

The lesser rhea Rhea pennata pennata is a large flightless herbivorous bird (height 1.2 m; weight 15–25 kg) that inhabits the steppes of Argentine Patagonia and southern Chile (Del Hoyo et al., Reference Del Hoyo, Elliot and Sargatal1992). Its reproductive system combines harem polygyny and sequential polyandry; males incubate the eggs and care for the chicks until early winter (Sarasqueta, Reference Sarasqueta1997; F. Barri, pers. obs.).
The species is categorized as Near Threatened on the IUCN Red List (IUCN, 2007), and considered ecologically extinct in parts of its range (Novaro et al., Reference Novaro, Funes and Walker2000). Although rheas are protected throughout Argentina, intense illegal hunting and egg harvest pressure and habitat alterations may be the causes of the decline of both the greater (Rhea americana; Bellis et al., Reference Bellis, Martella and Navarro2004) and lesser rhea (Bellis et al., Reference Bellis, Navarro, Vignolo and Martella2006). Additionally, both species are farmed commercially for meat, leather and feathers (Martella & Navarro, Reference Martella and Navarro2006). Despite the conservation status of natural populations of lesser rheas, most available information about their demographic parameters refers to captive populations (Sarasqueta, Reference Sarasqueta1997; Navarro & Martella, Reference Navarro and Martella2002), and data on wild populations are scarce.
Considering that knowledge of demographic variables and the mechanisms that regulate wild populations is essential to develop effective plans for the conservation and/or management of threatened species (Sutherland et al., Reference Sutherland, Pullin, Dolman and Knight2004; Butchart et al., Reference Butchart, Stattersfield and Collar2006), the objective of this study was to evaluate the impact of hunting, egg harvest and livestock grazing intensities on density and reproductive success of wild lesser rhea populations. We studied lesser rhea during two successive reproductive seasons (August–March, 2004–2005 and 2005–2006), in three adjacent ranches (1, 7,400 ha; 2, 3,200 ha; 3, 3,400 ha) in a homogeneous habitat of north-west Patagonia, in the Pilcaniyeu region, Río Negro province, Argentina (Fig. 1).
The location of the three ranches in the Pilcaniyeu region (a rural area of the north-western Patagonia steppe). The solid rectangle on the inset map indicates the location of Pilcaniyeu in Argentina.
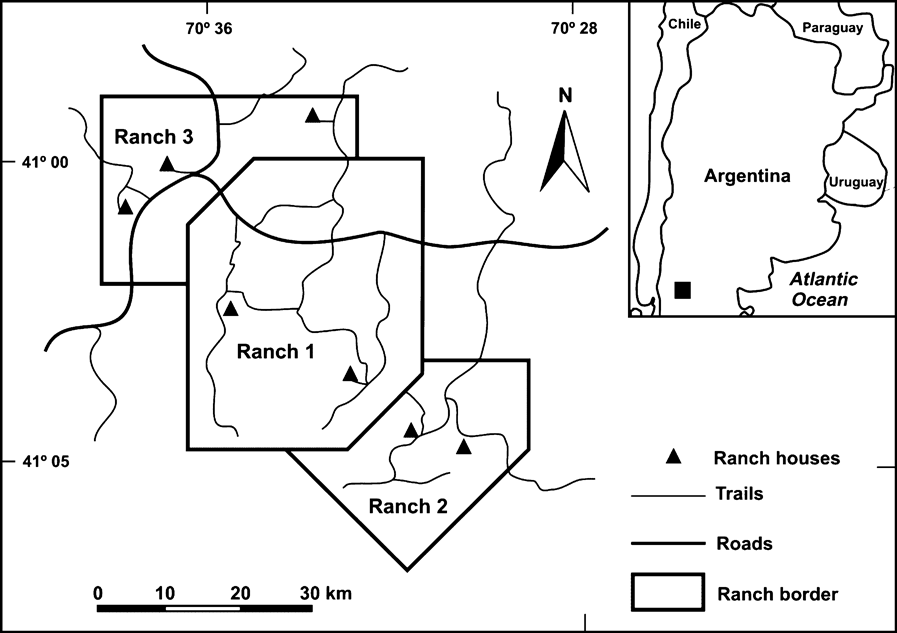
On these ranches there is extensive production of merino sheep and, to a lesser extent, goats, cows and horses. These are the most common farming activities on the Patagonia steppe, where agriculture is largely absent (Somlo et al., Reference Somlo, Pelliza, Willems, Nakamatsu and Manero1997). As in other ranches on the Patagonia steppe (Bellis et al., Reference Bellis, Martella and Navarro2004) all the study paddocks were available to rheas because they can cross the six-wire fences delimiting plots and ranches (F. Barri, pers. obs.). The three ranches differed in their livestock density and in the intensity of use of lesser rheas (i.e. hunting pressure and egg harvest). On ranch 1 there is sustainable land management comprising a livestock density <0.25 sheep per ha, which prevents overgrazing (Bonvissuto & Somlo, Reference Bonvissuto and Somlo1998), grasses are the predominant vegetation, and vegetation biomass production is high (500–750 kg ha-1 yr-1). On ranches 2 and 3 there is a high density of livestock (>1 sheep per ha) and consequently lower vegetation biomass production (300–400 kg ha-1 yr-1) and increased shrub encroachment (Bonvissuto & Somlo, Reference Bonvissuto and Somlo1998; F. Barri, pers. obs.). On ranch 1 rheas are actively protected by the owner. On ranch 2 there is moderate use of rheas (≤5 rheas hunted per year; ≤1 nest harvested per year). On ranch 3 there is intensive use of the species (≥10 rheas hunted per year and harvesting of all nests found). This information was obtained by surveys of landowners and workers (on average 4 surveys per ranch). The answers obtained were cross-checked weekly during the research by seeking for evidence of individuals hunted and/or eggs harvested.
Ranch workers in the study area each own 1–3 dogs, which are used mainly for the management of cattle. As there is approximately one worker per 1,000 ha, the density of dogs is ≤ 0.3 dogs km2. They are usually maintained in the vicinity of the houses, and only go to the field with their owners. For this reason, encounters between dogs and rheas are infrequent. During the study we recorded only two such events, with non-serious consequences for the rheas (F. Barri, pers. obs.).
In both reproductive seasons rheas were counted by line transect sampling conducted weekly by truck (at 10–20 km h-1) throughout the available internal roads and trails of the study area. This time interval guaranteed independent sampling because it enabled rheas to move amongst areas. The length of each transect was 10 km, totalling 380 km in 2004–2005 and 420 km in 2005–2006. We also carried out general surveys covering the entire study area by systematic daily inspections on foot or horseback, to record the number of active nests, eggs per nest, hatched eggs, and chick survival. The final recruitment (number of individuals that reached the juvenile stage) was recorded after the winter in August.
The density of rheas was estimated for each ranch using the software Distance v. 4.1 (Thomas et al., Reference Thomas, Laake, Derry, Buckland, Borchers and Anderson2001) and compared amongst ranches by a Kruskal-Wallis test and a multiple comparison a posteriori test. Nest densities, clutch sizes and brood sizes were compared by Mann-Whitney tests between the two ranches that exhibited reproductive events, and hatching success, chick survival and final recruitment were compared with χ2 tests. In both years ranch 3, with intensive hunting and egg harvest of lesser rheas, had the lowest density of rheas and no nests were recorded. Densities of rheas and nests did not differ between ranches 1 and 2 (Table 1).
Density (± SE) of individuals and nests of lesser rhea during two breeding seasons (2004–2005 and 2005–2006) on three ranches in north-west Patagonia (Fig. 1), and probabilities from Kruskal-Wallis tests of differences amongst ranches and amongst years.
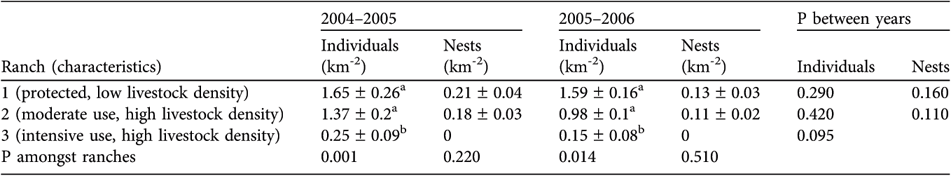
a,b Different superscripted letters indicate significant differences within columns (at P<0.05; multiple comparison a posteriori Kruskal-Wallis test)
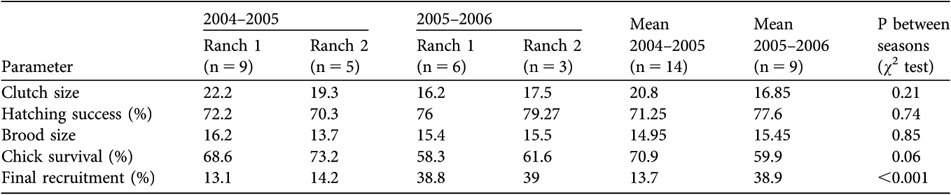
On the two ranches (1 and 2) where nests were observed there was no difference in clutch size, hatching success, brood size, chick survival, and final recruitment (Table 2). Differences between years (both ranches pooled) were also not significant for all reproductive parameters except final recruitment, which was higher in the second reproductive season (2005–2006).
Demographic parameters of wild lesser rheas during two breeding seasons (2004–2005 and 2005–2006), on ranches 1 and 2 (Fig. 1), and averaged for the two seasons, and probabilities from a χ2 test of averaged parameters between seasons.
The density and reproductive parameters we recorded are within the known ranges for lesser rheas (Navarro et al., Reference Navarro, Cardón, Manero and Clarke1999; Navarro & Martella, Reference Navarro and Martella2002). At ranch 1, with no hunting, density was similar to that observed by Bellis et al. (Reference Bellis, Navarro, Vignolo and Martella2006) in another Patagonian ranch where rheas are protected, showing the potential contribution that protected rural areas of Patagonia have for maintaining populations of lesser rheas. Hatching success and initial chick survival were generally higher than those recorded in captive populations of this species (Navarro & Martella, Reference Navarro and Martella2002), suggesting that productivity on lesser rhea farms could be improved. Our results show that hunting and egg harvesting appear to reduce the density of lesser rheas and their reproductive success. We did not record reproductive events on the ranch with intense use of the species and we therefore conclude that the risk of local extinction is increased in such areas.
Reproductive parameters of lesser rhea populations seem not to be directly affected by overgrazing, at least at the levels observed in this study, which are common in most of Patagonia (Somlo et al., Reference Somlo, Pelliza, Willems, Nakamatsu and Manero1997). This is probably a consequence of the low overlap in diet composition between rhea and domestic livestock. Sheep eat mostly grasses (mainly Festuca spp.), whereas the diet of rheas mainly includes herbs (35%, such as Cichorium spp.) and shrubs (45%, such as Mulinun spinosum), which are less affected by overgrazing, and under intense overgrazing pressure rheas can expand consumption of shrubs up to 75% (Somlo et al., Reference Somlo, Pelliza, Willems, Nakamatsu and Manero1997; Bonvissuto & Somlo, Reference Bonvissuto and Somlo1998).
Higher summer precipitation and winter temperatures improve production of vegetation biomass (Bonvissuto & Somlo, Reference Bonvissuto and Somlo1998), which in turn enhances chick survival because of the higher probability of reaching the winter in a better body condition (Sarasqueta, Reference Sarasqueta1997). Therefore, more favourable climatic conditions (i.e. higher temperatures during critical stages of chick development and more rainfall, which enhances food availability before winter) could be responsible for the higher final recruitment observed in the second reproductive season: monthly averages in 2004–2005 were 0.3 mm summer rainfall (December to February) and 1.3ºC winter temperatures (June–August), whereas averages in 2005–2006 were 31.7 mm and 2.3ºC, respectively (Bustos, Reference Bustos2006).
Our results suggest that the decrease of wild lesser rhea populations on the Patagonian steppe (Novaro et al., Reference Novaro, Funes and Walker2000) is more influenced by hunting and egg harvest than by overgrazing. We conclude that, as with other ratite species (Pople et al., Reference Pople, Cairns and Grigg1991; Bellis et al., Reference Bellis, Martella and Navarro2004), the lesser rhea could coexist with traditional livestock raising, as long as suitable habitats for reproduction (flooded areas locally called mallines) are not disrupted (Bellis et al., Reference Bellis, Navarro, Vignolo and Martella2006; F. Barri et al., unpubl. data). Effective conservation of the lesser rhea in Patagonia largely depends on the effective prevention of illegal uses of the species and the development of sustainable harvest programmes.
Acknowledgements
We are grateful to EEA INTA Bariloche, N. Roldan, J. Von Thüngen, D. Sarasqueta and the personnel in Pilcaniyeu who collaborated during the fieldwork. The map of the study area was provided by the Laboratorio de Teledetección y SIG from the EEA INTA Bariloche. L. Bellis provided comments on the manuscript. Financial support was provided by Secretaría de Ciencia y Técnica of the Universidad Nacional de Córdoba, and the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). FRB is a fellow and MBM and JLN are researchers at CONICET.
Biographical sketches
Fernando R. Barri has been working on the management and conservation of greater and lesser rheas since 2001, and is currently studying population dynamics of reintroduced wild guanaco Lama guanicoe in a national park in the central mountains of Argentina. Mónica Martella's main research interests are the ecology, behaviour, management and conservation of birds, and since 1990 she has focused on rheas. She is presently Director of a research project on the management and conservation of rheas in Argentina. Joaquín L. Navarro has been working in ecology and management of other birds and vertebrates. He is currently joint Director of the project on rhea conservation, and responsible for studies of sustainable use, captive breeding and farming production of both species of rheas.



