


Working memory denotes the mental processes involved in short-term maintenance and manipulation of task-relevant information. In daily life, it can be expressed as the ability to briefly hold information in the mind and use it to solve mental tasks such as adding numbers or following a set of instructions. Reference Chai, Abd Hamid and Abdullah1 Importantly, many complex higher-order functions rely on intact working memory, including learning, problem-solving and decision-making. Reference Cowan2 Impaired working memory can therefore severely affect everyday functioning Reference Aretouli and Brandt3,Reference Spencer-Smith and Klingberg4 and interfere with academic and professional achievements. Reference Gathercole, Pickering, Knight and Stegmann5,Reference Blankenship, ONeill, Ross and Bell6 Working memory deficits have been reported across a wide range of neuropsychiatric disorders, Reference Alderson, Kasper, Hudec and Patros7–Reference Wang, Zhang, Liu, Cui, Wang and Shum9 including major depressive disorder (MDD) Reference Nikolin, Tan, Schwaab, Moffa, Loo and Martin10 where patients frequently experience difficulties even after so-called remission of the depressive episode. Reference Roca, Monzón, Vives, López-Navarro, Garcia-Toro and Vicens11,Reference Kaneda12 At present, we have a very limited understanding of the neurobiological mechanisms of working memory deficits in mood disorders and, consequently, no efficient clinical strategies for treating them. Functional magnetic resonance imaging studies have implicated multiple brain regions in normal working memory functioning, Reference Emch, von Bastian and Koch13 yet the most consistent finding is enhanced activation of the prefrontal cortex (PFC) during tasks with increasing working memory load. Reference Lara and Wallis14 The importance of the PFC is further supported by lesion studies showing that frontal brain damage leads to working memory impairment in both humans Reference Müller, Machado and Knight15 and non-human primates. Reference Curtis and D’esposito16 A prevailing theory posits that the primary function of the PFC in working memory is coordinating and regulating other brain regions involved in maintaining and manipulating task-relevant information. Reference Lara and Wallis14 Notably, both structural and functional abnormalities in the PFC have reliably been shown in MDD, Reference Pizzagalli and Roberts17 which could help explain the presence of working memory deficits in patients. Although molecular imaging and pharmacological intervention studies have consistently linked dopaminergic neurotransmission, and in particular the dopamine 1 (D1) receptor, to working memory functioning, Reference Khan and Muly18 much less is known about the potential role of other neurotransmitter systems including serotonin (5-HT) signalling. The 5-HT system consists of seven receptor families, including several subreceptors and the 5-HT transporter (5-HTT), which facilitates reuptake of 5-HT from the synaptic cleft. 5-HT is produced in the raphe nuclei in the brain stem from where serotonergic neurons project to both subcortical and cortical regions, including the PFC. Reference Olivier19 Importantly, 5-HT has been strongly implicated in MDD pathology, as most standard antidepressant treatments target the 5-HT system, Reference Pereira and Hiroaki-Sato20 and recent studies have demonstrated altered 5-HT signalling in patients with MDD compared with healthy individuals. Reference Köhler-Forsberg, Dam, Ozenne, Sankar, Beliveau and Landman21,Reference Wang, Zhou, Zhu, Wang, Fang and Zhong22 In addition, serotonergic neurotransmission is involved in a wide range of cognitive domains, including learning and memory functions, Reference Švob Štrac, Pivac and Mück-Šeler23 and recent lines of preclinical evidence also support a link to working memory functioning. Reference Ellis and Nathan24 Pooling data from multiple neuroimaging studies from the Cimbi database, Reference Knudsen, Jensen, Erritzoe, Baare, Ettrup and Fisher25 we used positron emission tomography (PET) brain imaging to investigate the link between working memory function in both healthy individuals and patients in a depressed state and the expression of multiple 5-HT receptor systems in the frontal cortex, including 5-HT 1B receptor (5-HT1BR), 5-HT 2A receptor (5-HT2AR) and 5-HT 4 receptor (5-HT4R), as well as presynaptic 5-HTT levels.
Method
Participants
Working memory scores and neuroimaging data from 432 PET scans, including 332 scans collected from 269 healthy individuals and 100 scans collected from 92 patients with mood disorders, were available from the Cimbi database. Reference Knudsen, Jensen, Erritzoe, Baare, Ettrup and Fisher25 Data were only included if time between PET scan and cognitive test date was ≤100 days. The clinical cohort consisted of patients diagnosed with MDD (n = 81) and seasonal affective disorder ((SAD) n = 11). Data was pooled across 16 independent neuroimaging studies conducted at the Neurobiology Research Unit, Copenhagen University Hospital Rigshospitalet, between 2004 and 2019, and included PET scans assessing the 5-HT1BR (healthy controls: 24), 5-HT2AR (healthy controls: 97), 5-HT4R (healthy controls: 89, patients: 89) and 5-HTT (healthy controls: 122, patients: 11). Inclusion criteria for all participants were age between 18 and 65 years, no significant head trauma, no significant somatic illness, no current use of psychotropic medication, fluency in Danish and no pregnancy or breastfeeding. An additional exclusion criterion for healthy individuals was history of DSM Axis I psychiatric illness. The patients with MDD and SAD were pooled from two clinical studies. Reference Köhler-Forsberg, Jorgensen, Dam, Stenbæk, Fisher and Ip26,Reference Mc Mahon, Nørgaard, Svarer, Andersen, Madsen and Baaré27 All patients were antidepressant-free (≥2 months for MDD and ≥1 year for SAD) and met the criteria for a depressive episode (lasting <2 years) at the time of data collection; within the MDD group, 36 patients were diagnosed with first-episode depression and 45 with recurrent depression. Patients with SAD were screened with the Schedules for Clinical Assessment in Neuropsychiatry (SCAN) interview, and patients with MDD were screened with the Mini-International Neuropsychiatric Interview (M.I.N.I.); both groups had their clinical diagnoses confirmed by a trained psychiatrist.
The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional committees on human experimentation and with the Helsinki Declaration of 1975, as revised in 2013. The Cimbi database has been approved by the Danish Data Protection Agency (Capital Region protocol number: 2012-58-0004) and all data have been acquired with prior permission from the Danish Ethics Committee system and prior written informed consent from all enrolled individuals.
Measures
Working memory task
Working memory was assessed with the letter-number-sequencing task from Wechsler’s Adult Intelligence Scale III (WAIS-III). Reference Wechsler28 In the letter-number-sequencing task, a jumbled sequence of letters and numbers is read to the participant (e.g. 2-C-3-A-1-B), who is instructed to mentally sort and recite them back to the tester in numerical and alphabetical order (i.e. 1-2-3-A-B-C). The sequences increase in length and difficulty until an upper limit for working memory capacity has been found, with a maximum score of 21 points. Cognitive testing took place in standardised test rooms and was conducted by trained neuropsychological testers.
PET and structural magnetic resonance imaging
Detailed descriptions of PET data acquisition are available in previous publications. Briefly, 5-HT1BR levels were quantified with [11C]AZ10419369, Reference da Cunha-Bang, Hjordt, Perfalk, Beliveau, Bock and Lehel29 5-HT2AR levels with [18F]Altanserin (n = 32) Reference Kupers, Frokjaer, Naert, Christensen, Budtz-Joergensen and Kehlet30 and [11C]Cimbi-36 (n = 65), Reference Ettrup, Svarer, McMahon, da Cunha-Bang, Lehel and Møller31,Reference Madsen, Fisher, Burmester, Dyssegaard, Stenbæk and Kristiansen32 5-HT4R levels with [11C]SB207145 Reference Köhler-Forsberg, Jorgensen, Dam, Stenbæk, Fisher and Ip26,Reference Haahr, Fisher, Jensen, Frokjaer, Mahon and Madsen33,Reference Madsen, Marner, Haahr, Gillings and Knudsen34 and 5-HTT levels with [11C]DASB. Reference Frokjaer, Vinberg, Erritzoe, Svarer, Baaré and Budtz-Joergensen35,Reference Mc Mahon, Andersen, Madsen, Hjordt, Hageman and Dam36 PET scans were acquired on either a 3D High Resolution Research Tomography (CTI/Siemens) PET scanner ((HRRT) 388 scans) or a PET2 GE Advance scanner ((GE Advance) 90 scans) with scan time ranging between 40 and 120 min depending on radioligand scan protocol. Magnetic resonance imaging (MRI) T1-weighted images were acquired on 3T MR scanners (Magnetom Trio, Verio or Prisma; Siemens Healthcare Sector, Erlangen, Germany) and were segmented into grey matter, white matter and cerebrospinal fluid and co-registered with the PET images. PVElab (run on Matlab R2024a, Linux OpenSUSE 15.6, EU PVEout project (QLG3-CT-2000-00594), Copenhagen, Denmark, and Naples, Italy; https://nru.dk/pveout/) was used to automatically outline regions of interest from the structural MRI scan and extract PET time-activity curves for the frontal cortex. Reference Svarer, Madsen, Hasselbalch, Pinborg, Haugbøl and Frøkjær37
Outcome measure was defined as non-displaceable binding potential (BPND) for all tracers except [18F]Altanserin, for which we used binding potential (BPP). BPND was modelled with the simplified reference tissue model for [11C]AZ10419369, [11C]Cimbi-36 and [11C]SB207145 scans, employing cerebellum as a reference region, and was here defined as BPND = f ND×B avail×(1/ K D), where f ND is the free fraction of ligand in the non-displaceable tissue compartment, K D is the dissociation constant and B avail is the concentration of receptors available for binding. Reference Innis, Cunningham, Delforge, Fujita, Gjedde and Gunn38 For [11C]DASB scans, BPND was calculated using the Ichise multilinear reference tissue method 2 (MRTM2), Reference Ichise, Liow, Lu, Takano, Model and Toyama39 employing cerebellum as a reference region with the k 2’ clearance rate constant for cerebellum fixed by applying the MRTM2 on time-activity curve data from a composite high-binding region (volume weighted averages of thalamus, putamen and caudate). Lastly, for [18F]Altanserin, scan data was acquired with a bolus injection followed by continuous infusion to produce tracer steady state in the tissue and the blood. The outcome parameter was the BPP, defined as BPP = f P × B avail /K D, where f P is the free fraction of ligand in plasma. BPP was calculated as (C ROI – C ND)/C p, where C ROI and C ND are steady-state mean count density in the region of interest and the reference region (cerebellum), respectively; C P is the steady-state activity of non-metabolised [18F]Altanserin in plasma. Plasma from venous blood samples was used for the calculation of C P, including correction for radioactive metabolites passing the blood-brain barrier. Reference Adams, Pinborg, Svarer, Hasselbalch, Holm and Haugbøl40 C P was estimated using high-performance liquid chromatography as described by Pinborg et al. Reference Pinborg, Adams, Svarer, Holm, Hasselbalch and Haugbøl41 For a more detailed overview of scanner types and scan protocols, please see Supplementary Table 1.
Statistical analyses
We used multiple linear regression models to assess the association between working memory performance and frontal cortex binding for each of the three 5-HT receptors and the 5-HTT. For the 5-HT4R and 5-HTT analyses where patient data was also available, an interaction term (clinical status by binding) was included to test whether the association between working memory performance and frontal cortex binding differed between healthy individuals and patients with mood disorders.
All models were corrected for age, gender, injected radioligand mass per kilogram and, when relevant, PET scanner type (HRRT versus GE Advance). Major Depressive Inventory score, indexing depressive symptom severity, was also included in models for patients. Additionally, for the 5-HTT model, the serotonin transporter-linked promoter region (5-HTTLPR) genotype (s-carrier versus non-s-carrier) was included as a covariate, whereas radioligand type ([18F]Altanserin or [11C]Cimbi-36) was included as a covariate in the 5-HT2AR model. Significance threshold was set at α < 0.05 and all p-values are reported in uncorrected form.
Results
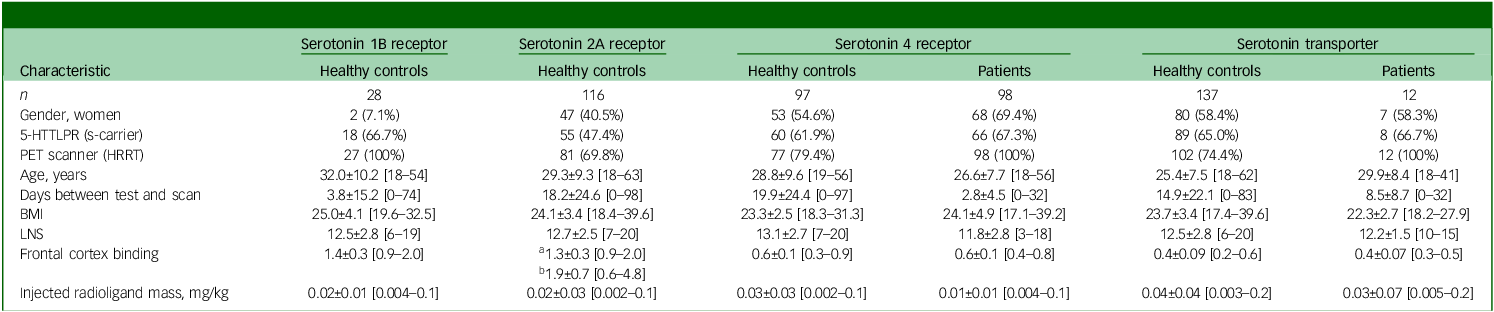
Table 1 provides demographic and neuroimaging characteristics of the study populations.
Descriptive and tracer information about participants

For continuous variables, mean ± s.d. and range is provided. Frontal cortex binding values represent non-displaceable binding potential (BPND), with one exception; for the serotonin 2A receptor group, two radioligand were used (a = [11C]Cimbi-36; b = [18F]Altanserin); for the [18F]Altanserin, binding potential (BPp) values are reported instead of BPND. 5-HTTLPR, serotonin transporter-linked promoter region; PET, positron emission tomography; HRRT, 3D High Resolution Research Tomography PET scanner; BMI, body mass index; LNS, letter-number-sequencing task.
Figure 1 shows associations between working memory scores and 5-HT1BR, 5-HT2AR, 5-HT4R and 5-HTT frontal cortex binding.
Association between working memory and serotonin brain markers. (a) Association between working memory and frontal serotonin 1B receptor binding (healthy individuals, n = 24). (b) Association between working memory and frontal serotonin 2A receptor binding (healthy individuals, n = 97). (c) Association between working memory and frontal serotonin 4 receptor binding (healthy individuals (teal circles), n = 89; patients (pink triangles), n = 89). (d) Association between working memory and frontal serotonin transporter binding (healthy individuals (teal circles), n = 122; patients (pink triangles), n = 11).

We found no evidence of an association between working memory capacity and 5-HT1BR (β = –4.83, p = 0.07) or 5-HT2AR (β = 0.20, p = 0.83) frontal cortex binding in healthy individuals. However, there was a significant interaction effect of clinical status on the association between 5-HT4R binding and working memory (p = 0.01), with patients showing a positive association (β = 6.74, p = 0.02) and healthy individuals showing a negative but statistically non-significant association (β = –3.59, p = 0.17). As a sensitivity analysis, we repeated the analysis for patients excluding patients with an SAD diagnosis, and found that the estimates for patients with MDD only remained robust (n = 81, β = 6.42, p = 0.03).
We found no evidence of a group effect on the association between working memory scores and 5-HTT frontal cortex binding (p = 0.89), nor evidence of a main effect across groups (β = 1.20, p = 0.77). As a final sensitivity analysis, we repeated all analyses with data from participants where the time between PET scan and cognitive testing was ≤30 days (see Supplementary Table 2); the results did not differ substantially. Gender was a significant covariate in the adjusted models for 5-HT2AR (p = 0.002) and 5-HT4R (p = 0.001).
Discussion
We report, for the first time, molecular brain imaging with PET to investigate the association between working memory capacity and multiple features of the 5-HT system in healthy individuals and patients in a depressed state. We observed a positive association between frontal cortex 5-HT4R binding and working memory performance in patients with mood disorders. In contrast, we found no evidence to support a direct link between serotonergic neurotransmission and working memory in healthy individuals for the 5-HT1BR, 5-HT2AR, 4-HT4R or 5-HTT in either healthy individuals or patients with MDD.
Until now, research into the role of 5-HT signalling in human working memory has been exceedingly sparse. Reference Khan and Muly18 The present PET cohort study is therefore not only one of the largest studies to investigate the molecular underpinning of any cognitive domain to date, but also the first to provide comprehensive insight into 5-HT contribution to human working memory.
We found no evidence that healthy-state variations in any of the 5-HT systems we investigated influence working memory performance. Yet, low 5-HT4R levels was associated with poorer performance in patients with mood disorders. Interestingly, the 5-HT4R has been highlighted as a promising therapeutic target in MDD, Reference Rebholz, Friedman and Castello42,Reference Frokjaer43 particularly for cognitive symptoms. Reference Murphy, de Cates, Gillespie, Godlewska, Scaife and Wright44 Pharmacological activation of the 5-HT4R has been shown to enhance hippocampus-dependent learning and memory in rodents, Reference Pascual-Brazo, Castro, Díaz, Valdizán, Pilar-Cuéllar and Vidal45 whereas intervention with the 5-HT4R partial agonist prucalopride has produced similar memory-enhancing effects in healthy individuals. Reference de Cates, Wright, Martens, Gibson, Türkmen and Filippini46,Reference Murphy, Wright, Browning, Cowen and Harmer47
Further, PET neuroimaging studies have shown associations between global 5-HT4R binding and verbal memory performance in both healthy individuals Reference Haahr, Fisher, Holst, Madsen, Jensen and Marner48,Reference Stenbæk, Fisher, Ozenne, Andersen, Hjordt and McMahon49 and unmedicated patients with MDD, Reference Köhler-Forsberg, Dam, Ozenne, Sankar, Beliveau and Landman21,Reference Dam, Köhler-Forsberg, Ozenne, Larsen, Ip and Jorgensen50 with the latter constituting a subgroup of the current study. We also recently reported that antidepressant-free patients with a moderate to severe depressive episode have around 7% lower cerebral 5-HT4R binding compared with healthy individuals, Reference Köhler-Forsberg, Dam, Ozenne, Sankar, Beliveau and Landman21 aligning with previous data showing that healthy individuals with a high familial risk of MDD also have lower striatal 5-HT4R binding. Reference Madsen, Torstensen, Holst, Haahr, Knorr and Frokjaer51 Although baseline 5-HT4R binding is typically interpreted as receptor availability, it has also been suggested to reflect longer term serotonergic tone, Reference Haahr, Fisher, Jensen, Frokjaer, Mahon and Madsen33 and thus group differences in MDD may reflect serotonergic dysregulation rather than solely altered receptor density. Together this indicates that altered 5-HT4R levels is part of depressive pathology, although it remains unclear whether low 5-HT4R is a driver of depressive symptoms, a compensatory mechanism or a downstream effect of other depression-related brain processes. In any case, our findings indicate that although variations in the 5-HT4R receptor settings are not directly linked to working memory functions in the healthy state, it becomes significant when 5-HT4R levels are pathologically altered. One explanation is that a still unidentified common upstream factor influences both 5-HT4R levels and working memory functioning. Alternatively, depression-related alterations in 5-HT4R signalling may modulate other neurotransmitter systems involved in working memory such as the dopamine system and thereby negatively affect working memory function. In support of this notion, Zhou et al Reference Zhou, Liang, Salas, Zhang, De Biasi and Dani52 showed that pharmacologically manipulating synaptic 5-HT levels through intervention with the selective serotonin reuptake inhibitor (SSRI) fluoxetine altered the spatial and temporal interaction between serotonergic and dopaminergic signalling in the striatum. The striatum together with the frontal cortex make up the frontostriatal circuit, which is crucial for cognitive control and adaptive manipulation of information during working memory tasks. Reference Chatham, Frank and Badre53 Interestingly, two independent PET studies have found that striatal D1 receptor binding is decreased in patients with MDD. Reference Cannon, Klaver, Peck, Rallis-Voak, Erickson and Drevets54,Reference Dougherty, Bonab, Ottowitz, Livni, Alpert and Rauch55 It is therefore plausible that striatal cross-talk between serotonergic and dopamine signalling could influence working memory in mood disorders, although future studies are needed to test this hypothesis.
Importantly, by establishing a link between 5-HT4R signalling and working memory function in mood disorders, our findings also highlight the potential of targeting the 5-HT4R as a therapeutic strategy for addressing working memory deficits in patients with mood disorders. In support of this, de Cates et al found that pharmacological stimulation of the 5-HT4R in healthy individuals enhanced the functional connectivity between brain networks involved in goal-oriented processes necessary for working memory. Reference de Cates, Wright, Martens, Gibson, Türkmen and Filippini46 In addition, there is evidence from animal models that direct stimulation of the 5-HT4R may increase 5-HT tonus in the brain by stimulating 5-HT-producing neurons in the dorsal raphe nuclei, which project to the rest of the brain, including the limbic system and the cerebral cortex. Reference Faye, Hen, Guiard, Denny, Gardier and Mendez-David56 Interestingly, we recently showed that 5-HT4R levels decrease over the course of SSRI treatment in patients with MDD, and that the patients who exhibit the largest fall in 5-HT4R also show the least improvement in cognitive symptoms. Reference Dam, Köhler-Forsberg, Ozenne, Larsen, Ip and Jorgensen50 Together, these findings provide a strong rationale that direct 5-HT4R stimulation holds promise as an augmentative strategy for treating cognitive symptoms in mood disorders.
In conclusion, we provide the first evidence to suggest that 5-HT4R signalling may be involved in disrupted working memory function in patients with mood disorders. Our findings help point to new avenues of research into targeted pharmacological treatment of cognitive dysfunction in MDD, and highlight the importance of assessing molecular mechanisms in cognitive functions in both the healthy and pathological state, as these mechanisms appear state specific.
Methodological considerations
Although our study is one of the largest PET studies to investigate molecular neurotransmission and cognitive functioning to date, there are several important methodological considerations that should be mentioned. First, we only looked at 5-HT1BR, 5-HT2AR, 5-HT4R and 5-HTT, and so did not cover all key 5-HT receptor systems that could be directly or indirectly involved in working memory functioning. Second, the healthy participant sample size for the 5-HT1BR and the patient sample size for the 5-HTT were both very low, and we therefore cannot exclude that we have missed a true association because of low statistical power. Third, we only had patient data for the 5-HT4R and 5-HTT systems; future studies are therefore needed to determine if the association between 5-HT signalling and working memory in patients with mood disorder is unique to the 5-HT4R or is systemic across different 5-HT receptors. Fourth, the use of different scanners and PET scan protocols across the different pooled studies may have introduced noise and obscured small effects, although this was partly mitigated by correcting for scanner type in the analysis. Finally, although SAD and MDD share core symptoms and treatment approaches, differences in underlying mechanisms mean that pooling them may introduce some variability. However, excluding patients with SAD in a sensitivity analysis had minimal impact on the 5-HT4R–working memory association, reducing concern about diagnostic heterogeneity. Future studies should explore diagnosis-specific effects more directly.
Supplementary material
The supplementary material is available online at https://doi.org/10.1192/bjo.2026.11045
Data availability
All study data are available upon request from the Cimbi database at Neurobiology Research Unit, Copenhagen University Hospital Rigshospitalet (https://nru.dk/).
Acknowledgements
We would like to thank all the participants for their time. In addition, we thank Lone Freyr and Peter Jensen for their assistance with the data collection and data management.
Author contributions
V.H.D. conceived and designed the study, conducted statistical analyses and wrote the first draft. A.J., N.G., C.S., G.M.K. and V.G.F. contributed to data collection and consulted on neuroimaging methods. A.J., C.S. and V.G.F. consulted on the statistical analysis, and K.W.M. and D.S.S. consulted on the interpretation of the findings. All authors contributed to manuscripts revision, read and approved the submitted version and agreed to be held accountable for all aspects of the work.
Funding
The study was supported by the Independent Research Fund Denmark (grant: 1057-00052B). We would also like to acknowledge and thank The John and Birthe Meyer Foundation for the donation of the Cyclotron and PET scanner.
Declaration of interest
V.H.D. has received honorarium as a speaker for H. Lundbeck Pharma. V.G.F. has received honoraria for consulting or as a speaker from SAGE Therapeutics, Lundbeck Pharma A/S, Janssen Cilag A/S, Gedeon-Richter A/S and Ferring Pharmaceuticals. G.M.K. has served as consultant for Onsero, Gilgamesh, Pure, Pangea and Sanos, and additionally served as speaker for Abbvie, Angelini and H. Lundbeck. K.W.M. has served as consultant for Janssen and Angelini. All other authors have no conflicts of interest.




 Open access
Open access
eLetters
No eLetters have been published for this article.