reduce chemical inputs, and delay resistance. Future research should focus on testing the integration of chemical and thermal tools under field conditions, refining application methods, understanding soil interactions, and evaluating multiyear emergence dynamics.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/wet.2026.10092
Funding
Funding for this research was provided via grant 20-02-0076 from the Chief Scientist of the Israel Ministry of Agriculture.
Competing Interests
The authors declare they have no conflicts of interest.



 Open access
Open access