



Introduction
Cyst nematodes are obligate biotrophic endoparasites that penetrate the roots of their host and subsequently induce the formation of complex feeding structures. Cyst nematodes move intracellularly through the root cortex and thereafter inject a suite of secretory compounds into an initial feeding cell near or within the vascular tissue. The secreted compounds modify host cells leading to a partial breakdown of cell walls and subsequent merging of neighbouring cells (Sobczak and Golinowski, Reference Sobczak, Golinowski, Jones, Gheysen and Fenoll2011). This process results in a structure known as a syncytium that acts as a nutrient source for the nematode. The plant-derived auxin indole-3-acetic acid (IAA) is considered to play a pivotal role in syncytium formation of cyst nematodes (Goverse et al., Reference Goverse, Overmars, Engelbertink, Schots, Bakker and Helder2000). For example, the tomato auxin mutant DIAGEOTROPICA and the Arabidopsis double mutant of AUXIN1 (AUX1) and LIKE-AUX3 (LAX3) are significantly less susceptible to cyst nematode infection (Goverse et al., Reference Goverse, Overmars, Engelbertink, Schots, Bakker and Helder2000; Lee et al., Reference Lee, Chronis, Kenning, Peret, Hewezi, Davis, Baum, Hussey, Bennett and Mitchum2011). Additionally, the auxin responsive DR5 reporter has shown that syncytium formation is accompanied by a localized and transient auxin accumulation in Arabidopsis, as early as 18 hours post-infection (Goverse et al., Reference Goverse, Overmars, Engelbertink, Schots, Bakker and Helder2000; Karczmarek et al., Reference Karczmarek, Overmars, Helder and Goverse2004). Auxin accumulation seems to be present in the entirety of the feeding site and during this process both reporter gene and transcriptomic studies show the expression of differentially expressed auxin-related genes such as auxin response factors (ARFs), GRETCHEN HAGEN 3 (GH3) and auxin transport-related genes like PIN-FORMED (PIN) and LIKE-AUX1 (LAX) (Hutangura et al., Reference Hutangura, Mathesius, Jones and Rolfe1999; Grunewald et al., Reference Grunewald2009; Cabrera et al., Reference Cabrera, Bustos, Favery, Fenoll and Escobar2014; Tarek et al., Reference Tarek, Piya, Richard and Rice2014; Kyndt et al., Reference Kyndt, Goverse, Haegeman, Warmerdam, Wanjau, Jahani, Engler, de Almeida Engler and Gheysen2016). In addition, upregulation of auxin biosynthesis genes, such as tryptophan decarboxylase (TDC), AMIDASE (AMI) and YUCCA (YUC), is observed in host plant cells. From this, it can be inferred that auxin accumulation in syncytia likely arises, at least partially, through changes in local auxin biosynthesis and auxin transport in plant roots.
Auxin can be found in a multitude of different species ranging from plants to bacteria, fungi and even animals. Auxin biosynthesis is best studied in plants and bacteria. When considering auxin biosynthesis in plants, IAA is mainly produced from tryptophan (Trp), a product of the shikimate metabolic pathway (Maeda and Dudareva, Reference Maeda and Dudareva2012). The majority of de novo IAA is synthesized from amino acids in the aerial part of the plant as well as the meristematic zone of the root (Ljung et al., Reference Ljung, Hull, Celenza, Yamada, Estelle, Normanly and Sandberg2005). Four routes are known for the biosynthesis of IAA in plants. For Trp-dependent IAA synthesis these pathways are: (1) the indole-3-acetamide (IAM) pathway, (2) the indole-3-pyruvic acid (IPyA) pathway, (3) the tryptamine (TAM) pathway and (4) the indole-3-acetaldoxime (IAOx) pathway (Mano and Nemoto, Reference Mano and Nemoto2012). The presence of a non-Trp dependent biosynthesis pathway has been postulated but little is known hereof (Wang et al., Reference Wang, Chu, Tianying, Qian, Sun, Yuan, Xiong, Wang, Wang and Jiayang2015). The IAM biosynthesis pathway was thought to be a bacteria-specific pathway until IAM was found naturally occuring in all plant species and plant genes were identified for the conversion of IAM to IAA (Mano and Nemoto, Reference Mano and Nemoto2012; Gao et al., Reference Gao, Dai, Aoi, Takebayashi, Yang, Guo, Zeng, Hanchuanzhi, Kasahara and Zhao2020). The IPyA pathway is the main biosynthesis pathway in plants and the majority of all IAA is produced in this manner (Mashiguchi et al., Reference Mashiguchi, Tanaka, Sakai, Sugawara, Kawaide, Natsume and Hanada2011). Aside from the main biosynthesis pathway IAA can alternatively be produced through the TAM pathway and requires a 3-step conversion from Trp to IAA involving TDC, YUCCA and AAO genes (Mano and Nemoto, Reference Mano and Nemoto2012). The IAA intermediary compound IAOx has only been found in Brassicaceae species and as such it has been concluded that the IAOx biosynthesis pathway is exclusive to this family (Sugawara et al., Reference Sugawara, Hishiyama, Jikumaru, Hanada, Nishimura, Koshiba, Zhao, Kamiya and Kasahara2009). It is assumed that over 80% of the bacteria isolated from the rhizosphere are capable to synthesize IAA and at least 5 different pathways have been described for this (Patten and Glick, Reference Patten and Glick1996; Khalid et al., Reference Khalid, Tahir, Arshad and Zahir2004; Spaepen et al., Reference Spaepen, Vanderleyden and Remans2007). In bacteria, the conversion of Trp to IAA can occur through the same intermediary compounds that are found in plants (Spaepen and Vanderleyden, Reference Spaepen and Vanderleyden2011). The pathways themselves show similarity to those described in plants, although the order of intermediates can differ. An example of this is the conversion of IPyA to indole-3-acetaldehyde (IAD) by indole-3-pyruvate decarboxylase (ipdC) in bacteria (Spaepen and Vanderleyden, Reference Spaepen and Vanderleyden2011). IPyA and IAD are found in the IPyA and the TAM pathways, respectively.
In plants, auxin is a versatile molecule with a myriad of functions in both growth and development (Vert and Chory, Reference Vert and Chory2011; Weijers and Wagner, Reference Weijers and Wagner2016). In addition, auxin related processes have also been shown to play an important role during plant-parasite interactions and symbiotic relationships (Boivin et al., Reference Boivin, Fonouni-Farde and Frugier2016; Kohlen et al., Reference Kohlen, Ng, Deinum and Mathesius2018). For example, auxin plays a role in haustorium formation during infection by the hemiparasitic plant Phtheirospermum japonicum. Auxin is also found in animals and has been shown to be present in various gall inducing insects (Yamaguchi et al., Reference Yamaguchi, Tanaka, Hasegawa, Tokuda, Asami and Suzuki2012; Tanaka Reference TANAKA, Okada, ASAMI and SUZUKI2013) auxin in insect galls. IAA in these insects is produced de novo from tryptophan through the IAOx and IAD intermediates (Suzuki et al., Reference Suzuki, Yokokura, Ito, Arai, Yokoyama, Toshima, Nagata, Asami and Suzuki2014). Auxin is produced in a similar manner in silkworms (Bombyx mori), which are non-galling insects, thereby suggesting that it is not parasite exclusive (H. Suzuki et al., Reference Suzuki, Yokokura, Ito, Arai, Yokoyama, Toshima, Nagata, Asami and Suzuki2014). The synthesis of auxin by plant-parasitic nematodes to manipulate its host has been discussed as well. Already decades ago, using bioassays and chemical techniques, indications were found for the occurrence of auxin in plant-parasitic nematodes (Yu and Viglierchio, Reference Yu and Viglierchio1964; Viglierchio and Yu, Reference Viglierchio and Yu1968). However, these findings did not lead to more detailed studies in recent years using modern techniques to investigate whether plant-parasitic nematodes can produce auxins de novo to facilitate plant parasitism.
Here, we investigated the ability of plant-parasitic nematodes to synthesize auxin on a genomic, transcriptomic and metabolomic level. Metabolomic analyses using liquid chromatography-tandem mass-spectrometry demonstrated the presence of IAA in pre-parasitic juveniles of the potato cyst nematode Globodera pallida. Furthermore, it was shown that the amount of IAA measured in our analyses increased upon stimulation by plant root diffusate. Subsequently, potential homologues of known auxin biosynthesis genes were identified in a genomic screen of the G. pallida genome. Several candidate genes were discovered forming 5 potential pathways for the conversion of Trp to IAA based on both plant and microbial pathways described in other studies. The transcriptomic profiles of these candidate genes further supported their expression during infection of plant roots, indicating that they could play a role as potential auxin biosynthesis genes in plant parasitism. Genomic analysis of bacterivorous, fungivorous, entomopathogenic and animal parasitic nematodes, however, revealed a wide-spread occurrence of potential auxin biosynthesis pathways within the phylum Nematoda. Our data in conjunction with data in the literature indicates that auxin might have multiple functions in plant-parasitic nematodes. The potential role of auxin biosynthesis in plant parasitism as well as in endogenous (physiological) functions in plant-parasitic nematodes are discussed.
Materials and methods
In silico analysis of auxin biosynthesis gene identification and their expression in G. pallida
To identify potential auxin biosynthesis genes in G. pallida, a comparative genomics approach was used to create Hidden Markov Model (HMM) conserved protein models for each gene in the biosynthesis pathways of plants and bacteria. The amino acid sequences of known auxin biosynthesis genes were collected from 10 plant and 10 bacteria species (Supplementary Table S1). Genes from different species that facilitate the same step in auxin biosynthesis were grouped together. Each such group was then aligned using MAFFT. If only one protein sequence was found that facilitates a step in auxin biosynthesis it was supplemented to form a group with the top 5 hits from a BLASTp search of that target in the National Centre of Biotechnology (NBCI) database. Potential auxin biosynthesis genes were identified by performing a Hidden Markov Model (HMM) scan on the G. pallida genome (van Steenbrugge et al., Reference van Steenbrugge, van den Elsen, Holterman, Lozano-Torres, Putker, Thorpe, Goverse, Sterken, Smant and Helder2023) with each gene group of auxin biosynthesis genes in order to find patterns of homologous domains. Cygwin was used as Linux extension to perform the HMM scan using HMMER (v3.3.1). On these results a domain search was performed to select for potential G. pallida homologues that contain the same predicted protein domains as known auxin biosynthesis genes. CLC Main was used to perform this domain search using the Pfam database. This resulted in a list of potential homologues in G. pallida that have a similar structure and predicted protein domains as known auxin biosynthesis genes (Supplementary Table S2). To determine the expression of the auxin biosynthesis homologues the reads from the life-stage specific transcriptomes of G. pallida from Cotton et al. (Reference Cotton, Lilley, Jones, Kikuchi, Reid, Thorpe and Tsai2014) were mapped to the candidate genes. Expression is normalized to transcripts per million (TPM).
Nematode sample preparation
Cysts of G. pallida (population Rookmaker) originated from greenhouse culture and were long term stored at −80°C. A users batch was transferred from −80°C to 4°C until use. An amount of 100 000 cysts was taken from this user batch and hatched as described below to assess and quantify the potential production of IAA by G. pallida. Cysts were exposed to tap water or tomato root diffusate (TRD) at 20°C in the dark, to obtain water or TRD hatched second stage preparasitic juveniles for further use in our experiments in this study. The TRD stock had been stored for over 2 years at 4°C since it was collected from the roots of tomato plants in a hydroponic growth chamber with ½ Hoagland solution as nutrients.
Juveniles were purified with 35% sucrose solution with a layer of water on top in conical glass centrifuge tubes. After centrifugation at 2000 rpm in a Hereaus Multifuge 3 S-R for 3 minutes the juveniles collected at the interface of sucrose and water from which they were aspirated with a glass Pasteur pipet and transferred to a new centrifuge tube. Juveniles’ pellets were resuspended and washed 3 times in tap water, followed by a wash in sterile tap water, by centrifugation at 3000 rpm. Juveniles were transferred to a sterile 1.5 mL Eppendorf tube containing 4 µL of 10% Tween-20, to prevent juveniles from sticking to the tube wall, and washed with sterile tap water 3 more times.
To surface sterilize the juveniles, they were incubated in the following solutions consecutively: 0.5% (w/v) streptomycin–penicillin solution for 20 minutes, 0.1% (w/v) ampicillin–gentamycin solution for 20 minutes, sterile tap water for 5 minutes, 0.1% (v/v) chlorhexidin solution for 3 minutes. Finally, juveniles were washed at least 3× in sterile tap water before their surface sterility was checked. To do so, a small aliquot of the juvenile suspension after surface sterilization was transferred to a B5 medium plate (containing 3.29 g/L Gamborg B5 including Vitamins and salts, 20 g/L Sucrose, pH 6.2; 1.5 % Bacto-agar, Difco) and was incubated in the dark at 20°C for 1 week. After this 1 week, the plate was checked for bacterial and fungal growth. Besides this, the viability of the nematodes was visually assessed to ensure that the amount of dead or ruptured nematodes was minimal (<1%).
A series of nematodes samples consisting of an increasing number of hatched pre-parasitic juveniles was prepared containing 3915, 8100, 39 150, 81 000 and 233 000 J2 individuals. Measurements of IAA (as well as SA and IPR) in water and TRD-hatched nematodes were done with 33 000 nematodes. For each of these measurements, 3 biological replicates were used with the nematodes collected all from the same user batch of cysts. For the collection of nematode secretion, 33 000 nematodes were used for water-hatched samples and 34 666 nematodes for TRD-hatched samples.
Collecting nematode secretions
TRD-hatched J2’s of G. pallida (population Rookmaker) were collected and surface sterilized as described above. Sterility check, vitality check and counting were performed as described above. Equal number of J2’s were incubated in 0.5 mL of either sterile tap water or TRD in sterile 1.5 mL Eppendorf tubes separately on a rotator (Agar scientific, AR 384) at 20°C in the dark for 24 hours. After 24 hours, a small amount of the suspension was visually inspected for nematode vitality again. The J2 were spun down and the supernatant, containing the secretions, was transferred to a new sterile 1.5 mL and stored at −80°C until auxin measurement by UPLC-MS/MS.
Extraction of phytohormones
Juveniles were lysed in a buffer according to Holterman et al. (Reference Holterman, van der Wurff, van den Elsen, van Megen, Bongers, Holovachov, Bakker and Helder2006) with some modifications. Lysis took place at 20°C and 1200 rpm in a Thermomixer (Eppendorf, Hamburg, Germany) during approximately 7 hours. Lysed juvenile samples were spun down at max. speed for 3 minutes and the supernatant transferred to a new sterile 1.5 mL Eppendorf tube. Lysed juvenile samples were aliquoted into 3 technical replicates for UPLC-MS/MS and stored at −80°C until use.
Per sample, 1 mL of 100% methanol (MeOH) containing stable isotope-labeled internal standards (IS, Supplementary Table S6) at an end concentration of 100 nM per standard per sample. Samples were extracted and measured as previously described (Schiessl et al., Reference Schiessl, Lilley, Lee, Tamvakis, Kohlen, Bailey and Thomas2019; Gühl et al., Reference Gühl, Holmer, Xiao, Shen, Wardhani, Geurts, van Zeijl and Kohlen2021). All solvents were evaporated in a speed vacuum system (SPD121P, ThermoSavant, Hastings, UK) at RT and the residue stored at −20°C until further analysis.
Detection and quantification of phytohormones by liquid chromatography-tandem mass-spectrometry
Sample residues were dissolved in 100 µL of acetonitrile/water (acid hormones) or MeOH/water (CKs) (0.1% formic acid) (20:80, v/v), and filtered through a 0.45 mm Minisart SRP4 filter (Sartorius, Goettingen, Germany). Analyses of plant growth regulators was performed by comparing retention times and mass transitions with those of unlabeled standards (Supplementary Table S6) using a Waters XevoTQs mass spectrometer equipped with an electrospray ionization source coupled to an Acquity UPLC system (Waters, Milford, USA) as previously described (Schiessl et al., Reference Schiessl, Lilley, Lee, Tamvakis, Kohlen, Bailey and Thomas2019; Gühl et al., Reference Gühl, Holmer, Xiao, Shen, Wardhani, Geurts, van Zeijl and Kohlen2021). Chromatographic separations were conducted on an Acquity UPLC BEH C18 column (100 mm; 2.1 mm; 1.7 mm; Waters, USA) by applying an acetonitrile/water (0.1% formic acid) or methanol/water (0.1% formic acid) gradient. The column was operated at 50°C with a flow rate of 0.5 mL•min−1. The column was equilibrated for 30 min using either solvent composition at the start of a run. The acetonitrile/water (0.1% formic acid) gradient started from 20% (v/v) acetonitrile, increasing to 70% (v/v) acetonitrile in 17 min. To wash the column, the water/acetonitrile gradient was increased to 100% (v/v) acetonitrile in a 1.0 min gradient, which was maintained for 1.0 min before going back to 20% acetonitrile using a 1.0 min gradient, prior to the next run. The methanol/water (0.1% formic acid) gradient started from 5% (v/v) methanol, increasing to 70% (v/v) methanol in 17 min. To wash the column, the water/methanol gradient was increased to 100% (v/v) methanol in a 1.0 min gradient, which was maintained for 1.0 min before going back to 5% methanol using a 1.0 min gradient, prior to the next run. The sample injection volume was 3/5 µL (acids/cytokinin, respectively). The mass spectrometer was operated in positive and negative electrospray ionization mode when required. Cone and desolvation gas flows were set to 150 and 1000/800 l•h−1, respectively. The capillary voltage was set at 3.5/3.0 kV, the source temperature at 150°C, and the desolvation temperature at 550°C. The cone voltage was optimized for each standard compound using the IntelliStart MS Console (Waters, Milford, USA). Argon was used for fragmentation by collision-induced dissociation. Multiple reaction monitoring (MRM) was used for quantification (Gühl et al., Reference Gühl, Holmer, Xiao, Shen, Wardhani, Geurts, van Zeijl and Kohlen2021). Parent–daughter transitions for the different (stable isotope labelled) compounds were set using the IntelliStart MS Console. MRM transitions, cone voltage and collision energy selected for compound identification and quantification are shown in Supplementary Table S6. To determine sample concentrations, a 10-point calibration curve was constructed for each compound ranging from 1 µM to 190 pM and each dilution also contained a known amount of an appropriate deuterium-labelled internal standard.
Results
Auxin is present in Globodera pallida and increases in response to root exudates
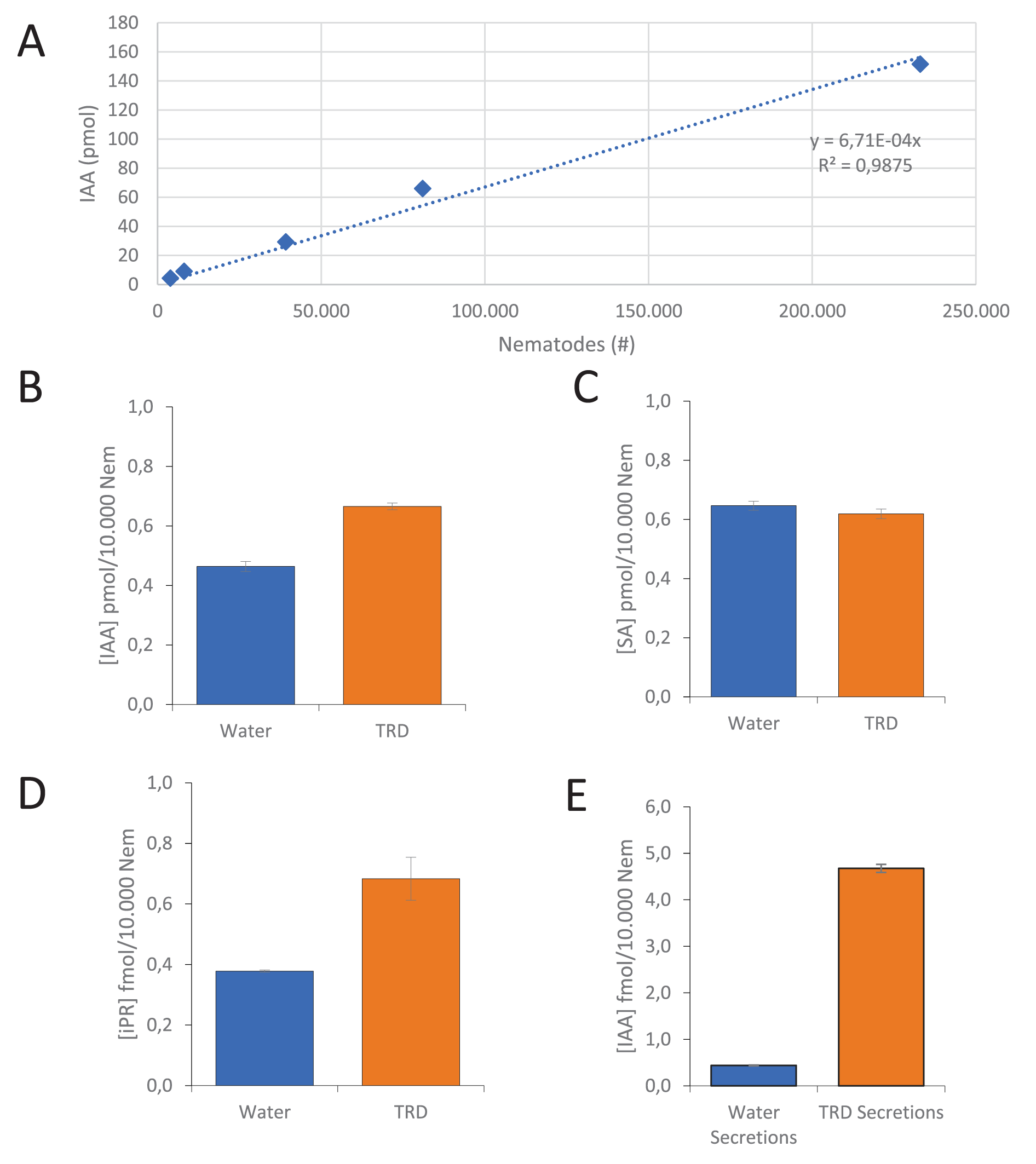
To assess and quantify the potential production of IAA by G. pallida, mass-spectrometry was used to identify this compound in infective second stage juveniles (pre-parasitic J2 stage). To ensure reliable auxin quantification under contamination-free and controlled conditions, nematodes were checked for contamination and viability upon collection (see M&M for details). Next, a series of 5 samples with increasing numbers of juveniles in the range of 4000–230 000 was prepared for mass-spectrometry, which revealed that IAA is present in pre-parasitic water-hatched second stage juveniles of G. pallida in detectable levels at 0.7 fmol per nematode (Figure 1A). In addition, this quantity of IAA linearly correlates with the number of nematodes in the samples (Pearson correlation, R 2 = 0.99). From these data, we conclude that IAA is already present in pre-parasitic J2 which are not yet exposed to host root diffusate and such plant stimuli are apparently not required to trigger auxin biosynthesis.
Auxin, cytokinin and salicylic acid quantities produced by pre-parasitic second stage juveniles of G. pallida as measured through MRM-UPLC-MS/MS. (A) Correlation of the concentration of IAA measured in an increasing number of infective nematodes obtained from a single batch of water-hatched pre-parasitic juveniles (J2). Effect of exposure to Tomato Root Diffusate (TRD) on the auxin (B), salicylic acid (C) and cytokinin (D) concentration shown as the average of 3 technical replicates of equal amounts of infective nematodes as compared to the water control. Auxin measurement is a representative dataset out of 3 independent biological replicates, while salicylic acid and cytokinin represent one biological sample per treatment. (E) Concentration of IAA measured in water-induced and TRD induced nematode secretions (one biological sample per treatment). Significance was determined using a Student’s t-test. *P < 0.05; ***P < 0.001.

Figure 1 Long description
Graph A shows a line graph with IAA concentration (pmol) on the y-axis and nematode count on the x-axis, ranging from 0 to 250,000. The graph displays a positive correlation with the equation y equals 6.71 times 10 superscript negative 4 x and R squared equals 0.9875. Graph B shows a bar graph comparing IAA concentration (pmol per 10,000 nematodes) in water and TRD treatments, with TRD showing higher values. Graph C presents a bar graph of SA concentration (pmol per 10,000 nematodes) in water and TRD treatments, with similar values for both. Graph D illustrates a bar graph of IPR concentration (fmol per 10,000 nematodes) in water and TRD treatments, with TRD showing higher values. Graph E shows a bar graph of IAA concentration (fmol per 10,000 nematodes) in water secretions and TRD secretions, with TRD secretions showing significantly higher values.
In order to determine whether the amount of auxin is influenced by the perception of a host plant, G. pallida cysts were also hatched in the presence of tomato root diffusate (TRD). In water-hatched juveniles a concentration was measured of 0.46 pmol IAA per 10 000 nematodes with a standard error of 1.7*10−2 (Figure 1B). In contrast, for TRD-hatched juveniles a concentration was measured of 0.67 pmol IAA per 10 000 nematodes with a standard error (SE) of 1.1*10−2 (Figure 1B). Thus, a comparative analysis of water-hatched and TRD-hatched juveniles demonstrated a 43% increase in the amount of IAA present in TRD-hatched nematodes compared to the water-hatched control with a significance of P = 5.7*10−4 (Student’s t-test). To exclude a plant-derived IAA contribution by TRD, we analysed its IAA content with mass spectrometry as well. No IAA was detected in any of the used TRD samples. For comparison, the cytokinin isopentenyl riboside (iPR) was included in our measurements as its presence in the cyst nematode Heterodera schachtii has been reported previously (Siddique et al., Reference Siddique, Radakovic, De La Torre, Chronis, Novák, Ramireddy and Holbein2015). Cytokinin was indeed detected in G. pallida juveniles at a level of 0.38 fmol/10 000 nematodes (SE = 3.2*10−3) for water-hatched nematodes and 0.68 fmol/10 000 nematodes (SE = 7.1*10−2) for TRD-hatched nematodes (Figure 1D). Similar to IAA, cytokinin increased by 81% upon hatching with TRD with a significance of P = 1.3*10−2 (Student’s t-test). Interestingly, iPR is present at lower concentrations than IAA by a factor thousand. The hormone salicylic acid (SA) was additionally included and was present in G. pallida juveniles at a level of 0.65 pmol/10 000 nematodes (SE = 0.015) for water-hatched nematodes (Figure 1C). However, no statistically significant change could be detected upon exposure to TRD with a concentration of 0.62 pmol/10 000 nematodes (SE = 0.016) for TRD hatched nematodes (Figure 1C), suggesting not a uniform increase of metabolism, but a more selective effect of TRD on a specific set of metabolites. Altogether, the data indicate the stimulation of auxin and cytokinin biosynthesis in pre-parasitic J2 stages of G. pallida as a response to plant stimuli.
Subsequently, it was assessed whether auxin is not only produced but also secreted by infective juveniles and for that purpose G. pallida secretions were collected. TRD-hatched juveniles were surface sterilized and thoroughly washed in several washing steps as described for water-hatched juveniles before an equal number was suspended in either water or TRD for 24 hours. Thereafter, the IAA concentration was measured in the nematodes as well as the liquid in which the nematodes were suspended. Mass spectrometry revealed that the auxin present in the nematodes was at comparable levels as measured before around 0.20 pmol/10 000 J2 (SD = 2.1*10−3). Our data obtained from one biological replicate and 3 technical replicates showed that auxin is present in the secretions of both water and TRD hatched nematodes (Figure 1E). Secreted auxin levels appear to be a factor thousand lower than endogenous auxin concentrations inside the nematodes at 0.44 fmol/10 000 J2 (SE = 2.4*10−3) with a significance of P = 3.1*10−9 (Student’s t-test) (Figure 1E). However, a more than tenfold increase in concentration is observed in the secretion of J2’s exposed to TRD at 4.67 fmol/10 000 J2 (SD = 2.8*10−3) compared to secretions of J2’s exposed to (tap) water with a significance of P = 9.9*10−9 (Student’s t-test). Overall, it can be concluded that G. pallida produces IAA of which the concentration increases within nematodes in response to host plant stimuli, and preliminary data also indicate that IAA is secreted and that IAA secretion is stimulated by root diffusate.
Identification of candidate auxin biosynthesis genes in G. pallida
The presence of auxin in pre-parasitic second-stage juveniles indicates the existence of an auxin biosynthesis pathway in G. pallida. However, no information is available yet about how auxin biosynthesis occurs in plant-parasitic nematodes. Auxin biosynthesis is best studied in plants and bacteria and involves multiple pathways. To identify potential auxin biosynthesis genes in G. pallida, a comparative genomics approach was used including 10 plant species and 10 bacteria species (Supplementary Table S1) to create Hidden Markov Model (HMM) conserved protein models for each gene in the biosynthesis pathways of plants and bacteria (63 protein sequences distributed over 11 different auxin biosynthesis genes). These HMM models were then used to scan the G. pallida genome to identify auxin biosynthesis candidate genes. The genes from the collection were grouped together for each enzyme in the plant and bacterial auxin biosynthesis pathway. In total, for the conversion of tryptophan to IAA, a total of 191 genes were identified in the G. pallida genome with 97 of these being annotated as cytochrome P450s.
Following the initial screening of the 191 genes, a comprehensive analysis utilizing the Pfam database was undertaken to ascertain the presence of catalytic domains associated with these genes. This analysis revealed that 47 of the 191 genes contain functional protein domains that correspond to their plant or bacteria counterpart. These 47 genes cover a total of 10 enzymatic steps in auxin biosynthesis (Figure 2C), each characterized by protein domains corresponding to their putative function (Figure 2A). It is noted that genes identified through the Hidden Markov Model (HMM) scan, and simultaneously found to contain expected functional protein domains, are categorized here as homologous. The majority of these genes are homologues of the Cytochrome P450 (CYP) gene family, which is not surprising considering its role with metabolites in general and the size and prevalence of this gene family in all (multi)cellular organisms (Nelson, Reference Nelson1999) (Figure 2B). Interestingly, putative homologous sequences for enzymes in all 4 major auxin biosynthesis routes from plants could be found in the genome of G. pallida (Figure 2C). Homologous sequences for the enzymes AMIDASE (AMI), YUCCA (YUC), Indole-3-acetaldehyde oxidase (AAO) and NITRILASE (NIT), all involved in the last step of their respective auxin biosynthesis route in plants, could be identified. From these, the largest number of homologues could be found for the AMI gene (8). While various YUC (5) and CYP (24) genes were identified, it was not possible to differentiate between the different YUC or CYPs in the biosynthesis pathway (Figure 2C). For example, the YUC responsible for the conversion of TAM to IAD or the YUC catalysing the reaction of IPyA to IAA both lead to the identification of the same potential homologues in G. pallida. In addition, these protein families have members that are not involved in auxin biosynthesis, which hampers a further differentiation within these families. Overall, 47 candidate auxin biosynthesis genes were identified in G. pallida on the basis of sequence similarity and functional domain prediction, providing evidence for the presence of potential auxin biosynthesis pathways in G. pallida (Supplementary Table S2).
Identification of potential auxin biosynthesis genes and possible pathways in G. pallida. (A) Overview of the different auxin biosynthesis gene homologues found in the G. pallida genome (D383, van Steenbrugge et al., Reference van Steenbrugge, van den Elsen, Holterman, Lozano-Torres, Putker, Thorpe, Goverse, Sterken, Smant and Helder2023) after a Hidden Markov Model search followed by a functional domain analysis. Potential homologues are named after their original search query and possess identical functional domains as predicted through Pfam with the indicated E-value for both the HMM scan and Pfam analysis (highest and lowest found). (B) Distribution of the number of auxin biosynthesis homologues detected in G. pallida as percentage of the 47 found genes. (C) Potential auxin biosynthesis pathways in G. pallida based on mapping of the identified homologues on the 5 main pathways in plants and bacteria. Precursor and end products are displayed with a green border and intermediate compounds with an Orange border. Blue shows the homologues identified in G.pallida that potentially facilitate the displayed conversion. Dotted line and question mark denote a hypothetical expected conversion step in nematode auxin biosynthesis.

Figure 2 Long description
The image consists of three parts related to auxin biosynthesis in Globodera pallida. A shows a table listing homologues, HMM E-value, Pfam E-value and domain for various enzymes like AAO, AMI, TAA, NIT, YUC, CYP, TDC and ipdC. B is a pie chart illustrating the distribution of auxin biosynthesis homologues, with CYP at 51 percent, AMI at 17 percent, YUC at 11 percent, TDC at 6 percent, ipdC at 4 percent, TAA at 7 percent and both AAO and NIT at 2 percent each. C is a diagram depicting potential auxin biosynthesis pathways, showing conversions between compounds like Trp, TAM, IAOx, IAN, IAM, IPyA, IAD and IAA, with enzymes such as CYP, TDC, TAA, YUC, AMI, NIT, AAO and ipdC facilitating these conversions. The diagram includes a hypothetical conversion step marked with a question mark.
Our bioinformatic pipeline was not only able to detect candidate auxin biosynthesis genes belonging to the last step of the 4 auxin biosynthesis routes (i.e. IAOx, TAM, IAM and IPyA (Figure 2C)), but it could also identify homologues that could facilitate the conversion of tryptophan to the intermediate compounds thereby forming a complete pathway. It is noted that for the IAM pathway a candidate gene for the last step (AMI) was detected, but that the homologue for the iaaM gene was absent in G. pallida. Several candidate iaaM genes were discovered in the HMM scan but did not have a matching predicted functional domain in our Pfam analysis. The iaaM gene facilitates the conversion of tryptophan to IAM in bacteria. IAM occurs also in plants while no iaaM gene has been detected and it is assumed that in plants the conversion of tryptophan to IAM is catalysed by another gene than in bacteria (Kasahara, Reference Kasahara2016). Similar to plants, it is here postulated that the absence of the iaaM gene in G. pallida does not necessarily obstruct the conversion of tryptophan via IAM into IAA. Potential auxin biosynthesis genes for conversion between intermediate compounds from bacterial pathways were also found. Two putative homologues for indole-3-pyruvate decarboxylase (ipdC) have been identified which, in bacteria, produce the auxin intermediate indole-3-acetaldehyde (IAD) from the auxin intermediate (IPyA) (Spaepen and Vanderleyden, Reference Spaepen and Vanderleyden2011). This enzyme could account for an additional biosynthesis route next to the 4 pathways known in plants by following the IPya pathway and half way branching off into the TAM pathway. In conclusion, our genomic analyses indicate that in theory 5 auxin biosynthesis pathways could be active in G. pallida and may contribute to the conversion of tryptophan into auxin as described (Figure 2C).
Expression and upregulation of potential auxin biosynthesis genes in different life stages
To investigate whether the auxin biosynthesis candidate genes detected in the G. pallida genome are also expressed, transcript abundance of the 47 candidate auxin biosynthesis genes was analysed in silico. Therefore, their transcription dynamics were retrieved from a publicly available expression atlas of various G. pallida life stages (i.e. egg, J2, 7dpi, 14dpi, 21dpi, 28dpi and 35 dpi (Cotton et al., Reference Cotton, Lilley, Jones, Kikuchi, Reid, Thorpe and Tsai2014)). For all 47 candidate genes, transcripts were found thereby confirming the expression of the selected putative auxin biosynthesis genes (Supplementary Table S3). Hierarchical clustering shows that the expression patterns of auxin biosynthesis candidate genes in the parasitic stages are more similar to each other than to eggs and pre-parasitic juveniles (Figure 3). In addition, the expression patterns of the genes in males form a clearly separated group, in line with the distinctive migratory lifestyle of males as compared to immobile sedentary females that develop on the roots during late stages of parasitism like dpi 21, 28 and 35. Within these groupings, some heterogeneity is observed: one male sample deviates from the other 2 male samples and at 3 time points the parasitic stages do not cluster together, indicating the experimental and/or biological variation. Similarly, heterogeneity is observed in expression within a group of similar genes. For example, YUC, TAA and CYP genes do not all cluster together based on the expression profile in different stages. Altogether, it is concluded that the candidate auxin biosynthesis genes are expressed during the life cycle of G. pallida and expression levels vary between life stages.
Identification and hierarchical clustering of transcripts from the 47 potential auxin biosynthesis genes present in the G. pallida genome. Gene expression levels are shown as log2 transcripts per million (TPM) values in a heatmap. Transcript data were retrieved from the gene expression atlas of Cotton et al. (Reference Cotton, Lilley, Jones, Kikuchi, Reid, Thorpe and Tsai2014). Samples obtained from different life stages and time points (days post infection (dpi)) as well as the gene transcripts are ordered according to a hierarchical clustering The household gene cyclic AMP-dependent transcription factor ATF-4 (g00617) is included as a visual reference (marked HOUSEKEEPING).

Figure 3 Long description
The heatmap displays gene expression levels of 47 potential auxin biosynthesis genes, organized by hierarchical clustering. The x-axis represents different life stages and time points, including egg, J2, 7 days post infection (dpi), 14 dpi, 21 dpi, 28 dpi and 35 dpi. The y-axis lists the gene identifiers, such as GpalD383g07189.t1 and CYP6GpalD383g02154.t1. Expression levels are shown as log transcripts per million values, with a color gradient indicating the intensity of expression from low to high. The housekeeping gene GpalD383_g00617.t1 is included as a visual reference. The clustering shows distinct expression patterns among parasitic stages, eggs and pre-parasitic juveniles, with notable differences in male samples. Some heterogeneity is observed within gene groups, such as YUC, TAA and CYP genes, which do not cluster together consistently across stages.
Next, the correlation between expression patterns of candidate genes within the same pathway was studied across different life stages. Therefore, candidate genes were grouped together within the same pathway based on similarities in their expression pattern using a Pearson correlation (Supplementary Table S4). The expression profile of genes within each pathway with the highest Pearson correlation to each other were selected as the most promising candidates for that pathway (Figure 4). The potential IAOx pathway consists of 2 CYP450P genes and one NIT gene where the NIT1 gene and CYP8 cluster close together in terms of expression (Figure 4A). Although CYP9 differs significantly from the other 2 in terms of expression level, the changes in expression follow a similar trend over life stages as NIT1 and CYP8. The potential TAM pathway consists of 3 genes of which AAO1 and YUC2 share a similar expression pattern, while the expression profile of TDC3 deviates (Figure 4B). The 3 genes in the TAM pathway seem to be lowly expressed during the egg and J2 phases, whereas specifically TDC3 is induced during infection with a sharp drop at 28 dpi which recovers slightly at 35 dpi. The potential IAM pathway lacks a possible gene for the conversion of Trp to IAM and as such only the expression of the potential AMI genes is considered, which facilitates the conversion of IAM to IAA. The different potential AMI genes display a variety in expression levels, with AMI1 showing relatively high transcripts per million (TPM) values compared to the other 7 AMI genes (Figure 4C). The most similar expression patterns between potential auxin biosynthesis genes are found within the IPyA pathway (Figure 4D). The expression levels of both YUC3 and TAA3 are low in eggs and pre-parasitic second-stage juveniles after which a steep rise is observed during the first 14 days followed by a decline (Figure 4D). Additionally, the potential IPyA pathway contains the branching point for what we refer to here as the IAD pathway and consists of TAA1, ipdC1 and AAO1 (Figure 4E). The selected genes in this pathway display a repetitive rise and decline in expression across the various life stages. The correlation between the expression of ipdC1 and AAO1 is one of the lowest (0.39) together with AAO1 and TDC3 (0.37) (Figure 4F). These correlations further show that genes in the IAOx and IPyA pathway correlate better in terms of expression when compared to the TAM and IAD pathway. Overall, this analysis shows that there is a level of similarity in expression patterns within potential auxin biosynthesis pathways, especially for the putative IAOx and IPyA pathways.
Expression pattern of potential auxin biosynthesis genes in various life stages of G. pallida relative to the household gene ATF-4 (g00617). Candidate genes with the highest Pearson correlation are shown for an auxin biosynthesis pathway based on similarity in transcript abundance. (A) the IAOx pathway, (B) the TAM pathway, (D) the IPyA pathway and (E) the IAD pathway. (C) For the IAM pathway only AMI genes were found and their expression pattern is displayed in various life stages of G. pallida. (F) Highest positive Pearson correlations between potential auxin biosynthesis genes per pathway.

Figure 4 Long description
The image contains six graphs labeled A to F, each depicting relative gene expression across different pathways. A) IAOx Pathway: The graph shows relative gene expression for NIT1 (g01223), CYP9 (g02859) and CYP8 (g02612) across various life stages, including egg, J2, 7dpi, 14dpi, 21dpi, 28dpi, 35dpi and male. B) TAM Pathway: The graph displays relative gene expression for AAO1 (g00915), YUC2 (g04073) and TDC3 (g00915) across the same life stages. C) IAM Pathway: The graph illustrates relative gene expression for AMI1 (g00632), AMI2 (g11146), AMI3 (g04644), AMI4 (g09834), AMI5 (g10146), AMI6 (g10908), AMI7 (g05406) and AMI8 (g06053) across the life stages. D) IPyA Pathway: The graph shows relative gene expression for YUC3 (g05685) and TAA3 (g00987) across the life stages. E) IAD Pathway: The graph presents relative gene expression for AAO1 (g02956), ipdC1 (g13700) and TAA1 (g00987) across the life stages. F) Correlation Data: The table shows correlation values between genes within the same pathway, including IAOx, TAM, IPyA and IAD pathways. Correlation values are provided for NIT1, CYP8, CYP9, AAO1, YUC2, TDC3, YUC3, TAA3, ipdC1 and TAA1.
Auxin biosynthesis genes are widespread within the phylum Nematoda
To explore how widespread the potential auxin biosynthesis genes and possible pathways are in the phylum Nematoda, HMM scans were conducted with the same models that were used to identify candidate auxin biosynthesis genes in G. pallida (Figure 2A). The HMM scans were carried out on a comprehensive panel of plant-parasitic, animal parasitic and free-living nematode proteomes ranging across all clades in the phylum for which genomes were available. By mapping these candidate biosynthesis genes on the known auxin biosynthesis pathways in plants and bacteria as a model, it was determined whether a species could have the required genes to form a hypothetical auxin biosynthesis pathway. Considering the best hits from the HMM scans, it is noteworthy that the E-values of the HMM hits for all nematode species closely approximate those of the candidate biosynthesis genes in G. pallida that were identified through our Pfam analysis (Supplementary Table S5, Figure 2A). This suggests that G. pallida is not an exception within the phylum Nematoda with regard to the occurrence of potential auxin biosynthesis genes. The HMM scan reveals that all nematode species tested contain candidate genes for potential auxin biosynthesis pathways (Figure 5). In total, 17 of the 30 nematode species have potential candidate biosynthesis genes for all 5 biosynthesis pathways. An exception among the plant-parasitic nematodes being the Meloidogyne species which seem to miss potential genes for the TAM and IAD pathways. This is due to the absence of potential candidates for the AAO gene that catalyses the last step in the auxin biosynthesis; the conversion of IAD to IAA. It is noted, that this gene seems to be absent in several animal parasitic nematodes as well. Soboliphyme baturini seems to be the least-equipped nematode species to synthesize auxin, and possesses only 2 potential auxin biosynthesis pathways; the IAM and IAOx pathway.
Overview of 5 potential auxin biosynthesis pathways in plant-parasitic (red highlight), free-living (blue highlight) and animal parasitic (yellow highlight) nematodes based on the detection of candidate auxin biosynthesis genes in genome sequences using an HMM scan. Green slots indicate the putative presence of a complete auxin biosynthesis pathway based on the identification of potential auxin biosynthesis genes for each gene within that pathway. Orange slots indicate that a potential pathway might be incomplete due to the absence of at least one candidate gene within that pathway (which gene is mentioned in text).

Figure 5 Long description
Table showing auxin biosynthesis pathways in various nematode species, categorized as plant parasitic, free-living and animal parasitic. Green indicates complete pathways; orange indicates incomplete pathways due to missing genes. See long description.","Long_Description":"The table presents an overview of auxin biosynthesis pathways across different nematode species, divided into plant parasitic, free-living and animal parasitic categories. The nematode species are listed in the left column, while the auxin biosynthesis pathways (IAM, IPyA, IAOx, TAM, IAD) are listed across the top row. Green slots indicate the presence of a complete auxin biosynthesis pathway, while orange slots indicate an incomplete pathway due to missing genes. For plant parasitic nematodes, species like Meloidogyne arenaria and Meloidogyne enterolobii show incomplete pathways due to the absence of the AAO gene. Free-living nematodes such as Acrobeloides nanus have complete pathways. Among animal parasitic nematodes, Soboliphyme baturini shows incomplete pathways with missing TAA and YUC genes. Other species like Onchocerca volvulus and Romanomermis culicivorax also have incomplete pathways due to missing AAO genes.
To study whether candidate auxin biosynthesis genes in G. pallida might have been recruited via horizontal gene transfer from e.g., bacteria or plants—a typical feature shown for specific genes involved in plant parasitism—a BLAST search was performed. This BLAST search was carried out for the nucleotide sequences of the 47 potential auxin biosynthesis enzymes against the NCBI database of plants, bacteria, nematodes, fungi and non-redundant protein sequences. The highest similarities for the 47 candidate genes from G. pallida were found with genes in other nematode species. BLAST searches cross-kingdom against plants, bacteria, fungi, and non-redundant protein sequences databases did result in hits, but with a significantly lower similarity, indicating that horizontal gene transfer from other kingdoms is an unlikely scenario. Moreover, analysing the candidate genes from G. pallida that might facilitate the last step in auxin biosynthesis (AMI, NIT, YUC, AAO) did not result in the detection of any predicted signal peptide for secretion, another hallmark for genes involved in plant parasitism. So, no evidence was found for evolutionary adaptations typical for plant parasitism.
In conclusion, our data indicate that potential auxin biosynthesis pathways are wide-spread within the phylum Nematoda and are present in plant-parasitic, bacterivorous, fungivorous, entomopathogenic and animal parasitic nematodes, indicating a broader role in nematode biology.
Discussion
Here, we investigated the ability of plant-parasitic nematodes to synthesize auxin on a genomic, transcriptomic and metabolomic level. We show that IAA can be detected in G. pallida using mass-spectrometry and that its concentration increases upon contact with plant stimuli. Preliminary data suggests that auxin is secreted and that exposure of nematodes to root exudates may result in a tenfold increase of auxin in secretions when compared to water hatched juveniles (Figure 1E). Potential auxin biosynthesis genes were identified using a collection of known homologues in bacteria and plants, leading to the identification of candidate genes for all steps in 5 potential auxin biosynthesis pathways (4 plant and 1 bacterial pathway). The expression of these candidate genes was verified in an RNA-seq dataset and varied for most genes between developmental stages. Our bioinformatic analysis revealed that potential auxin biosynthesis genes are widespread within the phylum Nematoda and occur in free-living, plant- and animal-parasitic nematodes.
The presence of IAA in its biologically active form in G. pallida is supported by previous data on plant-parasitic nematodes. Several decades ago the presence of auxin has been reported in egg masses and second-stage juveniles of the root-knot nematode M. incognita and larvae of M. hapla (Yu and Viglierchio, Reference Yu and Viglierchio1964; Viglierchio and Yu, Reference Viglierchio and Yu1968). This was shown with elementary verification methods such as chromatography, the Avena first internode test to determine if it is a growth substance, as well as Ehrlich and Salkowski chemical tests to determine whether it is an indole compound (Yu and Viglierchio, Reference Yu and Viglierchio1964; Viglierchio and Yu, Reference Viglierchio and Yu1968). Another preliminary report used mass-spectrometry to detect IAA and conjugated forms of IAA in secretions of M. incognita and H. schachtii as well as biosynthesis intermediates in their lysates (De Meutter et al., Reference De Meutter, Tytgat, Prinsen, Gheysen, Van Onckelen and Gheysen2005). In this research we have demonstrated the presence of IAA in G. pallida as well, but now supported by the identification of endogenous auxin biosynthesis candidate genes. Remarkably, our bioinformatic approach has shown that auxin biosynthesis candidate genes are widespread throughout the phylum Nematoda. Potential auxin biosynthesis genes are present in plant-parasitic nematodes but also in animal parasitic and non-parasitic free-living nematodes (Figure 5). With these data, it is tempting to speculate that these nematodes too might produce auxin. However, candidate genes for auxin synthesis steps could not always be found in each nematode species. This may be due to the absence of a particular biosynthesis route of which the AAO gene is the most absent auxin biosynthesis gene amongst the investigated species and is absent in all studied Meloidogyne species (Figure 5). AAO acts as an aldehyde oxygenase for the conversion of IAD to IAA in the last step of the TAM pathway. However, recently an aldehyde dehydrogenase (AO) was found to catalyse the same reaction of IAD to IAA in Pseudomonas syringae (McClerklin et al., Reference McClerklin, Lee, Harper, Nwumeh, Jez and Kunkel2018). In addition, AO mediated auxin biosynthesis contributed to virulence of P. syringae. AOs are conserved throughout evolution and are mainly known for their function in detoxifying endogenous aldehydes (Shortall et al., Reference Shortall, Djeghader, Magner and Soulimane2021). Moreover, AOs are studied in C. elegans to evaluate the effect of alcohol and alcoholism (Alaimo et al., Reference Alaimo, Davis, Song, Burnette, Grotewiel, Shelton, Pierce-Shimomura, Davies and Bettinger2012). The IAA synthesis properties of AO give rise to the possibility that not AAO is used in Meloidogyne species but instead AO catalyses the last step of the TAM pathway. A short BLAST search further supports this through the presence of several putative AO genes in M. graminicola and M. enterolobii.
Upregulation of the expression of potential auxin biosynthesis genes during infection was found in all 5 putative auxin biosynthesis pathways in G. pallida. However, changes in the expression levels of potential auxin biosynthesis pathways could not directly be associated with the rise and decline of auxin signalling during feeding cell development. Previous reports have shown that auxin signalling peaks during the initial phases of the infection (Karczmarek et al., Reference Karczmarek, Overmars, Helder and Goverse2004). Auxin signalling reaches its maximum level between 1 and 5 dpi and hereafter stays at a steady level for several days (Goverse et al., Reference Goverse, Overmars, Engelbertink, Schots, Bakker and Helder2000). At 10 dpi and later time points no signal can be detected (Karczmarek et al., Reference Karczmarek, Overmars, Helder and Goverse2004). Also considering the upregulation of individual candidate auxin biosynthesis genes no correlations are observed with reported levels of auxin signalling during the initial phases of syncytia development. Apart from pitfalls in such comparisons, there are more aspects to consider. The production rate of a metabolic compound in a pathway, such as IAA, is in part determined by its rate limiting step (Zhao, Reference Zhao2012). When the expression of a non-rate limiting enzyme is increased this does not necessarily affect the quantity of the produced compound. As such, if the auxin biosynthesis gene in G. pallida is not the rate limiting step, then its upregulation is a poor indicator of the production rate of IAA. Therefore, the expression of the biosynthesis genes should always be taken in consideration together with the other enzymes in the same pathway. The production of auxin is further complicated by substrate availability and the presence of multiple possible biosynthesis routes. While the substrates and intermediate compounds of all these auxin biosynthesis pathways can be converted to IAA, it is not necessarily their predetermined fate as the conversion to compounds other than IAA is possible. For example, tryptamine is an intermediary compound of the TAM auxin biosynthesis pathway but can alternatively be converted into serotonin by tryptamine 5-hydroxylase in rice (Kang et al., Reference Kang, Kang, Lee and Back2007). Additionally, IAOx and IAN of the IAN pathway are intermediates in the production of camalexin in Arabidopsis, which acts as a phytoalexin to deter bacterial and fungal pathogens (Mucha et al., Reference Mucha, Heinzlmeir, Kriechbaumer, Strickland, Kirchhelle, Choudhary and Kowalski2019). As such, it is clear that these metabolites should not be viewed as only intermediary compounds of auxin but as intermediary compounds for a diverse number of processes. The multifaceted nature of these compounds highlights the complexity of IAA metabolism. This further complicates investigations into the role of both the auxin biosynthesis genes as well as potential intermediary compounds in the biosynthesis of auxin in G. pallida. Taken together, it can be concluded that the interpretation of expression patterns of biosynthesis genes should be taken into the context of the entire pathway (e.g. rate limiting steps) and that auxin intermediates may have more functions than their conversion into auxin.
Auxin has long been thought to play a crucial role during nematode infection. During nematode infection auxin is suspected to be involved in processes such as hypertrophy, cell wall ingrowths and cell cycle activation (de Almeida Engler et al., Reference de Almeida Engler, De Vleesschauwer, Burssens, Jr Celenza, Inzé, Van Montagu, Engler and Gheysen1999; Gheysen and Mitchum, Reference Gheysen and Mitchum2011). The accumulation of auxin is thought to mainly be the result of manipulation of auxin transport and the induction of plant-derived local biosynthesis (Grunewald et al., Reference Grunewald2009; Kyndt et al., Reference Kyndt, Goverse, Haegeman, Warmerdam, Wanjau, Jahani, Engler, de Almeida Engler and Gheysen2016; Suzuki et al., Reference Suzuki, Kanno, Abril-Urias, Seo, Escobar, Tsai and Sawa2022). Our data suggest, however, that plant-parasitic nematodes might have an additional third option to increase auxin levels in plant roots through the secretion of nematode-derived auxin. The quantity of IAA measured in second stage juveniles of G. pallida (∼10−16 mol/J2) greatly exceeds that of a single plant cell as Arabidopsis root apex protoplast cells contain ∼10−19 mol IAA (Petersson et al., Reference Petersson, Johansson, Kowalczyk, Makoveychuk, Wang, Moritz, Grebe, Benfey, Sandberg and Ljung2009). This indicates that the nematode contains sufficient IAA to be biologically relevant. Preliminary data on the secretion of IAA suggest that the excreted quantity per juvenile is roughly 3 orders of magnitude lower than what is present in the body of the nematode (Figure 1E). However, these values stay within the range of ∼10−19 mol/J2 and thereby in the range of biological relevance. Taken together, a biologically relevant amount of IAA has been found in G. pallida, that may contribute to the development of syncytia via a yet unknown secretion mechanism.
Similar to auxin, the concentration of cytokinin (IPR) in second stage juveniles increases upon exposure to plant exudates (Figure 1D), but is 3 orders of magnitudes lower in G. pallida than the measured auxin quantities. Cytokinin has also been detected in H. schachtii and M. incognita and reported to have a secretion rate of ∼10−18 mol/J2/hr (De Meutter et al., Reference Meutter, Jan, Witters, Gheysen, Van Onckelen and Gheysen2003). This secretion rate is relatively high considering the low amount of cytokinin in second stage juveniles detected in this study (10−18 mol, Figure 1D), but may in part be explained by the measurement of more types of cytokinins (De Meutter et al., Reference Meutter, Jan, Witters, Gheysen, Van Onckelen and Gheysen2003). These secreted amounts of cytokinins were assumed to be physiologically relevant, which has been confirmed in a study in which silencing of isopentenyltransferase, a cytokinin biosynthesis gene, reduced the expansion of syncytia and reproduction of H. schachtii (Siddique et al., Reference Siddique, Radakovic, De La Torre, Chronis, Novák, Ramireddy and Holbein2015). The biosynthesis of the other measured phytohormone salicylic acid was not stimulated by plant exudates. The function of salicylic acid in G. pallida is not clear, but considering its occurrence and role in bacteria, fungi and malaria parasites (Matsubara et al., Reference Matsubara, Aonuma, Kojima, Tahara, Andrabi, Sakakibara and Nagamune2015; Mishra and Baek, Reference Mishra and Baek2021), this metabolite has most likely a function not directly related to plant parasitism but this needs further investigation.
Our data indicate that the ability of G. pallida to synthesize auxin is not a unique adaptation of plant-parasitic nematodes within the phylum Nematoda. Candidate auxin biosynthesis genes in G. pallida likely evolved through vertical evolution rather than horizontal gene transfer from bacteria or plants. This is supported by 2 main observations. First, potential auxin biosynthesis genes are widespread across nematodes, unlike cellulases, which are thought to have been acquired via horizontal gene transfer and are largely restricted to plant-parasitic species (Danchin et al., Reference Danchin, Rosso, Vieira, de Almeida-engler, Coutinho, Henrissat and Abad2010; Annelies et al., Reference Annelies, Jones and Danchin2011), with the recent exception of cellulases in Pristionchus nematodes likely acquired from slime molds (Han et al., Reference Han, Sieriebriennikov, Susoy, Wen-Sui, Igreja, Dong, Berasategui, Witte and Sommer2022). Second, G. pallida auxin biosynthesis genes more closely resemble those of bacterivorous, fungivorous and animal-parasitic nematodes than their equivalents in plants, bacteria or fungi.
Together, these findings suggest that auxin has an endogenous role in nematode growth and development. Indeed, mass-spectrometry confirmed auxin in Caenorhabditis elegans (Zhou et al., Reference Zhou, Zhang and Butcher2019), where isotope-labelled studies showed IAA acts as an intermediate in ascaroside production. Ascarosides regulate behaviours such as hermaphrodite repulsion, olfactory plasticity, and dauer formation (Ludewig and Schroeder, Reference Ludewig and Schroeder2013). Beyond this, auxin appears to extend lifespan in C. elegans (Loose and Ghazi, Reference Loose and Ghazi2021), consistent with observations that auxin promotes proliferation in mammalian cell lines (Cernaro et al., Reference Cernaro, Medici, Leonello, Buemi, Kohnke, Villari, Santoro and Buemi2015).
Auxin has also been detected in arthropods, including spiders, mites, crustaceans and millipedes, at concentrations often exceeding those in plants, sometimes by up to 50-fold (Tokuda et al., Reference Tokuda, Suzuki, Fujita, Matsuda, Adachi-Fukunaga and Elsayed2022). Although its exact function in arthropods is unclear, its presence across sexes and developmental stages suggests a general role in their life cycle. Altogether, auxin emerges as a molecule with diverse functions across taxa, of which acting as a plant hormone is only one.
To investigate the role of auxin secretion by G. pallida in syncytium induction, several experimental approaches can be considered. Analogous to classical effector studies, the spatial distribution of auxin within the nematode body could be examined, for instance, using antibodies to assess whether elevated auxin concentrations occur in secretory glands. In addition, the spatial and temporal dynamics of candidate auxin biosynthesis gene expression could be analysed at near-cellular resolution through RNA tomography (Ebbing et al., Reference Ebbing, Vértesy, Betist, Spanjaard, Junker, Berezikov, van Oudenaarden and Korswagen2018). Upregulation of these genes in secretory glands would suggest a potential role of auxin in external interactions. Functional studies could include knockdowns of candidate biosynthesis genes to assess their impact on the total auxin pool and infection success. However, given auxin’s involvement in both parasitic and non-parasitic processes, distinguishing between effects on endogenous functions and syncytium development may prove challenging. An alternative strategy would be to target genes associated with auxin secretion. A preliminary BLAST search revealed the presence of putative auxin exporters in the G. pallida genome. While PIN-type exporters appear absent, several potential ATP-binding cassette subfamily B (ABCB) transporters were identified. In plants, ABCB/MDR/PGP proteins are established auxin exporters that localize to the plasma membrane in a non-polar fashion, maintaining homeostasis and ensuring stable auxin distribution (Cho and Cho, Reference Cho and Cho2013). Investigating the expression, localization and functional disruption of these predicted ABCB transporters may therefore provide valuable insights into the contribution of auxin to syncytium formation. Overall, the study of auxin in G. pallida parasitism is more complex than the analysis of canonical effector proteins, owing to the various endogenous roles that auxins might play within nematodes.
In summary, our data demonstrate that G. pallida is able to synthesize and secrete IAA, and that plant stimuli seem to enhance these processes. The auxin concentrations appear to be biologically relevant, since the secreted quantities are in the same order of magnitude as biologically relevant concentrations in plant cells (Petersson et al., Reference Petersson, Johansson, Kowalczyk, Makoveychuk, Wang, Moritz, Grebe, Benfey, Sandberg and Ljung2009). However, the widespread occurrence of the auxin biosynthesis candidates within the phylum Nematoda also suggests an endogenous role for auxin outside of parasitism possibly related to vitality and ascaroside synthesis (Ludewig and Schroeder, Reference Ludewig and Schroeder2013; Loose and Ghazi, Reference Loose and Ghazi2021). Therefore, knocking down auxin biosynthesis and studying the potential auxin secretion mechanisms in e.g., secretory gland cells will be crucial in elucidating its relevance for the development of syncytia induced by G. pallida.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182025101480.
Author’s contribution
J.B. and A.G. and M.O. contributed to conception and design of the study. H.O. prepared samples for mass spectrometry. W.K. performed the mass spectrometry. A.B. provided the genomic, transcriptomic and proteomic sequences. A.B. and M.O. performed the HMM scan and subsequent gene selection. J.vS. and M.O. did the gene expression analysis. M.O. wrote the manuscript with input from J.B. and A.G. W.K. and H.O. provided input for the writing of the material and methods. W.K., J.vS., A.B. and H.O. provided insightful feedback for the revision of the manuscript.
Financial support
This research received no specific grant from any funding agency, commercial or not-for-profit sectors. Part of this work conducted by H.O. and A.B. was funded by TTW project 16873 of the Dutch Science Foundation NWO.
Competing interests
The authors declare there are no conflicts of interest.
Ethical standards
Not applicable







 Open access
Open access