


Non-technical Summary
The sedimentary rocks of the Grand Canyon, Arizona, have been used as a classic model of what happens when rising sea level causes continental flooding. Edwin McKee proposed this idea in the 1940s, documenting it using three iconic Cambrian (500- to 525-million-years old) rock units: the Tapeats Sandstone, Bright Angel Formation, and Muav Formation. A key part of McKee’s hypothesis is that the fossils and rock units that were deposited by this continental flooding are younger as one moves east, reflecting the progressive eastward movement of the shoreline onto the continent. For this study, we reexamined the Cambrian trilobites from the Grand Canyon described by previous workers and collected fossils from new Grand Canyon localities and adjacent portions of Nevada. This study reports two new species of fossils from these rocks as well as new occurrences of other trilobite species that were not previously known from the Grand Canyon (e.g., from California, Nevada, Utah, and Montana). This study shows that the rock formations do young eastward, and that this trend extends upwards to include younger intervals of time that were not previously recognized. However, the trend is not simple: the Cambrian sedimentary sequence is punctuated by previously unknown periods of erosion.
Introduction
The Cambrian of the Grand Canyon (Fig. 1) is often interpreted as a classic transgressive sequence (see review in Dehler et al., Reference Dehler, Sundberg, Karlstrom, Crossey, Schmitz, Rowland and Hagadorn2024). This model was formulated by McKee and Resser (Reference McKee and Resser1945), whereby the Tapeats Sandstone represents the initial shallow-water deposit that blanketed eroded Precambrian strata (thus forming the Great Unconformity), with the deeper water Bright Angel Formation (Shale) subsequently covering the Tapeats Sandstone, and the deepest water Muav Formation interfingering with the Bright Angel Formation and transgressing eastward across the shales. The genesis of this sequence and associated depositional environments has been questioned, with the most significant changes to the model often being that portions of the lower Tapeats and the Bright Angel formations represent braided delta to fluvial deposits (Rose, Reference Rose2003, Reference Rose2006, Reference Rose2011) and the Muav Formation represents intertidal to supratidal carbonate bank deposits (Wanless, Reference Wanless1973, Reference Wanless and Ginsburg1975). In addition, the Sixtymile Formation, previously thought to be Precambrian, is now known to represent basin fill of early Cambrian age (Karlstrom et al., Reference Karlstrom, Mohr, Schmitz, Sundberg, Rowland, Hagadorn, Foster, Crossey, Dehler and Blakey2020) and the ‘undifferentiated dolostones’ overlying the Muav Formation represent middle to late Cambrian subtidal to intertidal platform carbonate environments (Rowland et al., Reference Rowland, Korolev, Hagadorn and Ghosh2023). All of these stratigraphic units are considered part of the Tonto Group (Karlstrom et al., Reference Karlstrom, Mohr, Schmitz, Sundberg, Rowland, Hagadorn, Foster, Crossey, Dehler and Blakey2020).
Location map for the Western, Central, and Eastern transects of the Grand Canyon, Arizona, shown as white-outlined boxes. Measured sections illustrated in Figures 2–9 are the purple dots that are labeled in the white-background boxes. Additional isolated localities collected during this study are labeled in blue dots. McKee and Resser (Reference McKee1945) localities not used for measured sections are labeled in green, see Table 1 for abbreviations.

Figure 1. Long description
A satellite-style relief map of the Grand Canyon region spanning from Nevada in the West to Arizona in the East. The map is divided into three main regions labeled Western Transect, Central Transect, and Eastern Transect.
* Western Transect: Contains white boxes for Figure 4 and Figure 5. Purple dots mark Frenchman Mountain, Devil's Cove, Rampart Cave, Diamond Bar Ranch, Quartermaster Canyon, Meriwitica, and Bridge Canyon. Blue dots include G R C A 331, G R C A 384, and G R C A 382. Green dots are labeled C F, 90, G P, 212, D C, and P S.
* Central Transect: Contains white boxes for Figure 6 and Figure 7. Purple dots mark 190 Mile, Toroweap, Cove Canyon, Honga Springs, Pumpkin Springs, Peach Springs, Olo Canyon, Kanab Canyon, Fishtail, and Fossil Rapids. Blue dots include G R C A 377. Green dots are labeled G W and H M. The Havasupai Reservation is outlined in yellow dashed lines to the South.
* Eastern Transect: Contains white boxes for Figure 8 and Figure 9. Purple dots mark Hotauta, Blacktail Canyon, Dragon Head, Pipe Creek, Howlands Butte, Mile 41.5, Mile 50, Nankoweap Canyon, Malgosa Canyon, Palisades, and Tanner Trail. Blue dots include G R C A 252 and G R C A 272. Green dots are labeled T C, B A B, B E, H E, R S, X V, C V, L C, G r, C o, L T, and H G B A.
A North arrow points upward at the bottom center, and a scale bar indicates 0 to 50 kilometers in the bottom right corner.
Abbreviations of McKee and Resser’s (Reference McKee1945) localities and sections listed in Figures 1–5 and their corresponding stratigraphic section

Table 1. Long description
The table consists of three columns: Abbreviation, Locality/Section Name, and Data plotted on Section.
* 212: Mile 212, Pumpkin Springs.
* B A: Bright Angel Trail, Pipe Creek.
* B A S: Bass, no data listed.
* B C: Bridge Canyon, Bridge Canyon.
* C F: Columbine Falls, Rampart Cave.
* C V: Chuar Valley, Palisades.
* D C: Diamond Creek, Peach Springs.
* D R: Diamond Bar Ranch, Diamond Bar Ranch.
* G P: Granite Park, Pumpkin Springs.
* G W: Gateway Canyon, Gateway Canyon.
* H C: Havasu Canyon, Havasu Canyon.
* H E: Hermit, no data listed.
* H G: Havasu Gardens, Pipe Creek.
* H M: Hermit Creek, Pipe Creek.
* K C: Kanab Canyon, Kanab Canyon.
* K V: Kwagnunt Valley, Malgosa Canyon.
* L C: Lava Canyon, Palisades.
* L T: Lone Tree Canyon, Howlands Butte.
* M W: Meriwitica, Merwitica.
* N W: Nankoweap Valley, Nankoweap Valley.
* P S: Peach Springs Canyon, Peach Springs.
* Q M: Quartermaster Canyon, Quartermaster Canyon.
* R S: Roaring Springs, Palisades.
* S E: Serpentine, no data listed.
* T C: Tapeats Creek, Fossil Rapids.
* T W: Toroweap Valley, Toroweap Valley.
The time-transgressive model presented by McKee and Resser (Reference McKee and Resser1945) is based primarily on the occurrence of diagnostic fossils for the “Olenellus,” “Glossopleura,” and “Ehmaniella” biozones with correlations of minor lithologic units (e.g., intraformational conglomerate horizons). Subsequent discussions of the classic model have focused on depositional environment, but still relied on the original paleontological data. The intent of this paper is to focus on the trilobite paleontology of the Cambrian of the Grand Canyon and adjacent regions in Arizona and Nevada to determine the correlation of stratigraphic units and test and refine the McKee and Resser (Reference McKee and Resser1945) model. The data used for this reanalysis are from historical collections at the Smithsonian Institution (National Museum of Natural History – USNM) made by Walcott, Resser, McKee, and others; historical material collected by McKee in the Grand Canyon National Park collections (GRCA); and McKee’s field/lab notes (from the United States Geologic Survey – USGS, provided by M. Nebel, GRCA) that underpinned McKee and Resser’s Reference McKee and Resser1945 publication focused on at the GRCA and USNM collections.
Stratigraphic nomenclature
The Tonto Group consists of five formations: (1) the recently reassigned Sixtymile Formation (Karlstrom et al., Reference Karlstrom, Hagadorn, Gehrels, Mathews, Schmitz, Madronich, Mulder, Pecha, Geisler and Crossey2018, Reference Karlstrom, Mohr, Schmitz, Sundberg, Rowland, Hagadorn, Foster, Crossey, Dehler and Blakey2020); (2) the Tapeats Sandstone, which predominantly represents fluvial to coastal sands that mantled the majority of the Great Unconformity in the region; (3) the Bright Angel Formation and its associated marine shales and fossils; (4) the carbonate-dominated Muav Formation and associated fossils; and (5) the Frenchman Mountain Dolostone (Rowland et al., Reference Rowland, Korolev, Hagadorn and Ghosh2023), which is dominated by rhythmic and microbially influenced bedding (originally referred to as ‘Undifferentiated dolomites’ by McKee, Reference McKee1945). This paper focuses on the uppermost Tapeats Sandstone to basal Frenchman Mountain Dolostone. Figure 2 presents litho- and biostratigraphic correlations between two representative sections: our Diamond Bar Ranch section located in the western portion of our study area, and the Blacktail Canyon section located in the eastern portion of our study area (Fig. 1). Rose (Reference Rose2011) proposed Blacktail Canyon as the type section for the Tonto Group.
Generalized litho- and biostratigraphic correlation of Cambrian sections representative of the western portion (Diamond Bar Ranch) and eastern portion (Blacktail Canyon) of the Grand Canyon. See lithological legend in Figure 6. Intern. = International.

Figure 2. Long description
The chart is organized into vertical columns.
1. Chronostratigraphy and Biostratigraphy (Left):
- The leftmost columns define the time scales: Intern. (Miaolingian, Series 2) and Laurentian (Drumian Stage, Marjuman Stage, Wuliuan, Lincolnian, Delamaran, Stage 4, Waucoban, Dyeran).
- The Biozone column lists trilobite zones including Bolaspidella, Ehmaniella, Elrathiella, Proehmaniella, Glossopleura walcotti, Mexicella mexicana, Poliella denticulata, Amecephalus arrojoensis, Eokochaspis nodosa, and Nephrolenellus multinodus.
2. Lithostratigraphy (Center):
- The Muav Formation includes members: Havasu, Gateway Canyon, Peach Springs/Kanab, Spencer Canyon, Sanup Plateau, and Rampart Cave.
- The Bright Angel Formation includes Flour Sack, Merwitica, Tincanabits, Diamond Bar, and upper/lower slope units.
- The base is the Tapeats Sandstone (Lower Unit and Transition Zone).
3. Stratigraphic Sections (Right):
- Diamond Bar Ranch (~280 mile) and Blacktail Canyon (120 mi.) are shown as vertical columns with lithological patterns (bricks for limestone, dots for sandstone, dashes for shale).
- Correlation lines connect the two sections.
- Large diagonal color bands (yellow, green, blue) represent time-transgressive facies shifts.
- Three large white question marks are placed within the green and blue diagonal bands between the two sections, indicating uncertainty in the correlation of the lower Bright Angel Formation and Tapeats Sandstone transition.
- Vertical scales in meters are marked on the sections, reaching up to 300 meters.
Construction of the thickness and correlation diagram (Fig. 3) is based on 28 stratigraphic sections (Figs. 4–9), arranged by longitude from the palinspatically restored position of the Frenchman Mountain section (Fryxell and Duebendorfer, Reference Fryxell and Duebendorfer2005) to the west to the Palisades section to the east. From the Palisades section, the easternmost sections are displayed roughly south to north and spaced by distance given their same longitudinal position (Fig. 9). Fossils from adjacent sections were correlated to those illustrated in Figures 4–9. Furthermore, additional sections illustrated by Rose (Reference Rose2003) were utilized. Although stratigraphic sections that contained much more detail were used in our analyses, we simplified them in our fence diagrams in order to reduce the heterolithic complexity of the formations (e.g., Bright Angel Formation) and produce a relatively simplified fence panel (Fig. 3.1) that facilitates illustration of the overall relationships of the formations.
Fence and Wheeler diagrams of the Cambrian Tapeats Sandstone to Frenchman Mountain Dolostone illustrating paleontological control (modified from Dehler et al., Reference Dehler, Sundberg, Karlstrom, Crossey, Schmitz, Rowland and Hagadorn2024). Paleontological control point (circles) colors match the biozone colors. (1) Stratigraphic thickness panel showing total thickness of the Frenchman Mountain Dolostone. (2) Wheeler diagram of the same interval excluding most of the Frenchman Mountain Dolostone. The larger numbered circles represent sedimentological sequences (see Dehler et al., Reference Dehler, Sundberg, Karlstrom, Crossey, Schmitz, Rowland and Hagadorn2024). Biozone/subzone (in ascending order): Peach.–Neph. = Peachella iddingsi to Nephrolenellus multinodus; Eok. = Eokochaspis nodosa; Ame. = Amecephalus arrojosensis; Pol. = Poliella denticulata; Mex. = Mexicella mexicana; Gloss. = Glossopleura walcotti; Pro. = Proehmaniella; Elra. = Elrathiella; Ehm. = Ehmaniella; Bolas. = Bolaspidella; Ced. = Cedaria; Crep. = Crepicephalus. Member abbreviations (in ascending order): Transition = Transition zone; lower = lower slope unit (used by McKee, Reference McKee1945); Diamond = Diamond Bar member; upper = upper slope unit (used by McKee, Reference McKee1945); Lyndon = Lyndon Limestone; Rampart = Rampart Cave; Sanup = Sanup Plateau; Spencer = Spencer Canyon; Kanab = Kanab Canyon; Gateway = Gateway Canyon. Marj = Marjuman; Ss = Sandstone; Step = Steptoean.

Figure 3. Long description
Panel 1 is a stratigraphic thickness diagram. The vertical axis on the left shows biozones from bottom to top: Peach. dash Neph., Eok., Ame., Pol., Mex., Gloss., Pro., Elra., Ehm., Bolas., Ced., and Crep. Adjacent to this is the formation column: Tapeats S s, Bright Angel Fm. (including Transition, lower, Diamond, and upper members), Muav Formation (including Rampart, Sanup, Spencer, Peach Springs, Kanab, Gateway, and Havasu members), and Frenchman Mountain Dolostone. The main body of the diagram shows these units thinning and thickening across geographical locations labeled at the top, including Frenchman Mountain, Peach Springs, and Kanab Canyon. A legend identifies lithologies: grey for Shale forward slash Siltstone, tan for Bioturbated Sandstone, brown for Non-bioturbated Sandstone, red for Hematitic Sandstones, pink for Diamond Bar Sandstone, light blue for Carbonate, orange for Rust-brown Dolostone, and purple for Frenchman Mtn. Dolostone. Small colored circles indicate fossil control points corresponding to biozone colors.
Panel 2 is a Wheeler diagram representing the same interval in time-space. It is anchored by West (W) on the left and East (E) to North (N) on the right. Large numbered circles 1 through 5 mark sedimentological sequences. Sequence 1 at the base includes the Tapeats S s and lower Bright Angel Fm. Sequence 2 contains the Diamond and upper members. Sequence 3 includes the Lyndon Limestone and Flour Sack members. Sequence 4 encompasses the Muav Formation members. Sequence 5 is the Frenchman Mountain Dolostone. Grey vertical lines on the right represent Monadnocks and the Butte Fault. A horizontal scale bar at the bottom right indicates distance from 0 to 50 K m.
Cambrian westernmost sections of the Western Transect (see Fig. 1). Stratigraphic sections are hung on the base of the Frenchman Mountain Dolostone, which has only lower 15 m illustrated. Distances labeled are areal distances between sections. All thicknesses are in meters. See Figure 6 for lithologic symbols.

Figure 4. Long description
A stratigraphic cross-section featuring three vertical columns representing Frenchman Mountain (restored), Devil's Cove, and Diamond Bar Ranch. The columns are separated by horizontal distances of 12.7 k m, 18.7 k m, and 23 k m respectively.
Vertical Stratigraphy:
* The base is the Tapeats Sandstone, followed by the Bright Angel Formation (including Diamond Bar Ranch and Flour Sack members), and the Muav Formation (including Lyndon Limestone, Rampart Cave, Sanup Plateau, Spencer Canyon, Peach Springs, Kanab, Gateway, and Havasu members).
* A vertical scale on the left and center indicates thickness in meters, reaching up to 400 m.
Lithology and Biozones:
* Strata are correlated with solid and dashed lines across the sections.
* Major biozones labeled in the center and right include Ehmaniella, Elrathiella, Proehmaniella, Glossopleura walcotti, Mexicella mexicana, and Eokochaspis nodosa or Poliella denticulata.
* Three unconformities are marked with wavy colored lines: Disconformity 1 (blue/purple) below Nephrolenellus multinodus; Disconformity 2 (green/red) below Glossopleura walcotti; and Disconformity 3 (orange/red) above the Elrathiella subzone.
Trilobite Data:
* Vertical lines with colored circles indicate the stratigraphic range of specific trilobite species.
* A legend on the far right lists 13 biozones/subzones with corresponding colored circles, ranging from Bolaspidella at the top to Peachella iddingsi at the bottom.
* Specific sample codes like F H P R 008-7 and U C R 10029 are pinned to exact stratigraphic heights.
* Text labels for species names like Anoria tontoensis and Glossopleura bocar are oriented vertically next to their respective range lines.
Cambrian easternmost sections of the Western Transect (see Fig. 1). Stratigraphic sections are hung on the base of the Frenchman Mountain Dolostone, which has only lower 15 m illustrated. Distances labeled are areal distances between sections. All thicknesses are in meters. See Figures 4 and 6 for symbols.

Figure 5. Long description
A series of five vertical stratigraphic columns are arranged from West to East: Rampart Cave (275 miles), Quartermaster Canyon (260 miles), Bridge Canyon (235 miles), Peach Springs (226 miles), and an unnamed section 20 kilometers further East. The columns are hung on a horizontal baseline at the base of the Frenchman Mountain Dolostone.
* The vertical sequence from base to top consists of the Tapeats Sandstone (Lower portion and Transition Zone), the Bright Angel Formation (Diamond Bar, Flour Sack, and Rampart Cave members), and the Muav Formation (Sanup Plateau, Spencer Canyon, Peach Springs-Kanab, Gateway, and Havasu members).
* Correlation lines connect equivalent geological boundaries across the sections. A prominent dashed pink line marks the top of the Havasu member. Dashed yellow and orange lines correlate markers within the Muav and Bright Angel formations.
* Fossil biozones are indicated by colored dots and labels. Blue dots at the base represent the Nephrolenellus multinodus zone. Green dots in the middle represent the Mexicella mexicana zone. Yellow and orange dots higher up mark various trilobite species including Kootenia schenki and Spencella porcata.
* Specific sample identifiers such as U S N M 44345 and G R C A 393 are positioned next to their respective stratigraphic heights. Lithology is represented by patterns: stippled for sandstone, horizontal lines for shale, and block patterns for limestone and dolostone. Red wavy lines indicate unconformities or significant erosional surfaces between the Bright Angel and Muav formations.

Figure 6. Long description
A complex stratigraphic diagram showing six vertical columns representing geological sections at Pumpkin Springs, Toroweap, Honga Spring, Cove Canyon, and Gateway. The columns are spaced by distances ranging from 2 kilometers to 190 miles.
Vertical Succession:
* The base of all sections is the Tapeats Sandstone (Lower portion), characterized by cross-bedded siliciclastics.
* Above this is the Bright Angel Formation, which contains several fossil zones including Nephrolenellus multinodus (blue), Mexicella mexicana (green), Glossopleura walcotti (dark green), Proehmaniella (yellow), Elrathiella (light green), and Ehmaniella (orange). The Diamond Bar member is noted in the lower half.
* The upper section consists of the Muav Formation, subdivided into Peach Springs, Gateway, and Havasu members.
* The top boundary is marked by a dashed magenta line representing the Frenchman Mountain Dolostone.
Lithological Correlation:
Horizontal lines and shaded regions connect similar rock units across the transect. A prominent wavy red line indicates an unconformity or significant boundary within the Bright Angel Formation. Fossil localities are marked with colored dots and labels such as G R C A 381-1 and M & R 58.
Legend Categories:
* Glaucognitic: Shale (light green), Shale forward slash Sand (green with dots), and Sandstone (dotted green).
* Limestone and Dolostone: Symbols for bioturbated, silty, intraclastic, oncolitic, and red brown (R B D) varieties.
* Siliciclastic: Symbols for shale, siltstone, fine to coarse sandstone, and cross-bedded units.
* Specialized Units: Black Shale (flour sack pattern), Red Shale forward slash Sand, and Bioturbated and Cross-bedded units.
Cambrian westernmost sections of the Eastern Transect (see Fig. 1). See Figures 4 and 6 for symbols.

Figure 7. Long description
A geological cross-section diagram illustrating the stratigraphic correlation of Cambrian rock units. The Y axis represents vertical thickness in meters, while the X axis spans a West to East transect.
From West to East, the five columns are anchored at:
1. Olo Canyon at 146 miles.
2. Kanab Canyon at 144 miles, located 3 kilometers East of Olo.
3. Fishtail at 138.5 miles.
4. Fossil Rapids at 125.5 miles, located 17.5 kilometers East of Fishtail.
5. Blacktail Canyon at 120 miles, located 5.5 kilometers East of Fossil Rapids and 14.5 kilometers from the next section.
Vertical Stratigraphy:
* The base layer is the Tapeats Sandstone, divided into a Lower portion and a Transition Zone.
* Above this is the Bright Angel Formation, which contains the Glossopleura walcotti biozone at the bottom, followed by the Elrathiella biozone.
* The top layer is the Muav Formation, containing the Ehmaniella biozone. The Muav is further subdivided into members: Peach Springs, Gateway, and Havasu.
Key Features:
* A wavy red line indicates an unconformity or significant boundary between the Glossopleura and Elrathiella zones.
* A dashed orange line marks the boundary between Elrathiella and Ehmaniella.
* A dashed pink line runs horizontally across the top of the Havasu member.
* Fossil localities are marked with G R C A codes, such as G R C A 378-1 through 3 in the Peach Springs member and G R C A 341-1 through 4 in the Blacktail Canyon section.
* Specific fossil names are listed vertically next to their respective collection points, including Ehmaniella nitida, Kochina angustata, and Glossopleura boccar.
Cambrian easternmost sections of the Eastern Transect (see Fig. 1). See Figures 4 and 6 for symbols.

Figure 8. Long description
A geological cross-section diagram with six vertical columns representing locations from West to East: Hotauta, Dragon Head, Pipe Creek, Howlands Butte, and Tanner Trial. Distances between columns are marked in kilometers and miles.
Vertical Stratigraphy (Bottom to Top):
* Tapeats Sandstone: The base layer featuring cross-bedded patterns and a Transition Zone.
* Bright Angel Formation: A thick middle section containing green and grey shaded units. It is divided into biozones labeled Glossopleura walcotti (green text) and Elrathiella (yellow text). A wavy orange line indicates an unconformity or significant boundary within this formation.
* Muav Limestone: The uppermost major unit, subdivided into Peach Springs, Gateway, and Havasu members. An Ehmaniella biozone is marked in the upper Gateway member.
Key Features and Labels:
* Correlation Lines: Solid grey lines connect similar lithological units across columns. A dashed pink line marks a high-level datum across the Havasu member. A dashed brown line connects the top of the Peach Springs member.
* Fossil Localities: Numerous U S N M and G R C A labels with associated dots indicate fossil collection points. Specific taxa listed include Ptarmigania sp. indet., Elrathiella fonticola, Amecephalus althea, Anoria tontoensis, Glossopleura boccara, Kootenia sp. indet., and others.
* Vertical Scale: Each column includes a vertical meter scale from 0 to over 200.
* Horizontal Scale: Grain size indicators at the base of columns are labeled M, S, F S, C, and C S.
Cambrian far easternmost sections of the Eastern Transect (see Fig. 1) in a south–north profile. See Figures 4 and 6 for symbols.

Figure 9. Long description
A multi-panel stratigraphic profile displays six vertical columns representing different locations. From left to right, these are Palisades 66 miles, Malgosa Canyon 58 miles, Nankoweap Canyon 52 miles, 50 Mile, 3.5 kilometer, and 41.5 Mile. Distances between columns are marked at the top as 11.5 kilometers, 7 kilometers, 6.5 kilometers, 50 Mile, 3.5 kilometers, and 41.5 Mile.
Each column features detailed lithological symbols including limestone blocks, sandstone dots, and shale lines. Vertical scales on the columns range from 0 to over 200. Horizontal colored lines correlate biozones across the columns.
* A pink dashed line at the top marks the Bolaspidella zone.
* An orange dashed line marks the Ehmaniella zone.
* A solid red wavy line marks the Elrathiella zone.
* A green label at the bottom left identifies the Glossopleura walcotti zone.
Numerous labels with dots point to specific fossil occurrences. Labels include G R C A 362-2, G R C A 361-2, G R C A 357-7, and U S N M 44336. Species names listed vertically next to columns include Kootenia mckeii, Ehmaniella kwaguntensis, and Glossopleura sp. indet. The base of each column is marked with grain size indicators M, F S, C, and C S.
Tapeats Sandstone
This unit usually overlies Precambrian basement rock, also known as the Great Unconformity, and locally overlies the Sixtymile Formation in the eastern Grand Canyon. The lower portion of the unit is typically red sandstone, containing lens-shaped cross-bedded units representing stream deposits with minor overbank shale deposits that rarely contain bioturbation (Rose, Reference Rose2003, Reference Rose2006, Reference Rose2011; Hagadorn et al., Reference Hagadorn, Kirschvink, Raub and Rose2011; Dehler et al., Reference Dehler, Sundberg, Karlstrom, Crossey, Schmitz, Rowland and Hagadorn2024, table S1). The Transition Zone is the upper portion of the unit, and usually manifests as an upsection decrease in thicker, coarser sandstones and a change in weathering from cliffs to more recessive slopes of the Bright Angel Formation. The Transition Zone typically consists of brown, thinner bedded sandstones with common trace fossils including Arenicolites and Skolithos (Rose, Reference Rose2003, Reference Rose2006, Reference Rose2011; Hagadorn et al., Reference Hagadorn, Kirschvink, Raub and Rose2011; Dehler et al., Reference Dehler, Sundberg, Karlstrom, Crossey, Schmitz, Rowland and Hagadorn2024, table S1) and interbedded siltstones and mudstones with trilobites and inarticulate brachiopods. With the exception of eolian facies, these fossils and facies generally indicate an upsection increase in the amount of marine deposition in the Tapeats. This upper Tapeats grades into the overlying Bright Angel Formation, but has its upper boundary drawn at the last thick (≥ 0.5 m) sandstone layer.
Whereas the lower unit of the Tapeats Sandstone does not contain any Cambrian-diagnostic body fossils, detrital zircons from the lower Tapeats suggest an age of generally less than 508 Ma in the eastern sections to 506.5 Ma in the western sections (Karlstrom et al., Reference Karlstrom, Mohr, Schmitz, Sundberg, Rowland, Hagadorn, Foster, Crossey, Dehler and Blakey2020; Sundberg et al., Reference Sundberg2020). The upper Transition zone and lower Bright Angel Formation from Frenchman Mountain and the western Grand Canyon contain trilobites from the upper Dyeran Stage; (Series 2, Stage 4; Webster, Reference Webster2011a; Sundberg et al., Reference Sundberg, Karlstrom, Geyer, Foster, Hagadorn, Mohr, Schmitz, Dehler and Crossey2020) and in the eastern Grand Canyon from the upper Delamaran Stage; (Miaolingian Series, Wuliuan Stage; Fig. 2).
Bright Angel Formation
Rose (Reference Rose2011) and Karlstrom et al. (Reference Karlstrom, Mohr, Schmitz, Sundberg, Rowland, Hagadorn, Foster, Crossey, Dehler and Blakey2020) suggested changing the ‘Bright Angel Shale’ to ‘Bright Angel Formation,’ which is followed herein given the heterolithic nature of the package. In general, the formation is predominately composed of fine-grained siliciclastics (shales to siltstones) but contains common thin to thick beds of sandstones and carbonates. The lower portion of the formation is sandier, and the upper portion contains more carbonates. Sections exposed in the western Grand Canyon consist of proportionally more carbonates, whereas those in the eastern Grand Canyon consist of proportionally more sandstones. The interplay between these lithologies within the Bright Angel Formation is complex given the depositional environments represented (see Wanless, Reference Wanless1973; Rose, Reference Rose2003, Reference Rose2011; Dehler et al., Reference Dehler, Sundberg, Karlstrom, Crossey, Schmitz, Rowland and Hagadorn2024, table S1) and because the package contains several disconformities that are interpreted to increase in magnitude eastward. The Bright Angel Formation can be subdivided into several members, both formal and informal (McKee, Reference McKee1945). These units are, in ascending order (Figs. 2, 3): (1) lower slope unit, (2) Diamond Bar Sandstone, (3) upper slope unit, (4) Lyndon Limestone, (5) Tincan-ebits–Meriwitica interval, and (6) Flour Sack Member. Most of these units are exposed in the western Grand Canyon, but in the eastern Grand Canyon (Figs. 2, 3) these members either are not present or have transitioned laterally into other facies.
Deposition of the Bright Angel Formation took place from the late Dyeran (Epoch 2, Age 4) to medial Topazan, (medial Miaolingian Epoch, upper Wuliuan Age, Fig. 2). In the western Grand Canyon region, it ranges from the late Dyeran to the late Delamaran. In the eastern Grand Canyon, the Bright Angel Formation ranges from the late Delamaran to the early Topazan, indicating diachroneity of both the lower and upper contacts of the formation.
Muav Formation
This formation consists of various massive, cliff-forming carbonate members. These carbonates are separated by slope-forming shales and/or carbonates, which were suggested by McKee (Reference McKee1945) to be tongues of the Bright Angel Formation. In addition, McKee (Reference McKee1945) suggested that the cratonward portion of the massive carbonates change into “rusty-brown dolomites” (RBDs) that were given various names and assumed to be different ages. Established members of the Muav Formation include (in ascending order; McKee, Reference McKee1945): (1) Rampart Cave, (2) Sanup Plateau, (3) Spencer Canyon, (4) Peach Springs, (5) Kanab Canyon, (6) Gateway Canyon, and (7) Havasu members. Rose (Reference Rose2011), using the section at Blacktail Canyon (Fig. 2) as a proposed type section for the Tonto Group, suggested that the Rampart Cave to the Peach Springs members should be included into the Bright Angel Formation, that the Kanab Canyon and Gateway Canyon members should be restricted to the Muav Formation, and that the Havasu Member should be placed into the unclassified portion (undifferentiated dolostones) of the section. The reason for this regrouping was based on the hypothisized unconformities separating the proposed stratigraphic packages. Unfortunately, supporting biostratigraphic evidence of these unconformities is not robust, but that does not mean that such unconformities are not present.
This paper takes an approach more akin to what McKee (Reference McKee1945) did (see Dehler et al., Reference Dehler, Sundberg, Karlstrom, Crossey, Schmitz, Rowland and Hagadorn2024) in separating the members. Redefining the boundaries of the members and viewing them as sequences is a more practical way of determining correlation throughout the Grand Canyon and of conducting facies analyses. These sequences start with the bottom being more shaley/sandy (more slope forming), passing upward to thin-bedded, silty, bioturbated carbonates to massive-bedded, silty carbonates (cliff formers at the top; Dehler et al., Reference Dehler, Sundberg, Karlstrom, Crossey, Schmitz, Rowland and Hagadorn2024, table S1). In some instances, shales and sands do not dominate the lower portion of the sequence. The initial result is the elimination of the “tongues of the Bright Angel Formation” of McKee (Reference McKee1945) within the Muav. This practice is especially difficult to apply given the different ages of the Bright Angel in the western and eastern portions of the Grand Canyon and the unconformities that are present, especially in the eastern portion.
Similar to the Bright Angel Formation, the Muav Formation differs in age in the western to eastern portion of the Grand Canyon. In the western region, the formation ranges in age from late Delamaran to late Topazan (medial to upper Miaolingian). In the eastern region, the base of the formation is younger, late Topazan and ranges to early Marjuman (Fig. 2).
Frenchman Mountain Dolostone
This formation has been recently discussed by Rowland et al. (Reference Rowland, Korolev, Hagadorn and Ghosh2023) and is equivalent to the undifferentiated Cambrian dolostones of McKee and Resser (Reference McKee1945). The Frenchman Mountain Dolostone rests conformably on the Muav Formation and ranges in age from the early Marjuman to early Steptoean (Drumian to early Paibian) based on (1) fossils from the bottom two meters of the formation and underlying Muav Formation in the eastern Grand Canyon; (2) its chemostratigraphic profile recording the DICE (Drumian Carbon Isotope Excursion; Bolaspidella Biozone) near the base of the unit; and (3) the onset of the SPICE (Steptoean Positive Carbon Isotope Excursion), Aphelaspis Biozone (Fig. 3.1) at its top. In the more eastern sections, this formation has been truncated unconformably by Devonian and Mississippian strata. In these eastern outcrops, the Frenchman Mountain Dolostone probably only represents the basal Steptoean (Fig. 3.1).
Biostratigraphy
The Cambrian stratigraphy in the Grand Canyon typically ranges from the global Series 2, uppermost Stage 4 (Laurentian upper Waucoban Series, uppermost Dyeran Stage) to the Furongian Series, lowermost Paibian (Laurentian lower Millardan Series, Steptoean Stage). Within this interval, there are 16 biozones (mainly based on polymeroid trilobites) recognized in the inner shelf to cratonic regions and 14 biozones (Fig. 10) in the outer shelf settings (mainly based on agnostoids; Webster, Reference Webster2011a). Faunas typical of deeper water environments (agnostoids, pagetiids, and oryctocephalids; Sundberg, Reference Sundberg1994, Reference Sundberg2018, Reference Sundberg2020) are not represented in the Grand Canyon; thus, only 16 polymeroid biostratigraphic units potentially can be used for correlation of the Cambrian strata. To date, only seven of those trilobite biozones have been recognized in the Grand Canyon succession.
Biostratigraphic correlation of the Tapeats Sandstone to lowermost Frenchman Mountain Dolostone to age-equivalent strata in California, Nevada, Utah, and Montana. Fm. = Formation, Lms. = Limestone, Mbr. = Member, Sh. =Shale, Ss. = Sandstone. Modified from Schwimmer (Reference Schwimmer1973, Reference Schwimmer1975), Sundberg (Reference Sundberg1994), Webster (Reference Webster2011a), and Miller et al. (Reference Miller, Evans and Dattilo2012).

Figure 10. Long description
A complex stratigraphic chart organized into nine vertical columns.
1. The leftmost columns define time scales: Intern. Laurentian Epochs (Series 2 and Miaolingian), Stages (Waucoban, Dyeran, Delamaran, Wuluian, Lincolnian), and Biozones. The Biozones are color-coded: blue for the oldest (e.g., Arcuolenellus arcuatus), green for middle (e.g., Glossopleura walcotti), and yellow for the youngest (e.g., Ehmaniella).
2. Grand Canyon Column: At the base is Tapeats S s. Overlying this is the Bright Angel F m., which includes the Lower Unit, Transition Zone, and several members like Flour Sack Mbr. Above this is the Muav Formation containing members such as Rampart Cave, Sanup Plateau, and Havasu. The sequence tops with Frenchman Mountain Dolostone.
3. Marble Mountains, C A: Shows Latham Shale at the base, followed by Chambless L m s., Cadiz F m., and Bonanza King F m.
4. Death Valley Area, C A, N V: Features Eagle Mountain Sh. at the base, ascending through Carrara F m. (including Pyramid Sh. and Desert Range L m s.) to Bonanza King F m.
5. Pioche, N V: Starts with Delamar at the base, followed by Pioche F m. (including Combined Metals L m s. and Comet Sh.), Lyndon L m s., Chisholm F m., and Highland Peak L m s.
6. Drum Mountains, U T: Base is Prospect Mountain Qtz., followed by Pioche F m. (Tatow Mbr.), Howell L m s., Chisholm F m., Dome L m s., Whirlwind F m., Swasey F m., and Wheeler Sh.
7. Northwest Montana: Base is Flathead S s., followed by Gordon Sh., Damnation L m s., Dearborn L m s., Pagoda L m s., Pentagon Sh., and Steamboat L m s.
Horizontal lines and wavy unconformity lines correlate these units across the regions, showing how the Tapeats S s. in the Grand Canyon is age-equivalent to the Flathead S s. in Montana and the Prospect Mountain Qtz. in Utah.
Additional age control is based on the occurrence of the SPICE event and the Aphelaspis Biozone in nearby sections (Rowland et al., Reference Rowland, Korolev, Hagadorn and Ghosh2023; Fig. 3.1). The age distribution, which includes the generally unfossiliferous Frenchman Mountain Dolostone, ranges up to the lowermost Furongian Series, lower Paibian Stage (Laurentian lowermost Millardan Series, lowermost Steptoean Stage) at Frenchman Mountain. Another age determination occurs in the eastern Grand Canyon, where maximum depositional ages are based on the youngest detrital zircons in the Sixtymile Formation, which was deposited in a faulted basin and ranges from the Terreneuvian Series, upper Fortunian Stage to upper Series 2, Stage 4 (see Karlstrom et al., Reference Karlstrom, Hagadorn, Gehrels, Mathews, Schmitz, Madronich, Mulder, Pecha, Geisler and Crossey2018, Reference Karlstrom, Mohr, Schmitz, Sundberg, Rowland, Hagadorn, Foster, Crossey, Dehler and Blakey2020).
Paleontological information is derived from the 76 historical collections reported by McKee and Resser (Reference McKee and Resser1945) and an additional 122 new collections made as part of this study. The older collections were restudied and are housed in the Smithsonian Institution (USNM) and Grand Canyon Museum Collection (GRCA). Some of those collections, particularly those made by Walcott, contain specimens preserved in a variety of lithologies and thus appear to be pooled from broad stratigraphic intervals that might span different biozones. In addition, some identifications from McKee and Resser (Reference McKee and Resser1945) were incorrect (e.g., some Kootenia specimens were identified as Anoria), giving misleading biostratigraphic indications (see discussion below). In general, fossils are relatively rare and poorly preserved. No body fossils were found within the lower Tapeats Sandstone, and those from the Muav Formation are mostly preserved in the thinner shale of the ‘Bright Angel tongues’ or in thin bioclastic lags in carbonates (McKee, Reference McKee1945; this study).
In general, the rocks of the Tonto Group are not fossiliferous. Some horizons do contain abundant fossils, but these are generally limited in stratigraphic and geographic distribution and are sometimes only recovered as talus. As a result, no individual section contains a continuous fossiliferous sequence to determine the stratigraphic ranges of taxa and, in most cases, the boundaries between biozones cannot be accurately determined. As a result, boundaries between biozones are shown as dashed lines in Figures 4–9, and are tied in part to lithologic correlations. Nonetheless, the strata contain representatives of seven trilobite biozones.
The Laurentian traditional lower–middle Cambrian boundary is marked by extinction of the olenelloid trilobites and the first occurrence of the ptychoparioid trilobite Eokochaspis nodosa Sundberg and McCollum, Reference Sundberg and McCollum2000. This taxon delineates the boundary between the Waucoban and Lincolnian series. This boundary is older than the boundary between the global Series 2 (undefined) and Miaolingian, which is marked by extinction of the redlichid trilobites and the first occurrence of Oryctocephalus indicus (Reed, Reference Reed1910), an oryctocephalid trilobite. The boundary is defined in South China (Zhao et al., Reference Zhao, Yuan, Babcock, Guo and Peng2019) and appears to be two biozones above the last occurrence of olenelloids in Laurentia (see Lin et al., Reference Lin, Sundberg, Jiang, Montañez and Wotte2019; Sundberg et al., Reference Sundberg2020; Landing et al., Reference Landing, Schmitz, Westrop and Geyer2023).
Evidence for the biozones represented in the Cambrian of the Grand Canyon is discussed below. Each biozone is discussed separately in ascending order and includes a discussion of taxa reported by Resser (in McKee and Resser, Reference McKee1945) and material our group collected during field seasons in 2019–2024 (Fossil Locality Supplement; https://doi.org/10.5281/zenodo.14879978). Details about specimens are provided under the systematic sections. Correlation of biozones in the absence of fossils is based on lithologic and chemostratigraphic correlations.
Waucoban Series, Dyeran Stage
Six biozones were recognized by Webster (Reference Webster2011a; Fig. 10) in the upper portion of the Dyeran Stage. Based on the correlation of faunas from western Nevada to Siberia (Sundberg and Webster, Reference Sundberg and Webster2021; Sundberg et al., Reference Sundberg, Webster and Geyer2022), these biozones are apparently younger than the first occurrence paradoxidid trilobites in Avalonia at ca. 509 Ma (see Sundberg et al., Reference Sundberg2020). Of the six biozones, only three have been identified at Frenchman Mountain and the western portion of the Grand Canyon (Figs. 2–6): the Peachella iddingsi, Bolbolenellus euryparia, and Nephrolenellus multinodus biozones. The Peachella iddingsi Biozone has only been recognized from the Transition Zone of the Tapeats Sandstone at the Frenchman Mountain section, Nevada (Webster, Reference Webster2011b). The uppermost Bolbolenellus euryparia Biozone is also from the Transition Zone of the Tapeats Sandstone at Frenchman Mountain, based on the co-occurrence of Bristolia brachyomma Palmer in Palmer and Halley, Reference Palmer and Halley1979, Olenellus gilberti Meek in White, Reference White1874, and O. fowleri Palmer, Reference Palmer1998 (Webster, Reference Webster2011b). Biceratops nevadensis Pack and Gayle, Reference Pack and Gayle1971, was previously only known from this biozone, but its occurrence with Nephrolenellus sp. indet. at our Diamond Bar Ranch section suggests a slightly higher stratigraphic range of the species. The Nephrolenellus multinodus Biozone ranges from the Transition Zone of Tapeats Sandstone to the Bright Angel Formation at Frenchman Mountain to the Tapeats Transition Zone at the Quartermaster Canyon section. Most of the trilobites are preserved in sandstones, which generally prevent specific identifications. However, several taxa can be recognized, including Biceratops nevadensis, Mesonacis fremonti (Walcott, Reference Walcott1910), Nephrolenellus cf. N. multinodus (Palmer in Palmer and Halley, Reference Palmer and Halley1979), Olenellus gilberti, and O. terminatus Palmer, Reference Palmer1998. Other material is referred to as olenelloid specimens and occurs eastward toward our Pumpkin Springs section.
Nephrolenellus spp. are limited to the Nephrolenellus multinodus Biozone (Webster, Reference Webster2011b). Mesonacis fremonti ranges through the six upper Dyeran biozones and is not helpful in determining a specific biozone. The Nephrolenellus multinodus Biozone occurs at Frenchman Mountain, Rampart Cave, Mile 263, Peach Springs, Pumpkin Springs, and Diamond Bar Ranch sections. Localities at Meriwitica Canyon, Bridge Canyon, Peach Springs, and Pumpkin Springs do not have biozone-indicative taxa but are inferred to be N. multinodus Biozone based on their stratigraphic position.
Lincolnian Series, Delamaran Stage
Five biozones were recognized by McCollum and Sundberg (Reference McCollum and Sundberg2007; Figs. 2, 10) in the Delamaran Stage, of which only three are present in the Grand Canyon. The Eokochaspis nodosa Biozone has only been recognized from Frenchman Mountain, as indicated by Mexicella cf. Me. robusta Sundberg and McCollum, Reference Sundberg and McCollum2000 (Fig. 4; see Table 2 for generic abbreviations). These specimens are morphologically intermediate between Me. antelopea Sundberg and McCollum, Reference Sundberg and McCollum2000, and Me. robusta. The former occurs in the middle portion of the Eo. nodosa Biozone in the Pioche Formation and the latter occurs in the lower portion of the Am. arrojosensis Biozone also in the Pioche Formation (Sundberg and McCollum, Reference Sundberg and McCollum2000). If an evolutionary gradation occurs between Me. robusta and Me. antelopea, it may indicate that Mexicella cf. Me. robusta represents the middle to upper portion of the Eo. nodosa Biozone. The overlying Amecephalus arrojosensis and Poliella denticulata biozones have not been identified in the Frenchman Mountain section or the Grand Canyon area.
List of generic abbreviations used in text

Table 2. Long description
The table consists of two columns: Genus and Abbreviation. The entries are as follows:
* Albertella: Alb.
* Amecephalus: Am.
* Anoria: A.
* Biceratops: Bi.
* Blainia: B.
* Bolaspis: Bol.
* Bolbolenellus: Bo.
* Caspimexis: C.
* Ehmania: Ehm.
* Ehmaniella: Eh.
* Eldoradia: E.
* Elrathiella: El.
* Eokochaspis: Eo.
* Glossopleura: G.
* Glyphaspis: Gl.
* Kochina: Ko.
* Kootenia: K.
* Mesonacis: M.
* Mexicella: Me.
* Nephrolenellus: N.
* Olenellus: O.
* Pachyaspis: Pac.
* Parehmania: Par.
* Piochaspis: P.
* Proehmaniella: Pro.
* Solenpleurella: Sol.
* Spencella?: Spe.?
* Spencia: S.
* Trachycheilus: T.
* Trymataspis: Ty.
* Zacanthoides: Z.
The Mexicella mexicana Biozone has been recognized at Frenchman Mountain (approximately 100 m above the base of the Bright Angel Formation; Pack and Gale, Reference Pack and Gayle1971; Fig. 4), and at the Rampart Cave section (Fig. 5). Prior to this report, this biozone was recognized in the Grand Canyon based on one specimen of Albertella schenki Resser, Reference Resser1945, which, without the associated sclerites, made its identification questionable. This single occurrence lead Rose (Reference Rose2011) to suggest that the presence of the biozone is problematic in the Grand Canyon. We document the presence of this biozone in the Grand Canyon as indicated by the presence of several new specimens of Albertella schenki, Alb. nebeli n. sp., Me. mexicana Lochman, Reference Lochman1948, and Caspimexis cf. C. radiatus (Palmer in Palmer and Halley, Reference Palmer and Halley1979). Albertella schenki, Me. mexicana, and C. radiatus have been reported from the Albertella highlandensis subzone, upper Me. mexicana Biozone of the Pioche and/or Carrara Formation (Palmer and Halley, Reference Palmer and Halley1979; Eddy and McCollum, Reference Eddy and McCollum1998; Sundberg, Reference Sundberg2011; Fig. 10).
The Glossopleura walcotti Biozone occurs throughout the Grand Canyon area (Figs. 2–9), almost always below the Rampart Cave Member of the Muav Formation and its rusty brown dolostone correlative, the Elves’ Chasm Tongue. This biozone is characterized by the presence of Glossopleura boccar (Walcott, Reference Walcott1916b) and G. tuta Resser, Reference Resser1945. It is possible that the G. walcotti Biozone could be subdivided into two or more biozones or subzones. Little work has been done on this biozone from bottom to top, primarily because of the massive Bonanza King Formation and lithologic equivalents that occur in the middle of the biozone. The first occurrence of G. walcotti occurs just a few meters below the basal Bonanza King Formation in the uppermost Carrara Formation of California and Nevada (Palmer and Halley, Reference Palmer and Halley1979; Fig. 10). Associated with this species are G. lodensis (Clark, Reference Clark1921), G. tuta Resser, Reference Resser1945, and Alokistocarella brighamensis (Resser, Reference Resser1939b). Sundberg (Reference Sundberg2018) reported G. walcotti and a diverse trilobite fauna from the basal part of the Emigrant Formation, but the strata immediately overlying these faunas have not yielded faunas older than the Ehmaniella Biozone.
In contrast, faunas from the upper part of the G. walcotti Biozone contain G. boccar, Amecephalus laticaudum (Resser, Reference Resser1939a), Achlysopsis cf. A. liokata Fritz, Reference Fritz1968, and Kochina vestita Resser, Reference Resser1939b (Sundberg, Reference Sundberg2005). Morgan (Reference Morgan2021) suggested that the G. walcotti Biozone could be divided into a lower G. walcotti and an upper G. boccar biozones. However, there are no known published sections that cross the transition/boundary between the two potential biozones. Campbell’s (Reference Campbell1974) study of trilobites from the Brigham Quartzite to the Ute Formation, Utah, does cross a potential biozone boundary, but unfortunately he recognized neither G. walcotti nor G. boccar and the work remains unpublished (Campbell, Reference Campbell1974, fig. 4).
Taxa that represent this biozone include Amecephalus althea (Walcott, Reference Walcott1916a), Am. cf. Am. normale (Resser, Reference Resser1938b), Am. pioch-ensis (Walcott, Reference Walcott1884), Am. scapegoatensis (Deiss, Reference Deiss1939), Anoria tontoensis (Walcott, Reference Walcott1916b), A.? bessus (Walcott, Reference Walcott1916b), Athabaskia sp. indet., G. boccar, G. tuta, G. spp. indet., Kochina vestita, Kootenia tetraspinosa Resser, Reference Resser1945, Piochaspis nites (Resser, Reference Resser1945), and Zacanthoides cf. Z. typicalis (Walcott, Reference Walcott1886).
Lincolnian Series, Topazan Stage
The Topazan Stage was defined by Sundberg (Reference Sundberg2005) based on the first occurrence of Proehmaniella basilica (Resser, Reference Resser1945) (Fig. 10), which overlies the last occurrence of Glossopleura within a 10-cm-thick shale in the Drum Mountains, Utah. The Topazan Stage is equivalent to the upper half of the Wuluian Stage of the Miaolingian Series. This stage consists of a single biozone, the Ehmaniella Biozone, which occurs throughout the Grand Canyon area (Figs. 2–9), almost always above the Rampart Cave Member of the Muav Formation and its rusty brown dolomite correlative, the Elves’ Chasm Tongue.
Sundberg (Reference Sundberg1994) subdivided the Ehmaniella Biozone into the Proehmaniella, Elrathiella, Ehmaniella, and Altiocculus subzones based on distinct assemblages that occur across Utah and Nevada. These subzones were not defined as biozones due to the barren intervals at the boundary intervals of the Proehmaniella–Elrathiella subzones and the Ehmaniella–Altiocculus subzones.
The Proehmaniella subzone is represented by Kootenia havasuensis Resser, Reference Resser1945, Pro. basilica, Pro. moorei (Resser, Reference Resser1945), and Zacanthoides walapai Resser, Reference Resser1945, in the Elves’ Chasm Tongue, Bright Angel Formation, and upper Rampart Cave Member. Such fossils occur in sections in the eastern Grand Canyon (Figs. 4–6). The Elrathiella subzone is represented by Elrathiella tontoensis (Resser, Reference Resser1945), El. cf. El. decora Resser, Reference Resser1945, Trachycheilus granulosus Sundberg, Reference Sundberg1994, and T. whirlwindensis Sundberg, Reference Sundberg1994. Elrathiella tontoensis is commonly in the heavily glauconitic layers of the Bright Angel Formation in the eastern Grand Canyon, whereas the latter three taxa are from the Muav Formation in the western Grand Canyon.
Differentiating between the Ehmaniella and Altiocculus subzones in the Grand Canyon is difficult given the limited faunal distribution in the Muav Formation. The Altiocculus subzone as originally defined (Sundberg, Reference Sundberg1994) contrasts with the other subzones in its more oceanward depositional environment (Sundberg, Reference Sundberg, Cooper and Stevens1991); thus, the correlation of species in the Grand Canyon to the Ehmaniella and Altiocculus subzones is unclear. It should be noted, however that Ehmaniella spp. typically occur in open ocean settings (Rasetti, Reference Rasetti1951). Presently, there are no definitive criteria to separate the two subzones in the Grand Canyon given the stratigraphic ranges of trilobites found in the Peach Springs to Gateway Canyon members of the Muav Formation. As a result, only the Ehmaniella subzone is used (Fig. 10).
The Ehmaniella subzone is represented by Kootenia mckeei Resser, Reference Resser1945, K. barensis n. sp., K. erosa (Resser, Reference Resser1945) n. comb., Spencella? porcata (Resser, Reference Resser1945), Ehmania weedi Resser, Reference Resser1935, Ehmaniella kwaguntensis (Resser, Reference Resser1945) n. comb., Eh. hebes Resser, Reference Resser1945, Eh. nitida (Resser, Reference Resser1945) n. comb., and Glyphaspis vulsa Resser, Reference Resser1945. Such fossils occur in both the western and eastern Grand Canyon (Figs. 4–9).
Using genera to establish biozones or subzones limits precision. For example: Spencella? ranges from the Proehmaniella subzone to the Bolaspidella Biozone (Rasetti, Reference Rasetti1963; White, Reference White1973; Sundberg, Reference Sundberg1994) in other locations in the USA; Ehmaniella ranges from the Ehmaniella subzone to at least the basal Altiocculus subzone (Sundberg, Reference Sundberg1994); and Glyphaspis is known from the Altiocculus subzone to the Bolaspidella Biozone (Sundberg, Reference Sundberg1994; Melzak and Westrop, Reference Melzak and Westrop1994). Thus, it is possible that some of the trilobite taxa range up into the overlying Bolaspidella Biozone. Resolution of the boundary between the Ehmaniella and Bolaspidella biozones will need more study (see discussion below).
Lincolnian Series, Marjuman Stage
Faunas from the Marjuman Stage are representative of the Bolaspidella Biozone. This biozone is represented by the occurrence of Glyphaspis tetonensis Resser, Reference Resser1937, Crepicephalus? upis Walcott, Reference Walcott1916a, Solenopleurella? quadrata Rasetti, Reference Rasetti1963, and Modocia sp., which are known from the Bolaspidella-equivalent strata in Canada (Melzak and Westrop, Reference Melzak and Westrop1994), Montana (Schwimmer, Reference Schwimmer1973), and the eastern US (Rasetti, Reference Rasetti1963). This is first time the Bolaspidella Biozone has been reported from the Grand Canyon.
In the Drum Mountains, which is the type section for the global Drumian Stage (Babcock et al., Reference Babcock, Robison, Rees, Peng and Saltzman2007), the boundary between the Ehmaniella and Bolaspidella biozones is based on the first occurrence of Bolaspidella sp. (White, Reference White1973). However, the last occurrence of faunas typical of the Ehmaniella Biozone, diagnostic ptychoparioids, occurs 62 m below the occurrence of Bolaspidella Biozone; thus, the boundary could actually occur somewhere within this interval. The first occurrence of the Bolaspidella Biozone, at present, coincides with the base of the global Drumian Stage, which is based on the first-appearance datum (FAD) of Ptychagnostus atavus (Tullberg, Reference Tullberg1880). The DICE also occurs near the base of the Bolaspidella Biozone and the Drumian Stage (Babcock et al., Reference Babcock, Robison, Rees, Peng and Saltzman2007).
The Bolaspidella Biozone is recognized only in the easternmost Grand Canyon, at our Nankoweap, Buck Farm Canyon, and possibly Surprise Canyon sections (Fig. 9).
Conflicting biostratigraphic indicators
Some of the collections reported by McKee and Resser (Reference McKee and Resser1945) contain conflicting biostratigraphic indicators (e.g., a single locality that reportedly contains taxa representing both the G. walcotti and Ehmaniella biozones). These conflicts may result from taxa having longer stratigraphic ranges than previously recognized (range extension), from the pooling of specimens from several stratigraphic horizons into a single collection (e.g., as part of a preliminary survey, and/or from float material), or from taxonomic misidentification (e.g., due to poor preservation). The present study took several steps to alleviate these issues: specimens were marked as “f” for “Float” to indicate a surface collection; type and associated specimens were inspected for potential misidentification; and differences in lithology among slabs from a single collection were noted (e.g., red sandstone vs. bioclastic limestone).
McKee and Resser (Reference McKee and Resser1945, p. 172, loc. 19) reported from USNM loc. 74 (Fig. 9) Am. althea, Anoria tontoensis, G. boccar, Gl. vulsa, and El. arizonaensis (Resser, Reference Resser1945) n. comb. The representatives of the Ehmaniella Biozone, Gl. vulsa, are in limestone, and El. arizonaensis n. comb. are in micaceous, fine-grained sandstone, whereas the other species that represent the G. walcotti Biozone are preserved in medium-grained sandstone. This lithological heterogeneity suggests a mixed sample due to sampling at different horizons and/or through float samples.
A similar situation occurs at USNM loc. 73a (Fig. 9) where A. tontoensis, Pro. moorei?, Elrathiella fonticola (Resser, Reference Resser1945), and Olenoides? sp. indet. are reported from a coarse-grained, dark-red sandstone. No sclerites of Anoria were found with the type specimens. The type of El. fonticola from USNM loc. 44337 is reported with Am. piochensis? and A. tontoensis, but McKee and Resser (Reference McKee and Resser1945, p. 211) reported the type from a purple, glauconitic sandstone overlying the sandy shales that probably contain the latter two species. Another potential of pooling horizons is USNM loc. 74a, where specimens of Elrathiella? insueta Resser, Reference Resser1945, are tectonically distorted, but those of Ehmaniella hebes are not.
One of the difficulties with identifications in McKee and Resser (Reference McKee and Resser1945) is the similarity in pygidia between the corynexochids Anoria, Athabaskia, Glossopleura, and Kootenia and the ptychoparioid Glyphaspis. They all have similar subcircular pygidia that, when specimens are poorly preserved, can be confused. The pygidia of Anoria and Glossopleura cannot be distinguished without associated cranidia and librigenae. When such features are absent, the two taxa are lumped under Glossopleura sp. indet. herein. Kootenia generally has pygidial spines, but some forms in the Grand Canyon do not have spines. If the spines are absent or covered, a distinctive character of Kootenia is its narrow, well-defined pygidial border and longer axis. Pygidia of Glyphaspis have a narrower, more tapered, less inflated axis with the pleural furrows extending farther onto the border than Anoria or Glossopleura. The primary difference between the pygidia of the corynexochid Athabaskia and pygidia of the ptychoparioid Glyphaspis is that the pleural and interpleural furrows merge laterally in the latter. The cranidia of Athabaskia and Glyphaspis are distinctly different, and the association of the pygidia with the cranidia is the best way to distinguish the two taxa. Historical identifications of these taxa were revised based on these criteria for McKee and Resser’s USNM locs. 44338F, 44338G, 44343A (material not preserved well enough to determine taxon), 44344B, 44345F, 44345E, 443381 (Kootenia erosa, Spencella? sp. indet.), and GRCA loc. 356. Anoria sp. reported from USNM 44345B was probably float based on similar specimens found in the same stratigraphic position in the section (personal observations) and the lithology, preservation, and taxa similar to the sandstone interval between the Tincanebits and Meriwitica tongues in nearby sections.
Disconformities
McKee and Resser (Reference McKee and Resser1945) did not envisage major unconformities within the Cambrian of the Grand Canyon. In contrast, Rose (Reference Rose2003, Reference Rose2011, fig. 1A) separated the strata into four packages with proposed unconformities based on lithologic correlations and scant paleontological evidence. Herein, we recognize three “major” disconformities in the Cambrian of the Grand Canyon based on lithologic and paleontological data: (1) between the Dyeran and Delamaran stages, (2) within the Delamaran Stage, and (3) between the Delamaran and Topazan stages (Figs. 2–6). Other unconformities may exist higher in the Muav Formation and/or Frenchman Mountain Dolostone, but at present there is no paleontological evidence to constrain their scope beyond evidence for pronounced facies change or sequence boundaries (e.g., Brathovde, Reference Brathovde1986; Hardy, Reference Hardy1986; Korolev, Reference Korolev1997).
The unconformity between the Dyeran and Delamaran stages becomes more pronounced in an eastward direction in the Grand Canyon. In the Frenchman Mountain section, Mexicella cf. Me. robusta is present only 1 or 2 m above the local last occurrence of olenelloids (Fig. 4). The fossil material is preserved in red, glauconitic sandstone and as limonite molds and casts. The base of the red sandstones suggests the presence of an erosional surface (Fig. 11). Sample FHPR 008-5 was collected from the uppermost sandstone layer. Sample FHPR 008-4 consists of a small cranidium from a silty layer in the overlying shales exposed a few meters southward of the measured section. This sample is compatible with Me. cf. Me. robusta but is not preserved well enough to be accurately assigned to a species. These trilobite samples occur approximately 1–2 m above the last occurrence of olenellids, if the red sandstones correlate to the highest sands in Webster (Reference Webster2011b, figure 2-2; loc. ICS-10321) at the Frenchman Mountain section. This occurrence suggests that a hiatus occurred during the lower and possibly middle E. nodosa Biozone. The magnitude of the disconformity increases eastward, where in the central transect it appears to increase in duration to extend to the lower G. walcotti Biozone. This change is documented in part by the elimination of the red silty/sand unit (Diamond Bar Member) eastward. It is unknown how much, if any, of the Dyeran exists in the eastern transect.
Dyeran–Delamaran disconformable boundary (wavy red line) in the Bright Angel Formation, Frenchman Mountain, Nevada. Short stratigraphic section illustrating the location of trilobite-bearing beds (red circles) with the possible location of upper Dyeran olenellids (ICS-10321; from Webster, Reference Webster2011b) and Mexicella cf. Me. robusta from the Eokochaspis nodosa or Amecephalus arrojosensis biozones in the upper glauconitic sandstone. Section measured by Eben Rose.

Figure 11. Long description
A two-panel layout. On the left is a vertical stratigraphic column measured in meters from 0.0 to 3.5.
* From 0.0 to 1.6 meters, the column shows alternating layers of siltstone and sandstone with cross-bedding. A red circle at 0.6 meters is labeled I C S dash 10321 question mark.
* At 1.6 meters, a wavy red line indicates a disconformable boundary.
* From 1.6 to 3.0 meters, the section is shaded pink, representing glauconitic sandstone. Three red circles are clustered near the top of this unit, labeled F H P R 008 dash 4, F H P R 008 dash 5, and F H P R 008 dash 6.
* Above 3.0 meters, the section returns to a siltstone pattern.
In the center, vertical text identifies fossil zones. Olenelloid sp. is marked with a blue dot near the 0.5 meter mark. Mexicella cf. Me. robusta is marked with a green line and dots spanning from 2.8 to 3.3 meters. A thick black arrow points from the 1.6 meter boundary on the column to a specific layer in the photograph. A red arrow points from the upper red circles to the top layer of the photograph.
On the right, a field photograph shows the rock outcrop. A blue-handled rock hammer with a purple rope is placed against the rock for scale. The hammer head rests on a greenish-gray sandstone layer, which corresponds to the black arrow from the stratigraphic column. The rock above the hammer is darker and more massive, corresponding to the pink-shaded sandstone in the diagram.
The disconformity within the Delamaran Stage occurs between the Me. mexicella and G. walcotti biozones in the western portion of the Grand Canyon. In Nevada, these two biozones are normally conformable (Palmer and Halley, Reference Palmer and Halley1979), although in the Pioche area of eastern Nevada the two biozones have an unconformable contact (Eddy and McCollum, Reference Eddy and McCollum1998; McCollum and McCollum, Reference McCollum and McCollum2011; Fig. 10). At Rampart Cave in the Grand Canyon (Fig. 5) there is evidence of erosion having occurred between the two biozones in the upper Bright Angel Formation (Fig. 12). The erosion is visible at two localities at Rampart Cave (Figs. 12, 13) that are well exposed due to the overlying cliffs of sandstone and the Tincanebits Tongue, whereas these intervals are generally on slopes covered with a considerable amount of talus. One locality (GRCA loc. 391) illustrates a disconformity with a minimum relief of 1.5 m across a 5-m-horizontal interval (Fig. 12). Platy shale with minimal bioturbation containing Alb. schenki and underlying strata was eliminated from the west to the east due to the erosion. Deposited on top of the erosional surface are extensively bioturbated shales with sandstone interbeds (Fig. 12.3, 12.5). Thicker sandstone beds are deposited parallel to the erosional surface, and some are distorted (Fig. 12.1, 12.2). These beds may represent small channels that have been deformed due to shale compaction. Overlying this bioturbated shale is about 20 cm of bioturbated sandstone and a thin package of bioturbated shales that contain Glossopleura and Amecephalus. Albertella schenki represents the Me. mexicana Biozone, whereas Glossopleura represents the G. walcotti Biozone. No body fossils occur in the interval between the erosional surface and the first occurrence of Glossopleura.
Delamaran disconformity in the Bright Angel Formation, Rampart Cave section, illustrating the erosional contact between the Mexicella mexicana Biozone and overlying deposits bearing faunas of the Glossopleura walcotti Biozone. (1–3) Outcrop photograph and interpretive drawing showing erosional surface; note hammer (white circle) for scale; white rectangle in (1) marks the position of (3) close-up photograph of ~10 cm of platy shale and erosional surface; (4) outcrop of platy shale over 1 m thick taken a few meters to the right of the picture in (1); (5) bioturbation of shale and thin-bedded sandstones overlying the erosional surface.

Figure 12. Long description
A multi-panel display of geological outcrops and diagrams.
Panel 1: A wide-angle photograph of a rock face with horizontal layering. A white circle highlights a rock hammer for scale, and a white rectangle labeled Fig. 3 indicates the location of a close-up.
Panel 2: An interpretive line drawing of Panel 1. From bottom to top, it shows white Talus at the base, followed by a red Shale layer. A red dashed line marks the Erosional surface. Above this surface are white Sandstone distorted beds and a large green area labeled Shale/Sandstone. Labels include Glossopleura sp. on the left and Albertella schenki on the right.
Panel 3: A close-up photograph of the stratigraphy. From bottom to top, the layers are labeled Red and Green Sandstones, followed by a Pinch-out of Platy Shale with minimum bioturbation (marked by a double-headed red arrow), an Erosional surface, and Bioturbated Shales with thin Sand Layers at the top.
Panel 4: A photograph of a taller outcrop section. A person in a red shirt stands at the bottom right for scale. A long vertical double-headed red arrow spans a thick section of Platy Shale with minimum bioturbation, which sits above Red and Green Sandstones.
Panel 5: A high-detail close-up of rock texture with a 5 cm scale bar. It shows thin, dark, platy layers of shale interspersed with lighter, rounded sandy inclusions, illustrating bioturbation.
Delamaran disconformity in the Bright Angel Formation, Rampart Cave section, illustrating the flat erosional contact (white arrow) between the Mexicella mexicana Biozone and overlying deposits bearing faunas of the Glossopleura walcotti Biozone. (1) Exposure of the fossil locality showing the ranges of the Mexicella mexicana and Glossopleura walcotti biozones (double-ended line indicates that the biozones range down or up). (2) Contact between the uppermost Me. mexicella Biozone fauna and the overlying sandstones possibly representing a disconformity.

The second locality (Fig. 13) is approximately 200 m north of the section described above and shows no obvious erosional surface and intervening shales and sandstones. Instead, shales containing A. schenki and Caspimexis cf. C. radiatus are overlain by a 20-cm-thick bioturbated sandstone, which in turn is overlain by 30 cm of bioturbated shales that contain Glossopleura and Amecephalus (Fig. 13.1). If a disconformity exists, it is probably represented by the base of the 20-cm-thick sandstone (white arrow in Fig. 13.2).
Significant erosion between the two biozones is further supported by the eastward disappearance of the underlying Diamond Bar Member and the lower slope unit of the Bright Angel Formation. With the elimination of these two lithologic packages, the underlying disconformity and the Delamaran disconformity merge (Fig. 3.2).
The disconformity between the Me. mexicana and G. walcotti biozones indicates an erosional event in the Grand Canyon area that has gone unrecognized. Rose (Reference Rose2006, Reference Rose2011) suggested an unconformity at this horizon based on lithologic patterns. The paleontological and lithologic evidence here supports Rose’s unconformity hypothesis. In the Pioche area, Nevada, disconformities and/or shallowing events have been recognized between several biozones (see McCollum and McCollum, Reference McCollum and McCollum2011; Fig. 10). An example is at or near the upper boundary of the Nephrolenellus multinodus (although see discussion by Webster, Reference Webster2011a, of sequence development below the traditional lower–middle Cambrian boundary and the consistent thickness and presence of the uppermost sequence IV), Amecephalus arrojosensis, Poliella denticulata, and Me. mexicana biozones. Given the more cratonward position of the Grand Canyon Cambrian units, these disconformities and shallowing events should be more pronounced.
The magnitude of the disconformity is difficult to determine. In the Carrara Formation in Nevada and California (Palmer and Halley, Reference Palmer and Halley1979), the stratigraphically lowest Glossopleura species in the G. walcotti Biozone is G. walcotti, which does not occur in the Grand Canyon. In the upper Chisholm Formation in the Drum Mountains, Utah (Sundberg, Reference Sundberg2005), the stratigraphically highest species is G. boccar, which is common in the Bright Angel Formation. The absence of G. walcotti suggests that strata that could contain this species are missing in the Grand Canyon. This absence is further supported with the disappearance of the Me. mexicana Biozone faunas in the central transect. Unfortunately, detailed biostratigraphic ranges within the entire G. walcotti Biozone have not been published.
The disconformity between the Delamaran and Topazan stages is best represented in the eastern transect, where the G. walcotti Biozone is overlain by the Elrathiella subzone, Ehmaniella Biozone (Figs. 3, 8, 9). Missing is the Proehmaniella subzone, Ehmaniella Biozone, which is present in the Peach Springs and Gateway Canyon sections (Figs. 5, 6). This disconformity is represented at our Malgosa section (Figs. 9, 14), where shales are sharply overlain by ~5-m-thick, cross-bedded, arkosic sandstones, which in turn are overlain by glauconitic shale/siltstones (Fig. 14.1). The contact between the arkosic sandstones and underlying mudstones has small-scale channel features. Biostratigraphic constraints on the disconformity are not as refined as for the previously discussed disconformities but include faunas of the G. walcotti Biozone 22 m below the contact and Elrathiella subzone, Ehmaniella Biozone faunas 5 m above the arkosic sandstone (Fig. 14.3).
Delamaran–Topazan disconformity in the Bright Angel Formation, Malgosa Canyon section, illustrating the flat erosional contact (black and white arrow) between the Glossopleura walcotti Biozone and Elrathiella subzone, Ehmaniella Biozone. (1) Cliff exposure illustrating the sharp contact between the arkosic sandstone and underlying shale and the overlying glauconitic shales. (2) Overall view of Bright Angel Formation and overlying Muav Formation with arrow indicating (1). (3) Measured section of the Bright Angel Formation from the slope illustrated in (2) showing fossil locations. (4) Close-up of cross-bedded arkosic sandstones in the upper part of the measured section.

Figure 14. Long description
Panel 1: A photo of a cliff face showing a sharp horizontal contact between lower greenish-yellow shale and upper reddish-brown arkosic sandstone. A black and white arrow points from this contact toward Panel 2. A researcher stands at the bottom right for scale.
Panel 2: A wide-angle photo of a large mountain slope. The lower section consists of dark slopes, the middle section shows the Bright Angel Formation with distinct horizontal banding, and the top section is the massive Muav Formation cliff. An arrow from Panel 1 points to a specific dark ledge on the slope.
Panel 3: A stratigraphic column of the Bright Angel Formation. The vertical Y-axis measures height in meters from 0 to 75. The X-axis labels grain size from left to right: M, S, F S, C S, and C.
* The base at 0 meters is the Transition Zone.
* From 0 to 50 meters, the column shows repeating patterns of shale and sandstone with black dots.
* A vertical green line labeled Glossopleura sp. extends from 0 to 35 meters with three green circular markers.
* A vertical black line labeled Elrathiella arizonaensis extends from 0 to 55 meters, ending in a yellow circle.
* Between 50 and 60 meters, there are distinct pink and green shaded layers. An arrow points from the pink cross-bedded layer at 53 meters to Panel 4.
* The top of the section at 75 meters is labeled Muav Limestone.
Panel 4: A close-up photo of the reddish-brown arkosic sandstone. A yellow and black Jacob's staff is placed vertically against the rock for measurement. The rock shows clear cross-bedding structures where sediment layers are inclined at angles to the main horizontal strata.
Lithologic correlations
Additionally, there are three challenges in correlating finer-scale lithologies in the Tonto Group: (1) the inconstancy of the definition of lithologic units (cliff/slope weathering profiles do not necessarily reflect lithology or ages); (2) the paucity of paleontological data to constrain potential correlations; and (3) the rapid lateral changes in lithologies in the central and eastern portions of the canyon. Figures 2 and 3 illustrate this conundrum, and reflect an attempt to correlate the Tapeats Sandstone, Bright Angel Formation, Muav Formation, and Frenchman Mountain Dolostone across the Grand Canyon area.
The correlation in the western transect (Figs. 4, 5) is relatively straightforward, although its nomenclatural history is not. Due to the similarity of stratigraphic packages to the north (Pioche, Nevada) and to the west (southern Nevada, Carrara Formation; Fig. 10), it has been suggested that Tonto Group is represented by the Prospect Mountain Quartzite, Pioche Formation, Lyndon Limestone, Chisholm Shale, and Peasley Limestone (part of the Highland Peak Formation) (Schenk and Wheeler, Reference Schenk and Wheeler1942; Pack and Gale, Reference Pack and Gayle1971; Webster, Reference Webster2011a, Reference Websterb; Rowland, Reference Rowland2011; Rowland et al., Reference Rowland, Korolev, Hagadorn and Ghosh2023).
With our new paleontological data, more precise lithologic correlations can be made to test these hypotheses. For example, the upper Tapeats Sandstone and lower Bright Angel Formation contain lithologies and olenelloid faunas similar to the Delamar and Combined Metals members of the lower Pioche Formation and the Pyramid Shale member of the Carrara Formation (shales and sandstones, but lacking limestones). Lithologies above the olenelloid faunas (lower Delamaran) correlate in age to the upper Pioche Formation (Fig. 10), but are sandstone dominated as opposed to shale and limestone (McCollum and McCollum, Reference McCollum and McCollum2011), and to the upper Pyramid Shale member (Palmer and Halley, Reference Palmer and Halley1979). The platy shales below the Lyndon Limestone and correlative Tincanebits Tongue are equivalent in age (Me. mexicana Biochron) and general shale lithology to the Grassy Springs Member of the Pioche Formation (McCollum and McCollum, Reference McCollum and McCollum2011; Fig. 10) and the Pahrump Hills Shale of the Carrara Formation (Palmer and Halley, Reference Palmer and Halley1979; Fig. 10). The Lyndon Limestone (used as a member by Rowland et al., Reference Rowland, Korolev, Hagadorn and Ghosh2023, which is followed here) is the lowest limestone-dominated unit in the Bright Angel Formation. This limestone and overlying shales (Flour Sack Member) are age equivalent to the Lyndon Limestone and Chisholm Shale to the north in the Pioche area (Sundberg, Reference Sundberg1994, Reference Sundberg2005, Reference Sundberg2011; McCollum and McCollum, Reference McCollum and McCollum2011) and to the Jangle Limestone and Desert Range Limestone members of the Carrara Formation (Palmer and Halley, Reference Palmer and Halley1979). The Tincanebits and Meriwitica tongues appear to be eastward extensions of the Lyndon Limestone. It is also important to note that the Lyndon Limestone does not become physically attached to the Muav, as suggested by McKee and Resser (Reference McKee and Resser1945), but persists as a distinct interval into eastern Nevada. In central Nevada; however, what would be called the Lyndon Limestone does become part of a larger carbonate deposit (Sundberg, Reference Sundberg2011). The Muav Formation in the western transect correlates in age and lithology with the Highland Peak Formation in the Pioche area and the Papoose Lake Member of the Bonanza King Formation to the west (Palmer and Halley, Reference Palmer and Halley1979; Webster, Reference Webster2011a; Rowland et al., Reference Rowland, Korolev, Hagadorn and Ghosh2023). Precise correlation of these units is unclear due to the paucity of fossils in the Bonanza King Formation (Palmer and Halley, Reference Palmer and Halley1979). The Frenchman Mountain Dolostone compares in lithology and age to most of the Banded Mountain Member of the Bonanza King Formation (Rowland et al., Reference Rowland, Korolev, Hagadorn and Ghosh2023).
The character of Tonto Group rocks and ages changes in the central transect (Fig. 6), with the most prominent change being the elimination of the Lyndon Limestone and Flour Sack members of the Bright Angel Formation and the disappearance of the Rampart Cave to Spencer Canyon members of the Muav Formation. The central transect is very difficult to correlate and would benefit from discovery of additional fossil horizons in these strata.
The eastern transect is not as challenging to correlate, but there are areas of difficulty, especially in the easternmost section. One ongoing difficulty is the correlation of the Muav Formation. In the western transect, the members/lithologies are relatively consistent from one section to the next, but this situation is not true in the central and eastern transects. The Havasu Member, for example, is dolostone in some sections, but limestone in others. This lithologic heterogeneity is entirely possible, but with these shallow-water carbonates, widespread unconformities may explain lithologic changes. In addition, neither McKee (McKee and Resser, Reference McKee and Resser1945) nor Rose (Reference Rose2006, Reference Rose2011) documented lateral changes in these members, but rather documented changes in bioturbation, silt content, and bed thickness. Recognition of depositional packages similar to those recognized by Wanless (Reference Wanless1973) might clarify correlation of units in the Muav.
The supposed tongues of the Bright Angel Formation in the Muav are apparently represented by relatively thin-bedded, silty and/or bioturbated limestones. These lithologies are unlike the lithologies found in the Bright Angel. We recommend avoiding this nomenclature because it is misleading and indicates major changes in lithology within the Muav.
Preservation
Trilobites in the Cambrian of the Grand Canyon are preserved in a variety of lithologies ranging from coarse-grained sandstones to mudstones and carbonate intraformational conglomerates to lime mudstones. The vast majority of specimens are disarticulated sclerites in mudstone or coarser sediment; articulated specimens are rare and mostly found in mudstones (shales).
Mudstones are generally exposed as small fragments and generally have limited exposure in the Grand Canyon due to their relatively quick weathering. As a result, generally smaller fossil fragments are found in these weathered shales. However, trilobites and other fossils have been found, and quarrying has generated abundant specimens. Two classic localities in the Bright Angel Formation, the McKee Quarry and the Cameron-Walcott Quarry, are probably the most famous for their specimens (Foster, Reference Foster2011). Among quarried specimens, 19.7% and 16.7% of the fossils are articulated complete specimens (Fig. 15.2) and 5.2% and 9.3% are articulated thoracic segments, respectively (Foster, Reference Foster2011). In turn, nearly 75% of the specimens are isolated sclerites. These localities and other shale localities producing articulated specimens are listed in Table 3. Of the 50 trilobite species from the Grand Canyon, only 10 species are known from articulated material. Six other Grand Canyon species are also known from articulated material outside of the region (Table 3).
Siliciclastic preservation of trilobites in the Cambrian of the Grand Canyon. All scale bars are 2 mm unless otherwise listed. (1) Side profile of paratype cranidium of Anoria tontoensis (Walcott, Reference Walcott1916b), USNM 793400, preserved in sandstone showing the high convexity. (2) Large articulated Anoria tontoensis, USNM 794042, flattened in shale, arrows indicate crushed areas of the exoskeleton. (3) Pygidium of Albertella nebeli n. sp., GRCA 125398, preserved in bioturbated shales, arrow indicates bending of exoskeleton over a sand-filled burrow during compaction. (4) Tectonically distorted cranidium of Ehmaniella arizonaensis? Resser, Reference Resser1945, n. comb., GRCA 125187, in shale, white arrow illustrates the unusual orientation of the posterior margin. (5) Secondary calcite replacement of exoskeleton of the cranidium of Ehmaniella arizonaensis, n. comb., GRCA 125356, arrow points to broken edge of calcite and underlying internal mold; note the coarseness of the calcite surface, which does not represent the texture of the original exoskeleton. (6) Limonite replacement of the cranidium of Elrathiella sp. in a fine-grained sandstone. (7) Ventral surface of a cranidium of Ehmania weedi Resser, Reference Resser1935, in a calcite-cemented, medium-grained, glauconitic sandstone. (8) Internal mold of Glossopleura sp. in a hematitic, coarse-gained sandstone; the exoskeleton is not preserved in this or the counterpart specimen. (9) Cranidia of olenellids current-orientated perpendicular to ripple marks on the top of a 20-cm-thick sandstone bed. (10) Latex cast of cephalon of Olenellid sp. indet., GRCA 125250b, on the bottom surface of a similar 20-cm-thick sandstone bed.

Figure 15. Long description
A collection of ten fossil photographs.
1. A side profile of a paratype cranidium of Anoria tontoensis in sandstone, showing a highly convex, rounded top with a 5 mm scale bar.
2. A large, flattened Anoria tontoensis in shale with a 10 mm scale bar. Four black arrows point to crushed areas on the head and tail sections of the exoskeleton.
3. A pygidium of Albertella nebeli in bioturbated shale. A black arrow points to a bend in the exoskeleton over a sand-filled burrow. A 2 mm scale bar is at the bottom left.
4. A distorted cranidium of Ehmaniella arizonaensis in shale. A white arrow at the bottom right indicates an unusual orientation of the posterior margin.
5. A cranidium of Ehmaniella arizonaensis replaced by secondary calcite. A black arrow points to a broken edge revealing an internal mold. The surface appears coarse and granular.
6. A reddish-brown limonite replacement of an Elrathiella sp. cranidium embedded in fine-grained sandstone.
7. The ventral surface of an Ehmania weedi cranidium in a speckled, glauconitic sandstone containing green and brown grains.
8. A reddish internal mold of Glossopleura sp. in coarse hematitic sandstone with a 5 mm scale bar.
9. Two olenellid cranidia on a sandstone bed, oriented perpendicular to ripple marks. A 10 mm scale bar and a red transparent ruler are present for scale.
10. A latex cast of an Olenellid sp. cephalon on a sandstone surface with a 5 mm scale bar at the bottom left.
Trilobites found articulated in the Cambrian of the Grand Canyon and region. Taxa from the Grand Canyon and known from articulated material from other localities are also listed

Table 3. Long description
The table consists of five columns: Locality, Formation, Member, Taxa, and Biozone.
* At River Mile 263, Tapeats Sandstone, Transition Zone: Olenelus terminatus? within the Nephrolenellus multinodus biozone.
* At Rampart Cave area, Bright Angel F m., Transition Zone: Albertella nebeli and Albertella schenki within the Mexicella mexicana biozone.
* At McKee Quarry, Bright Angel F m., Transition Zone: Amecephalus althea, Anoria tontoensis, and Glossopleura boccar within the Glossopleura walcotti biozone.
* At Cameron-Walcott Quarry, Bright Angel F m., Transition Zone: Amecephalus althea, Anoria tontoensis, and Glossopleura boccar within the Glossopleura walcotti biozone.
* At Columbine Falls, Bright Angel F m., Flour Sack: Acrocephalops? arizonaensis = Amecephalus piochensis within the Glossopleura walcotti biozone.
* At Diamond Bar Ranch, Bright Angel F m., Flour Sack: Anoria bessus and Glossopleura boccar within the Glossopleura walcotti biozone.
* At Palisades, Bright Angel F m., Flour Sack: Anoria tontoensis within the Glossopleura walcotti biozone.
* At Bright Angel Canyon, Bright Angel F m., Flour Sack: Ehmaniella arizonaensis within the Ehmaniella biozone.
* At Peach Springs Wash, Muav F m., above Rampart Cave: Proehmaniella basilica within the Ehmaniella biozone.
* Other locations in the western U S include Pioche F m. with Biceratops nevadensis and Nephrolenellus multinodus; Lathum Shale with Mesonacis fremonti; Chisholm Shale with Zacanthoides typicalis and Amecephalus piochensis; and Meagher Limestone with Ehmania weedi.
Specimens preserved in mudstones have experienced taphonomic compaction, which can complicate identification. A prime example of this is Anoria tontoensis, which occurs in both sandstones and shales. This species has a relatively convex cranidium (Fig. 15.1) and pygidium when preserved in medium-grained sandstone. However, when preserved in shale, the cranidia and pygidia are flattened, and broken in the frontal portion of the cranidium and the axis and margin of the pygidium (Fig. 15.2). This flattening is not a problem for species known from both lithologic types but does complicate the identification of taxa found only in shale versus those found in limestone or sandstone. Additional problems with mudstone specimens are the occurrence of bioturbation and differential compaction that distorts morphological features (Fig. 15.3), tectonic distortion (Fig. 15.4), and secondary precipitation of calcite on trilobite exoskeletons that obscures the surface ornamentation (Fig. 15.5).
Sclerites are commonly preserved in fine- to coarse-grained sandstones. The lithologies range from micaceous fine-grained sandstones (Fig. 15.6) to medium-grained, calcite cemented, glauconitic sandstones (Fig. 15.7) to coarse-grained, hematitic arkoses (Fig. 15.8). Almost all of these specimens are preserved as limonite (Fig. 15.6), which unfortunately preserves few surface features. This mode of preservation is particularly challenging for the identification of ptychoparioids, where fine details such as surface ornamentation are important to specific identification. The thin-bedded to platy sandstone within the transition zone between the Tapeats Sandstone and Bright Angel Formation yields molds of olenelloid cephala preserved with three-dimensional relief on rippled upper surfaces (Fig. 15.9), but unfortunately they lack sufficient detail to determine their affinity. Molds of olenelloid cephala can be somewhat better preserved on the bottom surfaces of these sandstones (Fig. 15.10).
Fossils from the coarse-grained, hematitic arkoses are heavily fragmented, with most specimens enclosed or coated with a whitish, sandy clay material. This material also occurs as intraclasts in these sandstones (see Fig. 15.8). Trilobite fragments appear to represent Glossopleura sp. and are preserved as steinkerns composed of the same whitish, sandy clay, or “smooth” impressions in the sandstone. The common occurrence of specimens in clasts, coated with the whitish clay, or composed of this clay material (steinkerns) suggests that the fossils have been reworked from lower layers. These lower layers would have needed some time to lithify enough to have transported relatively delicate genal spines.
Trilobites from the Grand Canyon are also preserved in carbonate rocks ranging from micrites, to bioclastic limestone coquinas, to intraformational conglomeratic limestones and dolostones. Whereas most of these fossil-bearing carbonate rocks contain well preserved specimens (Fig. 16.1), specimens are commonly fragmented (Fig. 16.2) and sorted by size or sclerite type (e.g., primarily cranidia, Fig. 16.3) due to current activity. Although many of the carbonate specimens show full relief of the sclerites, some show cracking of sclerites due to compaction (Fig. 16.4). Tectonic distortion is rare in carbonate specimens (Fig. 16.5).
Carbonate preservation of trilobites in the Cambrian of the Grand Canyon. All scale bars are 2 mm. (1) Well-preserved specimen of Ehmaniella kwaguntensis Resser, Reference Resser1945, n. comb., FHPR 19619, in bioclastic limestone showing details of the cranidial ornamentation. (2) Specimen of Amecephalus cf. Am. normale (Resser, Reference Resser1938a), FHPR 19542, in bioclastic limestone showing pre-depositional breakage of the posterior portion of the librigena (arrows). (3, 4) Cranidia of Amecephalus piochensis (Walcott, Reference Walcott1886), illustrating (3) sclerites and size sorting and (4) post-depositional flattening and fracturing (arrows), FHPR 19555 lower and 19556 upper. (5) Tectonically distorted paratype cranidium of Elrathiella? insueta Resser, Reference Resser1945, USNM 794040, in sandy limestone. (6) Partially silicified librigena of Crepicephalus? upis Walcott, Reference Walcott1916a, GRCA 125431. (7) Holotype of “Bolaspis”? aemula Resser, Reference Resser1945, USNM 794055a, in sandy limestone showing the poor preservation of surface features. (8) Paratype of Ehmaniella kwaguntensis n. comb., USNM 794186, preserved as a weathered specimen in sandy limestone lacking the lateral glabellar furrows compared to (1), arrows show the partial preservation of the exoskeleton. (9, 10) Pygidium of Crepicephalus? upis, GRCA 125430, preserved as internal molds on surface of sandy limestone, dorsal and posterior views.

Figure 16. Long description
A multi-panel figure containing 10 photographs of trilobite fossils, each with a 2 mm scale bar.
Panel 1. A well-preserved cranidium of Ehmaniella kwaguntensis showing distinct surface ornamentation and a central glabella.
Panel 2. A cranidium of Amecephalus cf. Am. normale with a white arrow on the bottom-left and a black arrow on the bottom-right pointing to pre-depositional breakage on the librigena.
Panel 3. Two cranidia of Amecephalus piochensis showing size sorting within the limestone matrix.
Panel 4. Two cranidia of Amecephalus piochensis where a white arrow points to a fracture on the upper specimen and black and white arrows point to flattening and fractures on the lower specimen.
Panel 5. A large, tectonically distorted cranidium of Elrathiella? insueta with a stretched appearance.
Panel 6. A curved, partially silicified librigena of Crepicephalus? upis.
Panel 7. A poorly preserved holotype of Bolaspis? aemula with grainy, indistinct surface features.
Panel 8. A weathered paratype of Ehmaniella kwaguntensis where a white arrow at the bottom-right indicates partial preservation of the exoskeleton.
Panel 9. A dorsal view of a Crepicephalus? upis pygidium showing a wide, fan-like structure with segmented ribs.
Panel 10. A posterior view of the same Crepicephalus? upis pygidium showing the vertical profile and curvature of the fossil.
Fossils in sandy limestones have two aspects of preservation that present taxon identification problems. The first is the sand and the second is the surface weathering of the limestone. The sand often adheres to the surface of the exoskeleton (Fig. 16.6, 16.7), obscuring the morphological details needed to determine species identification. Moreover, weathering of these surfaces produces specimens that lack the details necessary for identification (Fig. 16.8, note that the exoskeleton is present in the posterior portion of the cranidium). In some instances, the exoskeletons are completely removed leaving only an internal mold (Fig. 16.9, 10).
Re-evaluation of historic taxonomic assignments is complicated by these specimen preservation constraints. For example, Resser (Reference Resser1945) commonly used type specimens that are preserved in sandy limestones, as well as distorted or weathered specimens, which makes it difficult to fully characterize their morphology. Examples include (using the original names of Resser, Reference Resser1945): Bolaspis aemula (Fig. 16.7); Elrathiella insueta (Fig. 16.5); Glyphaspis kwaguntensis; G. tecta Resser, Reference Resser1945; K. schenki; K. tetraspinosa; Parehmania kwaguntensis (Fig. 16.8); Solenopleurella erosa (cranidia only); and Sol. porcata Resser, Reference Resser1945. Fortunately, additional specimens of K. tetraspinosa, Parehmania kwaguntensis, Sol. erosa, and Sol. porcata were collected during this study, allowing a more complete assessment of Resser’s (Reference Resser1945) species.
Our assignment of new trilobite specimens from the Grand Canyon to species and our re-evaluation of other type and figured specimens is also complicated by specimen preservation issues. For some type specimens, characters typical of a genus or species (e.g., pygidial shape, surface ornamentation) are missing, preventing accurate placement of the type specimens and requiring their placement into nomen dubium (e.g., Elrathiella? insueta Resser, Reference Resser1945). In other cases, sclerites can be identified to a genus, but not a species and may represent multiple species. These specimens are identified as a genus spp. indet. (e.g., Zacanthoides spp. indet.). In one instance, the pygidium of Anoria tontoensis and Glossopleura boccar cannot be differentiated. When similar pygidia are found without the genus- or species-diagnostic features, these specimens are also listed at Glossopleura spp. indet.
In some cases, associated parts are found neither with the type specimens nor do they occur with new specimens, thus requiring the type specimens to be placed into nomen dubium. A prime example is Kootenia, which is generally identified to specific level using pygidial features. We initially included the Grand Canyon species K. mckeei Resser, Reference Resser1945, in K. quadriceps (Hall and Whitfield, Reference Hall and Whitfield1877), based on illustrations of the type specimens by Palmer (Reference Palmer1954, pl. 14, figs. 1–4) and additional specimens by Sundberg (Reference Sundberg1994, fig. 19). The pygidial features were very similar with six posteriorly directed spines including an anterior shorter pair. However, the three collections of illustrated cranidia associated with the pygidia had differently expanding glabellas—suggesting that three different species are represented: K. mckeei, K. quadriceps, and K. cf. K. quadriceps (see discussion under Kootenia).
Material and methods
There are three main periods during which fossils were collected from the Cambrian of the Grand Canyon: (1) an early period in the 1880s, (2) a middle period in the 1930s to 1940s, and (3) a late period in the 2010s and 2020s. There have been some other minor collections made between these periods, but the majority of the Cambrian fossils collected are derived from these three time intervals.
Early period
Collections in the early period were made primarily by C.D. Walcott, with additional material collected by or with N.J. Cameron and L.F. Noble, ranging from 1879 to 1915 (data from McKee and Resser, Reference McKee and Resser1945). Additional collection was made by R.C. Moore in 1924. Fifteen localities (USNM locs. 17c, 38h, 73, 73a–c, 73e, 73f, 74, 74a–e, 75; McKee and Resser, Reference McKee1945, locs. 10, 11, 13–25) from 11 sections were collected during this time period. This material was published by Walcott (Reference Walcott1898, Reference Walcott1901, Reference Walcott1903, Reference Walcott1905, Reference Walcott1912, Reference Walcott1916a, Reference Walcott1916b, Reference Walcott1924, Reference Walcott1925), Ulrich and Bassler (Reference Ulrich and Bassler1931), and Resser (Reference Resser1935, Reference Resser1945) and is presently stored in the Smithsonian Institution collections.
Middle period
Collections in the middle period were made primarily by E.D. McKee with additional specimens collected by or with W. Bryant, L.A. Goebel, A.R. Kerr, C.E. Resser, E.T. Schenk, R.P. Sharp, and R.B. Wheeler during the interval from 1930 to 1942 (Schenk and Wheeler, Reference Schenk and Wheeler1942; McKee and Resser, Reference McKee and Resser1945). From this time, 59 localities (USNM loc. 38i, McKee and Resser [M&R] locs. 1–8, 12, 22, 24, 26–28, 30, 31, 33, 35–69, 70–76; most now assigned to USNM 44338–44346, with several horizons designated with letter suffixes) were collected from 23 sections. This period of collection and that of the early period are the major sources of specimens used by McKee and Resser (Reference McKee and Resser1945). Selected material was sent to Resser by McKee for identification, the vast majority of which was returned to McKee, with only the type material retained at the Smithsonian. The supplemental specimens of McKee are presently stored in the GRCA collections. Some of the collections of McKee and Resser (Reference McKee and Resser1945) are from the Hualapai Tribe Nation (Fig. 1) and belong to the Tribe.
Late period
Collections made for this study date from 2019 to 2024. Material was collected from Frenchman Mountain, Nevada, Diamond Bar Ranch, Arizona, and from Grand Canyon National Park. Collections were primarily by F. Sundberg, J. Foster, J. Hagadorn, and A. Miller, with additional material collected by S. Bastien, H. Cothren, L. Crossey, C. Dehler, T. Farrell, L. Holmer, T. Karim, K. Karlstrom, K. Kirschner, L. McCollum, M. McCollum, M. Mohr, M. Nebel, M. Pulsipher, E. Rose, S. Rowland, and M. Schmitz. During this interval, 134 localities (Utah Field House of Natural History State Park Museum general [FHPR] locs. 006–008, with several horizons designated with number suffixes; GRCA general locs. 275–393, with several horizons also designated with number suffixes) were collected from 25 sections. Type and figured specimens from these localities were deposited in the GRCA collections at the South Rim, Grand Canyon, Arizona; FHPR, Vernal, Utah; Denver Museum of Nature & Science (DMNS), Denver, Colorado; and the University of California, Riverside (UCR), California.
The taxonomy for the olenelloids was done primarily by M. Webster, Anoria spp. and Glossopleura spp. by J. Foster, Albertella spp. by A. Miller, and the remaining trilobites by F. Sundberg.
Material
In several instances, Resser (Reference Resser1945) assigned holotype and paratype(s) to his new species. Designation of the type, if the taxon was represented by more than one specimen on a rock, was done by assigning an “a” suffix to the USNM specimen number. For example, the holotype of Ehmaniella hebes Resser, Reference Resser1945, is 108612a and the paratype that occurs on the same rock is assigned the suffix “b” (108612b). This labeling approach was done for all type specimens occurring on the same rock. Resser (Reference Resser1945) labeled many type specimens with the same number whether they were on the same rock or different rocks. To rectify this error, new USNM type numbers were assigned to the type trilobite specimens from the Cambrian of the Grand Canyon. Some of these specimens provide useful information about the taxon and are illustrated herein. These specimens were labeled using the same system as previously used (e.g., 108619d, e).
Localities
Samples span the Tapeats Sandstone, Bright Angel Formation, Muav Formation, and Frenchman Mountain Dolostone and are from: Frenchman Mountain, Garden Wash, and Devil’s Canyon, Nevada; Diamond Bar Ranch and Grand Wash Cliffs, Arizona; and several sections in Grand Canyon National Park Canyon, Arizona. Permits are required before any fossil collection can be made and as a general policy, publication of the location of specific fossiliferous horizons is not permitted. As a result, coordinates for these locations in the Grand Canyon have been shortened to only two decimal places. Contact the National Park Service before collecting any paleontological materials or if more specific information on the localities is needed. All localities used by Walcott (Reference Walcott1916a, Reference Walcottb, Reference Walcott1924, Reference Walcott1925), Schenk and Wheeler (Reference Schenk and Wheeler1942), McKee and Resser (Reference McKee and Resser1945), and this study are listed with geography, stratigraphy, biostratigraphy, and taxa in the Fossil Locality Supplement (https://doi.org/10.5281/zenodo.14879978). In total, 188 localities were used in this study, of which 128 localities are new sites.
Methods
All specimens were mechanically prepared under magnification. Some shale specimens processed with polyvinyl butyral in the field were treated with acetone during preparation to remove or reduce the amount of polyvinyl butyral on the surface of the specimen. Earlier trials on shale specimens from the Bright Angel Formation illustrated that water-based latex to make specimen casts was absorbed into the shale and destroyed the specimen when the latex was removed. Silicon casts of external molds provided a safer method for some specimens, especially those coated with polyvinyl butyral.
Illustrated specimens have been coated with colloidal graphite and then ammonium chloride sublimate. Specimen orientation for photography and measurements is primarily with the cranidial anterior border and/or palpebral lobes, librigenal border, or pygidial border in a horizontal plane. Some photographs of molds of specimens have been digitally inverted using Photoshop and are labeled in the figure captions as “inverted.” This inversion included changing positive images to negatives and flipping the image horizontally. This “inverted” image is utilized instead of casts where specimens are too fragile to cast in silicone, where the shale matrix breaks apart with water-based latex, or in cases where latex or silicone casting of museum specimens is not permitted. As a general USNM collections policy, type fossil specimens cannot be further prepared, coated, or casted unless prior permission is given. As a result, Resser’s (Reference Resser1945) type and associated specimens are not coated with colloidal graphite, but permission to coat with ammonium chloride sublimate was given. Without the colloidal graphite, the ammonium chloride sublimate needed a heavier coating to eliminate color patterns; but this approach generally reduced the contrast of different morphological features. No preparation of the type specimens was undertaken. Silicon molds were made of Antagmus arizonaensis type specimens as permitted by prior approval by USNM.
Resser’s (Reference Resser1945) photographs of several of the type specimens were trimmed so as to not display the matrix around the sclerites. This photographic trimming gives a misleading impression of what the specimens look like. For example, when the distal end of the posterior lateral area of the fixigena is covered and then trimmed, it gives the cranidium a more rectangular shape. Taxa where both preservation and/or trimmed photographs are misleading include Resser’s species that he assigned to Antagmus Resser, Reference Resser1937, Bolaspis Resser, Reference Resser1935, Elrathiella Poulsen, Reference Poulsen1927, Kochina?, Parehmania Deiss, Reference Deiss1939, Pachyaspis Resser, Reference Resser1939b, and Spencia Resser, Reference Resser1939a. In order to provide a more accurate representation of the specimens, such trimming of the photographs was not done in our work. Areas in the photographs where sand was used to support the specimens were black out using Photoshop.
Repositories and institutional abbreviations
Specimens discussed in this paper are housed in the National Museum of Natural History (NMNH), Smithsonian Institute (USNM), Washington, DC; Utah Field House of Natural History State Park Museum, Vernal, UT (FHPR); United States Geologic Survey (USGS); University of California, Riverside (UCR), CA; Denver Museum of Nature & Science (DMNS), Denver, CO; and the Grand Canyon National Park Museum Collections (GRCA), South Rim, AZ. See Table 2 for generic abbreviations.
The listing of localities is complex because of the multiple procedures that were used by different institutions and individuals to catalog the specimens. In general, specimens collected and discussed by Walcott were listed with USNM locality numbers, which consist of an isolated number (e.g., USNM loc. 74) or a number with a lower-case letter suffix (e.g., USNM loc. 74a). Collections by McKee andResser (Reference McKee and Resser1945) have only Walcott’s old locality numbers with one additional (USNM loc. 38i). McKee and Resser (Reference McKee and Resser1945) assigned numbers to each fossil locality (1–76, individual numbers are here referred to as M&R loc.), including the older Walcott localities. Resser did not assign USNM locality numbers to the type material housed at the USNM and the type collection has no notation as to which M&R locality they were derived from, although this information for the holotypes mostly can be retrieved from McKee and Resser (Reference McKee and Resser1945). To remedy this problem, new USNM locality numbers were assigned (44335–44346) for each general location with an upper-case letter suffix (e.g., 44545J) to indicate different horizons within McKee and Resser’s sections. The GRCA localities are defined on polygons of different shapes and sizes representing polygons of a fossiliferous area. The numbering has been shortened (originally, e.g., PALGRCA000123 to GRCA loc. 123) and does not separate different stratigraphic horizons. The locality numbers have been modified to reflect collections from different horizons by adding numeric suffixes (e.g., GRCA loc. 123-1, 123-2). In addition, GRCA does not assign locality numbers that are outside the park (e.g., Native American Lands and Diamond Bar Ranch). Some McKee and Resser (Reference McKee and Resser1945) localities fall outside of the National Park; these localities are designated by the original numbering system (e.g., M&R loc. 2). The UCR locality numbers are simple sequential numbers (e.g., 10027), although specimen numbers are represented by the locality number with a numbered suffix (e.g., 10027-1). The FHPR locality numbers are based on fossiliferous area (e.g., Diamond Bar Ranch) and the containing formation (Bright Angel Formation) in a simple number (e.g., 008). Specific horizons withing this area are denoted by a numbered suffix (e.g., 008-8).
All of the different locality formats and numbers are given in the Fossil Locality Supplement (https://doi.org/10.5281/zenodo.14879978). In all cases, if there is a USNM locality number present, it is taken as the primary locality in the paper (e.g., USNM loc. 44338D is equivalent to M&R loc. 35 and GRCA loc. 368-3).
Systematic paleontology
Order Redlichiida Richter, Reference Richter, Dittler, Joos, Korschelt, Linek, Oltmanns and Schaum1932
Suborder Olenellina Walcott, Reference Walcott1890
Superfamily Olenelloidea Walcott, Reference Walcott1890
Family Biceratopsidae Pack and Gayle, Reference Pack and Gayle1971
Genus Biceratops Pack and Gayle, Reference Pack and Gayle1971
Type species
Biceratops nevadensis Pack and Gayle, Reference Pack and Gayle1971, from the upper Dyeran (Cambrian Stage 4) of the western United States (monotypic).
Biceratops nevadensis Pack and Gayle, Reference Pack and Gayle1971
Biceratopsids from the Transition Zone, Bright Angel Formation, Grand Canyon Region, Arizona and Nevada. (1–5) Biceratops nevadensis Pack and Gayle, Reference Pack and Gayle1971: (1) latex cast of cephalon USNM 794086 (Resser, pl. 19, fig. 25) from USNM loc. 44343A; (2) latex cast of cephalon USNM 794085 (Resser, pl. 19, fig. 26) from USNM loc. 44343A; (3) shale cephalon URC 10031-12 from UCR locality 10031; (4) shale cranidium FHPR 19557 from FHPR loc. 006-7; (5) shale cephalon URC 10029-9 from UCR locality 10029. (6) Biceratopsid sp. indet. sandstone cephalon GRCA 125219 from GRCA loc. 382. (7) Nephrolenellus cf. N. multinodus (Palmer in Palmer and Halley, Reference Palmer and Halley1979), partial cephalon GRCA 125386 from GRCA loc. 388. (8–10) Nephrolenellus sp. indet.: (8) sandstone cephalon UCR 10031-22 from locality UCR 1003; (9) hypotype cephalon 794082 (Resser, Reference Resser1945, pl. 19, fig. 23) from USNM loc. 44345A, inverted photograph; (10) plastic cast of cephalon 794084 from USNM loc. 44345A.

Figure 17. Long description
A multi-panel photograph displaying ten fossil specimens labeled 1 through 10. A black scale bar labeled 4 m m is positioned in the center-right of the layout.
* Panel 1: Top left. A latex cast of a Biceratops nevadensis cephalon (U S N M 794086) showing a smooth, rounded outer margin and a raised central glabella.
* Panel 2: Top right. A latex cast of a Biceratops nevadensis cephalon (U S N M 794085) with visible eye ridges and a textured surface.
* Panel 3: Center. A large shale cephalon of Biceratops nevadensis (U R C 10031-12) showing a wide, semi-circular shape with a distinct central lobe.
* Panel 4: Middle left. A shale cranidium of Biceratops nevadensis (F H P R 19557) with a prominent, bulbous glabella and well-defined lateral furrows.
* Panel 5: Middle center. A small shale cephalon of Biceratops nevadensis (U R C 10029-9) showing symmetrical features.
* Panel 6: Middle right. A sandstone cephalon of Biceratopsid species indeterminate (G R C A 125219) with a grainy texture.
* Panel 7: Lower left. A partial cephalon of Nephrolenellus cf. N. multinodus (G R C A 125386) showing a curved outer rim and fragmented internal structures.
* Panel 8: Lower right, above panel 10. A sandstone cephalon of Nephrolenellus species indeterminate (U C R 10031-22) with a weathered appearance.
* Panel 9: Bottom left. An inverted photograph of a hypotype cephalon of Nephrolenellus species indeterminate (794082) showing detailed glabellar lobes.
* Panel 10: Bottom right. A plastic cast of a Nephrolenellus species indeterminate cephalon (794084) showing the most complete anatomical structure with wide lateral lobes and a central axis.
Reference McKee and Resser1945 Olenellus sp. undet.; Resser in McKee and Resser (part), p. 193, pl. 19, figs. 25, 26 only.
Reference Pack and Gayle1971 Biceratops nevadensis Pack and Gayle, p. 896–898, text-figs. 1, 2, pl. 102, figs. 1–5.
Reference Whittington1989 Biceratops nevadensis; Whittington, p. 131, 132, 139, fig. 11.
Reference Whittington1992 Biceratops nevadensis; Whittington, p. 124, pl. 58.
Reference Palmer and Repina1993 Biceratops nevadensis; Palmer and Repina, p. 23, fig. 4.1.
Reference Whittington, Chatterton, Speyer, Fortey and Owens1997 Biceratops nevadensis; Whittington in Whittington et al., fig. 44.
Reference Whittington, Chatterton, Speyer, Fortey and Owens1997 Biceratops nevadensis; Palmer and Repina in Whittington et al., p. 409, fig. 257.1.
Reference Webster2011b Biceratops nevadensis; Webster, p. 197, 201, fig. 4.1.
Reference Sundberg2020 Biceratops nevadensis; Sundberg et al., fig. 3a.
Holotype
USNM 168225, from the Bright Angel Formation, Frenchman Mountain, Las Vegas, Nevada.
Occurrence
Uppermost Bolbolenellus euryparia Biozone, Transition Zone, Tapeats Sandstone, Frenchman Mountain, Nevada (Webster, Reference Webster2011a). Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, Garden Wash, Indian Hills, Nevada; lowermost Bright Angel Formation, Diamond Bar Ranch, Grand Wash Cliffs; Transition Zone, Tapeats Sandstone, Quartermaster Canyon, Grand Canyon, Arizona (McKee and Resser, Reference McKee and Resser1945).
Remarks
Biceratops nevadensis is a distinctive biceratopsid characterized by its effaced cephalic features and the absence of genal spines. The species has previously only been reported from Frenchman Mountain (Pack and Gayle, Reference Pack and Gayle1971; Webster, Reference Webster2011b), although Resser (in McKee and Resser, Reference McKee and Resser1945, pl. 19, figs. 25, 26) figured two specimens as “Olenellus sp. undet.” from Quartermaster Canyon that represent Bi. nevadensis (Fig. 17.1, 17.2). The occurrences of Bi. nevadensis from the Indian Hills (Garden Wash), the Grand Wash Cliffs (Diamond Bar Ranch), and the western Grand Canyon (Quartermaster Canyon) documented herein expand the known geographic range of the species in a cratonward direction.
High-resolution stratigraphic sampling at Frenchman Mountain (Webster, Reference Webster2011b) found that the top of the local stratigraphic range of Bi. nevadensis was less than one meter below the local lowest occurrence of Nephrolenellus multinodus (Palmer in Palmer and Halley, Reference Palmer and Halley1979); those Biceratops-bearing strata are therefore assigned to the uppermost Bolbolenellus euryparia Zone (see also Webster, Reference Webster2011a). However, specimens of Nephrolenellus occur within the Biceratops-bearing collections at Diamond Bar Ranch (UCR 10031) and Quartermaster Canyon, suggesting that Bi. nevadensis locally ranges upwards into the Nephrolenellus multinodus Biozone (or that the Arizona specimens represent a previously unrecognized occurrence of a species of Nephrolenellus below the Nephrolenellus multinodus Biozone). However, collection UCR 10031 was pooled over a stratigraphic interval of greater than a meter, so it is conceivable that the co-occurrence of Biceratops and Nephrolenellus within that collection is an artifact of this coarser sampling resolution.
Despite intensive sampling of correlative strata in the depositionally oceanward Cadiz, Pioche, and Carrara formations of California and Nevada, Bi. nevadensis has not been recovered from those units. The species was apparently restricted to nearer-shore environments.
Genus Nephrolenellus Palmer and Repina, Reference Palmer and Repina1993
Type species
Olenellus multinodus Palmer in Palmer and Halley, Reference Palmer and Halley1979, from the upper Dyeran (Cambrian Stage 4) of the western United States and Canada.
Remarks
The two species within this genus (N. multinodus and N. geniculatus Palmer, Reference Palmer1998) are differentiated by relatively subtle cephalic features (see Webster, Reference Webster2007), and unambiguous species-level identification requires well-preserved material.
Nephrolenellus cf. N. multinodus (Palmer in Palmer and Halley, Reference Palmer and Halley1979)
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, River Mile 278, near Rampart Cave, Grand Canyon, Arizona.
Remarks
The posterior portion of the glabella is not preserved on a cephalon from River Mile 278, but the specimen exhibits a short preglabellar field and rounded adgenal angle and is identified as Nephrolenellus cf. N. multinodus.
Nephrolenellus sp. indet.
Reference McKee and Resser1945 Olenellus sp. undet.; Resser in McKee and Resser (part), p. 193, pl. 19, fig. 23 only.
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone; Rampart Cave (McKee and Resser, Reference McKee and Resser1945), Quartermaster Canyon (McKee and Resser, Reference McKee and Resser1945), Grand Canyon, Arizona.
Remarks
Several cephala are too poorly preserved for species-level assignment and are treated herein as an indeterminate species of Nephrolenellus. Resser in McKee and Resser (Reference McKee and Resser1945, pl. 19, fig. 23) figured one such specimen, but identified it as “Olenellus sp. undet.” Despite their low quality, the specimens are significant in that they constrain the fossiliferous strata to lie within the N. multinodus Zone.
Genus uncertain
Biceratopsid sp. indet.
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, Diamond Bar Ranch, Grand Wash Cliffs and Rampart Cave, River Mile 263 and Granite Park, Grand Canyon, Arizona.
Remarks
An internal mold of a cephalon preserved in a sandstone might represent either Nephrolenellus or Arcuolenellus Palmer and Repina, Reference Palmer and Repina1993. Also included are several poorly preserved specimens with similar cephalic outlines.
Family Olenellidae Walcott, Reference Walcott1890
Genus Olenellus Hall, Reference Hall1862
Type species
Olenus thompsoni Hall, Reference Hall1859, from the upper Dyeran (Cambrian Stage 4) of the Parker Formation of Vermont.
Olenellus gilberti Meek in White, Reference White1874
Olenellus spp. from the Transition Zone, Bright Angel Formation, Arizona and Nevada. Specimens (7, 11, 13, 14) preserved in sandstone, all others preserved in shale. (1) Olenellus gilberti Meek in White, Reference White1874, cephalon UCR 10027-4 from UCR loc. 10027, Devil’s Cove, Nevada. (2) Olenellus n. sp., cephalon UCR 10027-5 from UCR loc. 10027, Devil’s Cove, Nevada. (3–8) Olenellus terminatus Palmer, Reference Palmer1998: (3) cephalon UCR 10029-24 from UCR loc. 10029, Garden Wash, Nevada; (4) cephalon UCR 10029-4 from UCR loc. 10029, Garden Wash, Nevada; (5) cephalon UCR 10031-14 from UCR loc. 10031, Diamond Bar Ranch, Arizona; (6) cephalon UCR 10027-2 from UCR loc. 10027, Devil’s Cove, Nevada; (7) latex cast of cephalon GRCA 125556b from GRCA loc. 330-1, Mile 278, Arizona; (8) cephalon UCR 10029-13 from UCR loc. 10029, Garden Wash, Nevada. (9, 10) Olenellus cf. O. fowleri Palmer, Reference Palmer1998; (9) inverted photo of partial cephalon UCR 10029-6 from UCR loc. 10029, Garden Wash, Nevada; (10) partial cephalon UCR 10029-30 from UCR loc. 10029, Garden Wash, Nevada. (11) Olenellus? sp. A, latex cast of cephalon GRCA 125251 from GRCA loc. 388, near Rampart Cave, Arizona. (12–14) Olenellus sp. indet.: (12) Olenellus sp. indet. USNM 794083 of Resser (Reference Resser1945, pl. 19, fig. 24) from USNM loc. 44345A, near Rampart Cave, Arizona; (13) cephalon UCR 10030-1 from UCR loc. 10030, Garden Wash, Nevada; (14) latex cast of cephalon 125556a from GRCA loc. 330-1.

Figure 18. Long description
The plate contains 14 individual panels of fossilized trilobite heads, or cephala, numbered in the top-right or bottom-right corners of each panel.
* Panels 1 and 2: Semicircular cephala with distinct central glabellae and curved eye ridges.
* Panels 3 and 4: Similar to the first two, with panel 3 showing a vertical crack through the right side of the fossil.
* Panel 5: A high-relief, three-dimensional cephalon showing a bulbous glabella and deep furrows.
* Panel 6: A smaller, flatter specimen with a well-defined border.
* Panel 7: A larger, detailed view of a cephalon with a textured surface, preserved in sandstone.
* Panel 8: A wide, semicircular cephalon with symmetrical eye lobes.
* Panels 9 and 10: Partial specimens; panel 9 shows the right half of a cephalon with dark mineral staining, and panel 10 shows a large right pleural area.
* Panel 11: A weathered, light-colored latex cast of a cephalon.
* Panel 12: A small, central glabella specimen with a 4 mm scale bar located directly beneath it.
* Panel 13: A cephalon preserved in coarse sandstone with a rounded glabella.
* Panel 14: A latex cast showing the ventral or internal features of a cephalon with segmented structures.
Reference Webster2015 Olenellus gilberti; Webster, p. 11–68, figs. 6–15, 19–21, 25–29, 31–33, 36 [synonymy to date].
Reference Lerosey-Aubril and Peel2018 Olenellus gilberti; Lerosey-Aubril and Peel, p. 755.
Reference Pates and Bicknell2019 Olenellus gilberti; Pates and Bicknell, p. 297, 299, 301, 304, figs. 2, 3a, c, 4a, c.
Reference Sundberg and Webster2021 Olenellus gilberti; Sundberg and Webster, p. 1244, fig. 3.4.
not Reference Handkamer, Pratt and MacNaughton2022 Olenellus gilberti; Handkamer et al., p. 10, fig. 8.1–8.4, 8.6, 8.10 [= Mesonacis? sp.].
Lectotype
USNM 15411a, from USNM Locality 31a, the Combined Metals Member of the Pioche Formation, Pioche Hills, Nevada (see Webster, Reference Webster2015).
Occurrence
Nephrolenellus multinodus Biozone, lowermost Bright Angel Formation, Devil’s Cove, Indian Hills, Nevada.
Remarks
This cephalon conforms well to Olenellus gilberti, as recently described by Webster (Reference Webster2015), and represents a cratonward extension of the known range of the species. The Canadian specimens identified as O. gilberti by Handkamer et al. (Reference Handkamer, Pratt and MacNaughton2022) do not seem to represent that species. Most of the figured Canadian specimens have a relatively narrower cephalic border and a shorter, less robust genal spine that suggest closer affinity to taxa such as Mesonacis vermontanus (Hall, Reference Hall1859) (compare in particular Handkamer et al., Reference Handkamer, Pratt and MacNaughton2022, fig. 8.1 to Webster and Landing, Reference Webster and Landing2016, fig. 5C or Pari et al., Reference Pari, Briggs, Gaines, Roach and Webster2023, fig. 5a). Two of the figured Canadian specimens (Handkamer et al., Reference Handkamer, Pratt and MacNaughton2022, fig. 8.3, 8.4) have genal spines that appear to be more strongly anteriorly advanced than is typical for O. gilberti. More than one species might be represented by the Canadian material, but all specimens are herein provisionally reassigned to the genus Mesonacis Walcott, Reference Walcott1885 (see Webster and Landing, Reference Webster and Landing2016, and Webster and Caron, Reference Webster and Caron2024, for discussions of the problematic distinction between these genera).
Olenellus terminatus Palmer, Reference Palmer1998
Reference Palmer1998 Olenellus (Paedeumias) terminatus; Palmer, p. 670–671, figs. 5.2, 12.1–12.8.
Reference Webster2011b Olenellus terminatus; Webster, fig. 4.2.
Reference Webster2011c Olenellus terminatus; Webster, fig. 7.3.
Reference Pates and Bicknell2019 Olenellus terminatus; Pates and Bicknell, p. 297, 299, 301–304, figs. 2, 5b, c.
Reference Sundberg, Karlstrom, Geyer, Foster, Hagadorn, Mohr, Schmitz, Dehler and Crossey2020 Olenellus terminatus; Sundberg et al., fig. 3b.
?Reference Handkamer, Pratt and MacNaughton2022 Olenellus terminatus; Handkamer et al., p. 10, fig. 8.5, 8.7, 8.9.
Holotype
DMNS EPI 16085, from DMNS loc. 1196, Ruin Wash Lagerstätte, Combined Metals Member, Pioche Formation, Chief Range, Nevada.
Occurrence
Nephrolenellus multinodus Biozone, lowermost Bright Angel Formation, Devil’s Cove, Garden Wash, Indian Hills, Nevada; Transition Zone, Tapeats Sandstone and lowermost Bright Angel Formation, Diamond Bar Ranch, Grand Wash Cliffs, Arizona; Transition Zone, Tapeats Sandstone, River Mile 278 near Rampart Cave, Grand Canyon, Arizona.
Remarks
The cephalon of O. terminatus is most obviously characterized by development of only a weak adgenal angle, a proportionally narrow cephalic border that is less than half the length of the preglabellar field, Bertillon markings on the extraocular area, and moderately long ocular lobes with posterior tips transversely opposite the anterior portion of the lateral margins of LO or SO.
Olenellus terminatus is common in the Indian Hills and at Diamond Bar Ranch where—as at Frenchman Mountain (Webster, Reference Webster2011b)—it stratigraphically co-occurs with Bi. nevadensis. A single cephalon showing the distinctive Bertillon markings from Rampart Cave (Figure 18.7) represents the most cratonward definite occurrence of this species. The presence of Bertillon markings cannot be determined on the published images of Canadian specimens assigned to O. terminatus by Handkamer et al. (Reference Handkamer, Pratt and MacNaughton2022), rendering that identification ambiguous.
Olenellus cf. O. fowleri Palmer, Reference Palmer1998
cf. Reference Palmer1998 Olenellus (Olenellus?) fowleri Palmer, Reference Palmer1998, p. 669–670, figs. 5.5, 10.1–10.6.
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, Garden Wash, Indian Hills, Nevada.
Remarks
Two incomplete cephala recovered from Garden Wash show deep glabellar furrows, pit-like impressions of S2, long ocular lobes (with posterior tips transversely opposite the lateral margins of LO), and proportionally wide (tr.) extraocular areas. Such a combination of traits characterizes O. fowleri Palmer, Reference Palmer1998, although neither cephalon is sufficiently complete for unambiguous species assignment.
The cephala co-occur in a collection with Biceratops nevadensis. At Frenchman Mountain O. fowleri occurs less than a meter above the local highest occurrence of Bi. nevadensis (Webster, Reference Webster2011b). The co-occurrence of these species at Garden Wash may represent a previously unrecognized overlap in their stratigraphic ranges or could be an artifact of the coarser stratigraphic resolution (pooled from a stratigraphic interval spanning more than a meter) of the Garden Wash collection (see also remarks for Bi. nevadensis).
Olenellus n. sp.
Occurrence
Nephrolenellus multinodus Biozone, lowermost Bright Angel Formation, Devil’s Cove, Indian Hills, Nevada.
Remarks
A single incomplete cephalon from Devil’s Cove seems to represent a novel morphotype which, pending discovery of additional material, is not formally named herein. It is characterized by long ocular lobes with posterior tips transversely opposite the posterior portion of the lateral margins of LO; a short preglabellar field only slightly longer (sag.) than the length of the anterior cephalic border; a relatively strong adgenal angle located approximately three-fifths of the distance along the posterior cephalic margin between the axial furrow and the base of the genal spine, with the base of the genal spine transversely opposite the anteriormost portion of the lateral margin of LO; and a relatively narrow extraocular area. There is a suggestion of fine Bertillon markings on LA. The posterior cephalic margin is slightly worn, but an intergenal spine appears to be absent.
The specimen is similar to O. gilberti and to the poorly known O. buttsi Resser, Reference Resser1938a, from Alabama, but possesses longer ocular lobes than both those species. The long ocular lobes and short preglabellar field are reminiscent of O. puertoblancoensis (Lochman in Cooper et al., Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952), O. agellus Resser and Howell, Reference Resser and Howell1938, O. transitans (Walcott, Reference Walcott1910), and O. aff. O. transitans (Webster and Landing, Reference Webster and Landing2016) but those species have a weaker adgenal angle and/or possess intergenal spines. “Olenellus” macrophthalmus (Poulsen, Reference Poulsen1958) is known only from two fragmentary cephala collected from Marshall Bugt, Inglefield Land, northwestern Greenland (Poulsen, Reference Poulsen1958, pl. 3, figs. 2–4). The ocular lobes of those cephala are similar in length but not in orientation to those of the specimen from Devil’s Cove: the interocular area of the Greenland species is much wider (tr.).
Olenellus? sp. A
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, River Mile 278 near Rampart Cave, Grand Canyon, Arizona.
Remarks
The relatively long preglabellar field of this cephalon suggests that this specimen represents a distinct morphotype from those documented herein, but the preservation is too poor for confident assignment at even the generic level. The general form of the cephalon is reminiscent of O. nevadensis (Walcott, Reference Walcott1910) or O. chiefensis Palmer, Reference Palmer1998.
Olenellus sp. or spp. indet.
Reference McKee and Resser1945 Olenellus sp. undet.; Resser in McKee and Resser, p. 193, pl. 19, fig. 24 only.
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, Garden Wash, Indian Hills, Nevada; Transition Zone, Tapeats Sandstone, Rampart Cave and Quartermaster Canyon (McKee and Resser, Reference McKee and Resser1945), Grand Canyon, Arizona.
Remarks
These specimens are too poorly preserved for species-level assignment, but all are consistent with either O. terminatus or O. gilberti and therefore they do not necessarily increase the diversity of olenellids in the Grand Canyon region. They are figured herein for completeness. One of the specimens (Fig. 18.12) was previously figured by Resser (in McKee and Resser, Reference McKee and Resser1945, pl. 19, fig. 24).
Genus Mesonacis Walcott, Reference Walcott1885
Type species
Olenus vermontanus Hall, Reference Hall1859, from the upper Dyeran (Cambrian Stage 4) of the Parker Formation of Vermont.
Mesonacis fremonti (Walcott, Reference Walcott1910)
Mesonacis spp. from the Nephrolenellus multinodus Biozone, Transition zone and lower Bright Angel Formation, Grand Canyon Region, in Arizona unless otherwise noted. Specimens 2–5 are in shale; others are in sandstone. (1–3) Mesonacis fremonti (Walcott, Reference Walcott1910): (1) latex cast of cephalon GRCA 125555 from GRCA loc. 330-1 near Rampart Cave; (2) cranidium FHPR 19606 from FHPR 006-7, Diamond Bar Ranch; (3) cranidium UCR 10029-27 from UCR loc. 10029, Garden Wash, Nevada. (4–8) Mesonacis cf. M. fremonti (Walcott, Reference Walcott1910): (4), hypotype cranidium USNM 794081 reported as Olenellus cf. fremonti by Resser (Reference Resser1945, pl. 19, fig. 21) from USNM loc. 44341, Peach Springs Canyon; (5) hypotype cranidium USNM 794080 reported as Olenellus cf. fremonti by Resser (Reference Resser1945, pl. 19, fig. 22) from USNM loc. 44341, Peach Springs Canyon; (6) cephalon UCR 18031-25 from UCR loc. 18031, Diamond Bar Ranch; (7) latex mold of cephalon GRCA 125250 from GRCA loc. 388 near Rampart Cave; (8) partial cephalon GRCA 125217 from GRCA loc. 382, near Rampart Cave. Scales: (1–3) = 10-mm scale bar; (4, 5, 7, 8) = 5-mm scale bar; (6) = 10-mm scale bar.

Figure 19. Long description
A collection of eight fossil specimens arranged in a grid-like fashion.
* Panel 1: A latex cast of a cephalon showing a central glabella with distinct lobes and wide lateral borders.
* Panel 2: A large, wide cranidium with prominent crescent-shaped ocular lobes. A 10 mm scale bar is positioned below it.
* Panel 3: A wide cranidium similar to panel 2, showing a vertical crack through the center of the glabella.
* Panel 4: A hypotype cranidium with a more weathered, granular texture.
* Panel 5: A smaller hypotype cranidium showing the central glabella and partial right side.
* Panel 6: A large cephalon preserved in a dark, cracked matrix. A 10 mm scale bar is located in the top right corner of this panel.
* Panel 7: A small latex mold of a cephalon with a 5 mm scale bar positioned above it.
* Panel 8: A partial cephalon showing the right lateral side and posterior border.
All specimens exhibit the characteristic semi-circular head shield of the trilobite genus Mesonacis, with varying degrees of preservation in shale and sandstone matrices.
Reference Walcott1910 Olenellus fremonti Walcott (part), p. 320–323, pl. 37, fig. 2 only.
Reference Sundberg1999 Mesonacis fremonti; Lieberman, p. 41–43, figs. 8.1, 8.2, 9.1 [synonymy to date].
Lectotype
USNM 56819a, from the “Pioche Formation” at USNM Locality 52, Prospect Mountain, Eureka district, Nevada, designated by Harrington (Reference Harrington1956).
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, Garden Wash, Indian Hills, Nevada; Transition Zone, Tapeats Sandstone and/or lowermost Bright Angel Formation, Diamond Bar Ranch, Grand Wash Cliffs, Rampart Cave, Grand Canyon, Arizona.
Remarks
Mesonacis fremonti is characterized by a relatively broad glabella that is only slightly constricted (tr.) at S1; relatively short ocular lobes with posterior tips transversely opposite the middle of the lateral margin of L1 or S1; a very short or no preglabellar field; a relatively strong adgenal angle, with the base of the genal spine transversely opposite the middle or anterior half of the lateral margin of LO, SO, or L1; a genal spine that is as broad at its base as the adjacent lateral cephalic border; and a dorsally arched interocular area, with an intergenal ridge extending outwards from opposite L1 to the posterior cephalic border distal to the adgenal angle. The species closely resembles O. gilberti but has proportionally shorter ocular lobes and a proportionally shorter (or no) preglabellar field. A phylogenetic analysis of various species of Olenellus and Mesonacis will help resolve whether M. fremonti represents a basal member of Mesonacis (as recovered in the phylogenetic analysis by Lieberman, Reference Lieberman1999, although that analysis did not include O. gilberti), or should be reassigned back into Olenellus (e.g., Walcott, Reference Walcott1910; Palmer in Palmer and Halley, Reference Palmer and Halley1979), or represents a distinct genus (e.g., Fremontia Raw, Reference Raw1936, as proposed by Raw, Reference Raw1936, and Harrington, Reference Harrington1956).
Resser (in McKee and Resser, Reference McKee and Resser1945, p. 193, pl. 19, figs. 21, 22) identified some incomplete cephala from the Grand Canyon as “Olenellus cf. fremonti”. The better-preserved specimens documented herein represent the first unambiguous occurrence of the species in the Grand Canyon.
Mesonacis cf. M. fremonti (Walcott, Reference Walcott1910)
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, Diamond Bar Ranch, Grand Wash Cliffs; Transition Zone, Tapeats Sandstone, River Mile 263, Rampart Cave, Granite Park and, Peach Springs Canyon (McKee and Resser, Reference McKee and Resser1945), Grand Canyon, Arizona.
Remarks
The preserved portions of these specimens are consistent with M. fremonti, but the cephala are too poorly preserved for confident assignment. The two specimens from Peach Springs Canyon were previously figured as “Olenellus cf. fremonti” by Resser (in McKee and Resser, Reference McKee and Resser1945, p. 193, pl. 19, figs. 21, 22).
Genus Uncertain
Olenellid sp. indet.
Olenelloids indet. from the Nephrolenellus multinodus Biozone, Transition Zone, and lower Bright Angel Formation, Grand Canyon Region, in Arizona, unless otherwise noted. Specimen D is in shale, others are in sandstone. (1, 2) Olenellid sp. indet., latex casts: (1) cephalon GRCA 125250b from GRCA loc. 388; (2) cephalon GRCA 125216 from GRCA loc. 382. (3–9) Olenelloid sp. indet. from GRCA loc. 382, unless otherwise noted: (3) cephalon GRCA 125220b; (4) cephalon fragment UCR 10031-33 from UCR locality 10031, Diamond Bar Ranch; (5) cephalon GRCA 125249 from GRCA loc. 388; (6) partial cephalon USNM 125220a; (7) cephalon GRCA 125218; (8) partial cephalon GRCA 125222; (9) partial cephalon GRCA 125221. Scales: (1, 2, 5) = 10-mm scale bar; (3, 4, 6–9) = 4-mm scale bar.

Figure 20. Long description
A multi-panel collection of nine grayscale photographs showing fossilized trilobite remains. Two horizontal scale bars are positioned in the upper center, one labeled 10 mm and a shorter one labeled 4 mm.
* Panel 1 at the top left shows a wide view of a cephalon G R C A 125250b with a visible glabella and genal spines embedded in a coarse matrix.
* Panel 2 at the top right displays cephalon G R C A 125216, featuring a prominent, raised central glabella and a smooth outer border.
* Panel 3 in the middle left shows a partial cephalon G R C A 125220b with a curved lateral border.
* Panel 4 in the center shows a cephalon fragment U C R 10031-33 with a distinct crack running vertically through the fossil.
* Panel 5 in the middle right is a close-up of cephalon G R C A 125249, showing granular surface texture and a segmented glabella.
* Panel 6 at the bottom left shows a partial cephalon U S N M 125220a with a sharp genal spine pointing downward.
* Panel 7 at the bottom center-left shows a small, weathered cephalon G R C A 125218.
* Panel 8 at the bottom center-right shows a partial cephalon G R C A 125222 with visible marginal ribbing.
* Panel 9 at the bottom right shows a partial cephalon G R C A 125221 with a well-defined ocular ridge.
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, River Mile 276 near Rampart Cave, Grand Canyon, Arizona.
Remarks
These cephala probably represent M. fremonti but are too poorly preserved and incomplete for confident assignment to even the generic level.
Olenelloid sp. indet.
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone and lowermost Bright Angel Formation, Diamond Bar Ranch, Grand Wash Cliffs; Transition Zone, Tapeats Sandstone River Mile 263, River Mile 278, River Mile 275 near Rampart Cave, Meriwitica Canyon, Bridge Canyon, Granite Park, Diamond Creek, and River Mile 212, Grand Canyon (McKee and Resser, Reference McKee1945), Arizona.
Remarks
These fragmentary, very poorly preserved cephala cannot be confidently assigned to genus or species. However, the preserved portion of each specimen is consistent with one or more of the species documented above. Therefore, these specimens document the existence of olenelloids in their respective collections, but they do not necessarily increase the total diversity of olenelloids documented from the Grand Canyon region. Included in this taxon is the Olenellus sp. assigned, but not illustrated, by McKee and Resser (Reference McKee1945).
Order Corynexochida Kobayashi, Reference Kobayashi1935
Family Dolichometopidae Walcott, Reference Walcott1916b
Genus Anoria Walcott, Reference Walcott1924
Type species
Dolichometopus tontoensis (Walcott, Reference Walcott1916b), from the upper Delamaran (Wuliuan Stage) Bright Angel Formation, Bright Angel Trail, Grand Canyon, Arizona.
Remarks
Separation of the genera Anoria, Sonoraspis Stoyanow in Cooper et al., Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952, and Glossopleura Poulsen, Reference Poulsen1927, is difficult if the entire exoskeleton is not preserved (Foster, Reference Foster2022). Lochman (in Cooper et al., Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952), Palmer (in Palmer and Halley, Reference Palmer and Halley1979), and Foster (Reference Foster2022) synonymized Sonoraspis and Glossopleura, which is accepted here. The type species of Anoria can be separated from Glossopleura only in its shorter palpebral lobe and corresponding longer posterior area of the fixigena. The original characters of seven thoracic segments and the fifth axial segment with an extended pleural spine are variable within dolichometopids (Foster, Reference Foster2022). The measure of palpebral lobe length relative to glabellar length (Fig. 21) shows significant overlap among most species referred to Anoria, Sonoraspis, and Glossopleura, suggesting that these species should be under Glossopleura, except Anoria tontoensis (Foster, Reference Foster2022). The two taxa A. tontoensis and G. boccar Poulsen, Reference Poulsen1927 (= G. mckeei) are both found in the Bright Angel Formation and can be separated based on the relative length of the palpebral lobe (Fig. 21) and corresponding length of the posterior area of the fixigena. Anoria tontoensis has palpebral lobes less than 38% of the glabellar length whereas G. boccar has lengths greater than 45%.
Bivariate plot of the relative palpebral lobe lengths versus cranidial length of representatives of Anoria (red): A. baton (Walcott, Reference Walcott1916b), A.? bessus (Walcott, Reference Walcott1916b), A. tontoensis (Walcott, Reference Walcott1916b); Glossopleura (black/gray): G. boccar (Walcott, Reference Walcott1916b); G. campbelli Robison and Babcock, Reference Robison and Babcock2011; G. gigantea Robison, Babcock, and Gunther, Reference Robison, Babcock and Gunther2015; G. lodensis (Clark, Reference Clark1921); G. mohavensis Resser, Reference Resser1935; G. producta (Hall and Whitfield, Reference Hall and Whitfield1877); G. similaris Resser, Reference Resser1939a; G. stephenensis Resser, Reference Resser1935; G. utahensis Resser, Reference Resser1935; G. walcotti (Poulsen, Reference Poulsen1927); G. yatesi Robison and Babcock, Reference Robison and Babcock2011; and taxa originally assigned to Sonorapsis (blue): G. californica (Stoyanow and Susuki, Reference Stoyanow and Susuki1955); G. gomezi (Stoyanow in Cooper et al., Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952); and G. nelsoni (Stoyanow, Reference Stoyanow1958).

Figure 21. Long description
A bivariate scatter plot with the x-axis representing Glabellar length in millimeters from 0 to 40 and the y-axis representing Palpebral lobe length all over Glabellar length from 0.30 to 0.70.
Three colored polygons enclose specific data clusters:
- The Anoria cluster (red outline) is located at the bottom, spanning glabellar lengths of 5 to 23 millimeters and ratios between 0.28 and 0.46. It contains red and pink circular markers for A. tontoensis, A. bessus, and A. baton.
- The Sonorapsis cluster (blue outline) is centrally located, overlapping with the other two. It spans glabellar lengths of 8 to 18 millimeters and ratios between 0.38 and 0.55. It contains blue and cyan circular markers for G. californica, G. gomezi, and G. nelsoni.
- The Glossopleura cluster (black outline) is the largest and highest, spanning glabellar lengths from 0 to 40 millimeters and ratios from 0.37 to 0.67. It contains various black, gray, and white markers including squares, circles, diamonds, and crosses representing species like G. bion, G. boccar, G. producta, and G. stephenensis.
A legend below the plot organizes the species by genus. Glossopleura species are listed on the left and center with black and white symbols. Sonorapsis and Anoria species are listed on the right with blue and red/pink circular symbols respectively.
Anoria tontoensis (Walcott, Reference Walcott1916b)
Figures 15.1, 15.2, 22–24
Anoria tontoensis (Walcott, Reference Walcott1916b), from the Glossopleura walcotti Biozone, Bright Angel Formation, Grand Canyon, Arizona. All material is preserved in medium-grained sandstones, from USNM loc. 44345I. (1–3) Hypotype cranidium USNM 793400 (Walcott, Reference Walcott1925, pl 18, fig. 20), dorsal, anterior, and lateral views; (4) hypotype cranidium USNM 793999 (Walcott, Reference Walcott1925, pl. 18, fig. 19; Resser, Reference Resser1945, pl. 21, figs. 9, 10); (5) hypotype cranidium USNM 793994 (Walcott, Reference Walcott1925, pl. 18, figs. 17, 18; Resser, Reference Resser1945, pl. 21, figs. 9, 10). (6) hypotype cranidium USNM 793995c (Resser, Reference Resser1945, pl. 22, fig. 4, very top); (7) hypotype cranidium USNM 793995a (Resser, Reference Resser1945, pl. 22, fig. 4, very bottom; (8) cranidium USNM 793995b (Resser, Reference Resser1945, pl. 22, fig. 4, top); (9) hypotype librigena USNM 793402 (Walcott, Reference Walcott1925, pl. 18, fig. 22); (10) paralectotype pygidium USNM 793997 (Walcott, Reference Walcott1916b, pl. 51, fig. 1b; Walcott, Reference Walcott1925, pl. 18, fig. 28; Resser, Reference Resser1945, pl. 21, fig. 11); (11) paralectotype hypostoma USNM 793993 (Walcott, Reference Walcott1916b, pl. 51, fig. 1b; Walcott, Reference Walcott1925, pl. 18, figs. 24, 25; Resser, 1954, pl. 21, fig. 15); (12) hypotype librigena USNM 793401 (Walcott, Reference Walcott1925, pl. 18, fig. 21; Resser, Reference Resser1945, pl. 21, fig. 14); (13) thoracic segment USNM 793996; (14–16) paralectotype pygidium USNM 793998 (Walcott, Reference Walcott1916b, pl. 51, figs. 1g, 1g’; Walcott, Reference Walcott1925, pl. 18, figs. 26, 27; Resser, Reference Resser1945, pl. 21, figs. 12, 13), dorsal, posterior, and lateral views.

Figure 22. Long description
A collection of 16 fossil specimens numbered 1 through 16. A 5 mm scale bar is located near the center.
* Panels 1, 2, and 3 show hypotype cranidium U S N M 793400 in dorsal, anterior, and lateral views, displaying a rounded glabella and granular texture.
* Panel 4 shows hypotype cranidium U S N M 793999 in dorsal view.
* Panel 5 shows hypotype cranidium U S N M 793994.
* Panel 6 shows hypotype cranidium U S N M 793995c.
* Panel 7 shows hypotype cranidium U S N M 793995a.
* Panel 8 shows cranidium U S N M 793995b.
* Panel 9 shows hypotype librigena U S N M 793402, a long, curved cheek piece.
* Panel 10 shows paralectotype pygidium U S N M 793997, a broad, fan-shaped tail piece.
* Panel 11 shows paralectotype hypostoma U S N M 793993, a small, winged plate.
* Panel 12 shows hypotype librigena U S N M 793401 with a handwritten label 1399413 H.
* Panel 13 shows thoracic segment U S N M 793996, a narrow, pointed rib-like structure with a label 39 F.
* Panels 14, 15, and 16 show paralectotype pygidium U S N M 793998 in dorsal, posterior, and lateral views, highlighting the raised central axis and sloping pleural fields.
Anoria tontoensis (Walcott, Reference Walcott1916b), from the Glossopleura walcotti Biozone, Bright Angel Formation, Grand Canyon, Arizona. Specimens from USNM loc. 74e. (1–4) Lectotype shield USNM 794042 (Walcott, Reference Walcott1916b, pl. 51, fig. 1; Resser, Reference Resser1945, pl. 22, figs. 1, 3), dorsal of part and counterpart shield, axial spines, b/w and color. (5) Paralectotype shield USNM 794003 (Walcott, Reference Walcott1916b, pl. 51, fig. 1a; Resser, Reference Resser1945, pl. 22, fig. 2).

Figure 23. Long description
A multi-panel display of trilobite fossils.
* Panel 1: A black and white dorsal view of the lectotype shield U S N M 794042. It shows a broad cephalon, a segmented thorax with distinct pleurae, and a large pygidium. A black rectangular box outlines the central axial region.
* Panel 2: The counterpart shield of the same lectotype specimen in black and white, showing the impression of the fossil. A 1 cm scale bar is located at the bottom right.
* Panel 3: A high-magnification black and white view of the axial spines within the thorax and pygidium area highlighted in panel 1.
* Panel 4: A color version of the same high-magnification axial spine detail, showing greenish-grey rock matrix and reddish-brown mineral staining along the central axis.
* Panel 5: A black and white dorsal view of the paralectotype shield U S N M 794003. This specimen is smaller and more tapered than the lectotype, with a 1 cm scale bar at the bottom left.
All specimens exhibit the characteristic trilobite anatomy of a central axial lobe flanked by lateral pleural lobes.
Anoria tontoensis (Walcott, Reference Walcott1916b), from the Glossopleura walcotti Biozone, Bright Angel Formation, Diamond Bar Ranch and Grand Canyon, Arizona. (1) Articulated librigena and cranidium GRCA 123303, from GRCA loc. 363-2; (2) partial shield GRCA 123304 from GRCA loc. 363-2; (3) partial shield DMNS EPI. 42917 from FHPR loc. 006-5 / DMNS loc. 18384. Scale bars = 4-mm.

Figure 24. Long description
A multi-panel display of fossilized trilobite remains.
* Panel 1 at the top shows a wide view of a textured rock surface containing the articulated librigena and cranidium of specimen G R C A 123303. The fossil is low relief and blends with the surrounding matrix. A horizontal white scale bar is located in the bottom right corner.
* Panel 2 at the bottom left displays partial shield G R C A 123304. It features a segmented thorax with distinct horizontal ridges and a rounded tail section. A vertical white scale bar is positioned to the right of the fossil.
* Panel 3 at the bottom right shows a more detailed partial shield of specimen D M N S E P I dot 42917. This fossil exhibits a clear central axis with deep grooves separating the thoracic segments and a smooth, rounded pygidium at the base. A vertical white scale bar is located on the right edge.
All scale bars represent 4 millimeters.
Reference Walcott1916b Dolichometopus tontoensis Walcott, p. 373, pl. 51, figs. 1, 1a–1h.
Reference Walcott1924 Anoria tontoensis; Walcott, p. 54, pl. 9, fig. 2.
Reference Walcott1925 Anoria tontoensis; Walcott, p. 68, pl. 18, figs. 15–21, 23–28.
Reference Resser1945 Anoria tontoensis; Resser in Resser and McKee, p. 197–198, pl. 21, figs. 7–15, pl. 22, figs. 1–4.
Reference Stoyanow and Susuki1955 Sonoraspis californica Stoyanow and Susuki, p. 468, pl. 1, figs. 1, 2.
Reference Foster2011 “Anoria” tontoensis; Foster, p. 107–110, figs. 5, 6.
Reference Foster2022 Anoria tontoensis; Foster, p. 19, fig. 11F.
Lectotype
Shield USNM 794042 (Fig. 23.1–23.4) from USNM loc. 74e, Bright Angel Formation, Bright Angel Trail, Grand Canyon, Arizona.
Paralectotypes
USNM 794003 and 793993–793998 from USNM loc. 74, Bright Angel Formation, Nankoweap, Grand Canyon, Arizona.
Occurrence
Glossopleura walcotti Biozone, Bright Angel Formation, Frenchman Mountain (Flour Sack Member), Nevada; Diamond Bar Ranch (Flour Sack Member), Grand Wash Cliffs; Columbine Falls (Flour Sack Member); Peach Springs Wash; Palisades Section; Chuar Valley, Nankoweap Valley; Bright Angel Trail; Tapeats Creek.
Additional specimens
USNM 793999–794002 from USNM loc. 74, Bright Angel Formation, Nankoweap, Grand Canyon, Arizona.
Remarks
Foster (Reference Foster2011, Reference Foster2022) provided an extensive discussion of this species.
Anoria? bessus (Walcott, Reference Walcott1916b)
Anoria? bessus Walcott (Reference Walcott1916b), from the Glossopleura walcotti Biozone, Bright Angel Formation, Diamond Bar Ranch, Arizona. (1) Shield DMNS EPI. 42915 from FHPR loc. 006-12/ DMNS loc. 18384, missing librigena; (2) partial shield DMNS EPI. 42916 from FHPR loc. 006-15 / DMNS loc. 18384.

Reference Walcott1916b Dolichometopus? bessus Walcott, p. 332, pl. 51, figs. 3, 3a–3c.
Reference Resser1935 Anoria bessus; Resser, p. 10.
Reference Foster2022 Anoria? bessus; Foster, p. 19.
Lectotype
USNM 62699 from USNM loc. 62i, Wolsey Shale, Sixteen Mile Canyon, Meagher County, Montana.
Paralectotypes
USNM 62700–62701 from USNM loc. 62i, Wolsey Shale, Sixteen Mile Canyon, Meagher County, Montana.
Occurrence
Glossopleura walcotti Biozone, Flour Sack Member, Bright Angel Formation, Diamond Bar Ranch, Grand Wash Cliffs, Arizona. Glossopleura walcotti Biozone, Wolsey Shale, Sixteen Mile Canyon, Meagher County, Montana (Walcott, Reference Walcott1916b).
Remarks
This Grand Canyon species has the small, oval pygidium and long pleural spines on most segments like the material from Montana (Walcott, Reference Walcott1916b; Resser, Reference Resser1935; Schwimmer, Reference Schwimmer1973). The palpebral lobes in most specimens are rather long, similar to Glossopleura, suggesting that this species may in fact belong in that genus.
Genus Athabaskia Raymond, Reference Raymond1928
Type species
Athabaskia ostheimeri Raymond, Reference Raymond1928, from the middle Cambrian, Alberta, Canada.
Athabaskia sp. indet.
Miscellaneous corynexochids from Arizona. (1) Wenkchemnia? sp. indet., cranidium of FHPR 19623 from FHPR loc. 006-18, Glossopleura walcotti Biozone, Flour Sack Member, Bright Angel Formation, Diamond Bar Ranch. (2, 3) Olenoides? sp. indet.: (2) partial pygidium FHPR 19618 from FHPR loc. 006-21, Glossopleura walcotti Biozone, basal Rampart Cave Member, Muav Formation, Diamond Bar Ranch; (3) pygidium 794098 associated with the types of Pachyaspis sp. indet. from USNM loc. 73a, Elrathiella subzone, Ehmaniella Biozone Bright Angel Formation, Grand Canyon. (4) Athabaskia sp. indet., assigned to Clavaspidella sp. undet. by Resser (Reference Resser1945), pygidium USNM 794014 (Resser, Reference Resser1945, pl. 22, fig. 16), from USNM loc. 44344A, Glossopleura walcotti Biozone, Flour Sack Member, Bright Angel Formation, Grand Canyon. Scales: (1–3) = 2-mm scale bar; (4) = 5-mm scale bar.

Figure 26. Long description
A four-panel collection of grayscale photographs showing fossilized remains of corynexochid trilobites.
Panel 1, top left: A cranidium of Wenkchemnia? species indeterminate. The central glabella is elongated and slightly tapered, flanked by fixed cheeks with visible ocular ridges. A 2-mm scale bar applies to this and the next two panels.
Panel 2, top center: A partial pygidium of Olenoides? species indeterminate. It shows a segmented axis and pleural ribs extending toward a fragmented margin.
Panel 3, top right: A smaller, more complete pygidium of Olenoides? species indeterminate. The central axis is prominent with distinct horizontal segmentation, and the pleural fields show clear ribbing.
Panel 4, bottom: A large, wide pygidium of Athabaskia species indeterminate. This specimen is much broader than the others, featuring a wide, semi-circular shape with a central axis and faint radiating pleural furrows. A 5-mm scale bar is located in the bottom left corner of this panel.
Reference Resser1945 Clavaspidella sp. undet., Resser, p. 196, pl. 22, fig. 16.
Occurrence
Glossopleura walcotti Biozone, Flour Sack Member, Bright Angel Formation, Columbine Falls and Rampart Cave, Grand Canyon, Arizona.
Remarks
Resser placed this single pygidium into Clavaspidella, but the specimen is very similar to Athabaskia howelli (Walcott, Reference Walcott1886) with its posterior border nearly as wide as the pleural field. This specimen is left in open nomenclature due to the absence of cranidia, which are needed for species-level identification (see Palmer, Reference Palmer1954, p. 67). The reports of Glyphaspis from the Flour Sack Member, Bright Angel Formation at Columbine Falls and Rampart Cave by Resser (Reference Resser1945) are probably pygidia of Athabaskia. Specimens GRCA 2194 (not found in collection) and GRCA 11985 are both listed as Glyphaspis from the Columbine Falls locality; the latter specimen is probably a poorly preserved pygidium of Athabaskia although it has been coated with resin, which makes it difficult to identify. Resser (Reference Resser1945) also reported Clavaspidella sp. (USNM 794014) from the Gateway Canyon Member, Muav Formation from Havasupai Gardens (USNM loc. 44338F). Given its co-occurrence with Glyphaspis and the stratigraphic position, this specimen is probably Glyphaspis; however, Resser (Reference Resser1945) also reported Glossopleura sp. indet. from the same location.
Genus Glossopleura Poulsen, Reference Poulsen1927
Type species
Dolichometopus boccar Walcott, Reference Walcott1916b, from the upper Delamaran (Wuliuan Stage), Stephen Formation, Mount Bosworth, British Columbia, Canada.
Glossopleura boccar (Walcott, Reference Walcott1916b)
Glossopleura boccar (Walcott, Reference Walcott1916b), from the Glossopleura walcotti Biozone, Bright Angel Formation, Arizona. (1–6) Type specimens and additional material associated with types Glossopleura mckeei Resser, Reference Resser1945, preserved in shale unless otherwise noted: (1) paratype shield USNM 794043 (Resser, Reference Resser1945, pl. 21, fig. 1) from USNM loc. 74e; (2) paratype shield USNM 794044 (Resser, Reference Resser1945, pl. 21, fig. 3) from USNM loc. 74e; (3) paratype shield USNM 794047 (Walcott, Reference Walcott1916b, as Dolichometopus productus, pl. 53, fig. 4) from USNM loc. 74e; (4) paratype shield USNM 794045 (Walcott, Reference Walcott1916b, as Dolichometopus productus, pl. 53, fig. 4a), from USNM loc. 74e; (5) lectotype shield USNM 794046 (Resser, Reference Resser1945, pl. 21, fig. 2) from USNM loc. 74e; (6) paratype hypostome USNM 794049 internal mold (Resser, Reference Resser1945, pl. 21, fig. 5) from USNM loc. 74, preserved in sandstone. (7–14) Additional specimens from the Diamond Bar Ranch, Arizona, all preserved in limestone: (7) partially testate hypostoma FHPR 19560 from FHPR loc. 006-18; (8) exfoliated partial librigena FHPR 19558 from FHPR loc. 006-18; (9) mostly testate librigena FHPR 19565 from FHPR loc. 006-14; (10) exfoliated cranidium FHPR 19564 from FHPR loc. 006-14; (11) exfoliated cranidium FHPR 19562 from FHPR loc. 006-14; (12) partially exfoliated pygidium FHPR 19563 from FHPR loc. 006-14; (13) partially testate pygidium FHPR 19561 from FHPR loc. 006-18; (14) exfoliated pygidium FHPR 19559 from FHPR loc. 006-18. Scales: (1, 2, 5) = 1-cm white scale bars; (3, 4, 6, 7, 10, 12) = 5-mm scale bar; (8, 9, 11, 13, 14) = 2-mm scale bar.

Figure 27. Long description
A collection of 14 black and white photographs of trilobite fossils.
Panels 1 through 5 show complete or near-complete shields. Panels 1, 2, and 5 are large specimens with 1-centimeter scale bars. Panel 1 shows a paratype shield U S N M 794043. Panel 2 shows paratype shield U S N M 794044. Panel 5 shows the lectotype shield U S N M 794046. Panels 3 and 4 show smaller paratype shields U S N M 794047 and 794045 respectively, with a 5-millimeter scale bar.
Panels 6 and 7 show hypostomes. Panel 6 is an internal mold U S N M 794049 in sandstone. Panel 7 is a partially testate hypostoma F H P R 19560.
Panels 8 and 9 show librigena (free cheeks). Panel 8 is an exfoliated partial librigena F H P R 19558. Panel 9 is a mostly testate librigena F H P R 19565.
Panels 10 and 11 show cranidia (head sections). Panel 10 is an exfoliated cranidium F H P R 19564. Panel 11 is an exfoliated cranidium F H P R 19562.
Panels 12, 13, and 14 show pygidia (tail sections). Panel 12 is a partially exfoliated pygidium F H P R 19563. Panel 13 is a partially testate pygidium F H P R 19561. Panel 14 is an exfoliated pygidium F H P R 19559.
Scale bars are provided: 1 centimeter for panels 1, 2, 5; 5 millimeters for panels 3, 4, 6, 7, 10, 12; and 2 millimeters for panels 8, 9, 11, 13, 14.
Reference Walcott1916b Dolichometopus boccar Walcott, p. 363, pl. 52, figs. 1, 1a–f.
Reference Walcott1916b Dolichometopus productus; Walcott, (part), p. 369, pl. 53, figs. 4, 4a.
Reference Walcott1916b Dolichometopus tontoensis; Walcott, (part), p. 373, pl. 51, fig. 1h.
Reference Poulsen1927 Glossopleura boccar; Poulsen, p. 268.
Reference Resser1935 Glossopleura bosworthensis Resser, p. 31.
Reference Resser1935 Glossopleura nitida Resser, p. 31.
Reference Resser1935 Glossopleura mckeei Resser, p. 33.
Reference Resser1935 ?Glossopleura stephenensis Resser, p. 31.
Reference Resser1945 Glossopleura mckeei; Resser, p. 196, pl. 21, figs. 1–5.
Reference Rasetti1951 Glossopleura boccar; Rasetti, p. 164, pl. 24, figs. 1–6.
Reference Rasetti1951 Glossopleura mckeei; Rasetti, p. 165, pl. 24, figs. 9–12, 18.
Reference Sundberg2005 Glossopleura boccar; Sundberg, p. 65–66, figs. 6.10–6.14.
Reference Foster2011 Glossopleura boccar; Foster, p. 105–106, figs. 4.1–4.3.
Reference Foster2022 Glossopleura boccar; Foster, p. 16, fig. 11A, B, D, E.
Reference Handkamer, Pratt and MacNaughton2022 Glossopleura boccar; Handkamer et al., p. 21, fig. 15.1–15.6.
Cotypes
Cranidia USNM 62703, 62705, and 62707 from USNM loc. 15, Stephen Formation, Mount Bosworth, British Columbia, Canada.
Occurrence
Glossopleura walcotti Biozone. Bright Angel Formation, Frenchman Mountain (Flour Sack Member), Nevada; Diamond Bar Ranch (sandstones between the Tincanebits and Meriwitica tongues and Flour Sack Member), Grand Wash Cliffs; Havasu Creek, Nankoweap Valley, Bright Angel Trail, Little Colorado River Camp, ?Foster Canyon, Fossil Canyon, Tapeats Creek, Keyhole Canyon, Prospect Section, River Mile 200.8, Rampart Cave (Flour Sack Member). Glossopleura walcotti Biozone, Stephen Formation, Mount Bosworth, British Columbia, Canada (Rasetti, Reference Rasetti1951).
Remarks
See discussions by Sundberg (Reference Sundberg2005) and Foster (Reference Foster2011, Reference Foster2022) on this taxon. Identification of G. boccar (aka G. mckeei) in the Grand Canyon Bright Angel Formation is based on the relatively long length of the palpebral lobe and the narrow posterior area of the fixigena, relative to material from the Bright Angel assigned to Anoria tontoensis. Both of these features can be established on cranidia and librigena. At present, there are no other distinctive features that can be used to accurately separate cranidia of G. boccar and A. tontoensis. The presence of axial nodes on the pygidium is a possibility to differentiate the two species; however, this feature varies in both taxa (Sundberg, Reference Sundberg2005; Foster Reference Foster2011, Reference Foster2022).
Glossopleura tuta Resser, Reference Resser1945
Glossopleura tuta Resser, Reference Resser1945, from the Glossopleura walcotti Biozone, Rampart Cave Member, Muav Formation, Arizona. (1–3) Holotype pygidium USNM 794054 (Resser, Reference Resser1945, pl. 26, figs. 5, 6), USNM loc. 44340C, dorsal, lateral, and posterior views; (4) external mold of cranidium USNM 794051b (Resser, Reference Resser1945, pl. 20, fig. 22, bottom) from M&R loc. 44342B; (5) Glossopleura sp. undet. of Resser, Reference Resser1945, pygidium USNM 794051a (Resser, Reference Resser1945, pl. 20, fig. 22, center) from M&R loc. 44342B; (6) pygidium USNM 794052 from USNM loc. 44342B; (7) exfoliated partial cranidium GRCA 125242 from GRCA loc. 384; (8) mostly testate cranidium FHPR 19569 from FHPR loc. 006-23; (9, 10) partial cranidium GRCA 8558 from USNM loc. 44342B, dorsal and lateral views; (11) testate librigena FHPR 19566 from FHPR loc. 006-21; (12) testate partial cranidium GRCA 8564 from USNM loc. 44342B; (13) testate small librigena FHPR 19567 from FHPR loc. 006-21; (14) partial cranidium FHPR 19567 from FHPR loc. 006-23; (15) mostly testate, partial pygidium GRCA 8551 from USNM loc. 44342B, ventral view; (16) testate pygidium GRCA 8553 from USNM loc. 44342B, ventral view.

Figure 28. Long description
A collection of 16 monochrome photographs of trilobite fossils. A 5 mm scale bar is located at the top center.
* Panels 1 through 3 show the holotype pygidium U S N M 794054 in dorsal, posterior, and lateral views, featuring a smooth, semi-circular tail piece with a distinct central axis.
* Panel 4 shows an external mold of a cranidium, the central head part.
* Panel 5 displays a pygidium of Glossopleura species undetermined, showing a wide, fan-like structure.
* Panel 6 shows a pygidium U S N M 794052.
* Panel 7 shows an exfoliated partial cranidium G R C A 125242.
* Panel 8 shows a mostly testate cranidium F H P R 19569 with a rounded glabella.
* Panels 9 and 10 show a partial cranidium G R C A 8558 in dorsal and lateral views.
* Panel 11 shows a testate librigena, or free cheek, F H P R 19566.
* Panel 12 shows a testate partial cranidium G R C A 8564.
* Panel 13 shows a small testate librigena F H P R 19567.
* Panel 14 shows a partial cranidium F H P R 19567.
* Panels 15 and 16 show ventral views of pygidia G R C A 8551 and G R C A 8553, revealing the underside of the tail shield and the doublure.
Glossopleura tuta Resser, Reference Resser1945 from the Glossopleura walcotti Biozone, Rampart Cave Member, Muav Formation, Arizona. Articulated specimen FHPR 20141, float material from Diamond Bar Ranch (FHPR loc. 010), latex cast of mold in limestone. Scale = 1 cm.

Reference Resser1945 Glossopleura tuta Resser, p. 196–197, pl. 26, figs. 5, 6.
Reference Resser1945 Glossopleura sp. undet. Resser, p. 197, pl. 20, figs. 21, 22, pl. 21, fig. 6.
Reference Palmer and Halley1979 Glossopleura tuta; Palmer and Halley, p. 78, pl. 16, figs. 21–24.
Holotype
Pygidium USNM 794054 (Fig. 28.1–28.3) from USNM loc. 44340C, Bright Angel Formation, Diamond Creek and Peach Springs Wash junction, basal Rampart Cave Member, Muav Formation, Grand Canyon, Arizona.
Emended diagnosis
Cranidium with very shallow furrows on all parts except the hypostome; six thoracic segments; pygidium smooth, a longer axis (51–68%), and unscalloped, wide border (32–49%); pygidial axis only lightly furrowed.
Occurrence
Glossopleura walcotti Biozone, Rampart Cave Member, Muav Formation: Diamond Bar Ranch, Grand Wash Cliffs; Diamond Creek, Meriwitica Canyon, and River Mile 274, Grand Canyon, Arizona. Glossopleura walcotti Biozone, Carrara Formation, Striped Hills, Nevada.
Remarks
In addition to the type pygidium from the Diamond Creek section, additional specimens in the GRCA museum come from Meriwitica Canyon (McKee and Resser, Reference McKee1945; USNM loc. 44342B). These pygidia are large (18–21 mm) and preserved with the ventral surface exposed (exoskeleton preserved). There are also two cranidia (13.3 mm and 9.2 mm) from this site.
The new specimens (Fig. 29) collected from Mile 274 and Diamond Bar Ranch during this study are generally large and poorly preserved, but cranidia have the unique character of very shallow furrows on all parts except the hypostome. The only articulated specimen, from Diamond Bar Ranch (FHPR 20141), appears to possess only six thoracic segments.
The species is similar to G. bion (Walcott, Reference Walcott1916b) in having a smooth, unscalloped pygidial surface and a wide border relative to most other Glossopleura species (Resser, Reference Resser1945) but is differentiated by having a longer axis and slightly narrower border than G. bion (Schwimmer, Reference Schwimmer1973; Campbell, Reference Campbell1974). The pygidial axis is also only lightly furrowed (Palmer and Halley, Reference Palmer and Halley1979). Glossopleura tuta is broadly similar in the cranidium and pygidium to G. prona Resser, Reference Resser1939b, and G. arrecta Resser, Reference Resser1939b, from Utah, except in having shallower cranidial furrows; G. tuta is also similar to G. youngi Handkamer and Pratt in Handkamer et al., Reference Handkamer, Pratt and MacNaughton2022, from the Mount Cap Formation (Handkamer et al., Reference Handkamer, Pratt and MacNaughton2022) in the width of the pygidial border and in likely having six thoracic segments, but the pygidial axis in the latter species is noticeably longer.
All the material (type and non-type) is from the lower portion of the Rampart Cave Member of the Bright Angel Formation, except for one report from the Carrara Formation of Nevada (Palmer and Halley, Reference Palmer and Halley1979). This level of the Bright Angel contains a G. tuta and Kootenia tetraspinosa assemblage that also includes rarer elements of Amecephalus scapegoatensis, Am. piochensis, and Kochina vestita and abundant sclerites of Chancelloria sp.
Glossopleura sp. indet.
Occurrence
Glossopleura walcotti Biozone, below the Tincanebits Tongue, Bright Angel Formation to Rampart Cave Member, Muav Formation: Frenchman Mountain, Nevada; Diamond Bar Ranch, Grand Wash Cliffs; Rampart Cave, Columbine Falls, Malgosa Canyon, Palisades Section, Blacktail Canyon, Prospect Section, Granite Park, River Mile 274, Toroweap Valley, Diamond Creek (Spencer Canyon), Peach Springs Canyon (Sanap Plateau), Lava Canyon, Quartermaster Canyon (Peach Springs Member).
Remarks
When the diagnostic feature of palpebral length matching neither Glossopleura nor Anoria can be accurately determined, the specimens are referred to as Glossopleura sp. Specimens reported as Anoria sp. in McKee and Resser (Reference McKee and Resser1945) are included here as Glossopleura sp. Both genera are indicative of the G. walcotti Biozone.
Genus Wenkchemnia Rasetti, Reference Rasetti1951
Type species
Wenkchemnia walcotti Rasetti, Reference Rasetti1951, from the Delamaran (Wuliuan Stage), Mount Whyte Formation, Alberta, Canada.
Wenkchemnia kanabensis (Resser, Reference Resser1945)
Wenkchemnia kanabensis (Resser, Reference Resser1945), from the Ehmaniella subzone, Ehmaniella Biozone, Peach Springs Member, Muav Formation, Grand Canyon, Arizona. (1–6) Type and associated specimens of Clavaspidella kanabensis Resser, Reference Resser1945, from USNM loc. 75: (1) previously unfigured partial cranidium USNM 794013; (2) holotype cranidium USNM 794007 (Resser, Reference Resser1945, pl. 19, fig. 31); (3) paratype cranidium USNM 794011 (Resser, Reference Resser1945, pl. 19, fig. 29); (4) previously unfigured cranidium USNM 794009; (5) paratype hypostome USNM 794008 (Resser, Reference Resser1945, pl. 19, fig. 32); (6) paratype hypostome USNM 794010 (Resser, Reference Resser1945, pl. 19, fig. 20). (7–12) New material of Wenkchemnia kanabensis from near type locality in Kanab Canyon, in limestone: (7) exfoliated cranidium GRCA 125368 from GRCA loc. 378-1; (8) exfoliated partial cranidium GRCA 125374 from GRCA loc. 378-3; (9) partial pygidium GRCA 125370 from GRCA loc. 378-2; (10) exfoliated partial pygidium GRCA 125375 from GRCA loc. 378-3; (11) partially testate librigena GRCA 125367 from GRCA loc. 378-1; (12) exfoliated cranidium GRCA 125371 from GRCA loc. 378-2. (13) Ptarmigania sp. undet. of Resser Reference Resser1945, cranidium USNM 794135 (Resser, Reference Resser1945, pl. 19, fig. 33), from USNM loc. 74, Peach Springs Member, Muav Formation, Grand Canyon, Arizona. Scales: (1, 5, 7–13) 2-mm scale bar; (2–4, 6) 2-mm scale bar.

Figure 30. Long description
A collection of 13 fossil specimens labeled with white numbers in the top-right or bottom-right corners of each panel.
Panels 1 through 6 show type specimens of Clavaspidella kanabensis.
1. A partial cranidium showing the central glabella and one lateral wing.
2. The holotype cranidium, a complete central head shield with a bulbous glabella.
3. A smaller paratype cranidium with a 1 mm scale bar positioned below it.
4. An unfigured cranidium with distinct ocular ridges.
5. A paratype hypostome, a wing-like plate from the underside of the head.
6. A second paratype hypostome, smaller and more rounded.
Panels 7 through 12 show new material of Wenkchemnia kanabensis.
7. An exfoliated cranidium with a smooth, rounded glabella.
8. A partial exfoliated cranidium.
9. A partial pygidium, or tail shield, showing segmented ridges.
10. An exfoliated partial pygidium with visible pleural ribs.
11. A partially testate librigena, or free cheek, showing a curved outer margin.
12. A wide exfoliated cranidium with a horizontal crack across the center.
Panel 13 shows a cranidium of Ptarmigania species undetermined, featuring a highly convex glabella and deep furrows.
A 2 mm scale bar is located between panels 5 and 13 to provide physical dimensions for the larger specimens.
Reference Resser1945 Clavaspidella kanabensis Resser, p. 195–196, pl. 19, figs. 29–32.
?Reference Resser1945 Ptarmigania sp. indet.; Resser, p. 195–196, pl. 19, fig. 33.
Holotype
Cranidium USNM 794007 (Fig. 30.2) from USNM loc. 75, Peach Springs Member, Muav Formation, Kanab Canyon, Grand Canyon, Arizona.
Paratypes
Cranidia and hypostome USNM 794008–794013 from USNM loc. 75, Peach Springs Member, Muav Formation, Kanab Canyon, Grand Canyon, Arizona.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Peach Springs Member, Muav Formation, Kanab Canyon Grand Canyon, Arizona.
Remarks
Resser assigned three cranidia and one hypostome to this species; unfortunately, none of the sclerites is diagnostic for the genus. New materials found at Kanab Canyon, which is the type locality of the species, are small, but consist of cranidia, pygidia, and librigena. The cranidia are identical to Clavaspidella kanabensis, but the librigenal shape and boarder width and pygidial outline, three pleural furrows, and two or three axial rings (Fig. 30.9–30.11) are similar to Wenkchemnia swasensis Sundberg, Reference Sundberg1994, prompting the reassignment of the Grand Canyon specimens to Wenkchemnia. The two species differ in that the former has a more broadly expanding glabella with pronounced lateral glabella furrows.
Ptarmigania sp. indet. reported by Resser (Reference Resser1945; Fig. 30.13) is more than twice the size of C. kanabensis. This specimen has more pronounced lateral glabellar furrows and narrower (tr.) interocular fixigena than the latter specimens; however, these differences may be the result of ontogeny given that they are from the same locality (USNM loc. 75). The features of the single cranidium differ from the type species of Ptarmigania Raymond, Reference Raymond1928, Bathyuriscus rossensis Walcott, Reference Walcott1917, which has very faint S2–S4 glabellar furrows generating a frontal area of the glabella that looks egg-shaped (see Rasetti, Reference Rasetti1951, pl. 19, figs. 10, 14). Resser (Reference Resser1945) also listed Ptarmigania sp. indet. from near the base of the Bright Angel Formation in Bright Angel Canyon (GRCA loc. 367-1). It is unknown if this is the same taxon.
Family Dorypygidae Kobayashi, Reference Kobayashi1935
Genus Kootenia Walcott, Reference Walcott1889
Type species
Bathyuriscus (Kootenia) dawsoni Walcott, Reference Walcott1889, from the Topazan (Wuliuan Stage).
Remarks
The diagnosis presented by Sundberg (Reference Sundberg1994) is used herein. At present there are over 100 species of Kootenia, many that are based on the characteristics of the pygidium including the number of spine pairs (ranging from 1 to 6 pairs), length of spines, thickness of spines, and spine inflection (level, up, or down). In addition, the glabella of different species has different shapes ranging from parallel sided, expanded medially, expanding anteriorly to nearly egg-shaped. Some taxa have smooth exoskeletons, others have either fine to medium granules or a pitted surface, and sometimes these surface features are limited to only portions of the exoskeleton (e.g., spines). Kootenia has been extensively split; for example, Resser (Reference Resser1939b) named six species with five pairs of pygidial spines (four from the same locality), four species with six pairs of pygidial spines, one species with seven pairs of pygidial spines, and two species known only from cranidia. A major revision of the genus is needed, but at present large samples of different species are needed to assess the morphological variation within taxa.
Kootenia barensis new species
Kootenia barensis n. sp. from the Ehmaniella Biozone, Muav Formation, Diamond Bar Ranch. All specimens are from FHPR loc. 007-7 (= 007-8), preserved in limestone. (1, 2) Partial testate, paratype cranidium FHPR 19595 with occipital spine, dorsal and lateral views; (3) testate, paratype cranidium FHPR 19605; (4, 5) partial testate, paratype cranidium FHPR 19604, dorsal view and closeup view of pitted surface; (6) partial testate, paratype cranidium FHPR 19591; (7–9) testate, paratype cranidium FHPR 19601 with finely granulated surface, dorsal, lateral, and anterior views; (10) partial, mostly testate, paratype cranidium FHPR 19584; (11) mostly testate, paratype pygidium FHPR 19602; (12) partial, mostly testate, paratype cranidium FHPR 19590; (13–15) mostly testate, holotype pygidium FHPR 19580, dorsal, posterior, and lateral views; (16) partial, testate, paratype librigena FHPR 19597; (17) testate, paratype pleural segment FHPR 19597; (18) partly testate, paratype pygidium FHPR 19581; (19–21) mostly testate, paratype pygidium FHPR 19582, dorsal, posterior, and lateral views; (22, 23) partially testate, paratype librigena FHPR 19589, dorsal view and closeup of pitted surface; (24) exfoliated, paratype librigena FHPR 19599, dorsal view; (25–27) mostly testate, paratype pygidium FHPR 19593, dorsal, posterior, and lateral views; (28) testate, paratype pygidium FHPR 19603; (29, 30) mostly testate, paratype pygidium FHPR 19594, dorsal view and closeup of finely granulated surface; (31) partially testate pygidium FHPR 19596; (32) mostly testate, paratype pygidium FHPR 19583; (33) mostly testate, paratype pygidium FHPR 19598; (34) testate, paratype pygidium FHPR 19588. Scales: (1–4, 6–16, 18–22, 24–34) = 4-mm scale; (17) = white 2-mm scale bar; (5, 23) are enlargements of (4, 22) respectively.

Figure 31. Long description
The plate contains 34 panels numbered sequentially.
* Panels 1 through 10, and 12: Various views of cranidia (head segments). Panel 1 and 2 show dorsal and lateral views of a specimen with a prominent occipital spine. Panel 5 is a high-magnification closeup of the pitted surface texture of the cranidium in panel 4. Panels 7, 8, and 9 provide dorsal, lateral, and anterior views of a single cranidium with a finely granulated surface.
* Panels 11, 13 through 15, 18 through 21, and 25 through 34: Various views of pygidia (tail segments). Panels 13, 14, and 15 show dorsal, posterior, and lateral views of the holotype. Panels 19, 20, and 21 show dorsal, posterior, and lateral views of a paratype. Panel 30 is a high-magnification closeup of the granulated surface of the pygidium in panel 29.
* Panels 16, 22, and 24: Views of librigena (free cheeks). Panel 23 is a high-magnification closeup of the pitted surface of the librigena in panel 22.
* Panel 17: A single pleural segment.
Scale indicators include a black 4-mm scale bar located between panels 22 and 28, and a white 2-mm scale bar in panel 17. The fossils are light gray against a darker, textured limestone matrix.
Holotype
Pygidium FHPR 19580 (Fig. 31.13–31.15) from FHPR loc. 007-8, Kanab Canyon Member, Muav Formation, Diamond Bar Ranch, Arizona.
Paratypes
Cranidia, librigena, and pygidia FHPR 19581–19605 from FHPR loc. 007-8 (= 007-9), Kanab Canyon Member, Muav Formation, Diamond Bar Ranch, Arizona.
Diagnosis
Kootenia with pygidium having 6 pairs of very short, down-turned spines along edge of down-sloping border; 4 axial rings; no nodes; very shallow intrapleural furrows; and narrow border (9 ± 2% pygidial length). Cranidium width expanding anteriorly (115 ± 4% glabellar width), straight axial furrows, rounded frontal lobe, moderate length palpebral lobes (33 ± 3% glabellar length), and level, short occipital spine.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Kanab Canyon Member, Muav Formation, Diamond Bar Ranch, Grand Wash Cliffs, Arizona.
Description
Cranidium moderate size, length 8.8 ± 1.9 mm (n = 6); subpentagonal, length 71 ± 7% width; moderate sagittal and transverse convexity; anterior margin moderately curved 117 ± 6°, anterior margin width 67 ± 4% cranidial width; posterior margin, excluding occipital ring, straight to slightly backswept. Facial sutures parallel, 3 ± 7° from ends of palpebral lobes to anterior border; moderately convergent across anterior border; divergent 40 ± 6° posterior of palpebral lobes. Glabella elongate, length 97 ± 1% and width 52 ± 3% cranidial length; glabella width 37 ± 2% cranidial width; moderately expanding, width at anterior end 115 ± 4% glabellar width, frontal lobe has widest expansion at anterior end; high convexity (sag. and trans.); anterior end rounded, 3 lateral glabellar lobes. Axial furrows moderate depth, slightly shallower adjacent to lobe L3, slightly convex; preglabellar furrow moderately deep, moderately curved, shallower than axial furrow; lateral glabellar furrows very shallow, S1 furrow directed posteriorly, bifurcated; S2 furrow straight, S3 furrow directed anteriorly. Occipital ring sagittal length 15 ± 1% glabellar length, not elevated above glabella, moderately convex; very short, narrow, level occipital spine; furrow curved posteriorly, slightly deeper laterally, uniform depth medially, shallow depth; posterior margin curved. Frontal area length 3 ± 1% cranidial length. Preglabellar field absent. Anterior border moderately convex, down-sloping, widening laterally, evenly curved in front of glabella, straight laterally. Anterior border furrow evenly curved in front of glabella, less curvature laterally, moderately depth, shallower than axial furrows. Fixigena width, exclusive of palpebral lobe, 44 ± 5% glabellar width, moderately low convexity, down-sloping. Palpebral lobe narrow, width 19 ± 1% lobe length; short, length 33 ± 3% glabellar length; anterior margin located adjacent to 37 ± 3% glabellar length from anterior margin of frontal lobe; furrow moderate depth, wide. Ocular ridge very weak, joins with glabella at deep fossula, directed strongly posterolaterally from glabella at 43 ± 1° angle to axial trend. Posterior area of fixigena exsagittal width 25 ± 2% glabellar length; length 45 ± 6% glabella length; terminates moderately round. Posterior border moderately low, convex, wider distally; border furrow moderately deep shallowing laterally, straight.
Librigena moderate size, lengths 10–15 mm (n = 3); moderately narrow, width 42–44% length without spine; lateral margin moderately curved. Genal field moderately convex, width 20–30% librigenal width. Border width 36–38% librigenal width; moderately convex, level; lateral border furrow moderately deep, broad, uniform depth; posterior border furrow absent. Genal spine long, 64–72% librigenal length.
Hypostoma unknown.
Pleural segment with deep furrow, convex anterior and posterior bands, and strong fulcrum. Pleural spine short, extension of anterior band only, producing a notch anterior of spine.
Pygidium moderate size, length 6.9 ± 1.8 mm (n = 12); subcircular, length 64 ± 5% width; anterior margin directed laterally then moderately curved posteriorly; lateral margins evenly curved; posterior margin evenly curved; 6 pairs of spines, moderate width at base, down-sloping, most anterior positioned spine smallest, spines very short; moderate convexity (sag. and trans.). Axis moderately to slightly tapered, axial mid-width 86 ± 4% axial anterior width, axial anterior width 37 ± 2% pygidial width; axial length 91 ± 2% pygidial length, short low-relief postaxial ridge; 4 to possibly 5 axial rings, high convexity, nodes absent; terminal axial piece broadly rounded; axial furrows moderate depth; axial ring furrows moderate depth to very shallow, uniform depth. Pleural regions well defined, sagittally elongate, triangular shaped; moderate to high convexity, strongly down-sloping; anterior pleural furrow broad, moderate depth, extends to margin, curved slightly posterior; 3 additional pleural furrows, moderate depth to shallow; interpleural furrows very shallow. Border narrow, width 9 ± 2% pygidial length; slightly convex, down-sloping; border furrow shallow, broad, as deep as pleural furrows.
Exoskeleton thin; punctate to fine granular external surface and smooth internal surfaces.
Etymology
Species named after part of name the type location, Diamond Bar Ranch.
Remarks
Kootenia barensis n. sp. is most similar to K. tetraspinosa in possessing six pairs of relatively short pygidial spines that are directed ventrally and an anteriorly expanding glabella. The two species differ in the latter having a roughly triangle outline of the pygidium, slightly longer pygidial spines, and an occipital ring with a longer, upturned spine. The pygidia of K. barensis n. sp. and K. aff. K. incerta (Rusconi, Reference Rusconi1945) reported by Tortello (Reference Tortello2022) are similar, although the latter has longer marginal spines. The cranidia differ in the latter having a subquadrate glabella and a less curved anterior margin.
The exoskeleton of this species varies in its ornamentation between having finely pitted to finely granulated surfaces (Fig. 31.5, 31.7, 31.29). This appears to be consistent with sclerites having one or the other ornamentation. Both pitted and granulated specimens occur in the same horizon with no other morphological difference, suggesting that they represent the same species. Pitting and granulation of the exoskeleton have been used to define species of Kootenia. The co-occurrence of both in this species raises the question of whether surface ornamentation is a reliable species-defining character of Kootenia.
A small cranidium and a fragmented pygidium were found stratigraphically lower (FHPR loc. 007-5) in the same section as the types, which prevents accurate identification and is questionably assigned to the species.
Kootenia erosa (Resser, Reference Resser1945) new combination
Kootenia erosa (Resser, Reference Resser1945) n. comb., from the Ehmaniella subzone, Ehmaniella Biozone, Gateway Canyon Member, Muav Formation, Grand Canyon, Arizona; all material preserved in limestone. (1–7) Some of the type specimens of Solenopleurella erosa Resser, Reference Resser1945, from USNM loc. 73c, herein reassigned: (1–3) holotype testate pygidium USNM 794158 (Resser, Reference Resser1945, pl. 231, fig. 6), dorsal, posterior, and lateral view; (4) paratype pygidium USNM 794159 (Resser, Reference Resser1945, pl. 27, fig. 26), weathered specimen, exfoliated; (5, 6) paratype testate pygidium USNM 794164 (Resser, Reference Resser1945, pl. 23, fig. 5), posterior and dorsal views; (7) weathered large paratype pygidium USNM 794154; (8) weathered pygidium GRCA 8400c from GRCA loc. 368-8.

Figure 32. Long description
A multi-panel collection of eight black and white photographs showing fossilized trilobite pygidia.
* Panel 1 shows a dorsal view of the holotype U S N M 794158, featuring a segmented central axis and lateral lobes.
* Panel 2 shows the posterior view of the same holotype, highlighting the convex curvature of the shell.
* Panel 3 shows the lateral view of the holotype, illustrating the profile thickness.
* Panel 4 displays a weathered and exfoliated paratype U S N M 794159 with a rougher surface texture.
* Panel 5 and 6 show paratype U S N M 794164 in posterior and dorsal views respectively, with visible cracks across the testate surface.
* A scale bar located in the center indicates 4 mm.
* Panel 7 shows a large, heavily weathered paratype U S N M 794154 embedded in a coarse matrix.
* Panel 8 shows weathered pygidium G R C A 8400c, showing the outline of the fossil within the surrounding limestone rock.
Reference Resser1945 Solenopleurella erosa Resser, p. 214–215, pl. 23, figs. 5, 6; pl. 27, fig. 26 (not pl. 27, fig. 27, 28; = Spencella? sp. indet.).
Holotype
Pygidium USNM 794158 (Fig. 32.1–32.3) from USNM loc. 37c, Gateway Member, Muav Formation, Hermit Creek, Grand Canyon, Arizona.
Paratypes
Pygidia USNM 794154, 794159 and 794164 from USNM loc. 37c, Gateway Member, Muav Formation, Hermit Creek, Grand Canyon, Arizona.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Gateway Canyon Member, Muav Formation, Hermit Creek and Havasupai Gardens, Grand Canyon, Arizona.
Remarks
Resser (Reference Resser1945) assigned small cranidia and much larger pygidia from locality 37c to Solenopleurella erosa. However, these two different sizes of sclerites do not belong together, the pygidia represent Kootenia and the much smaller cranidia represent a small ptychoparioid here assigned to Spencella? sp. indet. Associated sclerites of K. erosa have not been identified. Resser (Reference Resser1945) also reported Anoria sp. and Solenopleurella erosa from USNM loc. 443381. These specimens belong to K. erosa (GRCA 8400a, pygidium) and S.? sp. A (GRCA 8400b, c, cranidia).
Kootenia erosa is also similar to K. havasuensis, but the latter species differs in having a stronger posterior notch in the pygidium (see below).
Kootenia aff. K. granulospinosa Palmer, Reference Palmer1968
Kootenia aff. K. granulospinosa Palmer, Reference Palmer1968, from the Ehmaniella subzone, Ehmaniella Biozone, Kanab Canyon Member, Muav Formation, Grand Canyon, Arizona. All specimens are from GRCA loc. 356. (1) Partial, partially testate cranidium GRCA 125270; (2) partial exfoliated cranidium GRCA 125271; (3) partial, partially testate cranidium GRCA 125272; (4) mostly exfoliated, fragmented cranidium GRCA 125281e, illustrating relatively large occipital spine; (5) testate librigena USNM 125273; (6, 7) testate librigena GRCA 125276b, boxed area in (6) is the close-up in (7) illustrating the fine-granule spine texture on the genal spine; (8) testate librigena GRCA 125284b; (9, 10) mostly exfoliated pygidium GRCA 125267b, dorsal and lateral views; (11) testate hypostome GRCA 125268; (12, 13) testate axial ring and spine GRCA 125269, posterior and lateral views; (14) fragmented, exfoliated pygidium GRCA 125276a illustrating only 4 pairs of spines. Scales: (1–8) 4-mm scale bar; (9–14) 2-mm scale bar.

Figure 33. Long description
A collection of 14 black and white photographs of trilobite fossil fragments.
Top Row:
1. Partial cranidium G R C A 125270 showing a smooth, rounded surface.
2. Exfoliated cranidium G R C A 125271 with a more textured, weathered appearance.
3. Partial cranidium G R C A 125272 showing the central glabella.
4. Fragmented cranidium G R C A 125281e featuring a prominent, thick occipital spine pointing downward.
Middle Row and Right Side:
5. Elongated librigena U S N M 125273 with a curved outer margin.
6. Librigena G R C A 125276b with a rectangular box highlighting the genal spine.
7. Close-up of the boxed area from panel 6, revealing a fine-granule texture on the spine surface.
8. Curved librigena G R C A 125284b.
Center and Bottom Row:
9. Dorsal view of pygidium G R C A 125267b, showing a central axis and five pairs of lateral spines.
10. Lateral view of the same pygidium, showing the elevation of the axial rings.
11. Small, shield-shaped hypostome G R C A 125268.
12. Posterior view of axial ring and spine G R C A 125269, showing a long, vertical projection.
13. Lateral view of the same spine, showing a slight posterior curvature.
14. Fragmented pygidium G R C A 125276a, notably showing only 4 pairs of spines.
Scale bars are located in the center: a 4-mm bar applies to panels 1 through 8, and a 2-mm bar applies to panels 9 through 14.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Kanab Canyon Member, Muav Formation, Nankoweap Canyon, Grand Canyon.
Remarks
Mostly fragments of Kootenia were found at the Nankoweap section; however, the fragments differ from most species of Kootenia and may represent a new species. One of the oddities is the presence of relatively long, thoracic ring spines (Fig. 33.12, 33.13). It is not uncommon for occipital rings to have spines, as this species does (Fig. 33.4), but the only known species with thoracic spines are K. randolphi Robison and Babcock, Reference Robison and Babcock2011, and K. youngorum Robison and Babcock, Reference Robison and Babcock2011, the former from the lower Bolaspidella Biozone, Wheeler Formation, and the latter from the G. walcotti Biozone, Spence Shale, Utah. It is interesting to note that like the pygidium illustrated here (Fig. 33.9, 33.10), axial spines are absent and axial nodes (if present) are poorly developed. The librigenal, axial ring, and pygidial spines of the specimens from Nankoweap a have very fine granulated to almost small spines covering the sclerites (Fig. 33.6–33.8, 33.12, 33.13). This granulated character is typical of K. granulospina from the “middle Cambrian-1 fauna” of Alaska reported by Palmer (Reference Palmer1968) and co-occurs with Altiocculus sp. and “Spencella” diligens (Resser, Reference Resser1945) of the Ehmaniella Biozone. This latter species is known from limited material but has smaller occipital and pygidial spines as well as six pairs of pygidial spines. Two forms of pygidia were found at this horizon, one with five pairs of spines (Fig. 33.9, 33.10) and one with only four pairs (Fig. 33.14). In the latter, the posterior pair of small spines are absent, similar to K. randolphi. Whether this variation represents one or two species is unknown. Kootenia youngorum differs from K. cf. K. granulospina in having a posterior spine that is very long relative to the other very short anterior and posterior spines.
Kootenia havasuensis Resser, Reference Resser1945
Kootenia havasuensis Resser, Reference Resser1945 from the Proehmaniella subzone, Ehmaniella Biozone, Elves Chasm Tongue and Sanup Plateau Member of the Muav Formation, Grand Canyon, Arizona. (1–7) Type and associated specimens from USNM loc. 38h, preserved in limestone: (1–3), paratype weathered cranidium USNM 794064 (Resser, Reference Resser1945, pl. 23, figs. 14, 15), dorsal, anterior, and lateral views; (4–6) holotype pygidium USNM 794063 (Resser, Reference Resser1945, pl. 23, figs. 16, 17), dorsal, lateral, and posterior views; (7) previously unfigured librigena USNM 794065. (8–10) Type and associated specimens of Kootenia simplex Resser, Reference Resser1945, here assigned as nomen dubium from USNM loc. 44345H, basal Sanup Plateau Member. All material preserved in calcareous sandstone, mostly exfoliated: (8) partial cranidium USNM 794077; (9) plastic cast of holotype pygidium 794076a (Resser, Reference Resser1945, pl. 20, figs. 16, top center, and 17); (10) plastic cast of paratype cranidium USNM 794076b (Resser, Reference Resser1945, pl. 21, fig. 16, left). Scales: (1–7) 2-mm scale; (8) 4-mm scale; (9, 10) 4-mm scale.

Figure 34. Long description
The photo contains ten panels labeled with numbers.
Panels 1 through 7 are associated with a 2-millimeter scale bar.
- Panel 1 shows a dorsal view of a weathered cranidium with a central oval glabella.
- Panel 2 shows the same cranidium from an anterior perspective.
- Panel 3 shows the cranidium in lateral view, highlighting its curvature.
- Panel 4 displays a dorsal view of a pygidium with distinct segmented lobes.
- Panel 5 shows the pygidium in lateral view.
- Panel 6 shows the pygidium from a posterior view.
- Panel 7 shows a curved librigena fossil.
Panels 8 through 10 are associated with a 4-millimeter scale bar.
- Panel 8 shows a partial cranidium with a granular surface texture.
- Panel 9 is a plastic cast of a pygidium showing a central axis and lateral pleural ribs.
- Panel 10 is a plastic cast of a cranidium featuring a prominent crack running diagonally across the glabella.
All specimens are preserved in either limestone or calcareous sandstone, with some showing signs of exfoliation.
Reference Resser1945 Kootenia havasuensis Resser, p. 202, pl. 23, figs. 14–17.
?Reference Sundberg1994 Kootenia cf. K. havasuensis; Sundberg, p. 26–27, fig. 17.
Holotype
Pygidium USNM 794063 (Fig. 34.4–34.6) from USNM loc. 38h, Elves Chasm tongue of the Muav Formation, Kwagunt Valley, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794064 and 794066 and librigena USNM 794065 from USNM loc. 38h, Elves Chasm tongue of the Muav Formation, Kwagunt Valley, Grand Canyon, Arizona.
Occurrence
Glossopleura walcotti Biozone? to Proehmaniella subzone, Ehmaniella Biozone, Meriwitica tongue? (Resser, Reference Resser1945) and Elves Chasm tongue, Bright Angel Formation to Rampart Cave Member, Muav Formation, Havasu Creek, and Rampart Cave, Grand Canyon, Arizona. Proehmaniella subzone, Ehmaniella Biozone Eldorado Formation, Egan Range, Nevada (Sundberg, Reference Sundberg1994).
Remarks
There are three species of Kootenia in the Grand Canyon that are unique in having a single pair of spines on the most anterior segment: K. havasuensis with smallest spines; K. simplex Resser, Reference Resser1945 (Fig. 34.9), and K. erosa (Resser, Reference Resser1945) n. comb. (Fig. 32) both with small spines. However, spine length in Kootenia can be variable (see discussion by Sundberg, Reference Sundberg1994, p. 23) and given the limited number of specimens and preservation, these three species could be synonymized. The type material of K. havasuensis is the best preserved, but differences do exist among the type material of the three assigned species. Kootenia havasuensis is most similar to K. simplex (Fig. 34.8–34.10) in overall features, but the latter species differs in having a slightly elongated pygidium with an undulating margin that may represent possible insipient spines (Fig. 34.9) and a cranidium with a less expanding frontal lobe (Figs. 24.8, 34.10). Until more, better preserved specimens of K. simplex are found the species should be considered nomen dubium. Kootenia havasuensis is also similar to K. erosa, but the latter species differs in having a stronger posterior notch in the pygidium. Kootenia cf. K. havasuensis identified by Sundberg (Reference Sundberg1994) differs from K. havasuensis in having a barrel-shape glabella instead of an expanding frontal lobe; but this glabellar shape is similar to the type material of K. simplex.
Similar cranidia and pygidia have been reported from the Ehmaniella Biozone of the Wolsey Shale, at Keegan Butt, Montana (Schwimmer, Reference Schwimmer1973). Although this species has not been formally named, the pygidia have the general outline of the K. havasuensis, but with a more pronounced posterior notch. Another similar species is K. georgiensis (Resser, Reference Resser1938a) from the Conasauga Formation, Georgia, which has only slight undulations for the pygidial anterior spines and a slight posterior notch; however, the single illustrated specimen is tectonically or compressionally distorted (Resser, Reference Resser1938a, pl. 7, fig. 13) that makes the present of the posterior notch questionable. Kootenia romensis Resser, Reference Resser1938a, from the Rutledge Limestone, Rome, Georgia, is also similar, but with a small pair of pygidial anterior spines. Both of the species from Georgia are poorly illustrated (Resser, Reference Resser1938b, pl. 7, figs. 13–15, 20, 21) where the photographs had the matrix trimmed away and/or they are small reproductions.
Kootenia mckeei Resser, Reference Resser1945
Kootenia mckeei Resser, Reference Resser1945, from the Ehmaniella subzone, Ehmaniella Biozone, Kanab Canyon Member of the Muav Formation, Grand Canyon and Diamond Bar Ranch, Arizona; all material preserved in limestone, mostly exfoliated. (1–5) Type and additional material of Kootenia mckeei Resser, Reference Resser1945, from USNM loc. 75: (1–3) holotype pygidium USNM 794067 (Resser, Reference Resser1945, pl. 20, figs. 7, 8), dorsal, oblique posterior, and lateral views; (4) paratype cranidium 794068 (Resser, Reference Resser1945, pl. 20, figs. 5, 6); (5) hypotype, partial pygidium 794069 (Resser, Reference Resser1945, pl. 20, fig. 10) questionably assigned to species, preserved in sandy limestone; (6–11) additional new specimens from FHPR loc. 007-9: (6) partial, mostly testate cranidium FHPR 19574; (7) partial pygidium FHPR 19575; (8) partly testate pygidium FHPR 19572; (9) mostly testate librigena from FHPR 19573; (10) partly testate pygidium FHPR 19571; (11) small cranidium FHPR 19570. Scales: (1–10) 4-mm scale bar, (11) white, 2-mm scale bar.

Figure 35. Long description
The photo contains eleven panels labeled 1 through 11.
* Panel 1 shows a dorsal view of a holotype pygidium with a central axis and six pairs of marginal spines.
* Panel 2 shows the same pygidium in an oblique posterior view, highlighting the elevation of the axis.
* Panel 3 shows the pygidium in lateral view.
* Panel 4 displays a paratype cranidium with a smooth, rounded glabella.
* Panel 5 shows a large, partial pygidium with prominent, elongated spines preserved in a coarse matrix.
* Panel 6 features a mostly testate cranidium with a wide glabella and distinct furrows.
* Panel 7 shows a large partial pygidium with a deeply segmented pleural field and long spines.
* Panel 8 shows a partly testate pygidium with a well-defined central axis.
* Panel 9 displays a mostly testate librigena, showing the curved outer margin and genal spine.
* Panel 10 shows a small, partly testate pygidium.
* Panel 11 features a small cranidium with a narrow glabella.
A black 4 mm scale bar is positioned in the center of the layout, and a white 2 mm scale bar is located at the bottom right under panel 11.
Reference Resser1945 Kootenia mckeei Resser, p. 200, pl. 20, figs. 5–10.
Holotype
Pygidium USNM 794067 (Fig. 35.1–35.3) Kanab Canyon Member of the Muav Formation, Grand Canyon, Arizona.
Paratype
Cranidium USNM 794068 Kanab Canyon Member of the Muav Formation, Grand Canyon, Arizona.
Occurrence
Ehmaniella Biozone, upper Bright Angel Formation? and Muav Formation, Diamond Bar Ranch (Kanab Canyon Member), Grand Wash Cliffs; River Mile 50, Nankoweap Canyon, Bright Angel Canyon, ?South Bass Trail, ?Blacktail Canyon, and Kanab Canyon, Grand Canyon, Arizona.
Remarks
The type specimens of K. mckeei Resser, Reference Resser1945, have a strongly curved anterior border, occipital spine?, and a pygidium with six pairs of relatively thin and short spines that increase in length posteriorly with the exclusion of the posteriormost pair that is shorter. The specimens of K. mckeei here are very similar to K. quadriceps (Hall and Whitfield, Reference Hall and Whitfield1877) figured by Palmer (Reference Palmer1954, pl. 14, figs. 1–4) from the Ute Limestone and by Sundberg (Reference Sundberg1994, fig. 19) from Altiocculus subzone, Ehmaniella Biozone in the Trailer Limestone, Utah, and the Eldorado Formation, Nevada. The major difference between the two species is that Kootenia mckeei has an expanded glabella where the anterior end is nearly 120–130% of the posterior end whereas K. quadriceps type specimens (Palmer, Reference Palmer1954, pl. 14, fig. 1) have about 115% and additional specimens have about 105% expansion (Sundberg, Reference Sundberg1994, fig. 19.1, 19.3). In addition, the anterior margin is strongly curved at 125° (smaller specimens) to 106° (larger specimen), whereas the type specimen illustrated by Palmer (Reference Palmer1954, pl. 14, fig. 1) is less curved at 136° and those illustrated by Sundberg (Reference Sundberg1994, fig. 19.1, 19.3) are strongly curved, ranging from 105° to 120°, similar to K. mckeei. This suggests that the specimens of Sundberg (Reference Sundberg1994) do not belong to either K. quadriceps or K. mckeei and may represent a new or different species.
At present the pygidia of K. mckeei and K. quadriceps illustrated by Palmer (Reference Palmer1954) and Sundberg (Reference Sundberg1994) cannot be separated into different species without associated cranidia. The pygidial spines of K. mckeei and K. quadriceps of Sundberg (Reference Sundberg1994) are directed nearly straight back (posterior) to slightly posterolaterally (Fig. 35.1). In the type specimens illustrated by Palmer (Reference Palmer1954) the spines are directed slightly posterolaterally. Variation in the character of the pygidial spines within a species is expected. Sundberg (Reference Sundberg1994) illustrated specimens from the same locality that have short spines to longer spines in both K. dawsoni and K. quadriceps.
Kootenia schenki Resser, Reference Resser1945
Kootenia schenki Resser, Reference Resser1945, from the Muav Formation, Grand Canyon, Arizona. All specimens are from impure or sandy limestones. (1–3) Holotype, mostly testate cranidium USNM 794070a (Resser, Reference Resser1945, pl. 20, figs. 11, 13), USNM loc. 44345J, from the base of the Peach Springs Member, Ehmaniella Biozone, dorsal, anterior, and lateral views; (4, 5) paratype specimens from USNM loc. 44345J: (4) mostly testate hypostome USNM 794070b (Resser, Reference Resser1945, pl. 20, fig. 13); (5) inverted photo of mold of pygidium USNM 794074 (Resser, Reference Resser1945, pl. 20, fig. 14); (6–9) paratypes from the base of the Rampart Cave Member, Glossopleura walcotti Biozone, USNM loc. 44342B; (6–8) paratype partially testate pygidium USNM 794075 (Resser, Reference Resser1945, pl. 20, fig. 17), posterior, dorsal, and lateral views; (9) partial exfoliated cranidium USNM 794073 (Resser, Reference Resser1945, pl. 20, fig. 16).

Figure 36. Long description
A multi-panel plate containing nine photos of fossilized trilobite parts. A scale bar labeled 4 m m is located near the bottom right.
* Panel 1: A large dorsal view of a cranidium showing a smooth, rounded central glabella and lateral fixed cheeks.
* Panel 2: An anterior view of the same cranidium showing the convex curvature of the glabella.
* Panel 3: A lateral view of the cranidium showing the profile of the glabella and the occipital ring.
* Panel 4: A dorsal view of a hypostome, an oval-shaped plate with a central body and lateral wings.
* Panel 5: An inverted photo of a pygidium mold showing a semi-circular tail shield with faint segmentation.
* Panel 6: A posterior view of a pygidium showing the vertical elevation of the axis.
* Panel 7: A dorsal view of the same pygidium showing a central axis and pleural regions with distinct marginal spines.
* Panel 8: A lateral view of the pygidium showing the downward slope of the pleural field.
* Panel 9: A dorsal view of a partial, exfoliated cranidium showing the internal texture of the glabella and a small occipital spine at the base.
Reference Resser1945 Kootenia schenki Resser, p. 200–201, pl. 20, figs. 11–14 (figs. 15–17 = K. tetraspinosa?)
Holotype
Cranidium USNM 794070a (Fig. 36.1–36.3) from USNM loc. 44345J, Peach Springs Member, Muav Formation, Peach Springs, Grand Canyon, Arizona.
Paratypes
Hypostome USNM 794070b and pygidium USNM 794075 from USNM loc. 44345J, Peach Springs Member, Muav Formation, Peach Springs; cranidium USNM 794073 and pygidium USNM 794074 from USNM loc. 44342B, Rampart Cave Member, Muav Formation, Rampart Cave, Grand Canyon, Arizona.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Peach Springs Member, Muav Formation, Rampart Cave, Grand Canyon, Arizona.
Remarks
Resser’s type material is from two different horizons. The holotype cranidium is from the basal Peach Springs at USNM loc. 44345J, which also contains Spe.? porcata and is from the Ehmaniella Biozone. Unfortunately, the type cranidium lacks characteristic to differentiate it from other species. In contrast, the paratypes are from the basal Rampart Cave at USNM loc. 44342B, which also contains “Alokistocare” and “Glossopleura” and belongs to the G. walcotti Biozone. Pygidia commonly are used to differentiate species of Kootenia, but only a single fragmentary pygidium (Fig. 36.6–36.8) is available from the type locality. As a result, the name K. schenki needs to be restricted to the holotype cranidium and paratype hypostome. In contrast, the other paratypes may belong to K. tetraspinosa given that they occur at the same locality (USNM loc. 44342B) and are here questionably assigned to the species because the pygidium has a more triangular outline.
Kootenia tetraspinosa Resser, Reference Resser1945
Kootenia tetraspinosa Resser, Reference Resser1945, from USNM loc. 44342B, the Glossopleura walcotti Biozone, lower Rampart Cave Member of the Muav Formation, Grand Canyon, Arizona; all material preserved in limestone, mostly exfoliated. (1–6) Type specimens of Kootenia tetraspinosa Resser, Reference Resser1945: (1–3) paratype cranidium USNM 794079 (Resser, Reference Resser1945, pl. 20, figs. 1, 2), dorsal, lateral, and anterior views; (4–6) holotype pygidium USNM 794078 (Resser, Reference Resser1945, pl. 20, figs. 3, 4), posterior, lateral, and dorsal views, arrows indicate small sclerites not part of pygidium. (7–14) Topotype material: (7–9) K. tetraspinosa?, testate cranidium GRCA 8711, dorsal anterior, and lateral views; (10) exfoliated, partial cranidium GRCA 8586; (11) mostly exfoliated, large cranidium GRCA 8560 illustrating occipital spine; (12) mostly exfoliated, partial pygidium GRCA 8567 illustrating two pairs of terminal spines not seen on holotype pygidium; (13, 14) testate, partial pygidium GRCA 2250 also illustrating two pairs of terminal spines, ventral view and inverted photograph. (15–25) Additional specimens: (15) testate, small, partial cranidium FHPR 19578 from FHPR loc. 006-21; (16, 17) partial exfoliated, cranidium GRCA 125228 from GRCA loc. 384, lateral and dorsal views; (18) testate partial, cranidium GRCA 125227a from GRCA loc. 384; (19) mostly testate pygidium GRCA 125229 from GRCA loc. 384; (20–22) mostly testate pygidium FHPR 19576 from FHPR loc. 006-21, lateral, dorsal, and posterior views; (23, 24) latex cast of pygidial axis and spines from FHPR 19576 from FHPR loc. 006-21, enlargement showing the Bertillon pattern; (25) exfoliated, partial pygidium FHPR 19577 from FHPR loc. 006-21. Scales: (1–14, 16–22), 4-mm scale bar; (15) white, 2-mm scale bar.

Figure 37. Long description
The figure consists of 25 numbered panels showing various fossil specimens.
Panels 1 through 3 show paratype cranidium U S N M 794079 in dorsal, lateral, and anterior views, featuring a rounded central glabella.
Panels 4 through 6 show holotype pygidium U S N M 794078 in posterior, lateral, and dorsal views. Small black arrows in panels 4 and 6 point to tiny sclerites near the tail piece.
Panels 7 through 9 show a testate cranidium G R C A 8711 in dorsal, anterior, and lateral views.
Panel 10 shows a partial, exfoliated cranidium G R C A 8586.
Panel 11 shows a large cranidium G R C A 8560 with a prominent occipital spine at the base.
Panel 12 shows a partial pygidium G R C A 8567 with two pairs of terminal spines.
Panels 13 and 14 show a ventral view and an inverted photograph of pygidium G R C A 2250.
Panel 15 shows a small cranidium F H P R 19578 with a 2-mm scale bar.
Panels 16 and 17 show lateral and dorsal views of cranidium G R C A 125228.
Panel 18 shows cranidium G R C A 125227a.
Panel 19 shows pygidium G R C A 125229.
Panels 20 through 22 show pygidium F H P R 19576 in lateral, dorsal, and posterior views, highlighting the segmented axis and marginal spines.
Panels 23 and 24 show a latex cast and a high-magnification view of the Bertillon pattern (fine fingerprint-like ridges) on the surface.
Panel 25 shows an exfoliated pygidium F H P R 19577.
A 4-mm scale bar is positioned between panels 8, 9, and 10.
Reference Resser1945 Kootenia tetraspinosa Resser, p. 199–200, pl. 20, figs. 1–4.
?Reference Resser1945 Kootenia schenki Resser, p. 200–201, pl. 20, figs. 16, 17 [not figs. 11–14].
Holotype
Pygidium USNM 794078 (Fig. 37.4–37.6) from USNM loc. 44342B, Rampart Cave Member, Muav Formation, Grand Canyon, Arizona.
Paratypes
Cranidium USNM 794079 from USNM loc. 44342B, Rampart Cave Member, Muav Formation, Grand Canyon, Arizona.
Emended diagnosis
Kootenia with pygidium having 6 pairs of short, down-turned spines along edge of down-sloping border; 4 axial rings; no nodes; very shallow intrapleural furrows; and moderate narrow border length (10–15% pygidial length). Cranidium expanding anteriorly (100–110% glabellar width), straight axial furrows, rounded frontal lobe, moderately short length palpebral lobes (20–30% glabellar length) and level, very short occipital spine.
Occurrence
Glossopleura walcotti Biozone, Rampart Cave Member, Muav Formation, Diamond Bar Ranch, Grand Wash Cliffs; Meriwitica Canyon and River Mile 274, Grand Canyon, Arizona.
Remarks
The type material reported by Resser (Reference Resser1945) is inadequate to name a species, with only a fragmented cranidium (Fig. 37.1–34.3) and pygidium (Fig. 37.4–37.6). Resser named this species based on the occurrence on only four pairs of pygidial spines; however, the posterior portion of the pygidium is covered by a small librigena. Topotype material from the GRCA collections clearly show that there are six pairs of pygidial spines (Fig. 37.12–37.14) and additional cranidial features to justify the naming of the species by Resser, although it is unfortunate that he used the name “tetraspinosa” to indicate the presences of only four pairs of pygidial spines.
Further complicating the identification of the species is Resser’s (Reference Resser1945) reporting of K. schenki material is from the “basal Rampart Cave” at USNM loc. 44342B, the type locality for K. tetraspinosa. It is likely that the cranidium USNM 794073 (Fig. 37.9) and pygidium USNM 794075 (Fig. 37.6–37.8), all from USNM loc. 44342B, belong to K. tetraspinosa given the pygidial spine number, thinness, length, and down-sloping features.
Additional specimens from other locations (Fig. 37.19–37.25) illustrate that the pygidial spines decrease in size from the anterior to posterior margin and the axis, anterior boarder, and spines have Bertillon marking and the pleural fields have a granular texture. The cranidia appear to have a smooth exoskeleton, but preservation is generally poor. A single cranidium from the topotype material has a more egg-shaped glabella (Fig. 37.7–37.9) and may represent a different species.
Kootenia spp. indet.
Occurrence
Glossopleura walcotti to Ehmaniella biozones, Bright Angel Formation and Muav Formation, Diamond Bar Ranch, Grand Wash Cliffs; River Mile 50, Kwagunt Valley, Havasupai Gardens, Peach Springs Canyon, Diamond Creek, Gateway Canyon, The Red Slide, Columbine Falls and Rampart Cave, Grand Canyon, Arizona.
Remarks
There are several occurrences of specimens that are recognizable as Kootenia, but it is unclear as to which species they belong because they lack specific diagnostic features. These specimens are left in open nomenclature and are not illustrated herein. These examples range from the G. walcotti to Ehmaniella biozones.
Genus Olenoides Meek, Reference Meek1877
Type species
Paradoxides? nevadensis Meek, Reference Meek1870, from the middle Topazan (Wuliuan Stage), Stephen Formation, British Columbia, Canada.
Olenoides? sp. indet.
Occurrence
Glossopleura walcotti Biozone, Bright Angel Formation to Rampart Cave Member, Muav Formation, Diamond Bar Ranch, Grand Wash Cliffs; Chuar Valley, Grand Canyon, Arizona.
Remarks
Two pygidial fragments have relatively narrow pygidial width, and an axis extending to nearly the posterior margin. These characteristics are similar to the pygidia of Olenoides, but not limited to this genus; thus, the questionable assignment. One specimen (Fig. 26.3) is associated with the type material of Pachyaspis sp. undet. by possessing six axial rings and a terminal piece. The other specimen (Fig. 26.2) is poorly preserved, internal mold in limestone from the base of the Rampart Cave Member.
Family Zacanthoididae Swinnerton, Reference Swinnerton1915
Genus Zacanthoides Walcott, Reference Walcott1888
Type species
Embolimus spinosa Rominger, Reference Rominger1887, from the middle Topazan (Wuliuan Stage), Stephen Formation, British Columbia, Canada (see discussion by Palmer, Reference Palmer1954, p. 70).
Zacanthoides cf. Z. typicalis (Walcott, Reference Walcott1886)
Zacanthoides cf. Z. typicalis (Walcott, Reference Walcott1886), from the Glossopleura walcotti Biozone, Diamond Bar Ranch, FHPR loc. 006-19 Flour Sack Member of the Bright Angel Formation, Arizona: (1) exfoliated, partial librigena FHPR 19639; (2) partial testate librigena FHPR 19638; (3) mostly exfoliated, fragmented pygidium FHPR 19640; (4) small pygidium FHPR 19642; (5) partial exfoliated pygidium FHPR 19641. Scales: (1, 3–5) 2-mm scale bar; (2) white, 2-mm scale bar.

Figure 38. Long description
The figure contains five panels labeled 1 through 5.
* Panel 1 at the top left shows a curved, crescent-shaped fossil fragment known as a librigena. It has a smooth, wide surface that tapers toward the bottom.
* Panel 2 to the right of panel 1 displays a more slender, elongated librigena with a sharp, needle-like posterior spine pointing downward. A small white horizontal scale bar is located at the bottom right of this panel.
* Panel 3 at the top right shows a segmented, oval-shaped pygidium. The segments are horizontal and slightly raised, though the lower portion of the fossil is fragmented and obscured by the matrix.
* Panel 4 at the bottom center shows a very small, complete pygidium with distinct horizontal ribbing and a tapered posterior end.
* Panel 5 at the bottom right shows a larger, partially exfoliated pygidium. It features clear horizontal segmentation and a rounded triangular overall shape.
A black horizontal scale bar labeled 2 m m is positioned in the bottom left corner of the overall figure layout.
Reference Walcott1886 Olenoides typicalis Walcott, p. 183, pl. 25, figs. 2, 2a.
Reference Pack1906 Zacanthoides typicalis; Pack, p. 299, pl. 3, figs. 2–2f.
Reference Walcott1913 Zacanthoides typicalis; Walcott, pl. 24, fig. 2.
Reference Shimer and Shrock1944 Zacanthoides typicalis; Shimer and Shrock, p. 619, pl. 257, figs. 20–22.
Reference Palmer1954 Zacanthoides typicalis; Palmer, p. 70–71, pl. 15, fig. 9.
Reference Robison and Babcock2011 Zacanthoides typicalis. Robison and Babcock, p. 19, 21, fig. 17.
Holotype
Shield USNM 15454 from USNM loc. 31, Chisholm Shale, Pioche, Nevada.
Occurrence
Glossopleura walcotti Biozone, Flour Sack Member, Bright Angel Formation, Diamond Bar Ranch, Grand Wash Cliffs, Arizona.
Remarks
Specimens tentatively assigned to this species are fragmented. The pygidia are similar to the type specimen in the broadness of the axis, narrowness of the pleural area, larger pleural spines directed posteriorly, and progressively smaller spines towards posterior margin.
Zacanthoides walapai Resser, Reference Resser1945
Zacanthoides walapai Resser, Reference Resser1945, from the Proehmaniella subzone, Ehmaniella Biozone. (1–10) Type and associated material from above the Rampart Cave Member, Diamond Creek, Arizona, preserved in shale and from USNM loc. 38i: (1) paratype cranidium USNM 794174a (Resser, Reference Resser1945, pl. 20, fig. 20, lower left); (2) holotype cranidium USNM 794173a (Resser, Reference Resser1945, pl. 20, fig. 19, center); (3) cranidium USNM 794176; (4) paratype cranidium USNM 794174b (Resser, Reference Resser1945, pl. 20, fig. 20, left); (5) cranidium USNM 794173b; (6) cranidium USNM 794177; (7) librigena USNM 794192; (8) paratype pygidium USNM 794179 (Resser, Reference Resser1945, pl. 20, fig. 19, inset); (9) pygidium USNM 794180; (10) pygidium USNM 794193. (11–16) Hypotypes of Zacanthoides sp. undet. of Resser, Reference Resser1945, from USNM loc. 38i, here assigned to Z. walapai; specimens preserved in limestone: (11) partial exfoliated cranidium USNM 794181 (Resser, Reference Resser1945, pl. 19, fig. 36); (12, 16) partial exfoliated cranidium USNM 794182 (Resser, Reference Resser1945, pl. 19, figs. 34, 35), dorsal and anterior views: (13–15) partial, mostly testate pygidium USNM 794184 (Resser, Reference Resser1945, pl. 19, figs. 37, 38), lateral, dorsal, and posterior views.

Figure 39. Long description
A multi-panel collection of fossil specimens with a 4 mm scale bar located in the center.
Panels 1 through 10 show specimens preserved in shale.
* 1, 2, 3, 4, 5, and 6: Various cranidia (head sections) showing the central glabella and fixed cheeks. Panel 2 is the holotype.
* 7: A single librigena (free cheek) with a long, slender genal spine.
* 8, 9, and 10: Pygidia (tail sections) showing a segmented central axis and lateral pleural ribs.
Panels 11 through 16 show specimens preserved in limestone, appearing more three-dimensional and lighter in color.
* 11: A partial exfoliated cranidium in dorsal view.
* 12 and 16: A partial exfoliated cranidium shown in dorsal (12) and anterior (16) views, highlighting the convex curvature of the head.
* 13, 14, and 15: A mostly testate pygidium shown in lateral (13), dorsal (14), and posterior (15) views, revealing the elevation of the axial rings and the downward slope of the posterior margin.
Reference Resser1945 Zacanthoides walapai Resser, p. 194, pl. 20, figs. 19, 20.
Reference Resser1945 Zacanthoides sp. undet.; Resser, p. 194, pl. 19, figs. 34–38.
?Reference Sundberg1994 Zacanthoides walapai; Sundberg, p. 51–52, fig. 31.1–31.4.
Holotype
Cranidium USNM 794173a (Fig. 39.2) from USNM loc. 38i, Sanup Plateau Member, Muav Formation, Diamond Creek, Grand Canyon, Arizona.
Paratypes
Cranidia and pygidium USNM 794173b–794193 from USNM loc. 38i, Sanup Plateau Member, Muav Formation, Diamond Creek, Grand Canyon, Arizona.
Occurrence
Proehmaniella subzone, Ehmaniella Biozone, Elves Chasm tongue, Bright Angel Formation, to Sanup Plateau Member, Muav Formation, Diamond Creek, Havasu Creek, Grand Canyon, Arizona. ?Proehmaniella subzone, Ehmaniella Biozone, Chisholm Formation, House Range, Utah; Eldorado Formation, northern Eagan Range, Nevada; and Pole Canyon Limestone, Member C, Snake Range, Nevada (Sundberg, Reference Sundberg1994).
Remarks
Resser (Reference Resser1945) differentiated Z. walapai from other species in possessing only one set of large, posterolateral margin, pygidial spines. In contrast, Sundberg (Reference Sundberg1994) identified limestone specimens showing one large posterolateral margin and two additional, very small, posterior pairs of pygidial spines and an undulatory posterior margin, which were assigned to Z. walapai. Given the crushed nature and shale preservation of the specimens from the Grand Canyon, it is possible that the collections of Resser and Sundberg are conspecific, and differences are due to differences in preservation. However, a shorter (tr.) preocular area and slightly longer pygidium suggests the specimens of Sundberg (Reference Sundberg1994, fig. 31) should be better assigned to Z. cf. Z. walapai.
The material illustrated by Resser as Zacanthoides sp. undet. are here assigned to Z. walapai given a pygidium with narrower axis and wider pleural field unlike pygidia of Z. typicalis.
Zacanthoides spp. indet.
Occurrence
Glossopleura walcotti to Ehmaniella biozones, Diamond Bar Ranch, Grand Wash Cliffs; Meriwitica Canyon, and Peach Springs Canyon, Grand Canyon, Arizona.
Remarks
Fragments of cranidia and librigena have been found, but the cranidial fragments are too small to confirm generic identification and no pygidia have been found. These specimens are not illustrated due to their fragmentary nature but are mentioned due to their relatively high stratigraphic range for Zacanthoides in the Grand Canyon. In addition, Resser (Reference Resser1945) reported Zacanthoides sp. from the Rampart Cave Member in Peach Springs Canyon (M&R loc. 69). Until additional material can be found, these specimens are left in open nomenclature.
Genus Albertella Walcott, Reference Walcott1908
Type species
Albertella helena Walcott, Reference Walcott1908, upper Delamaran (Wuliuan Stage), from the Gordon Shale near Scapegoat Mountain and Gordon Creek, Powell County, Montana (Resser, Reference Resser1936).
Remarks
The diagnosis of Albertella provided by Eddy and McCollum (Reference Eddy and McCollum1998) is used here.
Albertella nebeli new species
Figures 15.3, 40, 41
Larger specimens of Albertella nebeli n. sp. from GRCA loc. 331-1, Mexicella mexicana Biozone, Bright Angel Formation, Rampart Cave, Grand Canyon, Arizona; all specimens preserved in bioturbated shales. (1) Silicon cast of mostly complete shield, paratype GRCA 125383; (2) cranidium and partial thorax, paratype GRCA 125399a; (3) slightly disarticulated shield with hypostome impression, paratype GRCA 125391; (4) partial shield, holotype GRCA 125394; (5) partial, articulated cranidia and librigena, paratype GRCA 125396; (6) partial cranidium, paratype GRCA 125395; (7) partial cranidium, paratype GRCA 12554a; (8) articulated shield with missing librigena, paratype GRCA 125402; (9) partial cranidium, paratype GRCA 125392b; (10) partial pygidium, paratype GRCA 125397; (11) silicon cast of articulated cranidium, librigena, and thoracic segments, paratype GRCA 123390.

Figure 40. Long description
A collection of eleven black and white photographs of trilobite fossils, each labeled with a number in the top right corner. A 5 millimeter scale bar is located between panels 4 and 8.
* Panel 1: A nearly complete, elongated shield showing a distinct central axis and segmented pleurae.
* Panel 2: A cranidium attached to a partial thorax with seven visible segments.
* Panel 3: A disarticulated shield where the cephalon is slightly separated from the thorax, showing a hypostome impression.
* Panel 4: A partial shield focusing on the thoracic segments and the rounded pygidium at the base.
* Panel 5: A close-up of articulated cranidia and a librigena, showing the curved outer margins.
* Panel 6: A wide view of a partial cranidium with clear glabella furrows.
* Panel 7: A small, triangular fragment of a partial cranidium.
* Panel 8: The largest specimen, showing an articulated shield with a missing librigena and a prominent crack running diagonally through the cephalon and thorax.
* Panel 9: A partial cranidium with a vertical crack through the right side.
* Panel 10: A small partial pygidium showing the terminal segments of the trilobite body.
* Panel 11: A silicon cast of an articulated cranidium, librigena, and several thoracic segments, showing high relief detail.
Smaller specimens of Albertella nebeli n. sp. from GRCA loc. 331-1, Mexicella mexicana Biozone, Bright Angel Formation, Rampart Cave, Grand Canyon, Arizona; all specimens preserved in bioturbated shales. (1) Mostly complete shields, paratypes GRCA 125384b (left) and 125384c (right); (2) complete shield, paratype GRCA 125382; (3) complete shield, paratype GRCA 125393; (4) nearly complete shield, paratype GRCA 125389; (5) complete shield with a disarticulated librigena, paratype GRCA 125392a; (6) nearly complete shield, paratype GRCA 125399a; (7) cranidium, paratype GRCA 125256; (8) hypostome, paratype GRCA 125401; (9) pygidium, paratype GRCA 125398; (10) articulated, partial shield, paratype GRCA 125385; (11) silicon cast of articulated partial shield, paratype GRCA 125400; (12) partial pygidium with articulated thoracic segments, paratype GRCA 125384a.

Figure 41. Long description
A collection of 12 fossil photographs arranged in a grid. A black scale bar labeled 3 m m is positioned between panels 4 and 12.
Panel 1: Two specimens on a rock surface. The left is a small, faint impression; the right is a horizontal, elongated shield with distinct thoracic segments.
Panel 2: A vertically oriented, complete shield showing a rounded cephalon and a tapered thorax.
Panel 3: A slender, complete shield with a prominent glabella.
Panel 4: A large, nearly complete shield with well-defined pleural spines along the thorax.
Panel 5: A complete shield where the right librigena is disarticulated and shifted.
Panel 6: A nearly complete shield with a wide cephalon and segmented body.
Panel 7: A close-up of a single cranidium, showing the central head structure.
Panel 8: A small, isolated hypostome fossil.
Panel 9: A pygidium, or tail section, showing a segmented central axis and rounded border.
Panel 10: A partial shield showing the posterior segments and pygidium.
Panel 11: A silicon cast of a partial shield, showing high-relief details of the thoracic segments.
Panel 12: A large, partial pygidium with several articulated thoracic segments attached at the top.
Holotype
Thorax and pygidium GRCA 125394 (Fig. 40.4) from GRCA loc. 331-1, Me. mexicana Biozone, Bright Angel Formation, near Rampart Cave, Grand Canyon, Arizona.
Paratypes
Shields, cranidia, librigenae, hypostoma and pygidia GRCA 125254a, 125255, 125256, 125382–125384c, 125385, 125389–125391, 125392a–125393, 125395–125399b and 125400–125402 from GRCA loc. 331-1, Me. mexicana Biozone, Bright Angel Formation, near Rampart Cave, Grand Canyon, Arizona.
Diagnosis
Cranidia with narrow, upturned anterior border, well-defined expanding glabella; occipital ring node absent; moderate, medially placed palpebral lobe; ocular ridge angled at 52° from exsagittal line; postocular fixigena long (31 ± 5% glabellar length.); posterior suture moderately divergent; no granular ornamentation. Librigena with obtuse angle between the posterior margin of the free cheek and the inner spine margin. Pygidium with relatively narrow axis (40 ± 3% pygidial width) containing 3–4 axial rings; no thin, linear transverse nodes on anterior two axial rings; four pleural furrows; well defined border furrow; marginal spines straight directed posteriorly to slightly lateral; no granular ornamentation; spine length nearly as long or longer than pygidial length.
Occurrence
Mexicella mexicana Biozone, Bright Angel Formation, Rampart Cave and Toroweap Valley?, Grand Canyon, Arizona.
Description
Complete shields range in size from 2.8–24.2 mm (n = 9), not including pygidial spine. The cephalon length consists of 41 ± 5% of the shield length, thorax 36 ± 5%, and pygidia 27 ± 13%. All parts of the exoskeleton are smooth with no fine granular ornamentation apparent.
Cranidium small to moderately large in size, length 1.4–12.2 mm (n = 10); outline subtrapezoidal, length 67 ± 5% width; slightly convex; anterior margin evenly curved, width 63 ± 6% cranidial width. Anterior branches of facial sutures divergent at 10 ± 11°; posterior branches of facial sutures divergent at 42 ± 10°. Glabella length 96 ± 2% of cranidial length; width 30 ± 3% cranidial width; expanding slightly at anterior, width at anterior end 128 ± 7% of posterior glabellar width; slightly convex (sag. and trans.); frontal lobe moderately curved. Glabellar furrows shallow to very shallow with S1 being the longest and deepest, oriented posterolateral, and the others very shallow and shorter, S2 orientated slightly posterior, S3 transversely, and S4 slightly anteriorly; axial furrows shallow and distinct; preglabellar furrow moderately deep. Occipital ring slightly convex with no median node; length 19 ± 2% glabellar length; posterior margin slightly convex. Preglabellar field absent. Anterior border slightly convex, level, length 4 ± 2% cranidial length, widening laterally. Fixigena slightly convex, width 51 ± 12% of posterior glabellar width. Posterior fixigena behind palpebral long length 31 ± 5% glabellar length, width 48 ± 3% glabella length. Palpebral lobe curved, situated opposite of the mid glabellar length, lobe length 28 ± 4% glabellar length; anterior margin 34 ± 6% below glabellar front of glabellar length. Ocular ridge barely distinct, angled at 52 ± 7° from exsagittal line.
Librigenae length 3.4–7 mm (n = 2); moderately wide at 51 ± 6% length without spine; lateral margin curved. Genal field slightly convex. Border width 31 ± 2% librigenal width; border furrows moderate depth. Librigenal spine moderately long, 62 ± 13% librigenal length; directed posterolaterally. Rostral plate and hypostome poorly preserved.
Thorax with 7 segments in specimens larger than 2.7 mm. Axial rings tapering posteriorly, 7th ring is 71 ± 10% 2nd ring. Pleurae width 75 ± 10% of axial ring width (n = 9), slightly posterolaterally; 7th pleural width is 75 ± 10% of 2nd pleural width; fulcrum very poorly developed, probably due to preservation in shales. Pleural furrow moderately deep, uniform in width; anterior pleural band expands laterally, posterior band contracts laterally. Spines are projections of the anterior pleural band, thinning posterolateral, third spine is macropleural that extends to the lateral position of the pygidium.
Pygidium size variable, length 0.4–11.4 mm (n = 13); length 62 ± 8% width; anterior margin nearly straight and directed laterally; anterolateral corners curved. Axis moderately tapered, mid-width 79 ± 6% anterior width, anterior width 40 ± 3% pygidial width; length 87 ± 3% pygidial length, extends to nearly posterior margin; three to four axial ring segments, slightly convex; terminal piece small, somewhat pointed; axial ring furrows shallow. Marginal spines straight, directed posteriorly to slightly lateral, 5° to exsagittal line, length 108 ± 36% pygidial length (n = 6). Pleural field triangular with three moderate depth pleural furrows and a possibly short forth posterior furrow. Anterior border broad, well defined by convexity, border furrow shallow and broad.
Etymology
The species is named after Dr. Mark Nebel, Geoscience Program Manager, Grand Canyon National Park, for his continued administrative and logistical support for research on the Cambrian of the Park and for his patience to sit and collect several new specimens of the species.
Remarks
Despite the compression of specimens and their preservation in bioturbated shales, Alb. nebeli n. sp. is recognizable as a new species primarily based on the length and direction of the pygidial spine. The most similar species is Alb. highlandensis Eddy and McCollum, Reference Eddy and McCollum1998, from the Grassy Springs Member, Pioche Shale, eastern Nevada in its nearly directly posterior direction of the pygidial spines. The cranidia of Alb. nebeli n. sp. differs from Alb. highlandensis (n = 2 cranidia and 4 pygidia illustrated in Eddy and McCollum, Reference Eddy and McCollum1998; values rounded for both taxa) in the former having a more expanded frontal lobe (~130% vs. ~110% glabellar width), longer palpebral lobe length (~30% vs. ~20% glabellar length), ocular ridge angled more posterolaterally (~50° vs. ~60° from exsagittal line), medial node absent on occipital ring, posterior portion of facial suture directed more laterally (~40° vs. ~75° from transverse line). The pygidia of the two species differ in Alb. nebeli n. sp. having a spine length nearly as long or longer than pygidial length (~110% vs ~65%), no linear transverse axial nodes, well-defined border furrow, marginal spines directed posteriorly to slightly lateral (~5° vs. approximately −10° [convergent] to exsagittal line), straight, and no granular ornamentation. The magnitude of some of these differences may be due to preservation of the Alb. nebeli n. sp. in shale whereas Alb. highlandensis is preserved in limestone.
Resser (Reference Resser1945) reported but did not figure Z. cf. Z. walapai from the Transition Zone, Tapeats Sandstone from the Toroweap section (GRCA loc. 381-1). The single specimen (GRCA 2189) is a partial articulated specimen of Albertella cf. Alb. nebeli n. sp. The specimen is preserved as a negative impression and the details of the projection of the pygidial spines are unclear; thus, the specimen is left in open nomenclature. The specimen documents the occurrence of the Mexicella mexicana Biozone in the Tapeats Sandstone.
Albertella schenki Resser, Reference Resser1945
Albertella schenki Resser, Reference Resser1945, from the Mexicella mexicana Biozone, Bright Angel Formation, Rampart Cave, Grand Canyon, Arizona; all specimens preserved in shales and from GRCA loc. 389-2, unless otherwise noted: (1) inverted photograph of partial cranidium GRCA 125483; (2) cranidium and partial thorax GRCA 125491; (3) cranidium GRCA 125490b; (4) cranidium GRCA 125492; (5) librigena GRCA 125518d from GRCA loc. 389-1; (6) ventral view of librigena GRCA 125489; (7) internal mold of partial pygidium GRCA 125488 from GRCA loc. 389-2; (8) holotype pygidium USNM 793987 from USNM loc. 44345C; (9) pygidia GRCA 125518a (bottom left) and 125518b (upper right) from GRCA loc. 389-1; (10) hypostome GRCA 125518c from GRCA loc. 389-1; (11) partial pygidium GRCA 125484; (12) hypostome GRCA 125490a; (13) partial pygidium GRCA 125487. Scales: (1–6, 9–13) 2-mm scale bar; (7, 8) 4-mm scale bar.

Figure 42. Long description
A collection of 13 fossil specimens labeled with numbers in the top-right or bottom-right corners of each panel.
* Panel 1: An inverted photograph of a partial cranidium showing a broad glabella and fixed cheeks.
* Panel 2: A cranidium attached to a partial thorax with visible segments.
* Panel 3: A wide, short cranidium.
* Panel 4: A well-preserved cranidium with a distinct glabella.
* Panel 5: A long, curved librigena with a slender genal spine.
* Panel 6: A ventral view of a librigena showing the interior surface.
* Panel 7: An internal mold of a partial pygidium showing the segmented axis.
* Panel 8: The holotype pygidium, larger than others, showing a long, tapering axis and pleural spines.
* Panel 9: Two pygidia preserved together; one at the bottom left and one at the upper right.
* Panel 10: A small, winged hypostome.
* Panel 11: A partial pygidium with clear axial rings.
* Panel 12: A second hypostome specimen.
* Panel 13: A small, partial pygidium.
Two scale bars are included: a 2-mm scale bar located between panels 2 and 8, and a 4-mm scale bar located between panels 6 and 7.
Reference Resser1945 Albertella schenki Resser, p. 195, pl. 20, fig. 18.
Reference Palmer and Halley1979 Albertella schenki, Palmer in Palmer and Halley, p. 88, pl. 9, figs. 13–15, 17, 18.
Holotype
Pygidium USNM 793987 (Fig. 42.8) from USNM loc. 44345C, from the Bright Angel Formation, Rampart Cave, Grand Canyon, Arizona.
Emended diagnosis
Cranidium with narrow, level anterior border; well-defined, expanding glabella; occipital ring node absent; small, posteriorly placed palpebral lobes, ocular ridge angled at 62–75° from exsagittal line; postocular fixigena long (exsag.); posterior suture moderately divergent; no granular ornamentation. Hypostome with no distinct difference with the rostral plate. Librigena with 90° angle between the posterior margin of the free cheek and the inner spine margin. Pygidium with relatively narrow axis containing 4–5 axial rings, without nodes; four pleural furrows; well-defined border furrow; pygidial spines directed approximately 30° posterolaterally from axis; no granular ornamentation; spine length longer than pygidial length.
Occurrence
Mexicella mexicana Biozone, Bright Angel Formation, Frenchman Mountain, Nevada; Rampart Cave, Grand Canyon, Arizona; Mexicella mexicana Biozone, Pahrump Hills Shale Member, Carrara Formation Desert Range, Nevada (Palmer and Halley, Reference Palmer and Halley1979).
Remarks
Resser (Reference Resser1945) reported this species from only one shale specimen. New collections from the type locality (or near it) illustrate that the species does belong to Albertella based on cranidial and pygidial characteristics. Resser differentiated the species from Alb. helena in it possessing a greater proportional width and subangular posterior margin. Palmer (in Palmer and Halley, Reference Palmer and Halley1979) provided a full description of his specimens from the Desert Range, Nevada, which agrees well with the topotype material reported here. Palmer separated Alb. longwelli Palmer in Palmer and Halley, Reference Palmer and Halley1979, in having a distinctly differentiated cranidial anterior border, less well-defined glabella and two pygidial axial segments. The cranidial specimens of Alb. schenki from the Grand Canyon have a differentiated anterior border, but like Palmer and Halley specimens (Reference Palmer and Halley1979, pl. 9, figs. 13, 17), the medial portion of the anterior border is very thin (sag.; Fig. 42.1, 42.3).
Palmer and Halley (Reference Palmer and Halley1979) stated that Alb. proveedora Lochman in Cooper et al., Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952, is similar to Alb. schenki, but the latter differs in having longer palpebral lobes that are more posteriorly placed, a wider pygidial axis, spines directed at 20–35° and an obtuse angle between the posterior margin of the librigena and the inner spine margin. Albertella spectrensis Palmer in Palmer and Halley, Reference Palmer and Halley1979, differs in a well-defined upturned cranidial border, slightly larger more posteriorly placed palpebral lobes, occipital node, a distinct differentiation between the hypostome and rostral plate, and obtuse angle between the posterior margin of the librigena spine margin, and pygidial axial nodes.
Genus Caspimexis Özdikmen, Reference Özdikmen2005
Type species
Mexicaspis stenopyge Lochman, Reference Lochman1948, from the Delamaran (Wuliuan Stage), Arrojo Formation, Caborca, Sonora, Mexico. Özdikmen (Reference Özdikmen2005) renamed the genus due to its junior homonym to the Coleoptera Mexicaspis Spaeth, Reference Spaeth1938.
Caspimexis cf. C. radiatus (Palmer in Palmer and Halley, Reference Palmer and Halley1979)
Caspimexis cf. C. radiatus (Palmer in Palmer and Halley, Reference Palmer and Halley1979), from the Mexicella mexicana Biozone, Bright Angel Formation, Rampart Cave, Grand Canyon, Arizona; all specimens preserved in shales and from GRCA loc. 389-2. (1) Cranidium GRCA 125527; (2) cranidium GRCA 125530a; (3) inverted photograph of cranidium GRCA 125530d; (4) pygidium GRCA 125530c; (5) pygidium GRCA 125530e; (6) pygidium GRCA 125530f; (7) inverted photograph of pygidium GRCA 125530b.

Figure 43. Long description
The photo contains seven panels of trilobite fossils preserved in shale. A scale bar in the center indicates 2 mm.
* Panel 1 (Top Left): A large cranidium showing a rounded glabella and distinct lateral furrows.
* Panel 2 (Top Center): A cranidium with a prominent anterior border and wide fixed cheeks.
* Panel 3 (Top Right): An inverted photograph of a cranidium, highlighting the relief of the glabella against the matrix.
* Panel 4 (Bottom Left): A small pygidium with a central axis and faint pleural ribs.
* Panel 5 (Bottom Center-Left): A pygidium similar to panel 4, showing the posterior margin.
* Panel 6 (Bottom Center-Right): A pygidium with a more elongated central axis.
* Panel 7 (Bottom Right): An inverted photograph of a pygidium, showing the structural details of the axial rings and pleural spines in high contrast.
cf. Reference Palmer and Halley1979 Mexicaspis radiatus Palmer in Palmer and Halley, p. 92–93, pl. 10, figs. 22–25.
Occurrence
Mexicella mexicana Biozone, Bright Angel Formation, Rampart Cave, Grand Canyon, Arizona.
Remarks
Specimens from the Grand Canyon match those illustrated by Palmer and Halley (Reference Palmer and Halley1979) in that they have cranidia with a strongly curved, well-defined anterior border, long palpebral lobes, and relatively short (trans.) posterior area of the fixigena and the pygidia with three pairs of wide based pygidial spines that project nearly 90° to the border, and long pygidial axis. Differences of the specimens include the less expanding glabella, wider (trans.) anterior border, less pronounced pygidial border and the second set of spines being longer and wider as opposed to the third pair in C. radiatus. The differences in the cranidia and pygidia may be the result of the specimens being preserved in shale as opposed to the illustrated limestone specimens in Palmer and Halley (Reference Palmer and Halley1979). Alternatively, these specimens may represent a new species and until better preserved material is found, the specimens are left in open nomenclature.
Order Aulacopleurida Adrain, Reference Adrain2011
Remarks
Adrain (Reference Adrain2011) and Sundberg and Webster (Reference Sundberg and Webster2022) discussed the difficulty of recognizing the orders and families of the ptychoparioids. Adrain (Reference Adrain2011) placed the families Alokistocaridae Resser, Reference Resser1939b, Crepicephalidae Kobayashi, Reference Kobayashi1935, Ehmaniellidae Sundberg, Reference Sundberg1994 (= Alokistocaridae; Sundberg, Reference Sundberg1999; Esteve et al., Reference Esteve, Sundberg, Zamora and Gozalo2012), Marjumiidae Kobayashi, Reference Kobayashi1935, and Solenopleuridae Angelin, Reference Angelin1854, into the Order Aulacopleurida. Other ptychoparioid genera from the Grand Canyon are left in “Uncertain” as suggested by Adrain (Reference Adrain2011) due to a lack of phylogenetic analysis and incomplete knowledge of type species morphology (e.g., Antagmus Resser, Reference Resser1936; see Sundberg, Reference Sundberg, Landing, Milukic and Kluessendorf2007; Sundberg and Webster, Reference Sundberg and Webster2022).
Family Alokistocaridae Resser, Reference Resser1939b
Remarks
Sundberg (Reference Sundberg1999) discussed the revision of the Family Alokistocaridae based on the recognition that the type species Alokistocare subcoronatum (Hall and Whitefield, Reference Hall and Whitfield1877) had the characteristics typical of Ehmaniellidae Sundberg, Reference Sundberg1994.
Subfamily Alokistocarinae Hupé, Reference Hupé1955
Remarks
Sundberg (Reference Sundberg1994) recognized two subfamilies Ehmaniellinae and Altiocculinae. With the recognition that Ehmaniellidae and Alokistocaridae were synonymous, the subfamily Ehmaniellinae becomes synonymous with Alokistocarinae Hupé, Reference Hupé1955 (see Esteve et al., Reference Esteve, Sundberg, Zamora and Gozalo2012). It should be noted that the strict consensus cladogram of Esteve et al. (Reference Esteve, Sundberg, Zamora and Gozalo2012, fig. 1A) suggested that the two subfamilies cannot be differentiated. Further phylogenetic analysis of the subfamilies/family is needed but is out of the scope of this study.
Key cranidial characteristics used to diagnose genera within the family (derived from Sundberg, Reference Sundberg1994) include: (1) cranidial outline, (2) anterior facial suture direction, (3) palpebral lobe length, (4) anterior border length (sag.), tapering, and definition, (5) preglabellar field length (sag.) relative to frontal area, (6) glabella tapering and rounding of frontal lobe, and (7) posterior area of fixigenal length. Key pygidial characteristics include: (1) pygidial outline, (2) axis length and number of axial rings, (3) depth of pleural furrows, and (4) convexity of both posterior and anterior pleural bands.
Genus Ehmania Resser, Reference Resser1935
Type species
Ehmania weedi Resser, Reference Resser1935, from the upper Topazan (Wuliuan Stage), Meagher Limestone, Crowfoot Ridge, Wyoming.
Remarks
Ehmania has a subquadrate cranidium, divergent facial suture anterior of palpebral lobes; moderate to long palpebral lobes (40% glabellar length); well-defined anterior border, moderate width (25–33% cranidial length), flat, slightly downturned, moderately tapered; preglabellar field sagittally absent to narrow (0–25% frontal area length); slightly tapered glabella; bluntly rounded frontal lobe; exsagittally short posterior area of fixigena (≈ 25%). Pygidium subtriangular with elongated axis; 4–5 axial rings; moderate depth pleural furrow anterior pleural bands. Furrows and bands terminating near margin. This information is based on the type and additional specimens illustrated by Schwimmer (Reference Schwimmer1973).
Ehmania weedi Resser, Reference Resser1935
Ehmania weedi Resser, Reference Resser1935, from the Ehmaniella subzone, Ehmaniella Biozone, Gateway Canyon Member, Muav Formation, Grand Canyon, Arizona; all specimens are from GRCA loc. 357-3 and probably represent testate material, unless otherwise noted. (1) Partial cranidium GRCA 125408; (2) partial cranidium GRCA 125407; (3) partial cranidium GRCA 125409 with a thin coat of calcite; (4, 5) partial cranidium GRCA 125412, dorsal and anterior views; (6, 7) partial cranidium GRCA 125410, dorsal and lateral views; (8) partial cranidium GRCA 125411; (9) partial pygidium GRCA 125421, inverted photograph; (10) librigena GRCA 125414; (11) librigena GRCA 125415; (12) partial cranidium GRCA 125413a, exfoliated; (13) partial pygidium GRCA 125420; (14, 15) partial pygidium GRCA 125419, dorsal and posterior views; (16) partial pygidium GRCA 125417; (17, 18) partial pygidium GRCA 125418, dorsal and lateral views; (19) partial pygidium GRCA 125422. (20) Exfoliated partial cranidium USNM 794121 from USNM loc. 75, Peach Springs Member, Muav Formation.

Figure 44. Long description
A multi-panel plate of fossilized Ehmania weedi specimens. A 2 millimeter scale bar is located in the lower right quadrant.
* Panels 1 through 3: Three partial cranidia showing the central glabella and fixed cheeks.
* Panels 4 and 5: A single partial cranidium shown in dorsal view (4) and anterior view (5) highlighting the frontal curvature.
* Panels 6 and 7: A partial cranidium in dorsal view (6) and lateral view (7) showing the profile height.
* Panel 8: A larger, detailed dorsal view of a partial cranidium.
* Panel 9: An inverted photograph of a partial pygidium showing segmented pleural ribs.
* Panels 10 and 11: Two separate librigenae (free cheeks) showing the curved outer margin.
* Panel 12: An exfoliated partial cranidium with visible internal texture.
* Panel 13: A small partial pygidium.
* Panels 14 and 15: A partial pygidium in dorsal view (14) and posterior view (15).
* Panel 16: A partial pygidium showing distinct segmentation.
* Panels 17 and 18: A partial pygidium in dorsal view (17) and lateral view (18).
* Panel 19: A partial pygidium viewed from the rear.
* Panel 20: An exfoliated partial cranidium from a different geological member, showing a more granular surface texture.
Reference Walcott1899 Ptychoparia antiquata Walcott (not Salter), p. 456–457, pl. 65, figs. 7, 7a.
Reference Resser1935 Ehmania weedi Resser, p. 25.
Reference Resser1935 Ehmania walcotti Resser, p. 25.
Reference Deiss1939 Ehmania brevis Deiss, p. 79, pl. 13, figs. 20, 21.
Reference Deiss1939 Ehmania grandis Deiss, p. 80, pl. 14, figs. 6, 7.
Reference Deiss1939 Ehmania inconstans Deiss, p. 81, pl. 14, figs. 8–10.
Reference Deiss1939 Ehmania planioranta Deiss, p. 82, pl. 14, fig. 11.
Reference Deiss1939 Ehmania transversa Deiss, p. 83, pl. 14, figs. 20–21.
Reference Deiss1939 Ehmania sexannulata Deiss, p. 82, pl. 15, fig. 15.
Reference Handkamer, Ichaso, Pratt, Mángano and Buatois2023 Ehmania weedi; Handkamer et al. p. 1320–1322, pl. 9, figs. 1–3.
Holotype
Shield USNM 35234 from USNM loc. 151d, Meagher Limestone, Crowfoot Ridge, Wyoming.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone Gateway Canyon Member, Muav Formation, Nankoweap Canyon; Bright Angel Formation or Peach Springs Member, Muav Formation, Kanab Canyon, Grand Canyon, Arizona. Ehmania Biozone, Pagoda Limestone and Meagher Limestone, Wyoming and Montana, (Deiss, Reference Deiss1939; Schwimmer, Reference Schwimmer1973), or alternatively, the upper Altiocculus subzone, Ehmaniella Biozone, Earlie Formation, Saskatchewan (Handkamer et al., Reference Handkamer, Ichaso, Pratt, Mángano and Buatois2023, p 1320).
Remarks
The general cephalic and pygidial characteristics fit well with the species, but the presence of coarse granules on the pygidial pleural lobes are not obvious on the type specimens of Walcott and Deiss, but they are on the species from the Earlie Formation (Handkamer et al., Reference Handkamer, Ichaso, Pratt, Mángano and Buatois2023, fig. 9.1–9.3). The vast majority of this material has been broken during sediment transport and coated with a thin spar calcite layer that obscures surface ornamentation. All of the material appears to be replaced with phosphatic material. Schwimmer (Reference Schwimmer1973) and Handkamer et al. (Reference Handkamer, Ichaso, Pratt, Mángano and Buatois2023) discuss the variability of the species and synonymized several of the species assigned by Deiss (Reference Deiss1939). Handkamer et al. (Reference Handkamer, Ichaso, Pratt, Mángano and Buatois2023) synonymies are followed here.
Genus Ehmaniella Resser, Reference Resser1937
Type species
Crepicephalus (Loganellus) quadrans Hall and Whitfield, Reference Hall and Whitfield1877, from the Topazan (Wuliuan Stage), Spence Shale, Wasatch Range, Utah.
Remarks
Resser (Reference Resser1945) assigned three species to Ehmaniella. Ehmaniella arizonaensis is here assigned to Elrathiella (see below), Eh. basilica was established as the type species of Proehmaniella (Sundberg, Reference Sundberg1994), Eh. hebes was assigned to Proehmaniella by Sundberg (Reference Sundberg1994) but is here restored to Ehmaniella. In addition, Resser (Reference Resser1945) also assigned three new and one unnamed species to Parehmania: Par. tontoensis, Par. kwaguntensis, Par. nitida, and Par. sp. indet. The type species, Parehmania princeps Deiss, Reference Deiss1939 (re-illustrated by Schwimmer, Reference Schwimmer1973), differs from all of these species in possessing a glabella with a strongly rounded frontal lobe and relatively deep S1 lateral glabellar furrow with shallow connection to the axial furrows and the other lateral glabellar furrows very shallow to obscure. In addition, where the pygidia of the four Resser taxa are known, Par. princeps differs in having a suboval to sub-diamond with elongated axis; 4 axial rings; shallow pleural furrows; moderately convex pleural bands; weak posterior and anterior pleural bands. Of these taxa, Par. kwaguntensis and Par. nitida are here reassigned to Ehmaniella based on the bluntly rounded frontal lobe, longer palpebral lobes, exsagittally narrow posterior area of fixigena, and pygidial shape. Parehmania tontoensis and Par. sp. indet. fall within the morphological range of Elrathiella and are assigned to El. arizonaensis n. comb. (see below).
The strict consensus of Esteve et al. (Reference Esteve, Sundberg, Zamora and Gozalo2012, fig. 1.A) does not support the monophyly of Ehmaniella, although other analyses (Sundberg, Reference Sundberg1999; Esteve et al., Reference Esteve, Sundberg, Zamora and Gozalo2012, fig. 1B, 1.C) do.
Ehmaniella hebes Resser, Reference Resser1945
Ehmaniella hebes Resser, Reference Resser1945, from the Proehmaniella subzone, Ehmaniella Biozone, Bright Angel Formation or Muav Formation, Grand Canyon, Arizona; all specimens are mostly exfoliated in limestone from USNM locality 74a [20]. (1–3) Holotype cranidium USNM 794030a (Resser, Reference Resser1945, pl. 24, figs. 3, 4, top, right), dorsal, lateral, and anterior views; (4, 5) paratype cranidium USNM 794030b (Resser, Reference Resser1945, pl. 25, figs. 3, 4, bottom left), dorsal and anterior views; (6) paratype cranidium USNM 794029; (7) paratype cranidium USNM 794031 (Resser, Reference Resser1945, pl. 25, fig. 5); (8) paratype cranidium USNM 794032 (Resser, Reference Resser1945, pl. 25, fig. 2); (9) paratype cranidium USNM 794028 (Resser, Reference Resser1945, pl. 25, fig. 6); (10–12) paratype cranidium USNM 794190, dorsal and anterior views with close-up (11) of boxed area of anterior view (12) showing pits on internal mold and granules on external surface of the exoskeleton.

Figure 45. Long description
The multi-panel photo displays 12 views of trilobite cranidia specimens.
* Panels 1 through 3 show holotype U S N M 794030a. Panel 1 is a dorsal view showing the central glabella and right fixed cheek. Panel 2 is a lateral profile. Panel 3 is an anterior view showing the curvature of the frontal margin.
* Panels 4 and 5 show paratype U S N M 794030b. Panel 4 is a dorsal view and panel 5 is an anterior view.
* Panel 6 shows paratype U S N M 794029 in dorsal view.
* Panel 7 shows paratype U S N M 794031 in dorsal view.
* Panel 8 shows paratype U S N M 794032 in dorsal view.
* Panel 9 shows paratype U S N M 794028 in dorsal view.
* Panels 10 through 12 show paratype U S N M 794190. Panel 10 is a dorsal view. Panel 12 is an anterior view with a black box on the right side. Panel 11 is a high-magnification close-up of that boxed area, revealing fine pits on the internal mold and small granules on the external surface of the exoskeleton.
A black scale bar labeled 4 mm is positioned below panel 1 and above panel 6.
Reference Resser1945 Ehmaniella hebes Resser, p. 206–207, pl. 24, figs. 2–6.
Holotype
Cranidium USNM 794030a (Fig. 45.1–45.3) from USNM loc. 74a, Bright Angel Formation or Muav Formation, Nankoweap Valley, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794028, 794029, 794030b–794033, and 794190 from USNM loc. 74a, Bright Angel Formation or Muav Formation, Nankoweap Valley, Grand Canyon, Arizona.
Occurrence
Elrathiella or Ehmaniella subzones, Ehmaniella Biozone: Bright Angel Formation or Muav Formation, Nankoweap Valley, Grand Canyon, Arizona.
Remarks
Resser (Reference Resser1945, p. 206) differentiated this species based on the larger size, faint ocular ridges, shorter and less tapered glabella, “narrower anterior brim” and strongly down-sloping and “wider” preglabellar area. It is unclear if he was referring to narrower and wider in sagittal or transverse directions. The morphological features mentioned are present in other species of Ehmaniella, such as the faint ocular ridges (e.g., Eh. fronsplanata Sundberg, Reference Sundberg1994), “narrow” anterior margin (tr.; e.g., Eh. waptaensis Rasetti, Reference Rasetti1951 [illustrated in Sundberg, Reference Sundberg1994], Eh. angustigena Sundberg, Reference Sundberg1994), and glabellar tapering (e.g., Eh. fronsplanata, Eh. angustigena). However, distinctive characteristics of Eh. hebes are the relatively short glabella and corresponding long preglabellar area. The preglabellar area is steeply down-sloping medially, which is not present in other species of the genus, although other species do have steeply down-sloping anterior portion of the fixigena, but they have shorter, less sloped preglabellar areas (see Sundberg, Reference Sundberg1994). Variation in the type lot includes a more strongly curved (Fig. 45.1, 45.6, 45.7, 45.10) to less curved (Fig. 45.4, 45.8) anterior border. Although not mentioned by Resser (Reference Resser1945), the exoskeleton has a fine granular surface (Fig. 45.11, 45.12).
Sundberg (Reference Sundberg1994) assigned Eh. hebes to Proehmaniella, primarily based on the similarity of Pro. hebes? from the Pole Canyon Limestone, Snake Range, Nevada. However, these specimens are tectonically distorted, have more posteriorly placed and shorter palpebral lobes, and a rounded frontal lobe unlike the species from the Grand Canyon. The longer palpebral lobes and the broadly flattened frontal lobe indicate that the Grand Canyon specimens belong to Ehmaniella. Only one cranidium (Fig. 45.9) has the characteristics of Proehmaniella.
Resser (Reference Resser1945) reported types of Eh. hebes and El.? insueta) from the same locality collected by Walcott in 1882, but the horizon is stated to be in siltstones in the Bright Angel Formation or Muav Formation. Both type collections are preserved in limestone; however, El.? insueta specimens are tectonically distorted whereas the specimens of Eh. hebes are not. This suggests that the type lots were collected from two different horizons.
Ehmaniella kwaguntensis (Resser, Reference Resser1945) new combination
Figures 16.1, 16.8, 46, 47
Ehmaniella kwaguntensis (Resser, Reference Resser1945) n. comb., from USNM loc. 73b, Ehmaniella subzone, Ehmaniella Biozone, upper part of the Bright Angel Formation or lower Muav Formation, Grand Canyon, Arizona; all specimens are internal molds, mostly or entirely exfoliated, and preserved in fine-grained sandstone. (1, 2) Holotype cranidium USNM 794191b (Resser, Reference Resser1945, pl. 25, fig. 9, top right); (3, 4) paratype cranidium USNM 794105 (Resser, Reference Resser1945, pl. 26, fig. 7, left center), dorsal and anterior views; (5) paratype cranidium USNM 794191c (Resser, Reference Resser1945, pl. 25, fig. 9, top center right); (6) paratype cranidium USNM 79411d (Resser, Reference Resser1945, pl. 26, fig. 9, top center, pl. 25, fig. 11, top right center); (7) paratype cranidium USNM 794107 (Resser, Reference Resser1945, pl. 25, fig. 10, bottom left); (8) paratype cranidium USNM 794112a (Resser, Reference Resser1945, pl. 25, fig. 13, bottom left); (9) additional cranidium USNM 794112b; (10) additional cranidium USNM 794112d; (11) inverted photograph of USNM 794191d (Resser, Reference Resser1945, pl. 25, fig. 9, top left); (12) paratype librigena USNM 794111a (Resser, Reference Resser1945, pl. 25, fig. 11, center left, pl. 26, fig. 9, center left margin); (13) additional cranidium 794111b; (14–16) paratype pygidium USNM 794106a (Resser, Reference Resser1945, pl. 26, fig. 10); (17) paratype pygidium USNM 794111c (Resser, Reference Resser1945, pl. 25, fig. 10, top center); (18) paratype pygidium USNM 794109 (Resser, Reference Resser1945, pl. 26, fig. 9, bottom right corner); (19) paratype cranidium 794191e (Resser, Reference Resser1945, pl. 25, fig. 9, top left). Scales: (1–13, 17) 3-mm scale bar; (14–16, 18–22) 2-mm scale bar.

Figure 46. Long description
A collection of 19 photographs showing fossilized trilobite parts. Panels 1 through 11, 13, and 19 display cranidia (head shields) of varying sizes and preservation states. Panel 1 shows the holotype cranidium U S N M 794191b in dorsal view, featuring a prominent central glabella and wide fixed cheeks. Panels 2 and 4 provide anterior (front) views of cranidia, highlighting the convex curvature of the cephalon. Panel 11 is an inverted photograph of a cranidium. Panel 12 shows a paratype librigena (free cheek), which is elongated and curved. Panels 14, 15, 16, 17, and 18 show pygidia (tail shields). Panel 14 is a dorsal view of a pygidium with a distinct segmented axis and pleural ribs. Panels 15 and 16 show the pygidium in lateral or oblique views. Two scale bars are present: a 3 mm bar located between panels 8 and 9, and a 2 mm bar located near panel 16. All specimens are internal molds with a granular texture, typical of fine-grained sandstone preservation.
Ehmaniella kwaguntensis (Resser, Reference Resser1945) n. comb., from the Ehmaniella subzone, Ehmaniella Biozone, below Kanab Canyon Member, Muav Formation, Arizona; all specimens are preserved in limestone. (1) Cranidium FHPR 19620 from FHPR loc. 007-9; (2) partial cranidium FHPR 19621 from FHPR loc. 007-9; (3) partial, mostly testate cranidium FHPR 19619 from FHPR loc. 007-9; (4) testate cranidium GRCA 125439 and hypostome in upper right from GRCA loc. 357-5; (5) partial librigena FHPR 19622 from FHPR loc. 007-9; (6–8) exfoliated cranidium GRCA 125437 from GRCA loc. 357-5, dorsal, anterior, and lateral views; (9–11) pygidium GRCA 125440 from GRCA loc. 357-5, lateral, dorsal, and posterior views; (12) cranidium GRCA 125438 from GRCA loc. 357-5, external mold, inverted; (13) pygidium GRCA 125442 from GRCA loc. 357-5; (14) pygidium GRCA 125443 from GRCA loc. 357-5. Scales: (1–6, 12) 2-mm scale bar; (7–11, 13, 14) 1-mm scale bar.

Figure 47. Long description
The photo contains 14 numbered panels showing fossilized trilobite parts preserved in limestone.
* Panels 1 through 4: Located at the top, these show various cranidia (head shields). Panel 4 includes a small, oval hypostome in the upper right corner. A 2 mm scale bar is positioned between panels 2 and 6.
* Panel 5: Shows a partial librigena (free cheek) with a curved outer margin.
* Panels 6, 7, and 8: Feature a single exfoliated cranidium (G R C A 125437) in dorsal, anterior, and lateral views respectively, showing the convex glabella and surrounding fixigenae.
* Panels 9, 10, and 11: Show a pygidium (tail piece) in lateral, dorsal, and posterior views. A 1 mm scale bar is located to the left of panel 11.
* Panel 12: Displays an inverted external mold of a cranidium.
* Panels 13 and 14: Show two additional pygidia at the bottom, highlighting the segmented axis and pleural ribs.
The specimens vary in preservation from testate (with shell) to exfoliated (shell removed), revealing internal structural details of the exoskeleton.
Reference Resser1945 Parehmania kwaguntensis Resser, p. 208, pl. 25, figs. 9–13, pl. 26, figs. 7–10.
Holotype
Cranidium USNM 794191b (Fig. 46.1, 46.2) from USNM loc. 73b, Bright Angel Formation, Kwagunt Valley, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794105, 794107, 794111b, d, 794112a, b, d, and 794191b–e; librigena USNM 794111a; and pygidia USNM 794106, 794109, and 794111c from USNM loc. 73b, Bright Angel Formation, Kwagunt Valley, Grand Canyon, Arizona.
Emended diagnosis
Ehmaniella with narrow fixigena (35 ± 4% glabellar width), unequally divided frontal area (preglabellar region 34 ± 4% frontal area length), low convexity anterior border, tapered laterally, and suboval pygidium, 4 axial rings, and anterior and posterior pleural bands extending to near posterior margin.
Occurrence
Ehmaniella subzone Ehmaniella Biozone to Bolaspidella Biozone, upper Bright Angel Formation or Muav Formation, Diamond Bar Ranch (Kanab Member), Grand Wash Cliffs; Kwagunt Valley, River Mile 50, and Nankoweap Canyon (Gateway Canyon Member), Grand Canyon, Arizona.
Remarks
Resser (Reference Resser1945) identified this species based on the large size, short glabella, narrow preglabellar area, and broad anterior border, which differed from other previously recognized species of Parehmania. In overall character of the cranidium, this species is very similar to Eh. angustigena Sundberg, Reference Sundberg1994, in the relatively large glabella and frontal area construction, but differs in having a level and more tapered anterior border. One of the differences between Eh. kwaguntensis n. comb. and Eh. angustigena is that the former species has a suboval pygidium with 4 axial rings and anterior and posterior pleural bands nearly extending to the posterior margin. The latter has a more transversely elongated pygidium, shorter axis with 3 rings, and well-defined posterior border. The pygidium is similar to specimens of El. rectangularia Sundberg, Reference Sundberg1994.
Resser’s (Reference Resser1945) type material is preserved on the weathered surface of a sandy limestone with little or no exoskeleton preserved. Lateral glabellar furrows are generally absent although present on exterior molds (Fig. 46.11) and the curvature of the anterior border varies from slightly angled medially (Fig. 46.1) to smoothly curved (Fig. 46.5). New material assigned to this species (Fig. 47) is preserved in a purer limestone with some exoskeleton preserved. The similarity between the two assemblages is the tapered glabella in larger specimens (less tapered in smaller limestone specimens), curvature, length, and width of the anterior border, length of the preglabellar area, ocular ridge direction, palpebral lobe size and position, librigenal border and spine, and pygidial axial ring count, and variation in pygidial shape. The presence/absence of the lateral glabellar furrows appears to be related to the ontogeny of the species. The smaller limestone specimens illustrate well-defined S1 and S2 furrows (Fig. 47.1–47.3), whereas in the larger specimens (Fig. 47.6, 47.7) the furrows are very shallow. The type specimens are generally internal molds in a fine-grained sandstone and otherwise would not have had these very shallow furrows well preserved. The preserved exoskeleton shows scattered granules on the anterior border, preglabellar area, and fixigena (Fig. 47.3) and a pitted surface on the cranidial and pygidial internal molds (Figs. 47.6–47.12).
The axis of the pygidium (46.17) is wider than the other illustrated pygidia from the type collection and is more similar to the smaller pygidia from the limestone (Fig. 47.9–47.11). This specimen from the type collection may belong to a different taxon, possibly the co-occurring Trachycheilus typicalus Resser, Reference Resser1945.
Ehmaniella nitida (Resser, Reference Resser1945) new combination
Ehmaniella nitida (Resser, Reference Resser1945) n. comb., from the Ehmaniella subzone, Ehmaniella Biozone, Peach Springs Member, Muav Formation, Grand Canyon, Arizona. (1–15) Type and additional material from USNM loc. 75: (1–3) exfoliated holotype cranidium USNM 794113 (Resser, Reference Resser1945, pl. 24, figs. 8, 9), dorsal, anterior, and lateral views; (4–6) exfoliated paratype pygidium USNM 794115 (Resser, Reference Resser1945, pl. 24, figs. 13, 14), dorsal, posterior, and lateral views; (7–9) exfoliated paratype cranidium USNM 794118 (Resser, Reference Resser1945, p. 24, figs. 11, 12), dorsal, lateral, and anterior views; (10, 11) exfoliated cranidium USNM 794117, dorsal and anterior views; (12) exfoliated paratype pygidium USNM 794114 (Resser, Reference Resser1945, p. 24, fig. 15); (13) partially testate paratype cranidium USNM 794116 (Resser, Reference Resser1945, p. 24, fig. 7); (14) partially testate cranidium USNM 794119; (15) mostly testate cranidium GRCA 125377. (16–18) Additional material from Kanab Canyon GRCA loc. 378-1 preserved in limestone, exfoliated cranidium GRCA 125376, lateral, anterior, and dorsal views.

Figure 48. Long description
The plate contains 18 panels showing fossilized remains of the trilobite Ehmaniella nitida. A 2 mm scale bar is located near the center-right.
* Panels 1 through 3: Holotype cranidium U S N M 794113. 1 is a dorsal view showing a rectangular glabella and wide frontal border. 2 is an anterior view showing the curvature of the head. 3 is a lateral view showing the profile of the glabella.
* Panels 4 through 6: Paratype pygidium U S N M 794115. 4 is a dorsal view of the tail piece. 5 is a posterior view. 6 is a lateral view showing the segmented pleural ribs.
* Panels 7 through 9: Paratype cranidium U S N M 794118. 7 is dorsal, 8 is lateral, and 9 is anterior.
* Panels 10 and 11: Cranidium U S N M 794117 in dorsal and anterior views.
* Panel 12: Paratype pygidium U S N M 794114 in dorsal view.
* Panel 13: Partially testate paratype cranidium U S N M 794116 in dorsal view.
* Panel 14: Partially testate cranidium U S N M 794119 in dorsal view.
* Panel 15: Mostly testate cranidium G R C A 125377 in dorsal view.
* Panels 16 through 18: Cranidium G R C A 125376. 16 is a lateral view, 17 is an anterior view, and 18 is a large dorsal view showing fine surface details and the distinct furrows of the glabella.
Reference Resser1945 Parehmania nitida Resser, p. 207, pl. 24, figs. 7–15.
Holotype
Cranidium USNM 794113 (Fig. 48.1–48.3) from USNM loc. 75, Bright Angel Formation or Peach Springs Member, Muav Formation, Kanab Canyon, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794116–794122 and pygidia USNM 794114 and 794115 from USNM loc. 75, Bright Angel Formation or Peach Springs Member, Muav Formation, Kanab Canyon, Grand Canyon, Arizona.
Emended diagnosis
Ehmaniella with moderately narrow fixigena (44 ± 5% glabellar width), subequally divided frontal area (preglabellar region 46 ± 9% frontal area length), low convexity anterior border, tapered laterally, and suboval pygidium, 3 axial rings, and anterior and posterior pleural bands extending to anterior border.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Bright Angel Formation or Peach Springs Member, Muav Formation, Kanab Canyon, Grand Canyon, Arizona.
Remarks
This species is similar to Eh. kwaguntensis n. comb. but differs in the stronger curvature and flatter anterior border (Fig. 49) and the pygidium in having a shorter axis with three axial rings, pleural bands extending to the border, and the anterior lateral corners located adjacent to the anterior of the axis. Only two exfoliated pygidia are known, a larger collection of better-preserved specimens may show that some of the differences are either variable or not preserved in these specimens. Overall, the cranidia are similar and they overlap in morphologic space (Fig. 50.3).
Bivariate plot of cranidial size versus anterior border furrow angle degrees, of various taxa of Alokistocaridae (see Fig. 50) from the Grand Canyon. Solid red line indicates the mean; dashed red lines indicate standard deviation, 158 ± 6°, n = 23, for type and topotype specimens Elrathiella arizonaensis (Resser, Reference Resser1945) n. comb., which are preserved in shale. See text for discussion.

Figure 49. Long description
A scatter plot with the X axis representing Cranidial width in m m ranging from 1 to 14 and the Y axis representing Anterior border furrow angle in degrees ranging from 140 to 175.
Three horizontal red lines span the width of the graph. A solid red line at 158 degrees represents the mean. Two dashed red lines at 153 and 164 degrees represent the standard deviation.
Data is grouped into several overlapping shaded polygons representing different taxa.
- A large light yellow polygon labeled Ehmaniella arizonaensis Shale encompasses the majority of the central data points from 2 to 11 m m width.
- A smaller dark brown polygon labeled Ehmaniella arizonaensis Sandstone is centered vertically around 5.5 m m width.
- To the left, a brown polygon represents Spencia tontoensis and a light purple polygon represents Pachyaspis moorei.
- In the lower center, a dark purple polygon represents Parehmania nitida and a pink polygon represents Parehmania kweguntensis.
- A blue polygon at the top center represents Parehmania tontoensis.
- On the far right, a dark blue triangular polygon represents Elrathiella decora between 10.5 and 14 m m width.
- Individual points for Pachyaspis sp. and Pachyaspis fronticola are indicated with gray lines and black or gray diamonds.
A legend in the top right corner identifies Ehmaniella arizonaensis markers. A green diamond for type specimens, an orange diamond for other shale specimens, and a yellow diamond for sandstone.
PCA plots of various taxa of Alokistocaridae from the Grand Canyon. Most of the species are labeled with the taxonomic name given to them by Resser, Reference Resser1945, and do not represent their present taxonomic assignment (e.g., Parehmania kwaguntensis Resser, Reference Resser1945 = Ehmaniella kwaguntensis [Resser, Reference Resser1945], see text) and consist primarily of the type material. The data for Elrathiella arizonaensis (Resser, Reference Resser1945) n. comb. consists of the type, topotype, and new material to illustrate the morphological range of the taxon. PCA plot of (1) PC1 vs. PC2; (2) PC1 vs. PC3; (3) PC2 vs. PC3.

Figure 50. Long description
A three-panel vertical stack of P C A scatter plots.
Panel 1 at the top plots P C 1 on the horizontal axis against P C 2 on the vertical axis. A horizontal arrow labeled Size points right along the P C 1 axis. Data points are grouped into colored convex hulls. On the far left, clusters include Parehmania nitida and Spencia tontoensis. The center is dominated by a large yellow hull for Ehmaniella arizonaensis Shale and a smaller brown hull for Ehmaniella arizonaensis Sandstone. To the far right, a blue triangular hull represents Elrathiella decora.
Panel 2 in the middle plots P C 1 against P C 3. The distribution shifts, with Parehmania nitida moving to the bottom left and Elrathiella decora moving to the bottom right. The large yellow Ehmaniella arizonaensis Shale hull remains central but elongated horizontally.
Panel 3 at the bottom plots P C 2 against P C 3. The clusters are more overlapping in the center. Text labels around the perimeter of this panel define morphological variables: D 9 palpebral lobe position, F i fixigenal width, J 2 anterior cranidial width on the left; B 1 glabellar length, K 6 anterior glabellar width, G 1 anterior border length on the top right; and C Palpebral lobe length, K 2 posterior glabellar width, E occipital ring length, and H preglabellar length on the bottom right.
Legend in panel 1 identifies symbols: open diamonds for type specimens, solid diamonds for other shale specimens, and solid circles for sandstone.
Two new specimens from Kanab Canyon, which is near the type locality show some differences compared to the type material. One specimen (Fig. 48.16) indicates that the cranidia has a granulated surface similar to Eh. kwaguntensis n. comb. and Eh. hebes and has better defined glabellar furrows. A larger cranidium (Fig. 48.17–48.19) has a glabella that is more convex (tr.), less tapered, and a slightly rounded frontal lobe. Both specimens have a less tapered and longer anterior border and may represent a separate species; however, PCA (Fig. 50) illustrates that these specimens cluster with other representatives of Eh. nitida n. comb.
Genus Elrathiella Poulsen, Reference Poulsen1927
Type species
Elrathiella obscura Poulsen, Reference Poulsen1927, from the Topazan (Wuliuan Stage), Pemmican River Formation, Inglefield Land, Greenland.
Remarks
Resser (Reference Resser1945) assigned several taxa from the Grand Canyon to Elrathiella, Spencia, Resser, Reference Resser1939a, Parehmania, and Pachyaspis Resser, Reference Resser1939b. Unfortunately, most of the assigned specimens are preserved in medium- to fine-grained sandstone, internal molds, and limonite coated and mostly cranidia, with few librigenae or pygidia. Spencia, based on the type species S. typicalus Resser, Reference Resser1939a, illustrated with only small photographs (Resser, Reference Resser1939a, pl. 6, figs. 22–25, cranidia are 1 cm or less in length in photos); however, Campbell (Reference Campbell1974) in his unpublished thesis re-illustrated the type specimens from the Spence Shale. Spencia differs from Elrathiella in its parallel sided glabella, nearly effaced glabellar furrows, and small palpebral lobes and overall is very similar to Elrathina. These characteristics are not typical of Spencia tontoensis from the Grand Canyon.
Pachyaspis, based on the type species Pac. typicalis Resser, Reference Resser1939b, is well illustrated by Resser (Reference Resser1939b, pl. 11, figs. 15–20, pl. 12, figs. 1–3), although no librigena or pygidia are illustrated. Campbell (Reference Campbell1974) illustrated the new specimens from the Spence Shale, including pygidia. Pachyaspis differs from Elrathiella in its more inflated glabella, thin (tr.) palpebral lobes, faint ocular ridges, more tapered glabella with less defined glabellar furrows and a broadly transverse pygidium with the anterior lateral margins at the mid-axial level. These characteristics are not typical of either Pac. fonticola or Pac. sp. reported from the Grand Canyon.
Parehmania, based on the type species Par. princeps Deiss, Reference Deiss1939, is illustrated with retouched photos in Deiss (Reference Deiss1939, pl. 18, figs. 10–11) and again by Schwimmer (Reference Schwimmer1973), including complete shields. Schwimmer (Reference Schwimmer1973) and Handkamer et al. (Reference Handkamer, Ichaso, Pratt, Mángano and Buatois2023) synonymized 20 different species of Deiss (Reference Deiss1939) with Par. princeps, including eight species of Elrathiella. Parehmania differs from Elrathiella in its relatively deep S1 glabellar furrow isolated from the axial furrows, poorly developed S2 to S4 glabellar furrows and the pygidium has a more elliptical, diamond-shaped outline. As stated by Schwimmer (Reference Schwimmer1973, p. 288), the two genera are similar and Deiss’ (Reference Deiss1939) species of Elrathiella, which were (probably correctly) assigned to Par. princeps by Schwimmer. These glabellar characteristics are not typical of Par. tontoensis, Par. kwaguntensis, Par. nitida, or Par. sp. indet. reported from the Grand Canyon.
Elrathiella arizonaensis (Resser, Reference Resser1945) new combination
Elrathiella arizonaensis (Resser, Reference Resser1945) n. comb., from the Ehmaniella Biozone, Bright Angel Formation, Grand Canyon, Arizona; all specimens are preserved in shale from USNM loc. 44338A and are exfoliated. (1–5) Holotype, paratype, and associated material of Elrathiella arizonaensis (Resser, Reference Resser1945) n. comb.: (1) holotype cranidium USNM 794015a (Resser, Reference Resser1945, pl. 23, fig. 4, bottom); (2) associated cranidium USNM 794016a; (3) associated cranidium USNM 794016b; (4) associated poorly preserved shield USNM 794017, inverted image; (5) partial paratype pygidium USNM 794015b (Resser, Reference Resser1945, pl. 23, fig. 4, top). (6–29) Topotype material collected at the same time as type material: (6) small cranidium GRCA 8382a; (7) small, nearly complete shield GRCA 8361 with pygidium and frontal part of cranidium missing, silicon cast; (8) small cranidium GRCA 8382b; (9) pygidium GRCA 8389; (10) small cranidium GRCA 8387, silicon cast; (11) cranidium GRCA 8375b; (12) cranidium GRCA 8371c; (13) cranidium GRCA 8374; (14) cranidium GRCA 8383a; (15) cranidium GRCA 8378b, silicon cast; (16) cranidium GRCA 8375c; (17) cranidium GRCA 8373, silicon cast; (18) cranidium GRCA 8381; (19) cranidium GRCA 8378a, silicon cast; (20) cranidium GRCA 8383b, silicon cast; (21) cranidium GRCA 8372; (22) cranidium GRCA 8375a; (23) cranidium GRCA 8385, silicon cast; (24) cranidium GRCA 8376, silicon cast; (25) cranidium GRCA 8379; (26) cranidium GRCA 8371a; (27) cranidium GRCA 8370; (28) librigena GRCA 8382c; (29) librigena GRCA 8382d. Scales: (1–5, 11–29) 4-mm scale bar; (6–10) 2-mm scale bar.

Figure 51. Long description
The 29 panels are arranged as follows:
* Panels 1 through 5: Located at the top. 1 is a holotype cranidium. 2 and 3 are associated cranidia. 4 is a poorly preserved shield shown as an inverted image. 5 is a partial paratype pygidium.
* Panels 6 through 10: Middle-top section. 6 and 8 are small cranidia. 7 is a nearly complete shield silicon cast. 9 is a pygidium. 10 is a small cranidium silicon cast. A 2 mm scale bar is positioned between panels 6 and 7.
* Panels 11 through 16: Middle section. 11, 12, 13, 14, and 16 are cranidia. 15 is a cranidium silicon cast.
* Panels 17 through 20: Middle-bottom section. 17, 19, and 20 are cranidia silicon casts. 18 is a cranidium. A 4 mm scale bar is positioned between panels 18 and 19.
* Panels 21 through 24: Bottom section. 21, 22, and 25 are cranidia. 23 and 24 are cranidia silicon casts.
* Panels 25 through 29: Final row and right edge. 25, 26, and 27 are cranidia. 28 and 29 are librigenae located on the far right.
The fossils are preserved in shale, showing varying degrees of exfoliation. The cranidia typically feature a central glabella with surrounding fixed cheeks. The shields show segmented thoracic regions.
Holotype and associated material of “Spencia” tontoensis Resser, Reference Resser1945, and Parehmania tontoensis Resser, Reference Resser1945, here assigned to Elrathiella arizonaensis (Resser, Reference Resser1945) n. comb., from the Elrathiella subzone, Ehmaniella Biozone, Bright Angel Formation, Grand Canyon, Arizona; all specimens are internal molds in medium-grained sandstone from USNM loc. 74. (1–12) Spencia tontoensis: (1–3) holotype cranidium USNM 794166a (Resser, Reference Resser1945, pl. 24, fig. 27), dorsal, anterior, and lateral views; (4, 5) paratype cranidium USNM 794167 (Resser, Reference Resser1945, pl. 24, fig. 24), dorsal and anterior views; (6) paratype librigena USNM 794171 (Resser, Reference Resser1945, pl. 25, fig. 1); (7) paratype cranidium USNM 794172a (Resser, Reference Resser1945, pl. 24, fig. 26); (8) paratype cranidium USNM 794172b (Resser, Reference Resser1945, pl. 24, fig. 26); (9) cranidium USNM 794166b (Resser, Reference Resser1945, pl. 24, fig. 27); (10) paratype cranidium USNM 794169 [(Resser, Reference Resser1945, pl. 24, fig. 25); (11) paratype cranidium USNM 794170 (Resser, Reference Resser1945, pl. 24, fig. 23); (12) paratype cranidium USNM 794168 (Resser, Reference Resser1945, pl. 25, fig. 2). (13–22) Parehmania tontoensis: (13) cranidium USNM 794132a; (14–16) holotype cranidium USNM 794130 (Resser, Reference Resser1945, pl. 24, fig. 22), dorsal, anterior, and lateral views; (17, 18) cranidium 794131, dorsal and anterior views; (19) cranidium USNM 794132c; (20) cranidium USNM 794134; (21) cranidium USNM 794132b; (22) broken cranidium USNM 794133.

Figure 52. Long description
A collection of 22 photographs of trilobite fossils preserved as internal molds in sandstone.
* Panels 1 through 12 show Spencia tontoensis.
* Panels 1, 2, and 3 display the holotype cranidium U S N M 794166a in dorsal, anterior, and lateral views respectively, showing a rounded glabella and distinct ocular ridges.
* Panels 4 and 5 show paratype cranidium U S N M 794167 in dorsal and anterior views.
* Panel 6 shows a paratype librigena U S N M 794171.
* Panels 7 and 8 show paratype cranidia U S N M 794172a and 794172b.
* Panel 9 shows cranidium U S N M 794166b.
* Panels 10, 11, and 12 show paratype cranidia U S N M 794169, 794170, and 794168.
* Panels 13 through 22 show Parehmania tontoensis.
* Panel 13 is cranidium U S N M 794132a.
* Panels 14, 15, and 16 show the holotype cranidium U S N M 794130 in dorsal, anterior, and lateral views.
* Panels 17 and 18 show cranidium 794131 in dorsal and anterior views.
* Panels 19, 20, and 21 show cranidia U S N M 794132c, 794134, and 794132b.
* Panel 22 shows a large broken cranidium U S N M 794133.
A scale bar labeled 2 mm is located between panels 18 and 21. The fossils exhibit a granular texture characteristic of medium-grained sandstone molds.
New specimens of Elrathiella arizonaensis (Resser, Reference Resser1945) n. comb., from the Elrathiella subzone, Ehmaniella Biozone, Bright Angel Formation, Grand Canyon, Arizona. (1–15) specimens preserved in fine-grained sandstone as internal molds: (1, 2) cranidium GRCA 125153 GRCA loc. 355-4, anterior and dorsal views; (3) cranidium GRCA 125151 from GRCA loc. 355-4; (4) questionably assigned cranidium GRCA 125154 from GRCA loc. 355-4; (5) cranidium GRCA 125152 from GRCA loc. 355-4; (6–8) cranidium GRCA 125155b from GRCA loc. 355-4, dorsal, lateral, and anterior views; (9) cranidium GRCA 125145 from GRCA loc. 355-4; (10) librigena GRCA 125143 from GRCA loc. 355-1; (11) librigena cranidium GRCA 125143 from GRCA loc. 355-4; (12) small cranidium GRCA 125157 from GRCA loc. 355-1; (13–15) pygidium GRCA 125144 from GRCA loc. 355-1, dorsal, lateral, and posterior views. (16–20) Shale specimens from GRCA loc. 376: (16) cranidium GRCA 125356, coated with calcite; (17) internal mold of cranidium GRCA 125155a; (18) cranidium GRCA 125357, partially coated with calcite; (19) internal mold of cranidium GRCA 125361; (20) internal mold of pygidium GRCA 125358. (21–25) Tectonically distorted shale specimens questionably assigned to the species from GRCA loc. 341-4, 1 m below the Elves’ Chasm tongue: (21) partial cranidium GRCA 125185, latex cast; (22) partial cranidium GRCA 125186; (23) partial cranidium GRCA 125187; (24) partial pygidium GRCA 125188, (25) partial cranidium GRCA 125189. Scales: (1–11, 16–19, 21–24) 4-mm scale bar; (12–15, 20, 25) 2-mm scale bar.

Figure 53. Long description
The collection is organized into five rows of fossil specimens.
Row 1 (Panels 1 to 5): Features five cranidia preserved in fine-grained sandstone. Panel 1 shows a dorsal view, while Panel 2 provides an anterior view of the same specimen. Panels 3, 4, and 5 show dorsal views of individual cranidia with varying degrees of preservation.
Row 2 (Panels 6 to 11): Panels 6, 7, and 8 show dorsal, lateral, and anterior views of a single cranidium. Panel 9 is a dorsal cranidium. Panels 10 and 11 display librigenae (free cheeks). A 4-mm scale bar is positioned below Panel 2.
Row 3 (Panels 12 to 15): Panel 12 shows a small cranidium. Panels 13, 14, and 15 provide dorsal, lateral, and posterior views of a pygidium (tail piece). A 2-mm scale bar is located above Panel 14.
Row 4 (Panels 16 to 20): Displays shale specimens. Panels 16, 17, 18, and 19 show cranidia, some with calcite coating or as internal molds. Panel 20 is an internal mold of a pygidium.
Row 5 (Panels 21 to 25): Shows tectonically distorted specimens. Panels 21, 22, 23, and 25 are partial cranidia, including a latex cast in Panel 21. Panel 24 shows a partial pygidium.
Anatomical features across the cranidia include a distinct glabella, fixed cheeks, and palpebral lobes. The pygidia show segmented axial rings and pleural regions.
Reference Resser1945 Ehmaniella arizonaensis Resser, p. 207, pl. 23, fig. 4.
?Reference Resser1945 Parehmania tontoensis Resser, p. 207, pl. 24, fig. 22.
?Reference Resser1945 Spencia tontoensis; Resser, p. 213, pl. 24, figs. 23–30, pl. 25, figs. 1, 2.
Holotype
Cranidium USNM 794015a (Fig. 51.1) from USNM loc. 44338A, Bright Angel Formation, Bright Angel Trail, Grand Canyon, Arizona.
Paratypes
Shield USNM 794017; cranidia USNM 794016a, b; and pygidium USNM 794015b from USNM loc. 44338A, Bright Angel Formation, Bright Angel Trail, Grand Canyon, Arizona.
Occurrence
Elrathiella subzone, Ehmaniella Biozone, Bright Angel Formation, Bright Angel Trail, River Mile 143, ?Blacktail Canyon, River Mile 50, Nankoweap Valley, Nankoweap Canyon and Malgosa Canyon, Grand Canyon, Arizona.
Emended description
Cranidium of moderate size, length 7.4 ± 2.3 mm (n = 24); subtrapezoidal, moderately convexity (sag. and trans.); anterior margin moderately curved medially, straighter laterally; posterior margin (excluding occipital ring) directed laterally. Anterior branches of facial sutures moderately divergent by 15 ± 5° to anterior border, convergent to anterior margin; posterior branches moderately divergent at 40 ± 5°. Glabella moderately elongate, moderate convexity (sag. and trans.), length 76 ± 2% cranidial length; width 32 ± 3% cranidial width; moderately tapered, width at anterior end 74 ± 6% of posterior glabellar width; frontal lobe moderately to bluntly rounded. Axial furrows moderately deep, with slight constriction at the L2; preglabellar furrow moderately deep. Lateral glabellar furrows moderately shallow, S1 bifurcated, directed moderately posterior; S2 bifurcated directed transverse to slightly posteriorly; S3 shallow directed slightly anterior; S4 very shallow, directed anteriorly. Occipital ring slightly elevated above rest of glabella, moderately convex; length 18 ± 2% glabellar length; posterior margin moderately convex posteriorly, evenly curved; small occipital node absent. SO slightly curved anteriorly; deepest laterally forming deeper pits next to axial furrows, shallow depth medially. Frontal area subequally divided; length 24 ± 2% cranidial length. Preglabellar field slightly convex, down-sloping, length 9 ± 2% cranidial length, 40 ± 6% frontal area length. Anterior border slightly convex, level, flat to slightly arched dorsally, length 14 ± 2% cranidial length, slightly tapering laterally, no or slight medial inbend or swelling. Anterior border furrow narrow and of uniform and moderate depth, deeper than axial furrows. Fixigena moderately convex, level to slightly down-sloping, anterior area steeply down-sloping anteriorly, width 53 ± 7% of posterior glabellar width. Palpebral lobes nearly straight, level, moderately narrow and moderately long, length 35 ± 4% glabellar length; anterior margin located transversely opposite of S3 or L3. Ocular ridge weakly to moderately defined, very slightly curved anteriorly, directed moderately posterolateral from glabella at 69 ± 5° to sagittal axis. Posterior area of fixigena triangular, terminated with sharp corner; length 30 ± 4% glabellar length; width 73 ± 6% glabella length.
Librigenae moderate size, length approximately 5.2–13.2 mm (n = 3); moderately wide at about 30–55% length without spine; lateral margin moderately curved. Genal field slightly convex. Border moderately convex, uniform in width, width about 25–30% librigenal width; border furrows shallow and do not persist into the librigenal spine. Librigenal spine moderate length, about 45–60% librigenal length based on incomplete spine; directed posterolaterally.
Rostral plate and hypostome unknown.
Thorax with 12–15 segments (n = 2; 6.6 mm, 13.6 mm), tapering towards small pygidium. Pleurae width approximately 80% of axial ring width, inner portion oriented transversely, directed slightly posterolaterally and downwards? at weakly developed fulcrum. Anterior and posterior pleural bands moderately convex and uniform in length. Pleural furrow moderately deep and of uniform depth, narrows distally. Pleural spine very short, termination pointed.
Pygidium small, length 3.8–4.0 mm (n = 2); oval, length approximately 50% width; margin smooth, anterior margin nearly straight and directed slightly posterolaterally; anterolateral corners sharply rounded, transversely opposite near terminus of axis; no postaxial notch; moderately convex (sag.). Axis moderately tapered, mid-width 80–90% anterior width, anterior width approximately 40% pygidial width; length approximately 80% pygidial length; two or possibly three axial rings, moderately convex; terminal piece small, rounded posteriorly; axial ring furrows shallow. Pleural regions convex, down-sloping; most anterior pleural furrow narrow and moderate depth, extending to near margin; other pleural furrows very shallow. Border narrow and poorly defined.
Smooth surface on the entire exoskeleton. Faint genal caeca on preglabellar regions of larger specimens.
Remarks
The emended description above is based primarily on the type and topotype specimens that are preserved in shale. Convexity of different features is based on specimens preserved in sandstone (Figs. 52, 53); but due to the tentative assignment of these specimens, these convexities need to be considered as questionable.
Resser (Reference Resser1945) characterized this species by its long, tapering glabella, small eyes, wide fixigenas, and narrow occipital ring, but only compared it to Ehmaniella hebes and Eh. basilica. The morphological range of El. arizonensis derived from the two relatively large cranidia and the single mostly covered pygidium illustrated by Resser (Reference Resser1945) is inadequate for comparison to other similar taxa. Additional topotype specimens in the USNM type collection (2 specimens) and from the GRCA collections (21 cranidia, 2 librigena, and 1 pygidium) and GRCA loc. 376 (5 cranidia, 1 pygidium) (Fig. 51.6–51.29) provide additional information on the morphological range of the species. One disadvantage of these and the type specimens is that they are preserved in shale and are thus flattened whereas most other similar species from the Bright Angel Formation are preserved in sandstone. Compression of specimens would distort some linear measurements, with significant changes in the frontal area length and division, position of the palpebral lobes and glabellar tapering (FAS unpublished study on Amecephalus arrojosensis [Lochman, Reference Lochman1948]). However, given these difficulties, it appears that cranidial features of Parehmania tontoensis (Fig. 52.13–52.22), Spencia tontoensis (Fig. 52.11), and sandstone specimens (Fig. 53) assigned to El. arizonaensis n. comb. have significant overlap with the shale specimens of El. arizonaensis n. comb. both in size and morphology (Fig. 50). This overlap suggests that these species/specimens should be placed or questionably placed into El. arizonaensis n. comb.
The types of Par. tontoensis (Fig. 52.13–52.22) are preserved in sandstone and are similar to El. arizonaensis n. comb. in the tapering of the glabella with a bluntly rounded frontal lobe, more posterior placed palpebral lobes, relatively narrow intraocular width of the fixigena and the curvature of the anterior border in the larger specimens. Differences are generally between the holotype of El. tontoensis in its more posterior position palpebral lobes, and stronger curved anterior border; the specimens fall entirely within the variation of El. arizonaensis n. comb. (Fig. 50).
The types of S. tontoensis and Par. tontoensis are from Nankoweap Valley from the same horizon (USNM loc. 74; Resser, Reference Resser1945) and matrix type. Resser stated that this species is characterized by its extremely small preglabellar area, the inflated fixigenas, the truncated glabella, and abruptly steepened anterior slope in lateral profile; however, he only compared it to S. typicalis. Specimens in hand relatively small (PC1; Fig. 50); overlap with both El. arizonaensis n. comb. and Par. tontoensis.
One morphological aspect that could be used to separate species is the curvature of the cranidial anterior border, which is measured here as the degrees of the anterior border furrow from the lateral margins to the sagittal inflection (Fig. 49). For the type and topotype material of El. arizonaensis n. comb., this angle is 158 ± 6° (n = 23). The majority of specimens of S. tontoensis (n = 5) and Par. tontoensis (n = 4) fall within this range, with only one specimen of each taxon falling outside of that range. In contrast, only two specimens of the sandstone specimens of El. arizonaensis n. comb. (n = 5) and two specimens of shale specimens from GRCA loc. 376 (n = 5) fall within this range. It is possible that more than one species occurs at these locations, but this assumption cannot be supported without additional material. Taxa assigned by Resser (Reference Resser1945) to Pac. moorei (= Proehmaniella moorei; n = 2), Pac. sp. (= El. fonticola; n = 2), Parehmania sp. (= El. fonticola; n = 3) occur within the field of El. arizonaensis n. comb. type and topotype material linear features (Fig.50.3) but fall outside this range when looking at the curvature of the anterior border furrow. Parehmania nitida (= Eh. nitida n. comb.; n = 2), El. decora (n = 3), Par. kwaguntensis (= Eh. kwaguntensis n. comb.; n = 6), and Pac. fonticola (= El. fonticola; n = 1) fall outside the range of El. arizonaensis n. comb. linear measures (Fig. 50.3); however, the first two taxa overlap in anterior border furrow curvature, whereas the latter three do not overlap within the 158 ± 6° topotype range (Fig. 49).
Elrathiella decora Resser, Reference Resser1945
Elrathiella decora Resser, Reference Resser1945, from the Elrathiella subzone, Ehmaniella Biozone, Grand Canyon, Arizona. (1–3) Type specimens from the Bright Angel Formation?, preserved as internal molds in a piece of fine-grained sandstone float, from USNM loc. 44346: (1) paratype cranidium USNM 794035c (Resser, Reference Resser1945, pl. 24, fig. 1, upper left); (2) paratype cranidium USNM 794035b (Resser, Reference Resser1945, pl. 24, fig. 1, upper right), latex cast; (3) holotype cranidium USNM 794035a (Resser, Reference Resser1945, pl. 24, fig. 1, lower right), latex cast. (4–8) Elrathiella cf. E. decora Resser, Reference Resser1945, from the Elrathiella subzone, Ehmaniella Biozone, Spencer Canyon Member, Muav Formation, GRCA loc. 393, Grand Canyon, Arizona, preserved in limestone and mostly exfoliated: (4) librigena GRCA 125546; (5) partial cranidium GRCA 125551; (6) cranidium GRCA 125550; (7) librigena GRCA 125547; (8) librigena GRCA 125548. Scales: (1–3) 4-mm scale bar, (4–8) 2-mm scale bar.

Figure 54. Long description
The figure consists of eight panels labeled 1 through 8.
Panels 1 through 3 are grouped at the top and center, associated with a 4 millimeter scale bar.
* Panel 1 shows paratype cranidium U S N M 794035c, a fossilized head shield with a rounded central glabella and wide fixed cheeks.
* Panel 2 displays paratype cranidium U S N M 794035b, a latex cast showing similar anatomical features with higher surface detail.
* Panel 3 features holotype cranidium U S N M 794035a, a larger latex cast showing the complete semi-circular outline of the head shield.
Panels 4 through 8 are arranged at the bottom and right, associated with a 2 millimeter scale bar.
* Panel 4 shows librigena G R C A 125546, a curved lateral part of the head shield.
* Panel 5 shows partial cranidium G R C A 125551, embedded in a rocky matrix.
* Panel 6 shows cranidium G R C A 125550, viewed from a dorsal perspective showing the central lobe.
* Panel 7 shows librigena G R C A 125547, a narrow, elongated fossil fragment.
* Panel 8 shows librigena G R C A 125548, showing the textured surface of the fossilized shell.
Reference Resser1945 Elrathiella decora Resser, p. 209–210, pl. 24, fig.1.
Reference Sundberg, Cooper and Stevens1991 Elrathiella decora; Sundberg, fig. 2.17.
Reference Sundberg1994 Elrathiella decora; Sundberg, p. 61–63, fig. 38.1, 38.2, 38.4, 38.5 [fig. 38.3 possible E. cf. E. decora].
Holotype
Cranidium USNM 794035a (Fig. 54.3) from USNM loc. 44346, Bright Angel Formation? (float), Iceberg Canyon, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794035b and 794035c from USNM loc. 44346, Bright Angel Formation? (float), Iceberg Canyon, Grand Canyon, Arizona.
Occurrence
Elrathiella subzone, Ehmaniella Biozone, Bright Angel Formation?, Iceberg Canyon and Muav Formation, Rampart Cave, Grand Canyon. Elrathiella subzone, Ehmaniella Biozone, Whirlwind Formation, Drum Mountains and House Range, Utah (Sundberg, Reference Sundberg1994).
Remarks
Sundberg (Reference Sundberg1994) recognized El. decora Resser, Reference Resser1945, from the Elrathiella subzone, Ehmaniella Biozone of the Whirlwind Formation in the Drum Mountains and House Range, Utah. Unfortunately, the type specimens of El. decora from the Grand Canyon were found in a float sample of fine-grained sandstone and are relatively large (10.5 to 13.4 mm in cranidial lengths). The larger specimens from the Whirlwind Formation are very similar to the Grand Canyon specimens and were assigned to the species by Sundberg (Reference Sundberg1994). Partial cranidia and librigenae (Fig. 54.4–54.8) were found from the Spencer Canyon Member, Muav Formation, near Rampart Cave. These specimens are also similar to the limestone specimens (Sundberg, Reference Sundberg1994, fig. 38.5) found in the Whirlwind Formation, but are left in open nomenclature (cf.) due to the fragmented nature of the cranidia. Both assemblages have narrow intraocular fixigena typical of the species.
Elrathiella fonticola (Resser, Reference Resser1945)
Elrathiella fonticola (Resser, Reference Resser1945) from the Elrathiella subzone, Ehmaniella Biozone, Bright Angel Formation, Grand Canyon, Arizona; all specimens are internal molds in medium- to fine-grained sandstones from USNM loc. 44337. (1, 2) Pachyaspis fonticola Resser Reference Resser1945, holotype cranidium USNM 794087 (Resser, Reference Resser1945, pl. 24, figs. 31, 32), dorsal and lateral views; (3–11) mostly unfigured specimens associated with the type collection of Parehmania sp. of Resser, Reference Resser1945, reassigned herein: (3) partial pygidium 794126, (4–6) cranidium 794124 (Resser, Reference Resser1945, pl. 25, figs. 3, 4), dorsal, anterior, and lateral views; (7) cranidium 794123; (8) cranidium 794129; (9) small cranidium 794125; (10) small cranidium 794127; (11) small cranidium 794128. Scales: (1, 2, 4–8) 4-mm scale bar; (3, 9–11) 2-mm scale bar.

Figure 55. Long description
A collection of eleven fossil specimens preserved as internal molds in sandstone.
Top row:
1. Pachyaspis fonticola holotype cranidium in dorsal view showing a rounded glabella and wide fixed cheeks.
2. Lateral view of the same holotype showing the profile of the glabella.
3. A small partial pygidium with visible segmentation.
4. Cranidium 794124 in dorsal view.
5. Anterior view of cranidium 794124 showing the frontal margin.
6. Lateral view of cranidium 794124.
Bottom row:
7. Cranidium 794123 in dorsal view.
8. Cranidium 794129 in dorsal view.
9. Small cranidium 794125.
10. Small cranidium 794127.
11. Small cranidium 794128.
Scale bars:
A 4 mm scale bar is located to the right of panel 6, corresponding to specimens 1, 2, and 4 through 8.
A 2 mm scale bar is located above panel 11, corresponding to specimens 3 and 9 through 11.
Reference Resser1945 Pachyaspis fonticola Resser, p. 211, pl. 24, figs. 31, 32.
Reference Resser1945 Parehmania sp. undet. Resser, p. 209, p1. 25, figs. 3,4.
Holotype
Cranidium USNM 794087 (Fig. 55.1, 55.2) from USNM loc. 44337, Bright Angel Formation, Nankoweap Valley, Grand Canyon, Arizona.
Occurrence
Elrathiella subzone, Ehmaniella Biozone, Bright Angel Formation, Chuar Valley and Bright Angel Trail, Grand Canyon, Arizona.
Remarks
Resser assigned a single cranidium in a fine-grained sandstone matrix (Fig. 55.1, 55.2) to this new species. A distinctive feature of this cranidium is its prominent, nearly parallel sided glabella with a rounded frontal lobe. Specimens of Parehmania sp. indet. from a limonitic sandstone (Fig. 55.3–55.11) are here assigned to El. fonticola given their overall similarity in glabellar features. The single fragment of pygidium (Fig. 55.3) is also similar to other pygidia of Elrathiella (Sundberg, Reference Sundberg1994; this paper), but is mostly covered by rock, which limits an accurate determination. Elrathiella fonticola is most similar to El. eurthyopsis Sundberg, Reference Sundberg1994, in the glabellar shape and curved anterior border. The two differ in the latter having granular ornamentation, longer anterior border, and relatively wider intraocular fixigenal width.
Resser (Reference Resser1945, p. 210) named two new species, Pac. moorei and Pac. fonticola, and presented information related to Pac. sp. undet. and Pachyaspis with the quote “It seems to be fully characteristic of the genus, averaging a little smaller than the genotype” (Resser, Reference Resser1939b). However, the differences between these species (n = 5 measured specimens) and the type species (n = 6 measured specimens from Resser, Reference Resser1939b, and Campbell, Reference Campbell1974) include: (1) cranidium with a more rectangular shape (cranidial length 62 ± 5% vs. 72 ± 4% cranidial width), divergent anterior facial sutures (13 ± 10° vs. −12 ± 7°), wider interocular fixigena (59 ± 10% vs. 48 ± 6% of glabellar width), and posterior of fixigena wider (68 ± 3% vs. 57 ± 5% glabellar length); (2) glabella with less tapering (74 ± 4% vs. 68 ± 5% glabellar base width), relatively narrower glabellar base (33 ± 2% vs 39 ± 4% of cranidial width), and longer occipital ring (21 ± 4% vs. 16 ± 2% of glabellar length); (3) wider (tr.) palpebral lobe (24–37% [n = 3] vs. 23 ± 7% palpebral lobe length); and (4) pygidium (P. sp. indet. and Parehmania sp. indet. vs. specimens of Pac. typicalis illustrated by Campbell, Reference Campbell1974) relatively transversely narrow with anterior lateral corners at level of end of axis versus suboval outline with corners at level near mid-axial length and strongly developed anterior pleural bands versus poorly developed anterior pleural bands posterior of anterior pleural set.
The PCA results (Fig. 50) illustrate that Pac. fonticola, Pac. sp. undet, Pac. moorei, and Parehmania sp. indet. fall within the range of other taxa assigned to Elrathiella (e.g., El. arizonaensis n. comb.), although Pac. fonticola falls outside this field for PC2 versus PC3 (Fig. 50.3). However, the curvature of the anterior border furrow of Pac. fonticola, Pac. moorei, and Par. sp. indet. occur below the mean and standard deviation of El. arizonaensis n. comb. and form a linear trend around 145° (Fig. 49). One specimen of Pac. sp. indet. falls within this mean and standard deviation, but the other is outside that range.
Pachyaspis moorei and questionably Pac. sp. indet. are reassigned to Pro. moorei based on overall glabellar shape and relatively short palpebral lobes.
Elrathiella fonticola is from USNM loc. 44337 “in a purple layer overlying the green layer of impure sandy shale, containing considerable glauconite and iron oxide as well as mica flakes” (Resser, Reference Resser1945, p. 211); other samples are from this locality in the McKee Quarry, which contains fauna of the Glossopleura walcotti Biozone. This may suggest an unconformity at the top of the McKee Quarry. Parehmania sp. indet. is from USNM loc. 73a in Lone Canyon and co-occurs with Pac. sp. indet. (Ehmaniella Biozone) and members of the Glossopleura walcotti Biozone in a “dark-red sandy limonitic rock” (Resser, Reference Resser1945, p. 211). This mixture of biozone indicators may represent stratigraphic mixing of samples from this locality.
Elrathiella? insueta Resser, Reference Resser1945
Elrathiella? insueta Resser, Reference Resser1945, from the Ehmaniella subzone, Ehmaniella Biozone, Bright Angel Formation or Muav Formation, Grand Canyon, Arizona; all specimens are internal molds in a siltstone from USNM locality 74a. (1–3) Holotype cranidium USNM 794185 (Resser, Reference Resser1945, pl. 25, figs. 7, 8), dorsal, anterior, and lateral views; (4–6) paratype cranidium USNM 794186 (Resser, Reference Resser1945, pl. 25, figs. 5, 6), dorsal, anterior, and lateral views; (7) additional cranidium USNM 794040; (8) additional cranidium USNM 794039; (9) additional cranidium USNM 794038.

Figure 56. Long description
The multi-panel photo contains nine individual views of trilobite cranidia fossils.
* Panels 1 through 3 show the holotype U S N M 794185. Panel 1 is a dorsal view showing the central glabella and surrounding fixed cheeks. Panel 2 is an anterior view showing the frontal profile. Panel 3 is a lateral view showing the curvature of the head shield.
* Panels 4 through 6 show the paratype U S N M 794186. Panel 4 is a dorsal view with a well-preserved right palpebral lobe. Panel 5 is an anterior view highlighting the convex glabella. Panel 6 is a lateral view.
* Panel 7 shows an additional cranidium U S N M 794040 in dorsal view.
* Panel 8 shows an additional cranidium U S N M 794039 in dorsal view.
* Panel 9 shows an additional cranidium U S N M 794038 in dorsal view.
A black scale bar in the bottom right corner represents 5 millimeters. All specimens are preserved as internal molds within a textured siltstone matrix.
Reference Resser1945 Elrathiella? insueta Resser, p. 211, pl. 25, figs. 5–8.
Holotype
Cranidium USNM 794185 (Fig. 56.1–56.3) from USNM loc. 74a, Bright Angel Formation or Muav Formation, Nankoweap Valley, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794186–794040 from USNM loc. 74a, Bright Angel Formation or Muav Formation, Nankoweap Valley, Grand Canyon, Arizona.
Occurrence
Elrathiella subzone, Ehmaniella Biozone Bright Angel Formation or Muav Formation, Nankoweap Valley, Grand Canyon, Arizona.
Remarks
Resser (Reference Resser1945) reported the type material of both El.? insueta and Eh. hebes from the same locality, with the former in a silty horizon and the later in a limestone horizon. Key differences between the two species are that most specimens of El.? insueta are tectonically deformed (Fig. 56), larger, and poorly preserved. Despite these problems, there are strong similarities between the two species: (1) tapering glabella about 75% of cranidial length, (2) bluntly rounded frontal lobe, (3) nearly effaced lateral glabellar furrows, (4) long (sag) anterior border and preglabellar area, (5) anterior border curvature, and (6) convexity of the cranidium and the intraocular regions. It is possible that both species represent a single taxon, but due to the poor preservation of El.? insueta, the two species were not synonymized. At present this species should be considered nomen dubium.
Genus Proehmaniella Sundberg, Reference Sundberg1994
Type species
Ehmaniella basilica Resser, Reference Resser1945, from the lower Topazan (Wuliuan Stage), Sanup Plateau Member, Muav Formation, Peach Springs Canyon, Grand Canyon, Arizona.
Proehmaniella basilica (Resser, Reference Resser1945)
Proehmaniella basilica (Resser, Reference Resser1945), from the Proehmaniella subzone, Ehmaniella Biozone, above the Rampart Cave Member, Muav Formation, Grand Canyon, Arizona; all specimens are preserved in shale, without testate material, from USNM loc. 38i. (1) Holotype shield USNM 794018a (Resser, Reference Resser1945, pl. 23, fig. 10, left); (2, 3) paratype shield and close-up of pygidium, USNM 794019d (Resser, Reference Resser1945, pl. 23, fig. 11, top); (4) paratype cranidium USNM 794024; (5) paratype cranidium USNM 794019e (Resser, Reference Resser1945, pl. 23, fig. 11, top); (6) paratype cranidium USNM 794019b (Resser, Reference Resser1945, pl. 23, fig. 11, middle center); (7) paratype thorax and pygidium USNM 794021; (8) paratype thorax and pygidium USNM 794023; (9) paratype cranidium USNM 794018b (Resser, Reference Resser1945, pl. 23, fig. 10, right); (10) paratype cranidia USNM 794019f–h (top to bottom) (Resser, Reference Resser1945, pl. 23, fig. 11, top center, counterpart; (11, 12) paratype cranidium USNM 794019c with close-up of possible granulation on the anterior border; (13) paratype cranidium USNM 794019a (Resser, Reference Resser1945, pl. 23, fig. 11, middle center); (14) paratype librigena USNM 794194; (15) paratype librigena USNM 794195; (16) paratype cranidium USNM 794026 (Resser, Reference Resser1945, pl. 23, fig. 12), reported by Resser as being testate.

Figure 57. Long description
A collection of 16 fossil specimens and one scale bar. A horizontal black bar labeled 2 m m is located in the upper right quadrant.
* Panel 1: A complete holotype shield U S N M 794018a showing a rounded cephalon, segmented thorax, and small pygidium.
* Panel 2: A paratype shield U S N M 794019d with a wide thorax.
* Panel 3: A close-up of the pygidium from specimen 2.
* Panel 4: A paratype cranidium U S N M 794024 showing the central head region.
* Panel 5: A paratype cranidium U S N M 794019e.
* Panel 6: A paratype cranidium U S N M 794019b.
* Panel 7: A paratype thorax and pygidium U S N M 794021.
* Panel 8: A paratype thorax and pygidium U S N M 794023.
* Panel 9: A large paratype cranidium U S N M 794018b.
* Panel 10: Three paratype cranidia U S N M 794019f through h arranged vertically and intersected by a diagonal crack in the shale.
* Panel 11: A paratype cranidium U S N M 794019c.
* Panel 12: A high-magnification close-up of the anterior border of specimen 11 showing fine granulation.
* Panel 13: A paratype cranidium U S N M 794019a.
* Panel 14: A paratype librigena U S N M 794194, representing the free cheek.
* Panel 15: A paratype librigena U S N M 794195.
* Panel 16: A paratype cranidium U S N M 794026 with a well-defined glabella and wide anterior border.
Reference Resser1945 Ehmaniella basilica Resser, p. 206, pl. 20, figs. 19, 20; pl. 23, figs. 10–13.
Reference Sundberg, Cooper and Stevens1991 “Ehmaniella” basilica; Sundberg, fig. 2.20.
Reference Sundberg1994 Proehmaniella basilica; Sundberg, p. 73–75, fig. 45.
Reference Sundberg2005 Proehmaniella basilica; Sundberg, p. 70, fig. 6.2.
Holotype
Shield USNM 794018a (Fig. 57.1) from USNM loc. 38i, Sanup Plateau Member, Muav Formation, Peach Springs Canyon, Grand Canyon, Arizona.
Paratypes
Partial shields USNM 794019d, 794021, 794023, and 794189; cranidia USNM 794018b, 794019a–c, e–h, 794020, 794022, 794024, 794025, 794026, and 794027; and librigenae USNM 794194 and 794195 from USNM loc. 38i, top of Rampart Cave Member, Muav Formation, Peach Springs Canyon, Grand Canyon, Arizona.
Occurrence
Proehmaniella subzone, Ehmaniella Biozone Sanup Plateau Member, Muav Formation, Diamond Creek, Grand Canyon, Arizona. Proehmaniella subzone, Ehmaniella Biozone, Chisholm Formation, House Range and Drum Mountains, Utah; Eldorado Formation, northern Eagan Range, Nevada (Sundberg, Reference Sundberg1994).
Remarks
The collection that was studied includes material from the type locality and type specimens. In addition to some of the specimens illustrated by Resser (Reference Resser1945), articulated specimens and librigena are illustrated. A full description of the species was provided by Sundberg (Reference Sundberg1994).
Sundberg (Reference Sundberg1994) synonymized Pac. moorei with Pro. basilica, but reinvestigation of the type material indicates that they may represent two different species within the same genus, Proehmaniella. The cranidia of the two taxa differ in Pro. basilica having a more subsquare outline, longer anterior border, more rounded frontal lobe, and smooth exoskeleton. However, differences between these two species may be the result of ontogeny (Pro. moorei tend to be smaller) and preservation in shale versus limestone. Compression in shale tends to move the lateral margins of the cranidium abaxially (laterally) and anteriorly, with increasing intensity anteriorly (FAS, personal observation). This would generate differences in cranidial shape and possibly pygidial shape. When compared to limestone specimens, the compression in shale would generate small lengths in the occipital ring (E, −11%) distance of the palpebral lobe position from the top of the glabella (D9, −21%) and frontal lobe width (K6, −9%); wider lengths in cranidial width (L, 8%), and anterior cranidial width (J1, 5%); and ocular ridge angle (ER, 6%), anterior border curvature (AB, 1%), anterior branch of facial suture (AS, −4%), and posterior branch (PS, −3%) (see Sundberg and Webster, 2022, for abbreviations and how characters are measured). As a result, the ratios of cranidial length/cranidial width, glabellar width/cranidial width, glabellar tapering, occipital ring length of glabellar length, vertical position of the palpebral lobes relative to glabellar length, anterior portion of librigena width relative to glabellar length, and posterior margin of the cranidium relative to glabellar length changes with compression of specimens in shale. Application of these percentage changes to the database of Pro. moorei shifts the data points so that Pro. moorei has greater overlap with Pro. basilica. In summary, the measured aspect difference between the two species can be the result of compaction.
Another difference between the two species is the “absence” of a fine granular surface on Pro. basilica and its presence on Pro. moorei. However, this may also be the result of preservational differences between shale and limestone. Specimens of Pro. basilica have a roughen surface that may represent the fine granules (see enlarged anterior border area in Fig. 57.12), but again the preservation is not sufficient to clearly demonstrate their absence. As a result, we prefer to recognize two different species for the present.
Proehmaniella moorei (Resser, Reference Resser1945)
Proehmaniella moorei (Resser, Reference Resser1945) from the Proehmaniella subzone, Ehmaniella Biozone. (1–5) Type specimens of Pachyaspis moori Resser, Reference Resser1945, in limestone from USNM loc. 38h, from the Elves Chasm tongue, Muav Formation, Grand Canyon, Arizona: (1–3) holotype cranidium USNM 794088 (Resser, Reference Resser1945, pl. 23, fig. 9), dorsal, anterior, and lateral views; (4) paratype cranidium USNM 794089 (Resser, Reference Resser1945, pl. 23, fig. 8); (5) paratype cranidium USNM 794090 (Resser, Reference Resser1945, pl. 23, fig. 7). (6–12) Figured specimens of Pachyaspis sp. undet of Resser, Reference Resser1945, in sandstone from USNM loc. 73a, Bright Angel Formation, preserved in sandstone assigned to P. moorei: (6, 7) large cranidium USNM 794100 (Resser, Reference Resser1945, pl. 24, fig. 17) questionably assigned to the species, dorsal and lateral views; (8) cranidium USNM 794097 (Resser, Reference Resser1945, pl. 24, fig. 18); (9) pygidium USNM 794095 (Resser, Reference Resser1945, pl. 24, fig. 20); (10) pygidium USNM 794096 (Resser, Reference Resser1945, pl. 24, fig. 19); (11) cranidium USNM 794099 (Resser, Reference Resser1945, pl. 24, fig. 16); (12) cranidium USNM 794092 (Resser, Reference Resser1945, pl. 24, fig. 21). Scales (1–5) 1-mm scale bar; (6–8) scale bars in pictures represent 2-mm; (9–12) 2-mm scale bar.

Figure 58. Long description
A collection of 12 fossil photographs arranged in three rows.
Top row:
Panel 1 shows a dorsal view of a holotype cranidium with a smooth, rounded glabella and wide fixed cheeks.
Panel 2 shows the same specimen in an anterior view, highlighting the convex profile.
Panel 3 shows a lateral view of the holotype.
Panel 4 is a smaller paratype cranidium.
Middle row:
Panel 5 is another paratype cranidium in dorsal view. A 1 mm scale bar is positioned to the right of panel 5.
Panel 6 shows a larger, more textured cranidium in dorsal view with a 2 mm scale bar.
Panel 7 is a lateral view of the specimen in panel 6.
Panel 8 shows a highly textured cranidium in dorsal view.
Bottom row:
Panel 9 and 10 show small, rounded pygidia with visible segmentation.
Panel 11 and 12 show additional cranidia specimens with varying degrees of preservation and surface texture.
A 2 mm scale bar is located between panels 6 and 9. All specimens are embedded in a rocky matrix, with U S N M specimens 1 through 5 preserved in limestone and 6 through 12 preserved in sandstone.
Reference Resser1945 Pachyaspis moorei Resser, p. 210–211, pl. 23, figs. 7–9.
?Reference Resser1945 Pachyaspis sp. undet. Resser, p. 211–212, pl. 24, figs. 16–21.
Holotype
Cranidium USNM 794088 (Fig. 58.1–58.3) from USNM loc. 38h, Elves Chasm tongue of Muav Formation, Havasu Creek, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794089–794091from USNM loc. 38h, Elves Chasm tongue of Muav Formation, Havasu Creek, Grand Canyon, Arizona.
Emended diagnosis
Cranidium with anterior border moderately curved, moderately convex; moderately long preglabellar area, moderately intraocular fixigena, moderately posterolaterally directed ocular ridges; very fine granular ornamentation. Pygidium alate, blunt axis, with 2 axial rings; anterior pleural bands moderately developed; granules not apparent.
Occurrence
Proehmaniella subzone, Ehmaniella Biozone, Bright Angel Formation, Havasu Creek and Chuar Valley, Grand Canyon, Arizona.
Remarks
Cranidia and pygidia preserved in sandstone reported by Resser (Reference Resser1945) as Pac. sp. indet. from Chuar Valley are included within Pro. moorei based on fixigenal width and glabellar outline, and general pygidial features. A single large specimen (Figs. 58.6, 58.7) is questionably assigned to the species because of the narrower fixigena and is more similar to El. fonticola, which is also reported from USNM loc. 73a.
Genus Trachycheilus Resser, Reference Resser1945
Type species
Trachycheilus typicale Resser, Reference Resser1945, from the upper Topazan (Wuliuan Stage), Bright Angel Formation or Muav Formation, lower part of Kwagunt Valley, Grand Canyon, Arizona.
Remarks
Sundberg (Reference Sundberg1994) revised the generic diagnosis to include additional species from the Whirlwind Formation of Utah. Species within Trachycheilus are recognized based on characteristics of the cranidia (shape), glabella (axial and lateral glabellar furrow depth, convexity, and frontal lobe rounding), exoskeleton ornamentation (granular vs. smooth), anterior border width of frontal area, anterior inbend, and pygidial features (shape and convexity of anterior and posterior pleural bands).
Trachycheilus typicale Resser, Reference Resser1945
Trachycheilus typicale Resser, Reference Resser1945, from USNM loc. 73b, Ehmaniella subzone, Ehmaniella Biozone, upper part of the Bright Angel Formation or lower Muav Formation, Grand Canyon, Arizona. (1–3) Holotype cranidium USNM 794191a (Resser, Reference Resser1945, pl. 25, fig. 9, center), dorsal, anterior, and lateral views.

Reference Resser1945 Trachycheilus typicale Resser, p. 213, pl. 25, fig 9.
Holotype
Cranidium USNM 794191a (Fig. 59.1–59.3) from USNM loc. 73b, Bright Angel Formation or Muav Formation, lower part of Kwagunt Valley, Grand Canyon, Arizona.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Bright Angel Formation or Muav Formation, Blacktail Canyon (Havasu Member) and Kwagunt Valley, Grand Canyon, Arizona.
Remarks
This species is based on a single cranidium found in association with Eh. kwaguntensis n. comb. from a “thin limestone in upper part” (Resser, Reference Resser1945, p. 172). One morphological feature not mentioned by Resser (Reference Resser1945) is the presence of a moderately coarse granular surface of the exoskeleton. The type species differs from both subspecies of Trachycheilus whirlwindensis Sundberg, Reference Sundberg1994, and Trachycheilus aff. T. whirlwindensis (Foster et al., Reference Foster, Sundberg and Hagadorn2024) by its swollen anterior border, less pronounced lateral glabellar furrows and granular ornamentation.
Trachycheilus granulosus Sundberg, Reference Sundberg1994
Trachycheilus spp. from the Elrathiella subzone, Ehmaniella Biozone, Spencer Canyon Member, Muav Formation, GRCA loc. 393, Grand Canyon, Arizona; all material preserved in limestone and mostly exfoliated. (1–4) Trachycheilus whirlwindensis Sundberg, Reference Sundberg1994: (1–3) cranidium GRCA 125549, dorsal, lateral, and anterior views; (4) librigena GRCA 125539. (5–12) Trachycheilus granulosus Sundberg, Reference Sundberg1994: (5) librigena GRCA 125540; (6–8) partial cranidium GRCA 125544, dorsal, oblique, and lateral views; (9) cranidium GRCA 125542; (10) pygidium GRCA 125552 questionably assigned to the species; (11) cranidium GRCA 125543; (12), cranidium GRCA 125541. Scales: (1–8, 11) 2-mm scale bar; (9, 10, 12) 1-mm scale bar.

Figure 60. Long description
The multi-panel photo displays 12 fossil specimens numbered in the top-right corner of each panel.
Panels 1 through 4 show Trachycheilus whirlwindensis.
* Panel 1: Dorsal view of a cranidium with a rounded glabella and smooth surface.
* Panel 2: Lateral view of the same cranidium showing the profile curvature.
* Panel 3: Anterior view of the cranidium.
* Panel 4: A narrow, curved librigena.
Panels 5 through 12 show Trachycheilus granulosus.
* Panel 5: A librigena with a distinct granular texture.
* Panels 6, 7, and 8: Dorsal, oblique, and lateral views of a partial cranidium, highlighting prominent surface granules.
* Panel 9: A smaller cranidium in dorsal view.
* Panel 10: A small, fan-shaped pygidium with visible segmentation.
* Panel 11: A dorsal view of a cranidium with a heavily granulated surface.
* Panel 12: A larger, detailed dorsal view of a cranidium showing the glabella and fixed cheeks.
Two scale bars are present. A 2-millimeter scale bar is located between panels 3 and 8, applying to panels 1 through 8 and 11. A 1-millimeter scale bar is located between panels 10 and 11, applying to panels 9, 10, and 12.
Reference Sundberg1994 Trachycheilus granulosus Sundberg, p. 80–82, fig. 50.
Occurrence
Ehmaniella Biozone, Spencer Canyon Member, Muav Formation, Rampart Cave, Grand Canyon, Arizona. Elrathiella subzone, Ehmaniella Biozone, Whirlwind Formation, Drum Mountains and House Range, Utah; Shadscale Formation Dugway Range, Utah (Sundberg, Reference Sundberg1994).
Remarks
The coarse tubercles on the exoskeleton and the relatively deep lateral and axial glabellar furrows characterize Trachycheilus granulosus from the Elrathiella subzone, Ehmaniella Biozone in Utah (Sundberg, Reference Sundberg1994). The cranidial specimens from the Muav Formation are fragmented, but distinctive enough to be placed into this species. An associated pygidium (Fig. 60.10) is also similar to those illustrated by Sundberg (Reference Sundberg1994, fig. 50.4). In contrast, the smaller librigena (Fig. 60.5) has coarse granules, unlike those associated with T. granulosus (Sundberg, Reference Sundberg1994, fig. 50.3).
Trachycheilus whirlwindensis Sundberg, Reference Sundberg1994
Reference Sundberg, Cooper and Stevens1991 Trachycheilus n. sp., Sundberg, fig. 2.16.
Reference Sundberg1994 Trachycheilus whirlwindensis Sundberg, p. 82–85, figs. 51, 52.
Occurrence
Ehmaniella Biozone, Spencer Canyon Member, Muav Formation, Rampart Cave, Grand Canyon, Arizona.
Remarks
The specimens of this species are associated with Trachycheilus granulosus and Elrathiella cf. E. decora. The limited specimens are most similar to T. whirlwindensis whirlwindensis from the Elrathiella subzone, Ehmaniella Biozone, Whirlwind Formation, Drum Mountains, Utah (Sundberg, Reference Sundberg1994), in their wider frontal area, but do not show a median anterior border swelling or the nodes on the posterior border typical of the subspecies. Incomplete preservation of the specimens prevents assignment to a subspecies. A single librigena (Fig. 60.4) illustrates a deflected genal spine typical of the species. The cranidia do not illustrate the coarse granules typical of T. granulosa. Associated with the material is a single pygidium that is here assigned to T. granulosus based on its shorter axis compared to T. whirlwindensis.
Genus Spencella Rasetti, Reference Rasetti1963
Type species
Spencella montanensis Rasetti, Reference Rasetti1963, from the Topazan (Wuliuan Stage), Meagher Limestone, Montana.
Remarks
Rasetti (Reference Rasetti1963) separated Spencella from the type of Spencia by the former having rounded librigenal genal angle, thorax with 13 or 14 segments, a pygidium that is wider with a lower profile axis, and a smooth (non-granulated) surface. Rasetti (Reference Rasetti1963) placed all of Resser’s (Reference Resser1945) Solenopleurella from the Grand Canyon into Spencella; however, if the pygidia and librigena associated with Spencella? porcata belongs to the species, then the generic assignment needs to be changed. The pygidium is similar to those associated with the Alokistocaridae, particularly Elrathiella (see Sundberg, Reference Sundberg1994) in its multi-segmented axis and strongly posteriorly curved pleural furrows and bands towards the posterior anterior border, which is unlike the relatively smooth, wide, borderless pygidium of Spencella.
Trymataspis Robison, Reference Robison1964, which is only known from cranidia and librigena from the upper Bolaspidella Biozone from the Marjum Formation, House Range, Utah, is similar to the Grand Canyon taxa. This genus includes three species named by Robison (Reference Robison1964), Ty. depressa, Ty. lomaleie, and Ty. pristina. The species are characterized by slightly tapered and truncate glabella; geniculate posterior lateral glabella furrows; subequal length of preglabellar field and anterior border; depressant anterolateral corners; stubby occipital spine; fixigena of moderate width; and lack of a librigenal genal spine (Robison, Reference Robison1964, p. 546). In addition, the ocular ridges project slightly posteriolaterally (approximately 90°). The Grand Canyon specimens differ in having a narrower to absent preglabellar area (Spe.? erosa, Spe.? diligens), more tapered glabella with a slight constriction at the S2 position, having an occipital node rather than a spine, ocular ridge projected more posteriorly (approximately 72°), and possible short genal spine (Spe.? porcata).
Spencella? diligens (Resser, Reference Resser1945)
Spencella? diligens (Resser, Reference Resser1945), from USNM loc. 75, Ehmaniella subzone, Ehmaniella Biozone, Peach Springs Member, Muav Formation, Grand Canyon, Arizona, mostly exfoliated: (1–3) holotype cranidium USNM 794136 (Resser, Reference Resser1945, pl. 27, figs. 29, 30), dorsal, anterior, and lateral views; (4) cranidium USNM 794140; (5, 6) paratype cranidium USNM 794137 (Resser, Reference Resser1945, pl. 27, fig. 31), dorsal and lateral views; (7) large partial cranidium USNM 794138; (8) small cranidium USNM 794141; (9) small cranidium USNM 794142. Scales: (1–6, 8, 9) 1-mm scale bar; (7) black 1-mm scale in picture.

Figure 61. Long description
A collection of nine fossil photographs arranged in three irregular rows.
* Panel 1 shows a dorsal view of the holotype cranidium U S N M 794136, featuring a rounded glabella and distinct border.
* Panel 2 shows the same specimen in anterior view, highlighting the curvature of the frontal margin.
* Panel 3 shows the same specimen in lateral view, illustrating the profile height.
* Panel 4 displays cranidium U S N M 794140 in a dorsal-oblique view.
* Panel 5 shows paratype cranidium U S N M 794137 in dorsal view.
* Panel 6 shows the paratype in lateral view.
* Panel 7 is a large partial cranidium U S N M 794138, showing the right half of the fossil with a black 1 mm scale bar in the bottom left corner.
* Panel 8 shows a small cranidium U S N M 794141 in dorsal view.
* Panel 9 shows a small cranidium U S N M 794142 in dorsal view.
A 1 mm scale bar is positioned between panels 1, 5, and 7 to provide a size reference for the specimens in panels 1 through 6, 8, and 9. The fossils appear mostly exfoliated with a granular texture against a rocky matrix.
Reference Resser1945 Solenopleurella diligens Resser, p. 215, pl. 27, figs. 29–31.
?Reference Palmer1968 Spencella montanensis; Palmer, p. B98, pl. 5, figs. 14–16.
Holotype
Cranidium USNM 794136 (Fig. 61.1–61.3) from USNM loc. 75, Peach Springs Member, Muav Formation or underlying Bright Angel Formation, Kanab Canyon, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794137–794142 from USNM loc. 44345J, Peach Springs Member, Muav Formation or underlying Bright Angel Formation, Kanab Canyon, Grand Canyon, Arizona.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, upper most Bright Angel Formation or the base of the overlying Peach Springs Member, Muav Formation, Kanab Canyon, Grand Canyon, Arizona. Altiocculus subzone, Ehmaniella Biozone. ?Hillard Limestone, Alaska (Palmer, Reference Palmer1968).
Remarks
Material of this species is limited. Resser (Reference Resser1945, p. 215) separated Sol. diligens from Sol. porcata in the former having no preglabellar area, relatively larger glabellar with faint furrows, and occipital node (“neck spine”). The overlap in glabellar length (Fig. 61.1) and tapering (Fig. 61.3) of the two species suggests that some the differences in glabellar shape are ontogenetic, but not entirely given the samples with higher glabellar lengths compared to similar-sized specimens of Spe.? porcata. The occurrence of an occipital node on Spe.? porcata is much more pronounced than the small and slight node on Spe.? diligens. The presence of well-developed lateral glabellar furrows and preglabellar field length (Fig. 61.2) in the well-preserved specimens of Spe.? porcata appears to be the only consistent difference between the two species.
Spencella montanensis Rasetti, Reference Rasetti1963, reported by Palmer (Reference Palmer1968) from the “Middle Cambrian-1 Fauna” of the Hillard Limestone is very similar to Spe.? diligens in glabellar shape, shallow glabellar furrows, preglabellar area length, anterior border, and smooth exoskeleton. The locality containing Palmer’s assemblage also contains Kootenia granulospinosa and Altiocculus sp. suggesting that the locality represents the Altiocculus/Ehmaniella subzone of the Ehmaniella Biozone. The occurrence of Spe. montanensis in comparable strata in western Canada suggests a similar biostratigraphic assignment (Handkamer et al., Reference Handkamer, Ichaso, Pratt, Mángano and Buatois2023).
Spencella? porcata (Resser, Reference Resser1945)
Spencella? porcata (Resser, Reference Resser1945) from the Ehmaniella subzone, Ehmaniella Biozone, Muav Formation. (1–8) Type and additional specimens of Solenopleurella porcata Resser, Reference Resser1945, from USNM loc. 44345J, Peach Springs Member, all material exfoliated: (1–3) holotype cranidium USNM 794143 (Resser, Reference Resser1945, pl. 27, fig. 34), dorsal, anterior, and lateral views; (4) paratype cranidium USNM 794144 (Resser, Reference Resser1945, pl. 27, fig. 33, bottom center); (5) paratype cranidia USNM 794145a upper left, 794145b lower center, and 794145c right (Resser, Reference Resser1945, pl. 27, fig. 33, upper left); (6) paratype cranidium USNM 794145d (Resser, Reference Resser1945, pl. 27, fig. 33, bottom right); (7) paratype cranidium USNM 794145e (Resser, Reference Resser1945, pl. 27, fig. 33, bottom left); (8) paratype cranidium USNM 794145f (Resser, Reference Resser1945, pl. 27, fig. 33, upper right). (10) Hypotype of Solenopleurella sp. indet. of Resser, Reference Resser1945, questionably assigned here to S.? porcata, cranidium USNM 794165 (Resser, Reference Resser1945, pl. 27, fig. 32), from USMN loc. 75, Peach Springs Member or upper Bright Angel Formation, exfoliated. (9, 11) Topotype material of Solenopleurella porcata: (9) cranidium GRCA 8585a; (11) librigena GRCA 8588a. (12–19) Additional new material from the Kanab Member, Diamond Bar Ranch, Arizona, mostly exfoliated unless otherwise mentioned: (12) cranidium FHPR 19657 from FHPR loc. 007-9; (13) cranidium FHPR 19625 from FHPR loc. 007-9, (14–16) cranidium FHPR 19629 from FHPR loc. 007-10, dorsal, anterior, and lateral views; (17) testate cranidium FHPR 19628 from FHPR loc. 007-9; (18) cranidium FHPR 19624 from FHPR loc. 007-9; (19) cranidium FHPR 19626 from FHPR loc. 007-9. Scales: (1–10) 2-mm scale bar; (11–19) 1-mm scale bar.

Figure 62. Long description
The plate consists of 19 black and white photographs of trilobite fossil fragments.
Panels 1 through 10 are associated with a 2 mm scale bar located below panel 2.
- Panels 1, 2, and 3 show the holotype cranidium U S N M 794143 in dorsal, anterior, and lateral views respectively, displaying a rounded glabella and distinct furrows.
- Panel 4 shows a paratype cranidium in dorsal view.
- Panel 5 displays a cluster of three paratype cranidia (a, b, and c) embedded in matrix.
- Panels 6, 7, and 8 show individual paratype cranidia in dorsal view.
- Panel 9 shows a topotype cranidium G R C A 8585a.
- Panel 10 shows a cranidium of Solenopleurella species indet.
Panels 11 through 19 are associated with a 1 mm scale bar located below panel 14.
- Panel 11 shows a librigena (free cheek) G R C A 8588a.
- Panels 12 and 13 show individual cranidia in dorsal view.
- Panels 14, 15, and 16 show cranidium F H P R 19629 in dorsal, anterior, and lateral views, highlighting the convex profile and glabellar lobes.
- Panel 17 shows a testate cranidium with preserved shell material.
- Panels 18 and 19 show additional cranidia in dorsal view.
All specimens are mostly exfoliated, revealing internal molds of the cephalic structure. The glabellae are generally tapered and rounded anteriorly with three pairs of lateral glabellar furrows visible in the better-preserved specimens like 13 and 14.
Spencella? porcata (Resser, Reference Resser1945), pygidia questionably assigned from the Ehmaniella subzone, Ehmaniella Biozone, Kanab Canyon Member, Muav Formation, Diamond Bar Ranch, Arizona; all specimens are from FHPR loc. 007-9, unless otherwise mentioned and are preserved in limestone. (1) Partial, latex FHPR 19637; (2) partial, exfoliated FHPR 19632; (3) partial, mostly exfoliated FHPR 19636; (4) partial, exfoliated FHPR 19631; (5, 6) partial exfoliated FHPR 19635, latex and internal mold; (7) partial, mostly exfoliated FHPR 19630 from FHPR loc. 007-10; (8) exfoliated FHPR 19633; (9) exfoliated FHPR 19634. Scales: (1–4, 7, 8) 1-mm scale bar; (5, 6, 9) 2-mm scale bar.

Figure 63. Long description
The nine panels show fossilized pygidia of Spencella? porcata.
* Panel 1: Top-left, a small, well-defined pygidium with a segmented axis and distinct pleural ribs.
* Panel 2: To the right of 1, a partial specimen showing the central axis and left pleural field.
* Panel 3: Center-top, a larger, mostly exfoliated specimen with a prominent, rounded axis.
* Panel 4: Top-right, a large partial specimen showing deep pleural furrows and a smooth border.
* Scale Bars: Located in the center-left, two horizontal black bars indicate 2 m m and 1 m m lengths.
* Panel 5: Middle-left, a partial exfoliated specimen showing the internal structure of the axis.
* Panel 6: Bottom-left, an internal mold showing the broad, fan-like pleural fields and a missing central axis section.
* Panel 7: Bottom-center-left, a small, mostly exfoliated specimen with visible axial rings.
* Panel 8: Bottom-center-right, an exfoliated specimen showing the semi-circular posterior margin.
* Panel 9: Bottom-right, a larger exfoliated specimen with a damaged central axis and clear lateral ribbing.
All specimens are set against a rough, granular limestone matrix. The fossils vary from light gray to white against the darker gray rock.
Reference Resser1945 Solenopleurella porcata Resser, p. 214, pl. 27, figs. 33, 34.
?Reference Resser1945 Solenopleurella diligens Resser, p. 215, pl. 27, figs. 29–31.
?Reference Resser1945 Solenopleurella sp. undet., Resser, p. 215, pl. 27, fig. 32.
Reference Rasetti1963 Spencella porcata; Rasetti, p. 591.
?Reference Palmer1968 Spencella montanensis; Palmer, p. B98, pl. 5, figs. 14–16.
Reference Sundberg1994 Spencella? utahensa Sundberg, p. 112–113, fig. 72.
Holotype
Cranidium USNM 794143 (Fig. 62.10) from USNM loc. 44345J, Peach Springs Member, Muav Formation, Rampart Cave, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794144 and 794145a–g from USNM loc. 44345J, Peach Springs Member, Muav Formation, Rampart Cave, Grand Canyon, Arizona.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Kanab to Gateway members, Muav Formation, Diamond Bar Ranch, Grand Wash Cliff; Havasu Canyon, Gateway Canyon, Toroweap Valley, Granite Park, Diamond Creek, Rampart Cave, Grand Canyon, Arizona. Altiocculus subzone, Ehmaniella Biozone, Swasey Limestone, Drum Mountains and House Range and Trailer Limestone, Dugway Range, Utah; member E Pole Canyon Limestone, Patterson Pass, Nevada (Sundberg, Reference Sundberg1994).
Emended description
Cranidium of small size, length 3.8 ± 1.0 mm (n = 32); subtrapezoidal, moderately convexity (sag. and trans.); anterior margin moderately, evenly curved; posterior margin (excluding occipital ring) directed laterally. Anterior branches of facial sutures moderately convergent by 26 ± 6° to anterior border, convergent to anterior margin; posterior branches moderately divergent at 55 ± 20°. Glabella moderately elongate, moderate convexity (sag. and trans.), length 76 ± 2% cranidial length; width 40 ± 4% cranidial width; moderately tapered, width at anterior end 73 ± 5% of posterior glabellar width; frontal lobe moderately to bluntly rounded. Axial furrows deep, with no constriction at the L2; preglabellar furrow moderately deep. Lateral glabellar furrows moderately deep laterally to shallow medially, S1 bifurcated, directed slightly posterior; S2 bifurcated directed transverse to slightly posteriorly; S3 shallow not connected with axial furrows, directed laterally to slightly anterior; S4 very shallow, connected with axial furrows, directed anteriorly. Occipital ring elevated above rest of glabella, moderately convex; length 23 ± 2% glabellar length; posterior margin strongly convex posteriorly, flattened medially; moderate occipital node. SO slightly curved anteriorly; deepest laterally forming deeper slits next to axial furrows, moderate depth medially. Frontal area unequally divided; length 23 ± 2% cranidial length. Preglabellar field slightly convex, strongly down-sloping, length 6 ± 2% cranidial length, 26 ± 8% frontal area length. Anterior border strongly convex, level, moderately to strongly arched dorsally, length 17 ± 2% cranidial length, strongly tapering laterally, moderate medial inbend. Anterior border furrow narrow and moderate depth, shallower medially, shallower than axial furrows. Fixigena moderately convex, level to slightly down-sloping, anterior area steeply down-sloping anteriorly, width 53 ± 8% of posterior glabellar width. Palpebral lobes nearly straight, level, moderately narrow and moderately long, length 35 ± 4% glabellar length; anterior margin located transversely opposite of S3 or L3. Ocular ridge obscure to weakly defined, slightly curved anteriorly, directed moderately posterolateral from glabella at 72 ± 4° to sagittal axis. Posterior area of fixigena triangular, terminated with rounded corner; length 28 ± 9% glabellar length; width 57 ± 9% glabella length.
Librigenae small size, length approximately 3.3–3.4 mm (n = 2); moderately narrow at about 40% length without spine; lateral margin moderately curved. Genal field slightly convex. Border moderately convex, uniform in width, width about 40–50% librigenal width; border furrows moderate depth and do not persist into the librigenal spine. Librigenal spine moderate length? at about 40% of librigenal length, poorly preserved, directed slightly posterolaterally.
Rostral plate, hypostome, and thorax unknown.
Pygidium small, length 2.2 ± 0.8 mm (n = 8); subsquare, length 61 ± 5% width; margin smooth, anterior margin nearly straight then directed strongly posterolaterally; anterolateral corners sharply rounded, transversely opposite near terminus of axis; no postaxial notch; moderately convex (sag.). Axis slightly tapered, mid-width 89 ± 7% anterior width, anterior width 39 ± 2% pygidial width; length 87 ± 4% pygidial length, extends nearly to posterior margin; five axial rings, strongly convex; terminal piece small, rounded posteriorly; axial ring furrows moderately deep. Pleural regions convex, down-sloping; anteriormost pleural furrow narrow and deep, extending to near margin; other pleural furrows moderate depth. Border narrow and poorly defined.
Granulated surface on the entire exoskeleton. Faint genal caeca on preglabellar regions of larger specimens.
Remarks
Most of the type specimens and the material from GRCA are composed of recrystallized calcite attached to the underlying limestone and overlain by siliciclastic silt. The preservation of surface ornamentation in these specimens is questionable. In some specimens in both the type collection (Fig. 62.6) and new material preserved in limestone (Fig. 62.13, 62.14), the external surface of the exoskeleton is covered with relatively coarse granules (Fig. 62.13). In addition, the internal molds are pitted (Fig. 62.15), which indicates that the internal surface of the exoskeleton was also granulated. The generic identification is questionable due to the absence of well-preserved librigenae and the possible assignment of the alokistocarid pygidium (Fig. 63) to the species.
Pygidia are not associated with the type material. However, new specimens from Diamond Bar Ranch that contain specimens of Spe.? porcata have pygidia that consist of two morphologies: one group that is more subsquare (length-to-width ratio of 61 ± 5%), with strongly posterolaterally directed anterior margins and five axial rings; the second group is more rectangular (length-to-width ratio of 49 ± 1%; n = 4), with moderately posterolaterally directed anterior margins and three to four axial rings. This latter group is similar to the type specimens of Eh. kwaguntensis n. comb., of which cranidia are also found in the same locality and are assigned to this species (see above). The first group is questionably assigned to Spe.? porcata but potentially could belong to an unknown co-occurring taxon. If these pygidia actually belong to Spe.? porcata, then the species does not belong to Spencella.
Resser (Reference Resser1945, p. 214–215) named Solenopleurella porcata, Sol. erosa, and Sol. diligens, and reported one Sol. sp. undet. from the Grand Canyon. He differentiated cranidia of Sol. porcata from the weathered cranidia of Sol. erosa in the latter having larger glabellar with “normal” furrows that “stand” completely above the frontal area and straight and non-tapering anterior border. In general, specimens of Spe.? porcata are larger than Spe.? sp. indet. (= cranidia of Solenoplurella erosa of Resser, Reference Resser1945; Fig. 64), which suggests that some of the morphological differences may be the result of ontogeny. The sub-rectangular shape of the glabella of Spe.? sp. indet. is also seen in smaller specimens of Spe.? porcata (Fig. 62.3) and one paratype (Fig. 62.10) does have a curved and tapered anterior border. Spencella? porcata can be distinguished from Spe.? sp. indet. in that the latter has strong inflation of the glabella and a narrow preglabellar area (Fig. 62.2) The pygidia that Resser (Reference Resser1945) assigned to the species, including the holotype, represent a Kootenia species.
Bivariate plots of Spencella? taxa from the Grand Canyon. (1) Relative glabellar length vs. cranidial length. (2) Relative preglabellar length vs. cranidial length. (3) Glabellar tapering vs. cranidial length. GRCA samples include topotype material, additional samples are the material collected during this study, other data points are the type material of Resser (Reference Resser1945).

Figure 64. Long description
A multi-panel figure containing three scatter plots labeled 1, 2, and 3. All plots share a common x-axis representing Cranidial Length in millimeters, ranging from 0.0 to 6.0. A shared legend at the top identifies six groups: Spe.? porcata types (red open circles), Spe.? porcata G R C A (grey open circles), Spe.? porcata additional (pink solid circles), Spe.? sp. (teal solid circle), Spe.? diligens (yellow solid circles), and Spe.? sp. indet (black solid circles).
Panel 1: The y-axis is Glabellar length forward slash Cranidial length, ranging from 0.70 to 0.90. Black circles (Spe.? sp. indet) form a high-ratio cluster between 0.82 and 0.87. Yellow circles (Spe.? diligens) cluster between 0.80 and 0.83. A large pink shaded region contains the various Spe.? porcata samples, showing a broad distribution between 0.73 and 0.81 across the full x-axis range.
Panel 2: The y-axis is Preglabellar length forward slash Cranidial length, ranging from 0.00 to 0.15. The Spe.? porcata samples occupy a distinct upper cluster within a pink shaded region between 0.03 and 0.10. In contrast, Spe.? diligens and Spe.? sp. indet are clustered tightly at the bottom of the graph, with ratios near 0.00.
Panel 3: The y-axis is Glabellar Tapering K 6 forward slash K 2, ranging from 0.60 to 0.90. All taxa show a general downward trend as cranidial length increases. Spe.? diligens (yellow) and Spe.? sp. indet (black) occupy the upper-left area with higher tapering ratios (0.73 to 0.85), while the Spe.? porcata group (pink shaded) shows the widest variance, extending down to a ratio of 0.61 at larger cranidial lengths.
Solenopleurella sp. undet. of Resser (Reference Resser1945) falls in the parameters of both Spe.? porcata and Spe.? diligens (Fig. 61), but the specimen (Fig. 62.8) is too poorly preserved to be assigned with confidence to either species. This species is also questionably assigned to Spe.? porcata.
Spencella? buttsi (Resser, Reference Resser1938a) and Spe.? utahensa Sundberg, Reference Sundberg1994, illustrated by Sundberg (Reference Sundberg1994) from the Altiocculus subzone, Ehmaniella Biozone of Utah are very similar to Spe.? porcata in having a narrow preglabellar area, tapering convex anterior border, rounded to slightly flattened frontal lobe, and shallow to deeper lateral glabellar furrows. Spencella? buttsi differs in having a smooth exoskeleton and generally less centrally swollen anterior border. Spencella? utahensa differs in the higher arched anterior margin; however, this is considered a minor difference, and this species is synonymized with Spe.? porcata.
Spencella? sp. indet. (Resser, Reference Resser1945)
Spencella? sp. indet. from Ehmaniella subzone, Ehmaniella Biozone, below the Gateway Member, Muav Formation, Grand Canyon, Arizona. (1–7) Type and associated cranidial specimens referred to as Solenopleurella erosa by Resser, Reference Resser1945, from USNM loc. 73c, preserved in limestone, weathered and exfoliated: (1) paratype cranidium USNM 794155 (Resser, Reference Resser1945, pl. 27, fig. 27); (2) paratype cranidium USNM 794163; (3, 5, 6) paratype cranidium USNM 794160 (Resser, Reference Resser1945, pl. 27, fig. 28), dorsal, lateral and anterior views; (4) partial cranidium USNM 794157; (7) partial cranidium USNM 794156. (8, 9) Specimens from USNM loc. 443381: (8) cranidium GRCA 8400b; (9) cranidium GRCA 8400a.

Figure 65. Long description
A collection of nine macro photographs showing fossilized trilobite cranidia (head shields) preserved in limestone. A 1 m m scale bar is positioned in the upper center between panels 2 and 3.
* Panel 1 (top left): Dorsal view of paratype U S N M 794155, showing a rounded glabella and distinct anterior border.
* Panel 2 (top center): Smaller dorsal view of paratype U S N M 794163.
* Panel 3 (top right): Detailed dorsal view of paratype U S N M 794160, highlighting the granular surface texture.
* Panel 4 (middle left): Partial cranidium U S N M 794157 showing the left side and glabella.
* Panel 5 (center): Lateral view of paratype U S N M 794160, showing the profile curvature.
* Panel 6 (middle right): Anterior view of paratype U S N M 794160, showing the frontal elevation of the glabella.
* Panel 7 (bottom left): Partial cranidium U S N M 794156 with a weathered, pitted surface.
* Panel 8 (bottom center): Cranidium G R C A 8400b, showing a well-defined anterior border furrow.
* Panel 9 (bottom right): Cranidium G R C A 8400a, showing the glabella and fixed cheeks in high relief.
Reference Resser1945 Solenopleurella erosa Resser, p. 214–215, pl. 27, figs. 27, 28 (not pl. 23, figs. 5, 6; pl. 27, fig. 26; = Kootenia erosa [Resser, Reference Resser1945]).
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Gateway Canyon Member, Muav Formation, Havasupai Gardens and Hermit Creek, Grand Canyon, Arizona.
Remarks
Resser (Reference Resser1945) assigned several, relatively large pygidia and small weathered cranidia to this species. The pygidia are here assigned to Kootenia erosa (see discussion above). The cranidia are too poorly preserved to determine generic or specific identification and are left in open nomenclature. Added to this species are two cranidia (Fig. 65.8, 65.9) from Havasupai Gardens that are fragmented, but better preserved than the weathered cranidia from the type material.
Family Crepicephalidae Kobayashi, Reference Kobayashi1935
Genus Crepicephalus Owen, Reference Owen1852
Type species
Dikelocephalus iowensis Owen, Reference Owen1852, from the Marjuman (Wuliuan Stage), Eau Claire Formation, Minnesota (designated by Walcott, Reference Walcott1886, p. 206; see discussion by Pratt, Reference Pratt1992, p. 63).
Crepicephalus? upis Walcott, Reference Walcott1916a
Figures 16.6, 16.9, 16.10, 66
Crepicephalus? upia Walcott, Reference Walcott1916a, from the Bolaspidella Biozone, Gateway Canyon Member, Muav Formation, Mile 52, Grand Canyon, Arizona; all specimens from GRCA loc. 357-4 and are mostly exfoliated unless otherwise mentioned. (1) Small testate cranidium GRCA 125427a; (2) small testate librigena GRCA 125427b; (3) small testate pygidium GRCA 125425; (4) moderate-size librigena GRCA 125431; (5) partial testate librigena GRCA 125429b; (6) testate librigena GRCA 125429a; (7) incomplete pygidium GRCA 125433c; (8–10) latex cast of internal surface, partial cranidium GRCA 12526, dorsal, anterior, and lateral views; (11) partial cranidium GRCA 125433a; (12) latex cast of internal surface, partial cranidium GRCA 125425; (13) large librigena GRCA 125432a; (14) partial cranidium GRCA 125432b; (15) mostly complete pygidium GRCA 125433b; (16–18) larger pygidium GRCA 125430, dorsal, posterior, and lateral views. Scales: (1–3) 1-mm scale bar; (4–7, 15) larger 2-mm scale bar; (8–12, 14, 16–18) smaller 2-mm scale bar; (13) 4-mm scale bar.

Figure 66. Long description
A collection of 18 fossil specimens arranged in a grid.
* Top row: Panel 1 shows a small testate cranidium. Panel 2 is a small testate librigena. A 1 mm scale bar sits between panels 2 and 3. Panel 3 shows a small testate pygidium. Panel 4 is a moderate-size librigena. Panel 5 is a partial testate librigena. Panel 6 is a testate librigena. A 2 mm scale bar is positioned below panel 4.
* Middle section: Panel 7 is an incomplete pygidium. Panels 8, 9, and 10 show a latex cast of a partial cranidium in dorsal, anterior, and lateral views respectively. Panel 11 is a partial cranidium. Panel 12 is a latex cast of a partial cranidium. Panel 13 is a large librigena with a 4 mm scale bar. Panel 14 is a partial cranidium. A smaller 2 mm scale bar is located near panel 16.
* Bottom section: Panel 15 is a mostly complete pygidium. Panels 16, 17, and 18 show a larger pygidium in dorsal, posterior, and lateral views respectively.
The fossils are mostly exfoliated, appearing as light gray textured stone against a darker, rocky matrix. The cranidia feature a central glabella with distinct furrows, while the librigenae show curved outer margins and the pygidia exhibit segmented axial lobes.
Reference Walcott1916a Crepicephalus upis Walcott, p. 218, pl. 33, fig. 4–4d.
Reference Walcott1916a Crepicephalus unzia Walcott (part), p. 217, pl. 34, fig. 7a.
Reference Resser1935 Kochaspis upis; Resser, p. 38.
Reference Resser1935 Kochaspis unzia, Resser, p. 38.
Reference Deiss1939 Kochaspis upis Deiss, p. 99, pl. 17, figs. 6, 7.
Reference Deiss1939 Kochaspis unzia Deiss, p. 99, pl. 17, figs. 3–5.
Reference Deiss1939 Kochaspis resseri Deiss, p. 99, pl. 17, figs. 1, 2.
Lectotype
Cranidium USNM 61697 from USNM loc. 150b, Steamboat Limestone, White River-Indian Creek section, Montana.
Paralectotypes
Cranidium USNM 61695, 61696, and 61698 from USNM loc. 150b, Steamboat Limestone, White River-Indian Creek section, Montana.
Occurrence
Bolaspidella Biozone, Gateway Canyon Member, Muav Formation, Nankoweap Canyon, Grand Canyon, Arizona. Bolaspidella Biozone: Steamboat Limestone, Montana (Schwimmer, Reference Schwimmer1973).
Remarks
In the most recent review of this species, Schwimmer (Reference Schwimmer1973) placed material from the Steamboat Limestone of Montana into a new genus. The description of the genus has not been published; however, the name “Gandalfus upis” has been used in Schwimmer (Reference Schwimmer1975, fig. 2). The generic name is here considered a nomen nudum.
Walcott (Reference Walcott1916a) originally placed this species into Crepicephalus Owen, Reference Owen1852, and its morphology is generally similar to Crepicephalus. However, the cranidia of C.? upis illustrated by Deiss (Reference Deiss1939), Schwimmer (Reference Schwimmer1973), and herein have glabellar lateral furrows that are more defined than in species of Crepicephalus (data from Palmer, Reference Palmer1954; Derby, Reference Derby1965; Rasetti, Reference Rasetti1965; Eby, Reference Eby1981; Pratt, Reference Pratt1992), especially S2 and S3; a glabellar shape that is more rectangular (glabellar width 71 ± 3% of glabellar length, n = 7 vs. Crepicephalus spp. having 80 ± 5%, n = 18); and a less curved and more anteriorly placed palpebral lobes. The pygidia of C.? upis and Crepicephalus spp. are similar in their transversely rectangular outline with a single pair of marginal spines. In Crepicephalus? upis, the pygidial pleural furrows and bands extend onto the anterior border without the quick termination as seen in Crepicephalus. In addition, the lateral spines are composed of the anterior pleural segment with the anterior band, pleural furrow, and posterior band forming the spine. This is unlike Crepicephalus spp., Crepicephalina Resser and Endo in Kobayashi, Reference Kobayashi1935, Hsuchuangia Lu and Zhu in Qiu et al., Reference Qiu, Lu, Zhu, Bi and Lin1983, and Temnoura Resser and Endo in Kobayashi, Reference Kobayashi1935 (see Zhang and Jell, Reference Zhang and Jell1987, for discussions of the previous three genera), which have spines derived from the second and third pleural segment, with the anterior pleural segment terminating against the margin near the anterior of the pygidium before the well-developed spine. The segment arrangement seen in C.? upis; however, is similar to Hadrocephalites Sundberg and McCollum, Reference Sundberg and McCollum2002, in that the first pygidial segment forms the majority of the spine, but the latter genus differs in having the second pygidial segment also involved with forming the spine.
Deiss (Reference Deiss1939) placed C.? upis into Kochaspis Resser, Reference Resser1935, based on the assumption that Kochaspis had pygidial spines. However, the type species of Kochaspis, Crepicephalus liliana Walcott, Reference Walcott1886, does not have pygidial spines or the rectangular pygidial shape, subrectangular glabellar outline, narrow fixigena, or longer palpebral lobes (Sundberg and McCollum, Reference Sundberg and McCollum2002, Reference Sundberg and McCollum2003).
The specimens from the Muav Formation are nearly identical to the material from the Steamboat Limestone, Bolaspidella Biozone, illustrated by Schwimmer (Reference Schwimmer1973), with a minor difference of having shorter pygidial spines. This difference may be due to the relatively poor preservation of the material from the Muav, from which all of the material represents internal molds.
Family Marjumiidae Kobayashi, Reference Kobayashi1935
Genus Modocia Walcott, Reference Walcott1924
Type species
Arionellus (Crepicephalus) oweni Meek and Hayden, Reference Meek and Hayden1861, from the Marjuman (Wuliuan Stage) Deadwood Formation, South Dakota.
Modocia sp. indet.
Modocia sp. indet. from the Bolaspidella Biozone, Frenchman Mountain Dolostone, Grand Canyon, Arizona. Specimens from GRCA loc. 357-7, preserved in limestone. (1) Partially testate cranidium GRCA 125455; (2) testate, partial pygidium GRCA 125456.

Occurrence
Bolaspidella Biozone, Frenchman Mountain Dolostone, Nankoweap Canyon, Grand Canyon, Arizona.
Remarks
One partial cranidium and one partial pygidium were recovered. The general cranidial and glabellar shape is similar to species of Modocia, but without larger and more complete specimens, a specific identification cannot be made. Modocia is a common taxon in the Bolaspidella Biozone (e.g., Robison, Reference Robison1964).
Family Solenopleuridae Angelin, Reference Angelin1854
Genus Solenopleurella Poulsen, Reference Poulsen1927
Type species
Solenopleurella ulrichi Poulsen, Reference Poulsen1927, from the Steptoean (Drumian Stage), Cape Wood Formation, northwestern Greenland.
Solenopleurella? quadrata Rasetti, Reference Rasetti1963
Solenopleurella? quadrata Rasetti, Reference Rasetti1963, from the Bolaspidella Biozone, Frenchman Mountain Dolostone, Grand Canyon, Arizona; specimens from GRCA loc. 357-7, preserved in limestone. (1–3) Testate cranidium GRCA 125458, dorsal, anterior, and lateral views; (4) nearly complete, exfoliated cranidium GRCA 125457.

Figure 68. Long description
A multi-panel set of four grayscale photos showing fossilized trilobite cranidia.
* Panel 1, top-left: A dorsal view of testate cranidium G R C A 125458. It shows a central glabella with a rounded front, flanked by fixed cheeks with a granular surface texture. Deep furrows separate the glabella from the cheeks.
* Panel 2, top-right: An anterior view of the same specimen, showing the convex curvature of the frontal margin and the elevation of the glabella relative to the lateral lobes.
* Panel 3, middle-right: A lateral view of the same specimen, highlighting the profile of the glabella and the downward slope of the posterior portion.
* Panel 4, bottom-left: A dorsal view of a nearly complete, exfoliated cranidium G R C A 125457. This specimen appears smoother than the first, with the glabella tapering slightly toward the front and distinct ocular ridges extending toward the margins.
In the bottom-right corner, a horizontal black scale bar is labeled 1 m m.
Reference Rasetti1963 Solenopleurella? quadrata Rasetti, p. 590, pl. 70, figs. 9–12.
Holotype
Cranidium USNM 140301 from boulder G-6 within the Ordovician Lévis Formation, Grosses Roches, Gaspé Peninsula, Québec.
Paratypes
Cranidium USNM 140302 from boulder G-6 within the Ordovician Lévis Formation, Grosses Roches, Gaspé Peninsula, Québec.
Occurrence
Bolaspidella Biozone, Frenchman Mountain Dolostone, Nankoweap Canyon, Grand Canyon, Arizona. Bolaspidella Biozone, Lévis Formation, Québec (in a middle Cambrian boulder; Rasetti, Reference Rasetti1963), Abrigo Formation, southern Arizona (Łabaj, Reference Łabaj2015).
Remarks
This species is represented by only two small cranidia. They match well with the specimens illustrated by Rasetti (Reference Rasetti1963) from Québec and by Łabaj (Reference Łabaj2015) from southern Arizona. Both authors reported this taxon from the Bolaspidella Biozone.
Order Uncertain
Family Bolaspididae Howell in Harrington et al., Reference Harrington, Henningsmoen, Howell, Jaanusson, Lochman-Balk, Part and Moore1959
Remarks
Both Bolaspis and Eldoradia were included in the Family Bolaspididae by Jell and Adrain (Reference Jell and Adrain2003). Adrain (Reference Adrain2011) placed this family into an uncertain order.
Genus Bolaspis? Resser, Reference Resser1935
Type species
Alokistocare? labrosum Walcott, Reference Walcott1916a, from the Topazan (Wuliuan Stage), Wolsey Shale, near Neihart, Montana.
Remarks
Schwimmer (Reference Schwimmer1973) removed Bolaspis aemula Resser, Reference Resser1945, and Bol. sp. undet. of Resser (Reference Resser1945) from the genus based on the less-convex cranidia, non-triangular or absent frontal area swelling, and wider (tr.) frontal area, but provided no generic assignment. Given the limited number and generally poorly preserved specimens that are only known from cranidia in the Grand Canyon, the generic assignment is left open.
Bolaspis? aemula Resser, Reference Resser1945
(1, 2) Bolaspis? aemula Resser, Reference Resser1945, Ehmaniella subzone, Ehmaniella Biozone, Gateway Canyon Member, Muav Formation, Arizona: (1) holotype cranidium USNM 794055a (Resser, Reference Resser1945, pl. 22, fig. 15, upper left-hand corner; pl. 23, fig. 2) in sandy limestone from USNM loc. 44338F; (2) hypotype of Bolaspis? sp. undet. of Resser, Reference Resser1945, cranidium USNM 794004a (Resser, Reference Resser1945, pl. 19, fig. 4) from USNM loc. 73c, questionably assigned to “Bolaspis” aemula, in limestone. (3) Eldoradia cf. E. prospectensis Walcott, Reference Walcott1884, paratype cranidium of Bolaspis aemula Resser, Reference Resser1945, USNM 794055b (Resser, Reference Resser1945, pl. 22, fig. 15, lower right-hand corner; pl. 23, fig. 1, bottom center) in sandy limestone from USNM loc. 44338F.

Reference Resser1945 Bolaspis aemula Resser, p. 201, pl. 22, fig. 15 (lower right corner), pl. 23, fig. 2 (not fig. 1 = Eldoradia cf. E. prospectensis [Walcott, Reference Walcott1884]).
?Reference Resser1945 Bolaspis sp. undet. Resser, p. 204, pl. 19, fig. 4.
Holotype
Cranidium USNM 794055a (Figs. 16.7, 69.1) from the Gateway Canyon Member, Muav Formation at USNM loc. 44338F Bright Angel Trail, Grand Canyon, Arizona.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Gateway Canyon Member, Muav Formation Havasupai Gardens and Bright Angel Trail, Grand Canyon, Arizona.
Remarks
The two type specimens of Bol. aemula are here divided into Eldoradia cf. E. prospectensis (Walcott, Reference Walcott1884) (USNM paratype 794055b) and Bol.? aemula (USNM holotype 794055a). The paratype has the wide cranidium between the palpebral lobes, well-pronounced frontal area swelling, and relatively short (sag.), upturned anterior border typical of E. prospectensis. The holotype of Bolaspis? aemula occurs on the same slab of rock but illustrates a less-defined frontal area swelling and a longer anterior border. As discussed above, these features are not typical of Bolaspis. Bolaspis? sp. undet. (USNM 794004) is questionably assigned to this species based on cranidial outline, similar glabellar shape and characteristics of the anterior border. This single specimen differs from the holotype in lacking an apparent frontal area swelling.
Genus Eldoradia Resser, Reference Resser1935
Type species
Ptychoparia? linnarssoni Walcott, Reference Walcott1884, from the Topazan (Wuliuan Stage), Secret Canyon Shale, Eureka, Nevada.
Eldoradia cf. E. prospectensis (Walcott, Reference Walcott1884)
cf. Reference Walcott1884 Ptychoparia? prospectensis Walcott, p. 46, pl. 9, fig. 20.
cf. Reference Walcott1916a Alokistocare prospectensis; Walcott, p. 186, pl. 25, fig. 8.
Reference Resser1945 Bolaspis aemula Resser, p. 201, pl. 22, fig. 15 (upper left corner), pl. 23, fig. 1 (not fig. 2).
cf. Reference Palmer1954 Eldoradia prospectensis, Palmer, p. 77, pl. 16, fig. 8.
Holotype
Cranidium USNM 15441 from the Secret Canyon Shale (?), Eureka, Nevada.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Gateway Canyon Member, Muav Formation, Havasupai Gardens, Grand Canyon, Arizona.
Remarks
The paratype of Bol.? aemula (USNM 794055b) is similar to E. prospectensis, sharing a wide cranidium between the palpebral lobes, well-pronounced frontal area swelling, and relatively short (sag.) anterior border, This specimen differs from Bolaspis labrosa (Walcott, Reference Walcott1916a) (Handkamer et al., Reference Handkamer, Ichaso, Pratt, Mángano and Buatois2023, fig. 6.8, 6.9) in having a less tapered glabella and shorter anterior border. The specimen also differs from Eldoradia linnarssoni (Walcott, Reference Walcott1884) (Palmer, Reference Palmer1954, pl. 16, figs. 9, 10) in having a less tapered glabella, more defined anterior border, and longer palpebral lobes.
Family Uncertain
Genus Amecephalus Walcott, Reference Walcott1924
Type species
Ptychoparia piochensis Walcott, Reference Walcott1886, from the upper Delamaran (Wuliuan Stage), Chisholm Shale, Pioche Hills, Nevada.
Remarks
A modified diagnosis and discussion of the type species were presented by Sundberg (Reference Sundberg2020). Specimens preserved in limestone in the Flour Sack Member, Bright Angel Formation, at the Diamond Bar Ranch section are typically crushed and fractured. In most cases, this prevents an accurate assignment to a species.
Amecephalus althea (Walcott, Reference Walcott1916a)
Amecephalus althea (Walcott, Reference Walcott1916a) from the Glossopleura walcotti Biozone, Bright Angel Formation, Grand Canyon, Arizona. (1–4) Walcott’s (Reference Walcott1916a) type specimens: (1) lectotype shield USNM 793989 (Walcott, Reference Walcott1916a, pl. 25, fig. 4a; Resser, Reference Resser1945, pl. 22, fig. 7), from USNM loc. 74e, preserved in shale; (2) paralectotype partial shield USNM 793988 (Walcott, Reference Walcott1916a, pl. 25, fig. 4; Resser, Reference Resser1945, pl. 22, fig. 6), from USNM loc. 74e, preserved in shale; (3) paralectotype cranidium USNM 794152 (Walcott, Reference Walcott1916a, pl. 25, fig. 3; Resser, Reference Resser1945, pl. 22, fig. 10), from USNM loc. 74, preserved in sandstone; (4) paralectotype cranidium USNM 794146 (Walcott, Reference Walcott1916a, pl. 25, fig. 3a; Resser, Reference Resser1945, pl. 22, fig. 9), from USNM loc. 74, preserved in sandstone. (5–8) Resser’s (Reference Resser1945) hypotypes: (5) hypotype cranidium USNM 793990 (Resser, Reference Resser1945, pl. 22, fig. 8), from USNM loc. 73e, preserved in sandstone; (6–8) hypotype cranidium USNM 793991 (Resser, Reference Resser1945, pl. 22, fig. 11), from USNM loc. 73e, preserved in limestone, dorsal, lateral, and anterior views; (9) additional, mostly exfoliated cranidium FHPR 19532 from the Diamond Bar Ranch, Arizona, FHPR loc. 006-20, in limestone.

Figure 70. Long description
A multi-panel collection of nine fossil photographs. A black scale bar labeled 4 m m is positioned in the center.
* Panel 1: A complete, elongated lectotype shield U S N M 793989 preserved in shale, showing a wide cephalon and a long, segmented thorax tapering to a small pygidium.
* Panel 2: A paralectotype partial shield U S N M 793988 in shale, missing the lower right portion but clearly displaying the cephalon and upper thoracic segments.
* Panel 3: A paralectotype cranidium U S N M 794152 in sandstone, focusing on the central head region with a distinct glabella.
* Panel 4: A smaller paralectotype cranidium U S N M 794146 in sandstone.
* Panel 5: A hypotype cranidium U S N M 793990 in sandstone, showing a granular texture.
* Panels 6, 7, and 8: Three views of hypotype cranidium U S N M 793991 in limestone. Panel 6 is a dorsal view, Panel 7 is a lateral profile showing the curvature, and Panel 8 is an anterior view.
* Panel 9: An exfoliated cranidium F H P R 19532 in limestone, showing the internal mold of the head structure.
Reference Walcott1916a Alokistocare althea Walcott, p. 184, pl. 25, figs. 3, 3a, 4, 4a.
Reference Resser1935 Alokistocare althea; Resser, p. 6.
Reference Resser1939a Alokistocare septum Resser, p. 17, pl. 4, figs. 5–7.
Reference Resser1945 Alokistocare althea; Resser, p. 203, pl. 22, figs. 6–11.
?Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952 Alokistocare althea; Lochman in Cooper et al., p. 114–116, pl. 27, figs. 3–8.
Reference Foster2011 Alokistocare althea; Foster, p. 110–112, fig. 7.4–7.7.
Lectotype
Shield USNM 793989 (Fig. 70.1) from USNM loc. 74e, Bright Angel Formation, Bright Angel Trail near Havasupai Gardens, Grand Canyon, Arizona.
Paralectotypes
USNM 794146–794153 USNM loc. 74e, Bright Angel Formation, Nankoweap Valley; USNM 793988 from USNM loc. 74e, Bright Angel Formation, Bright Angel Trail near Havasupai Gardens, Grand Canyon, Arizona.
Occurrence
Glossopleura walcotti Biozone, Bright Angel Formation, Diamond Bar Ranch (Flour Sack Member), Grand Wash Cliffs; Nankoweap Valley, Kwagunt Valley, Lava Canyon, Bright Angel Trail, Tapeats Creek, Diamond Creek (Spencer Canyon Member, Muav Formation), and Rampart Cave (Flour Sack Member) Grand Canyon, Arizona.
Remarks
A single cranidium from limestone has the characteristics of Am. althea, including the triangular swelling in the frontal region, and is assigned to the species. Foster (Reference Foster2011) included two of Resser’s (Reference Resser1945) species, Am. laticaudum and Am. septum, from the Spence Shale into this species based on the long, broad-based genal spine.
Lochman (in Cooper et al., Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952, p. 116) assigned specimens from the Arrojos Formation to Am. althea; however, the palpebral lobes are longer and correspondingly the posterior area of the fixigena are shorter (exsag.) and the surface is “coarsely granulated”. These features are more similar to Am. scapegoatensis (Deiss, 1938), but the lack of visible fine details in Lochman (in Cooper et al., Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952, pl. 27) prevents a detailed assessment of the features present.
Amecephalus cf. Am. normale (Resser, Reference Resser1938b)
Amecephalus cf. A. normale (Resser, Reference Resser1938a) from the Glossopleura walcotti Biozone, Bright Angel Formation, Diamond Bar Ranch, Arizona; all specimens are mostly exfoliated. (1) Cranidium FHPR 19547 from FHPR loc. 006-19; (2) cranidium FHPR 19549 from FHPR loc. 006-19; (3) cranidium FHPR 19548 from FHPR loc. 006-19; (4) librigena FHPR 19550 USNM from FHPR loc. 006-19; (5) cranidium FHPR 19546 from FHPR loc. 006-19; (6) mostly testate cranidium FHPR 19540 from FHPR loc. 006-17; (7) cranidium FHPR 19541 from FHPR loc. 006-17; (8) mostly exfoliated cranidium FHPR 19542 from FHPR loc. 006-18; (9) partially testate cranidium FHPR 19544 from FHPR loc. 006-19; (10) cranidium FHPR 19543 from FHPR loc. 006-18; (11) cranidium FHPR 19545 from FHPR loc. 006-19; (12) cranidia FHPR 19538 larger, and FHPR 19539 smaller from FHPR loc. 006-17. Scales: (1–7, 9–11) 2-mm scale bar; (8, 12) white 2-mm scale bar.

Figure 71. Long description
The plate consists of 12 panels numbered in the top-right corner of each frame.
1. Top row, far left. Small exfoliated cranidium F H P R 19547.
2. Top row, second from left. Exfoliated cranidium F H P R 19549.
3. Top row, third from left. Exfoliated cranidium F H P R 19548.
4. Top row, fourth from left. A curved librigena F H P R 19550 U S N M.
5. Top row, far right. Large exfoliated cranidium F H P R 19546.
6. Middle row, far left. Mostly testate cranidium F H P R 19540 showing smooth surface texture.
7. Middle row, second from left. Cranidium F H P R 19541 with a prominent central glabella.
8. Middle row, third from left. Mostly exfoliated cranidium F H P R 19542 with a white 2-millimeter scale bar in the bottom-left corner.
9. Middle row, far right. Partially testate cranidium F H P R 19544 showing a crack across the right side.
10. Bottom row, far left. Cranidium F H P R 19543.
11. Bottom row, center. Cranidium F H P R 19545.
12. Bottom row, far right. Two cranidia, a larger F H P R 19538 and a smaller F H P R 19539, with a white 2-millimeter scale bar in the bottom-left corner.
A black 2-millimeter scale bar is positioned centrally between the middle and bottom rows, applicable to panels 1 through 7 and 9 through 11.
Reference Resser1938b Alokistocare normale Resser, p. 7, pl. 1, fig. 44.
Reference Sundberg2020 Amecephalus normale; Sundberg, p. 28–29, figs. 24, 25 (see for further synonymy).
Holotype
Cranidium USNM 95028 from USNM loc. 37n, Lakeview Limestone, Pend-Oreille Lake, Idaho.
Occurrence
Glossopleura walcotti Biozone, Flour Sack Member, Bright Angel Formation, Diamond Bar Ranch, Grand Wash Cliffs, Arizona.
Remarks
Overall, the specimens from Diamond Bar Ranch are a good match to the type material from the Lakeview Limestone, Idaho (see Sundberg, Reference Sundberg2020). However, if the librigena is correctly assigned to the cranidia, the genal spine has a narrower breadth of the base, is shorter, and is missing the furrow extending down the spine. In addition, some fragmented cranidia of the specimens have relatively short palpebral lobes (Fig. 71.10, 71.12) For these reasons, the specimens are considered closely related to A. normale.
Amecephalus piochensis (Walcott, Reference Walcott1886)
Amecephalus piochensis (Walcott, Reference Walcott1886), from the Glossopleura walcotti Biozone, Arizona. (1) Holotype shield USNM 793986 (Resser, Reference Resser1945, pl. 26, fig. 11) of Acrocephalops? arizonaensis Resser, Reference Resser1945, here assigned to A. piochensis from USNM loc. 44345F, Flour Sack Member, Bright Angel Formation, Grand Canyon. (2–6) Mostly exfoliated specimens in limestone from FHPR loc. 006-17 and unless otherwise mentioned: (2) cranidium FHPR 19552; (3) cranidium FHPR 19553; (4) pygidium FHPR 19554; (5) small cranidium FHPR 19551 from FHPR loc. 006-19, (6) cranidia FHPR 19555 lower and 19556 upper. (6) Alokistocare sp. undet. reported by Resser, Reference Resser1945, USNM 793992 (Resser, Reference Resser1945, pl. 22, fig. 5) questionably assigned to Amecephalus piochensis, from USNM loc. 44344A, Flour Sack Member, Bright Angel Formation, Grand Canyon; specimen appears to have been heavily damaged after photograph taken by Resser. Scales: (1, 4, 5) 2-mm scale bar; (2, 3, 6, 7) 5-mm scale bar.

Figure 72. Long description
A multi-panel collection of seven grayscale photographs of trilobite fossils.
* Panel 1 (Left): A large, nearly complete holotype shield U S N M 793986. It shows a rounded cephalon with a wide brim, a segmented thorax with approximately 15 segments, and a small pygidium at the base.
* Panel 2 (Top Center): A cranidium F H P R 19552 showing a smooth, rounded glabella and wide lateral fixed cheeks.
* Panel 3 (Top Right): Cranidium F H P R 19553, similar in structure to panel 2 but with more visible surface cracking.
* Panel 4 (Center): A small, wide pygidium F H P R 19554 with a distinct central axis and faint lateral ribbing.
* Panel 5 (Middle Right): A small cranidium F H P R 19551 with a prominent glabella.
* Panel 6 (Bottom Center): Two cranidia, F H P R 19555 (lower) and 19556 (upper), partially overlapping in the matrix.
* Panel 7 (Bottom Right): A heavily damaged specimen U S N M 793992 with a rough, pitted texture and less distinct anatomical features.
Two scale bars are positioned in the center right. The top bar is labeled 5 mm and applies to panels 2, 3, 6, and 7. The bottom bar is labeled 2 mm and applies to panels 1, 4, and 5.
Reference Walcott1886 Ptychoparia piochensis Walcott, pl. 9 fig. 1, pl. 28, fig. 1b (not pl. 28, fig. 1, 1a = possible Amecephalus kempi [Pack, Reference Pack1906]; not pl. 28, fig. 1c, 1d = Piochaspis packi Resser, Reference Resser1935).
Reference Pack1906 Ptychoparia piochensis; Pack, p. 297, pl. 2, fig. 4a, b (not 4c = corynexochid hypostome).
?Reference Walcott1924 Amecephalus piochensis; Walcott, p. 54 (not pl. 9, fig. 1 = possible Amecephalus kempi [Pack, Reference Pack1906]).
Reference Walcott1925 Amecephalus piochensis; Walcott, p. 66–67, pl. 15, fig. 9 (not pl. 15, fig. 8 = possible Amecephalus kempi [Pack, Reference Pack1906]).
Reference Resser1935 Alokistocare piochensis; Resser, p. 7.
Reference Resser1945 Acrocephalops? arizonaensis Resser, p. 203, pl. 26, fig. 11.
?Reference Resser1945 Alokistocare sp. undet. Resser, p. 203, pl. 22, fig. 5.
Reference Palmer1954 Alokistocare piochensis; Palmer, pl. 16, figs. 3, 4 (not fig. 1 = Piochaspis packi Resser, Reference Resser1935; figs. 2, 5 = possible Amecephalus kempi [Pack, Reference Pack1906]).
Reference Foster2011 Amecephalus piochensis; Foster, fig. 10.1 (not figs. 10.2, 10.3 = Piochaspis packi Resser, Reference Resser1935).
Reference Sundberg2020 Amecephalus piochensis; Sundberg, p. 26–28, figs. 22, 27 (not fig. 22.1, 22.4, 22.5 = Piochaspis packi Resser, Reference Resser1935; fig. 22.8, 22.10, 22.11 = possible Amecephalus kempi [Pack, Reference Pack1906]).
Lectotype
Shield USNM 793986 from USNM loc. 31, from the Glossopleura walcotti Zone, Chisholm Formation, Half Moon Mine, Pioche, Nevada.
Paralectotypes
Nearly complete shields, librigena, cranidia, and pygidium USNM 15434c–e, and USNM 15434 from the Glossopleura walcotti Zone, Chisholm Formation, Half Moon Mine, Pioche, Nevada.
Occurrence
Glossopleura walcotti Biozone, Flour Sack Member, Bright Angel Formation, Diamond Bar Ranch, Grand Wash Cliffs; Little Colorado River Camp, ?Foster Canyon, Fossil Canyon, Diamond Creek (Spencer Canyon Member, Muav Formation), Rampart Cave, ?River Mile 274, Grand Canyon, Arizona; Frenchman Mountain, Nevada. Glossopleura walcotti Biozone, Chisholm Formation, Half Moon Mine, Pioche, Nevada (Palmer, Reference Palmer1954).
Remarks
Sundberg (Reference Sundberg2020) discussed the complexity in recognition of the species and the lectotype from the type material and taxa reported by subsequent workers, which represents at least three species, Am. piochensis, Piochaspis packi (Resser, Reference Resser1935), and possibly Am. kempi (Pack, Reference Pack1906). Pack (Reference Pack1906, pl. 3, fig. 1) provided a description and only a drawing of his new species Ptychoparia kempi, which looks similar to some of the types presented by Walcott (Reference Walcott1886, Reference Walcott1924, Reference Walcott1925) and refigured by Palmer (Reference Palmer1954) and Sundberg (Reference Sundberg2020). Resser (Reference Resser1935, p. 7) assigned P. kempi to Glyphaspis and later (Resser, Reference Resser1937) to his new species G. nevadensis. Unfortunately, it is uncertain that the type of P. kempi and the type of G. nevadensis are the same specimen. Resser (Reference Resser1935) named Alokistocare packi based on Walcott’s specimens (1886, pl. 26, fig. 2a; pl. 28, fig. 1c, d) with little description. These specimens (see Sundberg, Reference Sundberg2020) are equivalent to Piochaspis sellata Levi-Setti, Reference Levi-Setti1993, which is here reassigned to Piochaspis packi. This complexity is reflected in the synonymy above.
Specimens of Amecephalus piochensis have not been reported from the Grand Canyon and region; however, specimens have been found in the Bright Angel Formation at Diamond Bar Ranch, Rampart Falls? (Alokistocare sp. indet., see below), Columbine Falls (Acrocephalops? arizonaensis, see below), Peach Springs Wash, Fossil Canyon, Lava Falls, and near the Little Colorado River camp. The specimens from Diamond Bar Ranch section are similar to the Am. piochensis specimens from the Pioche Shale illustrated by Sundberg (Reference Sundberg2020, figs. 22, 27). An exception is the librigena assigned to the species here, which has a more laterally projected genal spine. Specimens from Fossil Canyon and near the Little Colorado River, including articulated specimens, have cranidial and librigenal characteristics typical of Am. piochensis. Only small and poorly preserved specimens were found at the Peach Springs Wash section (mile 225) and cannot be assigned with confidence to this species.
A single cranidium possibly belonging to Am. piochensis was reported by Resser (Reference Resser1945) as Alokistocare sp. undet. (USNM 793992). This specimen is from the Flour Sack Member (USNM loc. 44344A) and would be equivalent to the material found at Diamond Bar Ranch. The placement of the specimen is questioned as a result of significant damage (Fig. 72.7) after Resser photographed the cranidium.
Acrocephalops? arizonaensis Resser, Reference Resser1945 (USNM 793986 from USNM loc. 44345F) is also from the Flour Sack Member and is here placed within Am. piochensis. The illustrated specimen (Fig. 72.1) differs from the specimens from the Pioche Shale due to its greater compression in shale, producing a more defined anterior border. Breakage of the palpebral lobes due to the movement of the librigenae below the cephalon, which probably occurred during compaction, gives the appearance of narrower intraocular area on the articulated specimens.
Amecephalus scapegoatensis (Deiss, Reference Deiss1939)
Amecephalus scapegoatensis (Deiss, Reference Deiss1939) from the Glossopleura walcotti Biozone, Muav Formation, Grand Canyon, Arizona; all specimens preserved in limestone. (1) Latex cast of partial cranidium FHPR 19533 from FHPR loc. 006-22 illustrating double border and granules in frontal area; (2) large librigena GRCA 8572 from M&R loc. 44342B; (3) ventral view of partial librigena FHPR 19536 from FHPR loc. 006-22; (4, 5) small weathered cranidium FHPR 19537 from FHPR loc. 006-22 questionably assigned to the species, oblique and dorsal views showing down-sloping frontal area; (6) partial exfoliated cranidium GRCA 8576 from USNM loc. 44342B; (7) weathered partial cranidium FHPR 19534 from FHPR loc. 006-22 showing faint granules in frontal area; (8) weathered, mostly testate cranidium FHPR 19535 from FHPR loc. 006-22 with granules on glabella. Scales: (1, 4–8) 2-mm scale bar; (2, 3) 4-mm scale bar.

Figure 73. Long description
A multi-panel collection of eight grayscale photographs of trilobite fossils.
1. Top left: A latex cast of a partial cranidium (F H P R 19533) showing a double border and granular texture in the frontal area. A 2-mm scale bar is positioned below it.
2. Top center: A large librigena (G R C A 8572) showing a curved, elongated shape. A 4-mm scale bar is positioned to the right.
3. Top right: A ventral view of a partial librigena (F H P R 19536).
4. Middle right: A small weathered cranidium (F H P R 19537) in an oblique view, showing a down-sloping frontal area. A 2-mm scale bar is shared with panels 5 through 8.
5. Far right: The same small cranidium (F H P R 19537) in a dorsal view.
6. Bottom left: A partial exfoliated cranidium (G R C A 8576) showing the central glabella and surrounding fixed cheeks.
7. Bottom center: A weathered partial cranidium (F H P R 19534) with faint granules visible in the frontal area.
8. Bottom right: A weathered, mostly testate cranidium (F H P R 19535) with distinct granules on the glabella.
Reference Deiss1939 Alokistocare? scapegoatensis Deiss 1939 p. 70, pl. 12, fig. 5.
Holotype
Cranidium T1006, Geological Museum, Montana State University, Damnation Limestone, Montana.
Occurrence
Glossopleura walcotti Biozone, Rampart Canyon Member, Muav Formation, Diamond Bar Ranch, Grand Wash Cliffs; Meriwitica Canyon, Arizona. Glossopleura walcotti Biozone, Damnation Limestone, Scapegoat Basin and near Dearborn River, Montana (Deiss, Reference Deiss1939; Schwimmer, Reference Schwimmer1973).
Remarks
All specimens are weathered, incomplete and preserved in limestone. However, specimens show the frontal area with an elongated medial swelling, paradoublural line and border furrow, and coarse, scattered granules. The holotype and additional material were figured by Schwimmer (Reference Schwimmer1973). Granules are also visible on the intraocular region of the fixigena (Fig. 73.1, 71.7) and possibly the glabella (Fig. 73.8). A single small specimen questionably assigned to the species (Fig. 73.4, 73.5) has a faint medial swelling and a strongly down-sloping frontal area.
Amecephalus spp. indet.
Occurrence
Glossopleura walcotti Biozone, Bright Angel Formation to Rampart Cave Member, Muav Formation, Blacktail Canyon, Tapeats Creek, River Mile 200.8, Columbine Falls, and Rampart Cave, Grand Canyon, Arizona.
Remarks
Several localities have fragmented specimens of Ameceph-alus , which may represent one or more species. These are left in open nomenclature until additional specimens are discovered. This includes specimens referred to as Alokistocare sp. or sp. undet. by McKee and Resser (Reference McKee1945).
Genus “Antagmus” Resser, Reference Resser1936
Type species
Ptychoparia teucer (Walcott, Reference Walcott1886), from the Dyeran (Cambrian Stage 4), Georgia Formation, Vermont.
Remarks
Recognition of the genus is presently not possible due to the type specimens being poorly preserved cranidia and the lack of associated sclerites (Sundberg Reference Sundberg, Landing, Milukic and Kluessendorf2007; Sundberg and Webster, 2022) and should be considered nomen dubium.
“Antagmus” arizonaensis Resser, Reference Resser1945
Lower Cambrian ptychoparioids from the Nephrolenellus multinodus Biozone, Tapeats–Bright Angel transition, Grand Canyon, Arizona; all specimens are in sandstone. (1–4) “Antagmus” arizonaensis Resser, Reference Resser1945, silicon cast of holotype cranidium USNM 794187 (Resser, Reference Resser1945, pl. 19, fig. 27) from USNM loc. 44343A, dorsal, anterior, slightly oblique lateral, and lateral anterior. (5, 6) Ptychoparioid sp. indet. 1, silicon cast of paratype cranidium USNM 794188 of “Antagmus” arizonaensis (Resser, Reference Resser1945, pl. 19, fig. 28) from USNM loc. 44345A, dorsal and anterior views.

Figure 74. Long description
A multi-panel photo displays silicon casts of trilobite cranidia. A central scale bar indicates 2 mm.
* Panel 1: Top left. Dorsal view of Antagmus arizonaensis holotype U S N M 794187, showing a rounded glabella and wide fixed cheeks with a granular texture.
* Panel 2: Top right. Anterior view of the same holotype, highlighting the convex elevation of the glabella above the frontal area.
* Panel 3: Middle right. Slightly oblique lateral view of the holotype, showing the profile of the glabella and the curvature of the palpebral lobes.
* Panel 4: Middle right, below panel 3. Lateral anterior view of the holotype, providing a low-angle perspective of the frontal margin.
* Panel 5: Bottom left. Dorsal view of Ptychoparioid species indet. 1, paratype U S N M 794188. It features a more pronounced, bulbous glabella and deeply defined furrows compared to the first specimen.
* Panel 6: Bottom right. Anterior view of the same paratype, showing the steep sides of the glabella and the horizontal extension of the fixigenae.
Reference Resser1945 Antagmus arizonaensis Resser, p. 205, pl. 19, fig. 27, (not fig. 28 = Ptychoparioid sp. indet. 1).
Holotype
Cranidium USNM 794187 (Fig. 74.1–74.4) from USNM loc. 44345G, Tapeats Sandstone, Quartermaster Canyon, Grand Canyon, Arizona.
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, Quartermaster Canyon, Grand Canyon, Arizona.
Remarks
This species is represented by a single, moderately preserved, external mold of a cranidium preserved in sandstone. No additional cranidia or associated sclerites are known for this species, thus the species should be considered nomen dubium and the name restricted to the holotype. Resser (Reference Resser1945) assigned a second cranidium (Fig. 74.5, 74.6) from USNM loc. 44345A as a paratype to the species; however, the partial specimen has a wider and more convex interocular portion of the fixigena, longer preglabellar area, and less pronounced ocular ridges. This specimen has been assigned to Ptychoparioid sp. indet. 1 (see below).
Genus Glyphaspis Poulsen, Reference Poulsen1927
Type species
Asaphicus capella Walcott, Reference Walcott1916b, from the Topazan (Wuliuan Stage), Wolsey Shale, Montana.
Remarks
An ongoing difficulty with the separation or lumping of Blainia Walcott, Reference Walcott1916b, and Glyphaspis Poulsen, Reference Poulsen1927 (see reviews by Bordonaro et al., Reference Bordonaro, Pratt and Robledo2013, and Peel, Reference Peel2020) makes it difficult to place these following species into one or the other genus. Bordonaro et al. (Reference Bordonaro, Pratt and Robledo2013) proposed that Blainia gregaria Walcott, Reference Walcott1916b, the type species of Blainia, is an extremely variable species and synonymized 30 species with B. gregaria, including the species of Glyphaspis reported from the Grand Canyon. Unfortunately, there are several problems with Bordonaro et al.’s (Reference Bordonaro, Pratt and Robledo2013) proposal: (1) as pointed out by Peel (Reference Peel2020), the materials illustrated by Bordonaro et al. (Reference Bordonaro, Pratt and Robledo2013) are only presented in dorsal view and many of the specimens are incomplete; (2) also pointed out by Peel (Reference Peel2020), the type material of B. gregaria from Alabama was not re-illustrated or redescribed; (3) the type material of B. gregaria needs to be restudied to ascertain if one or more species is present and what morphological features are present (e.g., is the axial furrow relatively deep as in USNM 62797, or shallow as in USNM 62802; is a genal spine present as in USNM 62797, or absent as in USNM 62802); and (4) Bordonaro et al. (Reference Bordonaro, Pratt and Robledo2013) collected over 2000 specimens, but these specimens were derived from at least 155 m of section and there is no information as to where the studied or figured specimens were derived from this interval. In addition, none of the illustrated specimens of Bordonaro et al. (Reference Bordonaro, Pratt and Robledo2013) has the nearly parallel sided glabella or the well-defined pygidial border furrow as present in the type material (Schwimmer, Reference Schwimmer1989). As a result, the synonymy of the two genera is still in flux and until the type species of Blainia is better understood, the retention of taxa in Glyphaspis is maintained.
The familial assignment of Glyphaspis is complicated by the assignment of the species to Blainia discussed above. Harrington et al. (Reference Harrington, Henningsmoen, Howell, Jaanusson, Lochman-Balk, Part and Moore1959) assigned Glyphaspis to the Superfamily Anomocaridae Poulsen, Reference Poulsen1927, and Blainia to Superfamily Asaphiscacea Raymond, Reference Raymond1924. Adrain (Reference Adrain2011) placed the latter (as Family Asaphiscidae Raymond, Reference Raymond1924) into the Order Olenida Adrain, Reference Adrain2011, whereas he placed the former (as the Family Anomocareidae Poulsen, Reference Poulsen1927) into “Order Uncertain”.
Glyphaspis? kwaguntensis Resser, Reference Resser1945
Glyphaspis spp. from the Ehmaniella subzone, Ehmaniella Biozone, Peach Springs Member, Muav Formation, Grand Canyon, Arizona. (1–12, 14) Glyphaspis? kwaguntensis Resser, Reference Resser1945, from USNM loc. 73b, all specimens are preserved in sandy limestone: (1) paratype cranidium USNM 794103 (Resser, Reference Resser1945, pl. 26, fig. 2) with thoracic segment crossing glabella; (2) cranidium USNM 794112c (Resser, Reference Resser1945, pl. 25, fig. 13, bottom center); (3) small pygidium USNM 794110a (Resser, Reference Resser1945, pl. 26, fig. 8, lower right corner); (4–6) paratype pygidium USNM 794102 (Resser, Reference Resser1945, pl. 26, fig. 3, bottom right), dorsal, posterior, and lateral views; (7) smaller cranidium USNM 794104; (8) small pygidium USNM 794110b; (9–11) holotype pygidium USNM 794101 (Resser, Reference Resser1945, pl. 26, fig. 6), dorsal, posterior, and lateral views; (12) previously unfigured pygidium USNM 794112e; (14) previously unfigured pygidium USNM 794108. (13, 15) Glyphaspis tecta Resser, Reference Resser1945, from USNM loc. 44338F, (13) holotype pygidium USNM 794055c (Resser, Reference Resser1945, pl. 22, fig. 15, bottom left); (15) paratype pygidium USNM 794055d (Resser, Reference Resser1945, pl. 22, fig. 15, top right). Scales: (1, 2, 3–7, 9–15) 5-mm scale bar; (3, 8) 2-mm scale bar.

Figure 75. Long description
A collection of 15 black and white photographs of trilobite fossils preserved in sandy limestone.
* Panel 1: Paratype cranidium of Glyphaspis? kwaguntensis showing a thoracic segment crossing the glabella.
* Panel 2: A larger cranidium with a distinct rounded glabella.
* Panel 3: A small pygidium with a visible central axis.
* Panels 4, 5, and 6: A paratype pygidium shown in dorsal, posterior, and lateral views respectively, illustrating its three-dimensional convex shape.
* Panel 7: A smaller cranidium.
* Panel 8: A small pygidium. A 2 m m scale bar is located between panels 5 and 8.
* Panels 9, 10, and 11: The holotype pygidium of Glyphaspis? kwaguntensis shown in dorsal, posterior, and lateral views. It features a broad, fan-like shape with a tapering central axis.
* Panel 12: A previously unfigured pygidium.
* Panel 13: Holotype pygidium of Glyphaspis tecta, showing a wider, more semi-circular margin.
* Panel 14: A large, previously unfigured pygidium of Glyphaspis? kwaguntensis.
* Panel 15: Paratype pygidium of Glyphaspis tecta. A 5 m m scale bar is positioned above panel 15.
The specimens are mostly light gray against a slightly darker, textured rock matrix. The fossils show varying degrees of surface detail, including pleural ribs on the pygidia and the defined borders of the cranidia.
Reference Resser1945 Glyphaspis kwaguntensis Resser, p. 199, pl. 25, figs. 12, 13; pl. 26, figs. 2–4.
Holotype
Cranidium USNM 794101 (Fig. 75.11) from USNM loc. 73b, Bright Angel Formation or Muav Formation, Nankoweap Valley, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794103 and 794104; pygidia USNM 794102, 794108, 794110a, b, and 794112c, e from USNM loc. 73b, Bright Angel Formation or Muav Formation, Nankoweap Valley, Grand Canyon, Arizona.
Occurrence
Ehmaniella Biozone, upper Bright Angel Formation or Muav Formation, Kwagunt Valley, Grand Canyon, Arizona.
Remarks
Specimens of Gl.? kwaguntensis are found on weathered surfaces of “impure limestone” and are not well preserved. Resser (Reference Resser1945, p. 199) stated “In its tumid axial region and lack of a well-defined concave posterior border, this species is unlike any yet described.” Overall, the cranidia and pygidia of Gl.? kwaguntensis and Gl. vulsa are similar, although similarities and differences are difficult to assess due to the former’s preservation, different sizes of specimens (e.g., pygidia of Gl. vulsa are 2.0–6.5 mm in length vs. Gl.? kwaguntensis range of 11.5–15.0 mm) and potential ontogenetic changes. One unique character state of Gl.? kwaguntensis is the cranidial anterior border furrow being nearly straight (164–176°) as opposed to the evenly curved furrow in Gl. vulsa (135–139°).
Some of the specimens of Gl.? kwaguntensis illustrate similarities to Asaphiscus wheeleri Meek, Reference Meek1873. One cranidium (Fig. 75.1) shows a similar glabellar outline, narrow intraocular region, and anterior border curvature, but lacks the relatively small palpebral lobes and well-defined anterior border furrow. The pygidia (Fig. 75.4–75.6, 75.9–75.12) have similar outlines of the pygidium, axis, and pleural fields, but differ in a poorly defined border furrow, wider border furrow, and shorter axis. These similarities could suggest that Gl.? kwaguntensis may actually belong to Asaphiscus. Given the poor preservation of the type material, it is suggested here that the species be restricted to the type material and left in open generic nomenclature.
Glyphaspis tecta Resser, Reference Resser1945
Reference Resser1945 Glyphaspis tecta Resser, p. 198–199, pl. 22, fig. 15.
Holotype
Pygidium USNM 794055c (Fig. 75.13) from USNM loc. 44338G, Gateway Canyon Member, Muav Formation, Havasupai Gardens, Grand Canyon, Arizona.
Paratype
Pygidium USNM 794055d (Fig. 75.15) from USNM loc. 44338G, Gateway Canyon Member, Muav Formation, Havasupai Gardens, Grand Canyon, Arizona.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Gateway Canyon Member, Muav Formation, Havasupai Garden, Grand Canyon, Arizona.
Remarks
A single large pygidium (13 mm in length) of Gl. tecta was illustrated by Resser (Reference Resser1945), although he did report the species from localities 44338F and 44338G and a paratype specimen. Preservation of the two type specimens on the same slab is poor and there are no cranidia assigned to this species to help evaluate the taxon. One unique character state of Gl. tecta is the relatively narrow pygidial axis compared to the pygidial width (about 20%) as opposed to Gl.? kwaguntensis and Gl. vulsa (about 25–30%), although this difference may be a result of specimen flattening.
Given the poor preservation of the type material, it is suggested here that the species be restricted to the type material.
Glyphaspis tetonensis Resser, Reference Resser1937
Glyphaspis tetonensis Resser, Reference Resser1937, from the Bolaspidella Biozone, Gateway Canyon and Havasu members, Muav Formation, Grand Canyon, Arizona: (1) partial cranidium GRCA 125557 from GRCA loc. 357-4; (2) latex cast of partial cranidium GRCA 125451 from GRCA loc. 357-5; (3) partial cranidium GRCA 125448 from GRCA loc. 357-5; (4) partial cranidium GRCA 125433d from GRCA loc. 357-4; (5) latex cast of partial cranidium GRCA 125445 from GRCA loc. 357-5; (6, 7) latex cast of partial cranidium GRCA 125444 from GRCA loc. 357-5, dorsal and oblique views; (8) poorly preserved, partial cranidium GRCA 125132 from GRCA loc. 352; (9, 10) latex cast of partial cranidium GRCA 125129 from GRCA loc. 352, dorsal and oblique views; (11–13) partial cranidium GRCA 125136 from GRCA loc. 352, dorsal, lateral, and anterior views; (14) librigena GRCA 125137 from GRCA loc. 352; (15) librigena GRCA 125130 from GRCA loc. 352; (16) librigena GRCA 125138 from GRCA loc. 352; (17) librigena GRCA 125141 from GRCA loc. 352; (18) pygidium GRCA 125447 from GRCA loc. 357-5; (19) pygidium GRCA 125450 from GRCA loc. 357-5; (20–22) partial pygidium GRCA 125134 from GRCA loc. 352, dorsal, lateral, and posterior views; (23, 24, 26) pygidium GRCA 125135 from GRCA loc. 352, posterior, lateral, and dorsal views; (25) latex cast of pygidium GRCA 125446c from GRCA loc. 357-5; (27) latex cast of pygidium GRCA 125446a from GRCA loc. 357-5, (28) latex cast of pygidium GRCA 125446b from GRCA loc. 357-5. Scales: (1–3) 2-mm scale bar; (4, 5, 8–28) 4-mm scale bar; (6, 7) 4-mm black bar in photos.

Figure 76. Long description
A collection of 28 monochrome photographs of Glyphaspis tetonensis fossils.
* Panels 1 through 13 show cranidia (head shields). Panels 1, 2, and 3 are small specimens with a 2 mm scale bar. Panels 4 through 10 show larger cranidia or latex casts, including dorsal and oblique views (6, 7 and 9, 10). Panels 11, 12, and 13 provide dorsal, lateral, and anterior views of a single specimen, showing a highly convex glabella and a 4 mm scale bar.
* Panels 14 through 17 display librigenae (free cheeks), which are crescent-shaped with a distinct border and a long genal spine.
* Panels 18 through 28 show pygidia (tail shields). These are semi-circular with a central axis and radiating pleural ribs. Panels 20, 21, and 22 show dorsal, lateral, and posterior views of one specimen. Panels 23, 24, and 26 show posterior, lateral, and dorsal views of another. Panels 25, 27, and 28 are latex casts of pygidia showing fine surface details.
Scale bars are provided: 2 mm for the first three panels and 4 mm for the remainder.
Reference Miller1936 Glyphaspis cf. G. perconcava; Miller, p. 30, p1. 8, fig. 17.
Reference Resser1937 Glyphaspis tetonensis Resser, p. 13.
Reference Lochman and Hu1960 Americare tetonensis; Lochman and Hu, p. 828, p1. 100, figs. 42–57.
Reference Melzak and Westrop1994 Glyphaspis tetonensis; Melzak and Westrop, p. 979–982, pl. 3, figs. 1–12; p1. 4, figs. 1–10.
Holotype
Pygidium Columbia University 12609 from the Death Canyon Limestone, South Fork, Teton Canyon Creek, Teton Range Wyoming.
Paratype
Cranidium Columbia University 12609a from the Death Canyon Limestone, South Fork, Teton Canyon Creek, Teton Range Wyoming.
Occurrence
Bolaspidella Biozone, Havasu and Gateway members, Muav Formation, Buck Farm Canyon and Nankoweap Canyon, Grand Canyon, Arizona. “Americare subzone” (= Bolaspidella Biozone) uppermost Death Canyon Limestone and Park Shale, Teton Range, Wyoming, (Lochman and Hu, Reference Lochman and Hu1960); Glyphaspis tetonensis fauna (= Bolaspidella Biozone) Pika Formation, Chaba River, Alberta, (Melzak and Westrop, Reference Melzak and Westrop1994).
Remarks
The specimens from the Grand Canyon are very similar to those preserved in limestone illustrated by Lochman and Hu (Reference Lochman and Hu1960) from Wyoming and by Melzak and Westrop (Reference Melzak and Westrop1994) from Chaba River, although their silicified material is tectonically distorted. The larger cranidium illustrated by Melzak and Westrop (Reference Melzak and Westrop1994, pl. 4, fig.1) is compressed, but the smaller cranidia appear to be comparable in convexity to the specimens from the Grand Canyon.
Glyphaspis vulsa Resser, Reference Resser1945
Glyphaspis vulsa Resser, Reference Resser1945, from the Ehmaniella subzone, Ehmaniella Biozone, Peach Springs Member, Muav Formation, Grand Canyon, Arizona, all specimens preserved in limestone, mostly exfoliated and from Nankoweap section, GRCA loc. 356 unless otherwise noted. (1) Holotype cranidium USNM 794056 (Resser, Reference Resser1945, pl. 23, fig. 14, top) from USNM loc. 74; (2, 3) cranidium GRCA 125280a, external mold inverted, external; (4) paratype pygidium USNM 794061 (Resser, Reference Resser1945, pl. 23, fig. 12) from USNM loc. 74; (5) pygidium GRCA 125288; (6, 7) cranidium GRCA 125281a, dorsal and oblique views; (8) cranidium GRCA 125287; (9) pygidium GRCA 125286; (10, 11) pygidium GRCA 125267a, dorsal and lateral views; (12) pygidium GRCA 125281b; (13) pygidium GRCA 125289; (14) testate librigena GRCA 125285; (15–17) pygidium GRCA 125282, dorsal, lateral, and posterior views; (18) pygidium GRCA 125283; (19) pygidium GRCA 125280b illustrating terrace lines on doublure; (20) librigena GRCA 125281d; (21) pygidium GRCA 125284a; (22) librigena GRCA 125281c.

Figure 77. Long description
The collection consists of 22 panels labeled with numbers in the top-right or top-left corners. A 2 mm scale bar is located between panels 13 and 17.
* Panel 1: Large dorsal view of a cranidium with a wide, rounded anterior margin.
* Panels 2 and 3: External mold and dorsal view of a cranidium showing a distinct glabella.
* Panel 4: Dorsal view of a semi-circular pygidium with a segmented axis.
* Panel 5: Pygidium with a damaged central axis.
* Panels 6 and 7: Dorsal and oblique views of a cranidium showing the three-dimensional curvature of the cephalon.
* Panel 8: Small dorsal cranidium.
* Panel 9: Wide pygidium with clear pleural ribs.
* Panels 10 and 11: Dorsal and lateral profile views of a pygidium, highlighting its convex shape.
* Panels 12 and 13: Two small, transverse pygidia.
* Panel 14: Elongated, curved librigena (free cheek).
* Panels 15, 16, and 17: Dorsal, lateral, and posterior views of a single pygidium, showing the elevation of the axis and the downward slope of the pleural fields.
* Panel 18: Large, well-preserved pygidium with a smooth border.
* Panel 19: Ventral view of a pygidium showing fine terrace lines on the doublure.
* Panel 20: Librigena with a long genal spine.
* Panel 21: Pygidium with a weathered central axis.
* Panel 22: Small librigena fragment.
Reference Resser1945 Glyphaspis vulsa Resser, p. 198, pl. 22, figs. 12–14.
Holotype
Cranidium USNM 794056 (Fig. 77.1) from USNM loc. 74, Bright Angel Formation, Nankoweap Valley, Grand Canyon, Arizona.
Paratypes
Cranidia USNM 794057, 794059, and 794060 and pygidia USNM 794058 and 794061 from USNM loc. 74, Bright Angel Formation, Nankoweap Valley, Grand Canyon, Arizona.
Emended diagnosis
Cranidium with low convexity, moderate length, evenly tapered glabella, frontal lobe bluntly rounded; moderately wide (tr.) anterior margin, moderately long frontal area; slightly concave, wide anterior border, shallow anterior border furrow. Librigena with narrower lateral border and shorter spine. Pygidium relatively transversely elongate, axis reaches margin of pleural field or extends onto border, faint postaxial ridge, weaker pleural furrows and band extend onto the border to nearly the margin of the pygidium, posterior lateral corners adjacent to the anterior portion of the pygidium. Exoskeleton with very fine granular surface.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Peach Springs and Kanab Canyon members, Muav Formation, River Mile 50, Nankoweap Valley, and Blacktail Canyon, Grand Canyon, Arizona.
Emended description
Cranidium 5.8 ± 2.6 mm long (n = 4); subsquare; anterior border width 78 ± 8% cranidial width, cranidial length 50 ± 4% cranidial width; moderately low convexity (sag. and trans.); anterior margin evenly curved, slightly concave; posterior margin, excluding occipital ring, distally curved slightly posterolaterally. Anterior branches of facial sutures divergent to anterior border; nearly parallel across anterior border; posterior branches strongly divergent. Glabella moderately elongated, length 69 ± 4% cranidial length; width 40 ± 5% cranidial width, moderately tapered, width at anterior end 68 ± 5% glabellar width; moderate convexity (sag. and trans.); frontal lobe rounded. Axial furrow moderately deep, slightly deeper medially, slightly constricted at L2; preglabellar furrow moderately shallow. Lateral glabellar furrows shallow to effaced, S1 directed slightly posterior; S2 directed slightly posterior; S3 projected laterally, S4 projected slightly anteriorly very faint or absent. Occipital ring slightly elevated above glabella, moderate convexity; small occipital node; length 21 ± 3% glabellar length; posterior margin convex posteriorly, slightly curved medially. SO gently curved anteriorly, moderately deep and arched anteriorly and deeper. Frontal area unequally divided; length 31 ± 4% cranidial length. Preglabellar field slightly convex, moderately down-sloping, length 30 ± 10% frontal area length. Anterior border slightly concave and level, uniformly curved, length 22 ± 2% cranidial length. Anterior border furrow shallow, shallower medially, shallower than axial furrows. Fixigena very slightly convex, level, anterior area moderately down-sloping; width 46 ± 9% glabellar width. Palpebral lobes moderately curved, sloping up, narrow, moderately long, length 40 ± 2% glabellar length; anterior margin located opposite L2. Ocular ridge moderate strength, slightly curved, directed moderately posterolaterally from glabella at 64 ± 3° to axis. Posterior area of fixigena strap-like, sharp termination; length (exsag.) 23 ± 5% glabellar length; width 63 ± 5% glabella length.
Librigenae 6.0–8.6 mm long excluding spine (n = 3); wide, width 54 ± 7% length without spine; lateral margin moderately curved. Genal field moderately convex. Border slightly convex, wider posteriorly, width about 37 ± 5% librigenal width; border furrows moderate, shallower posterior. Librigenal spine moderately long, 50–70% librigenal length, slightly curved, broad shallow furrow posteriorly, flattened.
Rostral plate, hypostome, and thorax unknown.
Pygidium 2.3–6.5 mm long (n = 11); suboval, length 59 ± 3% width; margin smooth, anterior margin curved slightly posterolaterally approximately half-way to anterolateral corners; anterolateral corners rounded, adjacent to anterior portion of axis; very slight posteromedial notch; moderately convex (sag.). Axis anterior width 27 ± 2% pygidial width; length 78 ± 4% pygidial length; postaxial ridge very slight; moderately tapered, medial width 81 ± 3% anterior width, five to six axial rings, moderately convex; terminal axial piece moderate size, rounded; axial ring furrows shallow. Pleural regions moderately curved laterally; pleural furrows and bands moderately developed and extend on the anterior border almost to pygidial margin. Border weakly defined; border furrow very shallow anteriorly.
Exoskeleton with a fine granular surface; preglabellar area and genal area of librigenae covered with genal caeca.
Remarks
The type locality of Gl. vulsa is listed as 300 ft. (92 m) above the base of the Tapeats at the head of Nankoweap Valley (Resser, Reference Resser1945, p. 172) and is listed to occur with several representatives of the G. walcotti and Ehmaniella biozones. This suggests that locality 74 consists of an assemblage from mixed horizons. Glyphaspis vulsa has been found in the same section in situ in the Peach Springs Member, Muav Formation at about 185 m above the base of the Tapeats.
Glyphaspis vulsa is similar to Gl. tetonensis in their granular ornamentation, glabellar outline, and relatively shorter preglabellar area and pygidial convexity. However, the two species differ in the latter’s: (1) cranidia having a higher convexity, more rounded glabellar frontal lobe, narrower interocular area; (2) librigena with wider lateral border and longer genal spine; and (3) pygidia with distinct anterior pleural bands and pleural furrows extending on to near the pygidial margin. Most pygidia of Gl. tetonensis have the anterior margins strongly curved posterolaterally with the posterolateral corners located adjacent to near the terminal piece.
Glyphaspis spp. indet.
Miscellaneous ptychoparioids. (1–3) Kochina angustata Resser, Reference Resser1945, holotype cranidium USNM 794062 (Resser, Reference Resser1945, pl. 26, fig. 11), from USNM loc. 75, Glossopleura walcotti Biozone?, Peach Springs Member? see discussion under species, Muav Formation, Grand Canyon, Arizona, dorsal, lateral, and anterior views. (4) Glyphaspis sp. indet., from the Ehmaniella subzone, Ehmaniella Biozone, Gateway Canyon Member, Muav Formation, Grand Canyon, Arizona; inverted photograph of external mold of cranidium GRCA 125242 from GRCA loc. 361-2. (5) Piochaspis nites (Resser, Reference Resser1945), cranidium USNM 794034 (Resser, Reference Resser1945, pl. 27, fig. 35), from USNM loc. 44344A, Glossopleura walcotti Biozone, Flour Sack Member, Bright Angel Formation, Grand Canyon, Arizona. (6) Kochina vestita Resser, Reference Resser1939b, partial cranidium GRCA 125510 preserved in shale from GRCA loc. 375, Bright Angel Formation, Grand Canyon. Scales: (1–3, 5, 6) 2-mm scale bar; (4) 2-mm scale bar in photo.

Figure 78. Long description
A multi-panel collection of six photographs showing fossilized trilobite cranidia.
* Panel 1: Top-left. A dorsal view of Kochina angustata holotype U S N M 794062, showing a rounded, shield-like head section with a prominent central glabella.
* Panel 2: Top-right. A lateral view of the same Kochina angustata specimen, highlighting the convex profile and elevation of the glabella.
* Panel 3: Middle-right, below panel 2. An anterior view of the Kochina angustata specimen, showing the frontal curvature.
* Panel 4: Middle-left. An inverted photograph of an external mold of Glyphaspis species indet G R C A 125242. It features a 2 mm scale bar in the top-left corner. The fossil shows a wide, flared cranidium with distinct ocular ridges.
* Panel 5: Middle-right, below panel 3. A dorsal view of Piochaspis nites U S N M 794034. A 2 mm scale bar is positioned below this panel. The specimen has a more elongated, narrow glabella compared to the others.
* Panel 6: Bottom. A large, partial cranidium of Kochina vestita G R C A 125510 preserved in textured shale, showing the right side of the fossilized head section.
All specimens are preserved in rock matrix with varying degrees of surface texture and relief.
Occurrence
Ehmaniella subzone, Ehmaniella Biozone, Kanab Canyon and Gateway Canyon members, Muav Formation, Surprise Canyon, Malgosa Canyon, Lava Canyon, Lone Tree Canyon, Havasupai Gardens, Grand Canyon, Arizona.
Remarks
Resser (Reference Resser1945) reported but did not describe or illustrate Glyphaspis sp. from the Gateway Canyon Member, Muav Formation from GRCA loc. 362-2 (Lava Canyon), USNM loc. 44338E (west side, Havasupai Gardens), USNM loc. 44338H (west of Havasupai Gardens), and GRCA loc. 356 (Lone Tree Canyon). Poorly preserved material in the GRCA museum from USNM locs. 44338E (32 ft, 9.8m above the base; GRCA 8355, 8405, 8406) and 44338H (97 ft, 29.6m; GRCA 8401–8412) west of Havasupai Gardens may represent possible Glyphaspis cranidium, librigena, and pygidia and Kootenia pygidia. Material from GRCA loc. 365, west of Lone Tree Canyon (35 ft, 10.7 m; GRCA 2187) above the base of the Muav Formation is poorly preserved in siltstone and may represent Glyphaspis cranidia and pygidia. Resser (Reference Resser1945) also reported specimens of Glyphaspis from the Flour Sack Member, Bright Angel Formation at Columbine Falls (USNM loc. 44344B) and Rampart Cave (USNM loc. 44345F) that probably are pygidia of Athabaskia (see discussion under this taxon).
Additional specimens of Glyphaspis were found during this study. Two cranidia resemble G. robusta Deiss, Reference Deiss1939, in its broad anterior border, well-defined ocular ridge, and tapered glabella, and thoracic segments are known only from GRCA loc. 361-2 (Fig. 78.4) in the Gateway Canyon Member; however, a pygidium is needed to properly identify the species. A single librigena similar to that of G. vulsa was found from the upper Kanab Canyon Member from GRCA loc. 361-1; and an articulated, but poorly preserved specimen from Surprise Canyon (GRCA loc. 275).
Genus Kochina Resser, Reference Resser1935
Type species
Olenopsis americanus Walcott, Reference Walcott1912, from the Delamaran (Wuliuan Stage), Gordon Shale, Montana.
Kochina angustata Resser, Reference Resser1945
Reference Resser1945 Kochina? angustata Resser, p. 212, pl. 23, fig. 3.
Holotype
Cranidium USNM 794062 (Fig. 78.1–78.3) from USNM loc. 75, Peach Springs Member, Muav Formation, Kanab Canyon, Grand Canyon, Arizona.
Occurrence
Ehmaniella Biozone, Bright Angel Formation or Muav Formation, Kanab Canyon. Grand Canyon, Arizona.
Remarks
Resser (Reference Resser1945) questioned the placement of the species into Kochina; however, the evenly tapered glabella, relatively narrow fixigenae, narrow occipital ring, and a distinctly posteriorly curved anterior border is characteristic of the genus. This species is most similar to Ko. vestita Resser, Reference Resser1939b (also see Sundberg, Reference Sundberg, Landing, Milukic and Kluessendorf2007; Fig. 78.4) in the curvature of the anterior border furrow and flattened anterior border, but differs in the narrower (tr.) width of the intraocular fixigena and anterior border.
Kochina is typically a member of the G. walcotti Biozone (see Rasetti, Reference Rasetti1951; Sundberg, Reference Sundberg, Landing, Milukic and Kluessendorf2007); however, the associated taxa listed by Resser (Reference Resser1945) at USNM loc. 75 are typically members of the Ehmaniella Biozone (Sundberg, Reference Sundberg1994). This mixed biozonal indication may be the result of the mixing sample horizons to generate the faunal list for this locality. This assertion is suggested by the lithology of the matrix of different taxa. The single specimen of Ko. angustata is preserved in a sandstone at USNM loc. 75, which is in contrast to the other taxa reported by Resser at this location. Wenkchemnia kanabensis, Spe.? diligens, and Kootenia mckeei are in a fine-grained limestone, whereas Eh. nitida is preserved in a sandy limestone.
Kochina vestita Resser, Reference Resser1939b
Reference Resser1939b Kochina vestita Resser, p. 53–54, pl. 12, figs. 7, 8.
Reference Gunther and Gunther1981 Kochina vestita; Gunther and Gunther, Reference Gunther and Gunther1981, pl. 27, figs. a–c.
Reference Sundberg2005 Kochina vestita; Sundberg, p. 66, fig. 6.5.
Holotype
Cranidium USNM 98550 from USNM loc. 54s, Langston Formation, Two Mile Canyon, Utah.
Occurrence
Glossopleura walcotti to ?Ehmaniella biozones, Bright Angel Formation to Sanup Plateau Member, Muav Formation, Keyhole Canyon and ?Peach Springs, Grand Canyon, Arizona. Glossopleura walcotti Biozone, Langston Formation (Resser, Reference Resser1939b); Chisholm Formation, Drum Mountains, Utah (Sundberg, Reference Sundberg2005).
Remarks
A partial shale cranidium (Fig. 78.6) matches the tapered glabellar with well-defined furrows, position of the palpebral lobes, wider intraocular region and laterally tapering anterior border characteristics illustrated by the specimens illustrated by Resser (Reference Resser1939b), Gunther and Gunther (Reference Gunther and Gunther1981), and Sundberg (Reference Sundberg2005).
Genus Mexicella Lochman, Reference Lochman1948
Type species
Mexicella mexicana Lochman, Reference Lochman1948, from the upper Delamaran (Wuliuan Stage), Arrojos Formation, Caborca area, Sonora, Mexico.
Mexicella mexicana Lochman, Reference Lochman1948
Mexicella mexicana Lochman, Reference Lochman1948, from the Mexicella mexicana Biozone, Bright Angel Formation, Rampart Cave, Grand Canyon, Arizona; all specimens preserved in shales and from GRCA loc. 331-1. (1, 2) Cranidium GRCA 125521, internal mold with flattened right side and silicone mold of external surface; (3) librigena GRCA 125254b questionably assigned to the species; (4) cranidium GRCA 125248; (5) cranidium GRCA 125246. Scales: (1, 2) 5-mm scale bar; (3–5) 2-mm scale bar.

Figure 79. Long description
A multi-panel display of five fossil specimens.
Panel 1 (top left): Cranidium G R C A 125521, an internal mold showing a central glabella and a flattened right side.
Panel 2 (top right): A silicone mold of the external surface of the same cranidium G R C A 125521, showing more defined surface texture.
Between the rows is a 5 mm scale bar.
Panel 3 (bottom left): Librigena G R C A 125254b, a triangular cheek piece with a grainy texture.
Panel 4 (bottom center): Cranidium G R C A 125248, a smaller specimen positioned above a 2 mm scale bar.
Panel 5 (bottom right): Cranidium G R C A 125246, showing a well-preserved central glabella and lateral lobes.
All specimens are preserved in shale and exhibit the characteristic rounded, semi-circular head shield structure of the trilobite genus Mexicella.
Reference Lochman1948 Mexicella mexicana Lochman, p. 457, pl. 69, figs. 12–22.
Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952 Mexicella mexicana; Lochman in Cooper et al., p. 150, pl. 24, figs. 1–25.
Reference Palmer and Halley1979 Mexicella mexicana; Palmer and Halley, p. 109, pl. 13, figs. 13–21.
Holotype
Cranidium USNM 115800, Arrojos Formation, Caborca area, Sonora, Mexico.
Paratypes
Cranidia, pygidia, thoracic segments, librigena USNM 115801–115822, Arrojos Formation, Caborca area, Sonora, Mexico.
Occurrence
Mexicella mexicana Biozone, Bright Angel Formation, Rampart Cave, Grand Canyon, Arizona. Mexicella mexicana Biozone, Arrojos Formation, Caborca area, Sonora, Mexico (Lochman, Reference Lochman1948; Lochman in Cooper et al., Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952); Pahrump Hills Shale and Jangle Limestone members, Carrara Formation, southern Nevada and eastern California (Palmer and Halley, Reference Palmer and Halley1979); Grassy Springs Member, Pioche Shale, eastern-central Nevada (Eddy and McCollum, Reference Eddy and McCollum1998).
Remarks
This species is known from specimens flattened in shale in the Bright Angel Formation. Despite their flattened nature, they match well with the limestone specimens illustrated by Lochman (Reference Lochman1948), Lochman in Cooper et al. (Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952), and Palmer and Halley (Reference Palmer and Halley1979). However, the larger specimen (Fig. 79.1, 79.2) has a more equally subdivided frontal area than the smaller cranidia (Fig. 79.4, 79.5). Most of the specimens illustrated by Lochman (in Cooper et al., Reference Cooper, Arellano, Johnson, Okulitch, Stoyanow and Lochman1952, pl. 24) and Palmer (in Palmer and Halley, Reference Palmer and Halley1979, pl. 13, figs. 14–16, 19, 20) are smaller specimens comparable to the smaller specimens illustrated herein. A single librigena is reported here and at first glance appears to be dissimilar to those previously illustrated by possessing a genal spine; however, this “spine” is the posterior portion of the genal area that has been flattened.
Mexicella cf. Me. robusta Sundberg and McCollum, Reference Sundberg and McCollum2000
Mexicella cf. M. robusta Sundberg and McCollum, 2000, from the Bright Angel Formation, Frenchman Mountain, Nevada; all specimens preserved as limonite in a medium-grain glauconitic sandstone from FHPR loc. 008-5. (1–3) Exfoliated cranidium FHPR 19608, dorsal, anterior, and lateral views; (4) exfoliated cranidium FHPR 19607; (5) exfoliated cranidium FHPR 19612; (6) exfoliated cranidium FHPR 19609; (7) partial exfoliated cranidium FHPR 19610; (8) mostly exfoliated cranidium FHPR 19611; (9) exfoliated librigena FHPR 19614 illustrating short, deflective spine; (10) partially testate librigena FHPR 19613; (11) exfoliated pygidium FHPR 19615, distorted on right posterior margin; (12–14) testate pygidium FHPR 19616, dorsal, posterior, and lateral views; (15) damaged, exfoliated pygidium FHPR 19617. Scales: (1–10) 4-mm scale bar; (11–15) 2-mm scale bar.

Figure 80. Long description
The photo plate contains 15 numbered panels showing various trilobite fossil parts.
* Panels 1 through 8 display cranidia (head shields). Panel 1 shows a dorsal view of an exfoliated cranidium with a prominent central glabella. Panels 2 and 3 provide anterior and lateral views of the same specimen, highlighting its convex profile. Panels 4, 5, 6, and 8 show additional exfoliated cranidia in dorsal view, while panel 7 shows a partial specimen.
* Panels 9 and 10 show librigenae (free cheeks). Panel 9 illustrates an exfoliated librigena with a short, outward-pointing spine. Panel 10 shows a partially testate (shell-bearing) librigena.
* Panels 11 through 15 display pygidia (tail shields). Panel 11 is an exfoliated pygidium with a distorted right margin. Panels 12, 13, and 14 show a single testate pygidium from dorsal, posterior, and lateral perspectives respectively. Panel 15 shows a damaged, exfoliated pygidium.
A 4-mm scale bar is positioned between panels 8 and 9, applying to panels 1 through 10. A 2-mm scale bar is located at the bottom right, applying to panels 11 through 15. The fossils appear as light-colored limonite against a dark, granular sandstone matrix.
cf. Reference Sundberg2020 Mexicella robusta Sundberg and McCollum, p. 622–624, fig. 13.1–13.10.
Occurrence
Middle to upper Eokochaspis nodosa Biozone, Bright Angel Formation, Frenchman Mountain, Nevada.
Remarks
The specimens reported here are intermediate between Me. robusta Sundberg and McCollum, Reference Sundberg and McCollum2000, and Me. antelopea Sundberg and McCollum, Reference Sundberg and McCollum2000, in possessing the effaced glabellar furrows and slightly inflated glabella of the former and the shorter (sag.) preglabellar area, more pronounced axial furrows, and a librigenal spine (although shorter) of the latter. Thus, the specimens may represent a new species needing a formal description.
The stratigraphic range of Me. antelopea is from the Comet Shale of the Pioche Shale, Nevada (see Sundberg and McCollum, Reference Sundberg and McCollum2000) and is representative of the middle Eokochaspis nodosa Biozone. Mexicella robusta is also from the Comet Shale, but also from the Susan Duster Limestone of the Pioche Shale, Nevada (see Sundberg and McCollum, Reference Sundberg and McCollum2000) and is representative of the stratigraphically higher Am. arrojosensis Biozone. If Mexicella cf. Me. robusta is part of an evolutionary sequence between Me. antelopea and Me. robusta, then the middle to upper Eo. nodosa Biozone position of this species is justified.
Genus Piochaspis Levi-Setti, Reference Levi-Setti1993
Type species
Piochaspis sellata Levi-Setti, Reference Levi-Setti1993, from the Delamaran (Wuliuan Stage), Chisholm Shale, Nevada.
Piochaspis nites (Resser, Reference Resser1945)
Reference Resser1945 Elrathia nitens Resser, p. 209, pl. 27, fig. 35.
Holotype
Cranidium USNM 794034 (Fig. 78.5) from USNM loc. 44344A, Flour Sack Member, Bright Angel Formation, Columbine Falls, Grand Canyon, Arizona.
Occurrence
Glossopleura walcotti Biozone, Flour Sack Member, Bright Angel Formation, Grand Canyon, Arizona.
Remarks
The single, poorly preserved, small cranidium is very similar to the cranidia of P. sellata Levi-Setti, Reference Levi-Setti1993, from the equivalent age in the Chisholm Shale, Nevada and may be conspecific. The tapered glabella, narrow intraocular region, relatively long palpebral lobes, frontal area construction, and slightly divergent anterior sutures of P. nites are very similar to P. sellata. The difference between the two species is the swollen anterior border of the latter.
Resser (Reference Resser1945) also reported Elrathia sp. from several localities in the upper Bright Angel Formation at Havasupai Gardens and Rampart Cave, Rampart Cave to Peach Springs members, Muav Formation at Peach Springs Canyon and Rampart Cave. These specimens were not found in the USNM or GRCA collections and it is unknown if they represent P. nites and or another ptychoparioid taxon.
Genus Unassigned
Remarks
There are several specimens of ptychoparioids that are not identifiable due to preservation, these specimens are referred to as ptychoparioid sp. indet. Only a single species is discussed below due to Resser’s (Reference Resser1945) original assignment of the taxon to Antagmus arizonaensis.
Ptychoparioid sp. indet. 1
Reference Resser1945 Antagmus arizonaensis Resser, p. 205, pl. 19, fig. 28 (not fig. 27 = “Antagmus” arizonaensis).
Occurrence
Nephrolenellus multinodus Biozone, Transition Zone, Tapeats Sandstone, Rampart Cave, Grand Canyon, Arizona.
Remarks
Resser (Reference Resser1945) assigned a second cranidium (Fig. 74.5, 74.6; USNM 794188) as a paratype of Antagmus arizonaensis; however, the partial specimen has (as discussed above) a wider and more convex interocular portion of the fixigena, longer preglabellar area, and less-pronounced ocular ridges. There are no other ptychoparioid cranidia or sclerites associated with this specimen, thus it is placed in open nomenclature.
Acknowledgments
We wish to thank S. Bastien (DMNS), J. Clevenger and H. Cothren (Utah State U.), L. Crossey (U. of New Mexico), C. Dehler (Utah State U.), T. Farrell (Boise State U.), T. Karim (U. of Colorado), K. Karlstrom (U. of New Mexico), K. Kirschner, M. Mohr (Boise State U.), E. Olson, M. Nebel (GRCA), J. Pollock (ARR), M. Pulsipher (GRCA), M. Schmitz (Boise State U.), E. Rose, S. Rowland (U. of Nevada Las Vegas) and M. Timmons (NMBGMR) for their assistance in the field. The detailed stratigraphic section for river mile 275 section was generously provided by C. Dehler. Extensive collection management was provided by K. Besom (GRCA), N. Drew (NMNH), M. Florence (NMNH), C. Labandeira (NMNH), H. Little (NMNM), J. Nakano (NMNH), and N. Neu-Yagle (DMNS); their efforts are greatly appreciated. This study was partially supported from NSF grant EAR 1955078 and 1954634 from the Sedimentary Geology and Paleobiology Program, by patrons of the Denver Museum of Nature & Science, and by the National Park Service (Paleontology Uprun River Trips 22-SRM014 and 23-SRM014). Fossils were collected under NPS permit GRCA-2022-SCI-0055. Fossils were collected under NPS permit GRCA-2022-SCI-0055 and BLM permit AZ-739. This manuscript benefited from the reviews by N. Handkamer, B. Pratt, and S. Westrop.
Competing interests
The authors declare none.
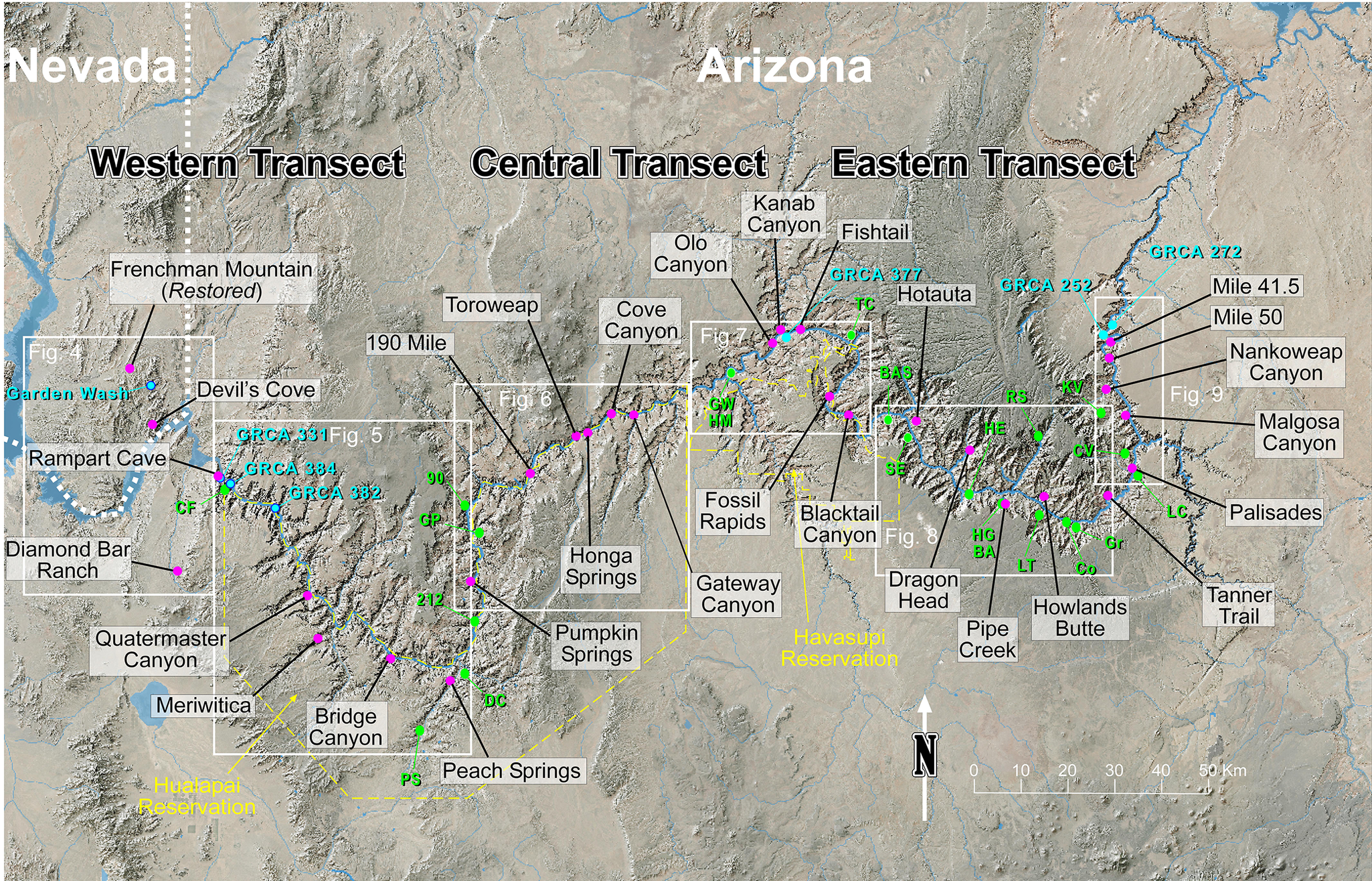
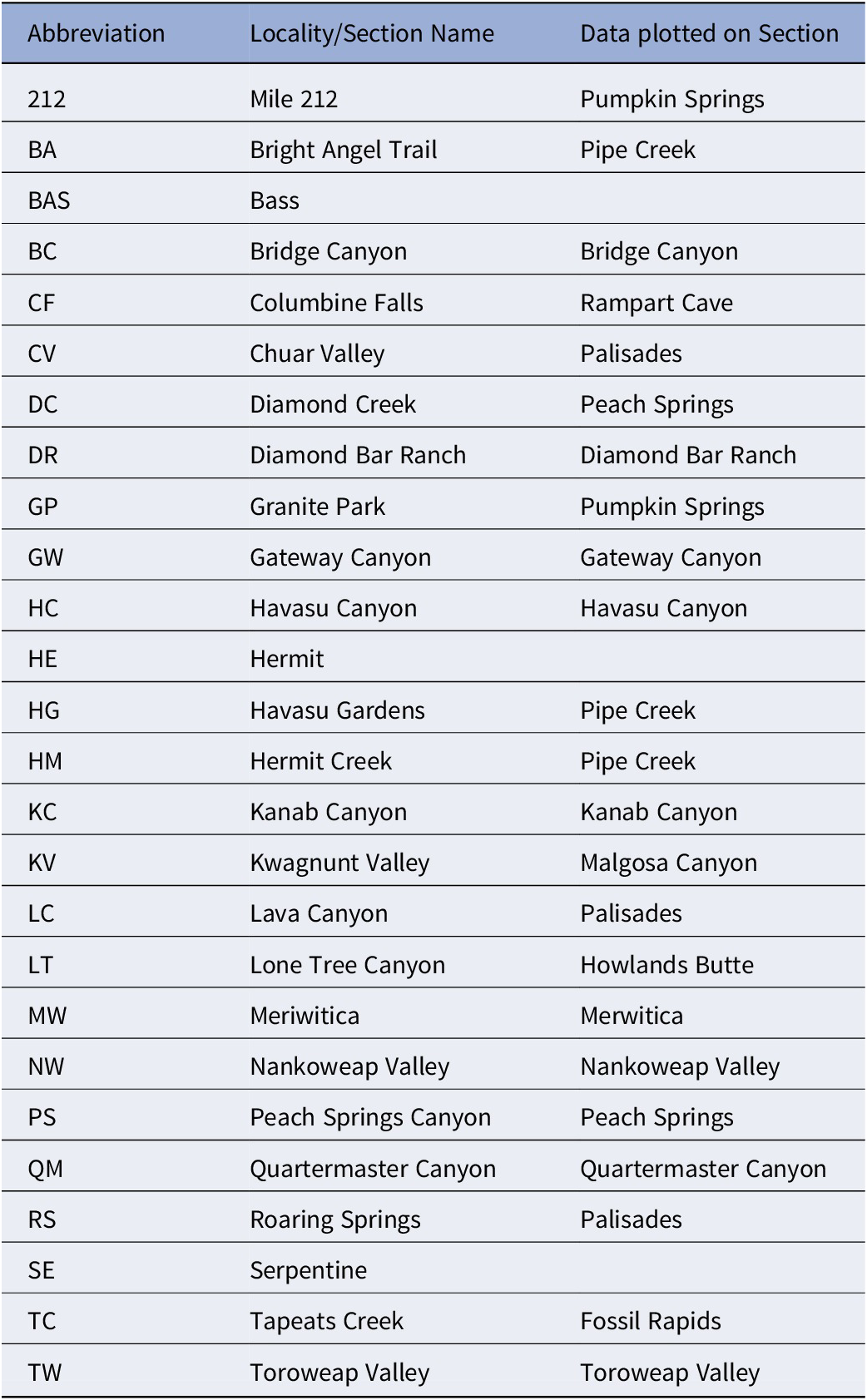
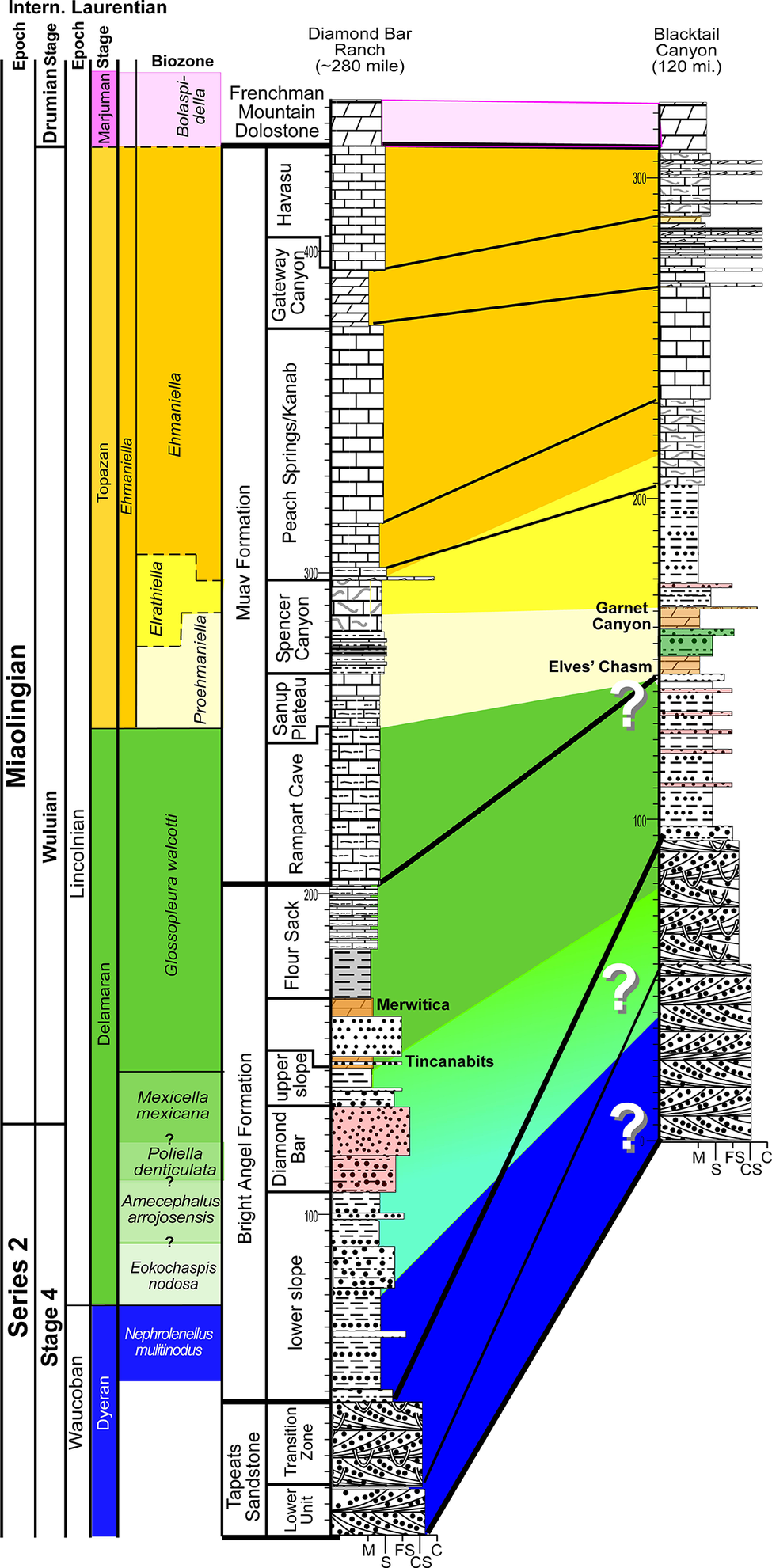
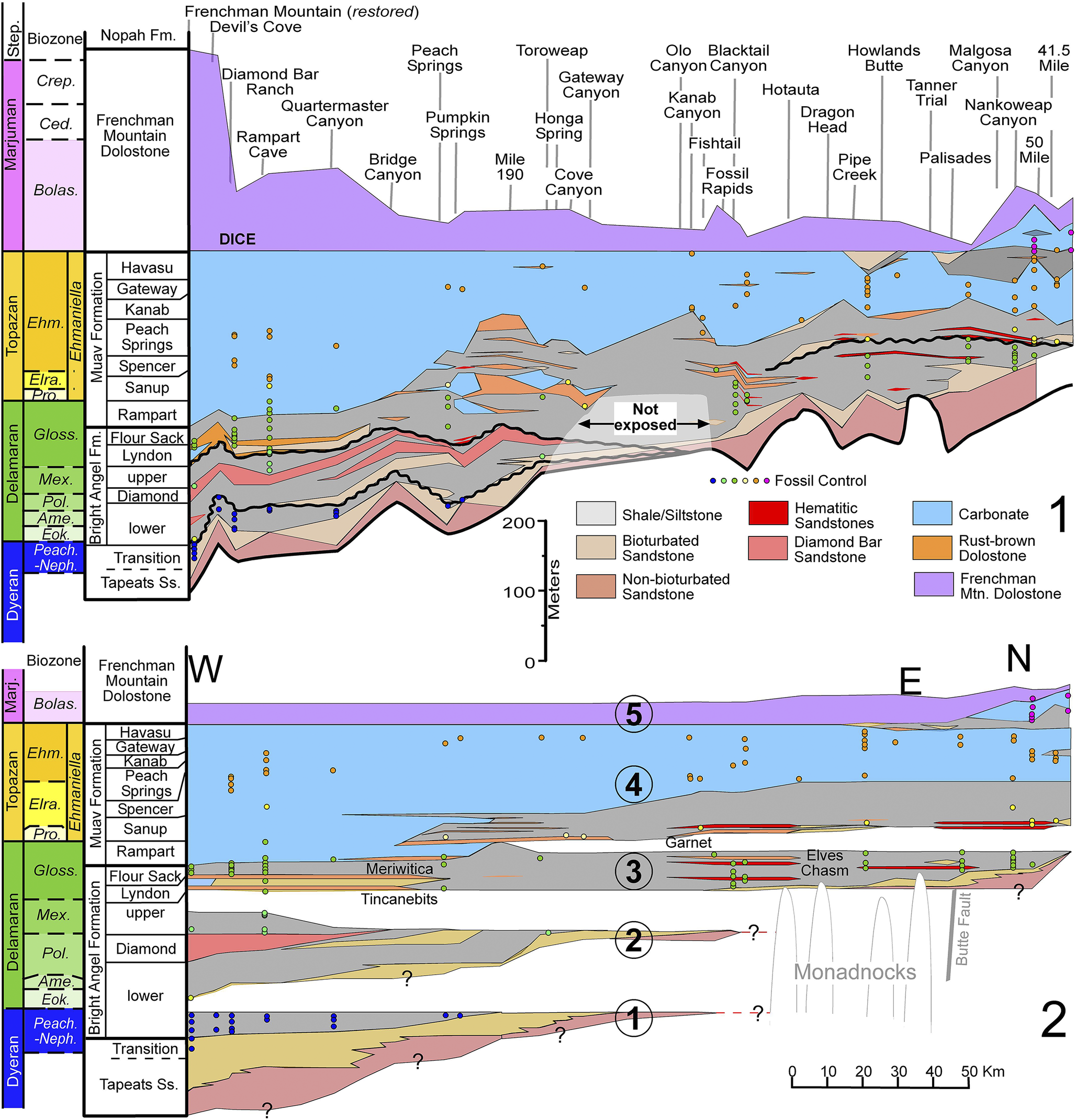
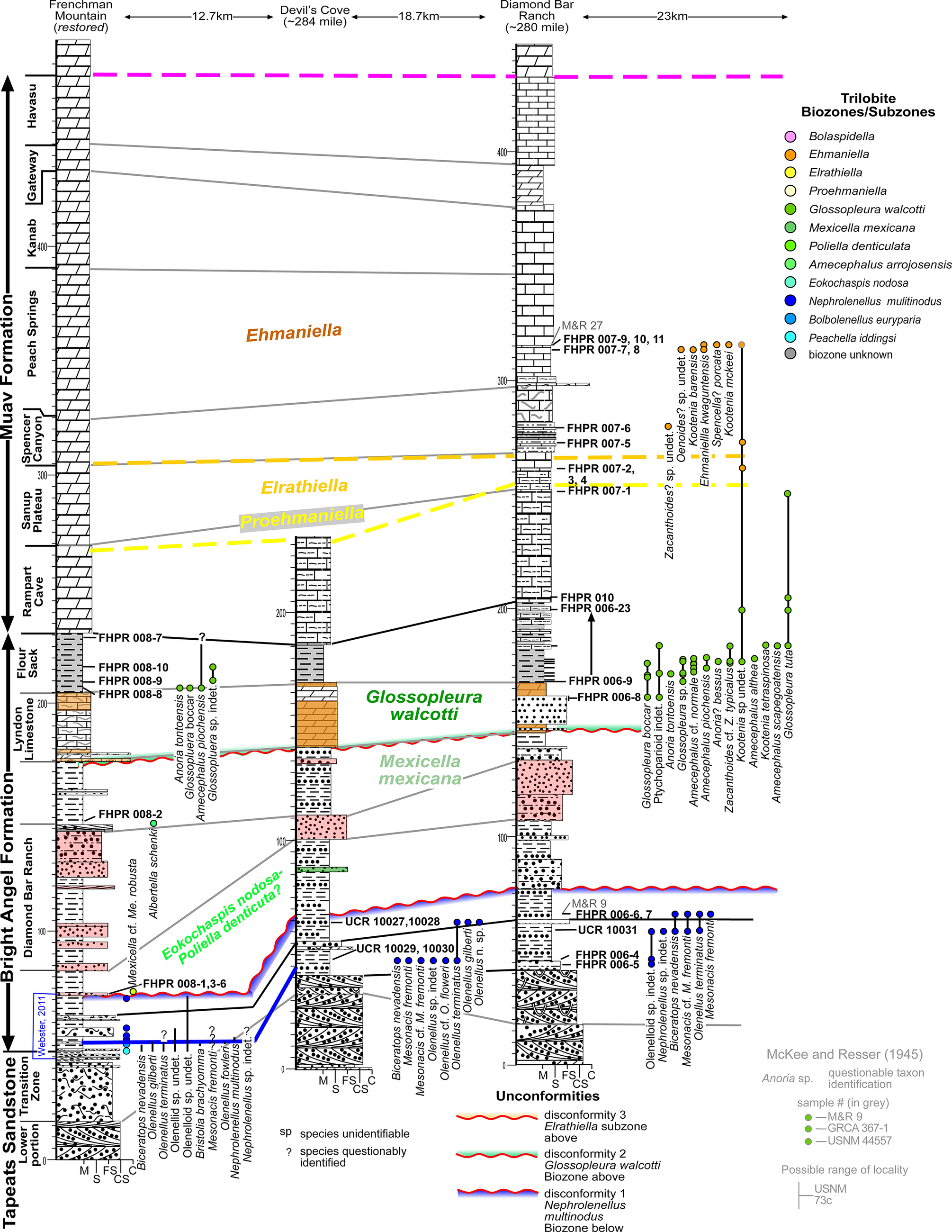
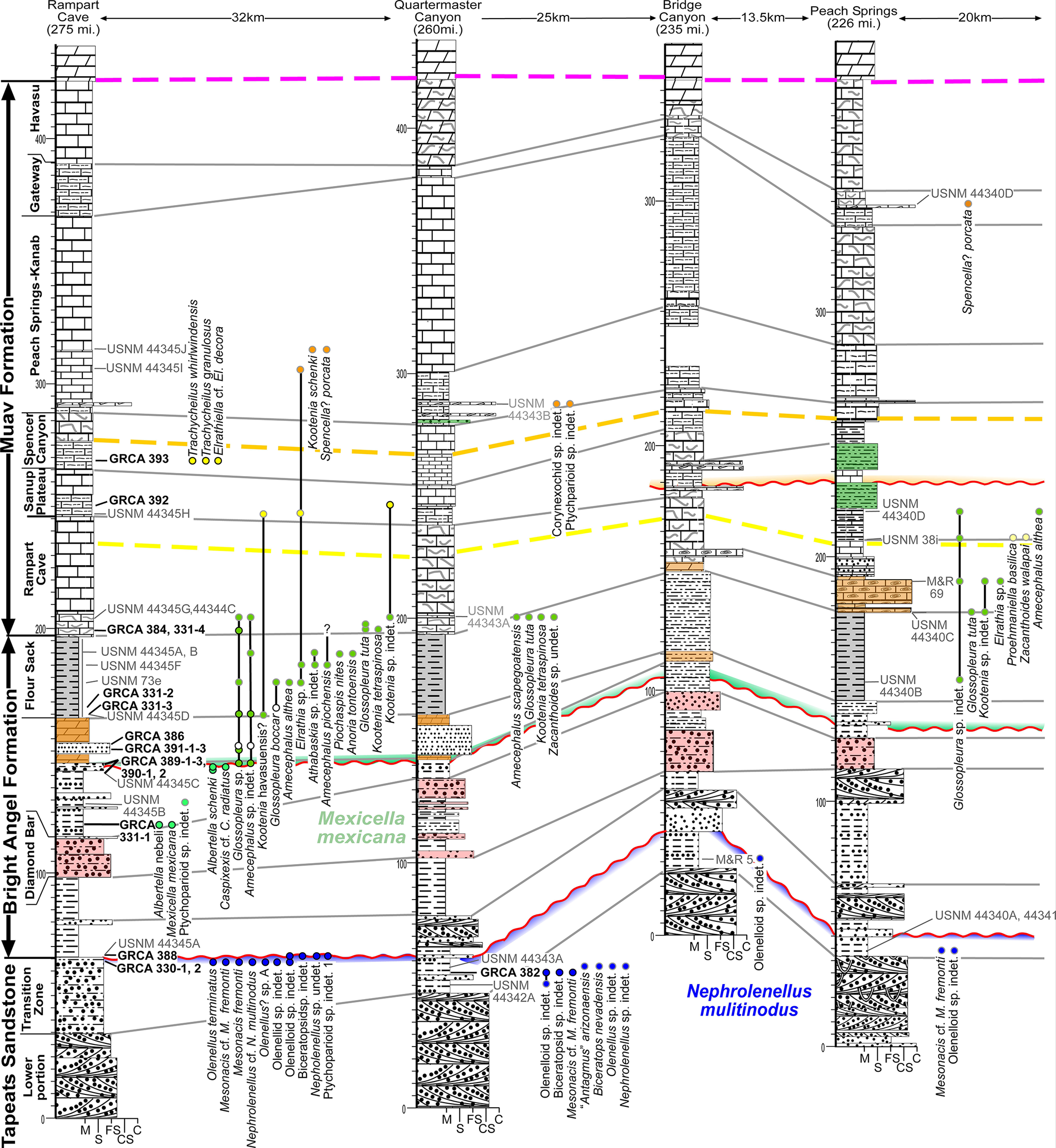
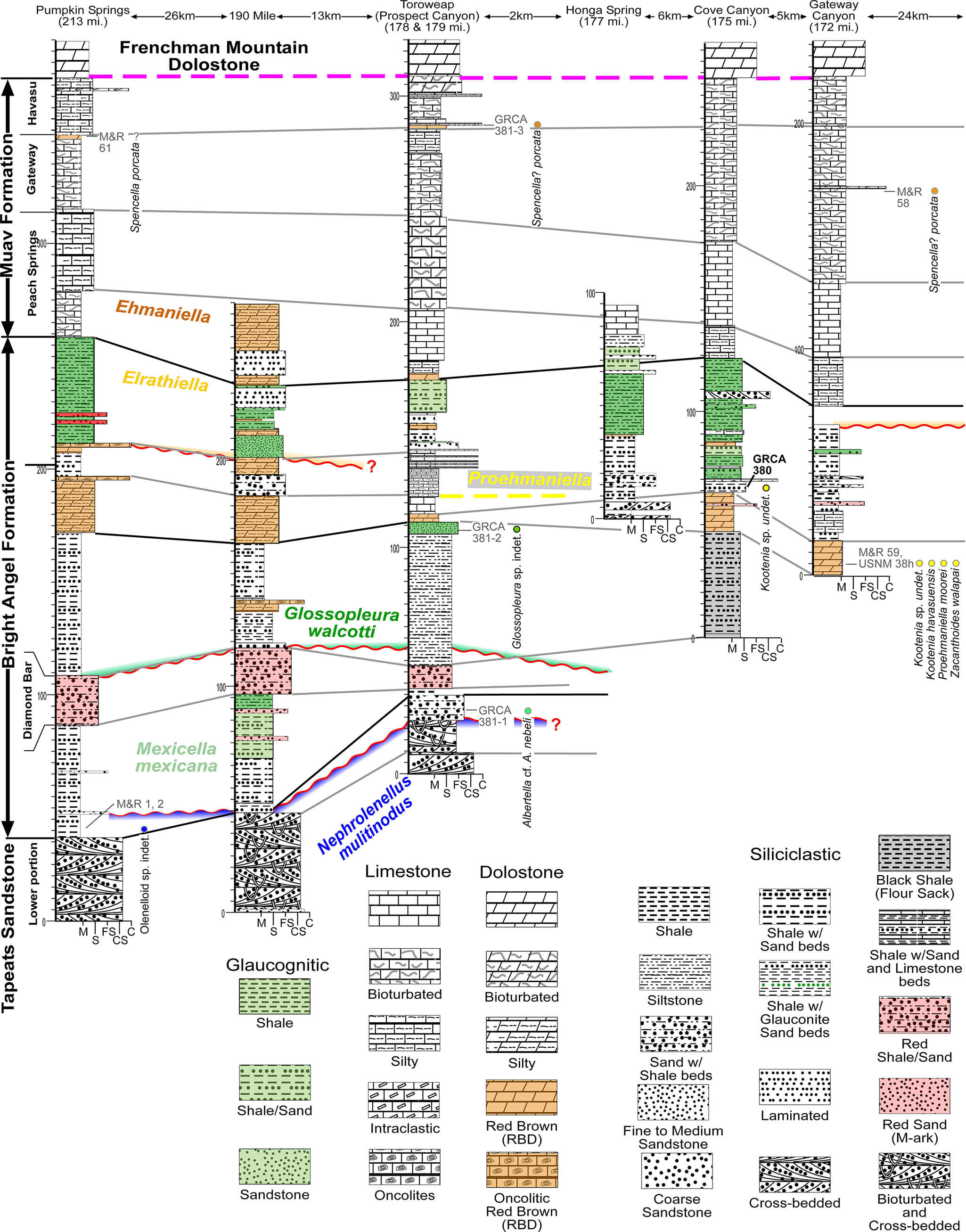
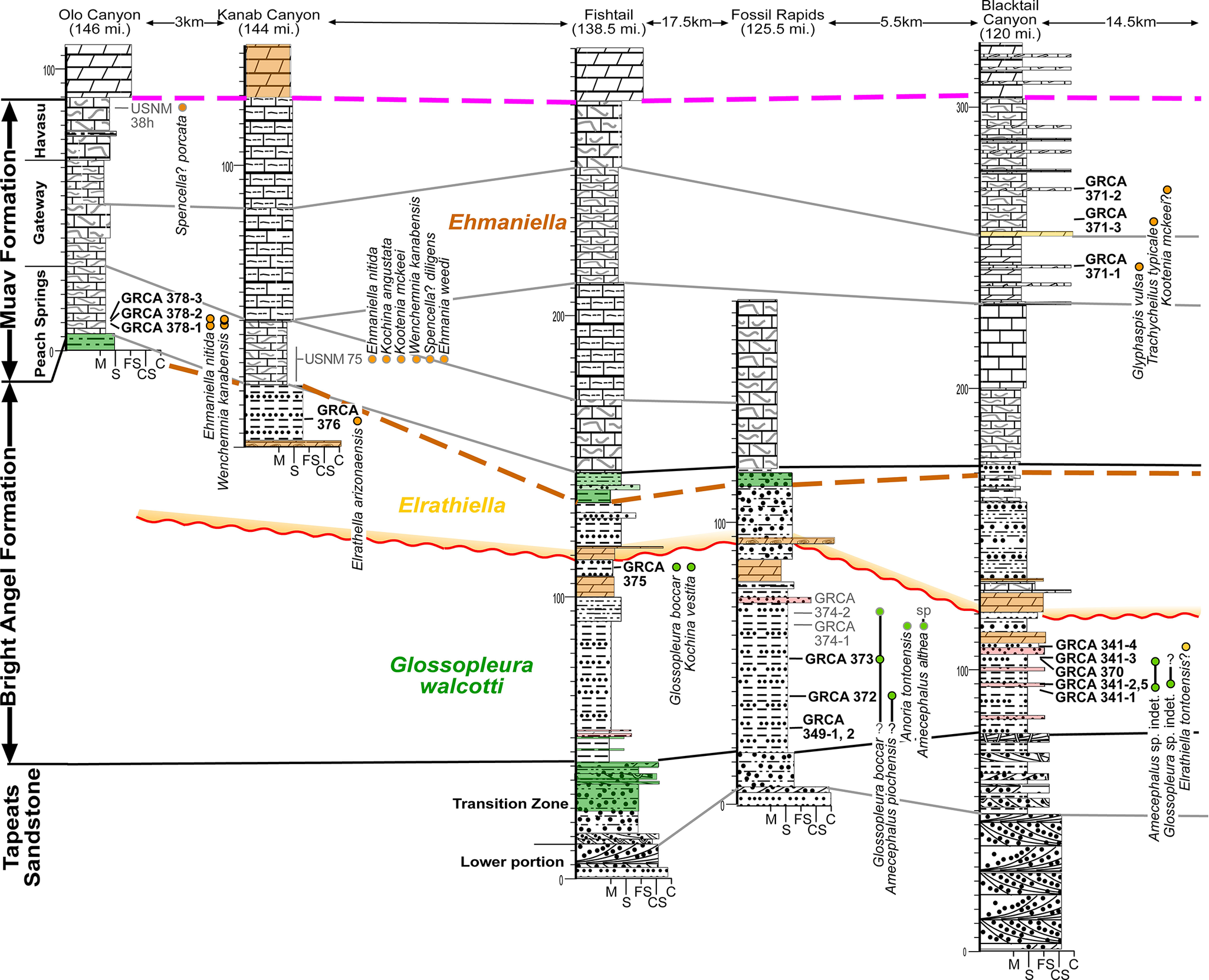
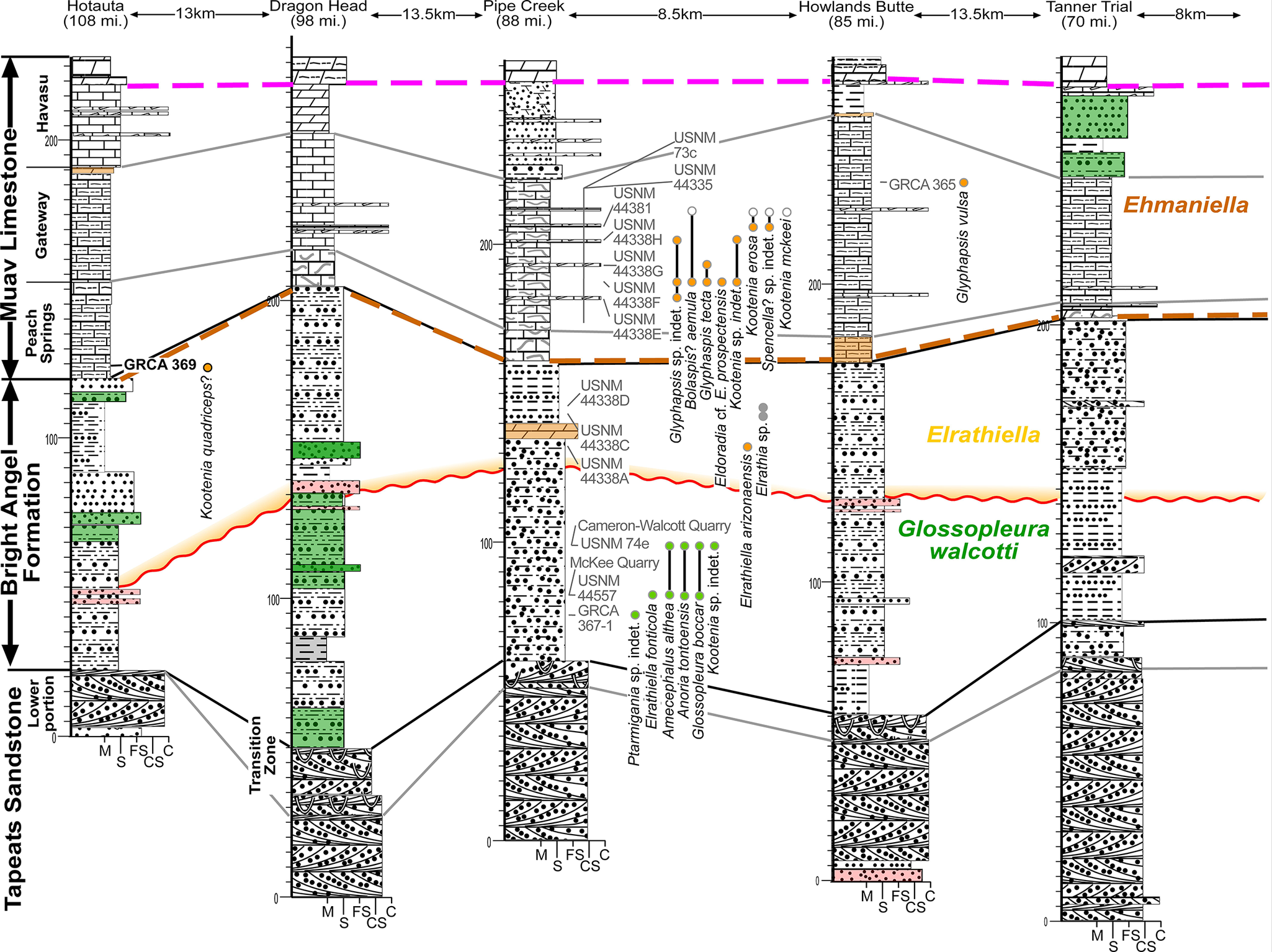
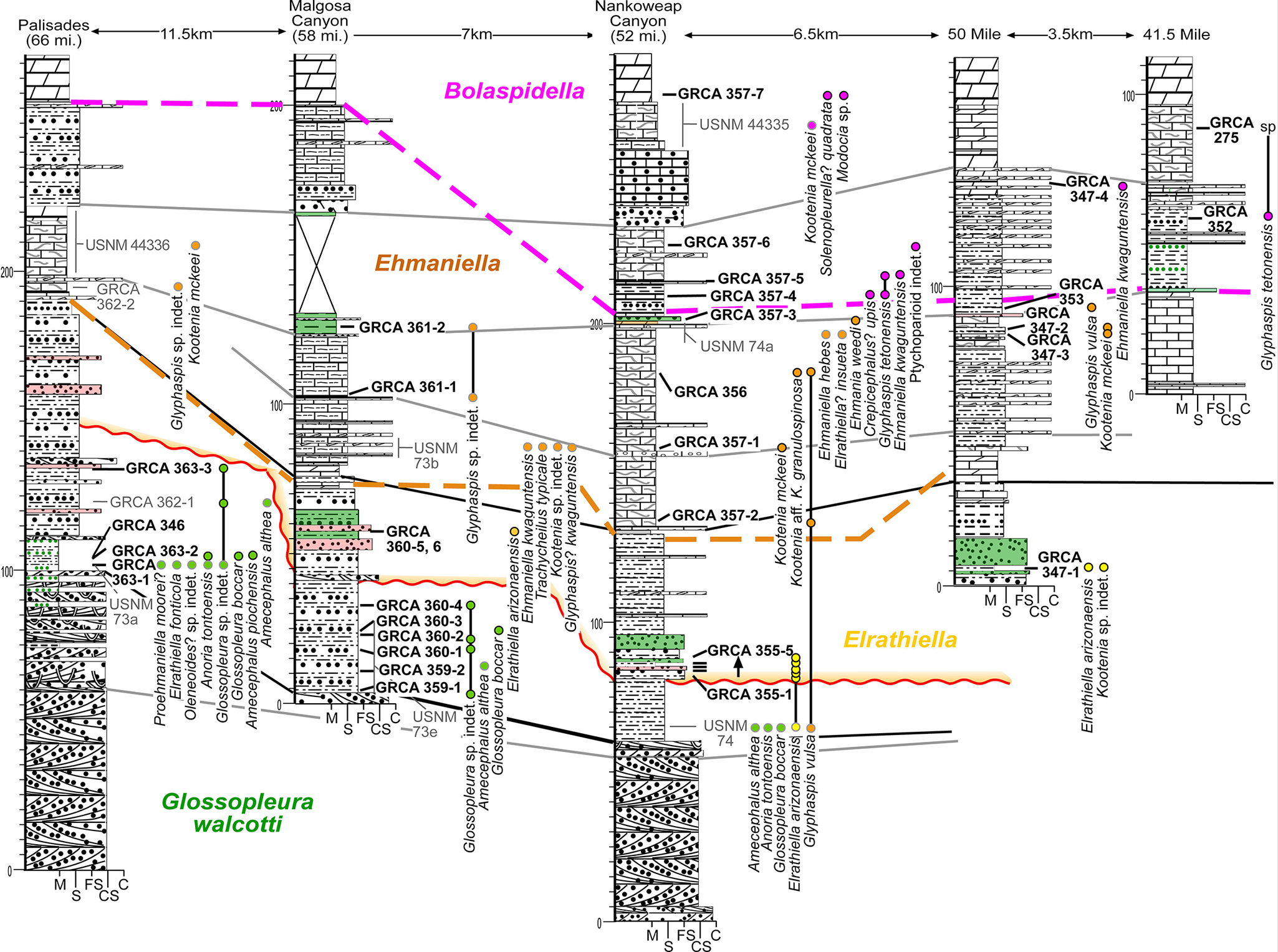
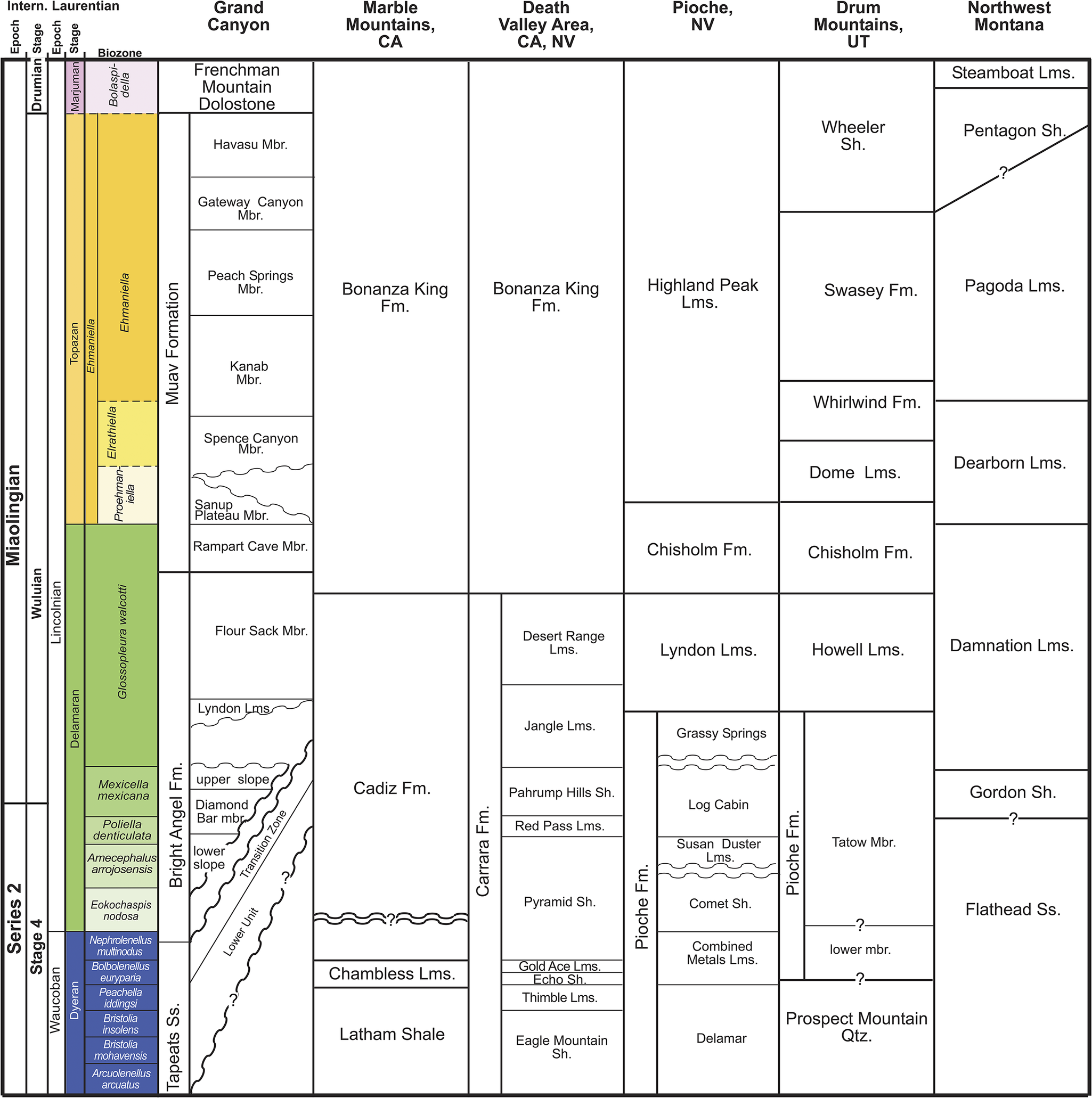
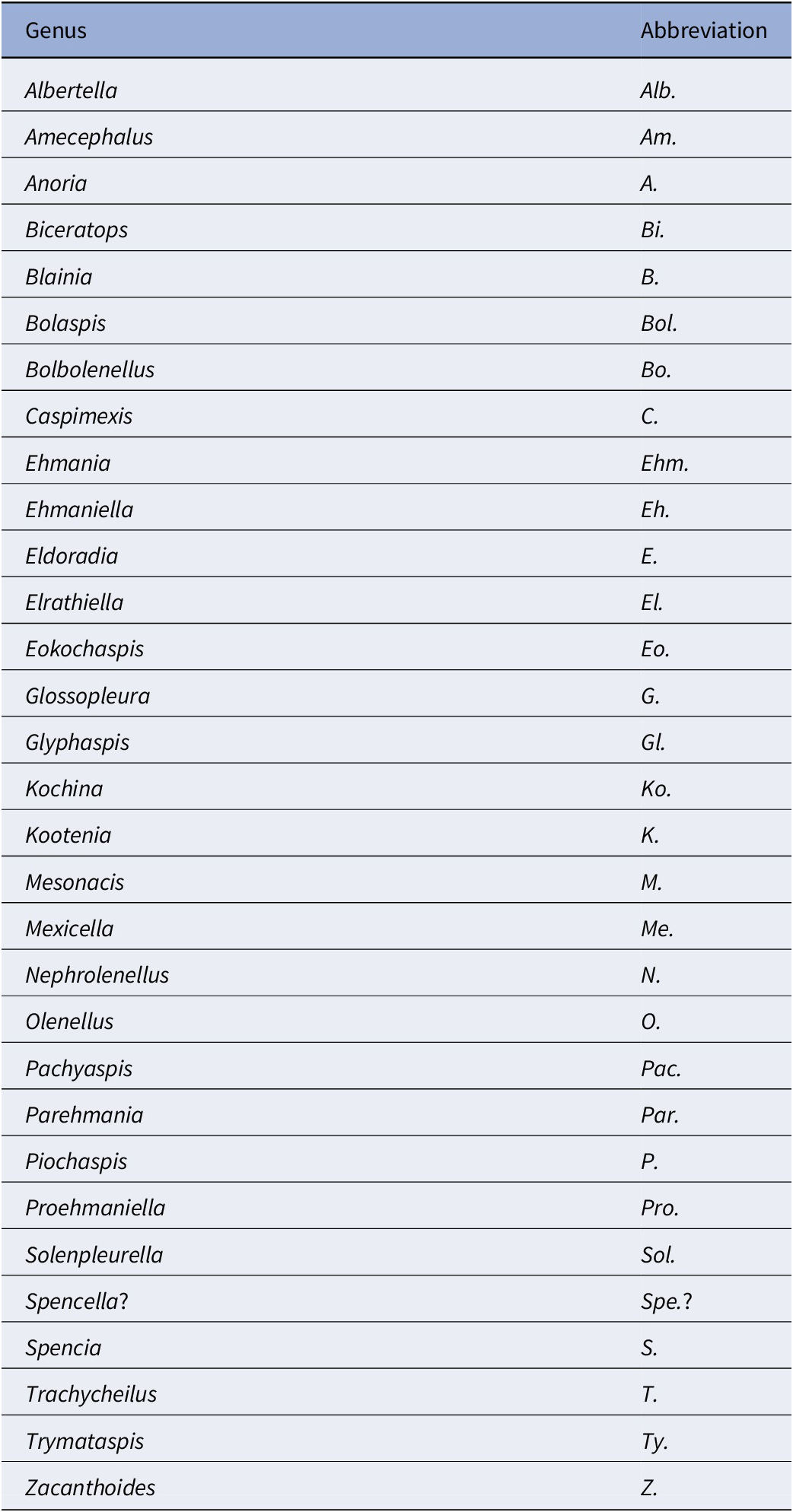

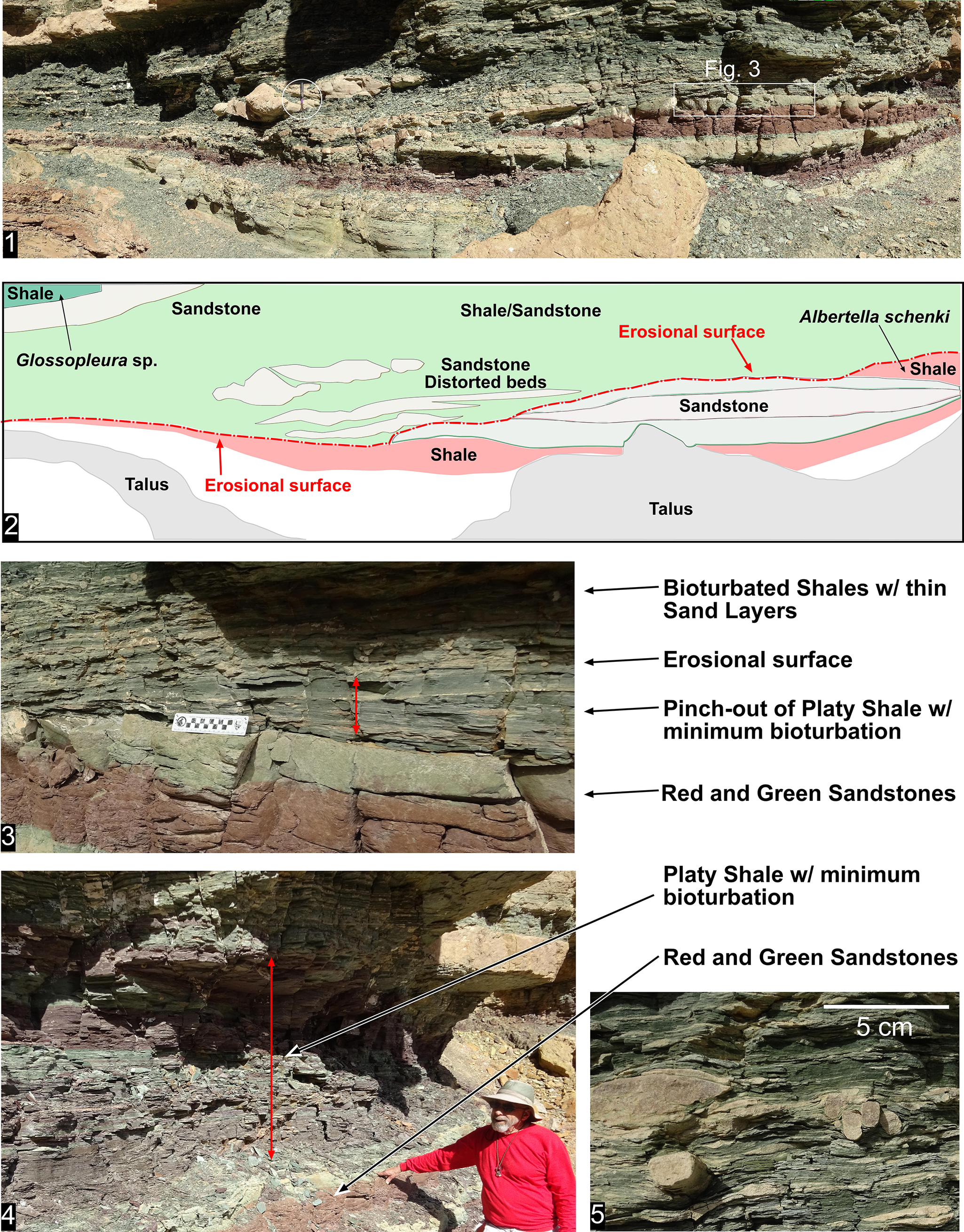


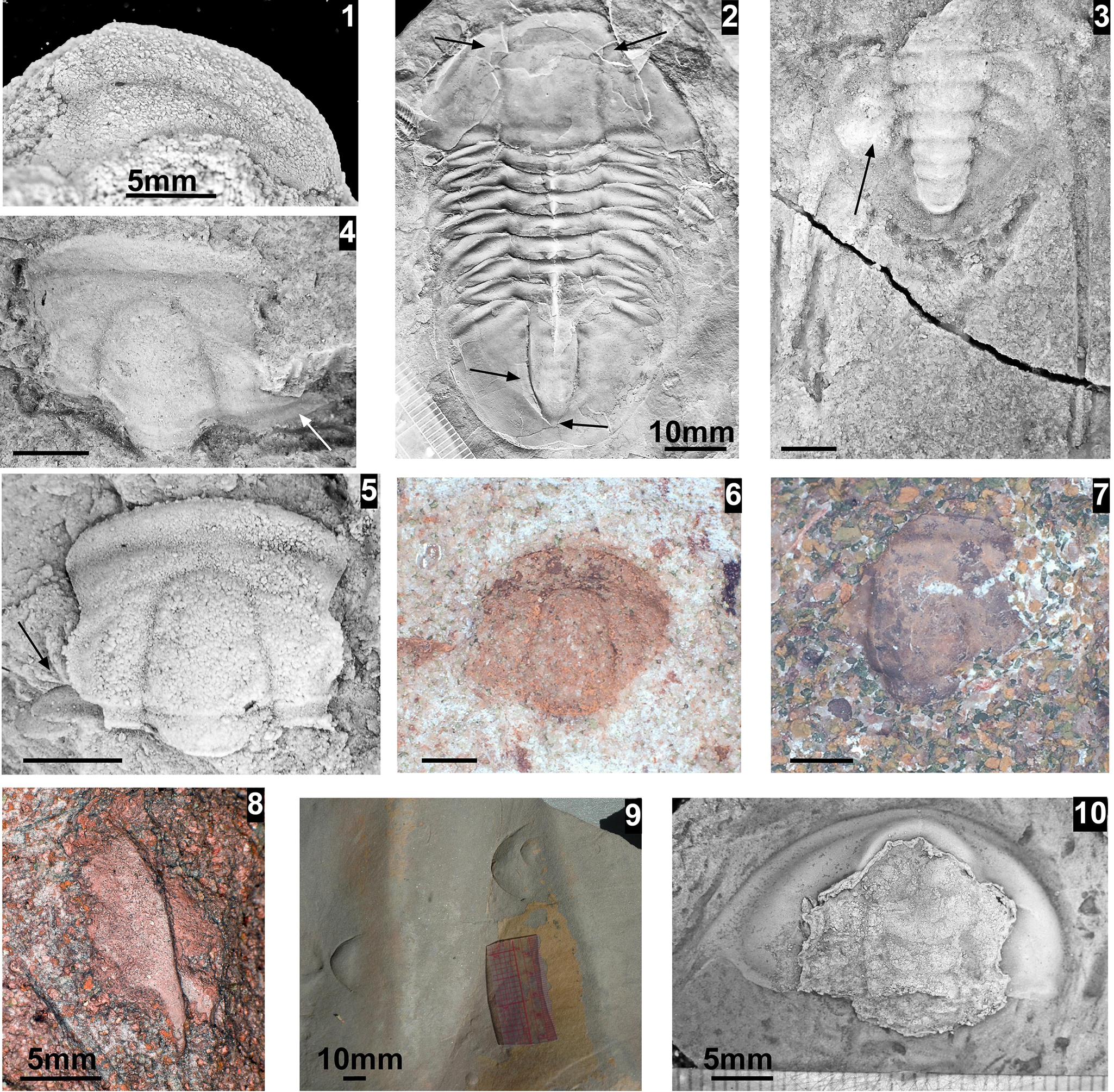
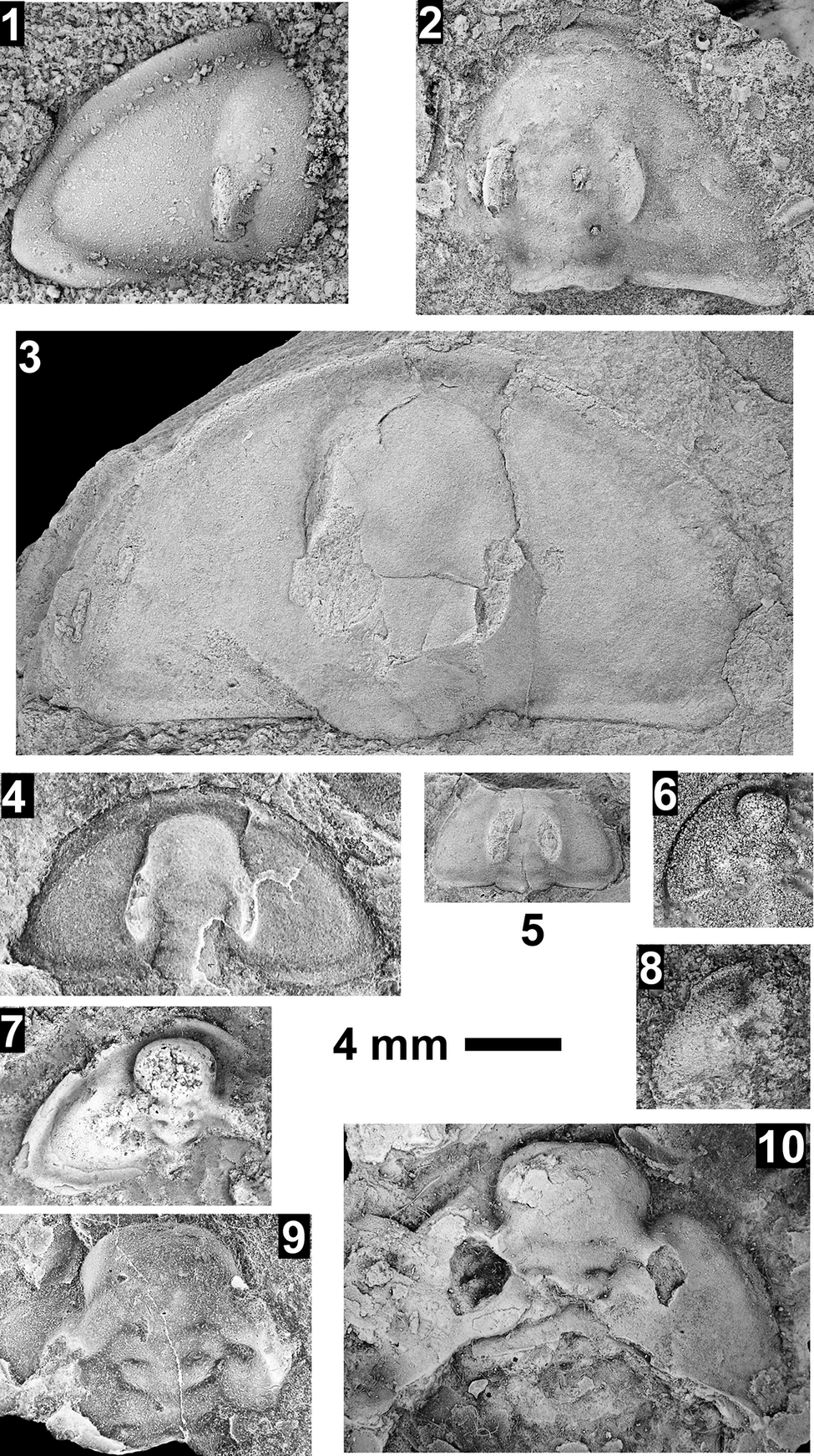
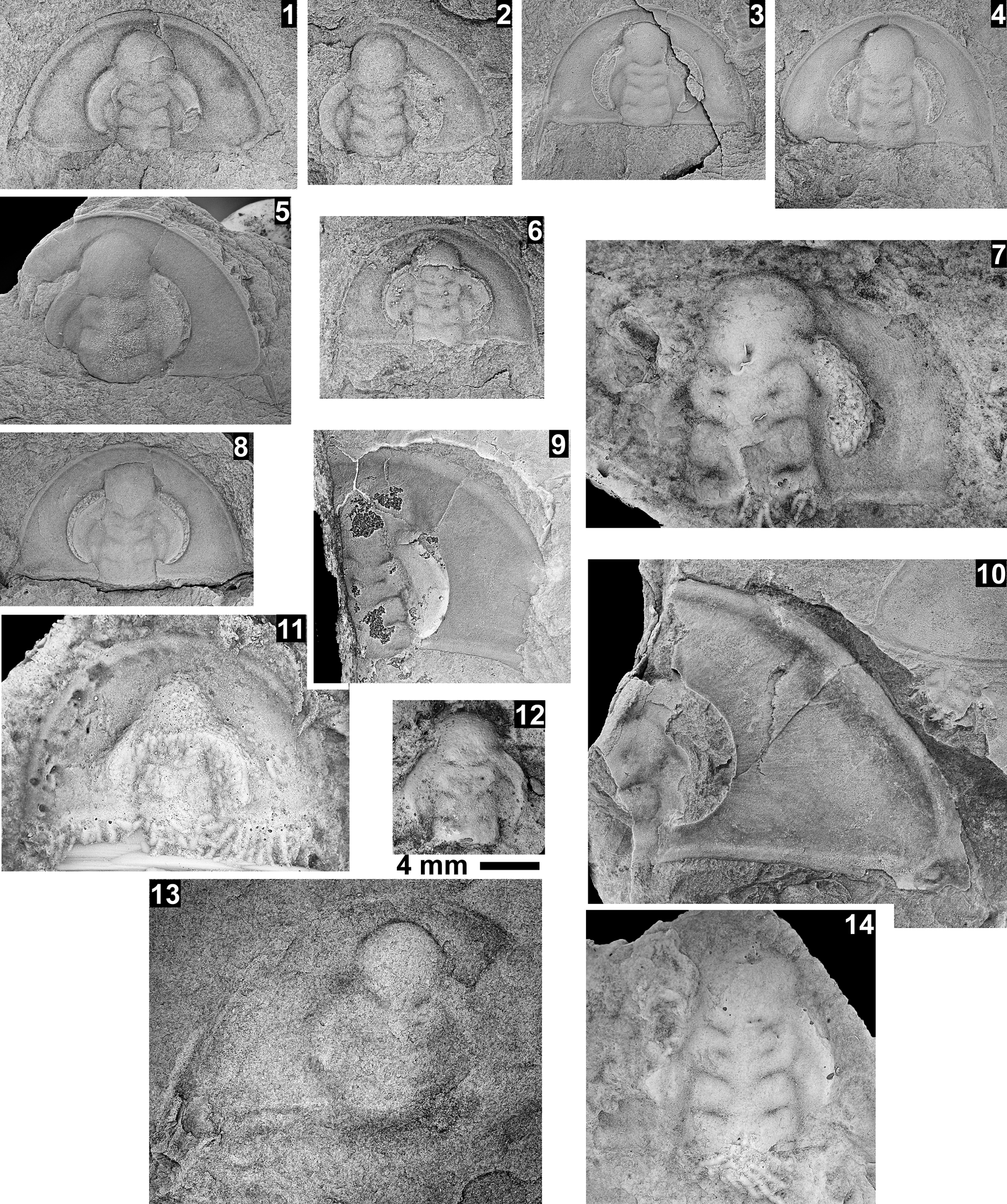
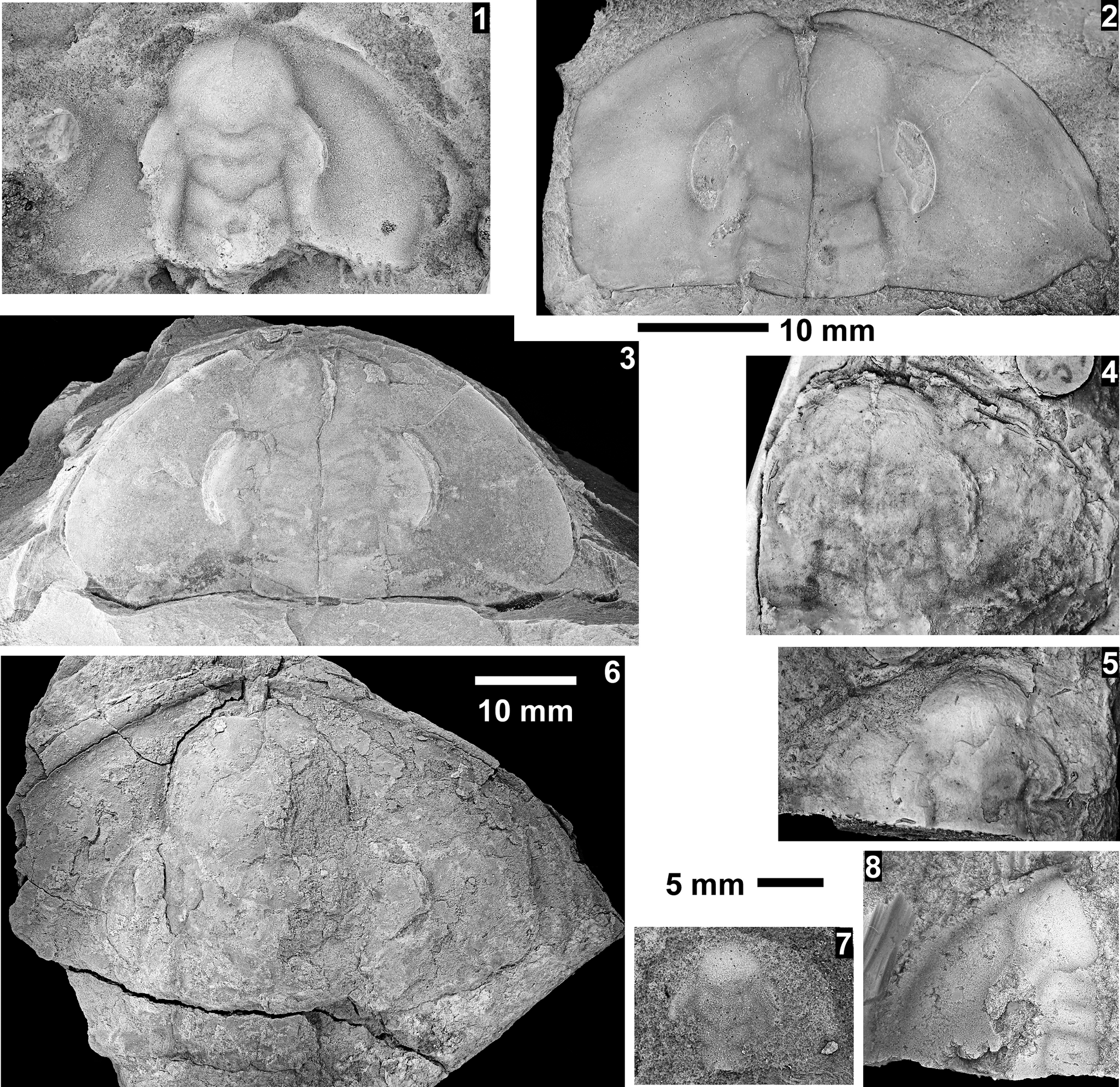

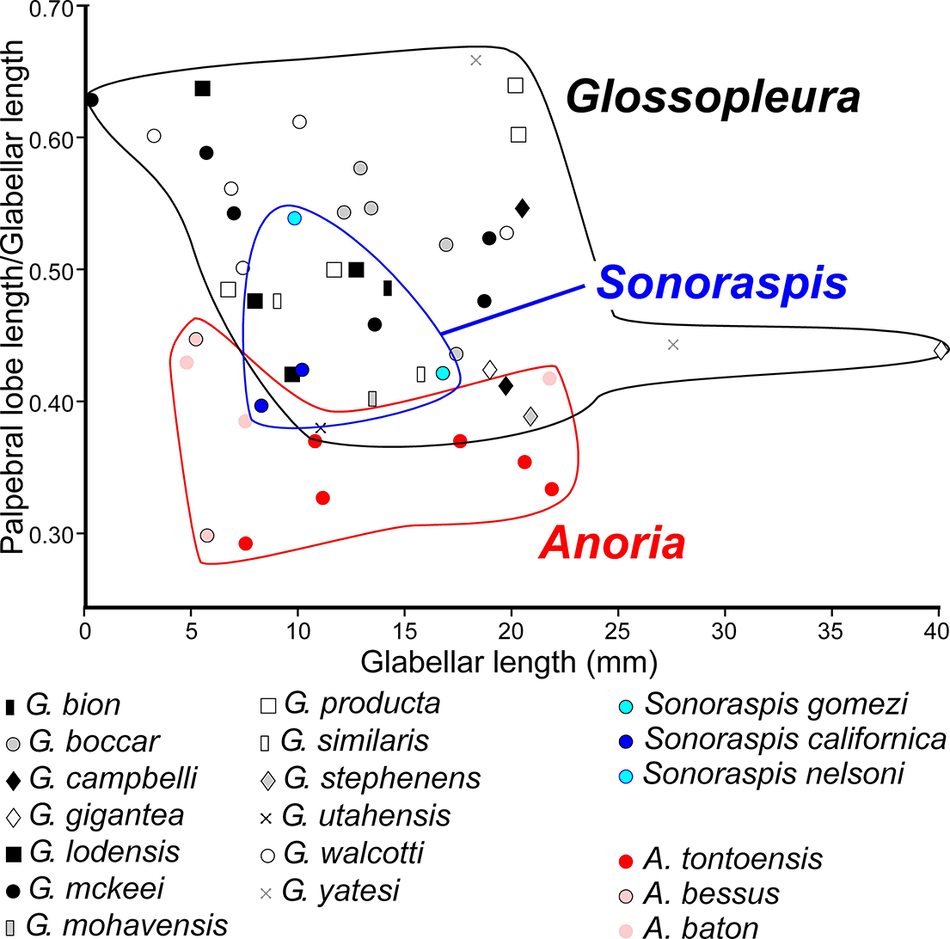
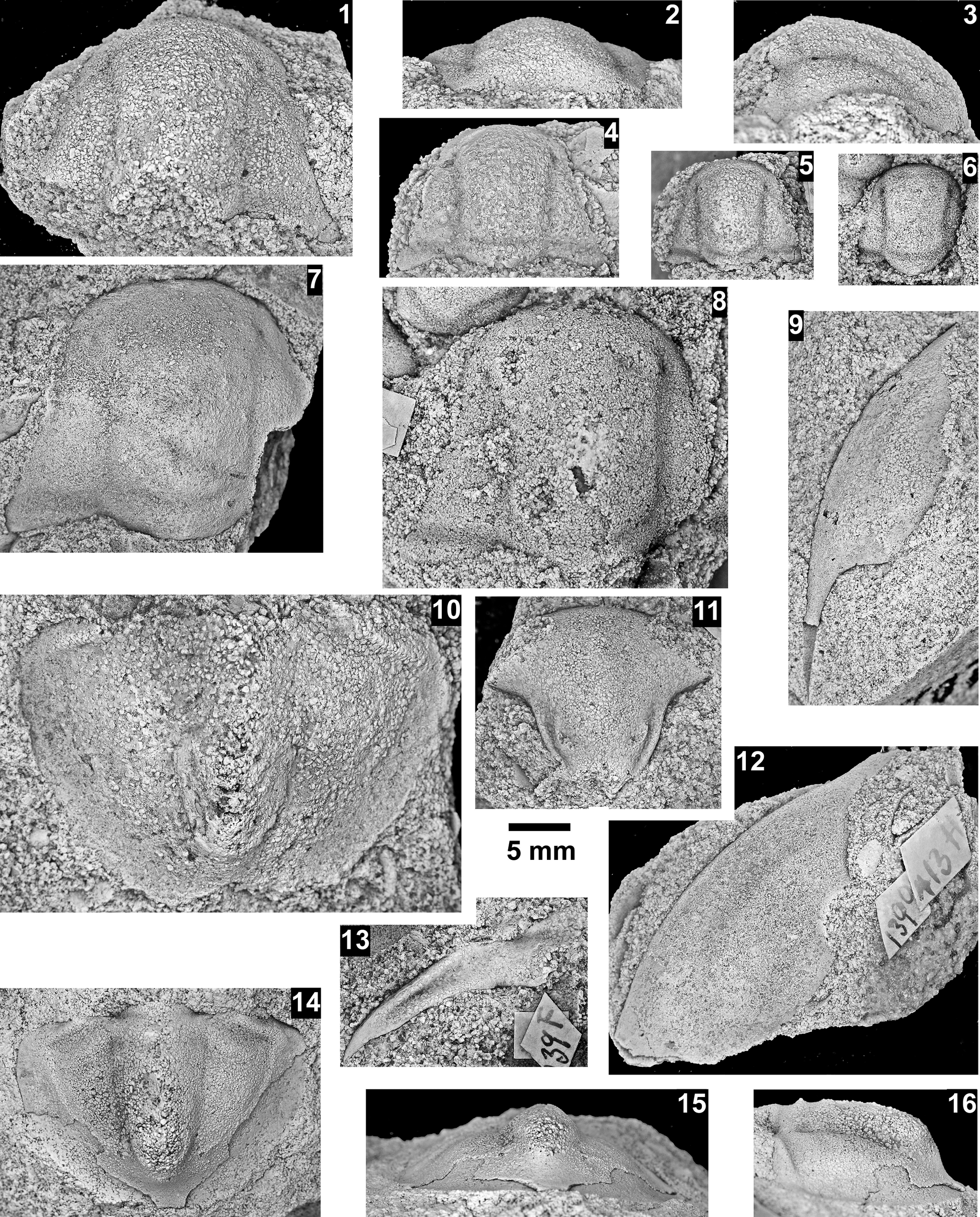
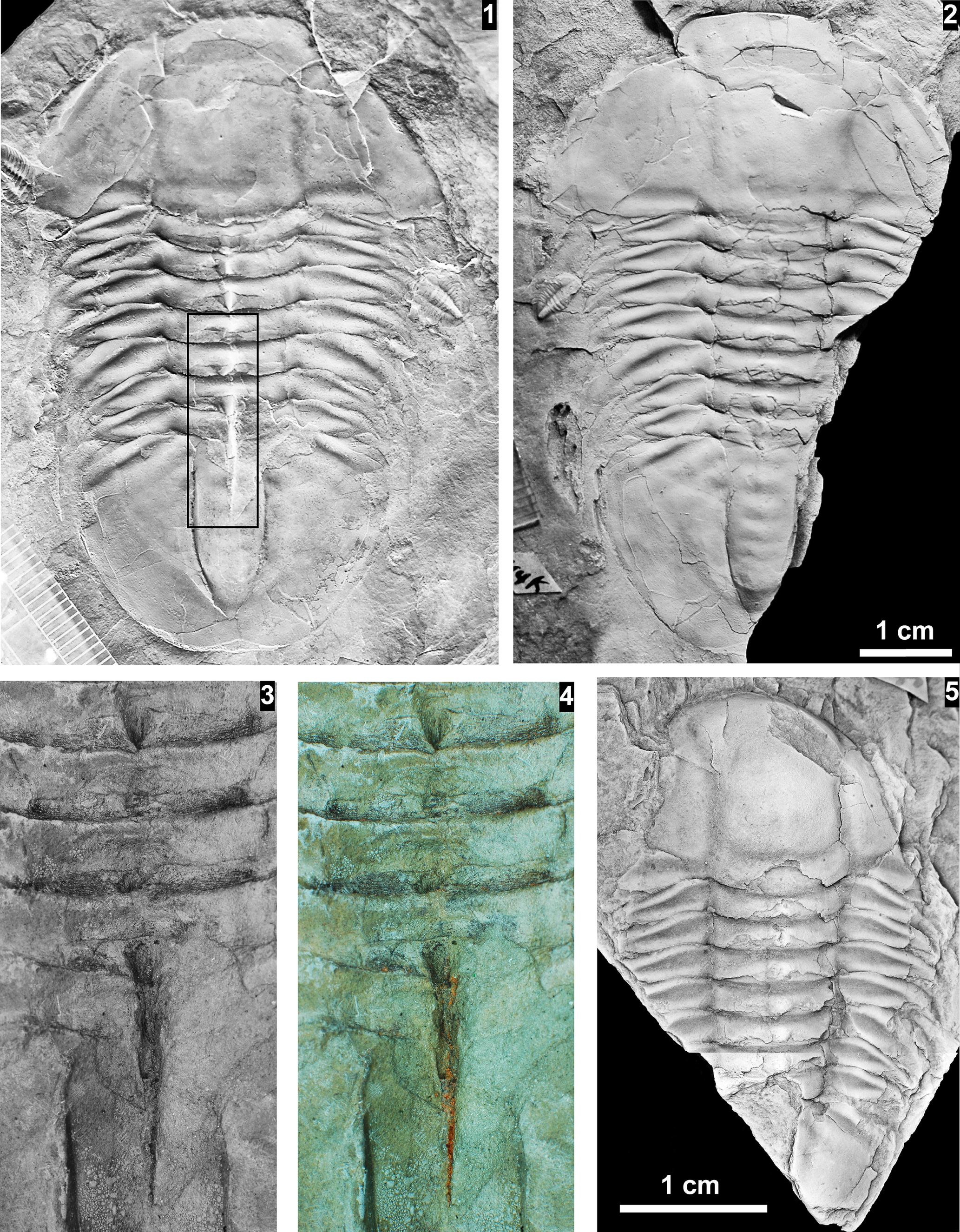
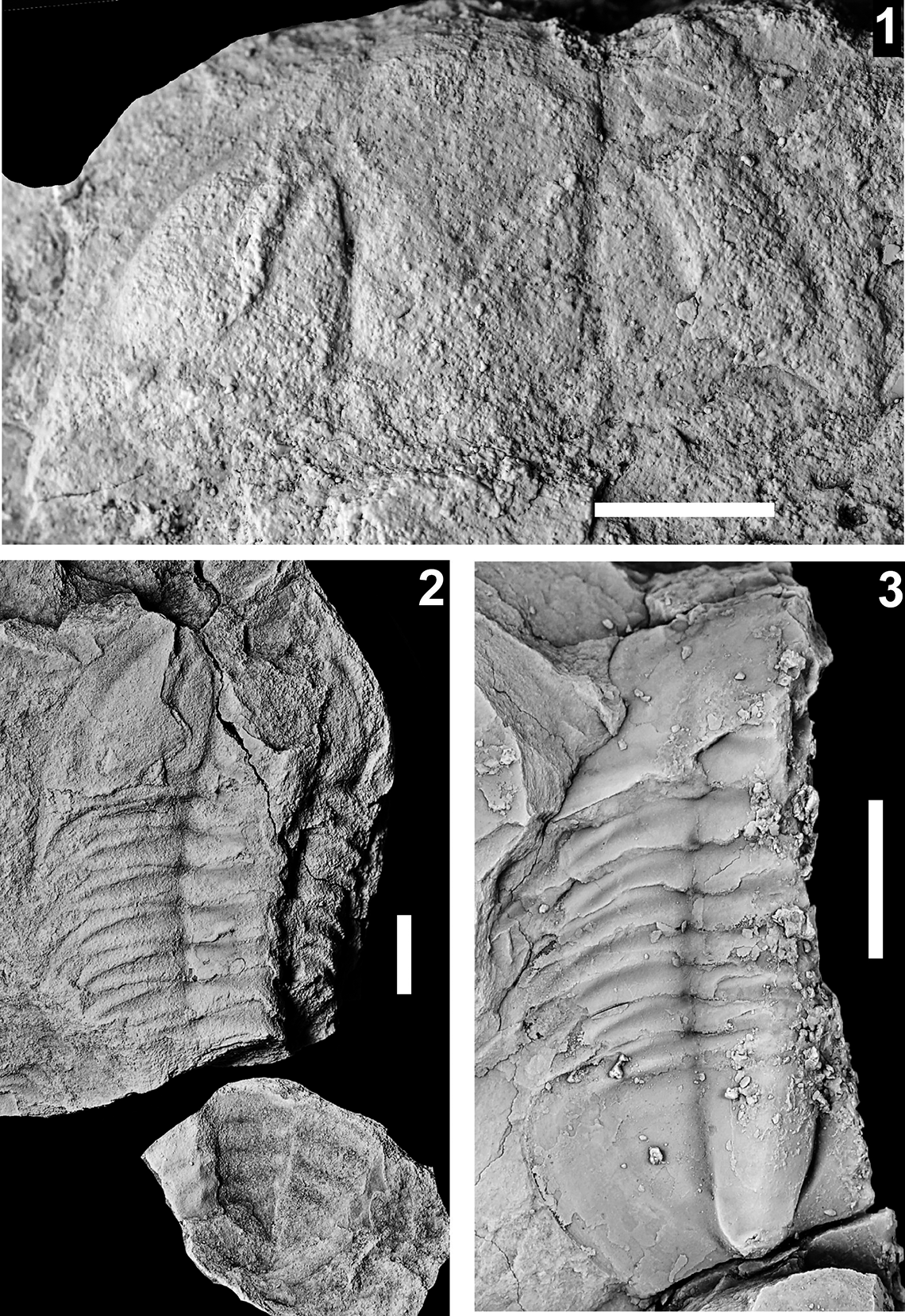
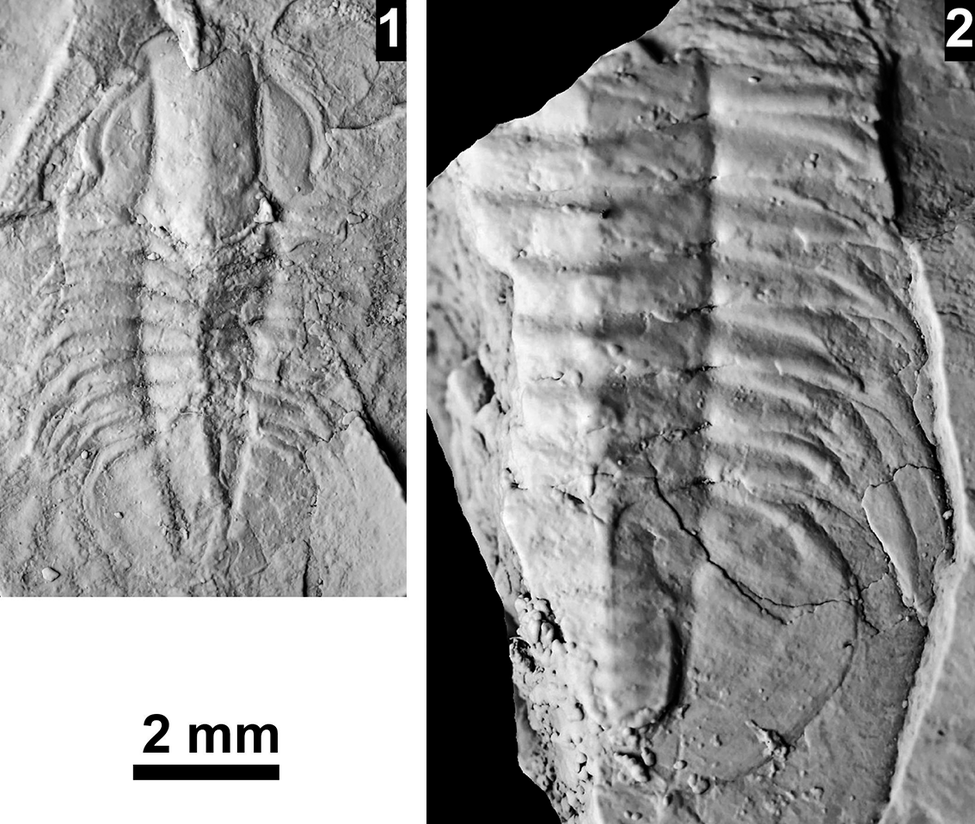
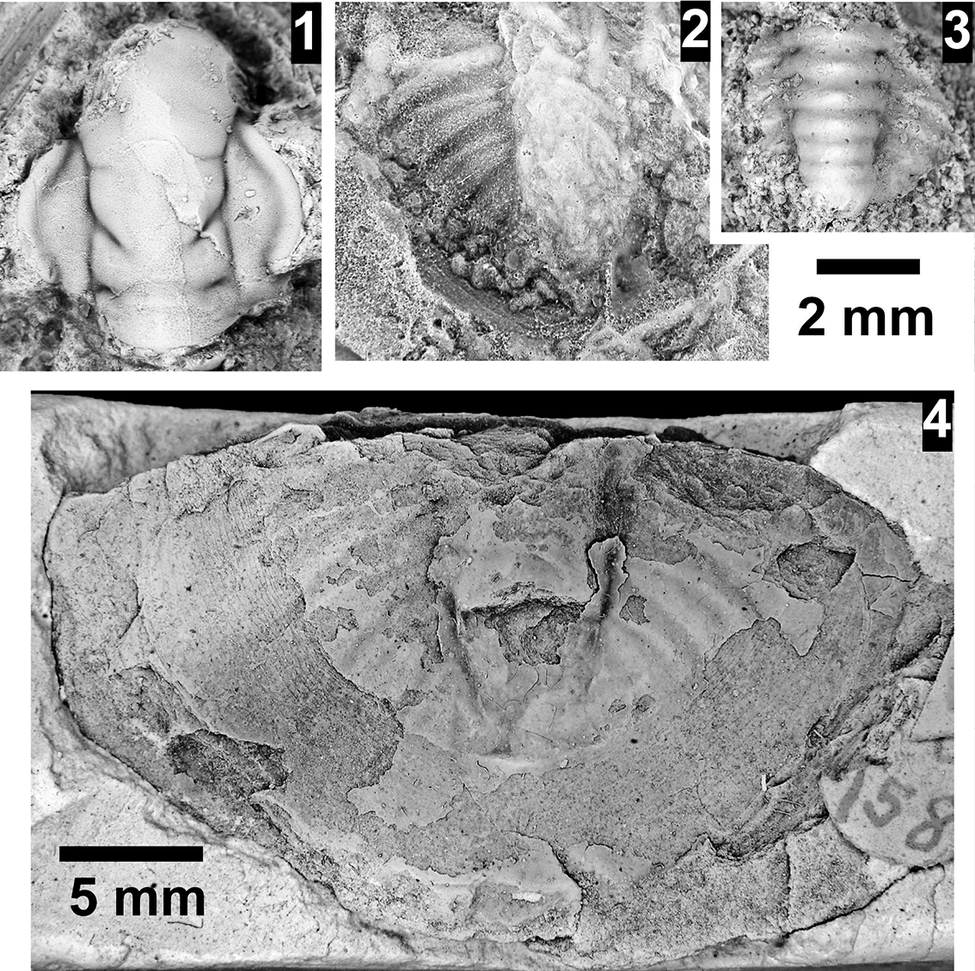
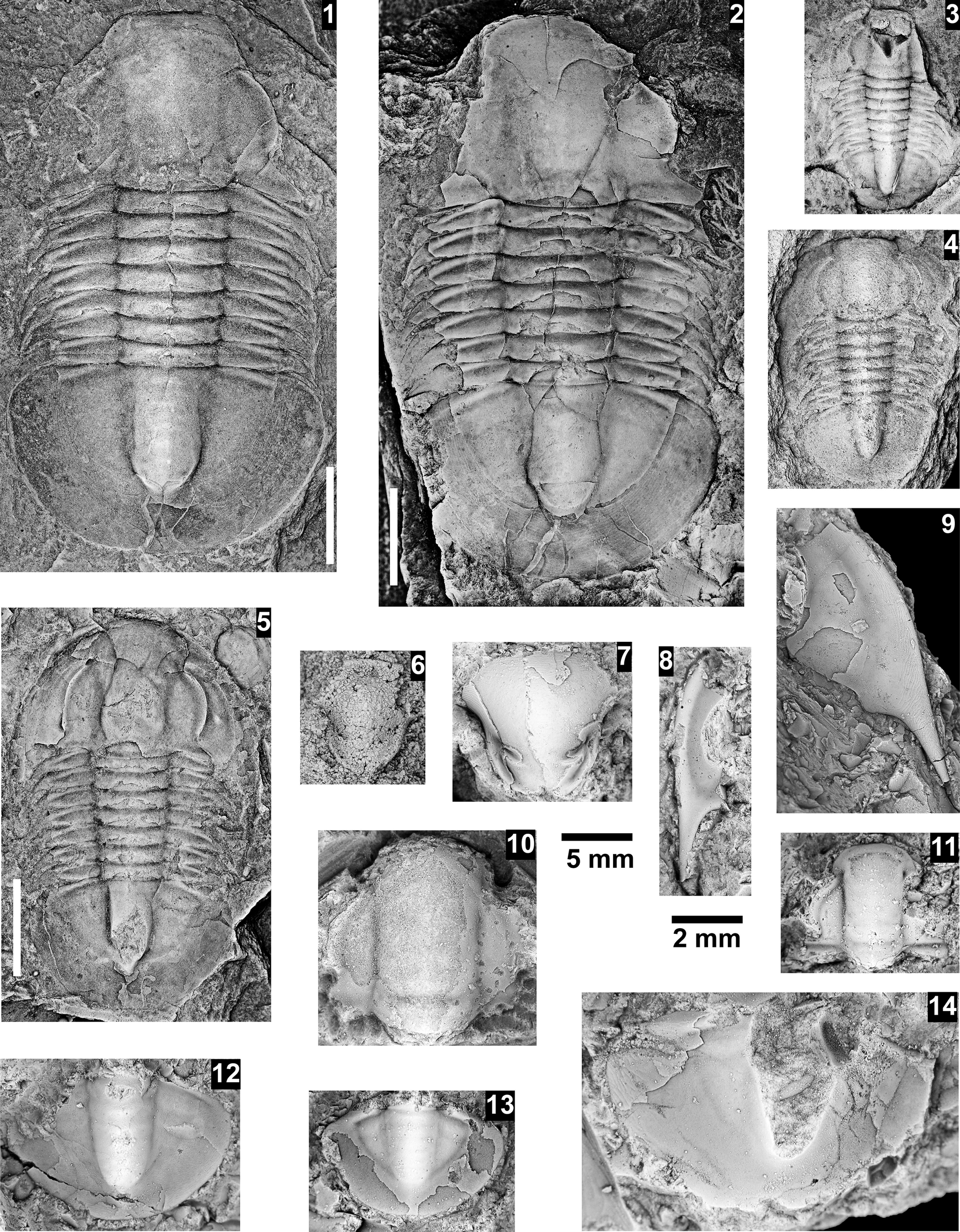
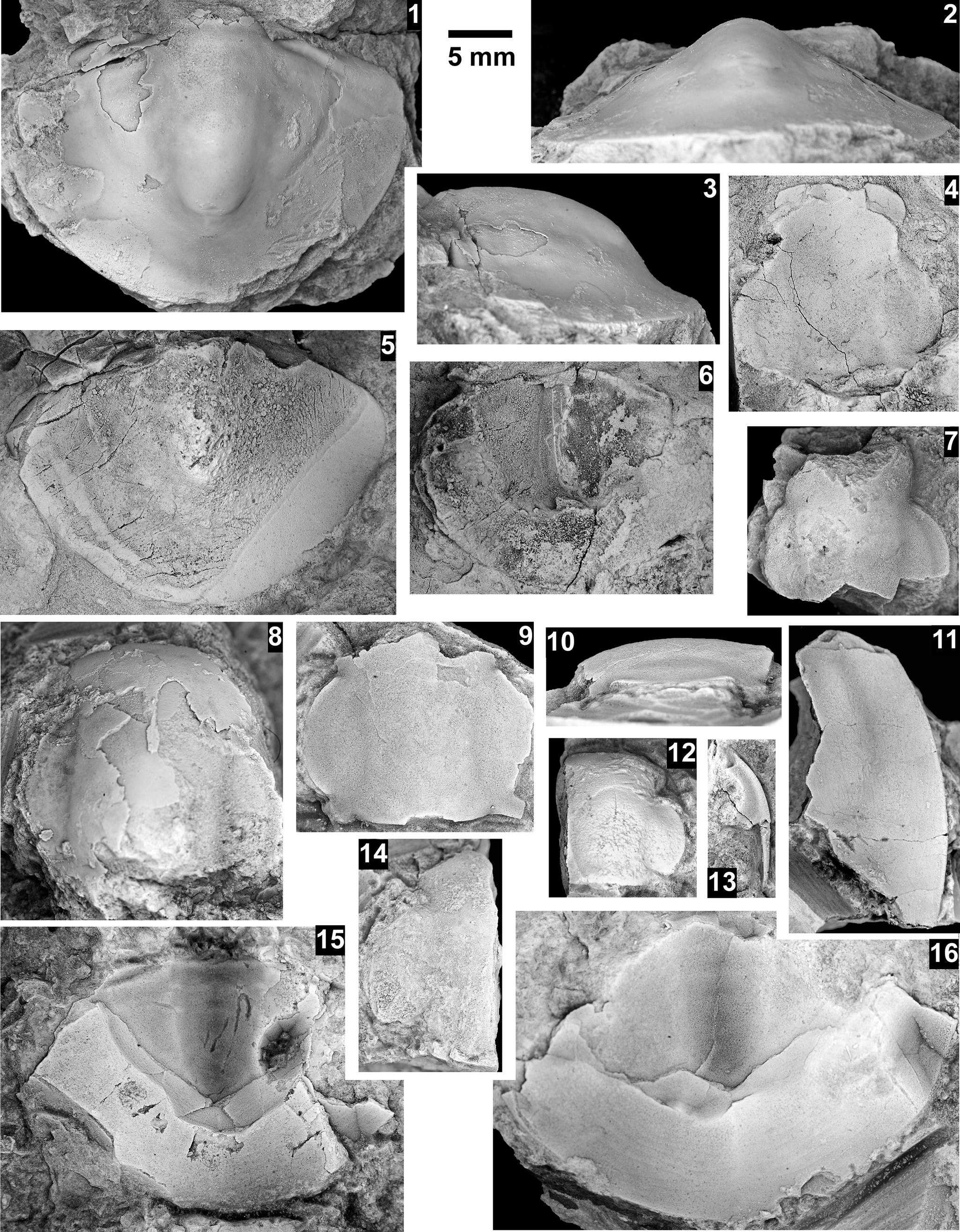
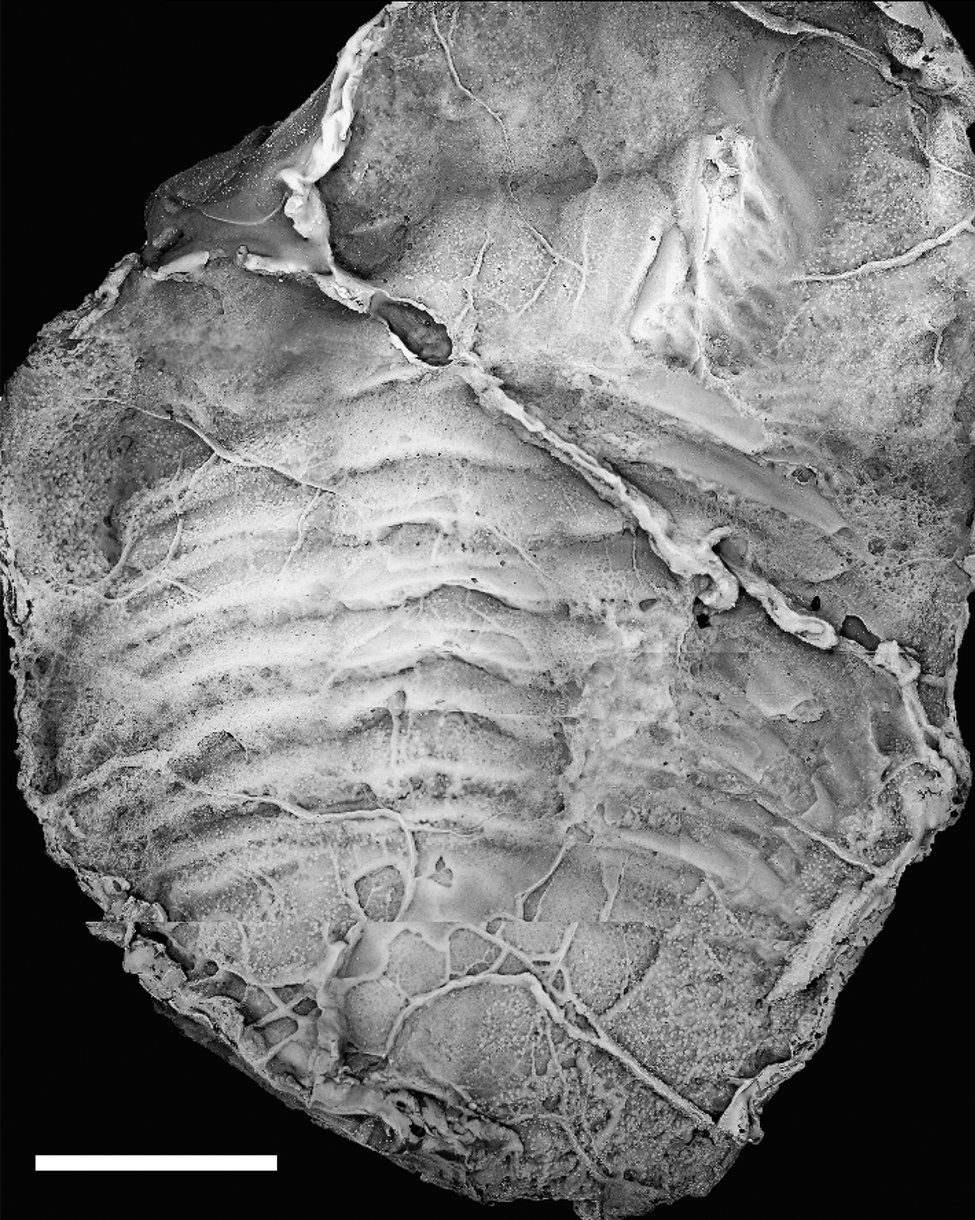

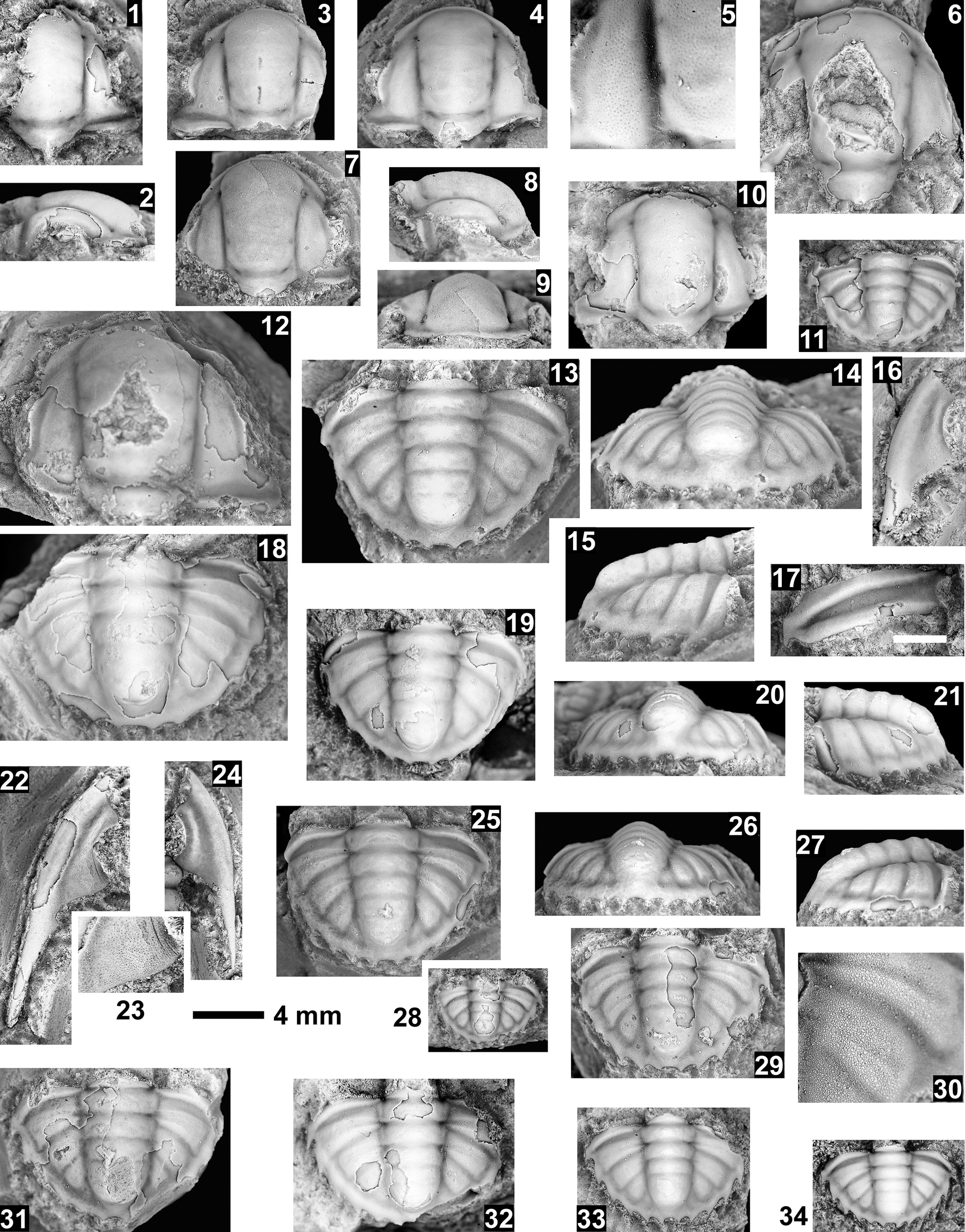
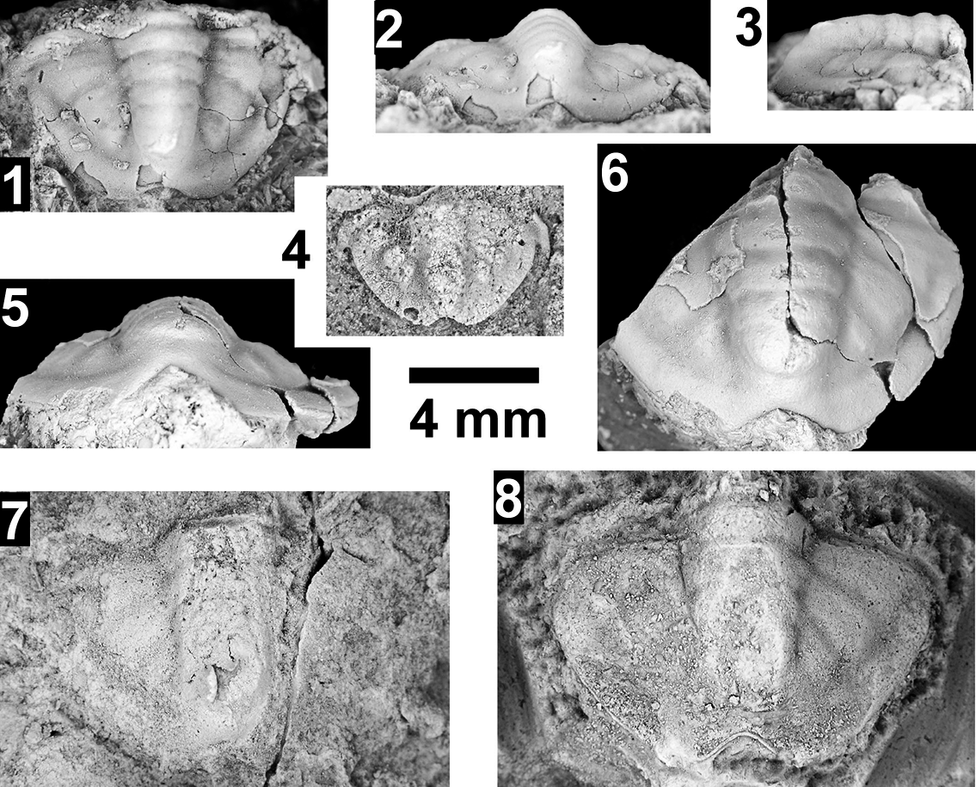
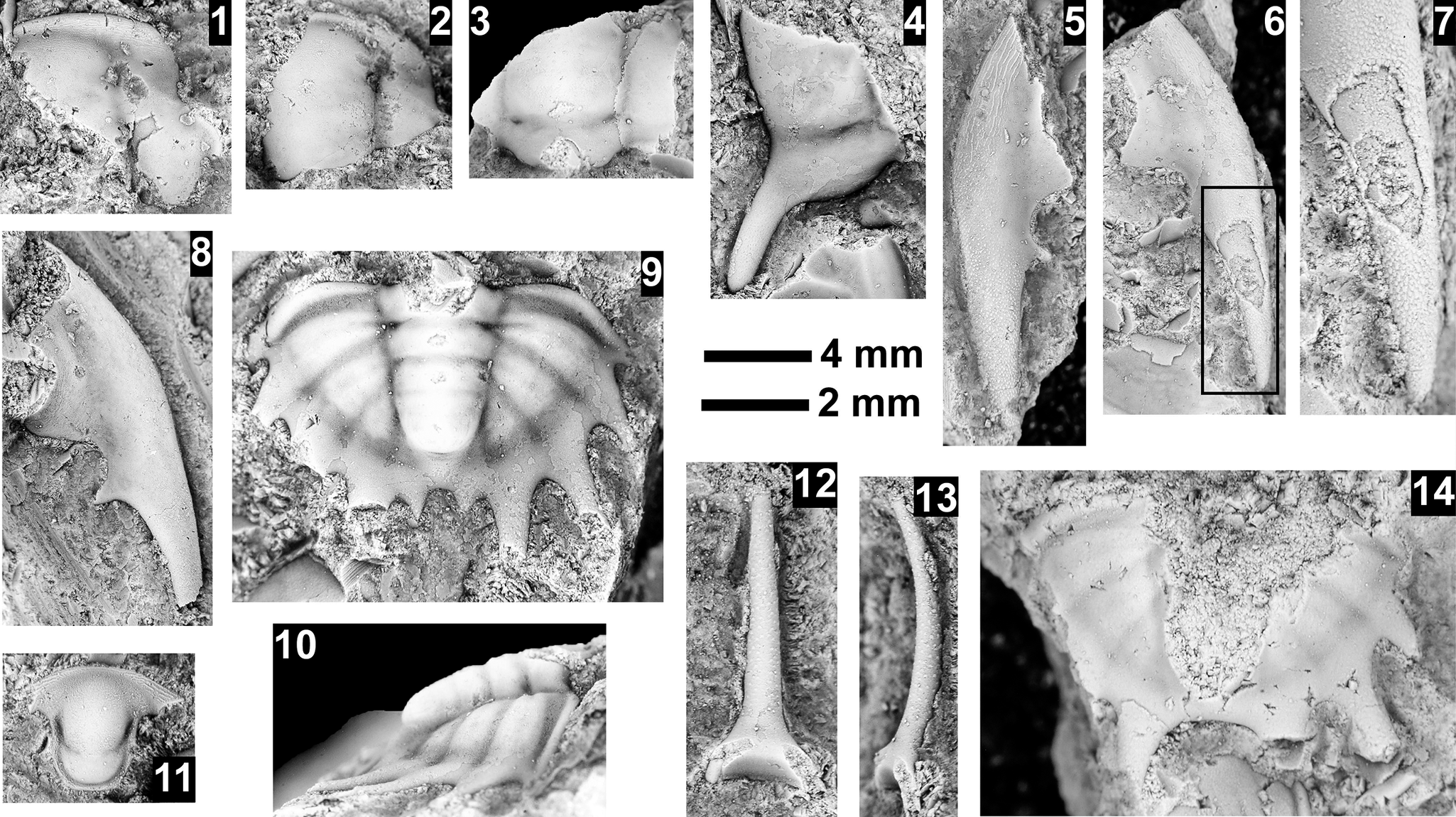
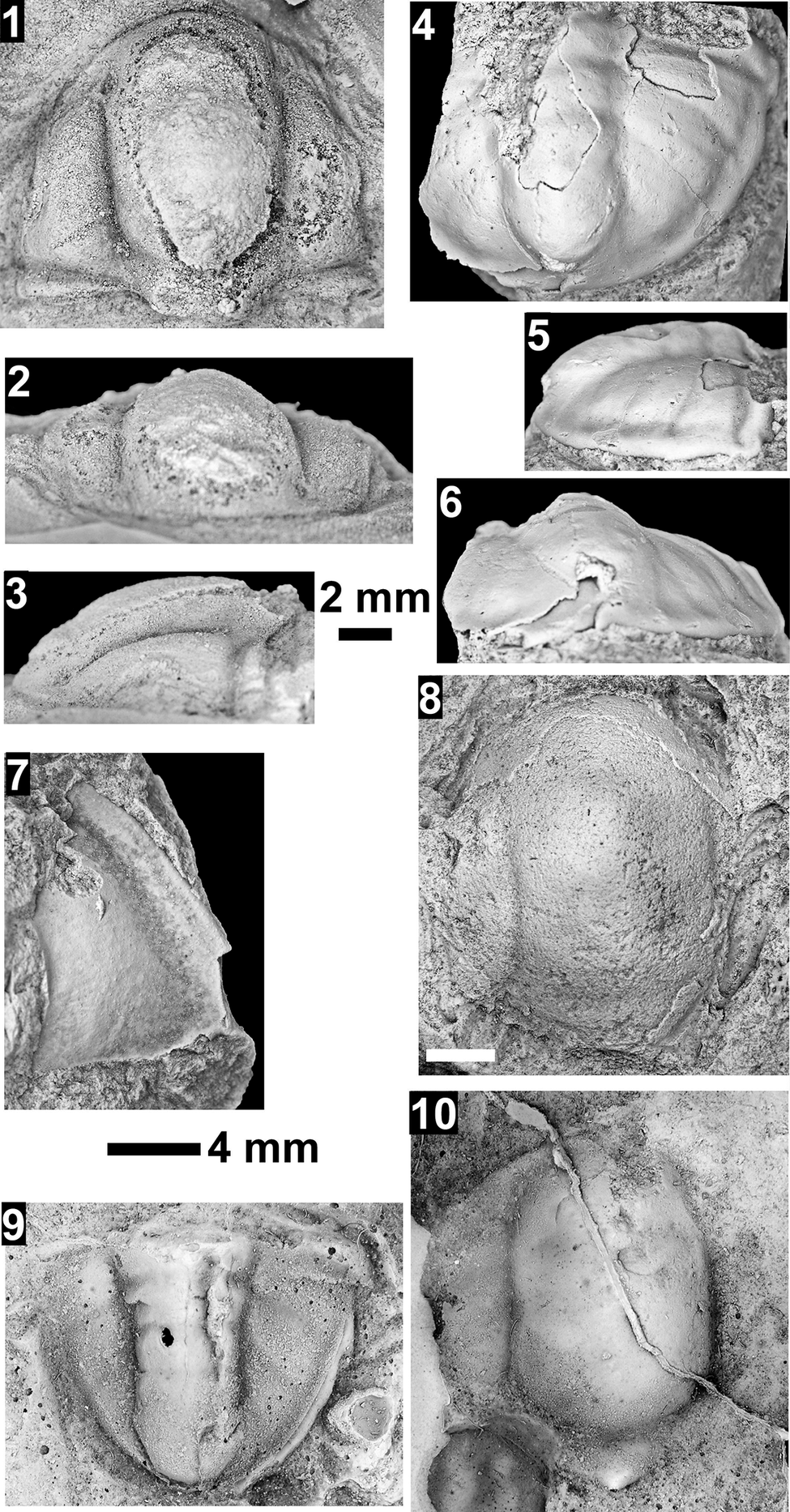
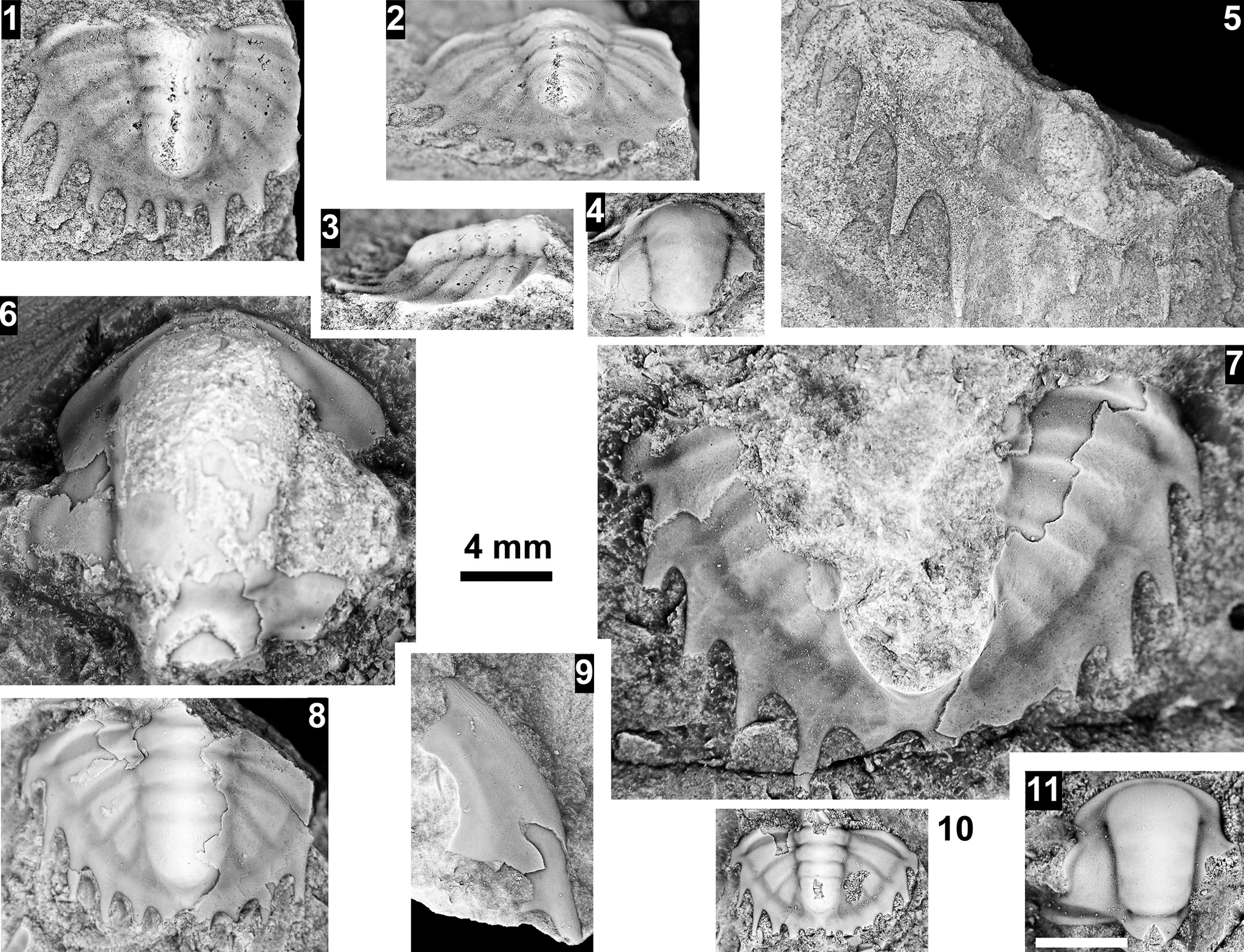
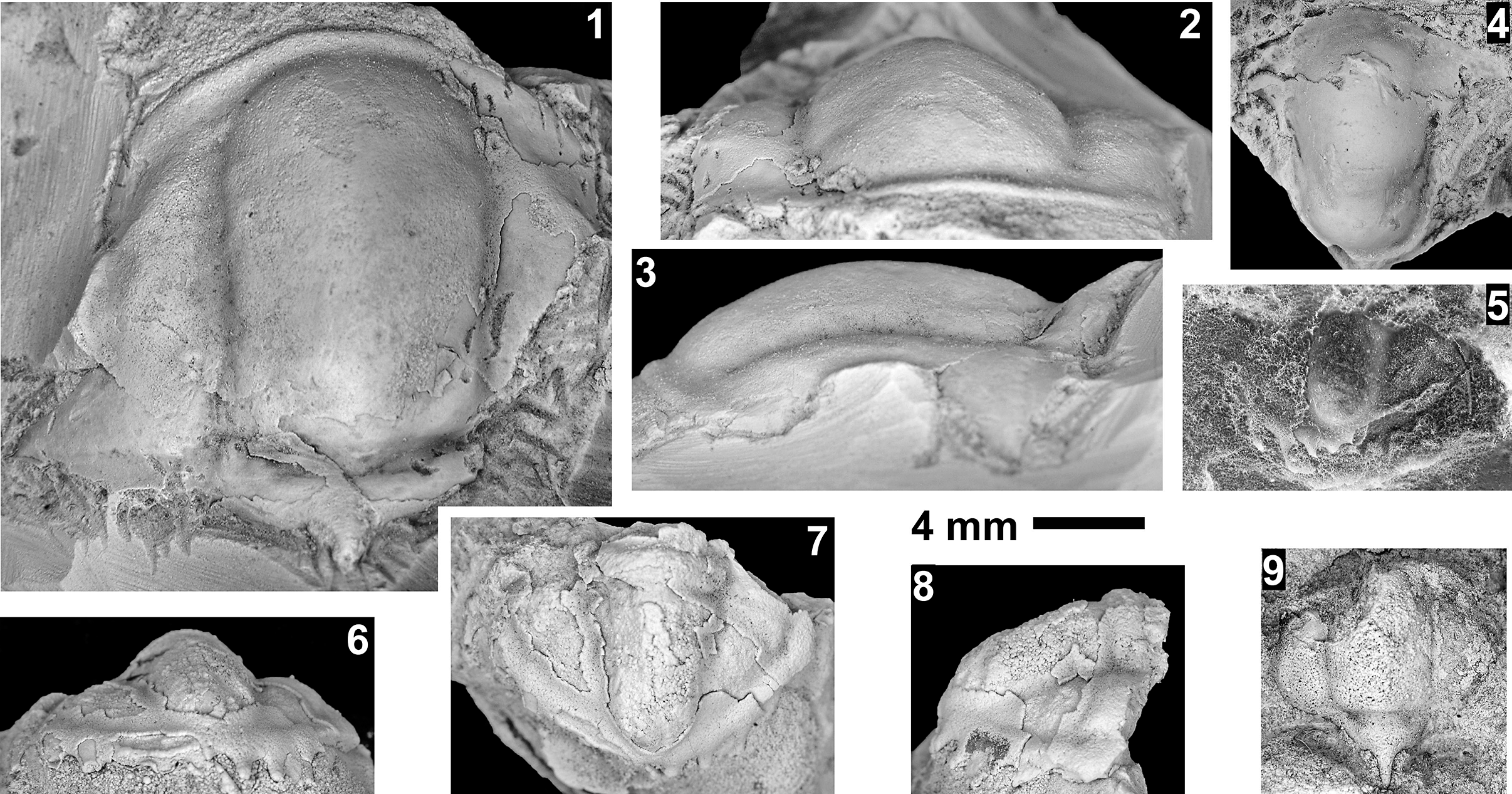
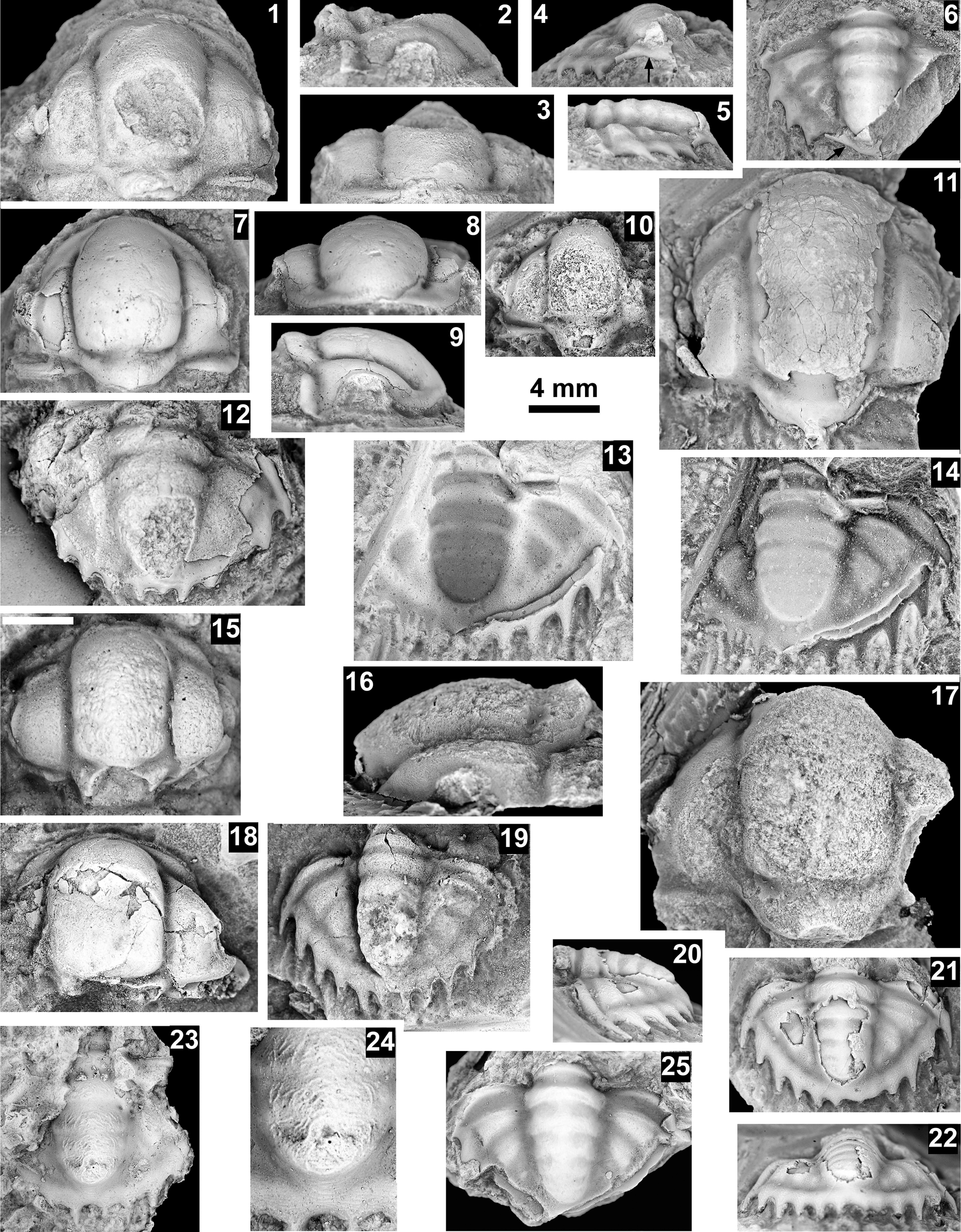
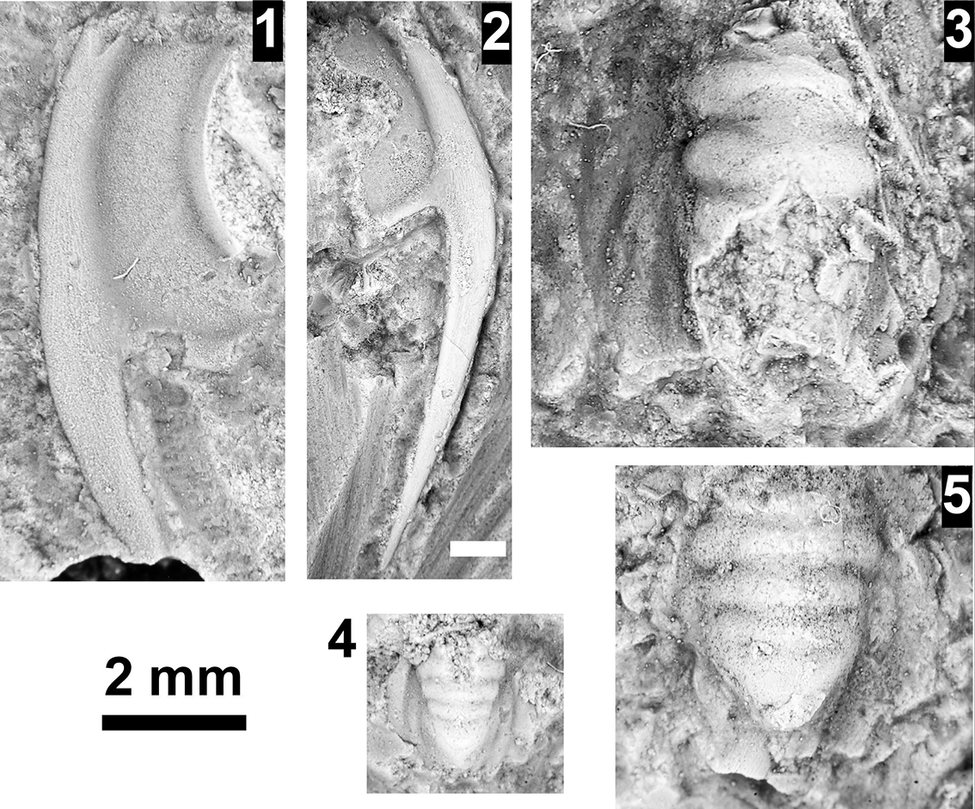
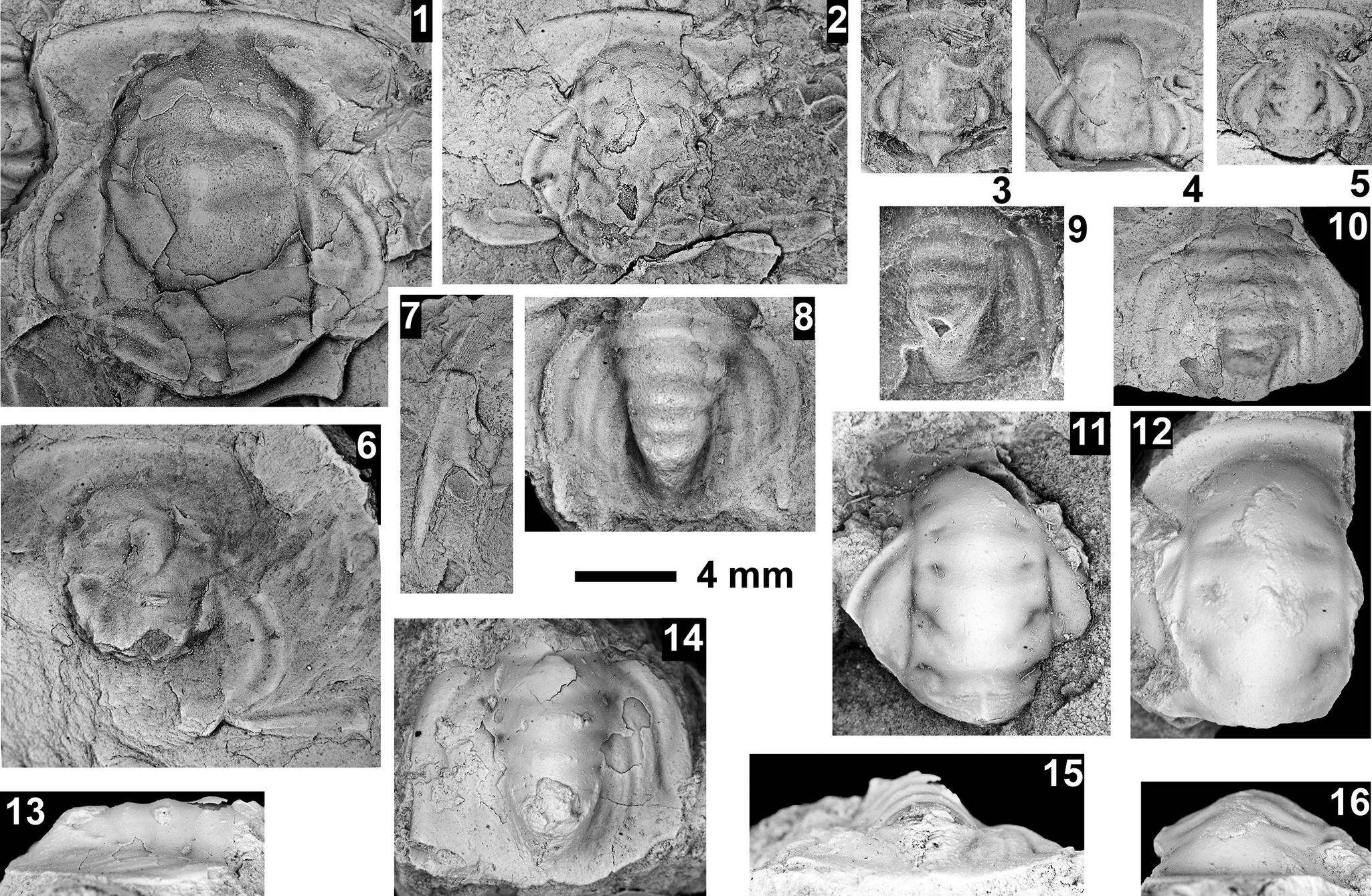
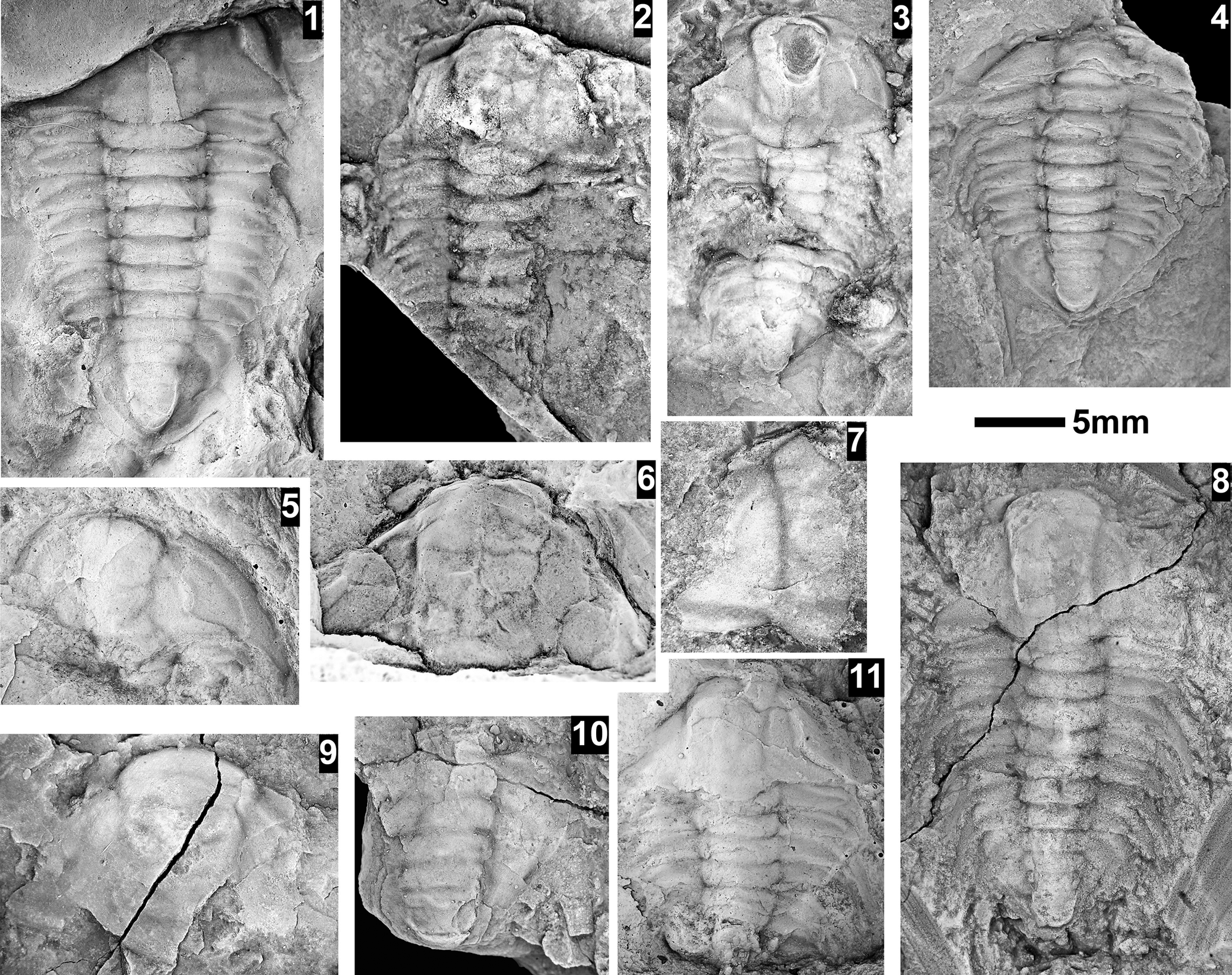
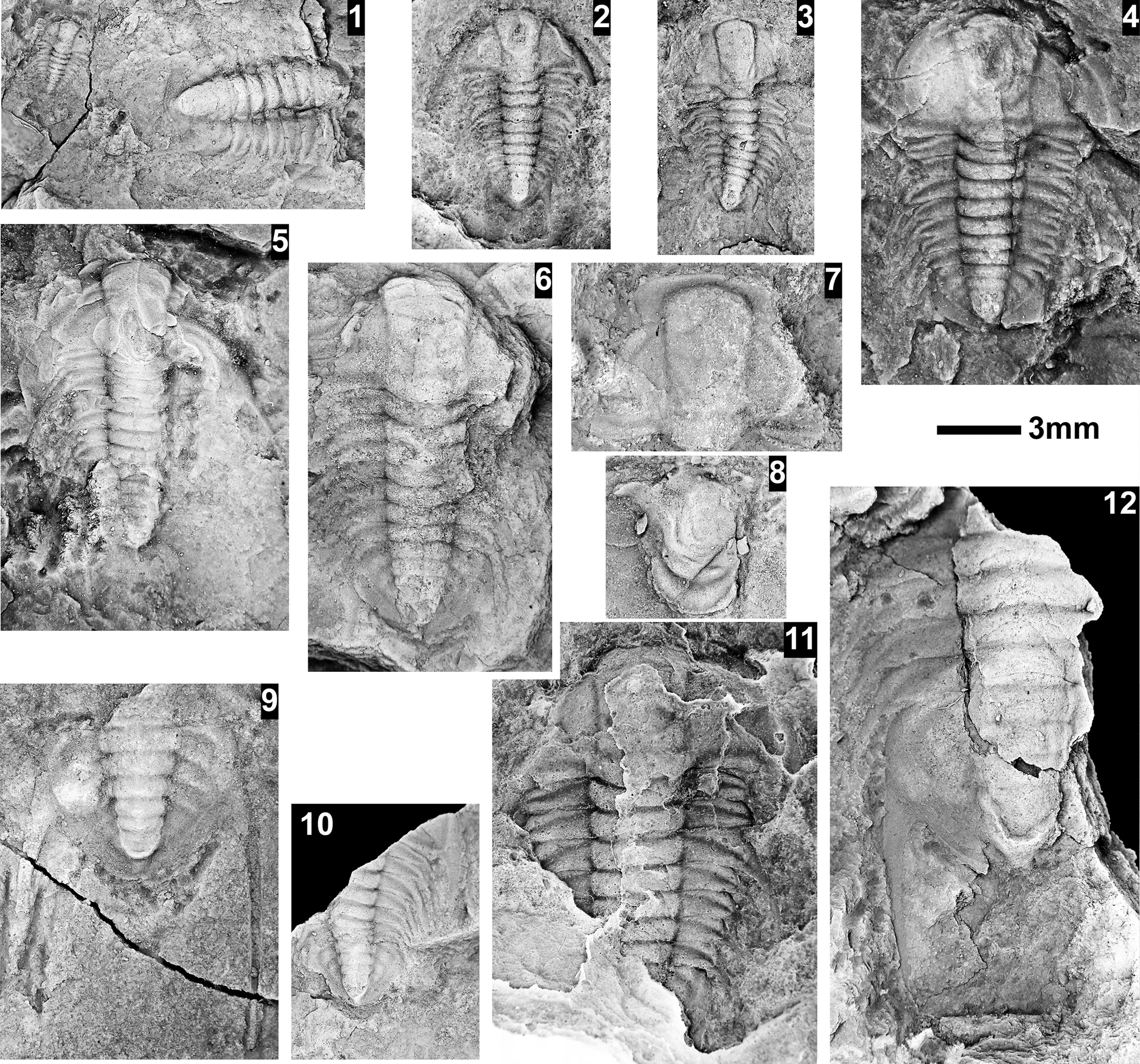
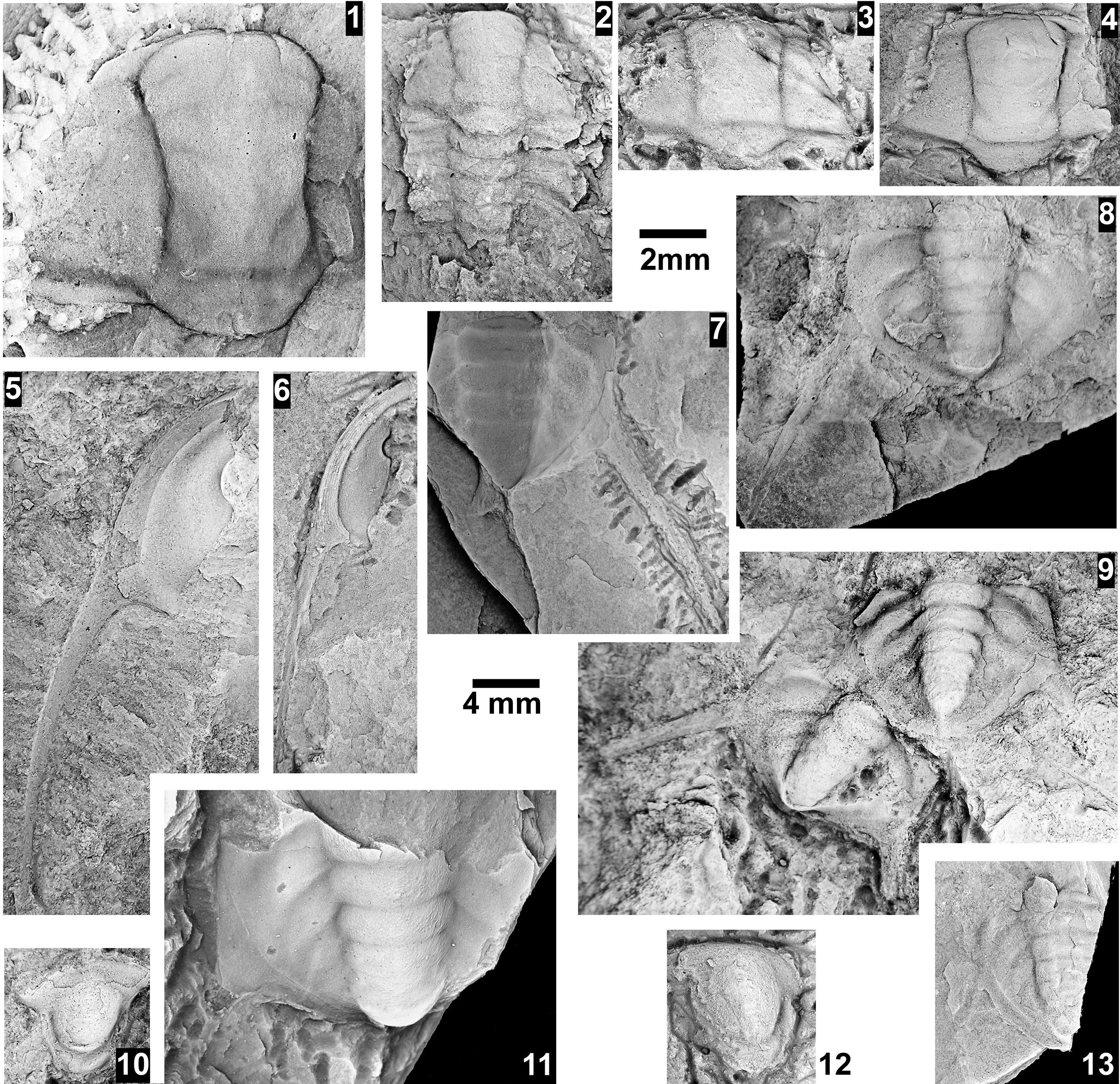
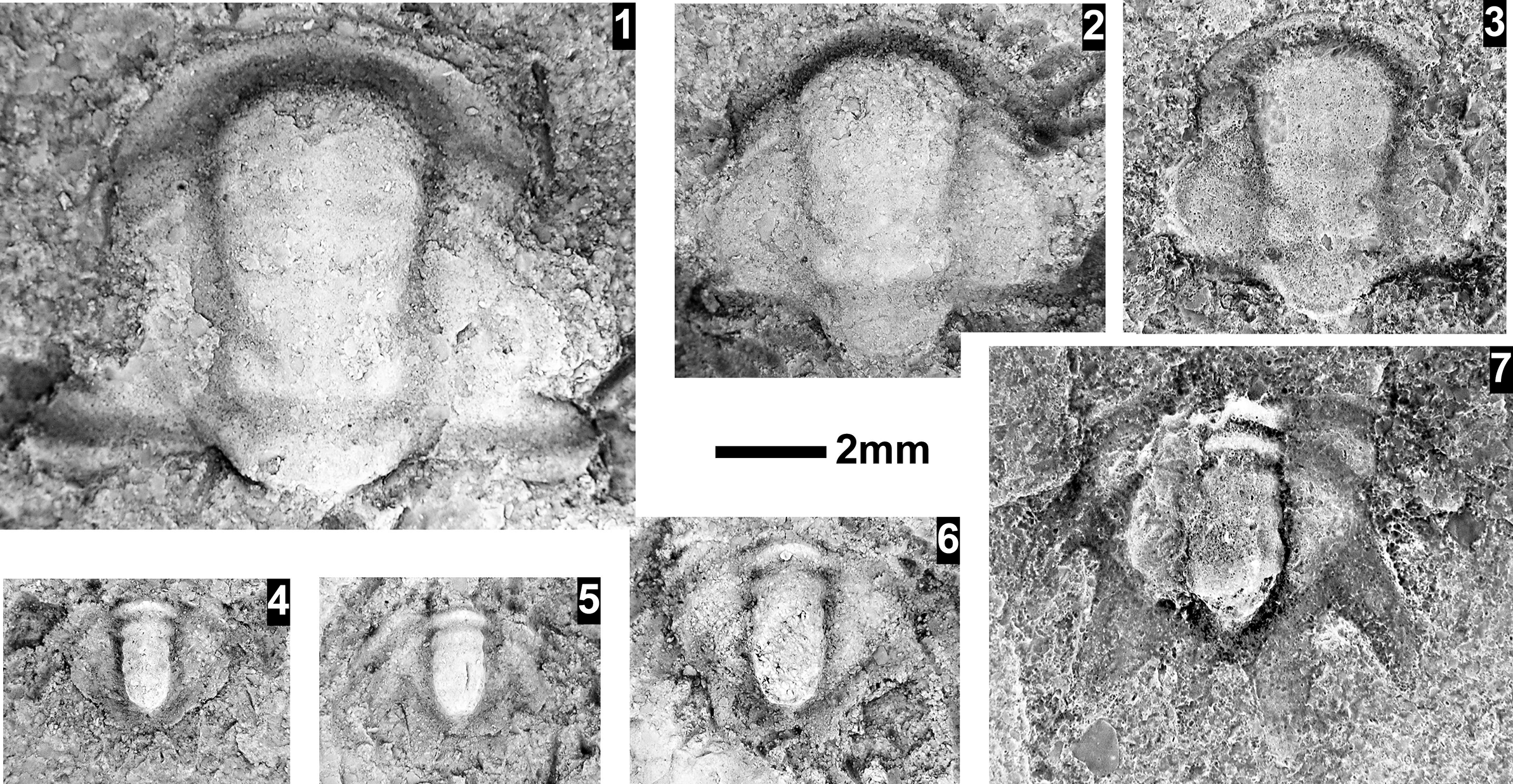
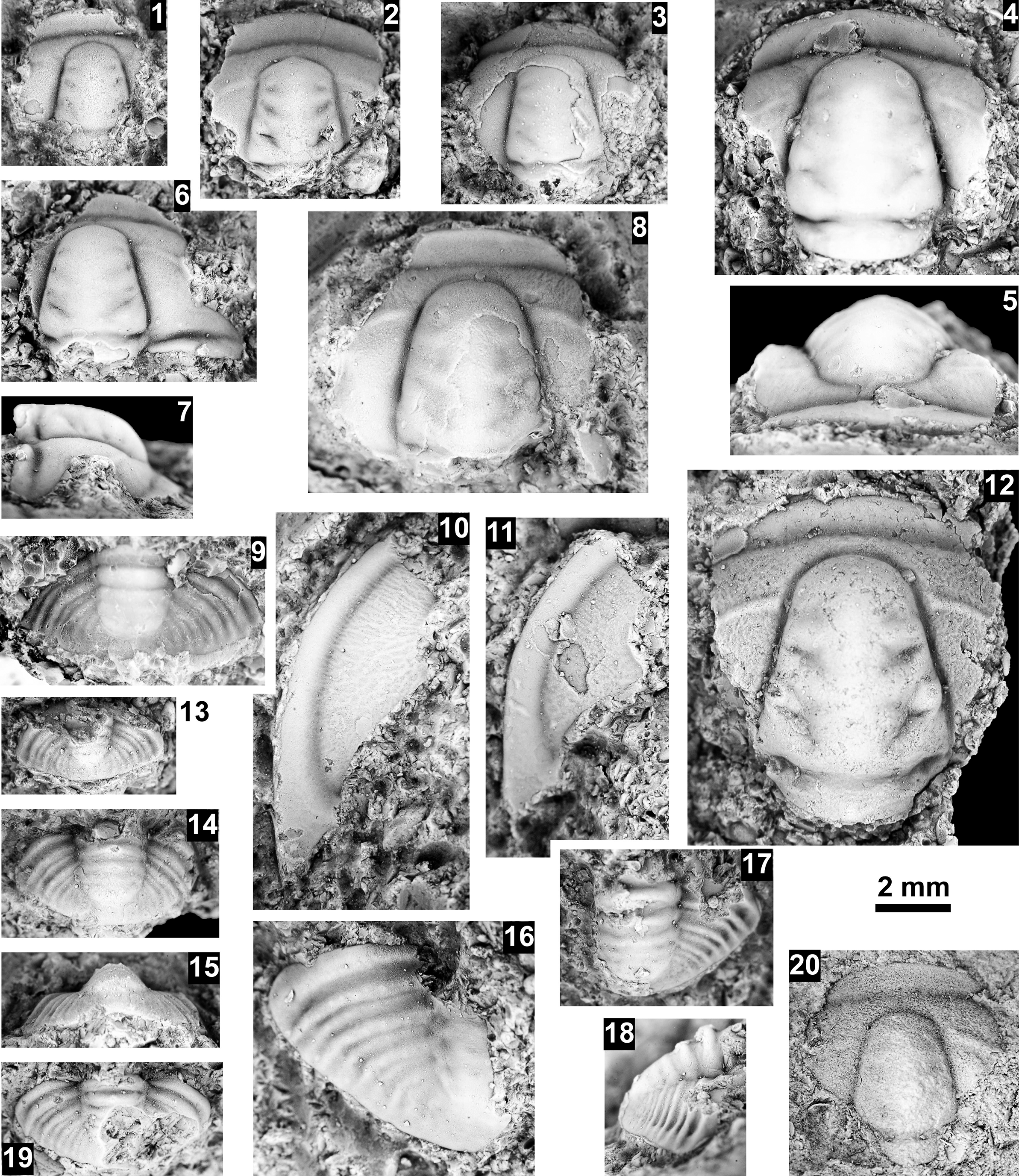
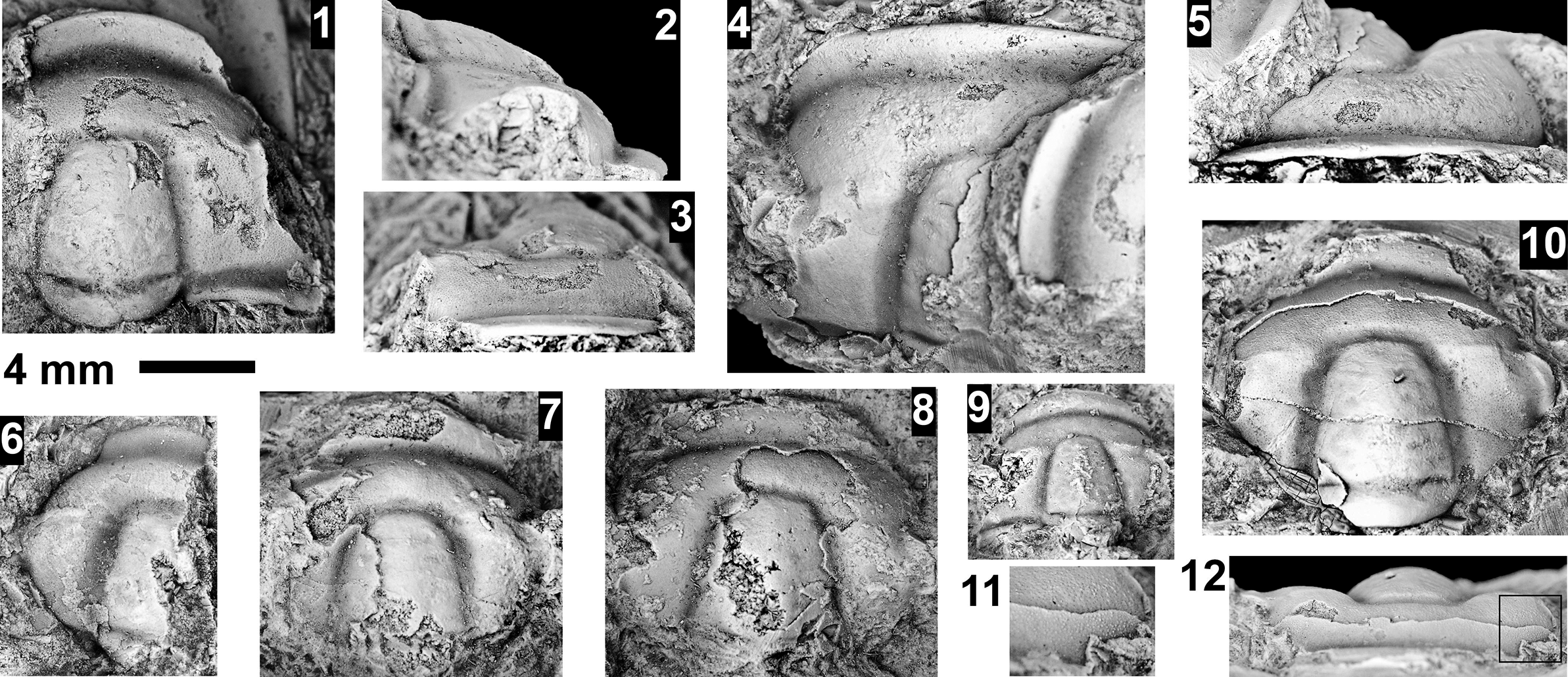
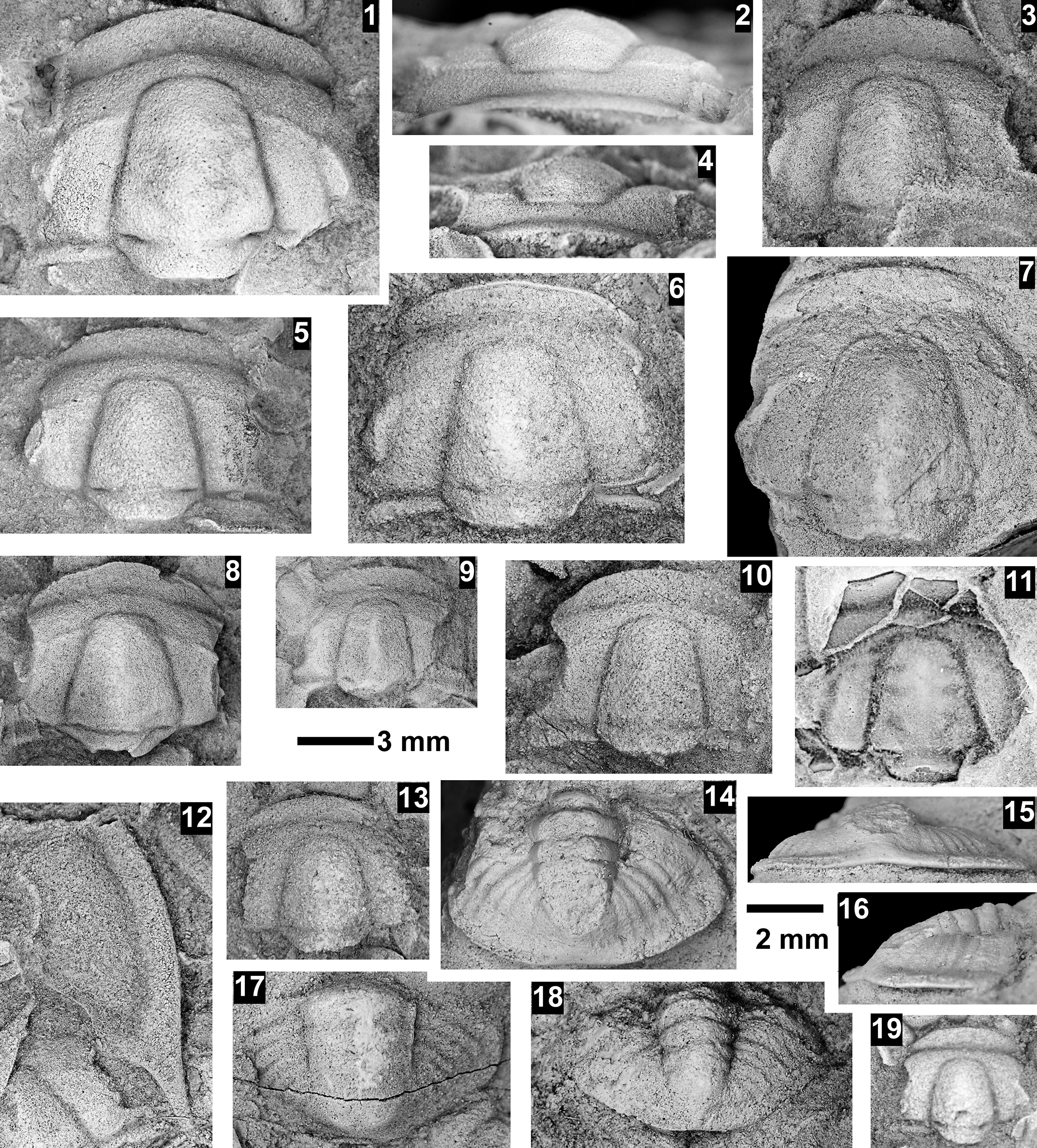
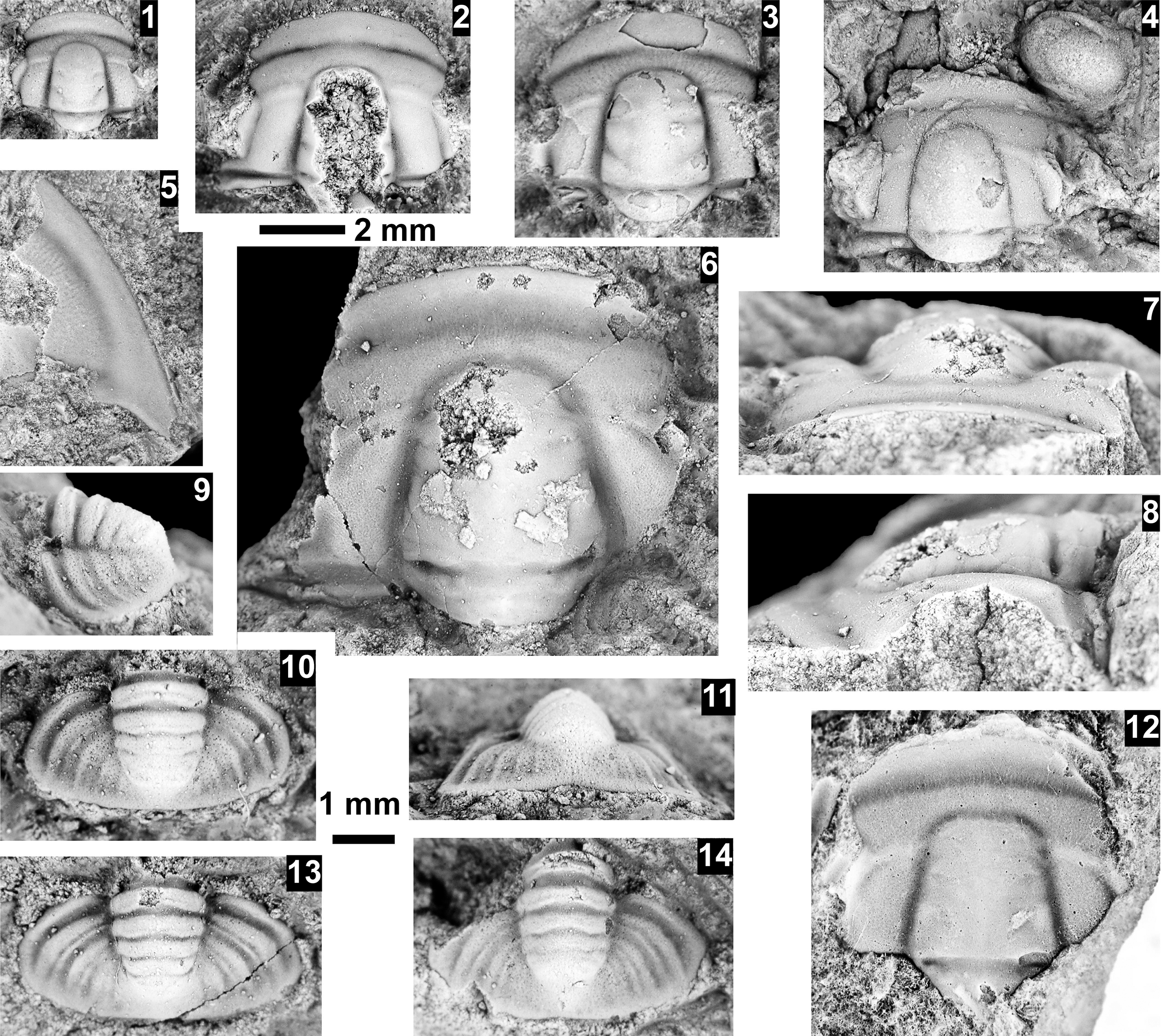
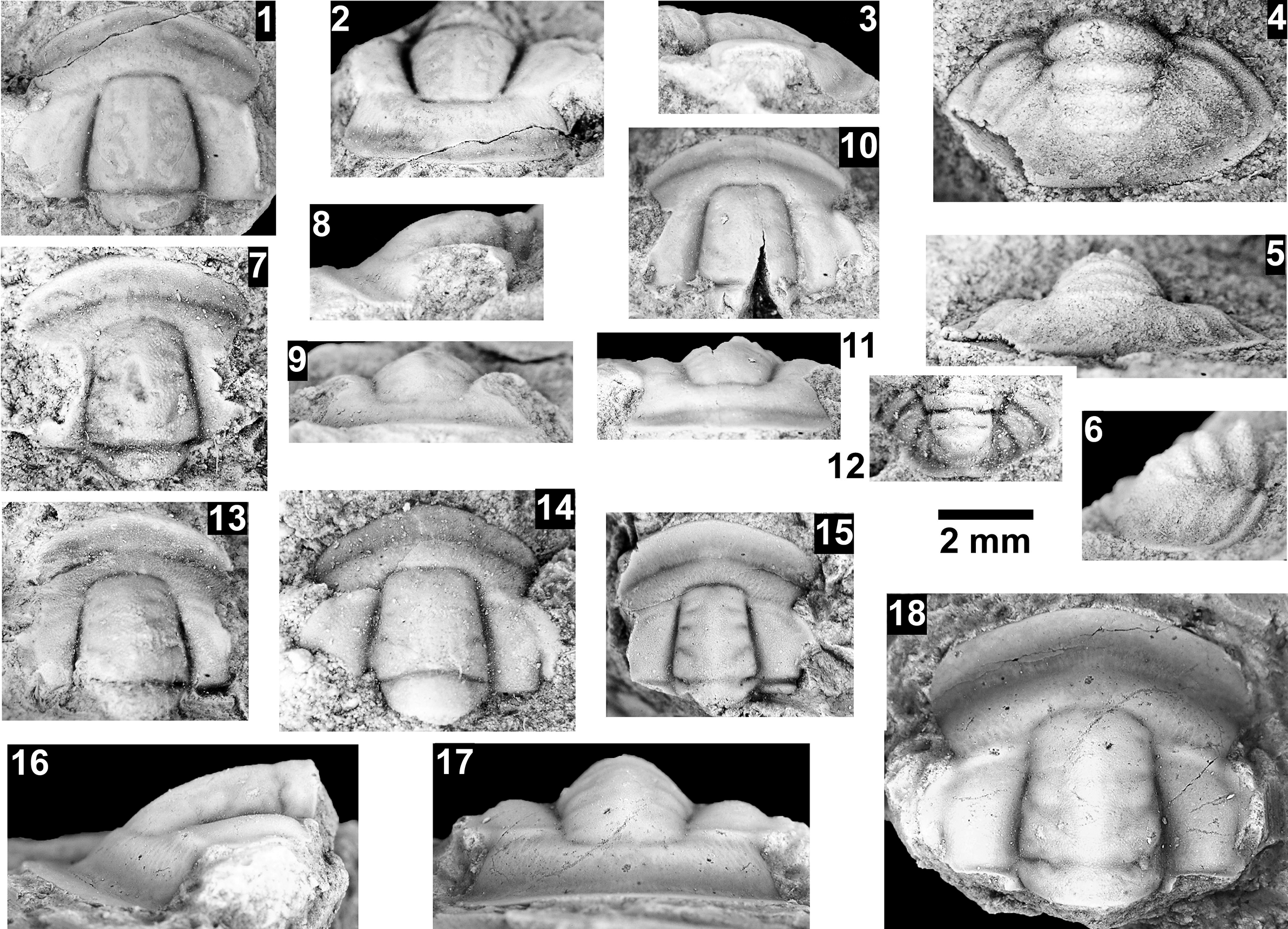
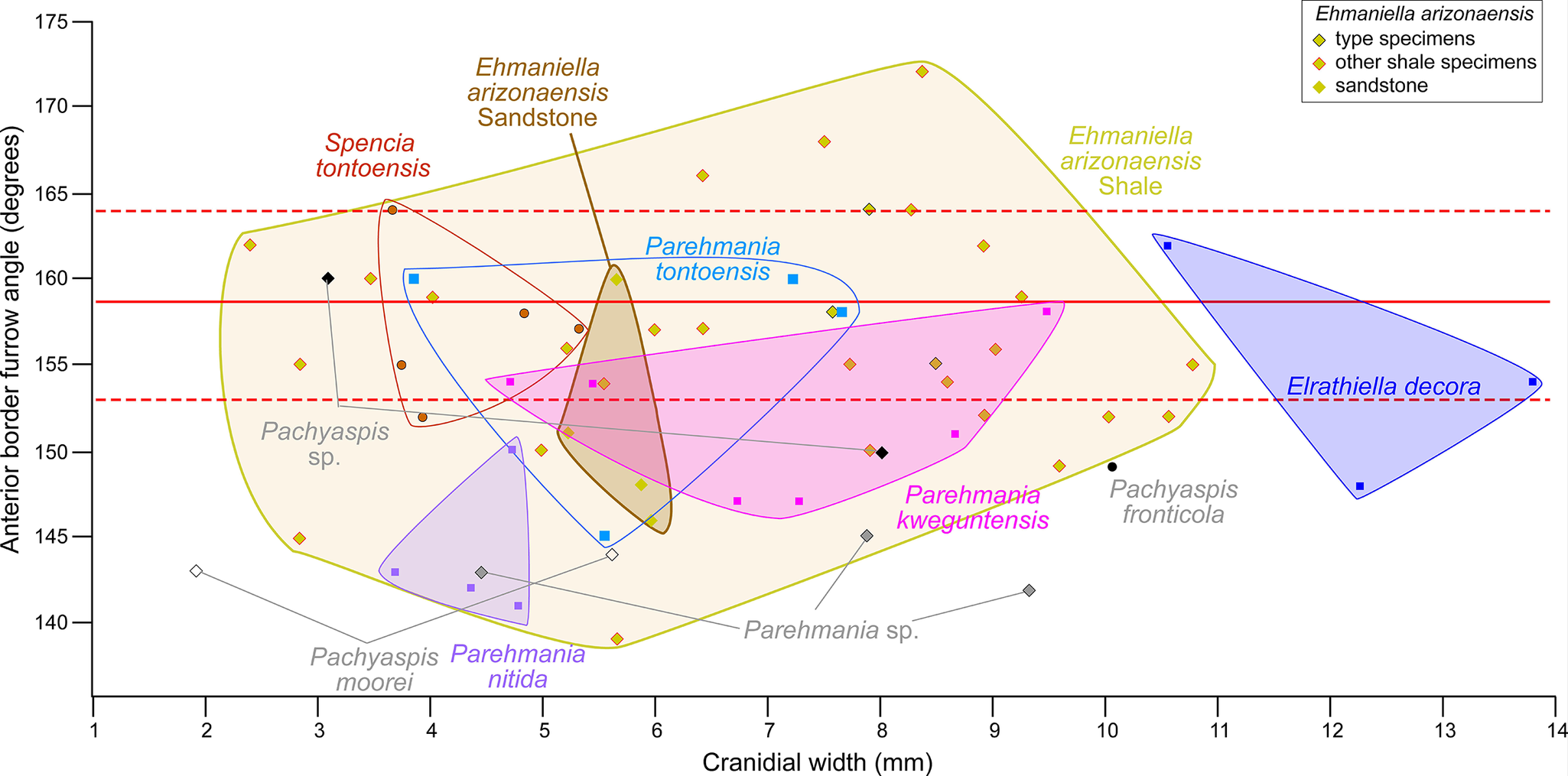
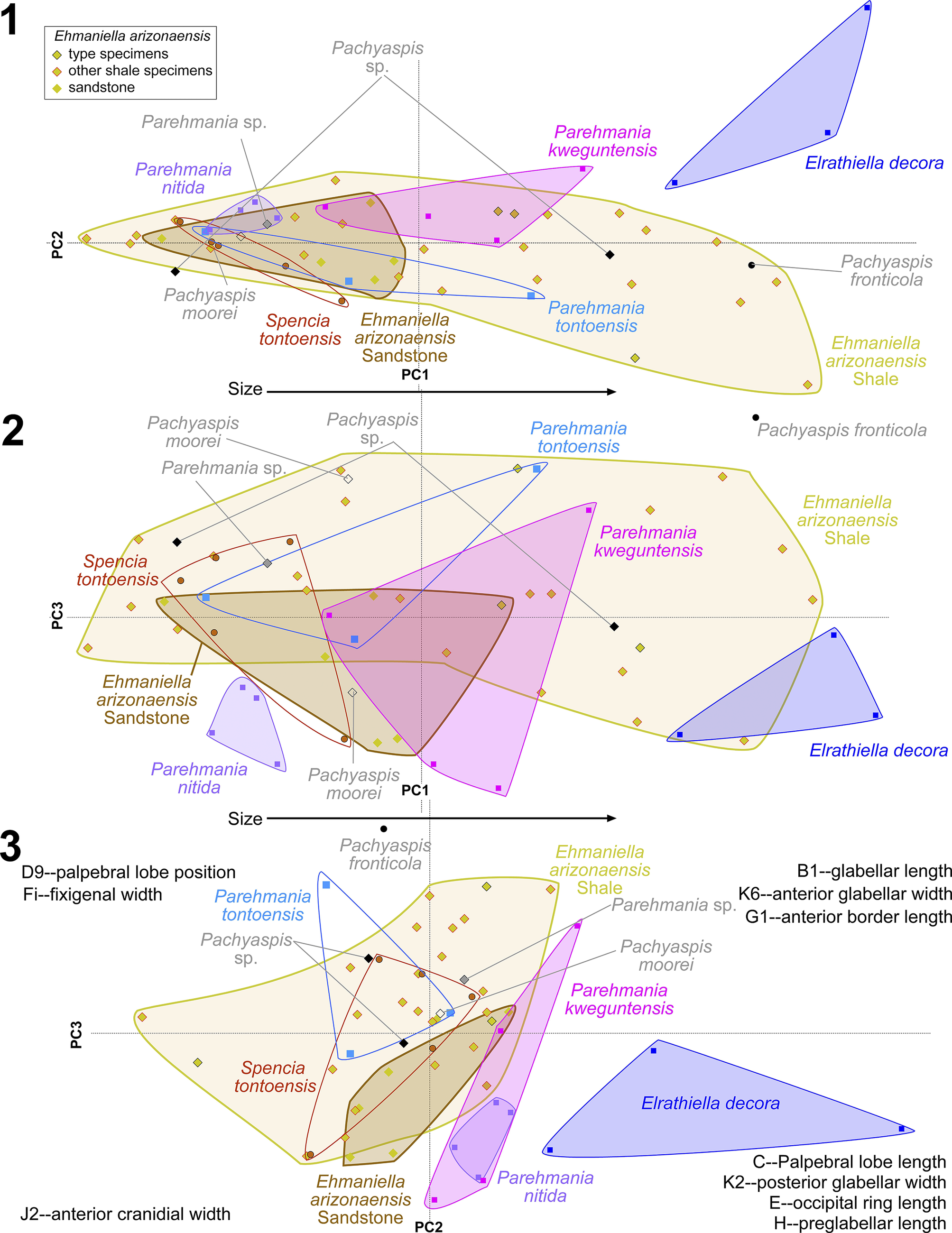
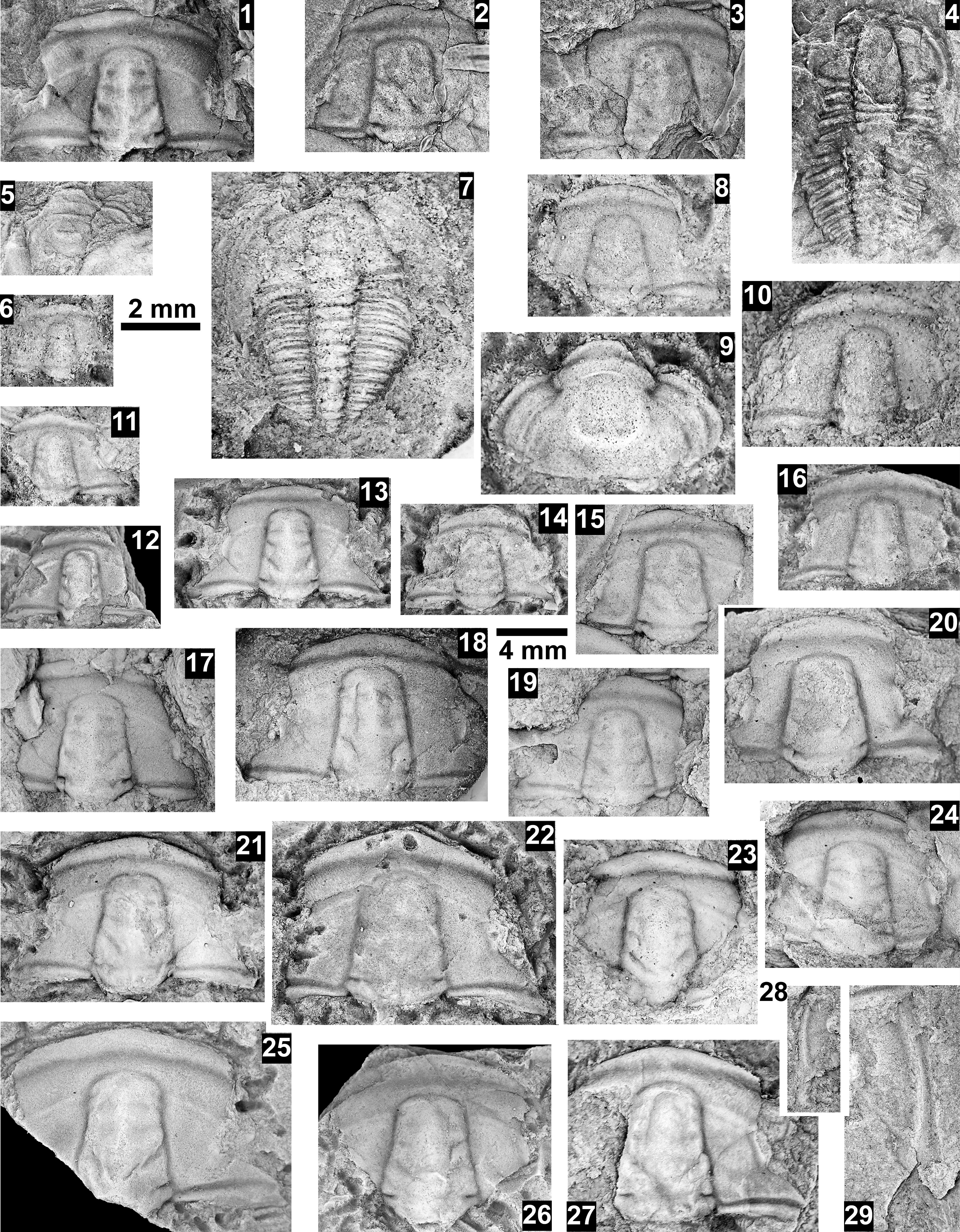
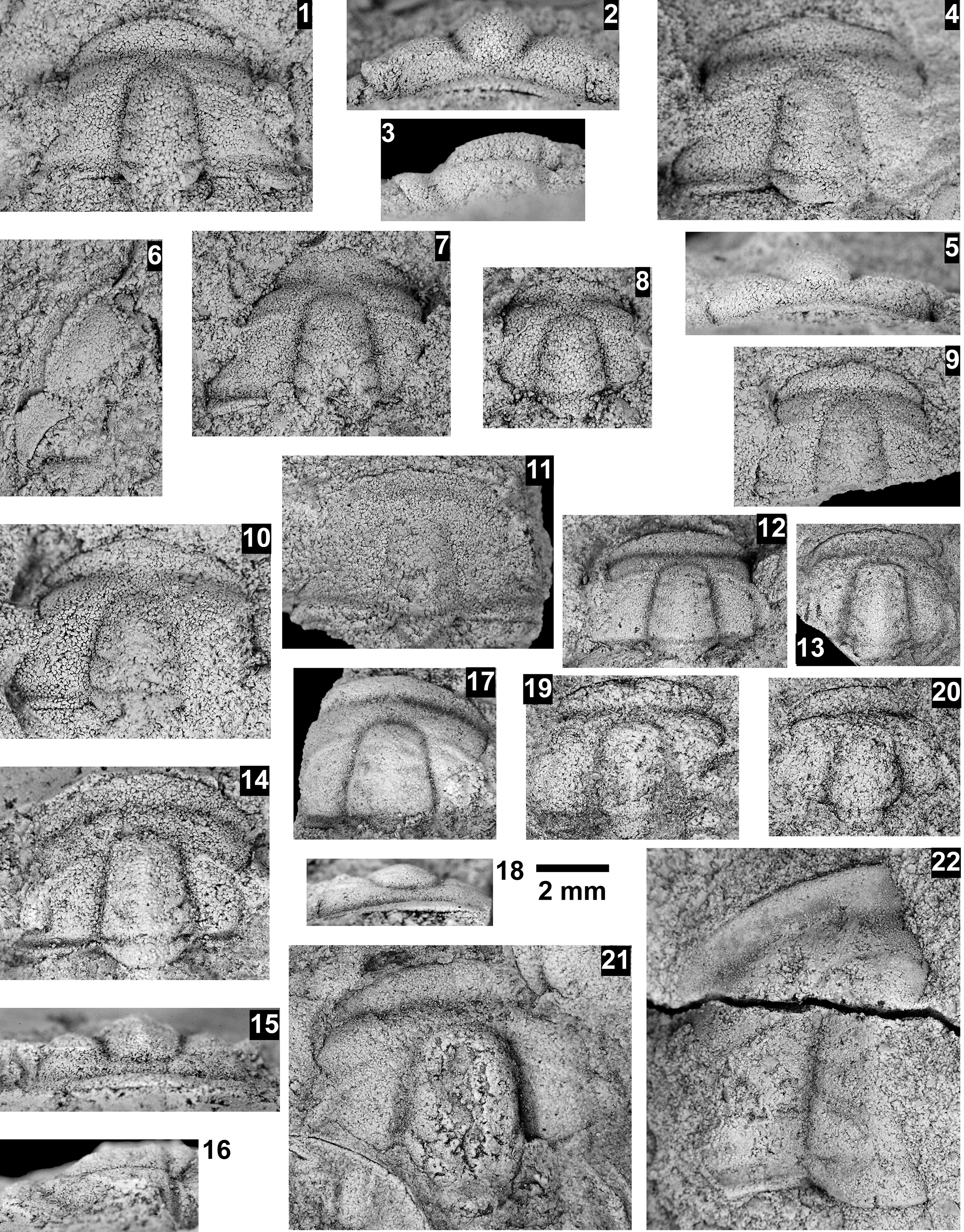
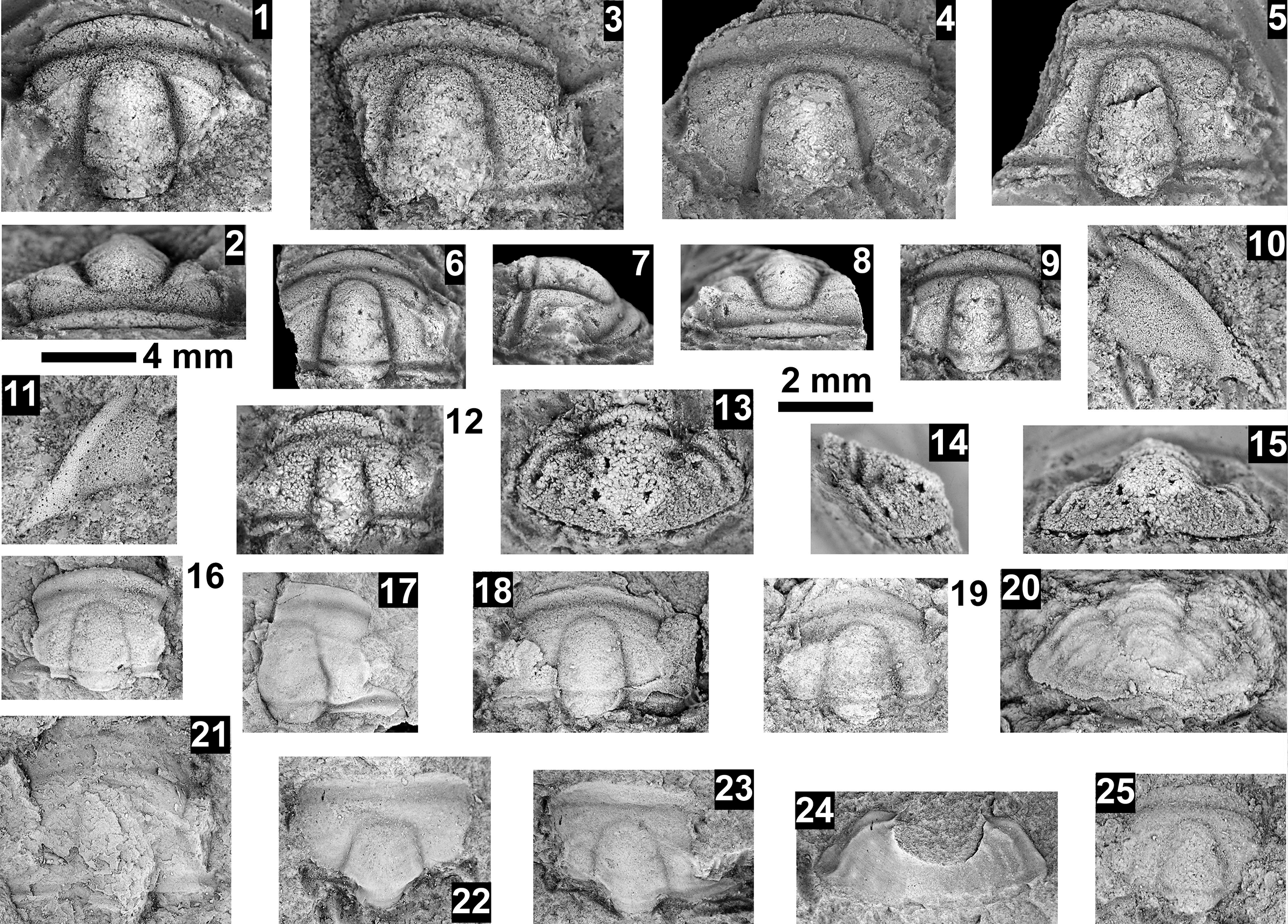
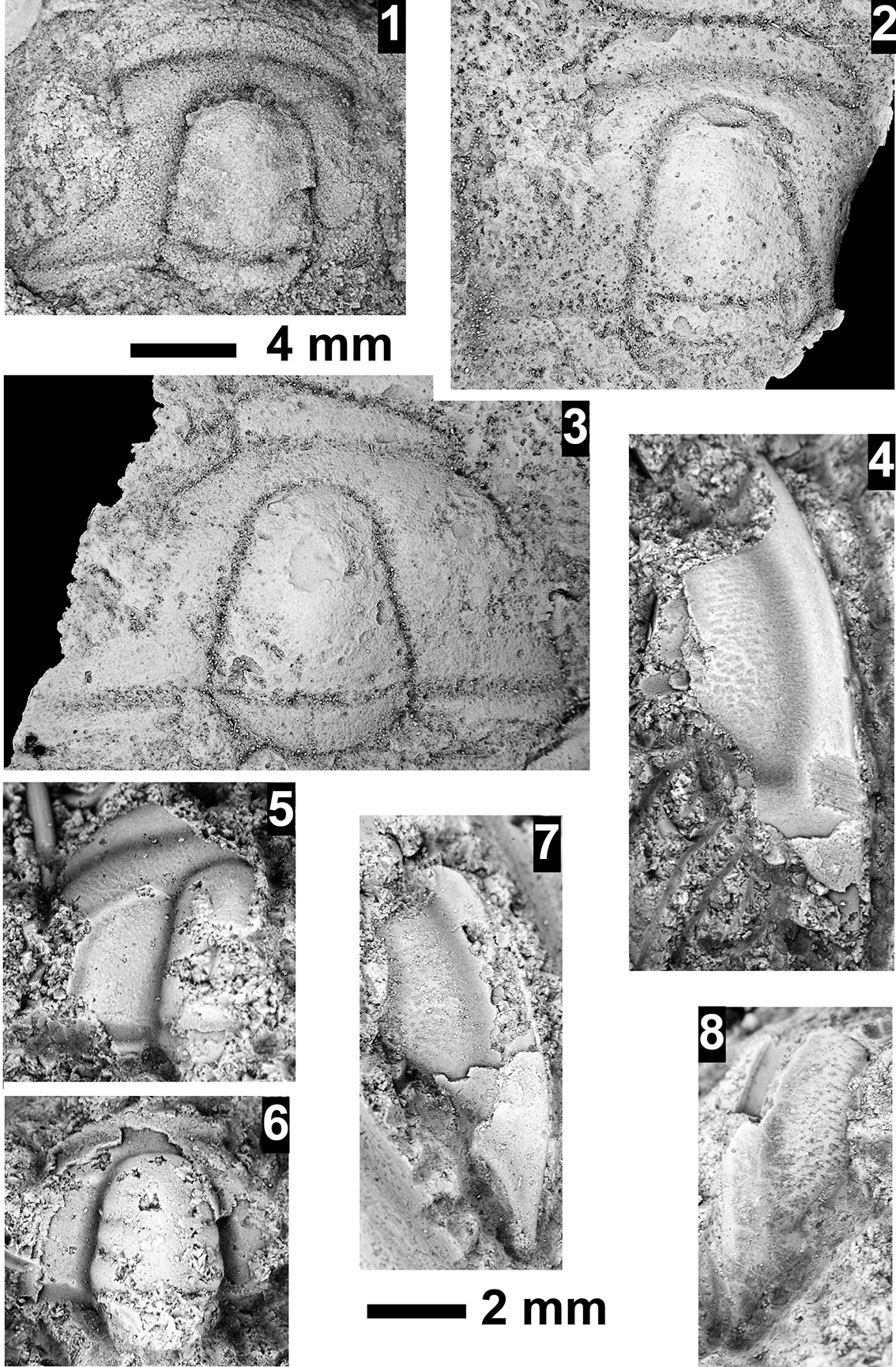
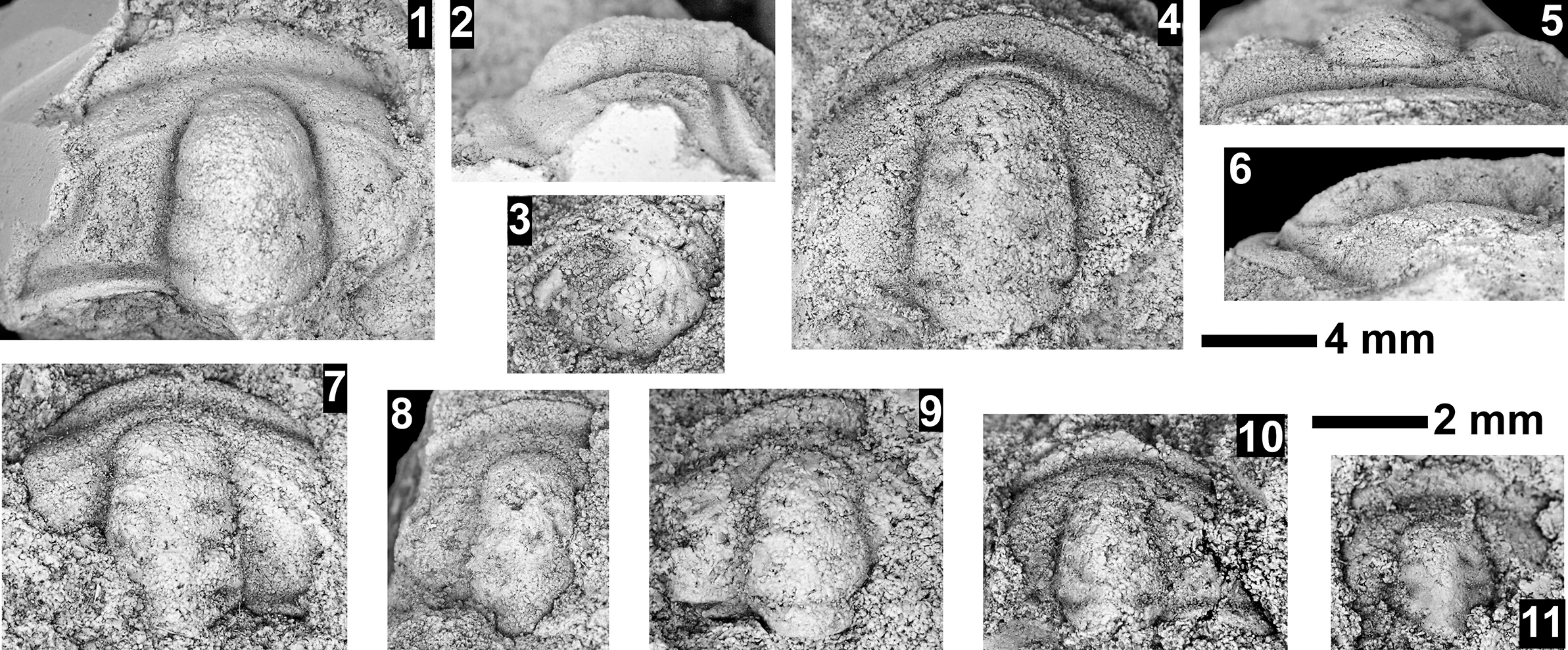
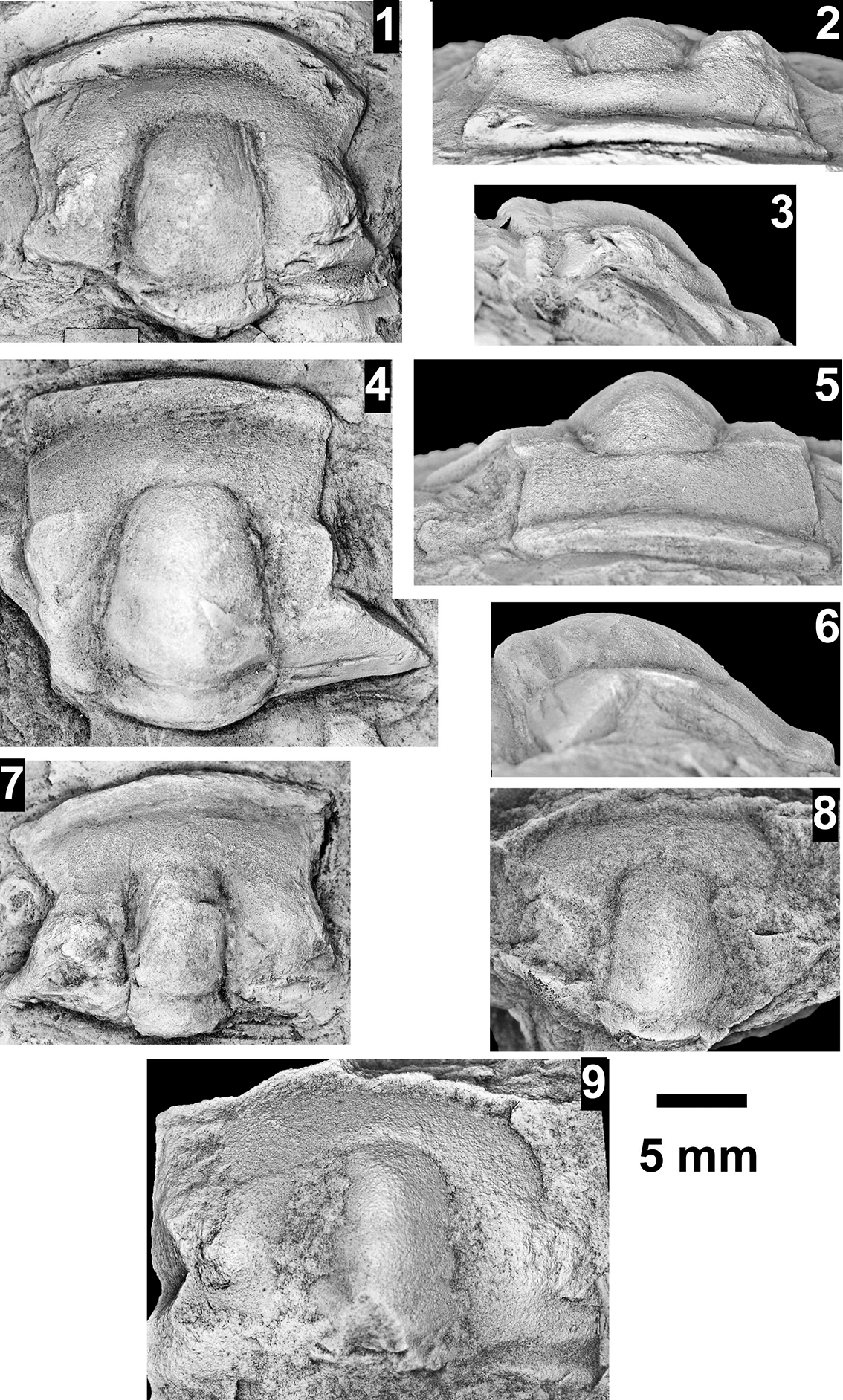
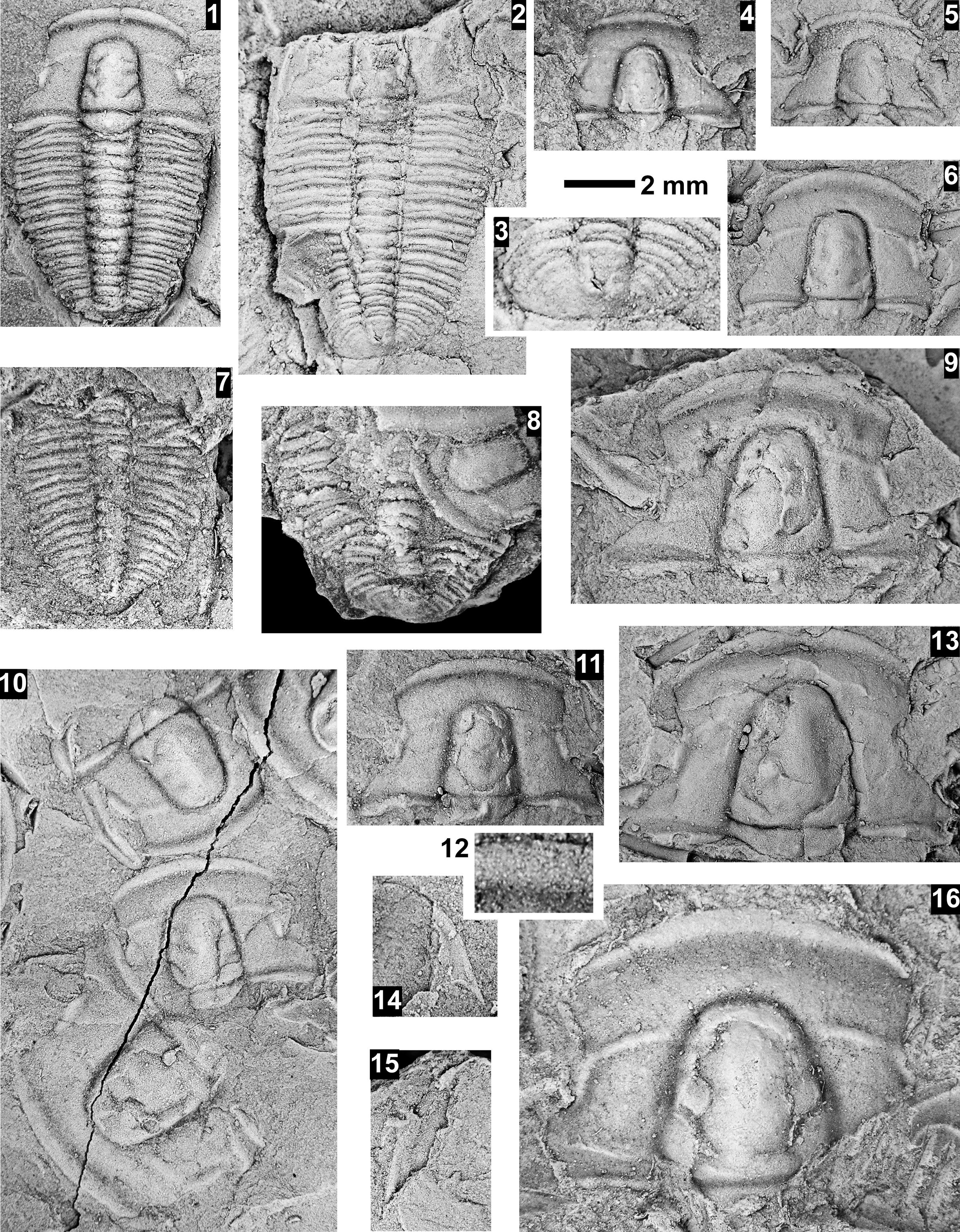
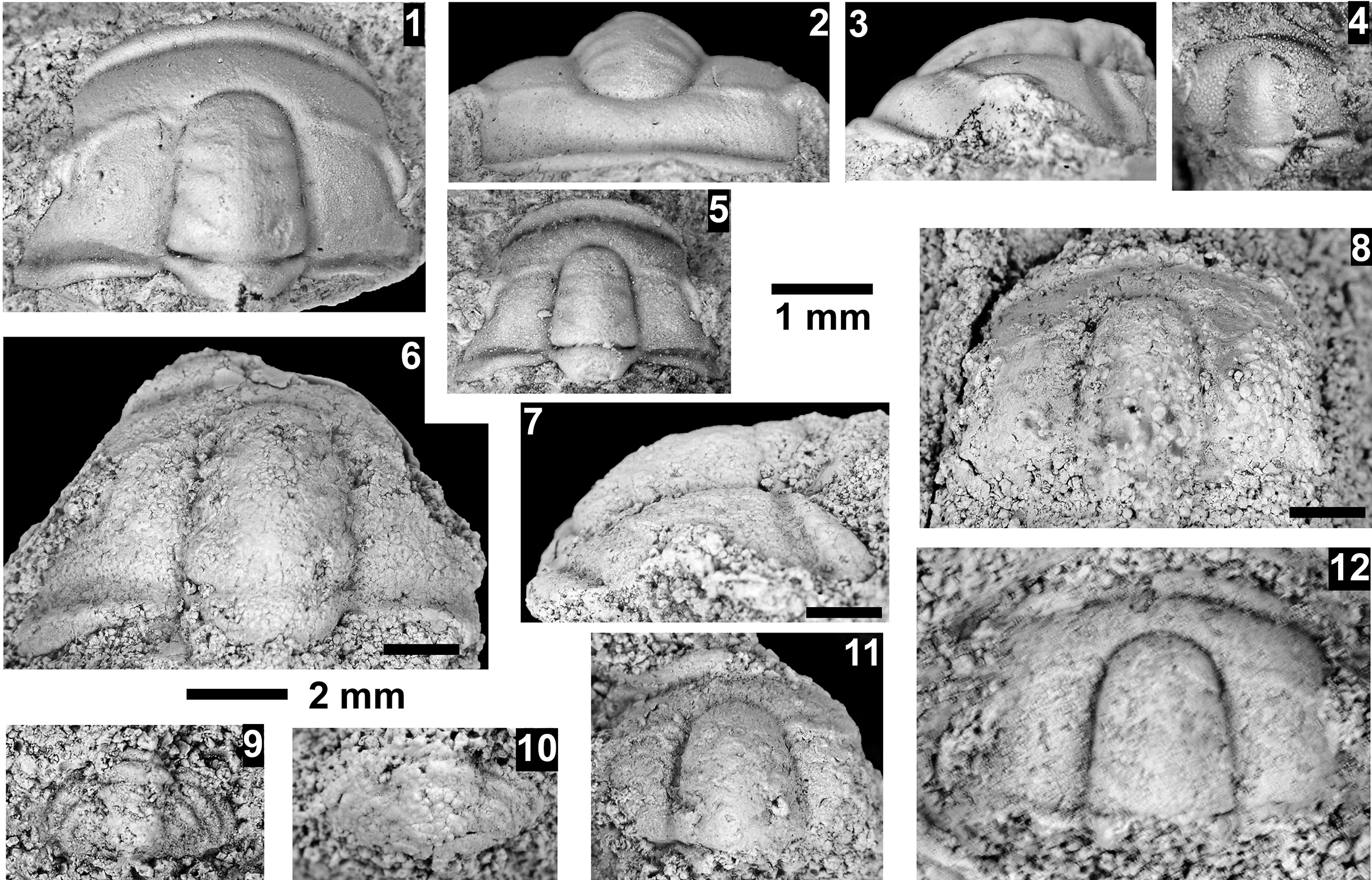

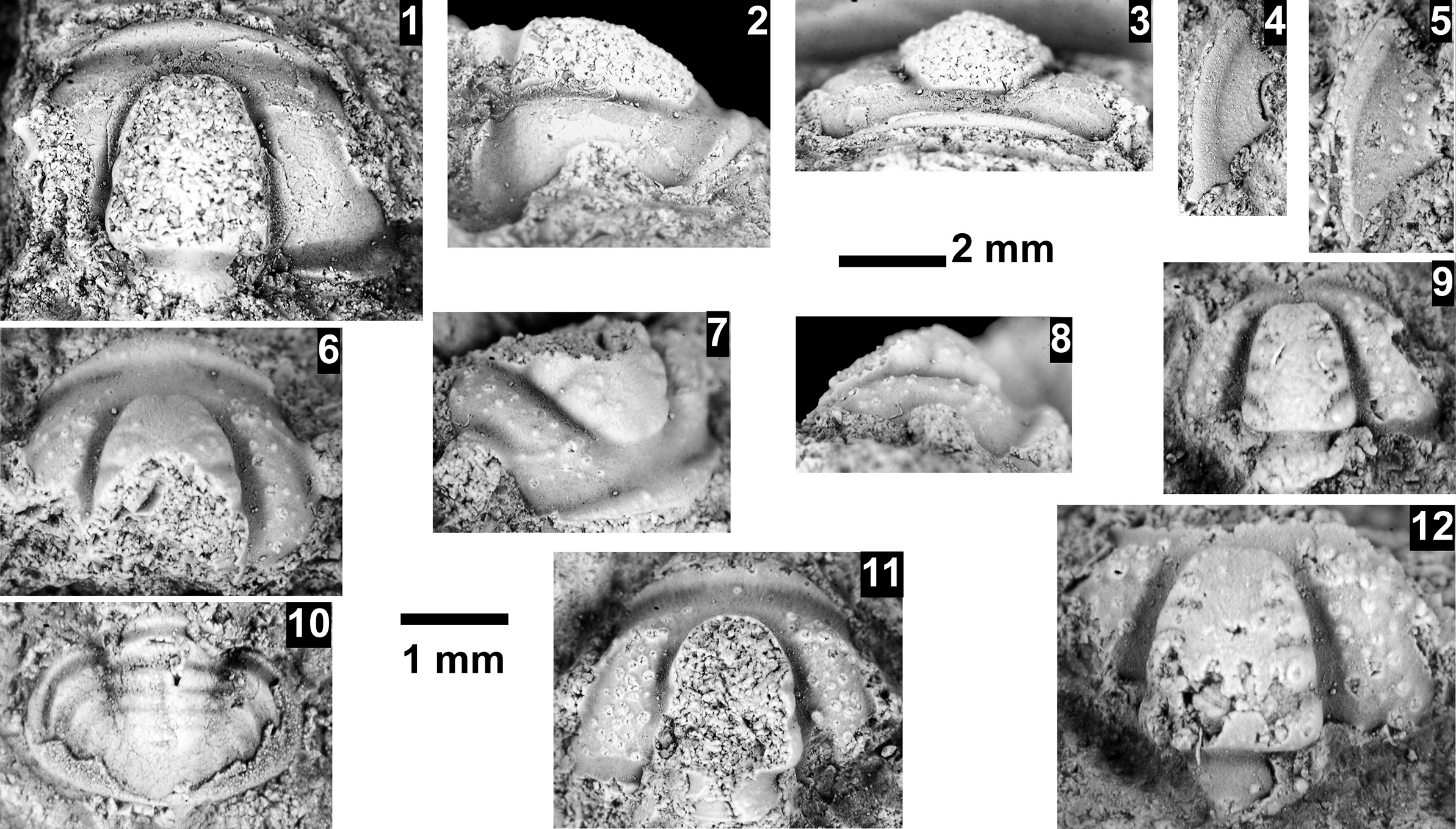

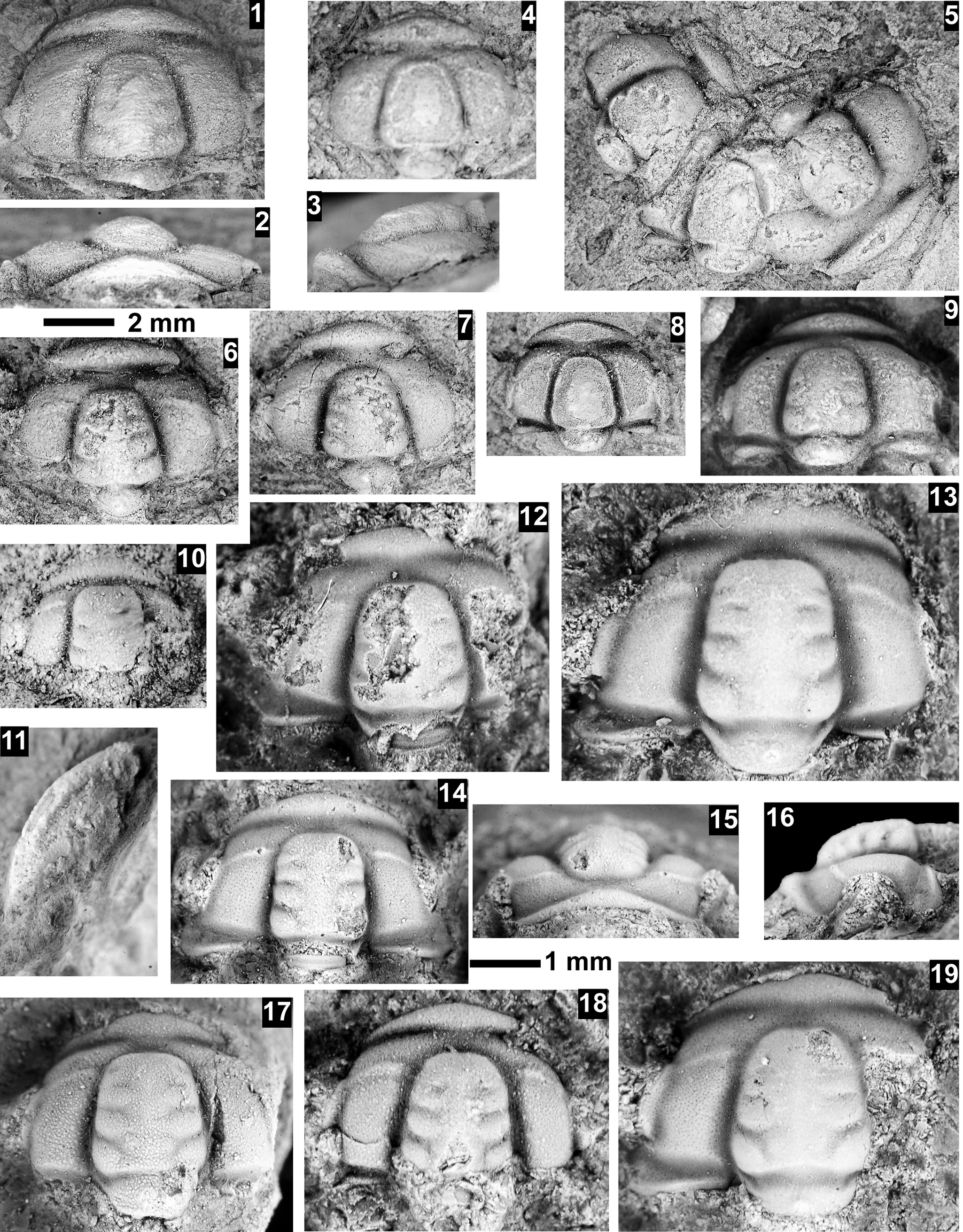
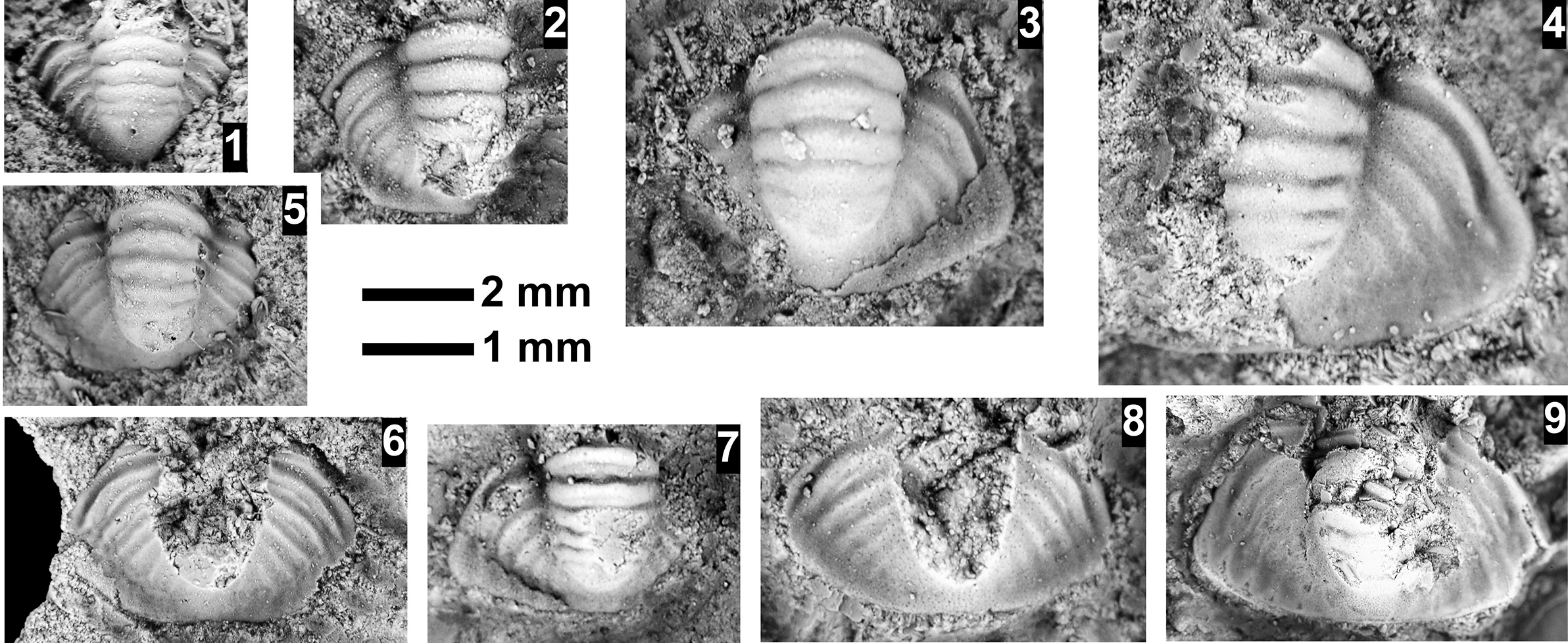
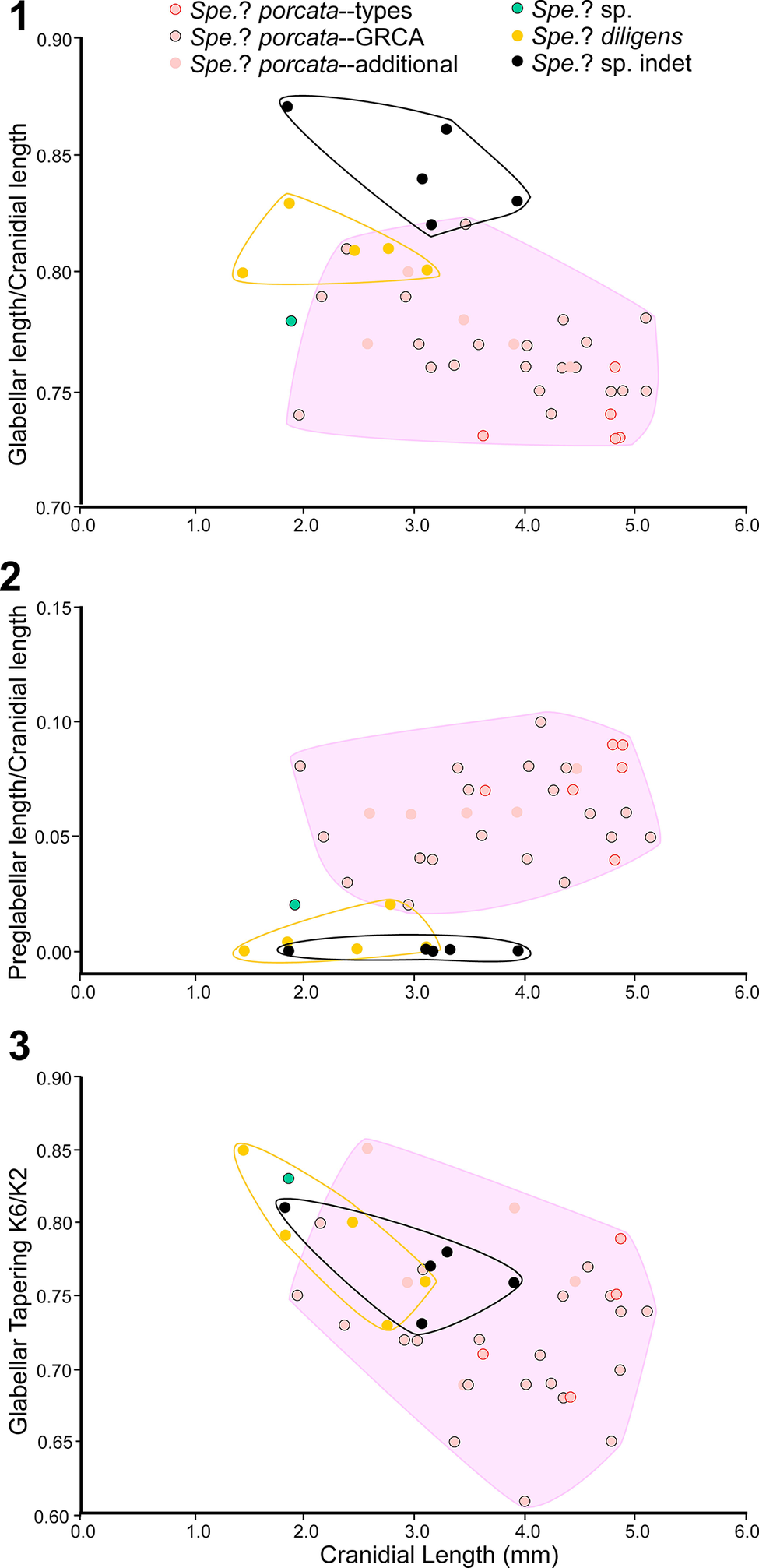
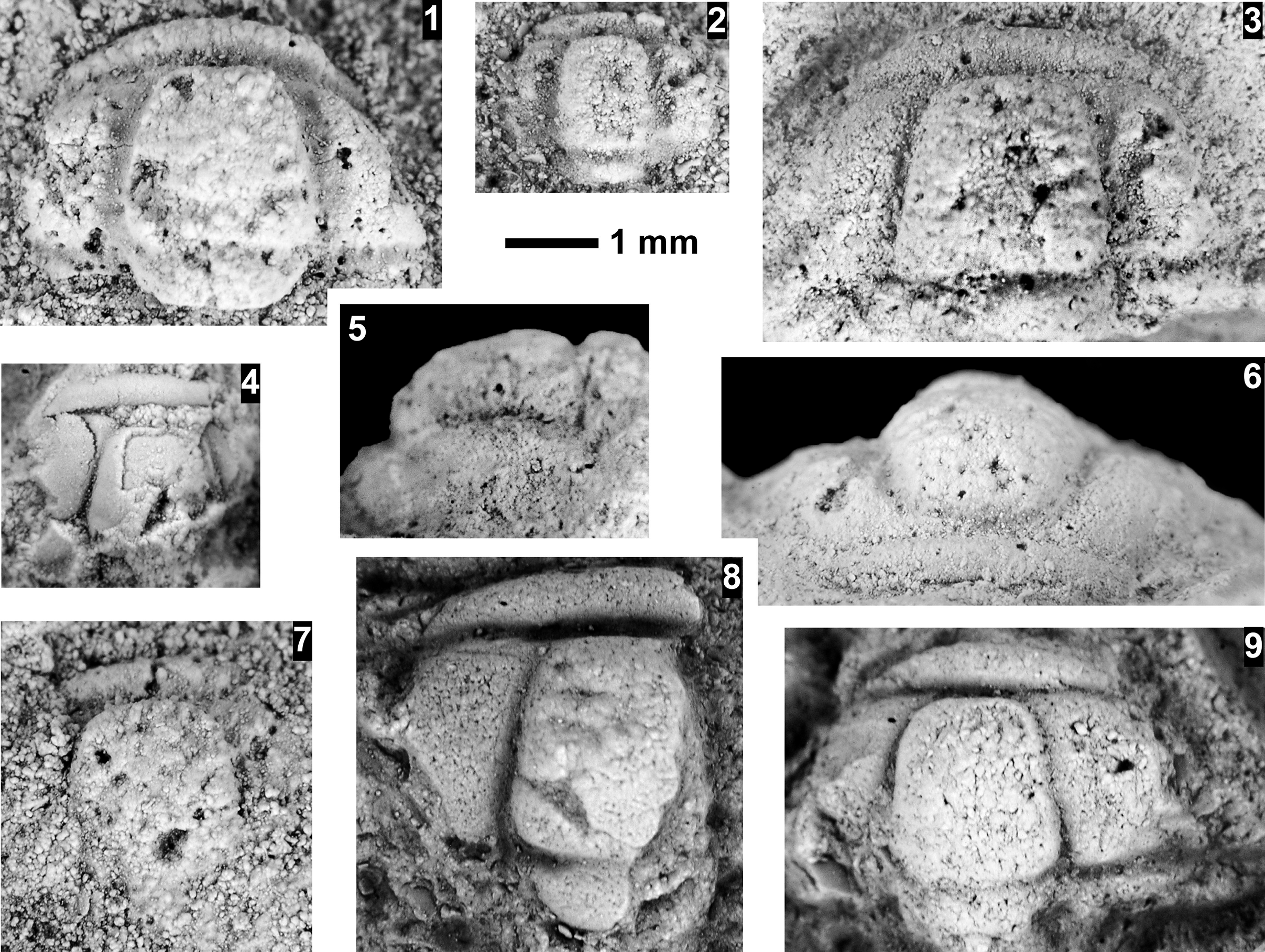
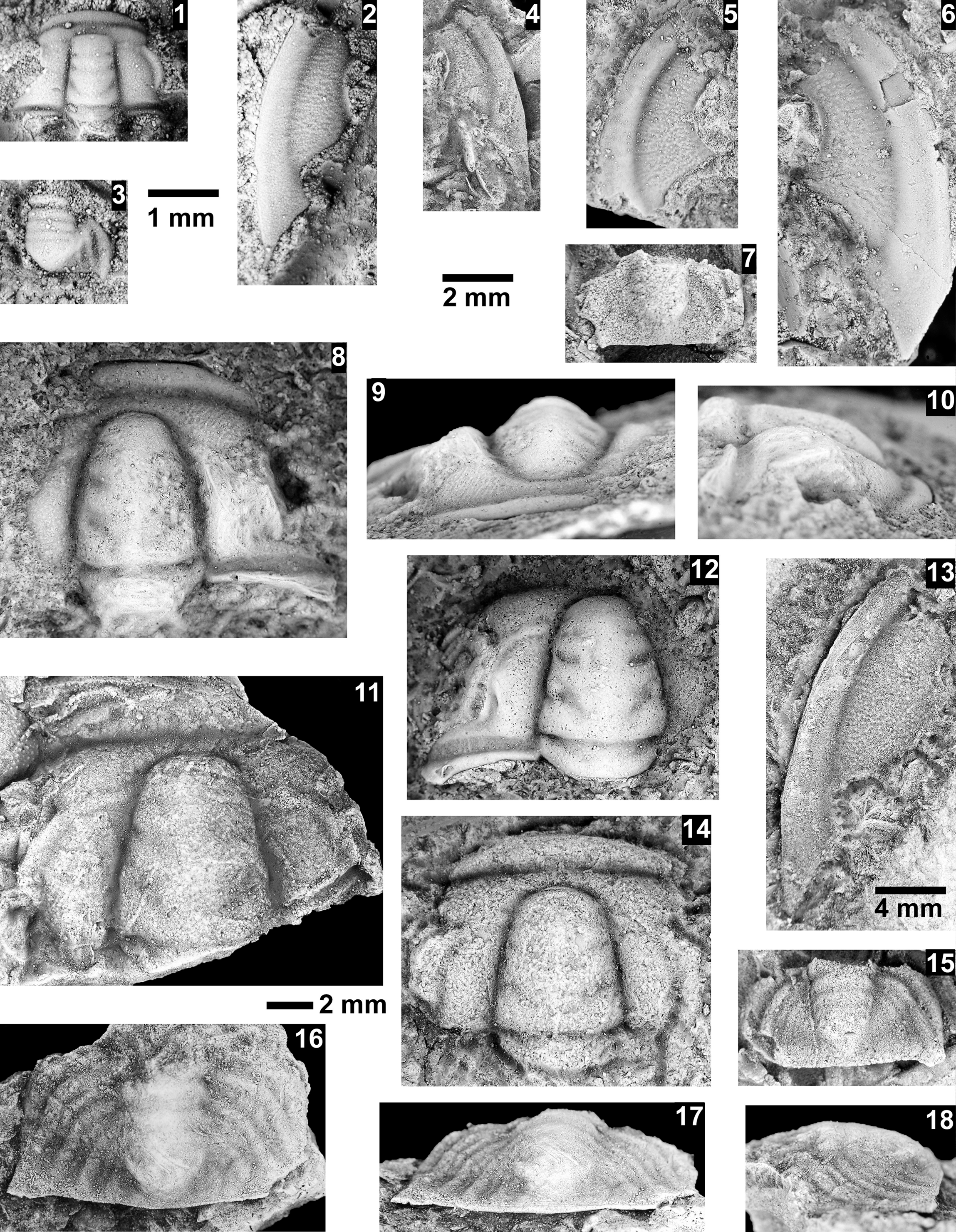
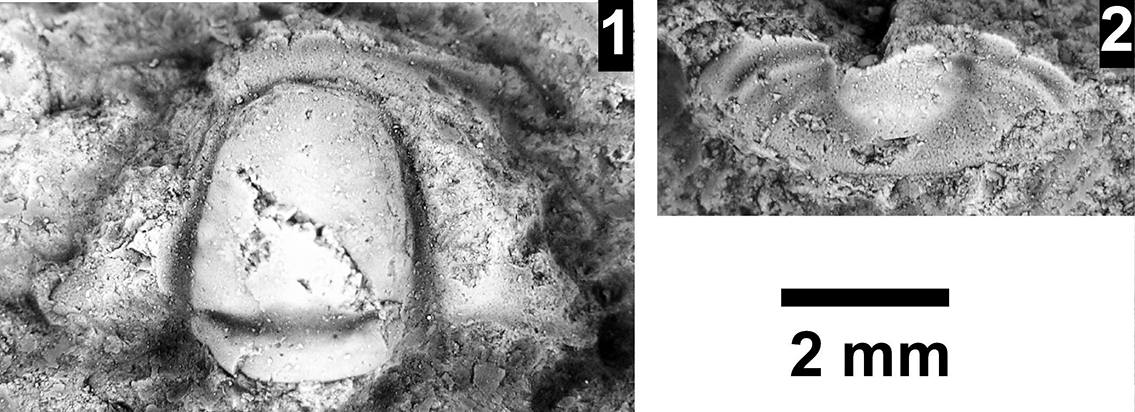
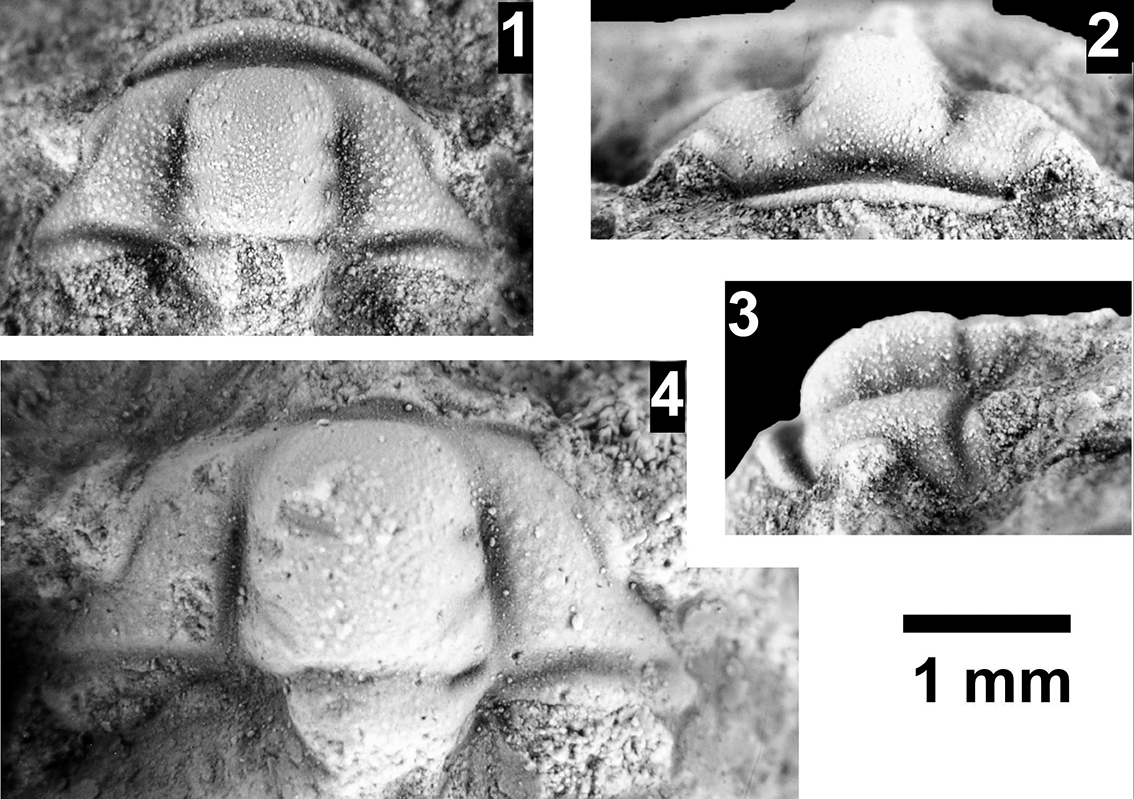
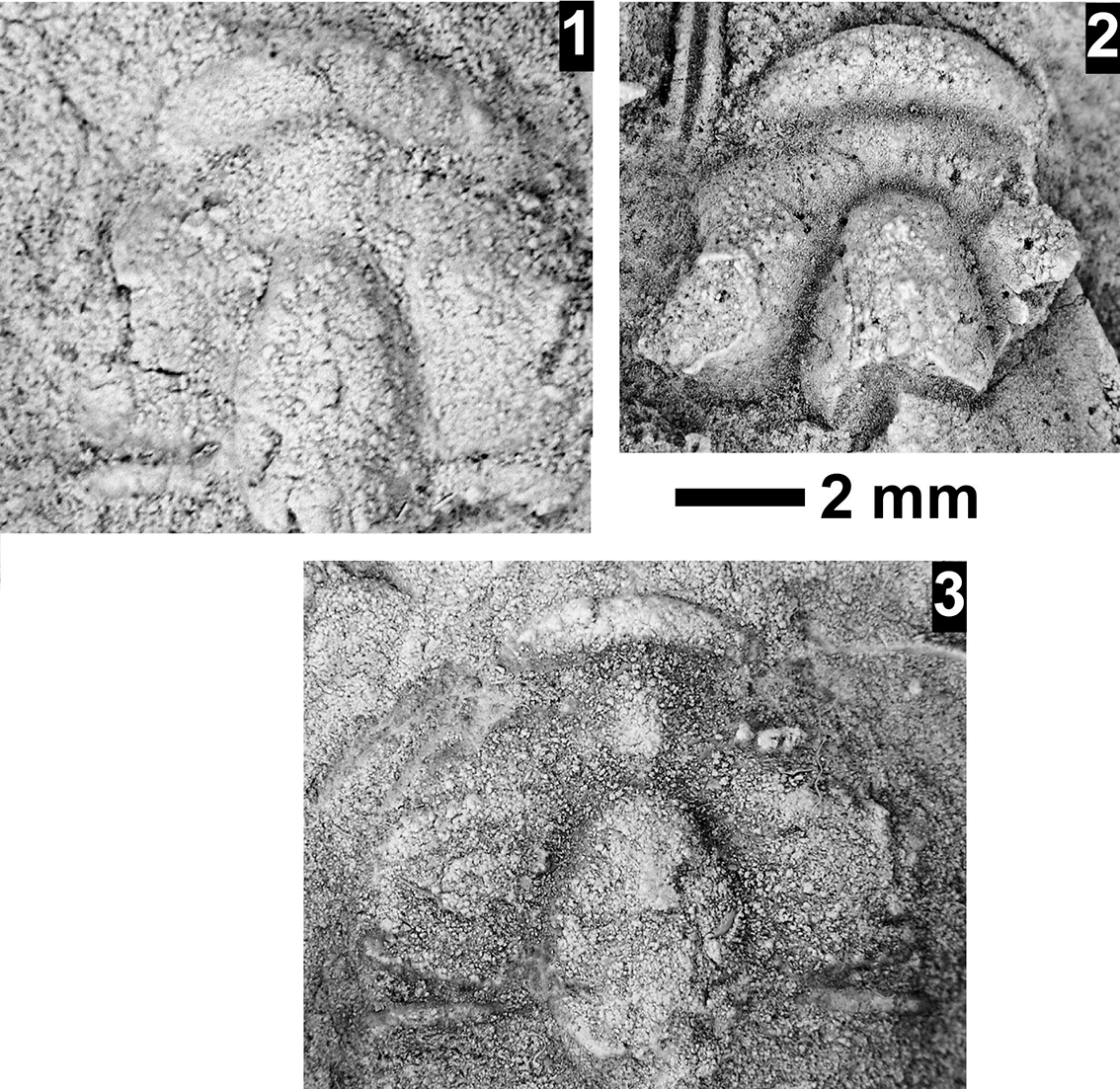
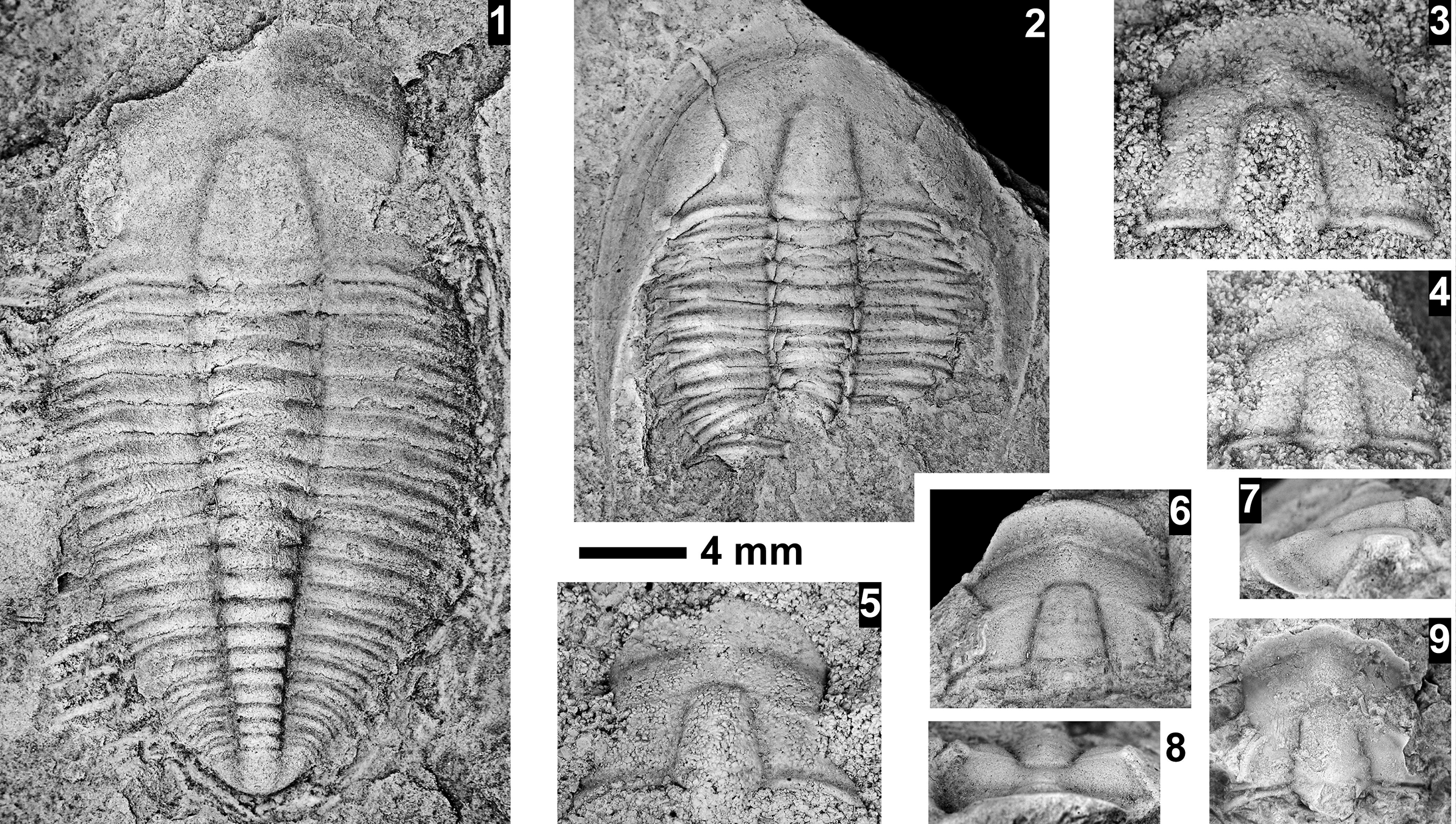
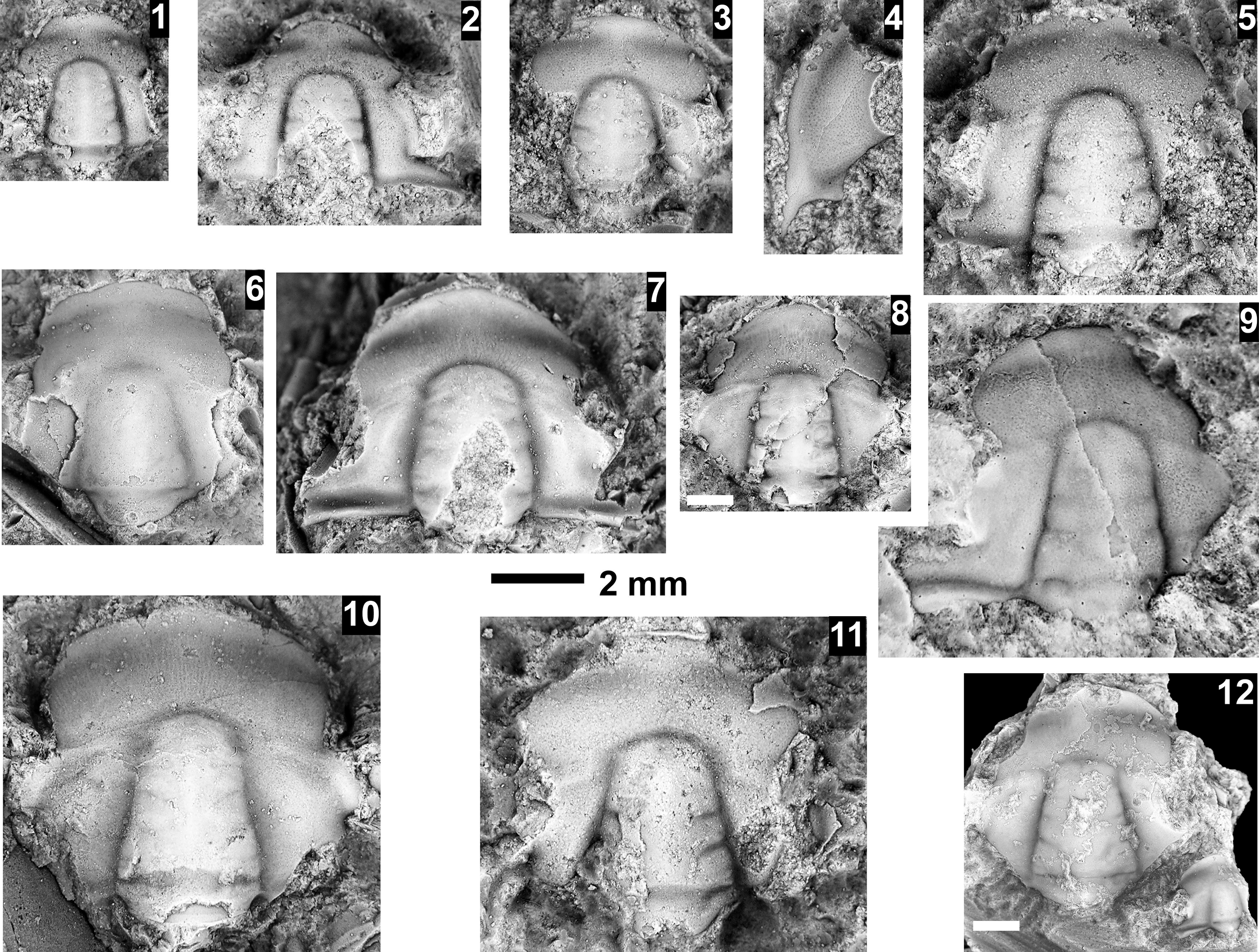
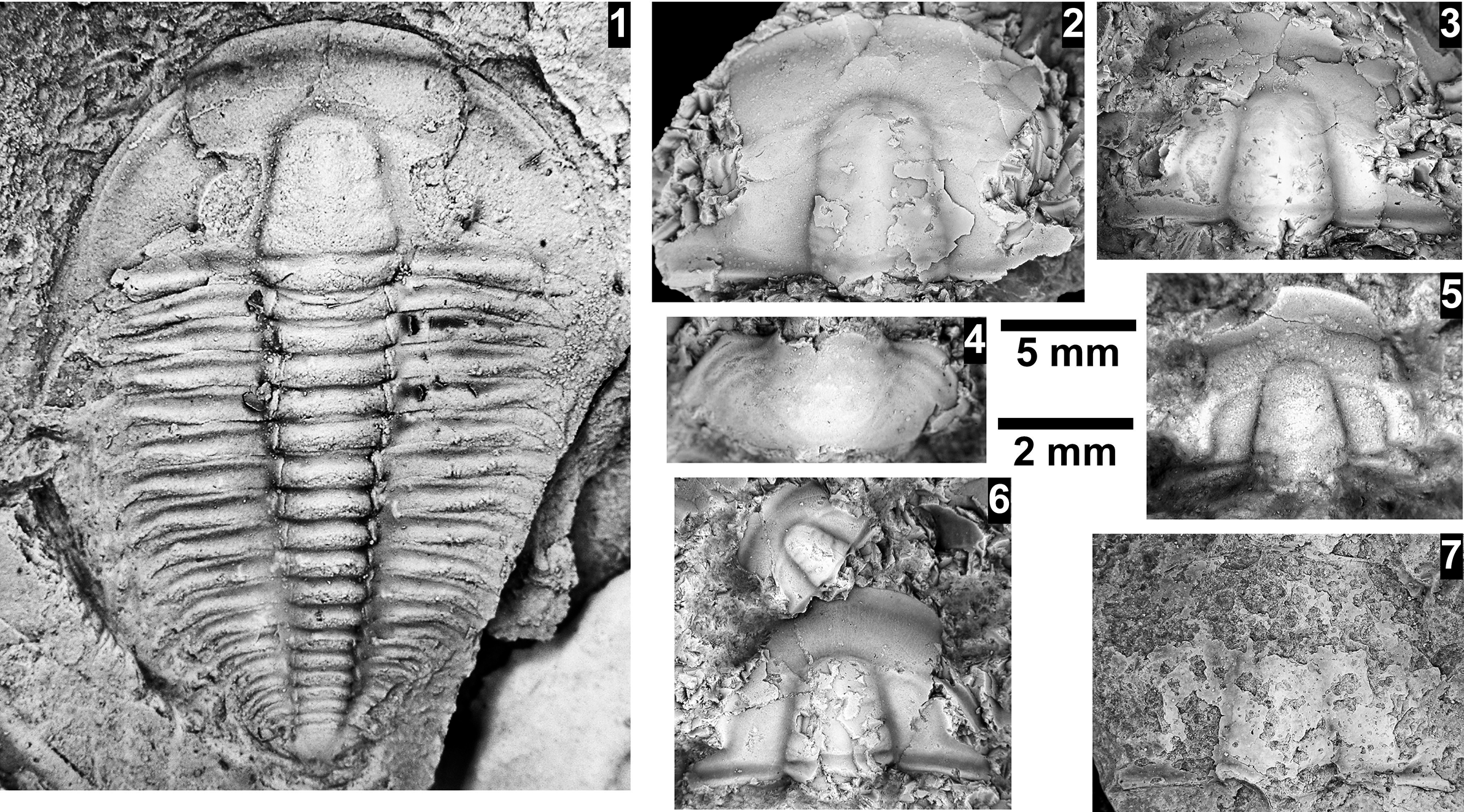

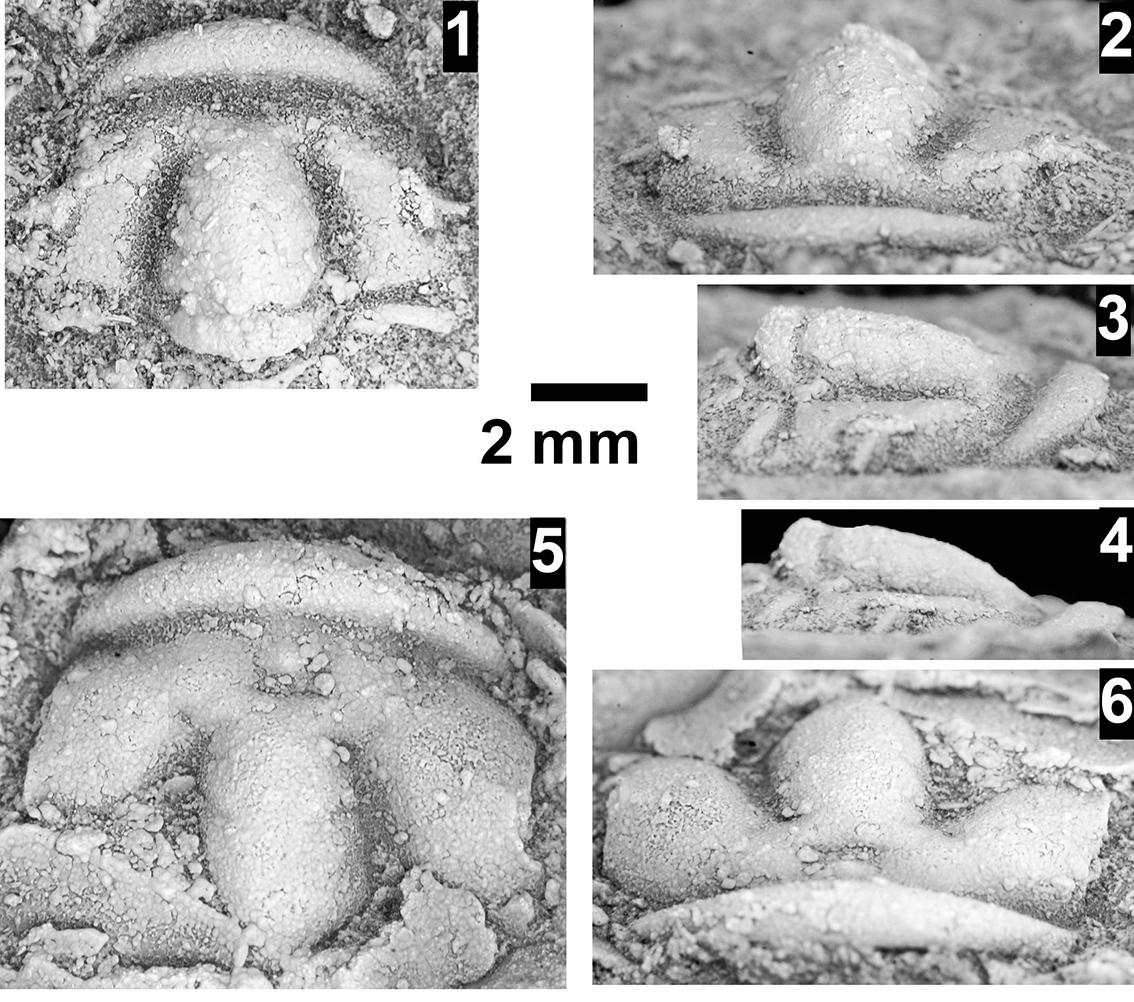
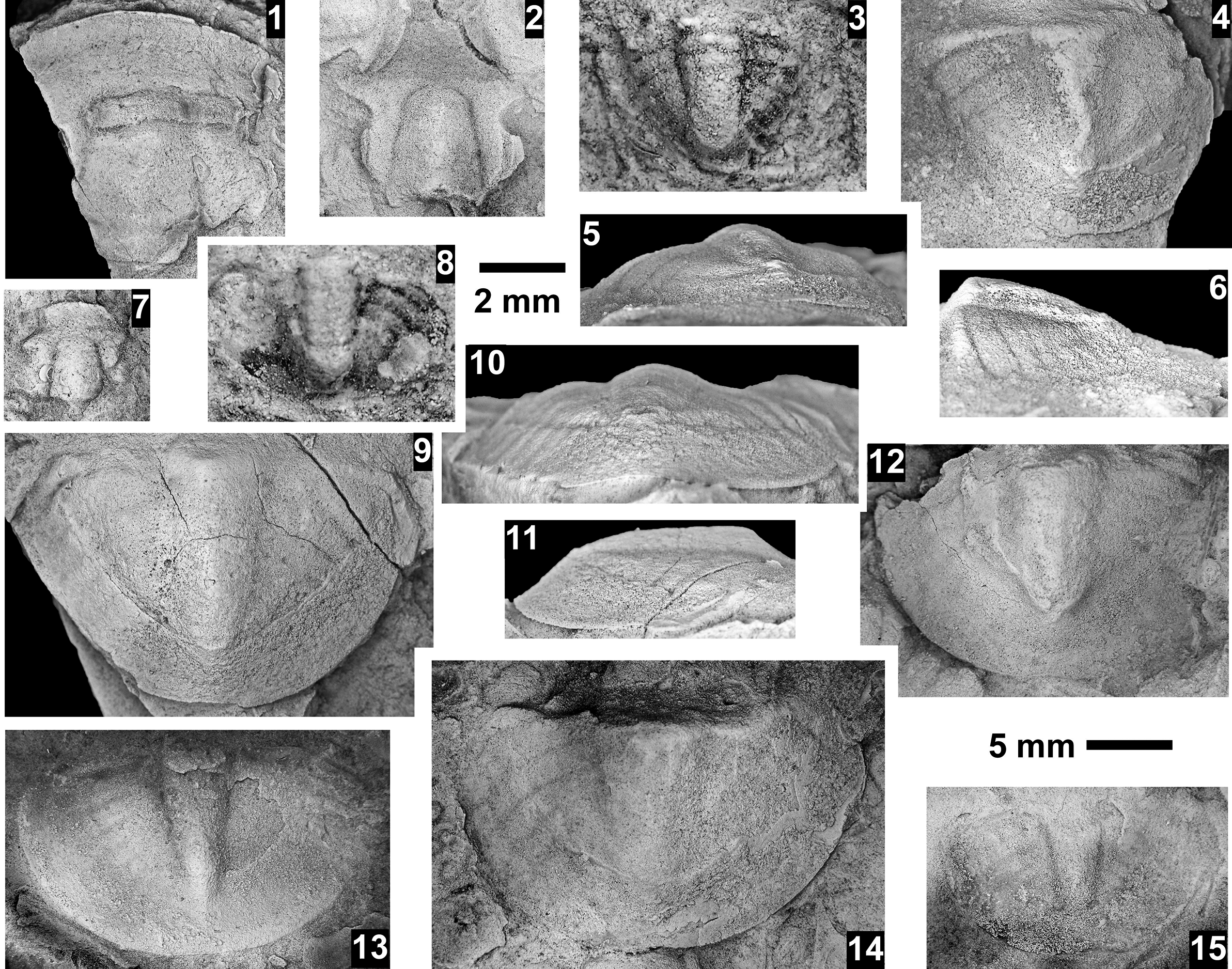
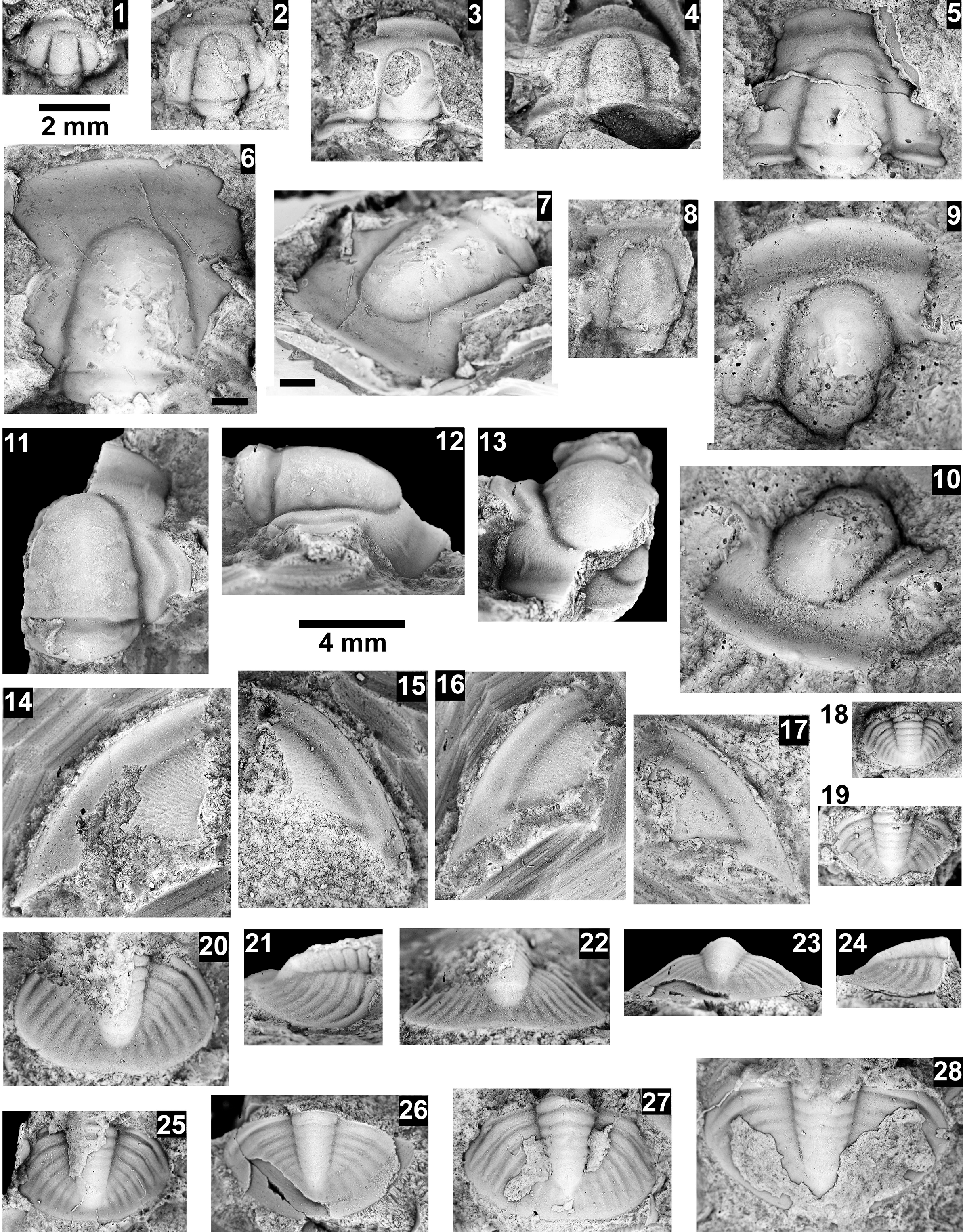
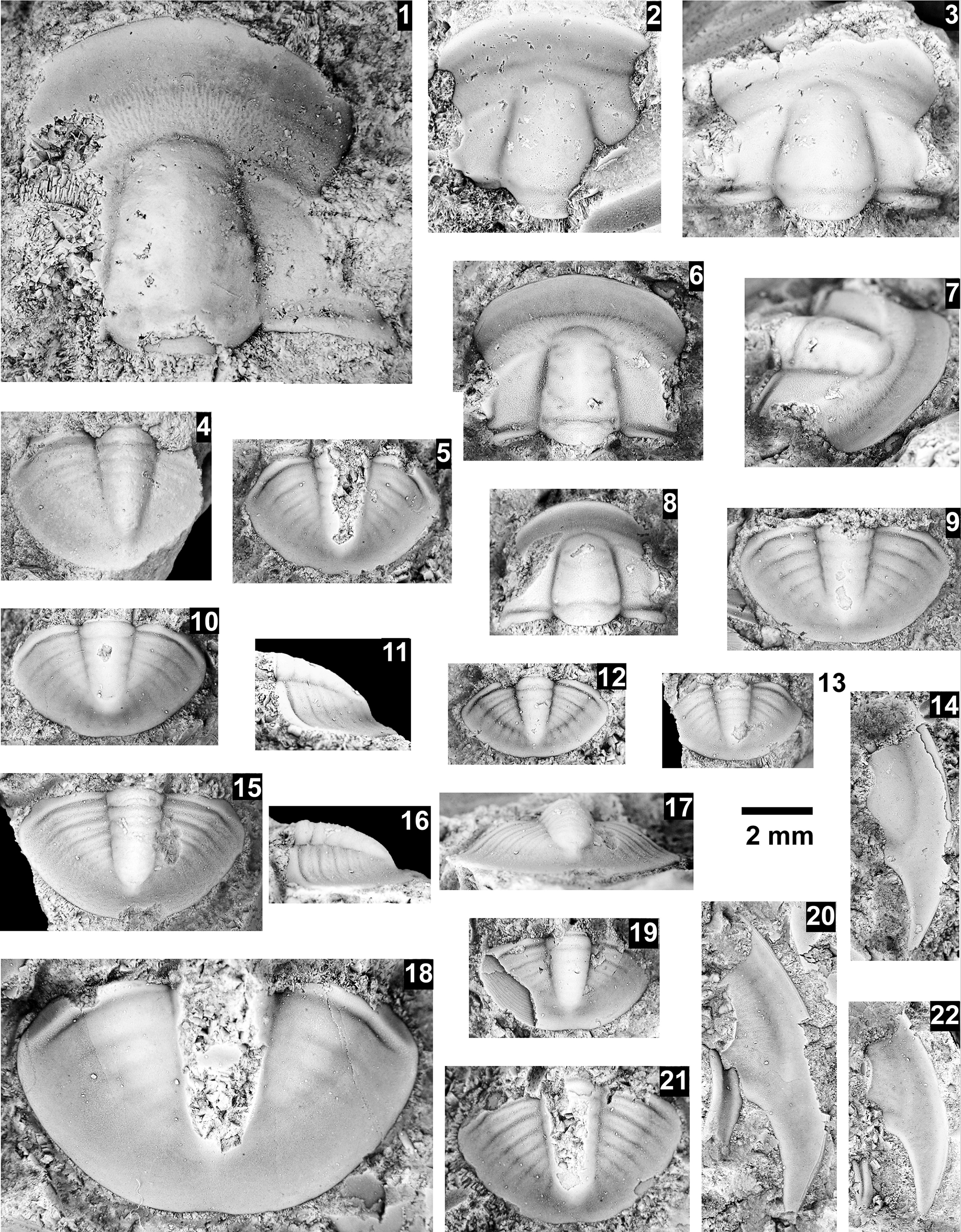
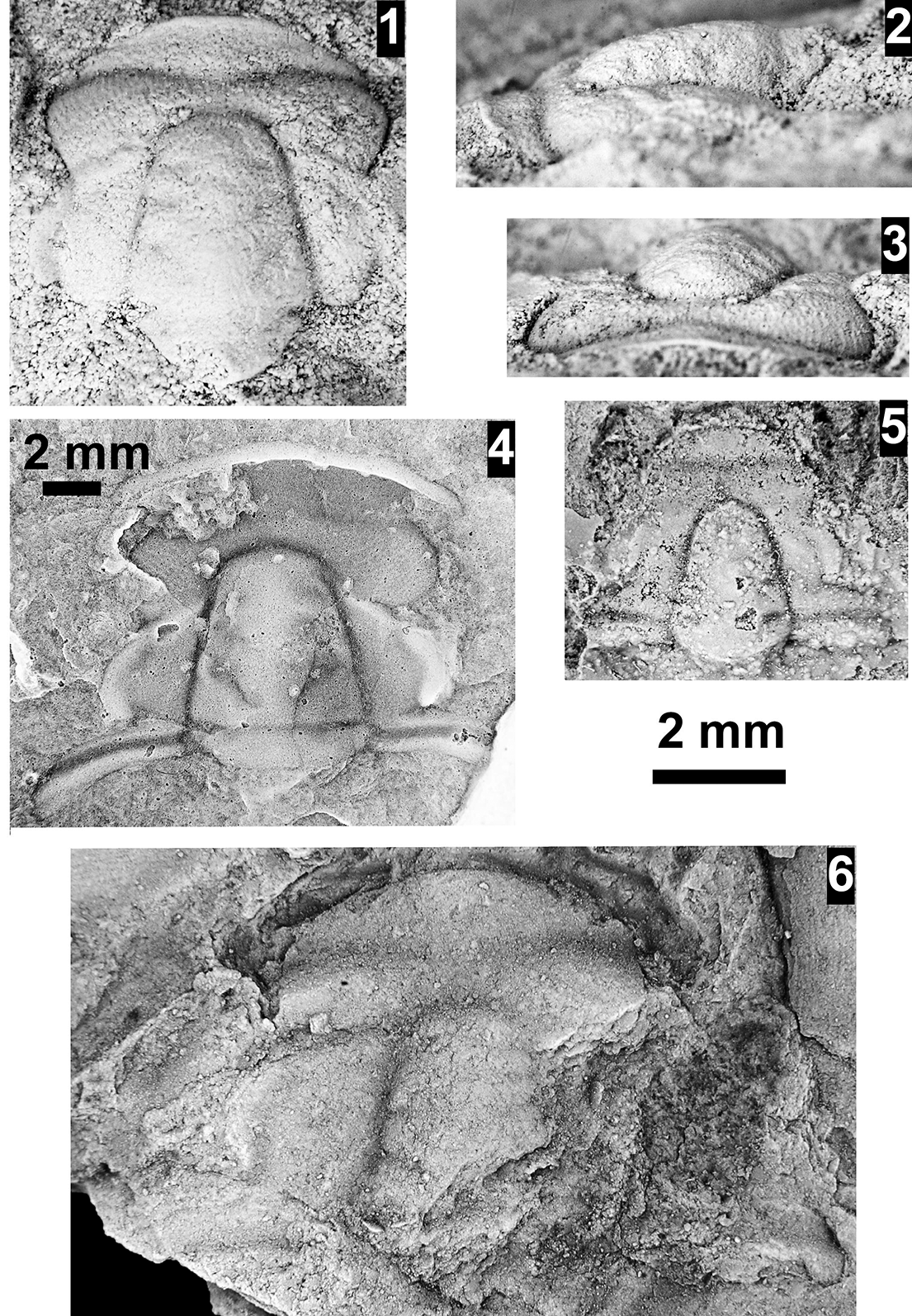

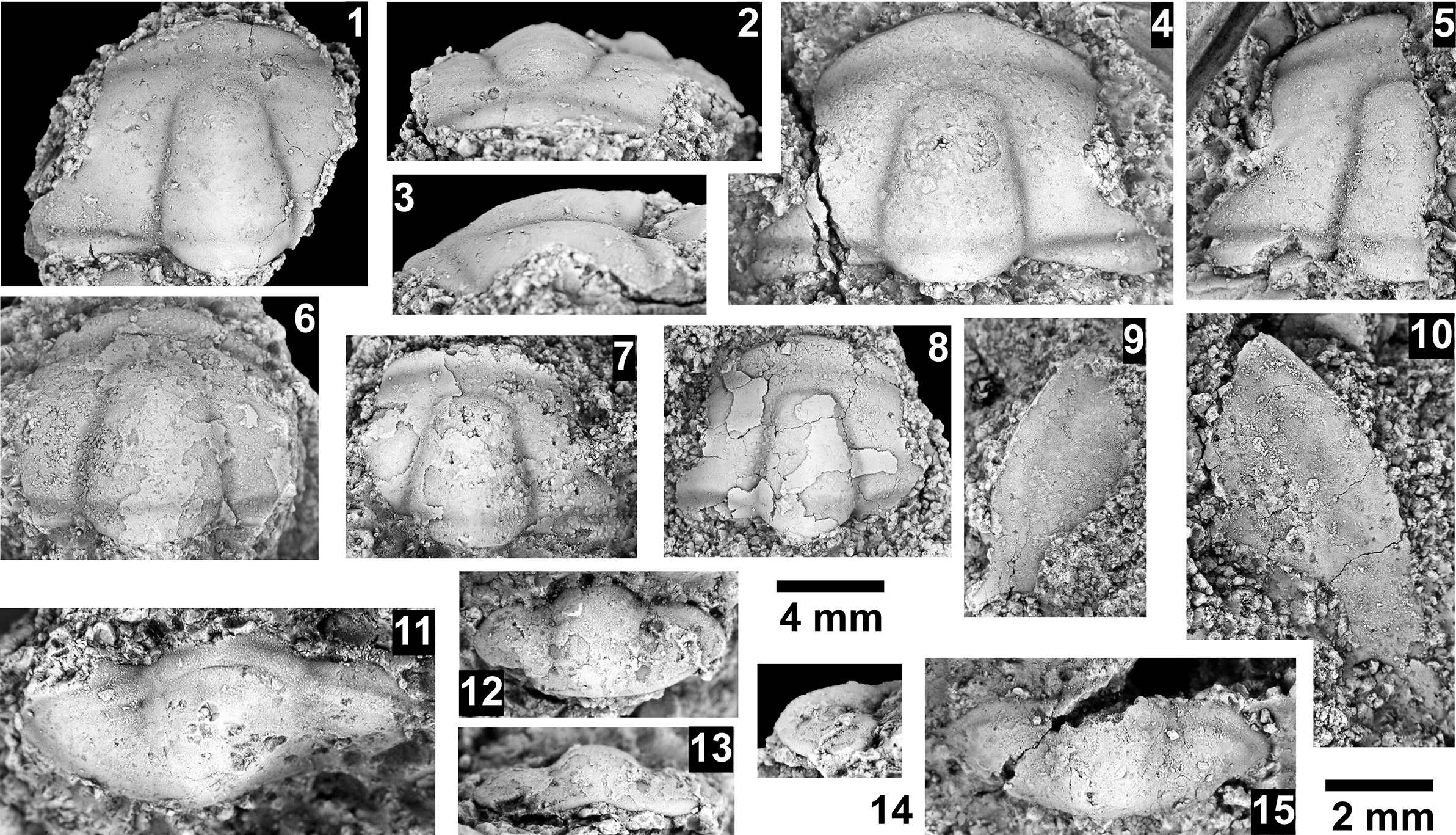

 Open access
Open access