


Non-technical Summary
Terrestrial ecosystems in Europe, but almost everywhere, changed significantly during the Cenozoic due to global climatic changes. The importance of their better understanding is magnified by present global climate change, which includes sea-level rise, as well as expansion in the distribution of thermophilic taxa, including parasites that cause serious infectious diseases. Squamates (lizards and snakes) are the largest and most successful group of living reptiles. They are globally distributed in different habitats and have a history dating back over 200 million years. The distribution, richness, and diversity of squamates is highly dependent on climatic conditions, and climate change has a marked impact on their life history. Thus, lizards are excellent indicators of past temperatures and precipitation. The appearance of many groups conceived to be thermophilic at high latitudes indicates that the tropics were much more expanded during the Eocene. We here describe pleurodontan iguanian (potentially Geiseltaliellus), scincoid and Squamata remains from Cos locality (France), one of the oldest sites of Phosphorites du Quercy. The age of the Cos deposit has been proposed to fit the MP 10−11 interval (late Ypresian). Thus, it either corresponds to the end of the Early Eocene Climatic Optimum (EECO) or slightly postdates it. In the last few years, our knowledge of early Eocene lizard faunas has increased substantially. This allows better comparison of the faunas and their changes due to temperature changes and migrations. Records are still very sketchy in European localities, but the overall picture is somewhat clearer. A small but visible drop in lizard diversity appears to be present in localities from MP 8−9, relative to those from MP 7, whereas at MP 10, the diversity slightly increased. This appears to correlate well with observed changes in temperature.
Introduction
In this paper, we describe the first evidence of iguanians and scincoid lizards from the Cos locality (44°13’11.20”N, 001°44’58.21”E) in France. The Cos fissure, in Quercy, was first discovered in 1865 by Jean André Poumarède (Pulou, Reference Pulou1980), but one of the present authors (DV) more recently discovered vertebrate fossils at the locality. The Cos fissure is the seventh, and one of the oldest, pre-upper Eocene localities known from Phosphorites du Quercy. Based on the studies of primates (Godinot et al., Reference Godinot, Blondel, Escarguel, Lézin, Pélissié, Tabuce and Vidalenc2021) and rodents (Vianey-Liaud et al., Reference Vianey-Liaud, Vidalenc, Orliac, Maugoust, Lézin and Pélissié2022), the Cos vertebrate fauna has been dated as MP 10−11 (MP 10b of Lihoreau et al., Reference Lihoreau, Yans, Benammi, Girard and Ballas2025; i.e., between MP 10 and 11) with an age estimated between ~50 and ~48 Ma, i.e., late Ypresian. Its age makes the Cos locality important, because the early Eocene terrestrial fossil record in Europe is poorly known. In fact, this locality represents one of the rare exceptions, serving as a window into that timeframe. The Cos age corresponds either to the end of, or slightly postdates, the Early Eocene Climatic Optimum (EECO; see Inglis et al. Reference Inglis, Bragg, Burls, Cramwinckel and Evans2020), which is estimated to be 53.3−49.1 Ma (see Lihoreau et al., Reference Lihoreau, Yans, Benammi, Girard and Ballas2025, fig. 12). The EECO was a time of major climatic and environmental change caused by perturbations to the global carbon cycle (Hyland et al., Reference Hyland, Sheldon and Cotton2017). The distribution, richness, and diversity of squamates, as ectothermic animals, are highly dependent on temperature and climatic conditions (Haller-Probst, Reference Haller-Probst1997; Rage, Reference Rage, Gardner and Nydam2013; Roll et al., Reference Roll, Feldman, Novosolov, Allison and Bauer2017). For this reason, they are excellent indicators of past temperature and precipitation, but data for the early Eocene is unfortunately rare. Regarding the squamates of Cos, several groups have been documented in the last few years, including anguimorphs (Čerňanský et al. Reference Čerňanský, Tabuce and Vidalenc2023a, for the geological setting and a map; Čerňanský et al., Reference Čerňanský, Tabuce and Vidalenc2023d), pan-gekkotan (Čerňanský et al., Reference Čerňanský, Daza, Tabuce, Saxton and Vidalenc2023b), and snake (Constrictores indet., Čerňanský et al., Reference Čerňanský, Georgalis, Tabuce, Vidalenc, Georgalis, Zaher and Laurin2025). In this study, we extend this faunal list by describing iguanian and scincoid specimens.
Iguanian lizards include two clades, Acrodonta and Pleurodonta (Iguanidae sensu Torres-Carvajal et al., Reference Torres-Carvajal, de Queiroz, Schulte, de Queiroz, Cantino and Gauthier2020). Today, Pleurodonta is primarily a New World group, with the exception of Madagascar and remote Pacific islands (Pough et al., Reference Pough, Andrews, Cadle, Crump, Savitzky and Wells2004). During the Eocene and Oligocene, however, Pleurodonta (e.g., Corytophanidae) was also present in Europe (Kuhn, Reference Kuhn1944; Estes, Reference Estes1983; Augé, Reference Augé1987, Reference Augé1990; Rossmann, Reference Rossmann2000; Smith, Reference Smith2009a; Augé and Pouit, Reference Augé and Pouit2012; Čerňanský et al., Reference Čerňanský, Klembara and Müller2016b, Reference Čerňanský, Smith, Smith and Folie2023c). Early Eocene pleurodontans from Europe include Geiseltaliellus Kuhn, Reference Kuhn1944 and Bifurcodentodon Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2023c documented from the earliest Eocene (MP 7) of the Dormaal locality in Belgium (Augé, Reference Augé1990, Reference Augé2005, Reference Augé2007; Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2023c). The former taxon is more spatially and temporally widespread, also reported from Silveirinha in Portugal (Rage and Augé, Reference Rage and Augé2003), Prémontré in France (~MP 10; Augé, Reference Augé2003a) and other sites. Moreover, beautifully preserved skeletons of Geiseltaliellus are known from early middle Eocene of Messel (MP 11; Smith, Reference Smith2009a, Smith and Wuttke, Reference Smith and Wuttke2012) and Geiseltal (MP 12 and 13, respectively; Kuhn, Reference Kuhn1944; Rossmann, Reference Rossmann2000; Smith, Reference Smith2009a). This taxon survived until the late Oligocene (MP 28) in France (Augé and Pouit, Reference Augé and Pouit2012) and potentially even later (MP 29) in Germany (Čerňanský et al., Reference Čerňanský, Klembara and Müller2016b). Besides Geiseltaliellus, there is also an exceptionally preserved, but undescribed, polychrotine specimen from Messel (see Smith et al., Reference Smith, Čerňanský, Scanferla, Schaal, Smith, Schaal and Habersetzer2018; MP 11). In later stages of the Eocene, several other taxa were recognized in Europe (see Augé, Reference Augé2005, Reference Augé2007), e.g., Cadurciguana Augé, Reference Augé1987 (MP 16−19) and Pseudolacerta De Stefano, Reference De Stefano1903 (?MP 16−19).
The Scincoidea clade includes Scincidae, Cordyliformes, and Xantusiidae (see Zheng and Wiens, Reference Zheng and Wiens2016). In Europe, scincoids are already known from the Paleocene (e.g., Čerňanský and Vasilyan, Reference Čerňanský and Vasilyan2024), with a well-documented record also from the early Eocene (e.g., Augé, Reference Augé2003a, Reference Augé2005).
Material and methods
Standard anatomical orientation system is used throughout this paper (Evans, Reference Evans, Gans, Gaunt and Adler2008; Rage and Augé, Reference Rage and Augé2010). All specimens were scanned at the microcomputed tomography (CT) facility at the Slovak Academy of Sciences in Banská Bystrica (Slovakia), using a Phoenix V|tome|x L 240 micro-CT. The CT datasets were analyzed using VG Studio Max 3.1 software (Volume Graphics GmbH, Heidelberg, Germany) and Avizo 8.1 (Visualisation Science Group). Specimens were also photographed under a scanning electron microscope (SEM; FEI Inspect F50) at the Slovak Academy of Sciences.
General abbreviations
Early Eocene Climatic Optimum (EECO); Eocene Thermal Maximum 2 (ETM 2); Mammal Paleogene intervals (MP); Paleocene-Eocene Thermal Maximum (PETM).
Repositories and institutional abbreviations
Cos collection, Université de Montpellier (UM-COS), France. The glyptosaurid parietal IPS 59567 discussed in this study is deposited at Institut Català de Paleontologia Miquel Crusafont (IPS), Barcelona, Spain.
Systematic paleontology
Squamata Oppel, Reference Oppel1811
Iguania Cope, Reference Cope1864
Pleurodonta Cope, Reference Cope1864 (sensu Frost et al., Reference Frost, Etheridge, Janies and Titus2001)
Genus Geiseltaliellus Kuhn, Reference Kuhn1944
Type species
Geiseltaliellus longicaudus Kuhn, Reference Kuhn1944.
?Geiseltaliellus sp. indet.
Pleurodontan iguanians (?Geiseltaliellus) from the early Eocene (MP 10−11) Cos locality, micro-CT visualizations and SEM. Right maxilla, UM-COS 1017, in: (1) lateral; (2) medial, with detail of teeth and tooth crowns; (3) dorsal; (4) ventral; and (5) dorsomedial views. Right dentary, UM-COS 1018, in: (6) lateral; (7) medial, with detail of tooth and tooth crown; (8) dorsal; (9) ventromedial; and (10) posteroventromedial views.

Occurrence
Cos, France, early Eocene.
Description
Of a right maxilla, only the posterior portion is preserved (Fig. 1.1−1.5). This element bears nine tooth positions, with five teeth still attached. The lateral surface is smooth and still covered by sedimentary matrix in some places. However, at least the posteriormost supralabial foramen can be observed, located at the level of the sixth tooth position (counted from posterior). CT revealed that another, smaller foramen is located at the level of the next anterior tooth position. This portion of the maxilla gradually narrows posteriorly to form a posteroventral process. There is no step at the dorsal margin. The medial side possesses a thin, but medially well-expanded supradental shelf. It is slightly dorsally convex, forms a contact with the palatine, and its maximum expansion is at the level between the sixth and seventh tooth positions (counted from posterior). It has to be noted, however, that the anterior portion of the bone is damaged and broken off. For this reason, a facet for the palatine cannot be recognized. The dental crest is moderately high, reaching approximately half of the general tooth height. On the dorsal surface of the supradental shelf, a region located laterally from the medial margin of the shelf close to the lateral wall of the posterolateral process is excavated and bears a longitudinal depression. At the approximate anterior third of the depression, at the level between the fourth and fifth tooth positions, a foramen forms the opening of the superior alveolar canal, which continues inside the bone and increases its size anteriorly. Although this foramen is atypically small, it most likely represents a superior alveolar foramen. Posterior to that, the depression is deeper and continues farther posteriorly, reaching the end of the bone. The depression forms (at least partly) a facet for the jugal. The depression diminishes anteriorly and reaches the approximate level of the seventh tooth position. Anterior to this, there is an additional tiny foramen. CT data reveal that this anterior foramen continues inside the bone as a narrow, moderately short canal that is connected to the large superior alveolar canal.
A right dentary fragment is also preserved (Fig. 1.6−1.10). Both anterior and posterior ends are broken off. The preserved portion bears 12 tooth positions, but only one tooth is preserved with a crown. The otherwise smooth lateral surface of the bone is pierced by three elliptical labial foramina. They are regularly distributed along the midregion of the bone. The medial side of the dentary is slightly expanded into the subdental shelf. The shelf is more or less straight in this portion, directed slightly posterodorsally. It roofs the Meckelian canal, which is fully open, but rather narrow anteriorly. The ventral margin of the bone is straight. Its posterior section forms a longitudinal facet for the splenial, dorsally to which lies an alveolar foramen. This foramen is in the posterior part of the bone, at the level directly posterior to the one preserved tooth. The intramandibular septum, which separates the Meckelian canal from the alveolar canal, is almost horizontal. The ventral margin of the septum forms a small and narrow crest (Fig. 1.9, 1.10). This crest is not free, but ventrolaterally fused to the bone, unlike the free posteroventral margin of the intramandibular septum (sensu Gauthier, Reference Gauthier1982). The above-mentioned opening of the alveolar canal (i.e., alveolar foramen) is located dorsal to that. The dental crest, which supports teeth laterally, is high. It reaches approximately half of the tooth height and, in fact, is higher than the ventral section of the dentary. This is better expressed in the anterior portion, because the whole dentary gradually narrows anteriorly.
Dentition
The tooth implantation is pleurodont. Both maxillary and dentary teeth are corroded (see Discussion) and have rather small interdental gaps. The only preserved dentary tooth is relatively high and surpasses the jaw parapet by approximtely half of its height. The overall size of the maxillary teeth is similar, except for the smaller, last preserved tooth. Teeth are more robust posteriorly, where they are tricuspid, with a dominant central cusp and mesial and distal accessory cusps of equal size, well separated from the main one. The tooth necks become broader toward the base. They are gradually mediolaterally expanded, but the tooth bases themselves are slightly compressed. Such a morphology is shared by both the dentary and maxillary teeth. Resorption pits are seen at the base of some teeth.
Materials
UM-COS 1017, right maxilla; UM-COS 1018, right dentary.
Remarks
Both maxillary and dentary teeth are tricuspid, except for the posteriormost maxillary one (the first preserved maxillary tooth has only incipient anterior cusps, but this can be caused by preservation). They have symmetrically developed mesial and distal cusps around the central dominant cusp as is typically seen in many pleurodontans (e.g., extinct Geiseltaliellus or extant Basiliscus Laurenti, Reference Laurenti1768). Although the Meckelian canal is open, it is also slightly restricted, especially in the anterior preserved section. This is also seen in some species of Basiliscus, Geiseltaliellus (Smith, Reference Smith2009a), and Suzanniwana Smith, Reference Smith2009b (Smith, Reference Smith2009b, fig. 3N), which might be closely related to Geiseltaliellus (Smith, Reference Smith2009a, Reference Smithb). Morphologically similar to the material from Cos, the latter taxon could represent a possibility regarding its taxonomic allocation, but this is unlikely biogeographically, because it is known from North America. Instead, Geiseltaliellus is widespread and common in the Eocene of Europe; known from the earliest Eocene of Dormaal in Belgium (MP 7; see Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2023c), the early Eocene of Prémontré in France (~MP 10, see Augé, Reference Augé2003a), and also from the later stages, e.g., early middle Eocene of Messel (MP 11; Smith, Reference Smith2009a) and Geiseltal (MP 12 and MP 13; Kuhn, Reference Kuhn1944 and Rossmann, Reference Rossmann2000, respectively) in Germany, and Grisolles (MP 16), Coânac (MP 19), and Escamps (MP 19) in France (Augé, Reference Augé2005). The material described here is similar to that described by Augé (Reference Augé2003a) from the locality of Prémontré and to that described by Čerňanský et al. (Reference Čerňanský, Smith, Smith and Folie2023c) from Dormaal and assigned to Geiseltaliellus. The matching characteristics include the distinctly restricted Meckelian canal and the well-developed subdental shelf. However, caution is needed because of its fragmentary condition, so we decided to assign the Cos material only questionably at the genus level.
Scincoidea Oppel, Reference Oppel1811 (sensu Zheng and Wiens, Reference Zheng and Wiens2016)
Scincoidea indet.
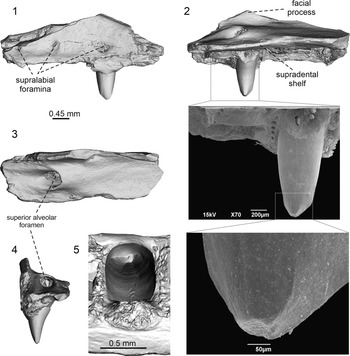
Scincoidea indet. from the early Eocene (MP 10−11) Cos locality, micro-CT visualizations and SEM. Left maxilla, UM-COS 1019, in: (1) lateral; (2) medial, with detail of tooth and tooth crown; (3) dorsal; and (4) posterior views. Tooth in: (5) ventromedial view.

Occurrence
Cos, France, early Eocene.
Description
Only a fragment of the left maxilla is preserved (Fig. 2). It bears five tooth positions, but only one tooth is preserved. The external surface is pierced by three supralabial foramina arranged in a single line. These foramina gradually increase in size posteriorly (Fig. 2.1). The dental crest is damaged, being partly preserved only around the tooth. The supradental shelf is medially well expanded, forming a slightly ventrally inclined table (Fig. 2.2, 2.4). Here, at the level of the preserved tooth, the table is pierced by a large superior alveolar foramen. The bone region, which frames the foramen anteriorly, has a swollen appearance. In contrast, the posterior region behind the foramen bears a depression. Only a small portion of the facial process is preserved, with only the dorsally rising base of its posterior portion visible.
Dentition
The tooth implantation is pleurodont. The preserved tooth is robust, slightly narrowing ventrally. In anterior and posterior (Fig. 2.4) views, the tooth base is medially expanded, whereas the neck and crown narrow distally. Thus, the whole lingual surface of the tooth is concave, being curved inward, whereas the labial surface is convex. The apical portion of the tooth is damaged, but the crown is clearly transversely bicuspid in occlusal view (Fig. 2.5). It has a larger labial edge, forming a labial cusp. Medially, there is a smaller, sharper, and well-defined lingual cusp. Both appear to be located at a mutual moderate distance from one another rather than being almost connected. Both cusps are distally displaced and thus, the cristae mesialis and distalis are not of the same length. These cristae of the labial cusp expand slightly at the base of the main apex, forming a tiny bulge (angulus mesialis and angulus distalis), slightly better developed on the anterior side. Further, the lingual margin of the crown is bordered by the weak culmen lateris anterior and culmen lateris posterior (Fig. 2.2). The lingual cusp is bordered by well-visible striae dominans anterior and posterior. These striae are almost parallel (especially in the apical portion), diverging only slightly farther in the direction of the tooth neck, where they gradually diminish. Thus, the region that they bound is narrow. No striations can be identified.
Materials
UM-COS 1019, left maxilla.
Remarks
The tooth crown is transversely bicuspid, with a lingual cusp together with striae dominans anterior and posterior. This indicates scincoid affinity (e.g., Caputo, Reference Caputo2004; Kosma, Reference Kosma2004; Folie et al., Reference Folie, Sigé and Smith2005; Čerňanský et al., Reference Čerňanský, Syromyatnikova, Kovalenko, Podurets and Kaloyan2020b; Čerňanský and Syromyatnikova, Reference Čerňanský and Syromyatnikova2021). Transversely bicuspid teeth can be present in some gekkotans, e.g., eublepharids (Sumida and Murphy, Reference Sumida and Murphy1987). Indeed, the pan-gekkotan Bauersaurus Čerňanský et al., Reference Čerňanský, Daza, Tabuce, Saxton and Vidalenc2023b was described from this locality (Čerňanský et al., Reference Čerňanský, Daza, Tabuce, Saxton and Vidalenc2023b). Yet, in contrast to the specimen described here, the tooth apices of this gecko have a rounded (blunt) appearance and, although they possess two cusps (labial and lingual), these are separated by a concave V-shaped sulcus. Moreover, the lingual cusp is broad (see Čerňanský et al., Reference Čerňanský, Daza, Tabuce, Saxton and Vidalenc2023b, fig. 4).
Squamata indet.
Squamata indet. from the early Eocene (MP 10−11) Cos locality, micro-CT visualizations and SEM. Left dentary, UM-COS 1020, in: (1) lateral; (2) medial, with detail of tooth; and (3) dorsal views.

Occurrence
Cos, France, early Eocene.
Description
A fragment of the midportion of the dentary is preserved (Fig. 3). It is C-shaped in cross section and bears eight tooth positions, with three more or less complete teeth and two partial ones still attached. The lateral surface of the bone is smooth and pierced by three large, more or less elliptical labial foramina along the midsection. In medial view, the Meckelian canal is fully open. It is roofed by a more or less straight (in its preserved portion) subdental shelf. Dorsally, the sulcus dentalis is absent. In fact, there is no bony elevation, e.g., a vertical lip of bone at the medial margin of the subdental shelf, which would protrude dorsally and thus, at least partly, cover the tooth bases medially. So, the whole dorsal surface of the shelf is exposed in medial view. The shelf is robust in the anterior preserved section, but it distinctly narrows posteriorly due to the presence of the facet for the splenial on its ventromedial surface. The splenial facet is medially exposed, reaching the level of the third preserved tooth position (counted from posterior). The splenial facet is also visible below this area, on the ventral margin of the bone, which is, however, damaged. Because of that, although it seems very likely that the Meckelian canal would narrow anteriorly, such a condition is difficult to access. The dental crest is high, being overarched by the teeth only by one-third of their lengths.
Dentition
The tooth implantation is pleurodont. Teeth are simple, tall, and conical. They are unicuspid and blunt in lingual aspect. The tooth crowns are slightly labiolingually compressed and thus, slightly concave lingually. In the two best-preserved teeth, the crowns appear to possess an elliptical surface, which fluently continues to the tooth neck (Fig. 3.2). However, the teeth look to be highly corroded and, thus, this does not seem to represent the original morphology. This is supported by the fact that the apices of these two teeth bear a stepped additional surface. The tooth necks are slightly swollen lingually, but the tooth bases are slightly compressed. Large elliptical resorption pits are present on the lingual aspects of tooth bases in some teeth. The narrow interdental gaps of the preserved teeth indicate that the teeth were rather closely spaced.
Materials
UM-COS 1020, left dentary.
Remarks
The specimen represents an animal of very small body size. However, it lacks potential diagnostic features, so that its allocation to any taxon is doubtful. The teeth seem to be highly corroded, hampering the identification of diagnostic traits. We can exclude an affinity to gekkotans, acrodont lizards, and, e.g., lacertids, but not other groups, e.g., skinks and anguimorphs. There is, however, no indication that this dentary belongs to the previously described taxa and it might plausibly indicate the occurrence of another lizard taxon in Cos. However, it also seems impossible to fully discard the possibility that it is an iguanian, and even another specimen of Geiseltaliellus. In the end, caution is needed in this case.
Discussion
Tooth corrosion as potential evidence of different predators in Cos
It must be noted that both maxillary and dentary teeth of the pleurodontan described here are corroded. In some maxillary teeth (Fig. 1.2), it seems that the crown is decalcified, but the shaft is not damaged (‘Type 1’ corrosion of Smith et al., Reference Smith, Comay, Maul, Wegmüller, Le Tensorer and Dayan2021, fig. 2e, f). In the dentary, it appears that the shafts have been corroded and only an enamel cap is left; note the ‘step’ both lingually and labially on the tooth crown (Fig. 1.6 and 1.7). That would correspond to ‘Type 2’ corrosion (potentially strigiform: low-acid, high-enzyme) of Smith et al. (Reference Smith, Comay, Maul, Wegmüller, Le Tensorer and Dayan2021, fig. 3i, k—experimental Anolis carolinensis Voigt, Reference Voigt1832). For this reason, it is very likely that the individuals, to which the maxilla and dentary belonged, were the prey of different predators. The ‘Type 1’ corrosion indicates a mammal predator (e.g., ferrets produce this type of corrosion in extant assemblages), with the gingiva already digested in the stomach. The tooth crown is ruined, whereas the shaft remains are almost undamaged (Smith et al., Reference Smith, Comay, Maul, Wegmüller, Le Tensorer and Dayan2021, fig. 1.1). In contrast, ‘Type 2’ corrosion could indicate the presence of birds of prey (e.g., owls produce this type of corrosion in extant assemblages) in Cos. In this case, dentine is the main subject of corrosion by acid and protease pepsin. The acidity is low and thus, the tooth shaft is corroded, whereas the tip is preserved (Smith et al., Reference Smith, Comay, Maul, Wegmüller, Le Tensorer and Dayan2021, fig. 1.2). The posterior teeth of UM-COS 1020 also look corroded (‘Type 1’ corrosion of Smith et al., Reference Smith, Comay, Maul, Wegmüller, Le Tensorer and Dayan2021).
The squamate paleobiodiversity in Cos during the early Eocene
Although the squamate fossils from the Cos locality are all represented by isolated and in most cases fragmentary individual bones, they can be taxonomically identified. Thus, they bring very important data about the paleobiodiversity of this reptile clade in Europe during a crucial time—the late early Eocene (MP 10−11), particularly during the end or directly after the Early Eocene Climatic Optimum (EECO). Among squamate lineages present in Cos, there were glyptosaurids, varanoids (Čerňanský et al., Reference Čerňanský, Tabuce and Vidalenc2023a, Reference Čerňanský, Tabuce and Vidalencd), a pan-gekkotan (Čerňanský et al., Reference Čerňanský, Daza, Tabuce, Saxton and Vidalenc2023b), and a constrictor snake (Čerňanský et al., Reference Čerňanský, Georgalis, Tabuce, Vidalenc, Georgalis, Zaher and Laurin2025). The present paper brings an additional report on new clades previously unknown in Cos—a pleurodontan iguanian, a scincoid, and one unidentified squamate (the latter potentially representing a further lizard lineage from this site, but one cannot fully exclude its iguanian affinity, see above). Overall, this shows that the paleobiodiversity of squamates at this locality was rather high and quite diverse.
Comment to the changes in paleobiodiversity of lizards during the early Eocene in Europe
The Cos locality represents a rare window into European paleoecosystems during MP 10−11. The lizard assemblage is quite unique in several aspects, yielding several taxa, e.g., Bauersaurus and Sullivanosaurus Čerňanský et al., Reference Čerňanský, Tabuce and Vidalenc2023d (see Čerňanský et al., Reference Čerňanský, Tabuce and Vidalenc2023a, Reference Čerňanský, Klembara and Müllerb, Reference Čerňanský, Tabuce and Vidalencd) currently known only from this locality. Comparison with other early Eocene assemblages in Europe is therefore interesting (Fig. 4). We can see that the paleodiversity of lizards during the Paleocene is low in Europe (Folie et al., Reference Folie, Sigé and Smith2005, Reference Folie, Smith and Smith2013; Rage, Reference Rage, Gardner and Nydam2013; Čerňanský et al., Reference Čerňanský, Augé and Phelizon2020a; Augé et al., Reference Augé, Dion, Phélizon, Steyer, Augé and Métais2021; Čerňanský and Vasilyan, Reference Čerňanský and Vasilyan2024), dominated by lacertoids (see Čerňanský and Vasilyan, Reference Čerňanský and Vasilyan2024), as is the case of snakes as well (see Smith and Georgalis, Reference Smith, Georgalis, Gower and Zaher2022). Although our knowledge about this geological epoch is limited (see Čerňanský and Vasilyan, Reference Čerňanský and Vasilyan2024), an important change happened around the Paleocene-Eocene boundary (see, e.g., Rage, Reference Rage2012, Reference Rage, Gardner and Nydam2013). As far as lizards are concerned, paleovaranids, scincoids, and lacertoids, as well as amphisbaenians (Folie et al., Reference Folie, Smith and Smith2013) survived the Paleocene-Eocene transition, but this was not the case of the typical Paleocene taxon Camptognathosaurus Folie, Smith, and Smith, Reference Folie, Smith and Smith2013. In the earliest Eocene sites of Dormaal, Belgium, and Silveirinha, Portugal, during the PETM, we have several new migrants (Rage, Reference Rage2012), e.g., agamids, pleurodontan iguanians, glyptosaurids, and varanids (see Augé and Smith, Reference Augé and Smith1997; Rage and Augé, Reference Rage and Augé2003; Augé et al., Reference Augé, Folie, Smith, Phélizon, Gigase and Smith2022; Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2023c, Reference Čerňanský, Smith, Smith and Folie2024), although some clades, e.g., paleovaranids (as mentioned above), appeared slightly earlier (Augé, Reference Augé, Wing, Gingerich, Schmitz and Thomas2003b).
Lizard-bearing European Paleocene and early Eocene localities (see Augé, Reference Augé1990, Reference Augé2003a, Reference Augé, Wing, Gingerich, Schmitz and Thomasb, Reference Augé2005, Reference Augé2012; Augé and Smith, Reference Augé and Smith1997, Reference Augé and Smith2002; Rage and Augé, Reference Rage and Augé2003; Folie et al., Reference Folie, Sigé and Smith2005, Reference Folie, Smith and Smith2013; Laurent et al., Reference Laurent, Adnet, Bourdon, Corbalan and Danilo2010; Bolet, Reference Bolet2017; Čerňanský et al., Reference Čerňanský, Augé and Phelizon2020A, Reference Čerňanský, Daza, Smith, Bauer, Smith and Folie2022, Reference Čerňanský, Tabuce and Vidalenc2023a, Reference Čerňanský, Daza, Tabuce, Saxton and Vidalencb, Reference Čerňanský, Tabuce and Vidalencd, Reference Čerňanský, Smith, Smith and Folie2024; Augé et al., Reference Augé, Folie, Smith, Phélizon, Gigase and Smith2022; Čerňanský and Vasilyan, Reference Čerňanský and Vasilyan2024) and their correlation to the temperature scale (modified from Zachos et al., Reference Zachos, Dickens and Zeebe2008). Presence of taxon indicated by +; absence indicated by -; Paleocene localities in blue column, earliest Eocene localities in dusty pink column (note that the exact age of Silveirinha within the PETM is unknown), later early Eocene localities in yellow columns. Arrows at right indicate lizard lineages that continue into the middle Eocene. Gekkota are highlighted in mustard yellow; Acrodonta in light gray; Pleurodonta in grass green; Glyptosauridae in blue; Varanidae in light salmon; Palaeovaranidae and Varanoidea in general in dark salmon; Eolacertidae in dark green; Lacertidae in mint green; Amphisbaenia in dark gray; Scincidae in light blue.

In the last few years, our knowledge of Paleocene and, especially early Eocene, lizard faunas increased thanks to finds in localities such as Walbeck (Čerňanský and Vasilyan, Reference Čerňanský and Vasilyan2024), Montchenot (Augé et al., Reference Augé, Dion, Phélizon, Steyer, Augé and Métais2021), Dormaal (Augé et al., Reference Augé, Folie, Smith, Phélizon, Gigase and Smith2022; Čerňanský et al. Reference Čerňanský, Daza, Smith, Bauer, Smith and Folie2022, Reference Čerňanský, Smith, Smith and Folie2023c, Reference Čerňanský, Smith, Smith and Folie2024), La Borie (Laurent et al., Reference Laurent, Adnet, Bourdon, Corbalan and Danilo2010), Cos (Čerňanský et al., Reference Čerňanský, Tabuce and Vidalenc2023a, Reference Čerňanský, Daza, Tabuce, Saxton and Vidalencb), and several sites in Spain (Bolet, Reference Bolet2017). Records are still too sketchy to allow much speculation regarding the reasons for missing groups in some European localities, but the overall picture of the changes is somewhat clearer (see Fig. 4).
Changes in lizard faunas during the early Eocene
Pleurodontan iguanians in the earliest Eocene (MP 7) are represented by two genera: Geiseltaliellus and Bifurcodentodon Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2023c, the latter with specialized dentition (Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2023c). However, only the first one is present during the later part of the early Eocene (EECO) in Masia de l’Hereuet (MP 8+9; see Bolet, Reference Bolet2017), Spain, Prémontré (Augé, Reference Augé2003a), and potentially in La Borie (~MP 8+9; Laurent et al., Reference Laurent, Adnet, Bourdon, Corbalan and Danilo2010) and Cos (MP 10−11), France, surviving until the late Oligocene (see Introduction). For European agamids (Tinosaurus Marsh, Reference Marsh1872), their oldest record is in Dormaal (Augé and Smith, Reference Augé and Smith1997; Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2023c), remaining present in Mutigny (MP 8+9; Augé, 2005) and Prémontré (~MP 10; Augé, Reference Augé2003a; Rage, Reference Rage2012), but with no evidence in Cos. Yet, we cannot exclude the possibility that the relatively small number of specimens and taxa in Cos might be a result of sampling or taphonomic bias—e.g., lacertids are absent in the material as well, although they are present in Prémontré (Fig. 4) and also in Viélase (personal observation of A.Č.; MP 10 or 11, Legendre et al., Reference Legendre, Marandat, Sige, Crochet, Godinot, Hartenberger, Sudre, Vianey-Liaud, Muratet and Astruc1992) in France.
As for glyptosaurids, Gaultia Smith, Reference Smith2009b is present in Dormaal, representing a connection to North America (Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2024; for USA record, see Smith, Reference Smith2009b). Gaultia is absent in Cos, but the somewhat similar Sullivanosaurus is present. This taxon appears to be more similar to Gaultia than to the late Eocene Placosaurus Gervais, Reference Gervais1848 in 1848–1852, although their exact phylogenetic relationship is currently unknown (see Čerňanský et al., Reference Čerňanský, Tabuce and Vidalenc2023a). Although glyptosaurids are known from MP 8−9 in Europe, their record is rather poor, with most of the material represented by isolated osteoderms from La Borie, France (Laurent et al., Reference Laurent, Adnet, Bourdon, Corbalan and Danilo2010), and Masia de l’Hereuet, Spain (Bolet, Reference Bolet2017). Therefore, more complete materials are needed to understand the evolution of this clade during this time interval. A partly preserved parietal, IPS 59567, from Escarlà (MP 10), was described by Bolet (Reference Bolet2017, fig. 1.12) and tentatively allocated to cf. Placosaurus sp. indet. The last author also compared it with Gaultia ragei (Sullivan et al., Reference Sullivan, Augé, Wille and Smith2012), previously described as ?Placosaurus ragei (see Sullivan et al., Reference Sullivan, Augé, Wille and Smith2012; Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2024). This parietal might plausibly represent a Sullivanosaurus-like glyptosaurid (known from the Cos locality, MP 10; Čerňanský et al., Reference Čerňanský, Tabuce and Vidalenc2023a, Reference Čerňanský, Tabuce and Vidalencd), showing much broader spatial distribution of this taxon. However, the parietal is incomplete and, moreover, no frontal (which is of major taxonomic importance) has been observed so far. New material from this locality could shed more light to this problem.
Varanoids are present in the early Eocene, but the occurrence of Saniwa Leidy, Reference Leidy1870 appears to be restricted to near the PETM, e.g., MP 7 of Dormaal, Belgium, and Le Quesnoy, France (Augé et al., Reference Augé, Folie, Smith, Phélizon, Gigase and Smith2022; Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2024), whereas paleovaranids are known through the whole Eocene, including Cos (Čerňanský et al., Reference Čerňanský, Tabuce and Vidalenc2023a) and even in the Oligocene (Augé and Smith, Reference Augé and Smith2009). Note that Saniwa feisti Stritzke, Reference Stritzke1983, known only from the early–middle Eocene of Messel, in Germany, is now assigned to the paleovaranid Paranecrosaurus feisti (see Smith and Habersetzer, Reference Smith, Habersetzer, Folie, Buffetaut, Bardet, Houssaye, Gheerbrant and Laurin2021).
In the European Paleogene, one of the clades that appeared in the early Eocene is Gekkota. Gekkotans are unknown in the Paleocene of Europe, although the Mesozoic record of this clade is well documented (Simões et al., Reference Simões, Caldwell, Nydam and Jiménez-Huidobro2017). However, gekkotans are relatively rare in the fossil record, probably because of their lightly built skeleton (e.g., Evans, Reference Evans, Gans, Gaunt and Adler2008). For this reason and because we still know very little about Paleocene paleobiodiversity, it is difficult to be sure whether geckos represent newcomers in Europe during the earliest Eocene or not. In fact, the lack of Paleocene gekkotans might be an artifact, given that most early Eocene localities share geckos of different types (Fig. 4). Their biodiversity is high in western Europe, including Dollogekko Čerňanský et al., Reference Čerňanský, Daza, Smith, Bauer, Smith and Folie2022 and Gekkota indet. from Dormaal (see Čerňanský et al., Reference Čerňanský, Daza, Smith, Bauer, Smith and Folie2022), the pan-gekkotan Bauersaurus from Cos (Čerňanský et al., Reference Čerňanský, Smith, Smith and Folie2023c), and Laonogekko Augé, Reference Augé2003a from Prémontré (Augé, Reference Augé2003a).
Conclusions
Terrestrial ecosystems in Europe, but basically in the entire world, changed significantly during the Eocene. The early Eocene climate experienced a sudden major warming (in a geologically brief time interval) during the Paleocene–Eocene Thermal Maximum at 56 Ma (Cramwinckel et al., Reference Cramwinckel, Huber, Kocken, Agnini and Bijl2018). The paleobiodiversity of lizards was then very high, represented by many thermophilic taxa, most of them newcomers. Shortly after this period, during a transient period corresponding to a significant and rapid drop in temperature (see Zachos et al., Reference Zachos, Dickens and Zeebe2008), the diversity of lizards appears to slightly decrease (Fig. 4). The earliest Eocene taxa, e.g., the iguanian Bifurcodentodon, the gecko Dollogekko, the varanid Saniwa, and others like the glyptosaurid Gaultia, seem to be restricted to the PETM, i.e., thermophilic and potentially more sensitive to the rapid environmental changes and temperature drops that occurred immediately after the PETM. This hypothesis is supported by the fact that members of these genera are already absent during the EECO (~53−49 Ma). The replacement of some members of these thermophilic groups by other members of the same groups can be observed and suggests a cooling event between the two assemblages. There are, of course, potential exceptions of some short-term survivors, e.g., Saniwa (documented based on caudal vertebrae from MP 8+9 of the Mutigny locality; see Augé et al., Reference Augé, Folie, Smith, Phélizon, Gigase and Smith2022). Some of them became extinct and some were possibly also replaced in certain areas by potentially related lineages (e.g., the glyptosaurid Sullivanosaurus).
In particular cases, some lizard populations might have shrunk during MP 8−9, occupying (probably small) suitable areas to recover, where lineages might have evolved to forms known from later time intervals (MP 10−11). For example, glyptosaurids are occasionally documented (mostly based on isolated osteoderms) from MP 8−9, e.g., La Borie, France (Laurent et al., Reference Laurent, Adnet, Bourdon, Corbalan and Danilo2010), and Masia de l’Hereuet, Spain (Bolet, Reference Bolet2017). As for scincoids, they are well documented from MP 10 in France, in the above-mentioned Spanish locality (see Bolet, Reference Bolet2017), but not in Mutigny or La Borie, France (Fig. 4). The thermophilic gekkotans are described from both Dormaal, Belgium (Čerňanský et al., Reference Čerňanský, Daza, Tabuce, Saxton and Vidalenc2023b Reference Čerňanský, Smith, Smith and Foliec), and Silveirinha, Portugal (Gekkonidae of Rage and Augé, Reference Rage and Augé2003), but appear to be absent in MP 8−9 localities, e.g., Mutigny and La Borie, France, and also Masia de l’Hereuet, Spain (MP 8+9; see Bolet, Reference Bolet2017). A different situation pertains to localities dated to MN 10, with gekkotans present in Prémontré, Cos (Fig. 4), and also Viélase (unpublished data, Čerňanský, 2025), France. In Spain, their occurrence is documented from La Morera (MP 10; Bolet, Reference Bolet2017).
Interestingly, some lineages, e.g., lacertoids, were successful in Europe through the entire Cenozoic (e.g., Augé, Reference Augé2005; Čerňanský and Augé, Reference Čerňanský and Augé2013; Čerňanský et al., Reference Čerňanský, Klembara and Smith2016a, Reference Čerňanský, Klembara and Müllerb, Reference Čerňanský, Bolet, Müller, Rage, Augé and Herrel2017; Čerňanský and Smith, Reference Čerňanský and Smith2018). Iguanians show a complex history, with the pleurodontan Geiseltaliellus surviving until the late Oligocene (see Introduction). The agamid Tinosaurus, which was present during the PETM, is also recorded at some localities during the EECO. The last documented occurrence of this lineage is at MP 10 (Augé, Reference Augé2003a), but it did not survive to the middle Eocene in Europe. In later stages, agamids are represented by Uromastyx Merrem, Reference Merrem1820 and Quercygama Augé and Smith, Reference Augé and Smith1997 (e.g., Augé, Reference Augé2005; Rage and Augé, Reference Rage and Augé2012).
We can conclude that the lizard fauna of MP 7 is different in many aspects from that of later reference levels of the early Eocene. Compared to amphibians, the relative number of salamanders appears to decrease at MP 8+9 relative to frogs (Rage, Reference Rage2012). According to Rage (Reference Rage2012), a warm interval (two optima: ETM 2, EECO) during MP 8+9 and MP 10 likely favored diversification of squamates relative to amphibians less dependent on temperature. However, there seems to be a small, but visible drop in lizard diversity in European localities from MP 8+9, relative to those from MP 7, whereas the diversity slightly increased at MP 10 (although some genera are different than those from MP 7; see Fig. 4). This appears to correlate with observed increases in temperature (Zachos et al., Reference Zachos, Dickens and Zeebe2008; Westerhold et al., Reference Westerhold, Marwan, Drury, Liebrand and Agnini2020) but could also reveal that the fossil record of MP 8+9 localities is simply poorer. Our knowledge about this geological epoch is still limited, especially considering that the early Eocene localities are nearly all known only from western Europe and the Iberian Peninsula (Rage, Reference Rage2012). New data can bring more robust conclusions in the future.
Acknowledgments
For access to fossil material from Cos, we thank M. Mouana (Institut des Sciences de l’Evolution de Montpellier, France). We are greatly indebted to K.T. Smith (Senckenberg Research Institute) for helpful advice. For critically reading the manuscript, we thank G. Georgalis (Polish Academy of Sciences) and one anonymous reviewer. This work was supported by the Scientific Grant Agency of the Ministry of Education of Slovak Republic and Slovak Academy of Sciences, Grant no. 1/0160/24.
Competing interests
The authors declare none.
Data availability statement
All specimens are cataloged and accessible in the fossil reptile collection of the University of Montpellier in France. Digital surface models of the figured fossil specimens are available on
Morphosource and Virtual Collections:
UM-COS 1017: https://www.morphosource.org/concern/media/000678361?locale=en
UM-COS 1018: https://www.morphosource.org/concern/media/000678371?locale=en
UM-COS 1019: https://www.morphosource.org/concern/media/000678382?locale=en
UM-COS 1020: https://www.morphosource.org/concern/media/000678388?locale=en




 Open access
Open access