


Non-technical Summary
In a shallow Ordovician sea, the shell of a brachiopod called Porambonites wesenbergensis became a host for a variety of smaller organisms, creating a bustling, miniature ecosystem. Some areas of the shell were covered in thin layers of an ancient problematic organism called Rothpletzella, preserved as molds. This organism, which harnessed sunlight to produce energy, lived alongside other creatures such as craniid brachiopods and bryozoans, tiny colonial animals that filtered food from the water. In addition, tiny tube-like creatures resembling Allonema formed small clusters on the shell. This discovery sheds new light on how diverse life was in ancient communities that grew on hard surfaces, such as shells, during the Ordovician period. It suggests that these communities may have been more complex than scientists originally believed, with different organisms playing various ecological roles. It is still unclear whether these creatures settled on the brachiopod’s shell while it was alive or after it died. The Rothpletzella and filter feeders did not seem to interfere with each other, as the algae did not compete for the same food as the filter feeders. Instead, the algae likely attracted grazers such as early snails, which may have left marks on the shell as they fed on the Rothpletzella.
Introduction
Hard substrates are commonly inhabited by various sclerobionts—organisms that live on or bore into hard surfaces, often using shells of other organisms as substrates (Taylor and Wilson, Reference Taylor and Wilson2002, Reference Taylor and Wilson2003). The most common hosts for sclerobionts are typically benthic suspension feeders with biomineralized skeletons, such as brachiopods, pteriomorph bivalves, crinoids, and hyolithids (e.g., Feldman and Brett, Reference Feldman and Brett1998; Galle and Parsley, Reference Galle and Parsley2005; Głuchowski, Reference Głuchowski2005; Barclay et al., Reference Barclay, Schneider and Leighton2013, Reference Barclay, Schneider and Leighton2015; Schneider, Reference Schneider2013; Freeman Peters et al., Reference Freeman Peters, Schneider and Leighton2024), as well as stromatoporoids and corals (e.g., Kershaw, Reference Kershaw1980; Baird and Brett, Reference Baird and Brett1983; Lebold, Reference Lebold2000). Both living and dead substrates were encrusted with organisms. However, sclerobionts also often colonized mobile organisms, such as echinoids, trilobites, and cephalopods, although these records are less common than compared with immobile substrates (Baird et al., Reference Baird, Brett and Frey1989; Brandt, Reference Brandt1996; Taylor and Brett, Reference Taylor and Brett1996; Key et al., Reference Key, Schumacher, Babcock, Frey, Heimbrock, Felton, Cooper, Gibson, Scheid and Schumacher2010; Borszcz et al., Reference Borszcz, Kuklinski and Zatoń2013).
Sclerobionts serve as a valuable tool for paleoecological studies as the communities they form are almost always preserved in situ, providing a unique insight into ancient ecosystems (e.g., Freeman Peters et al., Reference Freeman Peters, Schneider and Leighton2024). From the perspective of encrusting organisms, each host acts as a patch or “island,” allowing the analysis of individual sclerobiont assemblages on a single host as a community (Brett, Reference Brett1988). Although already present since the early Cambrian (Palmer, Reference Palmer1982; Brett et al., Reference Brett, Liddell and Derstler1983), many common sclerobiont taxa originated during the Ordovician (Lescinsky, Reference Lescinsky and Copper1996; Ma et al., Reference Ma, Taylor and Buttler2021), during or after the Great Ordovician Biodiversification Event (GOBE). It is essential to understand the evolutionary patterns and ecological distribution of sclerobiont communities during the Ordovician (Freeman Peters et al., Reference Freeman Peters, Schneider and Leighton2024) to better comprehend their role in the GOBE. The fossilization potential of encrusting sclerobionts varies depending on whether they possessed mineral skeletons or were lightly mineralized or even entirely soft-bodied. The latter types of taxa were usually not preserved, and when soft-tissue sclerobionts are preserved, it is usually through bioimmuration (Taylor, Reference Taylor1990; Wilson et al., Reference Wilson, Palmer and Taylor1994).
One exceptional example concerns Rothpletzella. This calcareous microproblematicum has previously been considered either a cyanobacterium or a green alga, although its exact biological affinities remain uncertain (Riding, Reference Riding and Riding1991). Sphaerocodium, first described by Rothpletz (Reference Rothpletz1890) from the rocks of Wenlock age from Gotland Island (Sweden), was later identified by Wood (Reference Wood1948) as an intergrowth of Rothpletzella and Wetheredella (Riding and Fan, Reference Riding and Fan2001, p. 804). During most of its research history, Rothpletezella was known exclusively from thin sections where it appeared as calcareous, bifurcating tubular filaments forming flat or undulose sheet-like layers (Zhou and Pratt, Reference Zhou and Pratt2019; Zatoń and Jarochowska, Reference Zatoń and Jarochowska2020). Recently, well-preserved three-dimensional Rothpletzella sheets encrusting brachiopod shells were discovered from the Upper Devonian of Russia (Zatoń and Jarochowska, Reference Zatoń and Jarochowska2020). Since then, well-preserved epibiotic Rothpletzella have also been found on Middle Devonian corals (Zatoń and Wrzołek, Reference Zatoń and Wrzołek2020) and brachiopods (Brychcy et al., Reference Brychcy, Zatoń, Nawrot, Halamski and Rakociński2023) from Poland, as well as on Middle Devonian brachiopods from Morocco (Zatoń et al., Reference Zatoń, Nawrot, Jakubowicz, Ernst, Rakociński, Berkowski and Belka2022). These records suggest that Rothpletzella may, in fact, be more common as an encruster of different biogenic substrates than previously thought (Zatoń et al., Reference Zatoń, Nawrot, Jakubowicz, Ernst, Rakociński, Berkowski and Belka2022); however, it could have been easily overlooked earlier due to its faint appearance and minute size.
Here we describe older, Upper Ordovician fossils from Estonia, assigned to Rothpletzella, and discuss their preservation and paleoecology.
Geological background and locality
During the Late Ordovician, the region that is today northern Estonia was submerged beneath a broad, generally warm, and shallow epicontinental sea. The conodont-based δ18Ophos studies suggest that the pre-Hirnantian Late Ordovician was characterized by a general cooling trend: a rapid climate cooling in the early Sandbian and a less intense cooling in the late Sandbian through the Katian (Männik et al., Reference Männik, Lehnert, Nõlvak and Joachimski2021). This extensive marine environment facilitated the deposition of a remarkably complete stratigraphic sequence, with all Ordovician stages well represented—predominantly by carbonate sedimentary rocks, which reflect the prevailing depositional conditions (Nõlvak, Reference Nõlvak, Raukas and Teedumäe1997). Throughout this time, the paleocontinent of Baltica experienced a profound climatic and paleogeographic transformation as it gradually migrated from the cool, high southern latitudes toward the equatorial belt (Torsvik and Cocks, Reference Torsvik and Cocks2013). This latitudinal drift ushered in a significant climatic warming, which in turn stimulated an upsurge in carbonate sedimentation—a response typical of tropical marine settings (Nestor and Einasto, Reference Nestor, Einasto, Raukas and Teedumäe1997).
The onset of these tropical conditions became increasingly evident in the fossil record during the early Katian, marked by the emergence of warm-water faunal elements, such as tabulate corals and stromatoporoids, which are often found in non-reefal settings (Nestor and Einasto, Reference Nestor, Einasto, Raukas and Teedumäe1997). In northeastern Estonia, the Oandu Regional Stage is particularly notable for its lithological composition, comprising clay-rich limestones and marls associated with the Hirmuse Formation (Hints and Meidla, Reference Hints, and Meidla, Raukas and Teedumäe1997). These fine-grained sediments were deposited in a deeper part of the carbonate ramp—yet still within the sunlit, photic zone—suggesting a relatively tranquil depositional environment with adequate light penetration to support benthic life. It is within these layers that the large shells of the brachiopod Porambonites are found (Nestor and Einasto, Reference Nestor, Einasto, Raukas and Teedumäe1997).
Rakvere swimming pool (59.34014°N, 26.361601°E) is currently a non-existing historical locality in the town of Rakvere (Fig. 1). The Rakvere outdoor swimming pools were built in 1937, adjacent to Soolikaoja. The swimming pool was excavated in the limestone of the Oandu Regional Stage. From the excavation site, Armin Öpik made a rich collection of fossils that included brachiopods, bryozoans, trilobites, rugosans, and gastropods (Rõõmusoks, Reference Rõõmusoks1970). The studied specimen originates from the historical collection of A. Öpik.
Locality map with Rakvere shown.

Material and methods
Teichert (Reference Teichert1930) described the brachiopod Porambonites wesenbergensis from an unknown site located in the southeast part of Rakvere. The described specimen originates from the excavation of the Rakvere swimming pool. The Estonian collection material of Porambonites wesenbergensis (141 specimens housed at the Natural History Museum, University of Tartu, and 31 specimens housed at Tallinn University of Technology) demonstrates that the species is more common in the sediments of the Oandu Regional Stage (112 specimens). The whole collection was examined, and a single specimen with Rothpletzella was found. P. wesenbergensis (width 38–50 mm, length 35–50 mm) is a large brachiopod relative to other brachiopod species from the Oandu Regional Stage. They are often encrusted by bryozoans and craniids; their surface may also be bioeroded. Seventy percent of P. wesenbergensis specimens are encrusted. As pentamerids have very thin shells (Balthasar et al., Reference Balthasar, Jin, Hints and Cusack2020), the specimens of P. wesenbergensis are quite often damaged. A single, densely encrusted brachiopod specimen from the Oandu Regional Stage (the collection number TUG 1766-136) was selected for a detailed scanning electron microscopy (SEM) study of microencrustation patterns. The specimen was prepared for imaging by coating it with a thin conductive layer of gold and palladium, ensuring optimal surface conductivity and image resolution. SEM analyses were conducted using TESCAN VEGA II and TESCAN VEGA III instruments at the Palaeontological Institute of the Russian Academy of Sciences in Moscow. All micrographs were captured using a backscattered electron (BSE) detector, which enhances compositional contrast and facilitates the visualization of internal structural details. In addition to morphological observations, elemental composition data were obtained using energy dispersive X-ray spectroscopy (EDS/EDX), which utilized the INCA energy system integrated with the SEM platform. This allowed for the qualitative and semi-quantitative analysis of the specimen’s elemental constituents, providing essential insights into its mineralogical composition and diagenetic history. Before the SEM examinations, the brachiopod and encrusting faunas were photographed using a Canon EOS 5Dsr digital camera and an apochromatic zoom system (Leica Z16 APO) in the Department of Geology at Tallinn University of Technology.
For comparisons, we also analyzed similar fossils encrusting brachiopod shells from the Middle Devonian of Madéne el Mrakib, eastern Anti-Atlas, Morocco (for details, see Zatoń et al., Reference Zatoń, Nawrot, Jakubowicz, Ernst, Rakociński, Berkowski and Belka2022).
Repository and institutional abbreviation
The specimen is housed at the Natural History Museum, University of Tartu (TUG).
Results
Characteristics of Rothpletzella
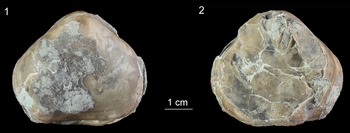
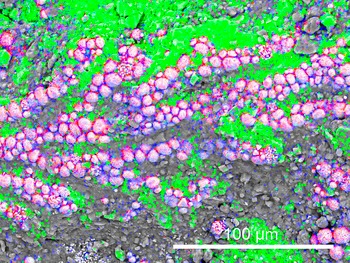
The study specimen of Porambonites wesenbergensis (Fig. 2) is encrusted in several places with thin sheets that exhibit a fan-shaped morphology (Figs. 3, 4). These fossils form either a single fan or several lobes that usually grow in the same general direction, forming a structure that is several millimeters in diameter. The fans are small, with a maximum width of 0.5 mm, and each individual lobe is typically around 0.2 mm wide. The surface of the lobes displays distinct longitudinal boundaries, which are slightly diverging and run almost parallel to each other, separating them into wider, closely spaced, branch-like structures. The branches are regularly split via bifurcation, creating secondary branches that further divide into two separate structures. These branches are either straight or gently curved, and they gradually widen until the bifurcation point, with maximal widths ranging from 18 to 38 μm. After bifurcation, the width of each branch ranges from 8 to 20 μm. The width of both the bifurcated branches and the single branches that follow the bifurcation shows considerable variation within the same specimen. The angle at which secondary branches emerge from the main branch ranges from 10° to 15°. The fossils are preserved as pyritized replacements of calcite. The EDS analysis reveals a high concentration of iron and sulfur in the branches of the studied specimens (Fig. 5). Pyrite is present in the form of small spheroids (Fig. 3.4).
Encrusted Porambonites wesenbergensis from the Oandu Regional Stage (lower Katian), Rakvere, northern Estonia (TUG 1766-136). (1) Ventral valve with sulcus. (2) Dorsal valve with fold.

Rothpletzella gotlandica from the Oandu Regional Stage (lower Katian), Rakvere, northern Estonia (TUG 1766-136-21). (1) General view of the sheet-like encruster. (2-3) Detail views of single branches. (4) Detail of a branch showing pyrite granules.

Rothpletzella gotlandica from the Oandu Regional Stage (lower Katian), Rakvere, northern Estonia (TUG 1766-136-21). (1-3) General views of the sheet-like encrusters. (4) Detail view of a single branch.

Elemental composition of Rothpletzella gotlandica from the Oandu Regional Stage (lower Katian), Rakvere, northern Estonia (TUG 1766-136-21). Green = calcium; red = sulfur; blue = iron.

Other encrusters and bioerosional traces
The large shell of P. wesenbergensis hosting Rothpletzella is also encrusted by three trepostome bryozoan colonies with sheet-like morphology (Fig. 6.1). In addition, 12 small craniid brachiopods are attached to the host’s shell. Eleven of them closely resemble specimens of the genus Petrocrania (Fig. 6.2). The surface of the brachiopod shell also contains a single Trypanites boring. Several small tubes with a smooth external surface and almost constant diameter form tiny aggregations (Fig. 6.3), somewhat resembling the ascodictyid Allonema (see, e.g., Jarochowska et al., Reference Jarochowska, Hierl, Vinn and Munnecke2016). The surface of the P. wesenbergensis shell also exhibits multiple tiny and shallow linear scratch marks, tentatively identified as Radulichnus (0.4–1.7 mm long and up to 0.26 mm wide; Fig. 6.4). These scratch marks were not observed on the surface of Rothpletzella.
Encrusters on Porambonites wesenbergensis. (1) Trepostome bryozoan (TUG 1766-136). (2) Small Petrocrania sp. near a larger Petrocrania specimen (TUG 1766-136-4). (3) Tubes of Allonema sp.(TUG 1766-136-17). (4) Scratch marks on the brachiopod exterior (TUG 1766-136).

Discussion
Affinity of the fossil
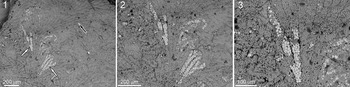
The studied fossils closely resemble those previously described from the Upper Devonian and interpreted as Rothpletzella (Zatoń and Jarochowska, Reference Zatoń and Jarochowska2020; Fig. 7). They are also similar to those reported from the Middle Devonian of Poland and Morocco (Zatoń and Wrzołek, Reference Zatoń and Wrzołek2020; Zatoń et al., Reference Zatoń, Nawrot, Jakubowicz, Ernst, Rakociński, Berkowski and Belka2022; Brychcy et al., Reference Brychcy, Zatoń, Nawrot, Halamski and Rakociński2023). The studied fossils match Rothpletzella in terms of size and the general architecture of sheets seen in thin section (i.e., fan shapes, branching; see, e.g., Zhou and Pratt, Reference Zhou and Pratt2019; Zatoń and Jarochowska, Reference Zatoń and Jarochowska2020). Like two-dimensional preservations of Rothpletzella, its branches are tightly packed, forming a coherent structure rather than loosely arranged filaments or filose growth. The main difference between the Upper Devonian, three-dimensionally preserved specimens and our material is the size of the branches, which are markedly larger in the Devonian specimens (the width of a single branch ranges from 37 to 88 μm). Although the width of the branches in the Middle Devonian specimens from Morocco is also larger (32 to 40 μm), the differences are smaller.
SEM microphotograph of Rothpletzella encrusting the Middle Devonian brachiopod shell from Morocco: (1) Fan-shaped colony with distinct bifurcation pattern. Some parts are pyritized (white arrows), while the rest of the thalli occur in the form of a calcareous sheath (black arrow). (2, 3) Magnified parts of the same colony showing pyritized bifurcating parts within the calcareous sheath.

Johnson (Reference Johnson1964) systematically compiled morphometric data used to delineate four distinct species within Rothpletzella. Among these, a maximum filament diameter of 40 μm was reported for both R. straeleni Lecompte, Reference Lecompte1936 and R. gotlandica Wood, Reference Wood1948. However, this species-level differentiation based on filament dimensions was later challenged by Riding and Soja (Reference Riding and Soja1993), who analyzed Rothpletzella specimens from Silurian deposits in Alaska. Their observations revealed a continuous range of filament widths, spanning from 13 to 37 μm, across specimens previously ascribed to multiple species. As a result, they advocated taxonomic consolidation, assigning all examined material to the type species R. gotlandica. Given that the dimensions of our specimens fall comfortably within this range, a tentative assignment to R. gotlandica appears justifiable.
The biological affinity of Rothpletzella has long been debated (Liu et al., Reference Liu, Wu, Yang and Riding2016, Reference Liu, Wu, Bao, Jiang, Zheng and Chen2021; Päßler et al., Reference Päßler, Jarochowska, Bestmann and Münnecke2018). It is frequently found within cyanobacteria (Zhou and Pratt, Reference Zhou and Pratt2019; Zatoń and Jarochowska, Reference Zatoń and Jarochowska2020), yet such classification raises contradictions, especially considering Devonian-aged specimens assigned to the genus. A comparison of the studied material with Girvanella is relevant as this calcified microfossil is widely regarded as a filamentous cyanobacterium and is commonly found in Paleozoic carbonates (e.g., Zhou and Pratt, Reference Zhou and Pratt2019). The filaments of Girvanella generally range from 10 to 40 μm in diameter, with most clustered around 20–30 μm (Riding, Reference Riding and Riding1991; Pratt, Reference Pratt2001), which is similar than the dimensions of the studied specimens. To analyze the phylogenetic affinities of Rothplezella, one should establish constraints on cyanobacterial cell size. While the upper limit for cyanobacterial cell diameter is observed in Oscillatoria—a free-living, non-branching freshwater genus reaching up to 100 μm—most large cyanobacteria, including Stigonema, do not exceed 60 μm in diameter (Nielsen, Reference Nielsen2006; Schulz-Vogt et al., Reference Schulz-Vogt, Angert and Garcia-Pichel2007). Although the dimensions of our specimens are consistent with cyanobacterial limits, their close morphological resemblance to larger forms from the Devonian makes the cyanobacterial affinities problematic. However, the size question cannot be separated from the temporal context. Microbial body sizes in the Ediacaran have often been significantly larger than their Phanerozoic counterparts, reflecting ecological and physiological differences across the Proterozoic–Phanerozoic transition (Knoll et al., Reference Knoll, Javaux, Hewitt and Cohen2006; Javaux, Reference Javaux2019). Thus, while Ediacaran microbial fossils include unusually large tubular and spheroidal forms, most Phanerozoic cyanobacteria, including calcified types, exhibit more constrained and typically smaller dimensions.
To address this discrepancy, Zatoń and Jarochowska (Reference Zatoń and Jarochowska2020) proposed that Rothpletzella was more plausibly a eukaryotic alga with a phototrophic lifestyle, possibly affiliated with the order Bryopsidales. This hypothesis accommodates both the filament size and the morphological complexity observed across different stratigraphic levels. Intriguingly, a temporal trend emerges in the size of Rothpletzella branches: the smallest forms are documented in Upper Ordovician and Silurian deposits (Johnson, Reference Johnson1964; Zatoń and Jarochowska, Reference Zatoń and Jarochowska2020), while progressively larger sizes are seen in Middle Devonian specimens, culminating in the largest dimensions during the Upper Devonian. By three-dimensional preservation similar to that of the Upper Devonian large forms, combined with dimensions like those of smaller Ordovician/Silurian representatives of Rothpletzella, the studied material provides additional evidence to support the identification of large Upper Devonian forms as Rothpletzella. This stratigraphic increase in branch diameter provides further support for the idea that these forms may represent a continuum or a series of closely related but distinct species that have evolved over time (Zatoń and Jarochowska, Reference Zatoń and Jarochowska2020). Moreover, one would expect the size evolution to proceed in the opposite direction, from larger to smaller, in the case of cyanobacterial affinities of Rothpletzella, to fit it into the general evolution of cyanobacteria from large Ediacaran forms to smaller forms in the Phanerozoic.
In conclusion, we admit that since the fossils consist only of the calcareous skeleton, or of pyrite coatings or infillings of this calcareous sheath, they do not provide any definitive information about cellular-level organization. Considering this, available evidence is broadly permissive of either a bacterial or algal affinity, without specifying any group of algae for the latter.
Preservation
Much like the specimens analyzed in this study, previously described Rothpletzella specimens from the Upper Devonian strata of Russia exhibit pyritization, as documented by Zatoń and Jarochowska (Reference Zatoń and Jarochowska2020). By contrast, related encrusting forms associated with Middle Devonian rugose corals from Poland are preserved in a distinctly different manner—manifesting as calcareous, thin laminae (Zatoń and Wrzołek, Reference Zatoń and Wrzołek2020). Similarly, Rothpletzella-like encrustations found on Middle Devonian brachiopods from Morocco show a mixed mode of preservation, with both calcareous and pyritized components present within the same specimens (Zatoń et al., Reference Zatoń, Nawrot, Jakubowicz, Ernst, Rakociński, Berkowski and Belka2022; Fig. 6). These variations in taphonomic pathways offer insights into the original biomineralization and post-depositional diagenetic history of these encrusting microorganisms. Rothpletzella possessed an originally thin, calcareous sheath, which was susceptible to early diagenetic dissolution, suggesting an original aragonitic or high-magnesium calcitic composition. However, the latter usually loses magnesium during diagenesis rather than suffer dissolution. In depositional settings where early mineralization did not occur or conditions favored dissolution—particularly in environments lacking rapid pyrite formation—these delicate thalli would likely have been poorly preserved or vanished from the fossil record altogether (Zatoń and Jarochowska, Reference Zatoń and Jarochowska2020).
Pyritization appears to have had a role in the preservation of Rothpletzella in certain sedimentary contexts. What is unusual is the spheroidal shape of the pyrite “crystals” and their apparent nucleation on the calcitic walls of Rothplezella (Brown, Reference Brown1996). After the burial of Rothplezella, sulfate-reducing bacteria respired organic matter and generated H₂S. Where dissolved Fe²⁺ was available, FeS was formed first and later converted into pyrite (FeS₂) via polysulfide or H₂S pathways (Thiel et al., Reference Thiel, Riedinger and Reimer2019; Duverger et al., Reference Duverger, Foucher, McLennan and Jensen2020). High supersaturation presumably caused the rapid nucleation of numerous microcrystals, which aggregated into raspberry-like spheroids. Extracellular polymeric substances (EPS) and microbial biofilms likely provided a template that promoted pyrite clustering (Borkow and Babcock, Reference Borkow and Babcock2003; MacLean et al., Reference MacLean, Scott, McDonald and Summons2008). The main question is why the pyrite nucleated on the walls of Rothpletzella. Presumably, its calcareous sheaths were coated with organic films that provided substrate for heterogeneous nucleation. Comparable wall-directed nucleation has been documented in the Ediacaran Conotubus, where pyritization initiates on the tube walls and progresses inward (Schiffbauer et al., Reference Schiffbauer, Xiao, Cai, Wallace, Hua and Wan2014). In semi-closed cavities (i.e., shell interiors), sulfide diffuses outward from decaying organics, while Fe²⁺ diffuses inward from pore waters. Their meeting zone at the wall produces FeS and eventually pyrite. This diffusion–precipitation behavior explains the rim-first mineralization observed in tube fossils and carbonate concretions (Schiffbauer et al., Reference Schiffbauer, Xiao, Cai, Wallace, Hua and Wan2014). The seawater was rich in sulfate and reactive iron (Thiel et al., Reference Thiel, Riedinger and Reimer2019). Moreover, microenvironments within carbonate shells often create reducing “mini-reactors” lined with pyrite (Tomasovych et al., Reference Tomasových, Berensmeier, Gallmetzer, Haselmair and Zuschin2021). A high abundance of pyrite-lined shells in the present-day northern Adriatic Sea has been interpreted as limited net exposure of labile tissues to O2 even when the seafloor is inhabited by abundant burrowing infauna (Tomasovych et al., Reference Tomasových, Berensmeier, Gallmetzer, Haselmair and Zuschin2021). There is a similarity between our material and pyrite-lined shells, and we hypothesize that the fossilization of Rothpletzella and the formation of pyrite-lined shells took place under similar geochemical conditions. Pyrite spheres have been described from the Devonian black shales of North America (Schieber and Baird, Reference Schieber and Baird2001), although the dimensions of these spheres as well as their paleoenvironment differ from those of Rothpletzella.
Paleoecology and environment
As mentioned, similarly preserved encrusting Rothpletzella specimens as described here have previously been reported so far only from a few Middle and Upper Devonian localities; in the Upper Ordovician, they are described for the first time. However, it is likely that they were not that rare; apart from their low fossilization potential, the fossils were simply overlooked due to their small size. Future studies should investigate the frequency of these encrusters in the Upper Ordovician. The paleoenvironment of our specimens was a shallow epicontinental sea with a soft carbonate mud bottom and depth range within the photic zone. The heavily encrusted shell of the Porambonites wesenbergensis reveals a high taxonomic diversity of epibionts. However, it is not clear whether the P. wesenbergensis was encrusted postmortem or during its life. The encrusters could represent a co-existing association, but the lack of spatial competition between the organisms makes it impossible to say that all the encrusters were alive at the same time. The phototrophic Rothpletzella did not compete with the filter feeders (i.e., brachiopods and bryozoans) for the food, although they may have preoccupied some surfaces. These, however, attracted possible grazers, which likely fed on Rothpletzella and left scratch marks on the shell surface, although the direct evidence in the form of Radulichnus crossing Rothpletzella is lacking.
Acknowledgments
O.V. was financially supported by the Institute of Ecology and Earth Sciences, the University of Tartu, the Estonian Research Council grant PRG2591, and a Sepkoski Grant from the Paleontological Society. We are grateful to G. Baranov, Department of Geology, Tallinn University of Technology, for photographing the specimens. We are also grateful to M.A. Wilson, B. Pratt, and associate editor S. LoDuca for constructive comments on the manuscript.
Competing interests
The authors declare none.





 Open access
Open access