



The gut–liver axis represents an integrated network of metabolic, immune and microbial communication essential for systemic homoeostasis. Through the portal vein, metabolites, cytokines and microbial products from the intestine reach the liver, directly modulating inflammatory, oxidative and metabolic processes. The reduction of intestinal barrier integrity leads to the translocation of lipopolysaccharides and other bacterial components, activating toll-like receptors 4 in hepatocytes and Kupffer cells. This triggers NF-κB activation and the release of pro-inflammatory mediators that impair hepatic metabolism and promote steatosis and fibrogenesis(Reference Zhu, Zong and Xiao1–Reference An, Wirth and Koch5).
The administration of dextran sulfate sodium (DSS) is widely used as an experimental approach to induce intestinal inflammation. DSS causes direct epithelial injury, reduces the expression of tight junction proteins such as claudins and zonula occludens (ZO-1/ZO-2) and disrupts the mucus layer by decreasing mucin-2 (MUC2) expression and goblet cell numbers(Reference Klitgaard, Kristensen and Venkatasubramanian6,Reference Chang, Wang and Liu7) . These alterations promote increased intestinal permeability and endotoxin translocation, amplifying systemic inflammation. NF-κB activation plays a central role in this process by regulating the expression of pro-inflammatory cytokines and contributing to redox and metabolic imbalance(Reference Liu, Huo and Gao8,Reference Kaur and Kushwah9) .
In the liver, DSS-induced intestinal inflammation promotes an intense redox imbalance, characterised by increased production of reactive oxygen species and activation of the NF-κB pathway(Reference Zhan, Peng and Wang10,Reference Kim, Kim and Kwon11) . Sustained activation of this transcription factor overloads the hepatic antioxidant system, stimulating a compensatory response with increased expression of antioxidant enzymes such as catalase (CAT), superoxide dismutase (SOD) and glutathione S-transferase, to neutralise excessive oxidative stress. However, this adaptive response is generally insufficient to restore redox homoeostasis(Reference Elsayed Azab, Adwas Almokhtar and Ibrahim Elsayed12–Reference Djordjevic, Djordjevic and Adzic14). Furthermore, NF-κB activation is associated with the induction of inducible nitric oxide synthase (iNOS), an enzyme that catalyses the production of nitric oxide under inflammatory conditions. The excessive production of nitric oxide can react with superoxide radicals to form peroxynitrite (ONOO⁻), a potent oxidising agent that damages lipids, proteins and DNA, exacerbating hepatocellular damage and contributing to metabolic dysfunction and fatty liver disease(Reference Lashgari, Khayatan and Roudsari15,Reference Cinelli, Do and Miley16) .
In this context, camu-camu (Myrciaria dubia), an Amazonian fruit rich in vitamin C, anthocyanins and phenolic compounds, has emerged as a valuable source of bioactive compounds with antioxidant, anti-inflammatory and modulatory properties(Reference Serrano, Ros and Nieto17,Reference García-Chacón, Marín-Loaiza and Osorio18) . In vivo studies demonstrated that camu-camu polyphenol extracts exert beneficial effects on the intestine by preserving intestinal morphology, stimulating MUC2 production and tight junction protein expression and strengthening the mucosal barrier(Reference Abot, Brochot and Pomié19,Reference Anhê, Nachbar and Varin20) Its bioactive compounds, such as polyphenols and anthocyanins, reduce NF-κB activation and pro-inflammatory cytokines, contributing to protection against intestinal inflammation and oxidative stress(Reference Anhê, Nachbar and Varin20–Reference Kaczmarczyk, Dziewiecka and Pasek22). Additionally, camu-camu polyphenols are metabolised by the gut microbiota into bioactive phenolic compounds (such as gallic and ellagic acids), which may act systemically to modulate immune responses and hepatic signalling, reducing NF-κB activation. These mechanisms contribute to reducing oxidative stress and preserving hepatic metabolic function(Reference Pulido-Mateos, Lessard-Lord and Desjardins23–Reference Kuntz, Kunz and Herrmann25).
The use of the in ovo paradigm (Gallus gallus) has become established as a valuable translational tool for studying the early effects of nutrients, extracts and pro-/antagonistic agents on intestinal development, epithelial morphology and microbial composition(Reference Hou and Tako26,Reference Ayalew, Wang and Wu27) . It also allows for the assessment of initial systemic effects following exposure during the embryonic period. Recent adaptations of this model have enabled the administration of inflammatory agents (such as DSS) and food extracts to investigate intestinal and peripheral outcomes post-hatch, providing a controlled system to study the gut–liver axis during early developmental stages(Reference Hou and Tako26,Reference Huang, Dugan and Tako28) .
Based on this background, the present study aimed to investigate the effects of spray-dried camu-camu phenolic extract (20 %) on the modulation of the gut–liver axis in Gallus gallus embryos post-DSS administration, focusing on morphological, inflammatory and antioxidant biomarkers. The findings of this study are expected to expand the understanding of the molecular and metabolic mechanisms by which camu-camu phenolic compounds modulate the gut–liver axis, supporting its potential as a nutraceutical agent in the prevention of hepato-intestinal disorders mediated by inflammation and oxidative stress.
Material and methods
Raw material and chemical characterisation
Camu-camu, being a typical fruit of the Amazon region, was obtained in powdered form from Cerealista Express, Brazil. The product used was camu-camu spray-dried extract 20 %, prepared from the fruit pulp (Myrciaria dubia H.B.K., Myrtaceae). Extraction was performed using water as the solvent, and the extract was dried using a spray dryer. All samples were subjected to the same cultivation conditions and were purchased in quantities sufficient for the study. The following analyses were performed on the acquired product: total phenolic content by Singleton et al. (1999)(Reference Singleton, Orthofer and Lamuela-Raventós29), monomeric anthocyanins by Giusti & Wrolstad (2003)(Reference Giusti and Wrolstad30) and antioxidant capacity using the 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging assay by Bloor (2001)(Reference Bloor31), the Trolox Equivalent Antioxidant Capacity (TEAC) assay by Pavan et al. (2014)(Reference Pavan, Sancho and Pastore32) and the Ferric Reducing Antioxidant Power (FRAP) assay by Oyaizu (1986)(Reference Oyaizu33).
Additionally, individual phenolic compounds were quantified by HPLC. Briefly, samples (20 mg) were diluted in 1 ml of ultrapure water and analysed using an LC-20A Prominence system (Shimadzu) equipped with a photodiode array detector (SPD-20A) at 270 and 320 nm and a Kinetex C18 column (250 × 4·6 mm). Elution was performed using a gradient of 0·1 % formic acid in water (A) and methanol (B) at a flow rate of 1·0 ml/min(Reference Ho, Hogg and Silva34,Reference Öztürk, Tunçel and Tunçel35) .
Anthocyanins, 3-deoxyanthocyanidins and flavones were also determined by HPLC after extraction with acidified methanol (1 % HCl). Analyses were carried out using a photodiode array detector at 520 nm (anthocyanins), 480 nm (3-deoxyanthocyanidins) and 340 nm (flavones) with reverse-phase C18 columns and gradient elution systems composed of acidified water and acetonitrile. Chromatographic conditions followed established protocols, with flow rates of 1·0 ml/min, injection volumes ranging from 20 to 50 µL and total run times of 45 min(Reference Cardoso, Montini and Pinheiro36,Reference da Costa Guerra, Maciel and de Abreu37) .
All analyses were conducted at the Universidade Federal de Viçosa, Minas Gerais, Brazil.
Biological assay
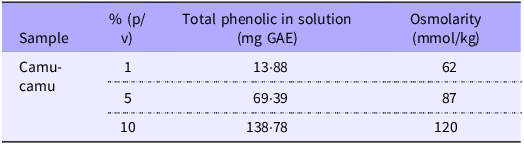
Prior to initiating the experiment, the camu-camu extract was diluted in ultrapure water (18 MΩ H2O) at three concentrations (1 %, 5 % and 10 %) to determine the appropriate dose that would maintain an osmolarity below 320 mOsm, ensuring that the chicken embryos would not experience dehydration upon solution injection(Reference Dias, Kolba and Hart38). The chosen dose was 10 %, which presented an osmolarity of 120 mOsm. In addition to osmolarity, the concentration of phenolic compounds in each solution was also considered for dose selection.
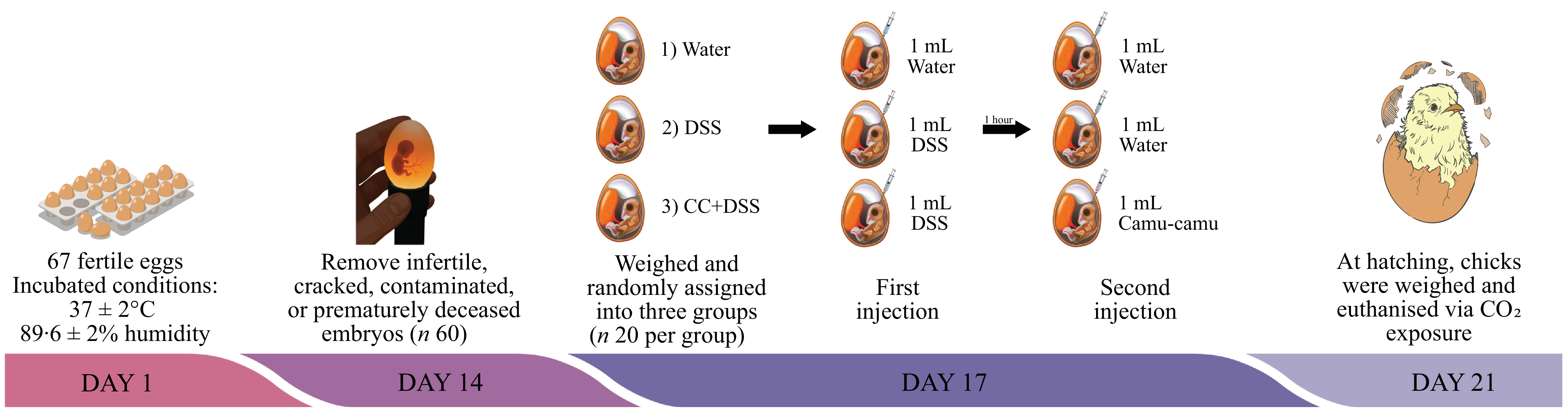
A total of sixty-seven fertile eggs were obtained from a commercial hatchery (Cobb) and incubated under controlled conditions (37 ± 2°C, 89·6 ± 2 % humidity) at the Cornell University Department of Animal Science Poultry Farm. On embryonic day 14, eggs were candled to remove infertile, cracked, contaminated or prematurely deceased embryos, leaving sixty viable eggs for the study. On day 17 of incubation, the eggs were weighed and randomly assigned into three groups (n 20 per group) to ensure similar weight distribution across treatments: (1) water (18 MΩ H2O); (2) DSS (1·5 %) and (3) camu-camu extract (10 %) + DSS (1·5 %) (Figure 1).
Experimental design. DSS, dextran sulfate sodium.

The injection site was identified using a specialised light, and the eggs were disinfected with 70 % ethanol. Eggs in group 1 received 1 ml of water, while groups 2 and 3 were injected with 1 ml of DSS using a 19 mm needle inserted vertically into the amniotic sac. One hour later, a second injection was performed: groups 1 and 2 received 1 ml of water, and group 3 was injected with 1 ml of the camu-camu extract. After each injection, eggs were disinfected with 70 % ethanol and sealed with cellophane tape. Following the second injection, eggs were placed in incubation baskets to evenly distribute treatments across incubator positions. At hatching (day 21), chicks were weighed and euthanised via CO₂ exposure. Liver and duodenum tissues were collected for subsequent analyses.
Analysis
Real-Time PCR
For analysis of liver and intestinal (duodenum) gene expression, total RNA was isolated using TRIzol Reagent (QIAzol Lysis Reagent, Qiagen®). Complementary DNA (cDNA) was subsequently synthesised with the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems®) on a C1000 Touch Thermal Cycler (Bio-Rad Laboratories®)(Reference Livak and Schmittgen39).
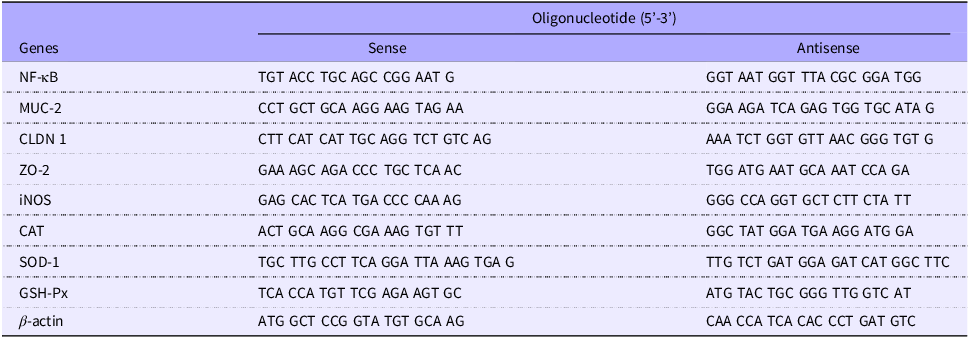
Gene expression levels were quantified by RT-qPCR using PerfeCTa® SYBR Green Fastmix® (Quantabio) on the CFX96 Touch Real-Time PCR Detection System (Bio-Rad Laboratories®). Specific primers (sense and antisense) were designed to amplify NF-κB, MUC-2, claudin and ZO-2 in intestine and NF-κB, iNOS, CAT, SOD-1 and GSH-Px in liver. Expression values were normalised to the endogenous β-actin reference gene (Table 1)(Reference Livak and Schmittgen39).
Sequence of primers used in the RT-qPCR analysis

RT-qPCR, real-time quantitative polymerase chain reaction; MUC-2, mucin 2; CLDN 1, claudin 1; ZO-2, zona occludens-2; iNOS, inducible nitric oxide synthase; CAT, catalase; SOD-1, superoxide dismutase 1; GSH-Px, glutathione peroxidase.
Histomorphometry
Liver and duodenal sections were removed immediately after euthanasia and stored in 10 % neutral buffered formalin for 72 h before sectioning. Formalin-fixed liver and duodenal segments were transversely sectioned to obtain sections suitable for morphological evaluation. Tissue sections were subsequently dehydrated, cleared and embedded in paraffin. Serial 5-µm-thick sections were prepared, mounted on glass slides, deparaffinised in xylene and rehydrated through a graded ethanol series. Duodenal sections were stained with Alcian Blue-Periodic Acid-Schiff (AB-PAS), and liver sections were stained with hematoxylin and eosin(Reference Suvarna, Layton and Bancroft40). Sections were photographed using an Olympus BX43 portable microscope equipped with a 2·8-megapixel DP22 digital camera (Olympus Life Science Solutions) with a 10× objective.
In the duodenum, morphometric measurements were collected of the villi (height, width and surface area), crypts (depth and width), number and area of goblet cells and longitudinal and circular thickness of the muscle layer. The villi surface area was calculated from the villi height and the average width of three villi, modelling each villus as a cylinder(Reference Huang, Smieska and Tako41). In the liver, the areas of cytoplasm and fat globules, inflammation and hepatic nuclei were analysed by tissue point counting using a 266-point grid, totalling 1064 points per animal(Reference de Paula Dias Moreira, Enes and de São José42). All measurements were performed using Image-Pro-Plus® software, version 4.5.0.29 (Media Cybernetics).
Statistical analysis
Statistical analysis and graph construction were performed using GraphPad Prism® software, version 10.1.2 (GraphPad Prism Inc.). Out of the twenty eggs allocated per group, eight successfully hatched in each group. Following outlier identification and exclusion, a final sample size of five animals per group was considered for each analysis performed. Data normality was assessed using the Shapiro–Wilk test. Intergroup differences (water, DSS and CC + DSS) were submitted to ANOVA, followed by the Tukey’s mean test. A P-value < 0·05 was adopted. The results are expressed as mean and standard deviation.
Results
All tested camu-camu solutions were within the acceptable osmolarity range. Notably, the 10 % camu-camu solution exhibited the highest concentration of phenolic compounds among the tested conditions, while maintaining an osmolarity of 120 mOsm. Therefore, the 10 % dose (120 mOsm) was selected as it combined appropriate osmolarity with a greater availability of bioactive compounds (Table 2).
Osmolarity and phenolic compound content of camu-camu solutions at different concentrations

GAE, gallic acid equivalent.
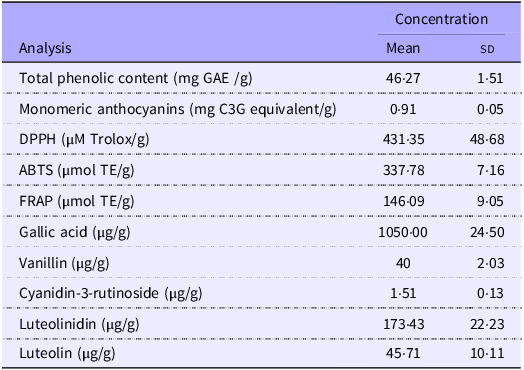
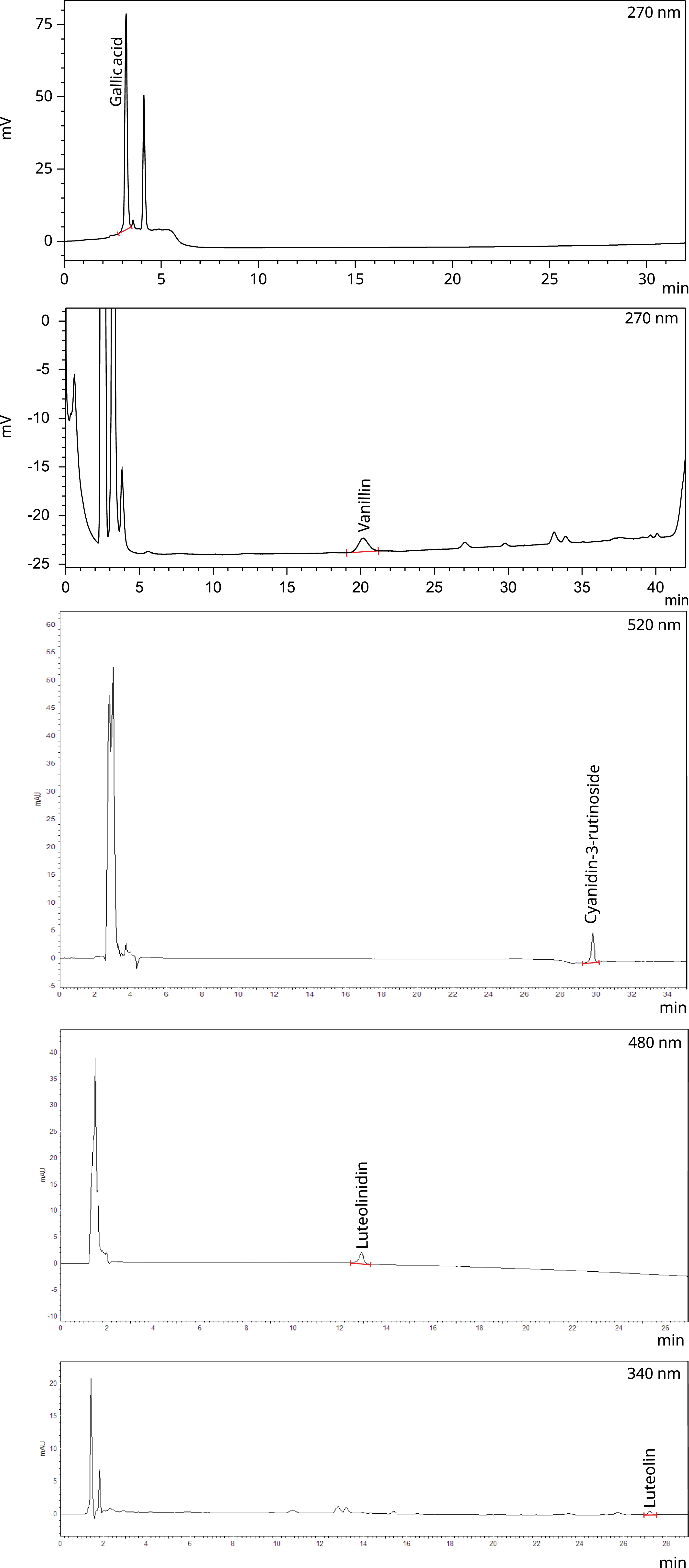
When analysing the concentration of phenolic compounds and antioxidant capacity, camu-camu presented 46·27 mg GAE/g of total phenolic content and 0·91 mg C3G equivalents/g of monomeric anthocyanins. Regarding antioxidant capacity by DPPH, the sample presented 431·35 µM Trolox/g, while the ABTS and FRAP assays yielded 337·78 and 146·09 µmol TE/g, respectively (Table 3). Regarding the presence of phenolic compounds, anthocyanins and flavones, it was possible to identify and quantify the following compounds by HPLC: 1·05 µg/g of gallic acid, 1·50 µg/g of cyanidin-3-rutinoside, 173·43 µg/g of luteolinidin and 45·71 µg/g of luteolin (Table 3, Figure 2).
Phenolic content, anthocyanin concentration and antioxidant capacity of camu-camu extract

GAE, gallic acid equivalent; C3G, cyanidin-3-glucoside; DPPH, 1,1-diphenyl-2-picrylhydrazyl; ABTS, 2,2′-azino-bis-3-ethylbenzthiazoline-6-sulphonic acid; FRAP, ferric reducing antioxidant power; TE, trolox equivalent.
HPLC chromatograms of camu-camu spray-dried extract 20 %. The identified compounds are indicated in each chromatogram based on retention time and comparison with analytical standards.

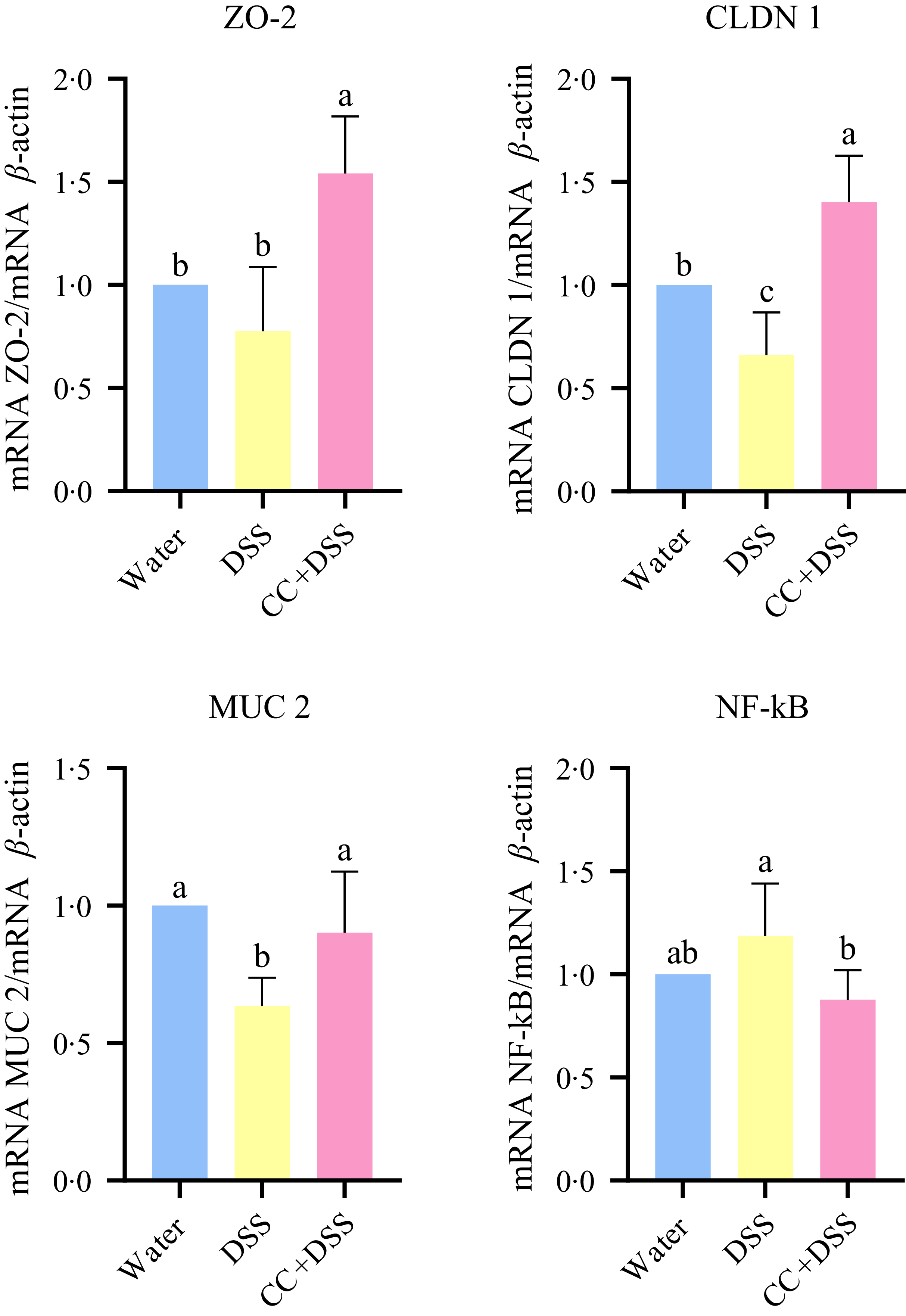
Gene expression in the duodenum of chicks revealed that DSS reduced claudin-1 (CLDN-1) and MUC-2 expressions compared with the healthy control group (water) (P = 0·0450 and P = 0·0040, respectively). The CC + DSS group showed higher expression of ZO-2 and CLDN-1 compared with both control and DSS groups (P = 0·0009 and P = 0·0007, respectively). MUC2 expression was higher in the CC + DSS group than in the DSS group (P = 0·0285), with no difference compared with the control group. NF-κB expression in the CC + DSS group did not differ from the control group but was lower than in the DSS group (P = 0·0337) (Figure 3).
Gene expression of ZO-2, CLDN 1, MUC-2 and NF-κB in duodenum. n 5/group. Different lowercase letters (a–b) indicate significant differences within groups, according to ANOVA, followed by the Tukey’s test, at 5 % probability. Data are expressed as mean and standard deviation. ZO-2, zona occludens-2; CLDN 1, claudin 1; MUC-2, mucin 2; water, 18 MΩ H2O; DSS, dextran sulfate sodium 1·5 %; CC + DSS, camu-camu extract (10 %) + DSS (1·5 %).

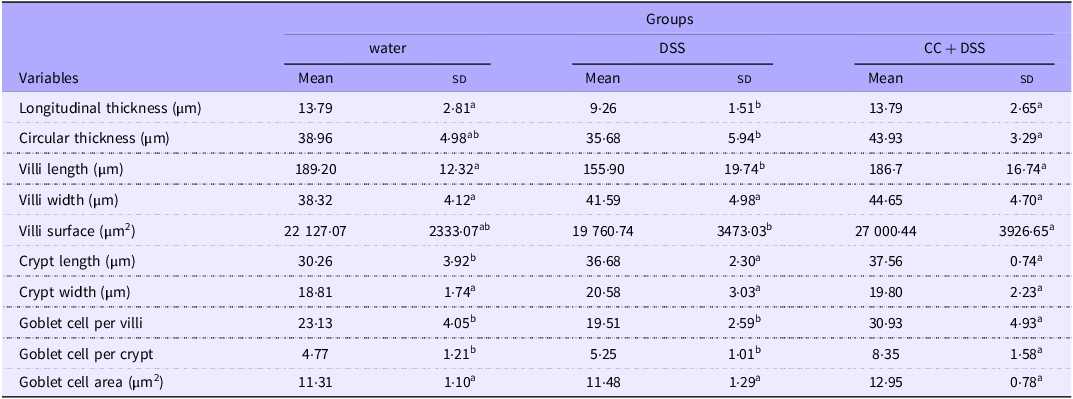
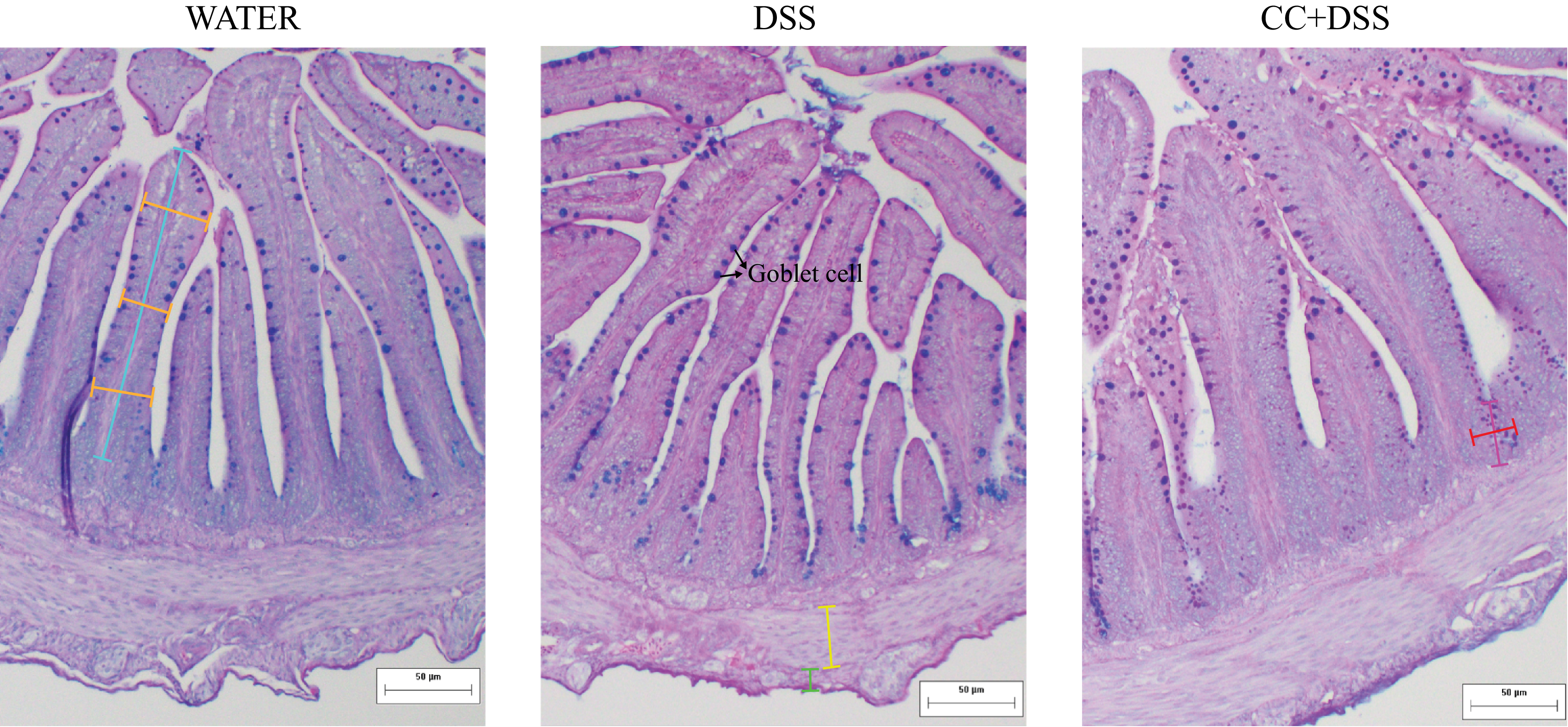
Histological analysis of the duodenum demonstrated that DSS reduced intestinal muscular layer and villus length compared with the healthy control group (P = 0·0281 and P = 0·0201, respectively). The CC + DSS group exhibited greater longitudinal and circular muscle thickness than the DSS group (P = 0·0281 and P = 0·0489, respectively), with no difference compared with the water group. Villus length and surface area were higher in the CC + DSS group than in the DSS group (P = 0·0308, and P = 0·0122, respectively), reaching values comparable to the control group. Villus width did not differ among experimental groups (P = 0·1368) (Table 4, Figure 4).
Duodenal histomorphometry analysis of chicks

Water, 18 MΩ H2O; DSS, dextran sulfate sodium 1·5 %; CC + DSS, camu-camu extract (10 %) + DSS (1·5 %). n 5/group.
Different lowercase letters (a–b) indicate significant differences within groups, according to ANOVA, followed by the Tukey’s test, at 5 % probability. Data are expressed as mean and standard deviation.
Representative photomicrographs of the duodenum stained with Alcian Blue-Periodic Acid-Schiff (AB-PAS). Blue line: villi length; orange line: villi width; green line: longitudinal thickness; yellow line: circular thickness; pink line: crypt length; red line: crypt width. Goblet cell was indicated in the image. Water, 18 MΩ H2O; DSS, dextran sulfate sodium 1·5 %; CC + DSS, camu-camu extract (10 %) + DSS (1·5 %).

Crypt length was higher in DSS and CC + DSS groups than in the control group (P = 0·0018), with no difference between DSS and CC + DSS (P = 0·8628). Crypt width presented no difference among treatments (P = 0·5223). The CC + DSS group had a higher number of goblet cells per villus and per crypt than the other groups (P = 0·0021 and P = 0·0017, respectively), although goblet cell area did not differ among treatments (P = 0·0631) (Table 4, Figure 4).
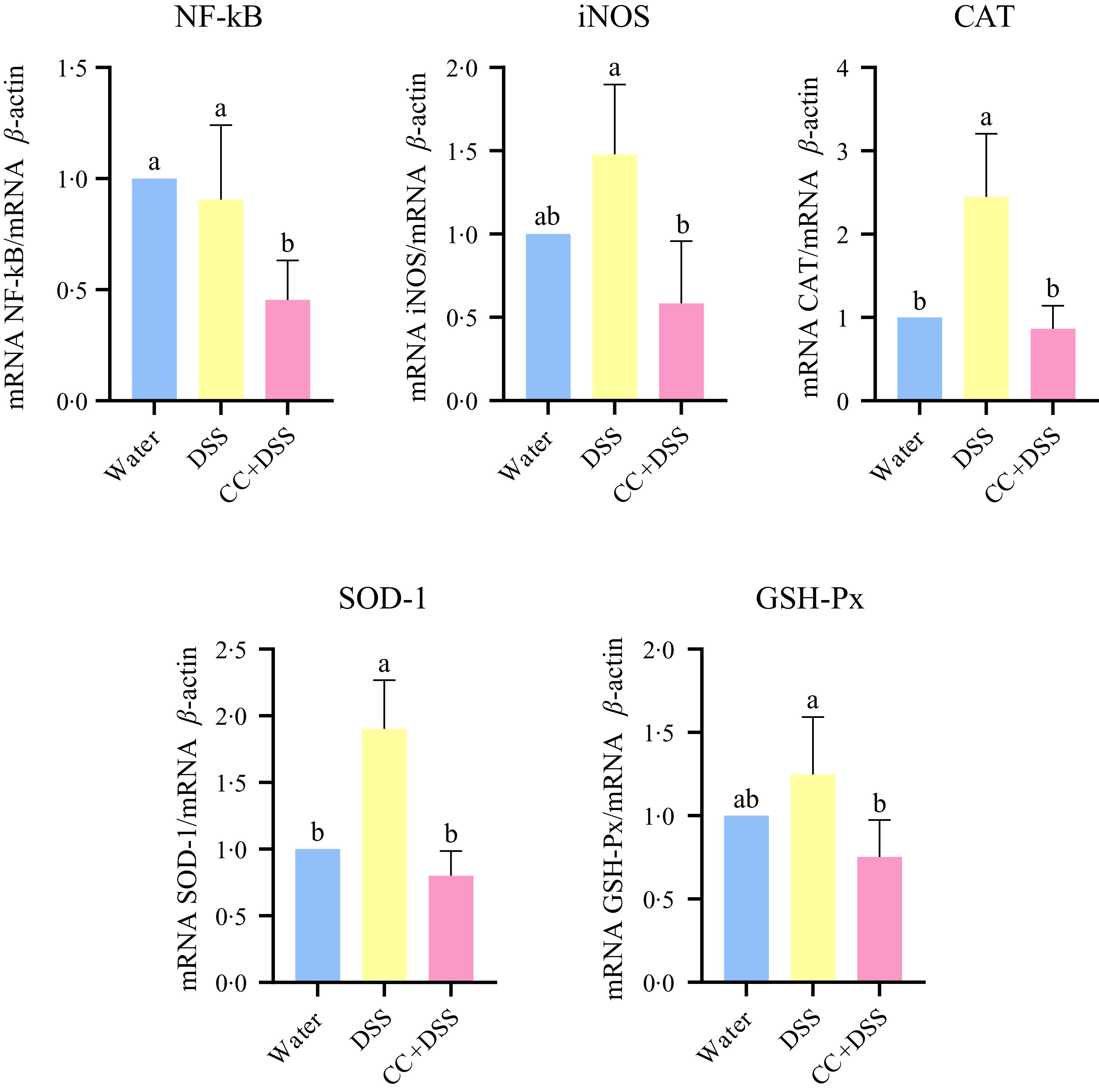
Gene expression in liver revealed that DSS did not affect NF-κB and iNOS expressions (P = 0·7793, and P = 0·1004, respectively) compared with the control group. CC + DSS group exhibited a lower NF-κB expression than both DSS and control groups (P = 0·0074). Expressions of iNOS, CAT and SOD-1 were lower in the CC + DSS group than in the DSS group (P = 0·0035, P = 0·0005 and P < 0·0001, respectively), with no differences compared with the control group. GSH-Px expression was lower in the CC + DSS group than in the DSS group (P = 0·0158), with no differences between control and the other groups (Figure 5).
Gene expression of NF-κB, iNOS, CAT, SOD-1 and GSH-Px in liver. n 5/group. Different lowercase letters (a–b) indicate significant differences within groups, according to ANOVA, followed by the Tukey’s test, at 5 % probability. Data are expressed as mean and standard deviation. iNOS, inducible nitric oxide synthase; CAT, catalase; SOD-1, superoxide dismutase 1; GSH-Px, glutathione peroxidase; water, 18 MΩ H2O; DSS, dextran sulfate sodium 1·5 %; CC + DSS, camu-camu extract (10 %) + DSS (1·5 %).

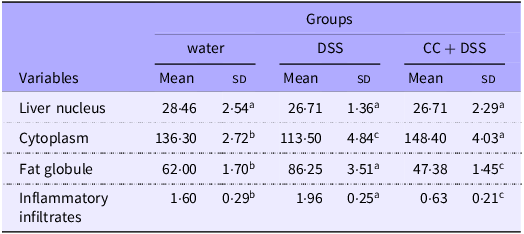
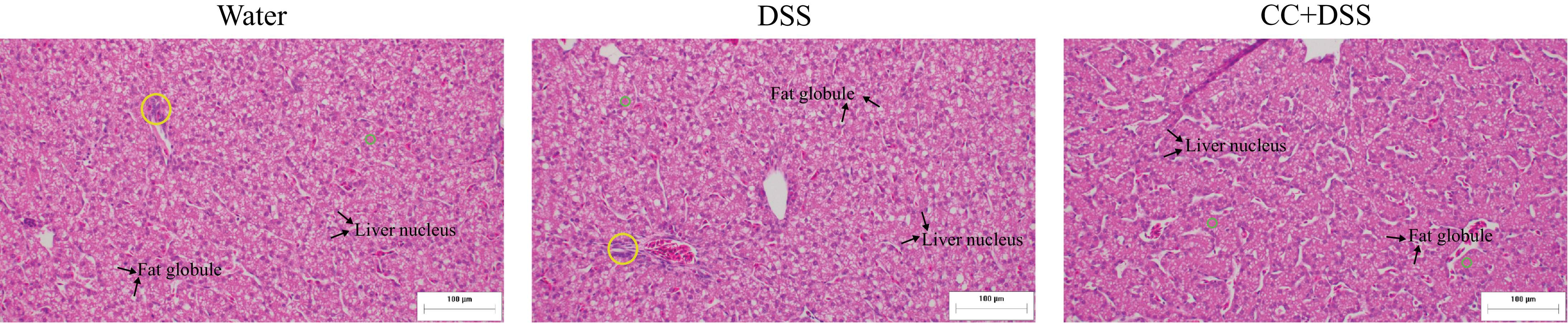
Histological analysis of liver revealed no differences among groups regarding the number of hepatic nucleus (P = 0·2864). DSS reduced cytoplasmic area and increased fat globules and inflammatory infiltrates compared with the control group (P < 0·0001, P < 0·0001 and P = 0·0019, respectively). The CC + DSS group showed higher cytoplasmic area and lower fat accumulation and inflammatory infiltrates than the DSS group (P < 0·0001), indicating improved hepatic morphology (Table 5, Figure 6).
Histomorphometry point-count analysis of liver tissue

Water, 18 MΩ H2O; DSS, dextran sulfate sodium 1·5 %; CC + DSS, camu-camu extract (10 %) + DSS (1·5 %). n 5/group.
Different lowercase letters (a–c) indicate significant differences within groups, according to ANOVA, followed by the Tukey’s test, at 5 % probability. Data are expressed as mean and standard deviation.
Representative photomicrographs of the liver stained with hematoxylin and eosin. Yellow circle: inflammatory infiltrates; green circle: cytoplasm. Fat globules and liver nucleus were indicated in the images. Water, 18 MΩ H2O; DSS, dextran sulfate sodium 1·5 %; CC + DSS, camu-camu extract (10 %) + DSS (1·5 %).

Discussion
The present study demonstrated that with the administration of camu-camu (Myrciaria dubia) extract attenuated DSS-induced alterations in intestinal and hepatic parameters of chicken embryos, highlighting the modulatory potential of this Amazonian fruit on the gut–liver axis. The chemical characterisation (Table 3, Figure 2) of the extract showed the presence of some phenolic compounds and anthocyanins, as well as a high antioxidant capacity, which may be related to the observed changes in the measured parameters. The observed changes in intestinal histomorphology, goblet cell parameters, barrier-related gene expression and hepatic gene expression markers are consistent with reported interactions between phenolic compounds and inflammatory- and oxidative stress-related pathways(Reference Boaru, Fraile-Martinez and De Leon-Oliva43,Reference Li, Peng and Ding44) .
In the duodenum, DSS exposure triggered a reduced number of goblet cells per villus and crypt and decreased expression of tight junction protein (CLDN-1) and mucin. The intestinal epithelium represents the first defensive interface that prevents the translocation of gut bacteria into systemic circulation(Reference Allam-Ndoul, Castonguay-Paradis and Veilleux45). The mucus layer, composed of mucins secreted by goblet cells, separates bacteria from the underlying intestinal epithelial cells, while tight and adherents’ junctions in endothelial cells regulate vascular barrier permeability, allowing nutrients and small molecules to pass into the enteral circulation while restricting the passage of viable bacteria and microbial antigens, such as lipopolysaccharides and toxic agents, such as DSS(Reference Di Tommaso, Gasbarrini and Ponziani46–Reference Hsu and Schnabl48).
In this study, co-administration of camu-camu extract was associated with increased expression of ZO-2, CLDN-1 and MUC2, along with changes in villus architecture and goblet cell numbers. These findings are consistent with alterations in epithelial morphology and mucus-related parameters, consistent with previous reports describing associations between polyphenols and barrier-related and inflammatory pathways(Reference Kong, Yan and Tong49–Reference Milev, Roglev and Kondeva Rogleva52). Similar findings were reported in chicks that received different jabuticaba extracts via intra-amniotic administration, which resulted in increased CLDN-1 expression and reduction in inflammatory markers(Reference Pereira, José and Huang53). Likewise, a study evaluating the effects of ellagic acid in E. coli-infected broilers also demonstrated beneficial effects of this phenolic compound on the expression of tight junction proteins and mucin(Reference Li, Zhao and Teng54). The restoration of these markers reinforces the protective role of camu-camu in maintaining intestinal homoeostasis, even under DSS-induced severe inflammation conditions.
An intact intestinal barrier is essential not only for nutrient absorption but also for retaining micro-organisms within the intestinal lumen, thereby preventing their systemic dissemination and preserving the health of other organs, such as the liver(Reference Plaza-Díaz, Solís-Urra and Rodríguez-Rodríguez55). Chronic inflammatory liver diseases are closely associated with gut dysbiosis, altered intestinal permeability and translocation of lipopolysaccharides to the liver(Reference Wang, Cao and Zhang56). Most venous blood from the intestine drains into the portal vein, which is rich in enteral metabolites(Reference Delzenne, Knudsen and Beaumont57). The liver is the first major organ to encounter these metabolites, which play key roles in hepatic immune and metabolic functions(Reference Hsu and Schnabl48,Reference Yang, Lu and Zhuo58) . For example, upon reaching the liver, endotoxins and exotoxins can trigger the overproduction of reactive oxygen species and reactive nitrogen species through the activation of oxidative and inflammatory pathways, particularly the NF-κB/iNOS axis(Reference Kim, Kim and Kwon11,Reference Albillos, de Gottardi and Rescigno59) . This activation leads to the excessive generation of nitric oxide and its reaction with superoxide anions to form peroxynitrite (ONOO⁻), a potent oxidant capable of damaging cell membranes, proteins and DNA(Reference Belenichev, Popazova and Bukhtiyarova60). The resulting oxidative stress promotes mitochondrial dysfunction, lipid peroxidation and hepatocellular injury, contributing to inflammation-induced metabolic disorders and the progression of hepatic steatosis and fibrosis(Reference Yang, Lu and Zhuo58,Reference Corbitt, Kimura and Isse61) .
The hepatic findings of this study further reinforce the interconnected nature of the gut–liver axis. In the DSS group, hepatic alterations were characterised by increased lipid accumulation and oxidative stress, consistent with changes in oxidative stress-related gene expression markers. Lipid overload in hepatocytes promotes excessive production of reactive oxygen species and free radicals, which in turn activate redox-sensitive pathways such as NF-κB and iNOS(Reference Manna and Jain62). This condition was accompanied by increased expression of antioxidant-related genes (CAT, SOD-1 and GSH-Px)(Reference Ma, Lee and Heo63,Reference Rives, Fougerat and Ellero-Simatos64) .
In contrast, embryos treated with camu-camu extract showed reduced lipid deposition and inflammatory infiltration, along with lower expression of NF-κB, iNOS and antioxidant-related genes. Specifically, the CC + DSS group presented reduced expression of NF-κB and iNOS compared with the DSS group, despite no significant increase of these markers in the DSS group relative to the water control. These findings indicate that the effects of camu-camu are better interpreted as a reduction in these markers relative to DSS exposure rather than a reversal of a DSS-induced activation (Figure 5). This protective effect may be partly attributed to decreased intestinal permeability and reduced translocation of pro-inflammatory and toxic molecules to the liver, as evidenced by improved intestinal barrier function (Figure 3). Moreover, camu-camu is rich in anthocyanins and other phenolic compounds with well-documented antioxidant and anti-inflammatory properties(Reference García-Chacón, Marín-Loaiza and Osorio18,Reference Baky, Elshahed and Wessjohann65) . These bioactives can be absorbed in the small intestine, an effect potentially enhanced by the increased villus surface area observed in the CC + DSS group, and reach the liver, which may be associated with their reported antioxidant properties.
Similar hepatoprotective effects have been reported in broilers supplemented with phenolic compounds such as resveratrol, which alleviated heat stress-induced liver injury by enhancing antioxidant defences and activating the Nrf2 signalling pathway, as well as in studies using polyphenol-rich grape by-products that modulated hepatic oxidative status and gene expression. Additionally, dietary polyphenols have been shown to reduce serum liver enzyme levels (e.g. ALT and AST) and mitigate toxin-induced hepatic damage, reinforcing their role in preserving liver function under stress conditions(Reference Mazur-Kuśnirek, Antoszkiewicz and Lipiński66–Reference Ding, Lu and Guo69). This dual mechanism, involving both reduced toxin exposure and direct antioxidant activity, likely contributes to the diminished demand for endogenous antioxidant enzyme production observed in the camu-camu-treated embryos(Reference Abot, Brochot and Pomié19,Reference Anhê, Nachbar and Varin20,Reference Agrinier, Morissette and Daoust24,Reference Do, Zheng and Park70) .
Moreover, morphological evaluation of liver tissue supports these molecular findings. Specifically, the CC + DSS group showed a larger cytoplasmic area and fewer lipid droplets compared with the DSS group, consistent with reduced lipid accumulation and altered hepatic morphology (Table 5; Figure 6). These findings are consistent with previous studies in broilers supplemented with phenolic compounds, such as resveratrol and phlorotannins, which demonstrated reduced hepatic lipid accumulation, inflammatory infiltration and overall histopathological damage under stress conditions, and are further supported by recent evidence showing that jabuticaba supplementation attenuates hepatic lipid deposition, inflammatory infiltrates and structural damage while restoring hepatocyte integrity in DSS-challenged models(Reference Pereira, José and Huang53,Reference Ding, Lu and Guo69,Reference Zhao, Yuan and Zhao71,Reference Sridhar, Suganthi and Thammiaha72) . This pattern parallels evidence that DSS-induced intestinal permeability promotes endotoxin translocation, leading to hepatic oxidative damage and lipid accumulation(Reference Kwon, Lee and Heo73–Reference Uno, Sekiguchi and Suzuki-Kemuriyama75). By improving intestinal barrier function, camu-camu likely reduced systemic exposure to antigens and toxic agents, limiting oxidative stress in hepatocytes.
Altogether, these findings reveal a dual mechanism of action by the investigated camu-camu extract, first, local intestinal effects that camu-camu extract is associated with changes in intestinal histomorphology, goblet cell parameters, barrier-related gene expression and hepatic morphology and gene expression markers. This integrated response underscores the functional interplay within the gut–liver axis, consistent with previous reports describing interactions between phenolic compounds and gut–liver axis-related parameters(Reference Mohammadhasani, Vahedi Fard and Mottaghi Moghaddam Shahri76,Reference Hu, He and Wang77) . Overall, the current study demonstrates that camu-camu extract exerts potent anti-inflammatory and antioxidant effects capable of mitigating DSS-induced dysfunctions in both intestinal and hepatic compartments. These protective effects likely stem from the action of phenolic compounds, which stabilise redox signalling, enhance barrier function and preserve metabolic integrity along the gut–liver axis, but these observations are limited to the measured morphological and gene expression outcomes.
Despite the relevance of the findings, some limitations should be acknowledged. The camu-camu used in this study consisted of a spray-dried extract containing only 20 % of the fruit pulp, without the peel, which is known to be a major source of phenolic compounds; this may have influenced the phytochemical composition and biological effects observed. Additionally, the absence of a group treated exclusively with camu-camu represents another limitation, as it is not possible to distinguish whether the observed effects are specific to the DSS challenge or reflect general effects of camu-camu on embryonic development, intestinal morphology or tissue viability. Moreover, the lack of a dose–response evaluation limits the ability to determine the relationship between camu-camu concentration and the observed outcomes.
Futhermore, the in ovo model represents an early developmental stage characterised by an immature immune system and a not yet fully established gut microbiota, which differs from the complex physiological and immunological conditions observed in humans. Additionally, the experimental design reflects an acute exposure to DSS, which may not fully recapitulate the chronic and multifactorial nature of inflammatory diseases affecting the gut–liver axis in humans. Although the in ovo model provides a controlled and valuable platform to investigate early mechanisms and direct effects of bioactive compounds, further studies in more complex and chronic in vivo models, as well as in humans, are necessary to confirm the translational applicability of these findings.
Conclusions
The present study shows that camu-camu is associated with changes in intestinal histomorphology, goblet cell parameters, barrier-related gene expression, hepatic histology and inflammatory- and oxidative stress-related gene expression markers. The bioactive compounds present in the extract, particularly anthocyanins and other phenolic compounds, were associated with modulation of oxidative stress-related gene expression markers (CAT, SOD-1 and iNOS). This modulation is consistent with the observed gene expression patterns, without direct assessment of enzymatic activity or functional outcomes.
Moreover, the findings indicate that camu-camu is associated with changes in inflammatory-related gene expression (e.g. NF-κB) and morphological features in the intestine and liver. Overall, these results are limited to the measured outcomes in this experimental model and do not allow conclusions regarding functional health effects or translational applications. Additionally, the camu-camu used in this study was a spray-dried extract containing only 20 % fruit pulp and excluding the peel, a major source of phenolic compounds, which may have influenced its phytochemical composition and observed effects.
Acknowledgements
The authors thank the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG, Brazil), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil), Federal University of Viçosa, and Cornell University.
The project already has financial resources from the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) (Finance Code APQ-01202–24) and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil) (Finance Code 001).
Conceptualisation: S. M. S. P., V. P. B. S. J., C. M. D. L. and E. T.; Data curation: S. M. S. P. and V. P. B. S. J.; Formal analysis: S. M. S. P. and V. P. B. S. J.; Investigation: S. M. S. P., V. P. B. S. J., M. Y. H. and L. C. S. A.; Methodology: S. M. S. P., V. P. B. S. J., M. Y. H. and E. T.; Project administration: C. M. D. L. and E. T.; Supervision: E. T.; Visualisation: S. M. S. P.; Roles/Writing – original draft: S. M. S. P. and V. P. B. S. J.; Writing – review and editing: S. M. S. P., V. P. B. S. J. and C. M. D. L, E. T.
The authors declare no conflict of interest.

 Open access
Open access