




Introduction
Rock doves or common pigeons (Columba livia) have historically been, and in some world areas continue to be, kept as pets or as sources of meat and fertiliser. They are also used as symbols, appearing in the bible and in association with various gods and goddesses (Allen Reference Allen2009: 57–87). Despite their importance to many societies, their long history of coexistence with humans and the identification of C. livia bones at some archaeological sites, the origin and development of this close relationship and the eventual domestication of the rock dove have yet to be explored in detail (Yeomans Reference Yeomans2025). Probably among the earliest birds to be brought into the domestic sphere, rock doves likely took the commensal pathway to domestication, living alongside and scavenging from human communities for many generations (Kabir Reference Kabir2025). However, the when, where and how of C. livia domestication is yet unknown.
Pigeon bones found in caves in Gibraltar associated with Neanderthal and anatomically modern human activity suggest that they were exploited as far back as 67 000 years ago (Blasco et al. Reference Blasco2014). Unequivocal evidence for domestication appears only in the first millennium BCE, when stone structures—columbaria—were constructed for nesting pigeons, although depictions of birds found in Iraq possibly suggest domestication was in process much earlier, c. 4500 BP (Yeomans Reference Yeomans2025). The earliest assemblage of pigeon bones identified as domesticated comes from Hellenistic Nea Helos in Greece (fourth–first centuries BCE) (Serjeanston Reference Serjeantson2009: 306–307).
Other than indirect evidence, such as the presence of fledgling bones and pigeon towers, it is difficult to differentiate between C. livia and C. livia f. domestica based on bone morphology alone (Jonsson Reference Jonsson and Åström1983; Boessneck Reference Boessneck1995; Studer Reference Studer and Prummel2008). The osteomorphology of wild and domestic pigeons is almost identical (Johnston Reference Johnston1992). It is generally accepted that pigeons increased in size with domestication, but while this is certainly true for modern specimens, early examples are difficult to identify (Johnston Reference Johnston1992), in part because pigeons show a great deal of morphological plasticity (Boessneck Reference Boessneck1995). Pigeons, especially domestic pigeons, show more physical variation than any other bird species, depending on environmental parameters and management styles, rendering size and general morphology ineffective as criteria for the identification of domestication (Kabir Reference Kabir2025). Moreover, frequent hybridisation between domesticated and wild populations confounds morphological analyses and suggests that interpretations of domestic status must also examine behavioural and lifestyle patterns.
This study analyses the abundant pigeon remains found at Late Bronze Age (1650–1050 BCE) Hala Sultan Tekke on Cyprus to assess the domestication status, management and significance of pigeons within the settlement, thereby contributing to the understudied prehistory of C. livia. Pigeons are native to much of the Mediterranean, including Cyprus, the Levant and northern Africa, and genomic analysis indicates that modern domestic pigeons are most closely related to wild pigeons living in the Middle East, suggesting at least one domestication event in this region (Hernández-Alonso et al. Reference Hernández-Alonso2023). As their name suggests, rock doves naturally make their homes in rocky outcrops and cliffs. The bones have been found in small numbers alongside other species at pre-Neolithic (12000–8800 BCE) sites on Cyprus, as at Akrotiri-Aetokremnos, suggesting opportunistic exploitation but no significant role in human diet at this time (Mourer-Chauviré Reference Mourer-Chauviré and Simmons1999). Starting in the Middle Bronze Age (1950–1650 BCE), pigeon/dove figurines became common on Cyprus, fitting wider trends in Aegean iconography (Binnberg Reference Binnberg2018; Zeman-Wisniewska Reference Zeman-Wisniewska, Bürge and Recht2024) (Figure 1), and an association between the island and pigeons later develops, as Cyprus is identified as the birthplace of the goddess Aphrodite, who had fondness for the birds (Knapp Reference Knapp2013: 372; Dissinger Reference Dissinger2017).
A small limestone pigeon sculpture from Cyprus dating to 600–480 BC (Cypro-Archaic II) (162 × 368 × 133mm) (from The Cesnola Collection of Cypriot Art at the New York Metropolitan Museum of Art (CC0)).

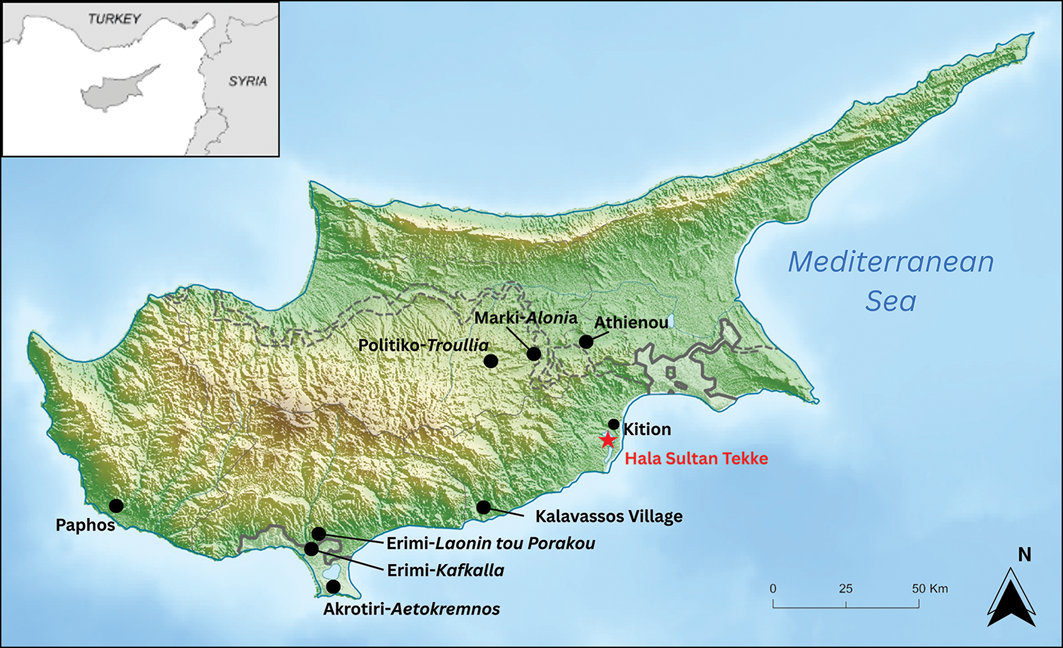
The archaeological site of Hala Sultan Tekke (Dromolaxia-Vyzakia), located on the south-eastern coast of Cyprus at the edge of the Larnaca Salt Lake (Figure 2), yielded a sizable collection of bird bones from its Late Bronze Age deposits (Jonsson Reference Jonsson and Åström1983). This is in line with the increasing quantities of avian remains recovered from the Late Bronze Age contexts in the Levant (Spiciarich Reference Spiciarich2020) and the visible symbolic roles of birds in the ancient Mediterranean (Binnberg Reference Binnberg2018; Zeman-Wisniewska Reference Zeman-Wisniewska, Bürge and Recht2024). Many of the bones are from various duck and waterfowl species, consistent with the site’s location in a wetland on an important migratory route (Devillers et al. Reference Devillers2015). After waterfowl, the most numerous species is C. livia (Jonsson Reference Jonsson and Åström1983; Reese Reference Reese and Åström1998). Only two other sites in the wider region have revealed similar high representation of C. livia remains dating to the Late Bronze Age: Athienou, also on Cyprus, where they are the most abundant bird (Reese Reference Reese2005), and Tell Qatna, Syria, where they are second to waterfowl (Vila & Gourichon Reference Vila, Gourichon and Morandi Bonacossi2007). At the time of excavation, the population at Hala Sultan Tekke was suggested to be Columba livia f. domestica, based on the presence and abundance of fledglings and the perceived importance of pigeons in prehistoric Cypriot society given their connection with the cult of Aphrodite (Jonsson Reference Jonsson and Åström1983).
Map of Cyprus showing the locations of Hala Sultan Tekke and settlements mentioned in the text (figure by authors).

The bones from Hala Sultan Tekke have now been subject to modern analysis, examining not only size through biometry as a proxy of domestication, but also diet, environment and context. The results provide insights into the journey of this often-underestimated domesticated bird, revealing a close relationship with humans.
The site
A harbour city at its peak in the Late Bronze/Late Cypriot Age (1650–1150 BCE), Hala Sultan Tekke (Figure 3) prospered from the large-scale Mediterranean trade and mobility (Fischer Reference Fischer2019a & Reference Fischerb). Although now enclosed, Larnaca Salt Lake was an open and semi-protected bay during this time (Devillers et al. Reference Devillers2015; Fischer Reference Fischer2019a). The city had a rich trading culture and benefitted from imports from all around the Mediterranean (Fischer Reference Fischer2019b). However, by c. 1200–1150 BCE, Hala Sultan Tekke had been destroyed twice (Fischer Reference Fischer2019a) and the bay had silted up, cutting the city off from the sea (Devillers et al. Reference Devillers2015). With its destruction and isolation, the city was quickly abandoned; an event that coincided with the ‘Crisis Years’ that affected many other Mediterranean societies (Cline Reference Cline2014).
Site map of Hala Sultan Tekke, with inset location relative to the modern coastline and current dimensions of Larnaca Bay (figure by authors).

Material and methods
Columbiform bones were identified using the Groningen Institute of Archaeology zooarchaeological reference collection (see Çakırlar et al. Reference Çakırlar2016 for full list) and a reference manual (Cohen & Serjeantson Reference Cohen and Serjeantson1996). A total of 183 Columbiform specimens were examined. Columba livia was distinguished from other possible Columbiformes, such as Columba palumbus (wood pigeon) or Streptopelia decaoto (Eurasian collared dove), with help from the reference collection and a published Columbiform guide (Tomek & Boucheński Reference Tomek and Boucheński2009). Columba oenas (stock dove), which is nearly indistinguishable from C. livia in archaeological material, was ruled out based on geographic range. Only specimens that had intact diagnostic zones (following Tomek & Boucheński Reference Tomek and Boucheński2009) were included in analysis.
Pigeon bones were examined and quantified by NISP (number of identified specimens), MNI (minimum number of individuals) and MNE (minimum number of elements) (Serjeantson Reference Serjeantson2009: 85–97). The MNI was calculated based on age categories, context and sided elements. Specimens were also examined to determine age-at-death, identify pathological or taphonomic changes (weathering, rodent gnawing, etc.), and reveal any signs of human processing (cut/chop marks, burning, etc.) (after Serjeantson Reference Serjeantson2009: 130–64). Birds mature quickly and many different terms are used interchangeably for young that are still nest-bound; here, ‘juvenile’ is used to indicate hatchlings, nestlings and fledglings, identified from any element that is not fully fused (following Studer Reference Studer and Prummel2008). Such juvenile specimens are identified as C. livia based on the urban context and the abundance of adult specimens of this species.
Biometry was also used in differentiating C. livia specimens from those of other possible species. This involved measuring the different portions and profiles of a bone to get its height, breadth and width (Albarella Reference Albarella, Dobney and O’Connor2002). A total of 154 long bones were measured in millimetres (mm) using a dial calliper following standard criteria (Fick Reference Fick1974). All measurements fell within the size range for C. livia (Fick Reference Fick1974; Tomek & Boucheński Reference Tomek and Boucheński2009), thus ruling out C. palumbus and smaller dove species. Greatest length and distal breadth were used to compare complete humeri and femora from Hala Sultan Tekke to published measurements from archaeological assemblages from the wider region and wild, feral and domesticated pigeons from Western Europe in the collections of the Natural History Museum London (NHML) (Marom et al. Reference Marom2018).
Stable isotope analysis of bone collagen was used to infer the diet and lifestyle of 37 individual pigeons. Stable carbon (δ13C) and nitrogen (δ15N) isotope values from teeth or bone may be used to infer diet and estimate trophic level and mobility (Sharpe & Krigbaum Reference Sharpe, Krigbaum, Sharpe and Krigbaum2022). Collagen samples were prepared at the Zooarchaeological Lab at the Groningen Institute of Archaeology (GIA), the Netherlands, and then analysed using the mass spectrometer facility at the Royal Netherlands Institute for Sea Research (see online supplementary material (OSM) 2 for procedures and methods). Isotope datasets from Bronze Age Cyprus are used for comparison (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019; Scirè-Calabrisotto et al. Reference Scirè-Calabrisotto2020; Pilaar Birch et al. Reference Pilaar Birch2022), as no isotopic data has yet been published for Hala Sultan Tekke. To infer pigeon dietary niche, the carbon and nitrogen data were statistically analysed for variation using the SIBER package in R (Jackson & Parnell Reference Jackson and Parnell2023) (data presented in OSM 3).
Results
Zooarchaeological analysis
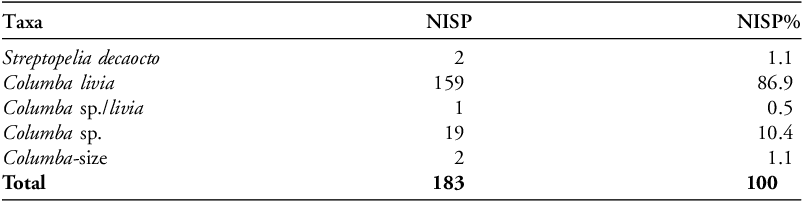
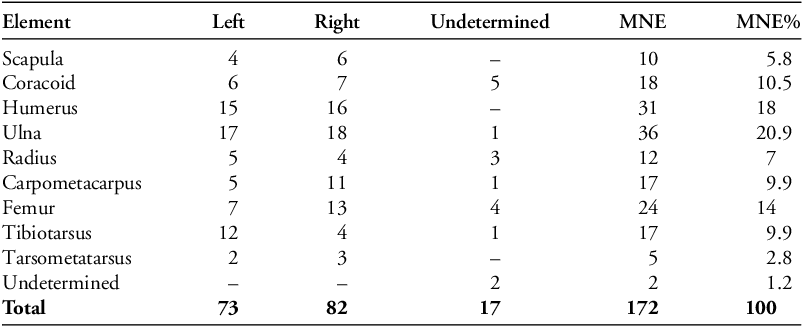
A total NISP of 183 Columbidae specimens were found, of which 159 were determined to be C. livia, 22 were unable to be identified down to species level and two were Streptopelia decaocto (Table 1). The C. livia population of Hala Sultan Tekke is represented by an MNE of 172, and an MNI of 55. Most (72%) identifiable elements belong to the wing portions, the rest (27%) belong to the leg portions (Table 2). No fragile elements from the main body (vertebrae, ribs and sternum) or the cranium could be confidently identified to Columbidae, given the fragmented and eroded nature of the bones.
Identified Columbiform specimens in the HST assemblage calculated by NISP.

Element representation (MNE) of C. livia.

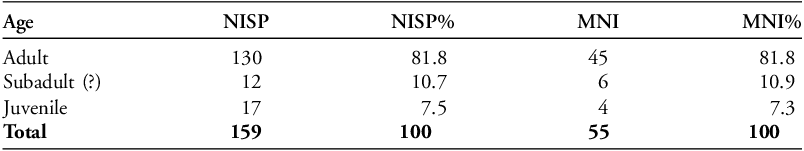
Most specimens (81.8% NISP) belonged to adult pigeons (Table 3). Twelve specimens (10.7% NISP), representing an MNI of six, were particularly porous at the distal and proximal ends, suggesting that these specimens were not fully mature (Serjeantson Reference Serjeantson2009: 36–47), though the porosity could also be due to a taphonomic effect. These specimens are labelled as potential subadults. The presence of juveniles in the assemblage is confirmed by 17 younger juvenile bones (7.5% NISP), representing an MNI of four, found in several different contexts across the site. Just over half of specimens (51% NISP) show some level of contact with fire (Table S1). It is difficult to determine the extent to which juveniles were burned; however, given that the juvenile bones were found among other burnt bones, we can assume that they also encountered fire. Six specimens are slightly discoloured green. These were found in the wells/refuse pits and discolouration could have been caused by the waterlogged environment, the sedimentary context or the presence of bronze objects. Chewing/gnawing marks, most likely made by rodents, are observable on 31 bones (19.5% NISP) (Table S2). There are no visible cut or chop marks, but, for birds of this moderate size, butchery is not necessary for consumption (Serjeantson Reference Serjeantson2009: 144–45). No pathological changes or healed fractures are identified.
Age representation calculated by NISP and MNI of C. livia.

Contextual analysis
Most of the specimens (91% MNI) are from the Late Cypriot II A–C sub-periods (fourteenth–thirteenth centuries) (Table S3). Eighty-two per cent (MNI) of specimens were found in City Quarter 1 (CQ1) and 14.5% (MNI) were found in Area 8 (Table S4). The CQ1 assemblage represents 45 pigeons, 33% (MNI) of which were in contexts L906 and L907 (str. 3), representing burnt animal bone deposits under a plaster floor (Fischer & Bürge Reference Fischer and Bürge2019). These contexts are in rooms 70 and 83, which are connected and appear to be one large space (Fischer & Bürge Reference Fischer and Bürge2020). Material from this location is labelled as feasting deposits (Table S5) and includes burnt bones from mammals and fish as well as other bird species. Other associated finds in these rooms include numerous ceramic vessels of different uses and origins (local, Mycenaean and Canaanite), a furnace, Egyptian faience beads, gold leaf, a Mycenaean figurine and botanical remains (olive, grape and cereals) (Fischer & Bürge Reference Fischer and Bürge2019, Reference Fischer and Bürge2020). At the centre of this larger space, there is a large stone structure that could be a table or workbench, and another stone platform sits to the side. This room is suggested to be a cultic centre used for ritual/religious purposes (Fischer & Bürge Reference Fischer and Bürge2020). C. livia bones were also found in other features in rooms 70/83, situated either above or below the main feasting deposits, potentially pointing to multiple instances of feasting in this location.
Area 8 was excavated two decades earlier, revealing a series of wells (Åström Reference Åström1998). These wells were reused as refuse pits and likely contained deposits from feasting (Bürge Reference Bürge2017). One C. livia individual was found in Tomb 21 in Area 23, which contained the burial of an adult male, and another was found in offering pit Z6 in Area A. In addition, a burnt Streptopelia decaocto humerus was found within a vessel in Tomb RR in Area A.
Biometry
A summary of metrics from comparative populations is presented in Table S6. Archaeological populations include material from Tell Hesban, Jordan (Late Hellenistic-Mamluk, 150 BCE–1517 CE) (Boessneck Reference Boessneck1995), Saadon and Shivta, Israel (Byzantine, 324–638 CE) (Marom et al. Reference Marom2018), and Qasr al-Hayr al-Sharqi, Syria (Early Islamic, 634–960 CE) (Studer Reference Studer and Prummel2008). Material from the latter three sites was found in association with pigeon towers. These assemblages represent C. livia f. domestica groups from later periods than the Hala Sultan Tekke assemblage, as no earlier material is available with published measurements.
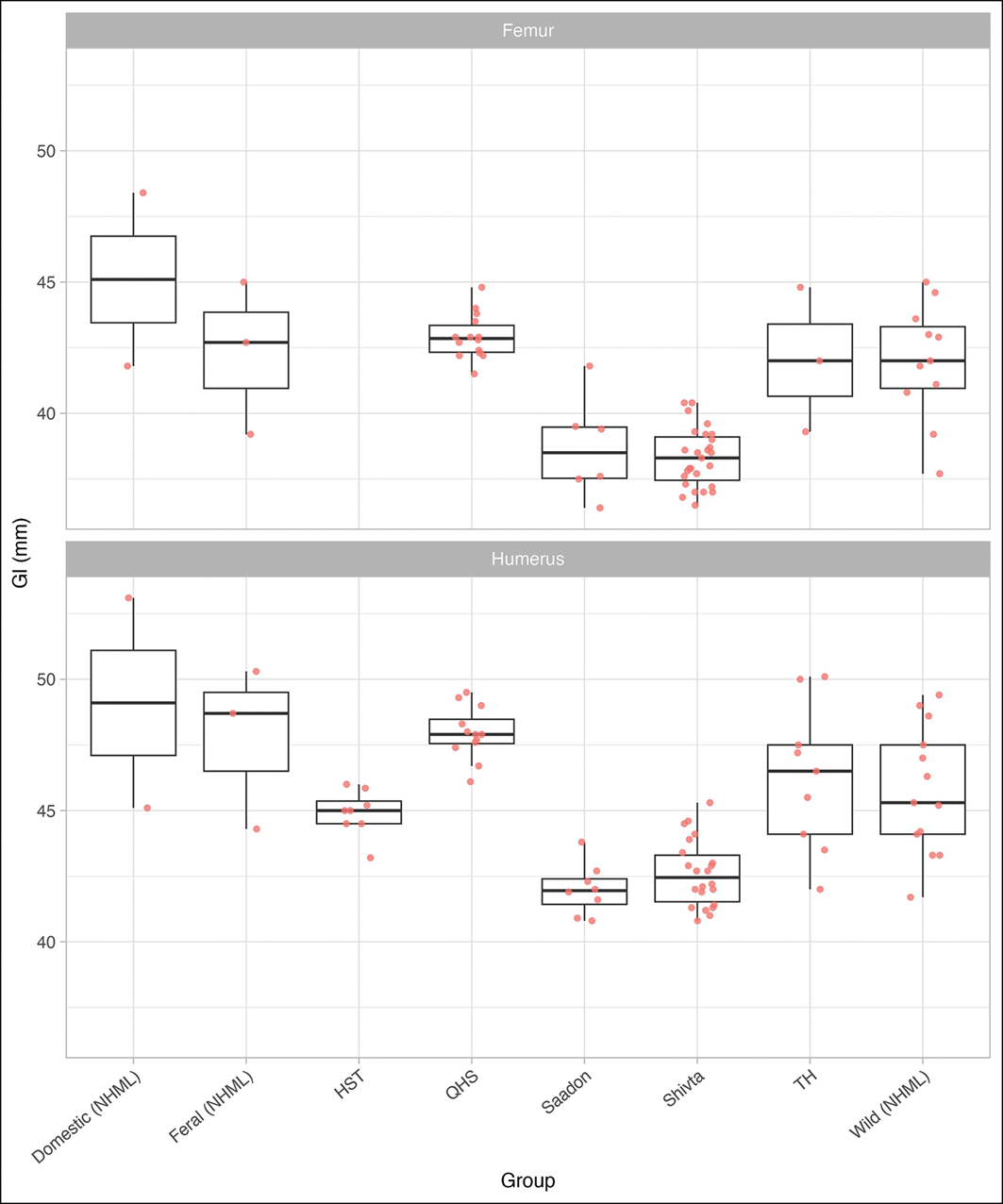
All archaeological assemblages had larger average body sizes than modern-day wild rock doves from the Levant and are therefore considered domesticated (Boessneck Reference Boessneck1995; Studer Reference Studer and Prummel2008; Marom et al. Reference Marom2018). When considering the greatest length of the femur (Figure 4), Hala Sultan Tekke seems to fall below Saadon and Shivta in Byzantine Israel, and even below the wild group. This reflects measurements taken from the only three complete femora in the Hala Sultan Tekke assemblage. When all the incomplete distal and proximal specimens are considered, the three complete femora are shown to sit at the smaller end of the range and are thus not accurate representations of the whole population. The humeri provide a better proxy for body size. When plotted alongside the other assemblages (Figure 4; see also Figures S2 & S3), Hala Sultan Tekke humeri are on average longer than specimens from Saadon and Shivta and shorter than specimens from Qasr al-Hayr al-Sharqi. They fall more in line with measurements from Tell Hesban and European wild rock doves (Figure 4). European specimens from the NHML show greater variation than the archaeological specimens. Ideally, the Hala Sultan Tekke assemblage would have been compared to smaller wild specimens from the Levant region, but such data are currently not available. This analysis exemplifies the great variation in size between different C. livia populations.
Humeral and femoral greatest length (Gl) distributions for C. livia and C. livia f. domestica (with means, extents and outliers). HST: Hala Sultan Tekke; QHS: Qasr al-Hayr al-Sharqi in Syria (Studer 2008); TH: Tell Hesban (Boessneck Reference Boessneck1995). Data for Saadon and Shivta from Marom et al. (Reference Marom2018), data for wild, feral and domestic groups from the NHML (Marom et al. Reference Marom2018) (figure by authors).

Stable isotope analysis
To date, isotopic analysis has been undertaken for several Cypriot Bronze Age zooarchaeological assemblages, including the coastal sites of Erimi-Laonin tou Porakou (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019), Erimi-Kafkalla (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019) and Kalavasos Village (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019), and the more inland sites of Marki-Alonia (Scirè-Calabrisotto et al. Reference Scirè-Calabrisotto2020) and Politiko-Troullia (Pilaar Birch et al. Reference Pilaar Birch2022). These analyses sample Tyto alba (barn owl), Homo sapiens (humans), Canis familiaris (dogs), Vulpes vulpes (foxes), Dama mesopotamica (Persian fallow deer), Sus scrofa (domesticus) (pigs), Ovis aries (sheep), Capra hircus (goats) and Bos taurus (cattle). A summary of the means, standard deviation and sample sizes of these datasets can be found in Table S7. The results from the Hala Sultan Tekke C. livia are presented in Table S8.
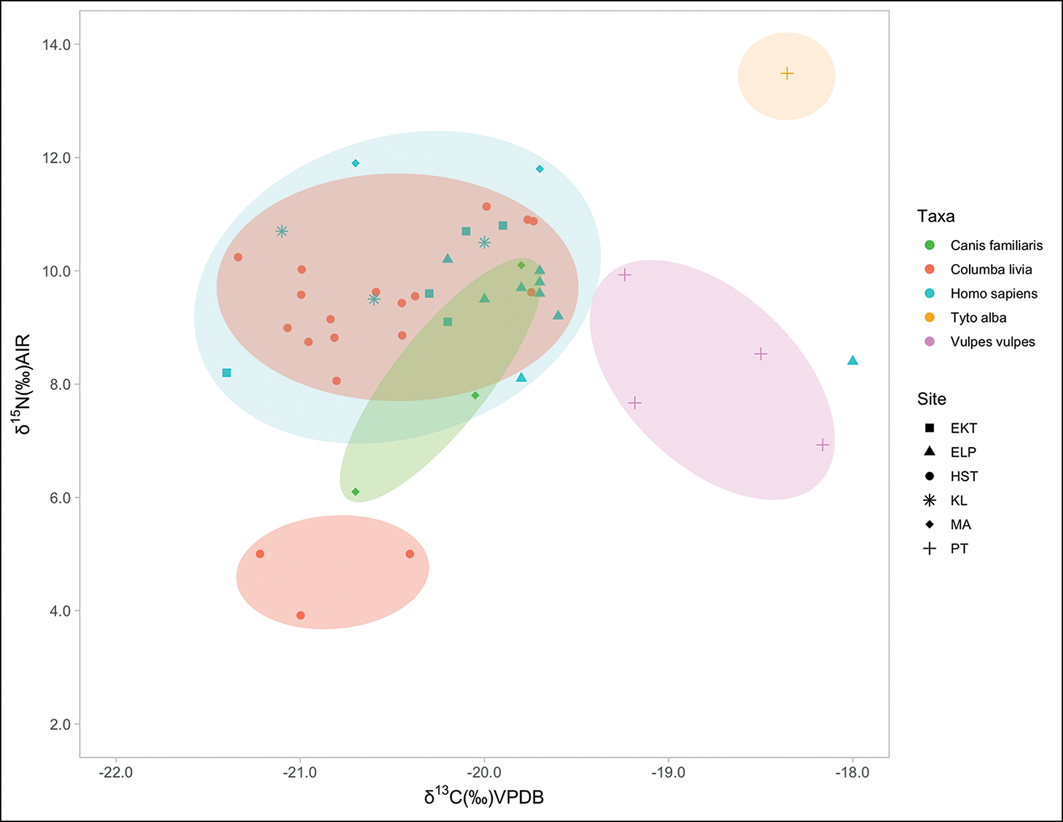
When plotted on a graph, the isotopic values of the C. livia samples from Hala Sultan Tekke form two groupings (Figures 5 & 6, red dots). The δ13C values fall within the range of values from other Cypriot fauna from this period, showing a reliance on C3 plants (C4 plants only arrived on the island later; Scirè-Calabrisotto Reference Scirè-Calabrisotto2019). However, the δ15N results show elevated values for the larger grouping of C. livia (mean δ15N of 8.82‰), indicating a trophic level that is higher than expected, sitting above typical scavenging populations like dogs (8‰) and foxes (8.26‰) and overlapping almost exactly with the human samples (9.92‰). Elevated δ15N values are common for synanthropic species with an urban diet/niche (Guiry & Buckley Reference Guiry and Buckley2018). A smaller grouping of three C. livia individuals has lower δ15N values, overlapping with wild fauna and with herbivores like sheep.
δ13C and δ15N values for Bronze Age Cypriot samples. EKT: Erimi-Kafkalla (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019); ELP: Erimi-Laonin tou Porakou (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019); HST: Hala Sultan Tekke; KL: Kalavasos Village (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019); MA: Marki-Alonia (Scirè-Calabrisotto et al. Reference Scirè-Calabrisotto2020); PT: Politiko-Troullia (Pilaar-Birch et al. Reference Pilaar Birch2022). Ellipses for visual purposes only (figure by authors).

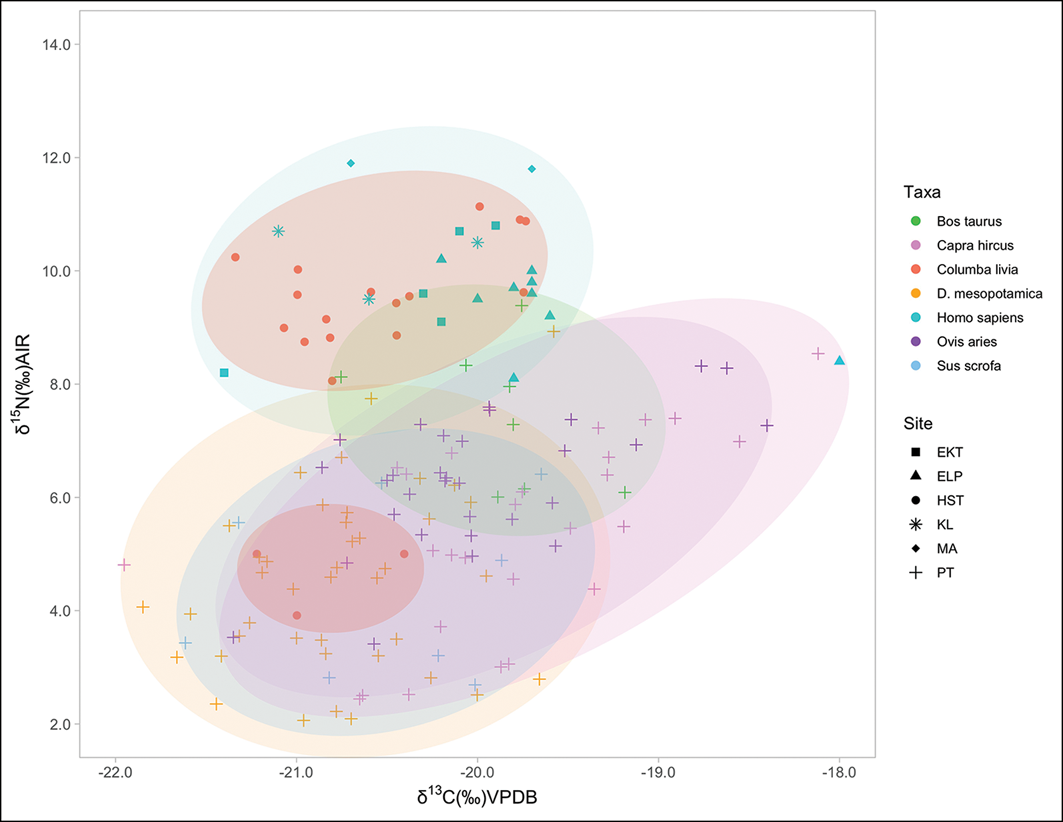
δ13C and δ15N values for Bronze Age Cypriot samples. EKT: Erimi-Kafkalla (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019); ELP: Erimi-Laonin tou Porakou (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019); HST: Hala Sultan Tekke; KL: Kalavasos Village (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019); MA: Marki-Alonia (Scirè-Calabrisotto et al. Reference Scirè-Calabrisotto2020); PT: Politiko-Troullia (Pilaar-Birch et al. Reference Pilaar Birch2022). Ellipses for visualisation only (figure by authors).

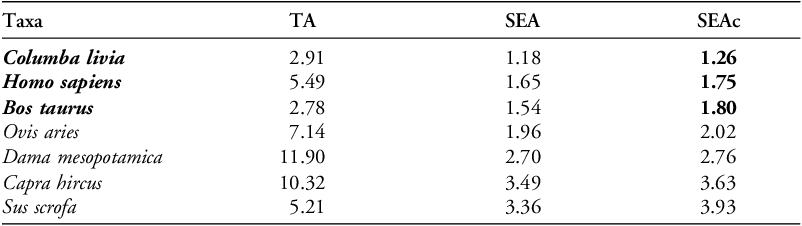
The human values used in this comparison represent multiple sites; although the similarities in δ15N are demonstrative, they cannot be taken to represent the situation in Hala Sultan Tekke directly. Consideration of the spread patterns of isotopic values for each taxa can be informative (see Figure S5). Species fully inhabiting a scavenger niche have highly variable diets that are reflected in greater isotopic ranges, particularly in their δ15N values, while domesticates subject to some level of management have a more homogeneous and consistent diet, showing tighter grouping both in δ15N and δ13C values (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019). This is apparent in the archaeological cattle samples (Figure 6), showing a different herd management style than goats and sheep. Cattle were likely foddered with leafy crops and legumes, while goats were herded in a less restricted manner resulting in a greater variation in δ13C values (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019; Pilaar Birch et al. Reference Pilaar Birch2022; Spyrou et al. Reference Spyrou2024). Fallow deer and feral pigs show a pattern consistent with more wild feeding habits in woodlands and thickets (Scirè-Calabrisotto Reference Scirè-Calabrisotto2019; Pilaar Birch et al. Reference Pilaar Birch2022). Application of SIBER, which fits bi-variate ellipses to isotopic data, indicates that the standard ellipse area (SEAc, acting as a proxy for variability) is lowest for pigeons (1.26), humans (1.75) and cattle (1.80), meaning these taxa have the most consistent diets (Table 4). Pigeons have the smallest SEAc, implying a limited and consistent diet on par with the patterns seen in managed species.
SIBER results representing a proxy for ecological niche size for each taxon.

Notes. TA = Total Area; SEA=Standard Ellipses Area; SEAc = Corrected Standard Ellipses Area (corrected for sample size).
Discussion
The archaeological visibility of pigeons seems to increase after the Neolithic in Cyprus and across the wider Mediterranean region, indicating growing synanthropy. Archaeological specimens that are bigger than modern wild rock pigeons are typically perceived as domestic or feral, as is the case for the Qasr al-Hayr al-Sharqi pigeons from Syria (Boessneck Reference Boessneck1995; Studer Reference Studer and Prummel2008). But this is not always the case. The pigeon bones from Shivta and Saadon in Israel are much smaller than expected for a domestic population, yet all age groups are represented in the assemblage, dovecotes are evidenced at the sites and pigeon guano seems to have been used as fertiliser, all of which would suggest that the bones belong to C. livia f. domestica (Marom et al. Reference Marom2018; Tepper et al. Reference Tepper2018). This disparity could be due to the desert location and the action of Bergmann’s rule, which broadly states that animals in hot, arid climates tend towards smaller body mass (Marom et al. Reference Marom2018; Yeomans Reference Yeomans2025). Alternatively, the smaller size of the specimens from Shivta and Saadon could indicate that their primary purpose was for fertiliser production, not consumption, and that they were not therefore fattened up with a rich diet (Marom et al. Reference Marom2018; Tepper et al. Reference Tepper2018). Thus, resources were prioritised in the desert, size was not a primary concern for the breeders, and the birds stayed small. This indicates a strong relationship between the roles of pigeons and the diets and management practices they were subjected to. Domesticity in pigeon morphology, therefore, is context and region specific, and reflects the plasticity of the species.
Overlap in the range of stable isotope values from the C. livia samples and the Homo sapiens samples from other Cypriot sites indicates that the Hala Sultan Tekke pigeons likely had an omnivorous diet that consisted of human-supplemented food including grains, seeds and possibly some protein or fat from animal products. The narrow dietary niche occupied by these pigeons is consistent with human control or confinement; a similar tight grouping of isotopic values is seen in ancient Cypriot cattle, which were prized livestock managed systematically. Although the comparative faunal data do not derive from Hala Sultan Tekke, they nevertheless indicate that pigeons were kept in close contact with humans and likely lived within the city. The presence of fledgling bones confirms that pigeons were also breeding within the settlement (Jonsson Reference Jonsson and Åström1983; Studer Reference Studer and Prummel2008). While no pigeon towers have been found, the consistency of isotopic values and their correspondence with the human dietary range point toward a commensal relationship. Pigeon towers are generally under-represented in the archaeological record due to the poor preservation of multi-level buildings (Yeomans Reference Yeomans2025). The use of Ashlar masonry is, however, recorded at Hala Sultan Tekke (Negbi Reference Negbi2005) and it is possible that multi-level buildings could have been constructed using this cut- and dressed-stone masonry technique in Late Bronze Age Cyprus. The presence of tall, sheer-sided buildings could have aided the human-pigeon commensal relationship by simulating the pigeons’ natural nesting sites.
The three specimens with lower δ15N values may represent pigeons from more rural contexts, who had a different diet. Rats from urban environments have also been shown to have higher δ15N values compared with rats from more rural settings (Guiry & Buckley Reference Guiry and Buckley2018). One plausible source for these outliers is the nearby site of Trypes, a rural settlement that was under the control of Hala Sultan Tekke and which might have functioned as a granary and supplier of managed livestock and game (Fischer Reference Fischer2019a). Alternatively, these pigeons could have come from outside of the anthropogenic niche.
Context is key when looking at the potential significance and role of pigeons at archaeological sites. The presence of burning and the selective representation of skeletal elements suggests that the birds at Hala Sultan Tekke were eaten (Serjeantson Reference Serjeantson2009: 163–64; Dissinger Reference Dissinger2017). Many pigeons (MNI = 55), mostly found in CQ1, were buried together with other burnt animal remains in an open room that contained a stone table and altar. These remains were marked as feasting deposits during excavation and were accompanied by abundant tableware, characteristic of feasting (Steel Reference Steel2004). In this context, the bones were likely burned either as a means of discarding waste after consumption or as part of an offering or ritual. Either way, the location and method of deposition—within covered pits in a ritual space—is intentional. Ritual feasting was an important part of Cypriot culture in the Bronze Age and examples can be found all around the island as well as in other cultures in the Mediterranean (Steel Reference Steel2004; Knapp Reference Knapp2013: 15). Feasts were socially, politically and religiously significant events that also formed part of mortuary rites, which could explain why some pigeons were found among the remnants of feasts deposited in wells close to tombs (Steel Reference Steel2004; Knapp Reference Knapp2013: 15; Bürge Reference Bürge2017).
Hala Sultan Tekke, with its trade links to the Levant and Syria-Palestine (Fischer Reference Fischer2019b), likely participated in a broader symbolic tradition where pigeons were associated with fertility goddesses, precursors to the classical cult of Aphrodite. Many coastal temples and sanctuaries were dedicated to her worship (Dissinger Reference Dissinger2017), but the centre of worship was in Paphos on Cyprus, where a large temple stood in her honour. This temple was built at the start of the Late Bronze Age, but another structure may have stood there before it, suggesting a continuity of worship that may have started in the Chalcolithic (3900–2300 BCE) (Maier & Karageorghis Reference Maier and Karageorghis1984: 81–102). While such connections cannot be attested with certainty at Hala Sultan Tekke, as no temples or sanctuaries are found there, they offer a wider interpretive framework for understanding the presence of pigeons in ritual contexts. Their presence in these contexts and their dietary entanglement with humans strongly suggests that C. livia were not just opportunistic urban inhabitants in Late Bronze Age Cyprus.
Conclusion
Examination of avian bones and their contexts of discovery at the Late Bronze Age site of Hala Sultan Tekke on Cyprus indicates that pigeons likely lived in sustained close contact with humans, where they were managed and probably bred on site and consumed during ritual feasting. Although no pigeon towers have been found at the site, biometry and stable isotope values suggest that C. livia on Cyprus were at least semi-domesticated by 1400 BCE and support a commensal pathway to domestication for pigeons. The study presents some of the earliest direct biomolecular evidence for niche-sharing among humans and pigeons. Case studies such as this exemplify the intricacies of the domestication process, highlighting the need for more context- and species-specific investigations. Further integrative studies, such as this one, will help fill the gaps in the story of pigeon domestication, contributing to our understanding of the prehistory of regions like Cyprus, where this bird was a significant and constant presence in everyday life.
Acknowledgements
We thank all our colleagues at the Groningen Institute of Archaeology who supported this study, especially Marcel van Tuinen, Willemien de Kock and Thom Brongers.
Online supplementary material (OSM)
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2026.10351 and select the supplementary materials tab.
Author contributions: CRediT categories
Anderson L. Carter: Conceptualization-Equal, Formal analysis-Lead, Investigation-Lead, Visualization-Lead, Writing - original draft-Lead, Writing - review & editing-Equal. David S. Reese: Investigation-Supporting, Resources-Supporting. Ronald van Bommel: Formal analysis-Supporting. Marcel T.J. van der Meer: Formal analysis-Supporting, Investigation-Supporting, Methodology-Equal, Validation-Equal, Writing - review & editing-Equal. Canan Çakırlar: Conceptualization-Equal, Data curation-Equal, Formal analysis-Supporting, Funding acquisition-Lead, Investigation-Equal, Methodology-Equal, Project administration-Lead, Resources-Equal, Supervision-Lead, Visualization-Supporting, Writing - original draft-Supporting, Writing - review & editing-Lead.





 Open access
Open access