


The genus Canoparmelia Elix & Hale contains c. 40 species, predominantly occurring in tropical and subtropical regions. The genus is characterized by having relatively narrow, subirregular lobes with rotund or subrotund eciliate margins, a pored epicortex, the presence of isolichenan in the cell walls, bifusiform conidia, and simple rhizines (Elix Reference Elix1993; Crespo et al. Reference Crespo, Kauff, Divakar, del Prado, Pérez-Ortega, Amo de Paz, Ferencova, Blanco, Roca-Valiente and Núñez-Zapata2010b). Phylogenetically, it belongs to the Parmotrema clade of parmelioid lichens (Crespo et al. Reference Crespo, Kauff, Divakar, del Prado, Pérez-Ortega, Amo de Paz, Ferencova, Blanco, Roca-Valiente and Núñez-Zapata2010b). Since its original circumscription (Elix et al. Reference Elix, Johnston and Verdon1986), species have been transferred to other genera, including Austroparmelina A. Crespo et al. (Crespo et al. Reference Crespo, Ferencova, Pérez-Ortega, Argüello, Elix and Divakar2010a), Crespoa (D. Hawksw.) Lendemer & B. P. Hodk. and Parmotrema A. Massal. (Crespo et al. Reference Crespo, Kauff, Divakar, del Prado, Pérez-Ortega, Amo de Paz, Ferencova, Blanco, Roca-Valiente and Núñez-Zapata2010b; Hawksworth Reference Hawksworth2011; Lendemer & Hodkinson Reference Lendemer and Hodkinson2012; Kirika et al. Reference Kirika, Divakar, Crespo, Leavitt, Mugambi, Gatheri and Lumbsch2016). In addition, some species of the genus Crespoa are morphologically similar to certain Canoparmelia species, which leaves uncertainty regarding the identification of the samples.
The type species of Canoparmelia, C. texana (Tuck.) Elix & Hale, is the most widespread species of the genus. It has been reported from tropical and subtropical regions throughout the world and extends into temperate areas of North America (Hale Reference Hale1976). The species has also been reported from the Azores and the Canary Islands (Østhagen & Krog Reference Østhagen and Krog1976; Berger & Aptroot Reference Berger and Aptroot2002), but it has not yet been recorded from mainland Europe. Recent studies, however, have shown that C. texana is polyphyletic (Kirika et al. Reference Kirika, Lumbsch, Huescar, Quedensley and Divakar2022). Most East African collections were found to belong to a separate species, C. albaniensis (C. W. Dodge) Divakar & Kirika, and both taxa appear to have broad, multi-continental distributions. This further complicates the identification of species within the complex.
During our studies on the parmelioid lichens in the Iberian Peninsula, we collected material morphologically resembling Crespoa crozalsiana (B. de Lesd. ex Harm.) Lendemer & B. P. Hodk. (≡Parmelia crozalsiana B. de Lesd. ex Harm. France, Herault, Adge, rochers volcaniques ombragés, mai 1909 (USNH 68779-lectotype!, designated by Hale Reference Hale1976: 27)), but showing some variation. Hence, we revisited the material using morphological, chemical and molecular methods to disentangle the complex of morphologically similar taxa.
Morphological and chemical studies were performed as described previously (Kirika et al. Reference Kirika, Lumbsch, Huescar, Quedensley and Divakar2022). For the molecular analysis, we added 30 newly generated sequences from 13 samples to a dataset used previously (Kirika et al. Reference Kirika, Lumbsch, Huescar, Quedensley and Divakar2022), which consisted of a DNA data matrix of nuclear large subunit (nuLSU), internal transcribed spacer region (ITS), and mitochondrial small subunit (mtSSU) rDNA sequences. This dataset was used to infer evolutionary relationships among the taxa studied. Information on the material studied, including collection information and GenBank Accession numbers, is provided in Table 1. Laboratory protocols (including DNA extraction, PCR amplification and amplicon sequencing) and sequence alignment (including alignment and removal of ambiguously aligned regions) were carried out as described previously (Kirika et al. Reference Kirika, Lumbsch, Huescar, Quedensley and Divakar2022). Phylogenetic relationships were inferred using maximum likelihood (ML) as implemented in IQ-TREE v. 2.4.0 (Minh et al. Reference Minh, Schmidt, Chernomor, Schrempf, Woodhams, von Haeseler and Lanfear2020). The concatenated supermatrix was partitioned according to locus boundaries, treating ITS1, 5.8S, ITS2, nuLSU and mtSSU as separate partitions. Model selection and partition optimization were performed using the ModelFinder framework with automatic model selection and partition merging (–m MFP+MERGE), allowing the identification of the best-fitting substitution model for each partition and merging of partitions with similar evolutionary patterns. Branch support was assessed using 1000 ultrafast bootstrap replicates. The resulting best-scoring ML tree was used for graphical representation.
Specimens of Canoparmelia, Crespoa and other Parmeliaceae species used in the present study, with collection information and GenBank Accession numbers. Newly obtained sequences for this study are in bold and missing data are indicated with a dash (—). Reference sequences from the type localities are also highlighted in bold.
Results and Discussion
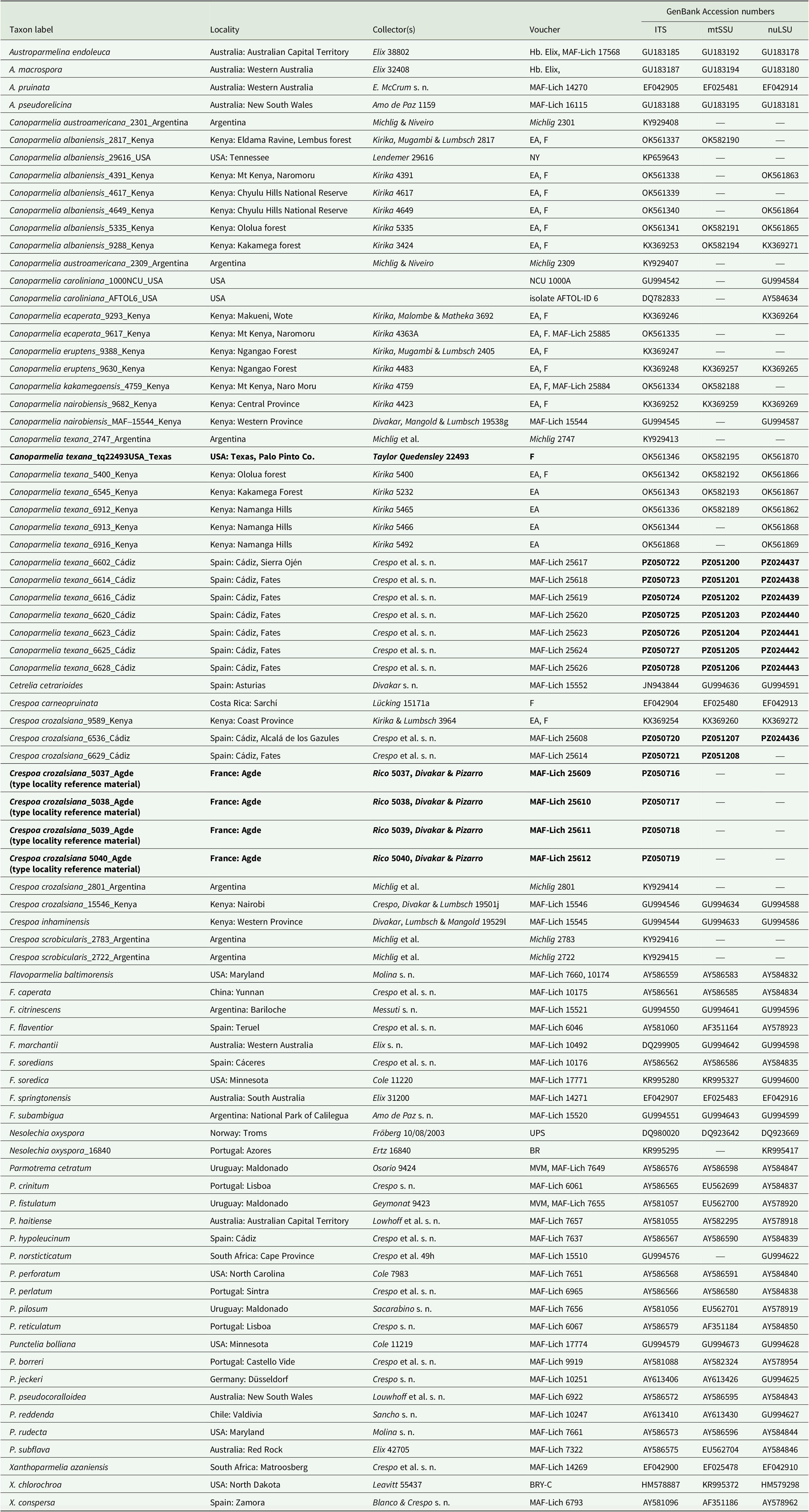
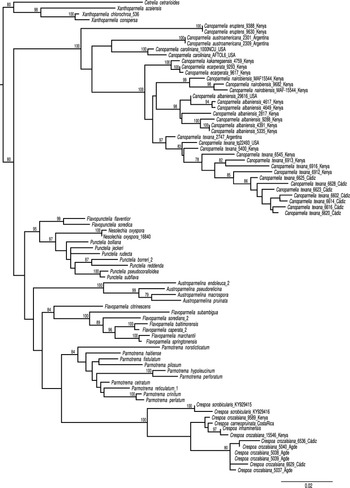
The phylogenetic analysis inferred from a three-marker dataset confirmed that the samples from southern Spain belong to Canoparmelia texana s. str. and Crespoa crozalsiana (Fig. 1), whereas C. albaniensis was not detected in the sampling. The Crespoa crozalsiana samples collected in Cádiz Province formed a monophyletic clade with samples from the Agde commune in southern France, the type locality of that species (Harmand Reference Harmand1910). In contrast, samples from Kenya clustered within a clade comprising Crespoa carneopruinata (Zahlbr.) Lendemer & B. P. Hodk., and Crespoa inhaminensis (C. W. Dodge) Lendemer & B. P. Hodk., indicating that the East African material requires further study. A specimen from Kenya (MAF-Lich 15546), previously annotated as C. schelpei (Hale) Lendemer & B.P. Hodk., is herein identified as C. crozalsiana, as it contains stictic acid complex rather than protocetraric acid. Consequently, all sequences generated from this specimen, including GenBank Accession numbers GU994546, GU994588, GU994634, KP888207 and KR995571 published in previous studies (Crespo et al. Reference Crespo, Kauff, Divakar, del Prado, Pérez-Ortega, Amo de Paz, Ferencova, Blanco, Roca-Valiente and Núñez-Zapata2010b, Divakar et al. Reference Divakar, Crespo, Wedin, Leavitt, Hawksworth, Myllys, McCune, Randlane, Bjerke and Ohmura2015) should be referred to as C. crozalsiana. Furthermore, the specimen of C. schelpei from the type locality (i.e., Mozambique), which contains protocetraric acid, does not belong to the genus Crespoa but instead to Parmotrema subgen. Africanae Kirika et al. (see Kirika et al. Reference Kirika, Divakar, Crespo, Leavitt, Mugambi, Gatheri and Lumbsch2016).
Phylogenetic relationships of Canoparmelia and related species based on maximum likelihood (ML) analyses of a concatenated, three-locus dataset (ITS, nuLSU and mtSSU rDNA). Maximum likelihood bootstrap values ≥ 70% are shown above branches. Cetrelia and Xanthoparmelia spp. were used as outgroup. Information for the specimens used in this analysis is given in Table 1.
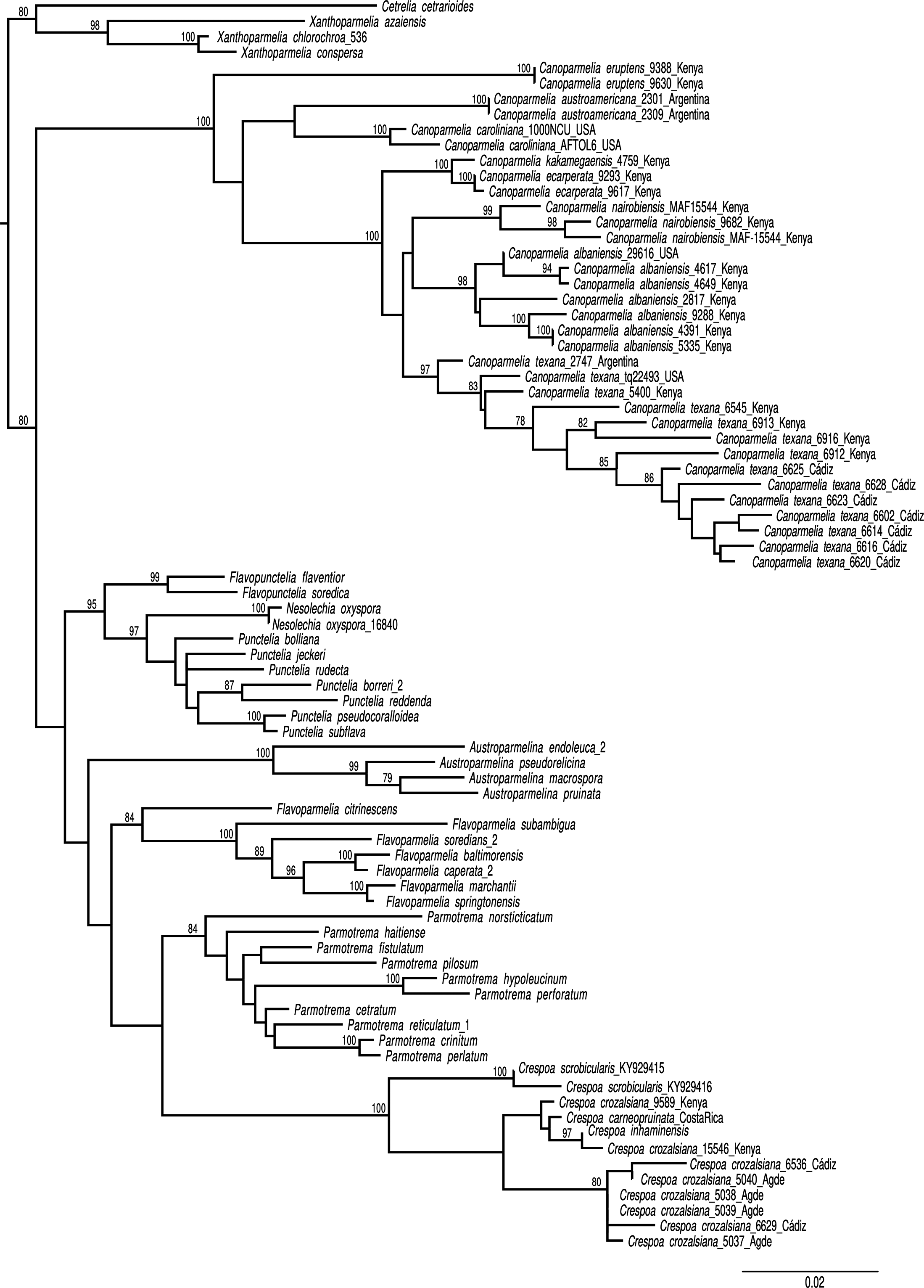
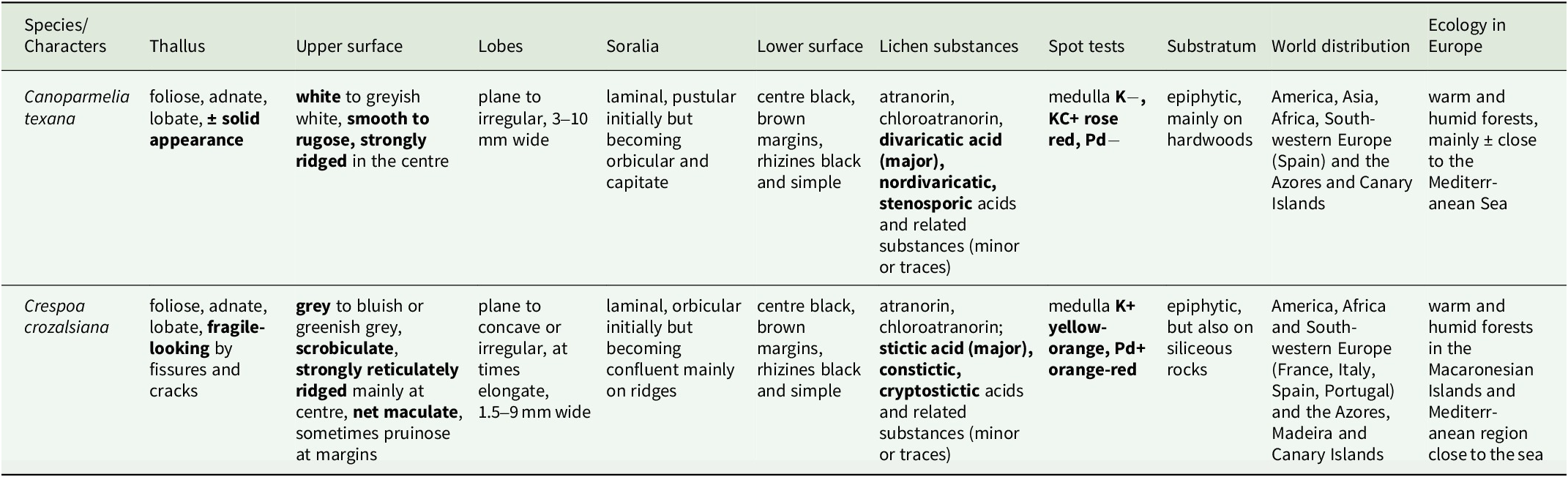
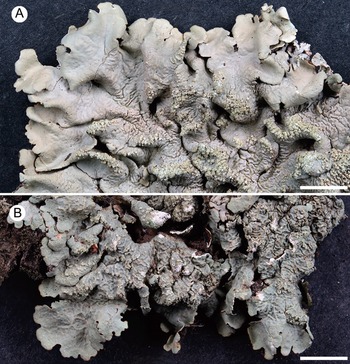
The seven samples of Canoparmelia collected in Cádiz Province (Table 1) formed a monophyletic group with Canoparmelia texana s. str. under its current circumscription (Kirika et al. Reference Kirika, Lumbsch, Huescar, Quedensley and Divakar2022). This is the first record of that species and the genus Canoparmelia s. str. from mainland Europe. The diagnostic features for distinguishing the morphologically similar species Canoparmelia texana and Crespoa crozalsiana (Fig. 2) are summarized in Table 2. Another superficially similar Crespoa species that also occurs in Europe (Hale Reference Hale1976; Hawksworth et al. Reference Hawksworth, Divakar, Crespo and Ahti2011; Jayalal et al. Reference Jayalal, Joshi, Oh, J-S and Hur2012) is C. carneopruinata. It is, however, readily distinguished from Canoparmelia texana by the presence of the stictic acid complex (stictic, constictic and cryptostictic acids).
A, Canoparmelia texana, from Cádiz, Spain (MAF-Lich 25623). B, Crespoa crozalsiana, from the French type locality in Agde (MAF-Lich 25611). Scales = 5 mm. In colour online.

Comparison of the morphological, chemical, and distributional diagnostic characters of Crespoa and Canoparmelia species in Europe. Diagnostic characters are shown in bold. Based on the specimens examined in this study.
We found a relatively high genetic variability within C. texana s. str., as shown in the branch lengths among samples of that species (Fig. 1). This indicates that additional species may be hidden under that name. However, this will require additional sampling through the entire distributional range of the species.
Data Accessibility
Newly generated sequences have been deposited in GenBank under Accession numbers PZ050716–PZ050728, PZ051200–PZ051208, and PZ024436–PZ024443.
Acknowledgements
This study was supported by the project PID2023-147388NB-I00 financed by the MCIN/AEI/10.13039/501100011033/FEDER, UE.
Author ORCIDs
Pradeep K. Divakar, 0000-0002-0300-0124; H. Thorsten Lumbsch, 0000-0003-1512-835X; Víctor J. Rico, 0000-0002-5064-2752.
Competing Interests
The author(s) declare none.


 Open access
Open access