


Introduction
The genus Cylindrobulla, with the type species by monotypy, C. beauii, was described from two empty shells from Guadeloupe in the Caribbean (Fischer, Reference Fischer1857). The original illustration shows a small, almost cylindrical shell with a deep apical funnel and a distinct apical keel and sutural slit, which are characteristic of this genus. At almost the same time, a species was described as Cylichna fragilis based on one tiny (<2 mm) shell from the Italian coast near Spezia (Jeffreys, Reference Jeffreys1856). This species was later transferred to the genus Cylindrobulla by Weinkauff (Reference Weinkauff1868: 185), and more recently to Ascobulla by Jensen (Reference Jensen1989). Unfortunately, Jeffreys’ figures of the shell (Plate 2, figs 16 and 17) do not show the apical area in sufficient detail to identify the genus, but all subsequent collections of the species from the Mediterranean, e.g., Thompson et al (Reference Thompson, Jarman and Zenetos1985), Marin and Ros (Reference Marin and Ros1988), Gavagnin et al (Reference Gavagnin, Marin, Castelluccio, Villani and Cimino1994), have clearly shown that it belongs to Ascobulla rather than Cylindrobulla.
The genus Cylindrobulla is externally almost indistinguishable from the genus Ascobulla. Both have thin, semi-transparent, nearly cylindrical shells with an apical keel and sutural slit surrounding a sunken spire, an anterior adductor muscle, a long gill attached to the mantle fold and composed of simple, ciliated folds, a bilobed ‘cephalic shield’, and a short, rounded foot (Jensen, Reference Jensen1989; Mikkelsen, Reference Mikkelsen1998; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). However, especially the radula and pharyngeal musculature of Cylindrobulla differ so much from those of other sacoglossans, including Ascobulla, that the inclusion of Cylindrobulla has been disputed (Marcus and Marcus, Reference Marcus and Marcus1970; Marcus, Reference Marcus1972, Reference Marcus1982; Jensen, Reference Jensen1989, Reference Jensen, Wells, Walker, Kirkman and Lethbridge1993b, Reference Jensen1996a, Reference Jensen and Taylor1996b). The uniseriate radula of Cylindrobulla has numerous small teeth of uniform size throughout the radula and with no pre-radular teeth (Jensen, Reference Jensen1989, Reference Jensen and Taylor1996b; Mikkelsen, Reference Mikkelsen1998; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). The teeth do not have a single long cusp, with or without lateral denticles, as found in all other sacoglossans. In Cylindrobulla, there is a central cusp, which is only slightly longer than the lateral cusps, which appear to be attached directly to the base rather than the sides of the central cusp. In other sacoglossans, the oldest used teeth are retained, either rolled up in a spiral where teeth remain attached to the radular ribbon, or in a densely packed heap (or heaps) surrounded by a membrane continuous with the epithelium lining the tube surrounding the descending limb of the radula. In Cylindrobulla, the oldest teeth just seem to fall off the radular ribbon and lie haphazardly inside the odontophore. Also, the odontophore has a long, unattached tip with many teeth exposed in both limbs, and the ventral, longitudinal ascus-muscle is thin and does not surround the descending limb of the radula throughout its length. The pharynx musculature in general is weaker than in other sacoglossans (Jensen, Reference Jensen and Taylor1996b). The presence of pharyngeal pouches as well as a ‘true’ ascus for discarded teeth has also been disputed (Jensen, Reference Jensen and Taylor1996b; Mikkelsen, Reference Mikkelsen1998). However, all recent phylogenetic analyses using molecular data (e.g., Maeda et al., Reference Maeda, Kajita, Maruyama and Hirano2010; Christa et al., Reference Christa, Händeler, Kück, Vleugels, Franken, Karmeinski and Wägele2015; Krug et al., Reference Krug, Vendetti, Ellingson, Trowbridge, Hirano, Trathen, Rodriguez, Swennen, Wilson and Valdés2015, Reference Krug, Caplins, Algoso, Thomas, Valdés, Wade, Wong, Eernisse and Kocot2022; Hirokane et al., Reference Hirokane, Miyamoto, Kitaura, Nakano, Hirano, Kawane and Yusa2022) unequivocally confirm the inclusion of the genus in the Sacoglossa.
Mikkelsen (Reference Mikkelsen1998) provided a historical overview of the systematic position of the genus. However, during the time interval from 1956, when Marcus and Marcus, (Reference Marcus and Marcus1956) described the anatomy of a species then called Cylindrobulla sp., and until Ascobulla Marcus, 1972, was formally included in the Volvatellidae by Marcus (Reference Marcus1982), species of Cylindrobulla were moving in and out of the Sacoglossa. Between 1972 and 1982, several species originally described as Cylindrobulla, but having radular teeth with a single, long cusp typical of sacoglossans and collected from Caulerpa beds, had been transferred to Ascobulla, e.g., C. japonica Hamatani, Reference Hamatani1969 and C. californica Hamatani, Reference Hamatani1971, and also Cylindrobulla fischeri A. Adams and Angas, 1864, was considered sacoglossan by Burn (Reference Burn1972) and was formally transferred to Ascobulla by Marcus (Reference Marcus1982).
Most of the early species descriptions were based on shells only, and therefore recent descriptions of new species have included comparisons of shells of species known to belong to either Cylindrobulla or Ascobulla to permit assignment of old species to the correct genus. In the present study, a new species of Cylindrobulla is described from Ghizo Island, Solomon Islands. The species is compared with other members of the genus, using original species descriptions and illustrations in addition to the characters described in Mikkelsen (Reference Mikkelsen1998: Table IV) and Laetz et al (Reference Laetz, Christa, Händeler and Wägele2014: Tables 2, 3, and identification key), as well as characters identified in the present study. Based on this, an emended diagnosis of the genus Cylindrobulla is presented. Finally, the feeding structures of Cylindrobulla are compared to those of other sacoglossans, shelled as well as non-shelled, focusing on the evolution of the unique sacoglossan piercing-sucking feeding method.
Materials and methods
As part of the Danish circumnavigation Galathea 3 (2006–2007), a team from the Zoological Museum, Natural History Museum Denmark, stayed at Ghizo Island in the western province of the Solomon Islands, from 30 December 2006 to 7 January 2007. We collected shallow water marine fauna from localities around the island and nearby islets, e.g., Kennedy Island, by snorkelling, scuba diving, and intertidal wading.
Specimens were collected by carefully sorting bags (1–5 l) of Halimeda and other green seaweeds collected during snorkelling or scuba diving. Most specimens were kept alive in Petri dishes with seawater for a few days before being preserved in 4% neutral formaldehyde and transferred to 80% ethanol after 24–48 h. Some specimens were photographed alive. Collection sites, dates, and number of specimens will be presented below. Type material has been deposited in the Mollusca collection of the Natural History Museum of Denmark.
Preserved specimens were photographed prior to dissection and at various stages during dissection, using a Nikon SMX-U stereomicroscope with a digital camera (Nikon DS-Fi1) attached. Due to its small size, the pharynx was only found in three of the five specimens dissected, one of these only after staining the cephalic organs in methylene blue. The first pharynx dissolved completely when placed in dilute NaOH. The subsequent ones were then placed in dilute household bleach, photographed under light microscopy (Nikon Optiphot-2) when partly dissolved, and the number of teeth was counted. The radula often broke into smaller pieces, which were difficult to rinse and dehydrate, so only one small piece was successfully transferred to an SEM (Scanning Electron Microscopy) stub and photographed in a JEOL JSM-6335F scanning electron microscope. Most of the descending limb of another radula was mounted in polyvinyl-lactophenol for light microscopy.
Slides of radula and penis used for the original description of Cylindrobulla phuketi Jensen, 1989, were re-examined for comparison. Preserved specimens of Ascobulla fragilis, Volvatella cf. candida (see Jensen, Reference Jensen2011 and Jensen, Reference Jensen and Wells1997a, respectively, for collection data of specimens), and an unidentified species of Volvatella collected in September 2018 from Singapore were used for comparison of apical areas of shells.
This work has been registered under ZooBank Accession: urn:lsid:zoobank.org:pub:D5D310CD-56B3-4C48-969D-64A4A8356025.
Systematics
Subclass HETEROBRANCHIA sensu Haszprunar, Reference Haszprunar1985
Superorder SACOGLOSSA von Ihering, 1876
Superfamily OXYNOOIDEA Stoliczka, 1868 (1847)
Family CYLINDROBULLIDAE Thiele, 1931
Genus Cylindrobulla Fischer, Reference Fischer1857
Type species Cylindrobulla beauii Fischer, Reference Fischer1857 by monotypy.
Diagnosis
An emended diagnosis will be presented after species comparisons.
Species Cylindrobulla ghizoensis sp. nov. (Figures 1–6)
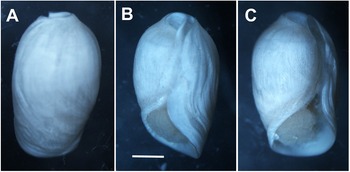
Two live specimens of Cylindrobulla ghizoensis sp. nov. photographed in outdoor laboratory space at Ghizo Island, Solomon Islands, both subsequently dissected. (A) specimen (from Kennedy Island), shell length approx. 6 mm; (B) specimen (from Titiana) with shell length approx. 3 mm. The debris on shells has been trapped in copious mucus secreted by live specimens.

ZooBank registration: urn:lsid:zoobank.org:act:3115090B-7C61-4301-A28C-B53ECFE52366
Synonym: Cylindrobulla sp. Jensen, Reference Jensen2011: Figs 1A, 2.
Material examined
Holotype. NHMD-2050150. 1 specimen, 4.5 mm shell length and 2.5 mm width (Figure 2), from mixed green seaweeds, mainly Halimeda, collected by J. Olesen on 3 January 2007 at Kennedy Island, Solomon Islands. Paratypes. 1 specimen from algae, mostly Halimeda and Caulerpa, collected in shallow water by the author near Titiana Village, Ghizo Island, on 31 December 2006, shell length 3 mm (Figure 1B; dissected); 2 specimens from Halimeda collected by divers at Kennedy Island on 3 January 2007, shell lengths 3.5 mm (Figure 3B) and 2.5 mm (Figures 5A, B, 6A, D; shell broken; dissected); 2 specimens same collection data as holotype, shell lengths 3 and 2 mm (both dissected; part of radula of 2 mm specimen used for SEM [Figures 5C, 6C]); 1 specimen, shell length approx. 6 mm (Figures 1A, 3A, 4, 6B; dissected), plus one empty shell from Halimeda collected by the author at Kennedy Island on 4 January 2007.
Preserved holotype (NHMD-2050150) of Cylindrobulla ghizoensis sp. nov. Shell length 4.5 mm. (A) dorsal view; (B) ventro-lateral view; (C) ventral view. Scale bar: 1 mm.

Apical area of two preserved specimens of Cylindrobulla ghizoensis sp. nov. (A) same specimen as Figure 1A, with nearly circular apical area; (B) specimen with shell length approx. 3.5 mm, with a typical teardrop-shaped outline. Both have a deep apical keel.

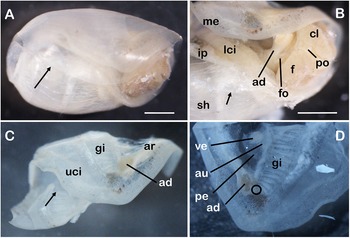
External and mantle features of Cylindrobulla ghizoensis sp. nov., shell length approx. 6 mm. (A) entire preserved specimen, lateral view; arrow points to transparent, overlapping outer lip of shell; mantle edge is not attached to edge of shell; (B) right anterior after removing part of shell; arrow points to thin reflected inner lip of shell (secreted by infrapallial lobe); (C) mantle fold cut off; arrow points to gill where it enters second whorl of shell; (D) close-up of anterior right corner of mantle fold viewed from inner side; black circle indicates position of presumed osphradium. Abbreviations: ad, adductor muscle; ar, adhesive ridge; au, auricle of heart; cl, lobe of head shield; f, foot; fo, female opening; gi, gill; ip, infrapallial lobe; lci, lower ciliated band; me, mantle edge; pe, pericardium; po, penial opening; sh, shell; uci, upper ciliated band; ve, ventricle of heart. Scale bars: 1 mm; A and C to the same scale and B and D to the same scale.

SEM of parts of Cylindrobulla ghizoensis sp. nov. (A) part of apical keel of specimen with shell length 2.5 mm; (B) upper whorl with protoconch of same specimen; (C) part of radula, specimen with shell length 2 mm.

Anatomical parts of Cylindrobulla ghizoensis sp. nov. (A) protoconch and uppermost whorl of specimen with shell length 2.5 mm; arrow points to growth discontinuity between protoconch and teleoconch; (B) pharynx of specimen with shell length 6 mm, partly dissolved in dilute bleach; several teeth dislodged from posterior end of radula ribbon forming almost empty circle. (C) radula of specimen with shell length 2 mm, stained with methylene blue; teeth at posterior end of descending limb in small circle; (D) penis of same specimen as (A); (E) preserved egg mass, partly disintegrated upon preservation, but single row of embryos in individual capsules within mass is distinct; (F) light micrograph of single egg capsule with embryo, apparently in early veliger stage. Abbreviations: a, ascending limb of radula; am, ascus muscle; c, capsule wall; d, descending limb of radula; e, oesophagus; em, embryo; pn, penis; ps, penial sheath; o, odontophore; rs, radula sac with large odontoblasts posteriorly; sm, dorsal septate muscle; vd, vas deferens. Scale bars: A, 250 µm; B, D, F, 100 µm; C, 50 µm; E, 1 mm.

Type locality: Kennedy Island, Solomon Islands: 08°06.78′S, 156°54.44′E and 08°06.74′S, 156°54.33′E.
Etymology: The species is named for Ghizo Island of the Solomon Islands, where it was first collected.
External features
Shell transparent in living specimens (Figure 1), becoming whitish opaque in most specimens when preserved. Shell of holotype is white with distinct growth lines (Figure 2). Soft parts are completely retractable into shell, and only foot and anterior lobes of head shield are visible in wide anterior opening, occupying about one-third of shell length. Anterior edge of body whorl with thickened margin. Behind this, outer lip slightly overhanging thin reflected inner lip, forming a narrow, slit-like aperture continuing as an apical slit (Figures 3, 4A). Shell is covered with irregular, short fracture lines in addition to fine growth lines. Apical keel forming a deep funnel (Figures 3, 5A). Protoconch is located at bottom of apical funnel (Figures 5B, 6A), though it is invisible in live specimens. Apical keel continuous with inner lip, which is deposited by shell glands of the infra-pallial lobe. Periostracum thin, flexible, and in some specimens extending beyond the shell edge, indicating that it is secreted before deposition of calcareous shell.
Head covered dorsally by a head shield divided longitudinally by a deep central groove. Lobes rounded anteriorly, overhanging the mouth area. Lobes are also rounded posteriorly, only slightly extending beyond the head. Neck short; can be extended when the animal tries to burrow or has been turned over accidentally (Figure 1B). Foot short, rounded in front and back, slightly longer than the head. Eyes small and black, visible laterally in groove separating head shield from foot (Figure 1B). Penial opening on the right side below and behind the eye (Figure 4B). A narrow ciliated groove running from the female genital opening and beyond the penial opening; this is the spawn groove.
Mantle organs
Mantle organs of this species have been illustrated in Jensen (Reference Jensen2011: Figure 2) but will be described in detail here. The mantle edge of preserved specimens is not attached to shell and can be retracted with head–foot part of body (Figures 2B, C, 4A). Anteriorly, the infra-pallial lobe merges with mantle edge on left side, and posteriorly with mantle edge at bottom of apical slit. Behind a broad anterior mantle edge, mantle is attached to neck by the adhesive ridge, which runs from adductor muscle on right side to left side, where mantle edge meets the infra-pallial lobe (Jensen, Reference Jensen2011: Figure 2C). Adhesive ridge also forms an attachment to shell. Tiny osphradium wedged between adductor muscle and right anterior corner of gill (Figure 4D). Behind adhesive ridge, mantle fold is thin-walled, and the underlying visceral organs (female glands and part of digestive gland) are visible (Jensen, Reference Jensen2011: Figure 2A, C). Gill lying across mantle fold, extending for more than 360°, entering second whorl (Figure 4C). Gill consists of a series of longitudinal, ciliated folds. Pericardium and heart are located in front of gill, slightly to the left of mid-dorsal line. Auricle short and thin-walled, ventricle almost spherical with muscular walls (Figure 4D). Kidney is located on top of gill. Behind gill is a narrow glandular band (Jensen, Reference Jensen2011: Figure 2C), possibly corresponding to hypobranchial gland in other gastropods. Dorsal (upper) ciliated ridge is located behind glandular band and, in combination with ventral (lower) ciliated ridge on mantle floor, forms a functional separation of anterior and posterior mantle cavities. Behind dorsal ciliated ridge is a second narrow glandular band (Jensen, Reference Jensen2011: Figure 2C). Ciliated ridges and glandular band extend into upper whorls of shell. Intestine emerging from digestive gland on mantle floor, and anus is located immediately in front of ventral ciliated ridge (Jensen, Reference Jensen2011: Figure 2E).
Internal anatomy
Mouth located under anterior lobes of head shield. Oral tube long, folded, and surrounded by glands. Pharynx very small, barely extending in front of cephalo-pleural ganglia. Pharynx of 6 mm specimen 257 µm long. Anteriorly, with a thick layer of circular muscles surrounding pharyngeal opening. Dorsal septate muscle (Figure 6B) is distinct, but thin relative to height of pharynx. Odontophore narrow with long, unattached tip containing very little musculature and many teeth exposed both dorsally (ascending limb) and ventrally (descending limb). The musculature of odontophore is weak. Only muscles extending diagonally from ascending limb to descending limb of radula (Figure 6B, C), probably functioning as radula protractor/ retractor muscles. Ventral layer of longitudinal muscle strands, the ascus-muscle, is very thin except for the anteriormost part. Radular sac with large odontoblasts extending posteriorly below oesophagus. Several teeth in radular sac have indistinct outlines, i.e., being in a formative stage. Long ascending limb of radula located very close to surface of odontophore, with many teeth exposed at long narrow tip. Descending limb almost equally long and attached to ventral surface of odontophore, ventrally covered by thin ascus-muscle. Number of teeth is high, and all teeth are of same size, but with some variation in number (two or three on either side of central cusp) and shape of cusps (Figure 5C). Tooth bases with large articulation knobs anteriorly and posteriorly. Specimen with shell length 3.5 mm had 42 teeth in ascending limb, 50 in descending limb, plus 16 teeth within odontophore, of which nine were still attached to a separate piece of radular ribbon. Specimen with shell length about 2 mm had 32 teeth in ascending limb, including six formative ones, 40 in the descending limb, and 10 attached to radular ribbon, bending upwards and forwards inside odontophore (Figure 6C). Teeth of this specimen measured from the SEM photo (Figure 5C) were about 6 µm long. Radula of specimen with shell length 6 mm broke into several pieces (Figure 6B); preliminary counting gave 33 teeth in ascending limb, including eight or nine formative ones, 42 in descending limb, and 13 mostly loose, but few attached to partly empty radular ribbon curling up inside odontophore. Subsequently, most teeth were accidentally lost, but what remained of descending limb, containing about 45 teeth of which 39 were attached to radular ribbon, was embedded in polyvinyl-lactophenol. Teeth measured 8.4 µm. This is slightly larger than in the 2 mm specimen, and thus there may be a barely recognizable increase in tooth size as the animal grows bigger.
The oesophagus is long and thin-walled, with two sharp bends where it passes through the ring of ganglia. No oesophageal pouch was observed in any dissected specimen. Due to difficulties in separating pharynx during dissections, salivary glands were not observed. Digestive gland a solid mass taking up most of the space of the visceral mass and extending into the protoconch (Jensen, Reference Jensen2011: Figure 2E, F). Intestine with several longitudinal folds.
Circum-oesophageal nerve ring consisting of elongate cerebro-pleural ganglia (210 × 151 µm in specimen with shell length 3 mm) connected by a short commissure, and pedal ganglia (189 × 168 µm in same specimen) also connected by very short commissure. Supra-oesophageal ganglion (189 × 126 µm) located rather close to right cerebro-pleural ganglion. Visceral chain broke in all dissected specimens when trying to extract pharynx so remaining ganglia were not observed.
Reproductive system was impossible to interpret. Penial aperture connecting to thin-walled penial sheath containing an unarmed, lobed, muscular penis (Figure 6D), which in specimen with shell length 3 mm was 281 µm long and 181 µm in diameter. Vas deferens broke close to penis in all dissected specimens, so it was impossible to trace backwards. Prostate not identified. On anterior left side of mantle floor, a large white glandular structure was visible (Jensen, Reference Jensen2011: Figure 2A, C). This is most likely the albumen gland; at least it is part of the female nidamental glands. Just behind and ventral to female genital aperture is a large sperm-filled structure, containing both sperm with heads attached to wall and free spermatozoa in lumen, thus apparently a combined bursa copulatrix and seminal receptacle. The free spermatozoa could also be from a recent copulation, so sperm not yet attached. In this case, the structure is a seminal receptacle.
Egg mass
One specimen produced an egg mass before it was preserved. The egg mass formed a string of single ova in individual capsules connected by chalazae (Figure 6E). Eggs had already cleaved when the egg mass was discovered, and it had to be preserved soon after, when embryos appeared to be early veligers (Figure 6F). Egg capsules preserved measured 214 ± 15 µm by 158 ± 10 µm (N = 8). Development is presumably planktotrophic; the protoconch in Figure 6A was about 180 µm in diameter.
Discussion
Comparison with other species of Cylindrobulla
Most species of Cylindrobulla are externally indistinguishable and also difficult to distinguish from species of Ascobulla. Mikkelsen (Reference Mikkelsen1998) lists 14 characters for comparing the two West Atlantic/Caribbean species of Cylindrobulla, C. beauii and C. gigas, and the sympatric Ascobulla ulla. Seven of these are shell characters. Laetz et al (Reference Laetz, Christa, Händeler and Wägele2014) list 19 characters, of which six relate to the shell, for separating species of Cylindrobulla and for distinguishing these from species of Ascobulla. Only three shell characters, outer lip shape, apex shape, and protoconch visibility, overlap in these two studies. Four characters of soft parts are shared: head shield furrow, shape and number of radular teeth, and intestinal typhlosole. In the present new species, C. ghizoensis, the shell shape (character 2 of Mikkelsen) apparently varies with size, small specimens being more cylindrical, larger specimens more bulloid, i.e., the maximum diameter is close to the apex. The shell length (character 1 of Mikkelsen) of C. ghizoensis sp. nov. was 2–6 mm with an average of 3.5 mm, which is smaller than the average length of C. gigas, but within the size range of most species of Cylindrobulla (Table 1). Also, transparency of the shell (character 2 of Laetz et al.) varies within C. ghizoensis sp. nov., some specimens having transparent shells, others opaque ones; furthermore, transparency was retained in a few specimens after preservation but became opaque in most. Unfortunately, only a single specimen or two have been described in detail for most species, so intraspecific variation of any character has been mentioned for just a few species. The apex of C. ghizoensis sp. nov. is what Mikkelsen calls teardrop (character 3) in smaller specimens (Figure 3B) but almost round in the largest specimen (Figure 3A; shell length 6 mm). The apical keel forms a steep cone rather than being rounded and flat (character 6 of Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). When viewed directly from above, the apical cone is oblique, with the bottom point (protoconch) being off-centre. This is what accounts for the teardrop shape. The anterior edge of the body whorl is thickened as in C. beauii. The outer shell lip shape (character 6 of Mikkelsen and char. 1 of Laetz et al.) of the present species differs among specimens from almost straight to highly rounded (convex). The protoconch (character 4 of Mikkelsen and char. 5 of Laetz et al.) is externally invisible even in the smallest specimen with a shell length of 2 mm. Hairline cracks in the shell (character 3 of Laetz et al.) occur in all examined specimens.
Comparison of species of Cylindrobulla with known soft parts

Table 1 Long description
The table compares seven Cylindrobulla species using shell size, protoconch diameter, external features (head shield, body color, eyes), feeding anatomy (radula counts, tooth size, cusp number), and selected internal and reproductive traits. Shell length varies widely: C. gigas reaches about 17.5 mm and C. beauii spans about 1 to 13.6 mm, while C. phuketi is smaller at about 1.2 to 4.6 mm and C. rozadora is listed at 3.5 mm. Protoconch diameter is reported for only three species, about 217 micrometers in C. beauii and about 176 to 180 micrometers in C. gigas and C. ghizoensis, with unknown values for the others. Head shields are described as longer than the shell in C. beauii, as long as the shell in C. schuppi, and shorter than the shell in several others; eye visibility also varies, with lateral visibility in C. beauii, C. phuketi, and C. ghizoensis but invisibility or deep embedding in others. Radula tooth totals range from low counts such as C. rozadora (32 ascending and 64 descending plus an ascus count) to high counts around 80 to 100 in C. gigas and 100 in C. schuppi; several species list separate ascending and descending limb counts and additional loose or ascus-associated teeth. Tooth size differs strongly, from about 6 to 8.4 micrometers in C. ghizoensis to about 200 micrometers for the ascending limb in C. xishaensis, and cusp patterns range from about 2 to 3 lateral cusps per side in C. phuketi and C. ghizoensis to about 7 in C. gigas. Where reported, all listed penes lack a stylet, and egg capsule size is provided only for C. gigas (about 300 by 200 micrometers) and C. ghizoensis (about 214 by 158 micrometers). Many cells are unknown or qualitative and several traits are referenced to figures in the cited sources, so cross-species comparisons are limited by missing data and differing reporting detail.
Abbreviations: a, ascending limb of radula; as, ‘ascus’; d, descending limb of radula; dg, digestive gland; H-F, head–foot; L, length; vm, visceral mass. ? indicates unknown or questionable.
a 0.2 mm is probably a mistake for 0.02 mm.
Soft parts and anatomy have been partially described for seven species of Cylindrobulla, summarized in Table 1. The type species, Cylindrobulla beauii, has been described anatomically in several papers (Marcus and Marcus, Reference Marcus and Marcus1970; Mikkelsen, Reference Mikkelsen1998; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014; Ortea, Reference Ortea2020). The descriptions of the shell are very similar, but descriptions of radular teeth vary among these descriptions. Cylindrobulla ghizoensis sp. nov. is externally similar to other species of Cylindrobulla for which morphology and anatomy of soft parts have been described. Eyes (character 7 of Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014) were visible laterally in living specimens, but not in preserved ones. The head shield (character 8 of Mikkelsen, Reference Mikkelsen1998) is shorter than the shell as in C. phuketi and C. rozadora and longer or as long as the shell in the other species. The furrow between head shield lobes (character 9 of Mikkelsen, Reference Mikkelsen1998 and char. 8 of Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014) is narrow with no gap, but not particularly shallow. The head, neck and foot of all species are opaque white, whereas various parts of the visceral mass and/or mantle may be coloured pale yellow, brown or orange, but in most species the digestive gland appears grey. Colouration in the mantle is restricted to clusters or bands of glands.
Tooth shape (character 10 of Mikkelsen, Reference Mikkelsen1998 and char. 11 of Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014) is the same in all species of Cylindrobulla described so far. They are small with broad bases and several short cusps, not much longer than the base. Tooth size of C. ghizoensis sp. nov. (character 12 in Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014) is very small even compared to other species of Cylindrobulla (Table 1). The slight variation in tooth size of C. ghizoensis may be an indication of an extremely small increase with the size of animal. The more noticeable variation in tooth size of C. beauii may also reflect differences in animal size, or that specimens of C. gigas were included in some studies. The number of lateral cusps is two or three in C. ghizoensis sp. nov., as well as in C. phuketi, but the size of the teeth is slightly bigger in the latter species, and the median cusp is distinctly larger than the first lateral cusps, whereas the lateral cusps in C. ghizoensis sp. nov. are almost the same length as the central one (Figure 5C). The teeth of C. schuppi have five or six lateral cusps, gradually decreasing in size laterally. The ascending (upper) limb of the radula of C. ghizoensis sp. nov. is slightly longer than the descending (lower) limb (character 14 of Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014), and the radular sac where new teeth are formed protrudes at the posterior end of the pharynx. The number of formative teeth (having indistinct outlines) in Cylindrobulla spp. is high. In C. ghizoensis sp. nov., 6–9 teeth with indistinct outlines are present. In C. beauii, seven or eight formative teeth can be seen in Figure 3A of Ortea (Reference Ortea2020), though only six are mentioned in the text. The number of formative teeth has not been mentioned in other descriptions of this species (Marcus and Marcus, Reference Marcus and Marcus1970; Mikkelsen, Reference Mikkelsen1998; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014), and none are visible in Figure 4E of the latter study. In C. schuppi, three formative teeth are visible in Figure 4D of Laetz et al (Reference Laetz, Christa, Händeler and Wägele2014). In C. rozadora, six formative teeth are mentioned, but seven are visible in Figure 5B of Ortea (Reference Ortea2020). Formative teeth have not been mentioned for other species, but re-examination of the original slide of C. phuketi showed eight or nine teeth with indistinct outlines (present study). The total number of teeth (character 12 of Mikkelsen, Reference Mikkelsen1998 and char. 13 of Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014) in all species is 80–100+ (Table 1). Pre-radular teeth, which occur in all other sacoglossans, are absent in all described species of Cylindrobulla. Thus, the anatomically described species seem to differ mainly in minute differences in size and number of cusps and/or denticles of the radular teeth.
Mikkelsen’s description of Cylindrobulla gigas includes a figure of a section of the pharynx (Mikkelsen, Reference Mikkelsen1998: Figure 3A) for comparison with a similar section of the pharynx of Ascobulla ulla. The figure shows a tall, narrow pharynx towards the posterior end where both the radular sac (a formative tooth) of the ascending limb of the radula and the descending limb of the radula with some loose teeth above it, called ascus by Mikkelsen (Reference Mikkelsen1998), are visible. It is noteworthy that there seems to be a layer of transverse muscles separating the ascending and descending limbs of the radula. This has not been observed in any other species of Cylindrobulla but is common in many other sacoglossans. The figure also shows a structure called ‘pharyngeal pouch’. However, no indication of an external muscular bolster is visible, and the lumen is connected to the oesophagus, which is not the case for pharyngeal pouches (see next section). The pharynx of C. ghizoensis sp. nov. is relatively taller than that of C. phuketi (compare Figure 6 of Jensen, Reference Jensen1989 and Figure 20.10D of Jensen,Reference Jensen and Taylor1996b with present study, Figure 6B). In both, only muscles running diagonally from ascending to descending limbs of the radula are seen within the long and narrow odontophore. In C. schuppi, a few superficial odontophore depressor muscles are present (Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014; Figure 4A). If scale bars are correct, the pharynx height of C. gigas and C. schuppi is at least twice that of C. phuketi and C. ghizoensis sp. nov.
An oesophageal pouch has been described for C. beauii, C. phuketi and C. schuppi (Marcus and Marcus, Reference Marcus and Marcus1970; Jensen, Reference Jensen1989; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014), but not for other species. Salivary glands have been described as bulky in C. beauii (Marcus and Marcus, Reference Marcus and Marcus1970) and as sac-like in C. schuppi (Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). Re-examination of C. phuketi showed that it has long, thin salivary ducts, as in C. schuppi, leading to large salivary glands close to body wall (present study). They are unknown for the remaining species.
The intestine of C. ghizoensis sp. nov., as seen on the surface of the digestive gland, has a number of longitudinal folds, but a typhlosole (character 14 of Mikkelsen, Reference Mikkelsen1998 and char. 17 of Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014) was not distinguishable. A typhlosole has been described in C. beauii and C. gigas (Mikkelsen, Reference Mikkelsen1998) but was not found in C. schuppi (Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). It is unknown for the remaining species.
The penis of Cylindrobulla spp. is unarmed, i.e., without a chitinous stylet. As in all sacoglossans, it is protrusible, enclosed in a penial sheath and connected by an internal, closed, and partly embedded in lateral cephalic musculature, vas deferens to the visceral hermaphrodite duct. The size of the penis differs among species (Table 1). Neither Mikkelsen (Reference Mikkelsen1998) nor Laetz et al (Reference Laetz, Christa, Händeler and Wägele2014) gave length measurements for penes of their new species. For C. schuppi, the penis was described as long and cylindrical, and the schematic drawing indicates that the penis is as long as the head (Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). The penis of C. phuketi is almost twice the length of that of C. ghizoensis sp. nov. for specimens of approximately the same shell length (Table 1).
Other parts of the reproductive system are difficult to interpret. A prostate gland has been identified in the three species that have been serially sectioned. It was described as having a ‘flower configuration’, i.e., as petals of a flower surrounding a narrow cylindrical lumen, for C. beauii and C. gigas (Mikkelsen, Reference Mikkelsen1998), and a similar structure was described for C. schuppi (Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). This is similar to the prostate of other sacoglossans (see, e.g., Baba, Reference Baba1961; Marcus, Reference Marcus1972; Sanders-Esser, Reference Sanders-Esser1984; Kohnert et al., Reference Kohnert, Brenzinger, Jensen and Schrödl2013). A prostate gland was not found in C. phuketi or in C. ghizoensis sp. nov. (Jensen, Reference Jensen1989, present study), but this probably just means that it is not located in the cephalic region. Mikkelsen (Reference Mikkelsen1998) indicated a pallial position, whereas Laetz et al (Reference Laetz, Christa, Händeler and Wägele2014) indicated a visceral position.
The number of allo-sperm receptacles also differs in species of Cylindrobulla. Two receptacula have been described for C. beauii, C. gigas, C. schuppi and tentatively for C. phuketi (Jensen, Reference Jensen1989; Mikkelsen, Reference Mikkelsen1998; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014), but only one receptacle was seen in C. ghizoensis sp. nov. (present study). The bursa copulatrix, located near the female gonopore, is large and contains irregularly arranged free spermatozoa. The receptaculum seminis, containing spermatozoa with heads embedded in epithelium, is located deeper in the viscera, and in three of the above species it is smaller than the bursa (Jensen, Reference Jensen1989; Mikkelsen, Reference Mikkelsen1998; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). In C. ghizoensis sp. nov., the single receptaculum is located close to the female gonopore, but contains attached as well as free, unoriented spermatozoa (present study).
The circum-oesophageal (i.e., post-pharyngeal) nerve ring consists of fused cerebro-pleural ganglia and separate pedal ganglia. The length of the cerebral commissure is described as long in C. beauii (Mikkelsen, Reference Mikkelsen1998), probably corresponding to a relatively large pharynx. The ganglia are illustrated as distinctly smaller than the pharynx by Marcus and Marcus (Reference Marcus and Marcus1970). A long cerebral commissure is also described for C. schuppi, and based on sections, pleural and cerebral ganglia were closely attached rather than fused (Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). In C. phuketi, the fused cerebro-pleural ganglia are almost as long as the pharynx (Jensen, Reference Jensen1989), and in C. ghizoensis sp. nov., the pharynx is completely surrounded by the ganglia (pharynx length 257 µm, cerebro-pleural ganglia 210 µm long), and the cerebral commissure was short (present study). The number of ganglia on the visceral loop and, particularly, their interpretation, differ among species. Mikkelsen (Reference Mikkelsen1998) mentions separate pallial ganglia, at least on the left side for C. beauii and C. gigas, but this has not been observed in any other species. In other shelled sacoglossans, the small ganglion close to the left cerebro-pleural ganglia has been interpreted as the sub-oesophageal ganglion (Jensen and Wells, Reference Jensen, Wells, Wells, Walker, Kirkman and Lethbridge1990; Jensen, Reference Jensen and Wells1997a).
Three of the known species of Cylindrobulla, C. beauii, C. gigas and C. rozadora, are from the Caribbean and the West Atlantic from Florida to Brazil (Marcus and Marcus, Reference Marcus and Marcus1970; Mikkelsen, Reference Mikkelsen1998; Ortea, Reference Ortea2020). Another three species, C. xishaensis, C. schuppi and C. ghizoensis sp. nov., are from the West Pacific (Lin, Reference Lin1978; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014; present study), and one, C. phuketi, is from the Andaman Sea in the Indian Ocean (Jensen, Reference Jensen1989). Hedley (Reference Hedley1899) lists C. sculpta from Funafuti, which is about the same latitude as Ghizo, and it may actually be C. ghizoensis sp. nov. rather than the Indian Ocean species. However, it may also have been a species of Ascobulla, since there is no illustration of the specimen. The present new species, C. ghizoensis, thus seems to be the first Indo-Pacific species anatomically described from the southern hemisphere.
In addition to the species mentioned above, there may be a number of species that remain undescribed. Carlson and Hoff (Reference Carlson and Hoff2003) mentioned a species labelled Cylindrobulla sp. 1 from Guam, but this is most likely the species described from Guam as C. schuppi by Laetz et al (Reference Laetz, Christa, Händeler and Wägele2014). Krug et al (Reference Krug, Vendetti, Ellingson, Trowbridge, Hirano, Trathen, Rodriguez, Swennen, Wilson and Valdés2015) included two undescribed species, one from Lord Howe Island, Australia, and the second taken from GenBank with no collecting data included. Krug et al (Reference Krug, Caplins, Algoso, Thomas, Valdés, Wade, Wong, Eernisse and Kocot2022) included a specimen from New Caledonia labelled Cylindrobulla sp. 2. Neither the specimen from Australia nor the one from New Caledonia has been described morphologically, and there is a chance that either one could be the species described here from Ghizo, Solomon Islands. Gosliner et al (Reference Gosliner, Valdés and Behrens2015) illustrated five unnamed species of Cylindrobulla from Hawaii, the Philippines, Papua New Guinea, and Aldabra, but based only on photos that do not show the apical area adequately, it is not possible to decide whether they belong to Cylindrobulla or Ascobulla.
Only the seven species listed in Table 1 have been confirmed by radular teeth as belonging to the genus Cylindrobulla. Five additional species are known only from more than 100-year-old descriptions of shells. The externally similar genus Ascobulla also contains very few species. In fact, only five species have been described anatomically, and four of these were originally described as Cylindrobulla. Two original descriptions are more than 100 years old and based only on shells. Molecular analyses have shown that the two genera are not closely related, Ascobulla being closely related to Volvatella and the two genera comprising the family Volvatellidae (Maeda et al., Reference Maeda, Kajita, Maruyama and Hirano2010; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014; Christa et al., Reference Christa, Händeler, Kück, Vleugels, Franken, Karmeinski and Wägele2015; Krug et al., Reference Krug, Vendetti, Ellingson, Trowbridge, Hirano, Trathen, Rodriguez, Swennen, Wilson and Valdés2015; Hirokane et al., Reference Hirokane, Miyamoto, Kitaura, Nakano, Hirano, Kawane and Yusa2022). In connection with more recent descriptions of new species of Cylindrobulla, some of these old species have been transferred tentatively to Ascobulla. As indicated above, the only shell character consistently separating Cylindrobulla from Ascobulla is the shape of the apex area, which is deep and conical with an invisible protoconch in Cylindrobulla and rounded with a low keel and visible protoconch in Ascobulla. Careful examination of old descriptions and illustrations (Figure 7) has now shown that a few additional shell characters are usable for distinguishing Cylindrobulla from Ascobulla, as described below.
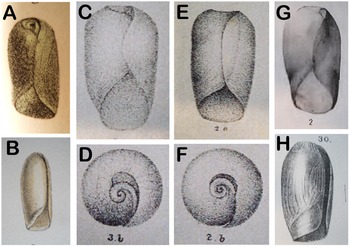
Scanned copies of published original illustrations of Cylindrobulla species. (A) Cylindrobulla beauii from Fischer, Reference Fischer1857; (B) Cylindrobulla souverbiei from Souverbie and Montrouzier, Reference Souverbie and Montrouzier1874; (C–D) Cylindrobulla sculpta from Nevill and Nevill, Reference Nevill and Nevill1869; (E–F) Cylindrobulla pusilla from Nevill and Nevill, 1869; (G) Cylindrobulla turtoni from Bartsch, Reference Bartsch1915; (H) Cylindrobulla systremma from Melvilll, Reference Melvill1918.

Nevill and Nevill (Reference Nevill and Nevill1869) described two species, C. sculpta and C. pusilla, both of which were transferred to Ascobulla by Gosliner (Reference Gosliner1987) and Mikkelsen (Reference Mikkelsen1998). Only the latter was tentatively moved to Ascobulla by Jensen (Reference Jensen1989). The original drawing of the apical area, reproduced here in Figure 7D, shows a ‘teardrop-shaped’ area for C. sculpta, as mentioned for C. beauii by Mikkelsen (Reference Mikkelsen1998), and illustrated for C. xishaensis by Lin (Reference Lin1978: Figure 3), C. phuketi by Jensen (Reference Jensen1989: Figure 3A), C. schuppi by Laetz et al (Reference Laetz, Christa, Händeler and Wägele2014: Figure 1E), as well as for C. ghizoensis sp. nov. (present study: Figures 2B, C, 3B). The latter five species all belong to Cylindrobulla based on radular morphology. Furthermore, Nevill and Nevill (Reference Nevill and Nevill1869) mentioned a ‘fattened’ inner lip for C. sculpta as also found in C. beauii and C. ghizoensis sp. nov. (Mikkelsen, Reference Mikkelsen1998; present study). Re-examination of the paratype of C. phuketi in the Natural History Museum Denmark showed a thickened inner lip in this species too, and this is also indicated for C. xishaensis (Lin, Reference Lin1978: Figure 1), and thus C. sculpta can be referred to this genus. The apical area of C. pusilla was round (Nevill and Nevill, Reference Nevill and Nevill1869: Figure 2B, reproduced here in Figure 7F), as described for Ascobulla ulla by Mikkelsen (Reference Mikkelsen1998), and also for C. gigas. A circular apical area is also found in Ascobulla fischeri (see Jensen, Reference Jensen and Taylor1996b: Fig. 20.1D) and A. fragilis (present study, Figure 8A). For C. pusilla, Nevill and Nevill (Reference Nevill and Nevill1869) mention similarity with both C. beauii and Volvatella candida Pease, 1868, a species externally similar to Ascobulla and Cylindrobulla, but with the distinct pharynx and radula of Volvatella (Jensen, Reference Jensen and Wells1997a). The original figures of C. pusilla show a much wider apical area than seen in V. candida, and the transfer of C. pusilla to Ascobulla seems justified.
Apical area of (A) Ascobulla fragilis and (B) Volvatella cf. candida. (C) Ventral view of Volvatella sp. from Singapore. Abbreviations: af, apical funnel; as, sutural slit; ca, calloused part of sutural slit; sp, posterior shell spout; u, anterior umbilicus. Scale bars: 1 mm.

Cylindrobulla souverbiei Montrouzier, 1874 has a very long (14 mm) and slender shell with a thickened anterior margin of body whorl. The apex is described as sunken with open suture, but a sutural keel is not mentioned (Montrouzier in Souverbie and Montrouzier, Reference Souverbie and Montrouzier1874). Shell length of more than twice the shell diameter has been mentioned for some specimens of C. beauii (Marcus and Marcus, Reference Marcus and Marcus1970) but not for any other species. The original description mentions eight specimens of approx. the same shell length, but variable widths. The original figure, reproduced here in Figure 7B, indicates a teardrop-shaped apical area. The original description mentions a yellowish periostracum that was also mentioned in the original description of C. beauii. The thickened anterior margin of body whorl and teardrop apical area indicate that it is most likely a Cylindrobulla.
Cylindrobulla turtoni Bartsch, 1915 has been synonymized with Volvatella laguncula Sowerby III, 1894 by Gosliner (Reference Gosliner1987). However, a distinct posterior ‘spout’ is mentioned for the latter species, both in the original description (with no illustrations) and in Thompson (Reference Thompson1979). A tiny drawing in Sowerby III (Reference Sowerby1897: plate VI, fig. 17), as well as personal observations of the type specimens (1899.4.14.3600-2) in the Natural History Museum, London (NHMUK), show a short and narrow apical spout, most distinct from the dorsal side. Sowerby III (Reference Sowerby1894, Reference Sowerby1897) mentions a specimen of Cylindrobulla sculpta, and that specimen was also examined in the NHMUK when I visited in 1988. My old notes identified this specimen as ‘definitely a Cylindrobulla with steep apical funnel’; it was 6 mm long and 4 mm wide, approximately the same size as C. turtoni and collected from Port Alfred, South Africa, whereas C. turtoni was from Port Elizabeth, South Africa, both within the same biogeographic zone (Gosliner, Reference Gosliner1987). Bartsch (Reference Bartsch1915) mentions specimens of V. laguncula from the same collection, so obviously he (or Turton, the collector) considered them different species. The original figure in Bartsch (Reference Bartsch1915), reproduced here in Figure 7G, does show a certain resemblance to the shell of Volvatella candida with a strongly convex outer lip and a wider apical area than other species of Volvatella (compare Figures 8B and C), although still narrower than Ascobulla and Cylindrobulla, including C. turtoni. Axial riblets on the apical shoulder are mentioned in the text, and these are also seen in V. cf. candida (Figure 8B), in C. ghizoensis sp. nov. (Figures 3 and 5A [towards lower left]), and more widely spaced in several species of Ascobulla (Marcus, Reference Marcus1972; Jensen and Wells, Reference Jensen, Wells, Wells, Walker, Kirkman and Lethbridge1990; Jensen, Reference Jensen2011; present study, Figure 8A). Species of Volvatella, including V. cf. candida, have an ‘umbilicus’, usually filled with debris, anteriorly, where the aperture widens (Figure 8C). This cannot be seen in the figure of C. turtoni (Bartsch, Reference Bartsch1915). As the soft parts of C. turtoni are still unknown, its correct generic affiliation remains uncertain.
Cylindrobulla systremma Melvill, 1918 has the diagonal line near the inner lip (original illustration reproduced here in Figure 7H), which is also seen in C. gigas, A. ulla (Mikkelsen, Reference Mikkelsen1998), A. fragilis (Jensen, Reference Jensen2011: Figure 4B) and A. fischeri (Hedley, Reference Hedley1903: plate 29, fig. 9; pers. obs.). This line may be a growth discontinuity line, indicating that growth after this point extends the inner lip and shortens the anterior aperture. As this line is found in several species of Ascobulla and in only one species of Cylindrobulla, it seems likely that C. systremma should be transferred to Ascobulla as previously suggested by Jensen and Wells (Reference Jensen, Wells, Wells, Walker, Kirkman and Lethbridge1990).
In conclusion, Cylindrobulla sculpta is here re-instated as a species of Cylindrobulla, and C. pusilla and C. systremma remain transferred to the genus Ascobulla. The specific synonymization of these two species remains uncertain, as does the generic placement of C. souverbiei and C. turtoni. Adding information on shells from the seven anatomically described species to the above descriptions, the thickened inner lip and the teardrop-shaped apical area, i.e., with acentric nucleus, are characters of the genus Cylindrobulla in addition to the steep apical keel and externally invisible protoconch.
Emended diagnosis of genus Cylindrobulla
Small shelled sacoglossans (<20 mm shell length). Shell thin, fragile, unpigmented, nearly cylindrical or somewhat bulloid. Anterior aperture wide, approximately one third of total shell length or less. Anterior edge of body whorl thickened. Inner lip thin and reflected. Apical area with sutural slit and deep sutural keel forming an oblique cone with externally invisible protoconch at bottom. Head, neck and foot white. Head covered dorsally by elongate muscular shield divided longitudinally by deep, narrow, median groove. Foot short, rounded anteriorly and posteriorly. Gill forming band around entire outer whorl, visible through shell. Pericardium and heart in front of gill. Kidney in mantle roof, covered by gill. Strong, diagonal adductor muscle attached to right anterior corner of shell, in front of gill. Small suctorial pharynx. Radula uniseriate with numerous (80–100+) small, uniformly sized teeth with several almost equally sized cusps on a squarish base. Penis unarmed.
Comparison of feeding structures of Cylindrobulla and other Sacoglossa
The Sacoglossa are characterized by being suctorial herbivores, feeding almost exclusively on siphonalean green algae, such as Caulerpa, Halimeda, or Codium. Obviously, the evolution and diversification of the Sacoglossa are intimately associated with the development of morphological adaptations of the feeding apparatus for increased feeding efficiency, as evidenced in the unique piercing-sucking feeding method, which has been observed in shelled as well as non-shelled sacoglossans (e.g., Gascoigne, Reference Gascoigne1956; Kawaguti and Yamasu, Reference Kawaguti and Yamasu1966; Jensen, Reference Jensen1980, Reference Jensen1981, Reference Jensen1994, Reference Jensen and Wells1997a). Remarkably, all shelled sacoglossans except Cylindrobulla feed exclusively on algae of the genus Caulerpa. There are no published observations on feeding in Cylindrobulla, but several species have been found associated with the often bioherm-forming, calcareous genus Halimeda, and molecular studies have identified genes of Halimeda in C. schuppi as well as C. beauii (Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014).
It has been known since the first anatomical description of Cylindrobulla beauii that its radula differed in several aspects from that of the ‘usual’ sacoglossan radula (Marcus and Marcus, Reference Marcus and Marcus1970), and for many years its inclusion in the Sacoglossa was contested (Marcus and Marcus, Reference Marcus and Marcus1970; Marcus, Reference Marcus1982; Jensen, Reference Jensen1989, Reference Jensen1996a), although a sister-group relationship was accepted (Jensen, Reference Jensen and Taylor1996b). Molecular studies have all shown that Cylindrobulla is firmly lodged within the Sacoglossa, and furthermore, it is included with the other shelled sacoglossans in the superfamily Oxynooidea (Maeda et al., Reference Maeda, Kajita, Maruyama and Hirano2010; Christa et al., Reference Christa, Händeler, Kück, Vleugels, Franken, Karmeinski and Wägele2015; Krug et al., Reference Krug, Vendetti, Ellingson, Trowbridge, Hirano, Trathen, Rodriguez, Swennen, Wilson and Valdés2015, Reference Krug, Caplins, Algoso, Thomas, Valdés, Wade, Wong, Eernisse and Kocot2022; Hirokane et al., Reference Hirokane, Miyamoto, Kitaura, Nakano, Hirano, Kawane and Yusa2022). In the next section, the differences in the feeding apparatus of Cylindrobulla from that of other sacoglossans will be outlined.
The unique adaptations associated with the piercing-sucking feeding method and their importance for the evolution of the Sacoglossa have been elucidated in a series of papers (Jensen, Reference Jensen1991, Reference Jensen1992, Reference Jensen1993a, Reference Jensen1993c, Reference Jensen and Morton1993d, Reference Jensen1994, Reference Jensen1996a, Reference Jensen1997b). These specialized characters include: (1) A uniseriate radula with articulating teeth; (2) a radula composed of an upper (or ascending) limb and a lower (or descending) limb, i.e., the used teeth are retained; (3) unicuspid, interlocking radular teeth, increasing in size with age; (4) pre-radular teeth (forming the radula of newly metamorphosed juveniles); (5) a muscular, suctorial pharynx composed of (i) a dorsal, septate muscle (alternating dorso-ventral (radial) and (semi)circular muscles), (ii) a muscular odontophore enclosing most of the ascending limb of the radula, and (iii) a ventral, longitudinal ascus-muscle more or less surrounding the descending limb of the radula; (6) a tissue-enclosed ascus for accumulating used teeth at the posterior end of the descending limb of the radula; (7) functional (or anatomical) separation of ascending and descending limbs of radula; and (8) muscular pharyngeal pouches attached postero-ventrally to pharynx. Only three of these complex characters (1, 2 and 5) are found in Cylindrobulla. The presence of characters 6–8 has been debated and will be discussed here.
The presence of an ascus for accumulating used radular teeth is the hallmark character of the Sacoglossa. Its presence in Cylindrobulla has been discussed almost since the genus was separated from Ascobulla by Marcus (Reference Marcus1972) (Marcus, Reference Marcus1982; Jensen, Reference Jensen1989, Reference Jensen1996a, Reference Jensen and Taylor1996b; Mikkelsen, Reference Mikkelsen1998; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). Unfortunately, the definition of an ascus has not been unequivocally fixed. Some authors (e.g., Fretter, Reference Fretter1940; Gabe and Prenant, Reference Gabe and Prenant1957; Mikkelsen, Reference Mikkelsen1998) consider the ascus to be the entire epithelium-lined structure surrounding the major part of the descending limb of the radula. Others, e.g., Gascoigne (Reference Gascoigne1956, Reference Gascoigne1978), Bleakney (Reference Bleakney1988), Wägele et al (Reference Wägele, Stemmer, Burghardt and Händeler2010), Krug et al (Reference Krug, Vendetti and Valdés2016), and the present author, consider the ascus to be only the posteriormost part of the descending limb, with the used teeth either rolled up in a spiral or forming a densely packed heap. Part of the confusion is probably due to the longitudinal muscles surrounding the descending limb of the radula being called the ascus-muscle, a term apparently coined by Tom Gascoigne (Reference Gascoigne1978). In all sacoglossans except Cylindrobulla, the teeth at the posterior end of the descending (lower) limb remain surrounded by a thin membrane visible in many SEM photos (e.g., Jensen, Reference Jensen1992: Fig. 15; Krug et al., Reference Krug, Vendetti and Valdés2016: Figures 8C, 26C, 50C, 63C, H, 69C; Moreno et al., Reference Moreno, Rico, Middlebrooks, Medrano, Valdés and Krug2024: Fig. 10; McCarthy-Taylor et al., Reference McCarthy-Taylor, Krug, Muro, Vendetti, Maestrati, Wong, Gosliner and Valdés2025: Figs 14C, 23C). This membrane may be the radular ribbon and/or the epithelium surrounding the descending limb of the radula. In some species, the sac with accumulated teeth is located between the ascending and descending limbs, e.g., in Plakobranchus spp. (Jensen, Reference Jensen1992: Fig. 23), and Gascoignella aprica (Kohnert et al., Reference Kohnert, Brenzinger, Jensen and Schrödl2013: Figure 2B), but in most species the ascus forms a thin-walled sac behind the ascus-muscle, e.g., most species of Elysia and Ercolania (Jensen, Reference Jensen1992, Reference Jensen, Wells, Walker, Kirkman and Lethbridge1993b). In a few species, e.g., Ascobulla and Volvatella, the descending limb is short, ending in an almost straight row (Jensen, Reference Jensen1993c, Reference Jensen1996a, Reference Jensen2015), in which case the ascus is only the pre-radular and intermediate teeth. In most species of Cylindrobulla, the teeth at the posterior end of the descending limb seem to fall off the radular ribbon and lie haphazardly within the odontophore (Jensen, Reference Jensen1989; Mikkelsen, Reference Mikkelsen1998; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). In C. rozadora, Ortea (Reference Ortea2020) described a spiral ascus containing four or five teeth, but a surrounding membrane was not mentioned. For C. beauii, a heap of eight loose teeth in an ascus was mentioned (Ortea, Reference Ortea2020), but the illustration shows, in addition, two or three pieces of radula broken off the posterior end of the descending limb. The posteriormost end of the descending limb curving upwards into the odontophore was also indicated for C. beauii by Marcus and Marcus (Reference Marcus and Marcus1970: fig. 29), but not mentioned in text or in any of the more recent descriptions of this species (Mikkelsen, Reference Mikkelsen1998; Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014). It was, however, noticed in the smallest specimen of C. ghizoensis sp. nov. (Figure 6C), whereas the posteriormost teeth of the larger specimens fell off the radular ribbon (Figure 6B), or pieces of the radular ribbon broke off, leaving teeth loose inside the odontophore. In C. schuppi, the structure labelled as ascus (Laetz et al., Reference Laetz, Christa, Händeler and Wägele2014: Figure 4B) appears to be the oesophagus with the salivary reservoirs next to it. It seems clear that all old teeth are retained in Cylindrobulla and not periodically shed as suggested by Marcus and Marcus (Reference Marcus and Marcus1970). However, the radular ribbon is apparently less resistant and easily broken when the musculature is chemically dissolved. This may indicate a different chemical composition from that of other sacoglossans. It remains difficult to identify an ascus as a distinct, separate structure in Cylindrobulla, except in the few cases where the posterior part of the descending limb rolls up, and the teeth remain attached.
Pre-radular teeth are another key innovation of the Sacoglossa (Jensen, Reference Jensen1996a). They are formed in metamorphosis-competent larvae, as observed in Elysia subornata (as E. cauze), Oxynoe azuropunctata and Berthelinia singaporensis (Clark et al., Reference Clark, Busacca, Stirts and Stancyk1979; Jensen, Reference Jensen1980; Jensen and Ong, Reference Jensen and Ong2018), and in the directly developing Limapontia senestra (Mikhlina et al., Reference Mikhlina, Khramova, Nikitenko and Vortsepneva2024). However, pre-radular teeth have not been observed in any species of Cylindrobulla. Pre-radular teeth differ between shelled and non-shelled genera. Shelled sacoglossans have a single rod-shaped pre-radular tooth (Baba, Reference Baba1961, Reference Baba1966; Gascoigne and Sartory, Reference Gascoigne and Sartory1974; Jensen, Reference Jensen1980; Kawaguti and Yamasu, Reference Kawaguti and Yamasu1982; Jensen and Wells, Reference Jensen, Wells, Wells, Walker, Kirkman and Lethbridge1990), although this has not yet been observed in Lobiger, where even the smallest specimen studied (shell length about 2 mm) had a dense heap of used teeth in the ascus (unpubl. pers. obs.). The Plakobranchacea have a small number (3–7) of tiny, cusp-less pre-radular teeth (see, e.g., Jensen, Reference Jensen1996a: Figure 6, Reference Jensen2014: Figures 3, 10), which are usually distinct in juvenile animals but may be obscured in larger animals if they have a densely packed heap of discarded teeth. Although pre-radular teeth have not yet been observed in several genera of Plakobranchacea, the fact that the ascus and/or descending limb of the radula contain teeth of decreasing sizes indicates that pre-radular teeth will be present and that more juvenile specimens should be examined. The length of the teeth of Cylindrobulla is nearly equal to that of pre-radular teeth and bases of early teeth of some Plakobranchacea (compare Table 1 of the present study with Tables 1 and 2 of Jensen, Reference Jensen2014). This may indicate that newly settled Cylindrobulla do not need specially formed teeth to initiate feeding.
In all sacoglossans except Cylindrobulla, the teeth of the main part of the radula are unicuspid with cusps longer than bases. Tooth bases are squarish with anterior and posterior articulation knobs, also seen in Cylindrobulla, but cusps are elongate with pointed tips and may have small denticles along one or two edges. The cusps of teeth still attached to the radular ribbon are more or less interlocked, with the cusp of one tooth fitting into a depression of the tooth in front or covering a keel of the neighbouring tooth. This means that the teeth in both limbs form rigid rods of interlocking teeth, except for a few teeth at the tip of the odontophore, which are not enclosed in musculature (Gascoigne and Sartory, Reference Gascoigne and Sartory1974; Gascoigne, Reference Gascoigne1977; Jensen, Reference Jensen1991, Reference Jensen1993a, Reference Jensen1993c, Reference Jensen and Morton1993d, Reference Jensen1996a). All the teeth of the descending limb have been considered ‘used’, except the anteriormost one, called the leading or active tooth. However, the entire descending limb of the radula and its surrounding ascus-muscle are in use during feeding. It works as a rod for pushing the leading tooth through the algal cell wall (Jensen, Reference Jensen and Morton1993d). In C. phuketi, C. schuppi, and C. ghizoensis sp. nov., the descending limb of the radula remains attached to the ventral surface of the odontophore throughout its length and is only surrounded by a thin layer of longitudinal muscle ventrally and laterally.
The radular sac of sacoglossans contains only one or two formative teeth, called ‘ghost teeth’ (Gascoigne and Sartory, Reference Gascoigne and Sartory1974; Gascoigne, Reference Gascoigne1978) for their indistinct outline. In Cylindrobulla, the radular sac is long, and there are many formative teeth (present study, Table 1). Except for Cylindrobulla, the size of the teeth increases as the animal grows larger, and this is true for the base as well as the cusp of all studied species, although the rate of increase may differ for both parts (see, e.g., Jensen, Reference Jensen2014). In Cylindrobulla, all the teeth appear to be the same size, although an imperceptible increase may occur with age (see above and Table 1). The multiple tiny cusps are approximately the same length, with the median one somewhat longer, and it seems the cusps are attached directly to the broad, squarish base, rather than forming lateral denticles on the central cusp. Although there is a shallow depression on the backside of the teeth of Cylindrobulla, it does not look as if the cusps of the succeeding tooth will fit into this depression.
The sacoglossan odontophore is muscular, completely surrounding the radular sac and most of the ascending limb of the radula in most species. Dorso-ventral odontophore depressor muscles are used in combination with the dorsal septate muscle for sucking in the liquid contents of algal filaments, and radula suspending muscles attached to the ascending limb of the radula protract, align, and retract the ascending limb of the radula for piercing or cutting through algal cell walls (Jensen, Reference Jensen1993c, Reference Jensen and Morton1993d, Reference Jensen1994). The odontophore of Cylindrobulla is very long and narrow, and the musculature is rather weak. The radular sac protrudes from the posterior end of the pharynx, below the oesophagus. Radula suspending muscles run diagonally from ascending to descending limbs of the radula throughout the length. In other sacoglossans, the radula suspending muscles only connect the ascending limb with the anteriormost teeth of the descending limb, the posterior part being completely surrounded by the longitudinal ascus-muscle and separated from the odontophore by transverse muscles.
The separation of the ascending and descending limbs, either functionally by a layer of transverse muscles or anatomically by the detachment of most of the ascus muscle from the ventral surface of the pharynx, is another specialization connected to the piercing-sucking feeding method. This is characterized by separate ‘rasping’ and ‘sucking’ phases, which are associated with separate muscles for moving the ascending limb of radula (radula suspending muscles) and for moving the descending limb of the radula (longitudinal ascus-muscle) (Jensen, Reference Jensen1993a, Reference Jensen1993c, Reference Jensen and Morton1993d, Reference Jensen1994). Although the development of the different muscle groups differs among different genera and species, the separation seems absent in Cylindrobulla except in C. gigas, where some transverse muscles occur (Mikkelsen, Reference Mikkelsen1998: Figure 3A), and they are present in all other families. In some species, the separation seems to be formed mainly by a densely packed ascus sac (Jensen, Reference Jensen1992; Kohnert et al., Reference Kohnert, Brenzinger, Jensen and Schrödl2013).
Pharyngeal pouches are muscular bolsters attached to the pharynx postero-ventrally. They have narrow lumina with indistinct connection to the pharyngeal cavity below the oesophagus (see, e.g., Jensen, Reference Jensen1991, Reference Jensen1992, Reference Jensen and Morton1993d). Muscular pharyngeal pouches occur in all shelled genera except Cylindrobulla, though Mikkelsen (Reference Mikkelsen1998) indicated pouches for C. gigas. However, these appear to be the lateral pockets of the pharyngeal cavity, which are connected to the oesophagus without any special muscular padding (Mikkelsen, Reference Mikkelsen1998: Figure 3A). Pharyngeal pouches are also present in some species of most plakobranchacean families, and a short, collar-like pharyngeal pouch is seen in shelled Ascobulla and Oxynoe as well as in some non-shelled species of Costasiella and Plakobranchus (Jensen, Reference Jensen1980, Reference Jensen1992, Reference Jensen1993c; Jensen et al., Reference Jensen, Krug, Dupont and Nishina2014). Pharyngeal pouches are assumed to be lost secondarily in many plakobranchaceans and may also have been lost in Cylindrobulla. The function of the pharyngeal pouches is unknown. It has been suggested that they support the ascending limb of the radula during feeding (Jensen, Reference Jensen and Morton1993d).
Of course, there are also morphological and behavioural characteristics not associated with this specialized feeding method. These include (1) shell adductor muscle; (2) length of visceral nerve cord and number of ganglia; (3) body colouration; (4) cephalic tentacles or head shield; (5) penial armature; (6) burrowing behaviour. However, these characters are not restricted to the Sacoglossa and apparently have evolved in parallel in several other heterobranch groups (see, e.g., Kano et al., Reference Kano, Brenzinger, Nützel, Wilson and Schrödl2016; Brenzinger et al., Reference Brenzinger, Schrödl and Kano2021), and thus should be compared to these groups, but this is beyond the scope of the present study.
In conclusion, Cylindrobulla has a suctorial pharynx formed by the same muscles as in other sacoglossans, though weaker. The radula of Cylindrobulla seems unsuitable for piercing or cutting through algal cell walls in the manner refined in other sacoglossan families (Jensen, Reference Jensen and Morton1993d, Reference Jensen1994). The numerous, small, equally sized teeth may be used to graze on the sediment-encrusted rhizoids of Halimeda in a chain-saw manner. This is supported by the long, exposed part of the ascending limb of the radula, rather than the muscle-embedded radula of other sacoglossans, where only a few teeth at the tip of the odontophore are exposed. The long radula and high number of formative teeth indicate that teeth are replaced rapidly, possibly in response to feeding on a hard substrate (Krings et al., Reference Krings, Brütt and Gorb2022), although no distinct signs of wear have been detected in radulae of Cylindrobulla.
Acknowledgements
My participation in the Galathea 3 Expedition was funded by a grant from the Villum Kann Rasmussen Foundation to R.M. Kristensen. The field collecting efforts and companionship of the research group from the Zoological Museum (now the Natural History Museum of Denmark) during the stay at Ghizo Island were greatly appreciated.
Declaration of interest
None.
Ethics and approach
Not applicable.
Funding statement
This research has received no special funding.







 Open access
Open access