



1. Introduction
Locomotion in complex media, such as colloids, polymer solutions and granular materials, is characterised by the interaction of swimmers with nonlinear rheological properties and non-continuum effects (Arratia Reference Arratia2022). In polymeric liquids, swimmers like cells, bacteria and micro-robots can take advantage of liquid properties including shear-rate-dependent viscosity (Fu, Wolgemuth & Powers Reference Fu, Wolgemuth and Powers2009; Qiu et al. Reference Qiu, Lee, Mark, Morozov, Münster, Mierka, Turek, Leshansky and Fischer2014; Riley & Lauga Reference Riley and Lauga2017; Han et al. Reference Han, Shields IV, Bharti, Arratia and Velev2020) and elasticity involving hoop stress and normal stress differences (Normand & Lauga Reference Normand and Lauga2008; Keim, Garcia & Arratia Reference Keim, Garcia and Arratia2012; Pak et al. Reference Pak, Zhu, Brandt and Lauga2012; Binagia & Shaqfeh Reference Binagia and Shaqfeh2021; Rogowski et al. Reference Rogowski, Ali, Zhang, Wilking, Fu and Kim2021; Datta et al. Reference Datta2022; Kroo et al. Reference Kroo, Binagia, Eckman, Prakash and Shaqfeh2022). Propulsion gaits that stretch polymers in shear flow, thereby reducing the local viscosity or inducing elasticity in the medium, can enhance swimming efficiency under these conditions (Fu, Shenoy & Powers Reference Fu, Shenoy and Powers2010; Wróbel et al. Reference Wróbel, Lynch, Barrett, Fauci and Cortez2016). These effects also enable propulsion schemes such as reciprocal swimming (Keim et al. Reference Keim, Garcia and Arratia2012; Qiu et al. Reference Qiu, Lee, Mark, Morozov, Münster, Mierka, Turek, Leshansky and Fischer2014; Han et al. Reference Han, Shields IV, Bharti, Arratia and Velev2020), that are otherwise not possible in Newtonian liquids (Purcell Reference Purcell1977; Lauga Reference Lauga2011).
For media with heterogeneous microstructures (Balin et al. Reference Balin, Zöttl, Yeomans and Shendruk2017; Zhang, Li & Ardekani Reference Zhang, Li and Ardekani2018), significant non-continuum effects emerge when the characteristic length scale of the swimmer is comparable to that of a medium’s constituents. For instance, in dense polymer solutions, a nanometre-scale beating flagellum may locally stretch out polymer coils, creating a polymer-depleted channel (Martinez et al. Reference Martinez, Schwarz-Linek, Reufer, Wilson, Morozov and Poon2014). This structural modification results in a significantly lowered propulsive force when the flagellum revisits the wake. While subtle at the nanoscale, such non-continuum effects may become more pronounced for locomotion in complex media with larger constituent particles, such as granular materials.
Locomotion in granular media is a particularly rich problem, with examples ranging from sandfish lizards that undulate through sand (Maladen et al. Reference Maladen, Ding, Li and Goldman2009; Goldman Reference Goldman2014), to worms that burrow in soil via peristalsis (Dorgan Reference Dorgan2018), and southern sand octopuses that fluidise the substrate to alter the resistance of submerged sand (Montana, Finn & Norman Reference Montana, Finn and Norman2015). These systems are challenging to understand because granular matter can behave either like a flowing liquid (Forterre & Pouliquen Reference Forterre and Pouliquen2008; Xiao et al. Reference Xiao, Ottino, Lueptow and Umbanhowar2017, Reference Xiao, Hruska, Ottino, Lueptow and Umbanhowar2018) or like a load-bearing solid (Majmudar et al. Reference Majmudar, Sperl, Luding and Behringer2007; Bi et al. Reference Bi, Zhang, Chakraborty and Behringer2011), depending on the conditions. The solid-like behaviour can arise from a jamming transition, which can be induced by isotropic pressure or shear (Liu & Nagel Reference Liu and Nagel1998; Bi et al. Reference Bi, Zhang, Chakraborty and Behringer2011; Behringer & Chakraborty Reference Behringer and Chakraborty2018), and has important implications for locomotion. For instance, particles ahead of an intruder in a static bed often remain jammed, effectively enlarging the penetrating object (Buchholtz & Pöschel Reference Buchholtz and Pöschel1998; Brzinski III et al. Reference Brzinski III, Mayor and Durian2013; Müller et al. Reference Müller, Formella and Pöschel2014; Kang et al. Reference Kang, Feng, Liu and Blumenfeld2018; Harrington, Xiao & Durian Reference Harrington, Xiao and Durian2020). At large stresses, collective particle rearrangements dissipate elastic energy, giving rise to elasto-plastic behaviour (Schall & van Hecke Reference Schall and van Hecke2010; Xiao et al. Reference Xiao, Ivancic and Durian2020, Reference Xiao, Zhang, Yang, Ivancic, Ridout, Riggleman, Durian and Liu2023). Most studies on locomotion, therefore, model granular materials as elasto-plastic or perfectly plastic continua, for which reduced-order approaches such as resistive force theories are applicable (Askari & Kamrin Reference Askari and Kamrin2016; Agarwal et al. Reference Agarwal, Karsai, Goldman and Kamrin2021, Reference Agarwal, Goldman and Kamrin2023).
Granular materials containing highly deformable, cohesive or nearly frictionless particles can display pronounced viscoelasticity, as in food products, powders, cosmetics and crude oil. Their flow behaviour differs markedly from that of rigid model materials, such as dry glass or plastic beads and poppy seeds, which are typically used in laboratory studies. Granular hydrogels provide a particularly interesting cohesive model system because they combine very low friction coefficients (
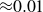 ${\approx} 0.01$
) with high elastic deformability (elastic moduli
${\approx} 0.01$
) with high elastic deformability (elastic moduli
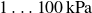 $1\ldots 100\,\text{kPa}$
) (Ashour et al. Reference Ashour, Trittel, Börzsönyi and Stannarius2017; Shakya et al. Reference Shakya, van der Gucht and Dijksman2024). Due to their high water content, liquid capillary bridges can form between hydrogel particles, inducing cohesive forces. These effects lead to distinct time-dependent behaviours in shear flows (Dijksman & Mullin Reference Dijksman and Mullin2022; Wang et al. Reference Wang, Farmani, Dijksman, Lübeck, Speck and Stannarius2022; Farmani et al. Reference Farmani, Wang, Stannarius and Dijksman2025) and silo discharge (Stannarius et al. Reference Stannarius, Martinez, Börzsönyi, Bieberle, Barthel and Hampel2019; Harth et al. Reference Harth, Wang, Börzsönyi and Stannarius2020; Wang et al. Reference Wang, Fan, Pongó, Börzsönyi, Hidalgo and Stannarius2023). Because granular hydrogels combine viscoelasticity, reminiscent of polymeric liquids, with reconfigurable heterogeneous packing, swimming in such media is of particular interest. Beyond fundamental physics, it may also be relevant for applications such as microrobots navigating among blood cells.
$1\ldots 100\,\text{kPa}$
) (Ashour et al. Reference Ashour, Trittel, Börzsönyi and Stannarius2017; Shakya et al. Reference Shakya, van der Gucht and Dijksman2024). Due to their high water content, liquid capillary bridges can form between hydrogel particles, inducing cohesive forces. These effects lead to distinct time-dependent behaviours in shear flows (Dijksman & Mullin Reference Dijksman and Mullin2022; Wang et al. Reference Wang, Farmani, Dijksman, Lübeck, Speck and Stannarius2022; Farmani et al. Reference Farmani, Wang, Stannarius and Dijksman2025) and silo discharge (Stannarius et al. Reference Stannarius, Martinez, Börzsönyi, Bieberle, Barthel and Hampel2019; Harth et al. Reference Harth, Wang, Börzsönyi and Stannarius2020; Wang et al. Reference Wang, Fan, Pongó, Börzsönyi, Hidalgo and Stannarius2023). Because granular hydrogels combine viscoelasticity, reminiscent of polymeric liquids, with reconfigurable heterogeneous packing, swimming in such media is of particular interest. Beyond fundamental physics, it may also be relevant for applications such as microrobots navigating among blood cells.
This study presents experiments with a scallop-inspired swimmer driven by two counter-rotating blades that flap reciprocally. The swimmer moves within a bed of hydrogel spheres and, as will be shown, generates propulsion only within a specific frequency range of flapping. This behaviour contrasts with swimmers in beds of frictional polystyrene spheres (Xiao et al. Reference Xiao, Torres, Sack and Pöschel2025). Both the swimming direction and its frequency dependence differ from those observed in the rigid and frictional particles.
2. Experimental methods and materials
The experimental set-up for the scallop-inspired swimmer in granular media is shown in figure 1 based on our previous study (Xiao et al. Reference Xiao, Torres, Sack and Pöschel2025). The swimmer consists of a pair of counter-rotating, square wings (
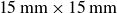 $15\,\text{mm}\times \text{15}\,\text{mm}$
, thickness
$15\,\text{mm}\times \text{15}\,\text{mm}$
, thickness
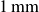 $1\,\text{mm}$
), 3D printed from resin. Each wing is mounted on a vertical carbon-fibre rod (diameter
$1\,\text{mm}$
), 3D printed from resin. Each wing is mounted on a vertical carbon-fibre rod (diameter
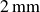 $2\,\text{mm}$
), supported by ball bearings, with the rods separated by
$2\,\text{mm}$
), supported by ball bearings, with the rods separated by
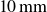 $10\,\text{mm}$
. A servo motor drives one rod directly, while the other is coupled through a pair of identical gears, ensuring symmetric counter-rotation of the wings. A Cartesian coordinate system is defined with
$10\,\text{mm}$
. A servo motor drives one rod directly, while the other is coupled through a pair of identical gears, ensuring symmetric counter-rotation of the wings. A Cartesian coordinate system is defined with
 $x$
the lateral direction,
$x$
the lateral direction,
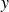 $y$
the swimming direction (figure 1 inset) and
$y$
the swimming direction (figure 1 inset) and
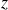 $z$
the vertical axis pointing upward. The wings flap between
$z$
the vertical axis pointing upward. The wings flap between
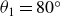 $\theta _1=80^\circ$
(opened) and
$\theta _1=80^\circ$
(opened) and
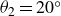 $\theta _2=20^\circ$
(closed), respectively.
$\theta _2=20^\circ$
(closed), respectively.
Experimental set-up for swimming in granular media. The apparatus consists of a pair of wings (
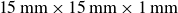 $15\,\text{mm}\times \text{15}\,\text{mm}\times 1\,\text{mm}$
) operating in a reservoir (
$15\,\text{mm}\times \text{15}\,\text{mm}\times 1\,\text{mm}$
) operating in a reservoir (
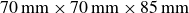 $70\,\text{mm} \times 70\,\text{mm} \times 85\,\text{mm}$
) filled with granular particles. The inset shows a photograph of the hydrogel particles.
$70\,\text{mm} \times 70\,\text{mm} \times 85\,\text{mm}$
) filled with granular particles. The inset shows a photograph of the hydrogel particles.

To prepare the hydrogel granulate, we started from dry powder sieved to a narrow size distribution
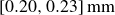 $[0.20, 0.23]\,\text{mm}$
. After swelling in distilled water, the particles assumed diameters of
$[0.20, 0.23]\,\text{mm}$
. After swelling in distilled water, the particles assumed diameters of
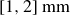 $[1,2]\,\text{mm}$
. The hydrogel particles were removed from the water bath with a strainer and placed on paper towels to drain excess surface water. Residual water formed liquid bridges between neighbouring particles, rendering the material slightly cohesive. The liquid bridge-induced cohesion and the deformation behaviours of bulk packing are described in Harth et al. (Reference Harth, Wang, Börzsönyi and Stannarius2020), Wang et al. (Reference Wang, Farmani, Dijksman, Lübeck, Speck and Stannarius2022) and Farmani et al. (Reference Farmani, Wang, Stannarius and Dijksman2025). The inset of figure 1 shows the bed surface. For comparison, we also used dry polystyrene particles with an average diameter of
$[1,2]\,\text{mm}$
. The hydrogel particles were removed from the water bath with a strainer and placed on paper towels to drain excess surface water. Residual water formed liquid bridges between neighbouring particles, rendering the material slightly cohesive. The liquid bridge-induced cohesion and the deformation behaviours of bulk packing are described in Harth et al. (Reference Harth, Wang, Börzsönyi and Stannarius2020), Wang et al. (Reference Wang, Farmani, Dijksman, Lübeck, Speck and Stannarius2022) and Farmani et al. (Reference Farmani, Wang, Stannarius and Dijksman2025). The inset of figure 1 shows the bed surface. For comparison, we also used dry polystyrene particles with an average diameter of
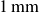 $1\,\text{mm}$
and a similar density to the hydrogel particles, as examined in greater detail in Xiao et al. (Reference Xiao, Torres, Sack and Pöschel2025).
$1\,\text{mm}$
and a similar density to the hydrogel particles, as examined in greater detail in Xiao et al. (Reference Xiao, Torres, Sack and Pöschel2025).
The granulate was contained in a cubic box with a cross-section of
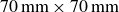 $70\,\text{mm} \times 70\,\text{mm}$
, filled to a height of
$70\,\text{mm} \times 70\,\text{mm}$
, filled to a height of
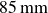 $85\,\text{mm}$
. The swimmer’s mid-
$85\,\text{mm}$
. The swimmer’s mid-
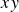 $xy$
plane was thus positioned at a depth of
$xy$
plane was thus positioned at a depth of
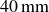 $40\,\text{mm}$
. The box rested on two linear rails, each supported by two air bearings. The total moving mass was
$40\,\text{mm}$
. The box rested on two linear rails, each supported by two air bearings. The total moving mass was
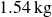 $1.54\,\text{kg}$
, including the reservoir and rails. This is far below the load-bearing capacity of the air bearings (4
$1.54\,\text{kg}$
, including the reservoir and rails. This is far below the load-bearing capacity of the air bearings (4
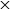 $\times$
4.5 kg), allowing frictionless motion in the
$\times$
4.5 kg), allowing frictionless motion in the
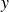 $y$
direction.
$y$
direction.
The displacement of the reservoir was recorded by a linear position Hall encoder and converted to the relative displacement of the swimmer,
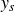 $y_s$
. A load cell measured the force
$y_s$
. A load cell measured the force
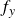 $f_y$
acting on the swimmer in the
$f_y$
acting on the swimmer in the
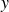 $y$
direction. Neither the load cell nor the motor was mechanically coupled to the moving granular bed. Swimming was tested in both hydrogel and polystyrene media over a range of cycle periods
$y$
direction. Neither the load cell nor the motor was mechanically coupled to the moving granular bed. Swimming was tested in both hydrogel and polystyrene media over a range of cycle periods
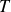 $T$
, always beginning with the closed angle
$T$
, always beginning with the closed angle
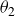 $\theta _2$
.
$\theta _2$
.
3. Locomotion in granular media of hydrogel and polystyrene particles
Figure 2 shows the swimmer displacement
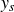 $y_s$
as a function of scaled time
$y_s$
as a function of scaled time
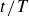 $t/T$
(dashed curves), averaged over two repetitions, with the positive direction (forward) defined in figure 1. In polystyrene granulate, the swimmer exhibits persistent locomotion in the
$t/T$
(dashed curves), averaged over two repetitions, with the positive direction (forward) defined in figure 1. In polystyrene granulate, the swimmer exhibits persistent locomotion in the
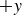 $+y$
direction, consistent with our previous results (Xiao et al. Reference Xiao, Torres, Sack and Pöschel2025). In contrast, locomotion in hydrogel granulate occurs in the
$+y$
direction, consistent with our previous results (Xiao et al. Reference Xiao, Torres, Sack and Pöschel2025). In contrast, locomotion in hydrogel granulate occurs in the
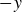 $-y$
direction and depends strongly on the flapping rate: at
$-y$
direction and depends strongly on the flapping rate: at
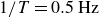 $1/T=0.5\,\text{Hz}$
, the net displacement vanishes, with the swimmer returning to
$1/T=0.5\,\text{Hz}$
, the net displacement vanishes, with the swimmer returning to
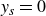 $y_s=0$
after each cycle; at
$y_s=0$
after each cycle; at
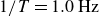 $1/T=1.0\,\text{Hz}$
, the swimmer develops persistent locomotion in the
$1/T=1.0\,\text{Hz}$
, the swimmer develops persistent locomotion in the
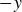 $-y$
direction, opposite to that observed in polystyrene granulate; and at
$-y$
direction, opposite to that observed in polystyrene granulate; and at
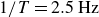 $1/T= 2.5\,\text{Hz}$
, the net displacement has vanished again. For each case, measurements from two individual runs are shown with lighter colours. Results of repeated runs coincide, except for
$1/T= 2.5\,\text{Hz}$
, the net displacement has vanished again. For each case, measurements from two individual runs are shown with lighter colours. Results of repeated runs coincide, except for
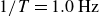 $1/T=1.0\,\text{Hz}$
, where a slight quantitative difference between the two runs exists. This indicates a slight dependence of the propulsion on the initial packing condition, which may have a stronger history dependence for the cohesive hydrogel granulates than for polystyrene.
$1/T=1.0\,\text{Hz}$
, where a slight quantitative difference between the two runs exists. This indicates a slight dependence of the propulsion on the initial packing condition, which may have a stronger history dependence for the cohesive hydrogel granulates than for polystyrene.
Swimmer displacement as a function of time (dashed curves) for several flapping periods:
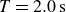 $T=2.0\,\text{s}$
(blue),
$T=2.0\,\text{s}$
(blue),
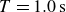 $T=1.0\,\text{s}$
(purple),
$T=1.0\,\text{s}$
(purple),
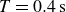 $T=0.4\,\text{s}$
(red). For comparison, the black curve shows data for polystyrene granulate at
$T=0.4\,\text{s}$
(red). For comparison, the black curve shows data for polystyrene granulate at
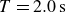 $T=2.0\,\text{s}$
. For each case, symbols, connected by solid lines, indicate the displacement at the end of each cycle, averaged over two repetitions, and the corresponding displacement in each repetition is shown as a solid curve with lighter colour.
$T=2.0\,\text{s}$
. For each case, symbols, connected by solid lines, indicate the displacement at the end of each cycle, averaged over two repetitions, and the corresponding displacement in each repetition is shown as a solid curve with lighter colour.

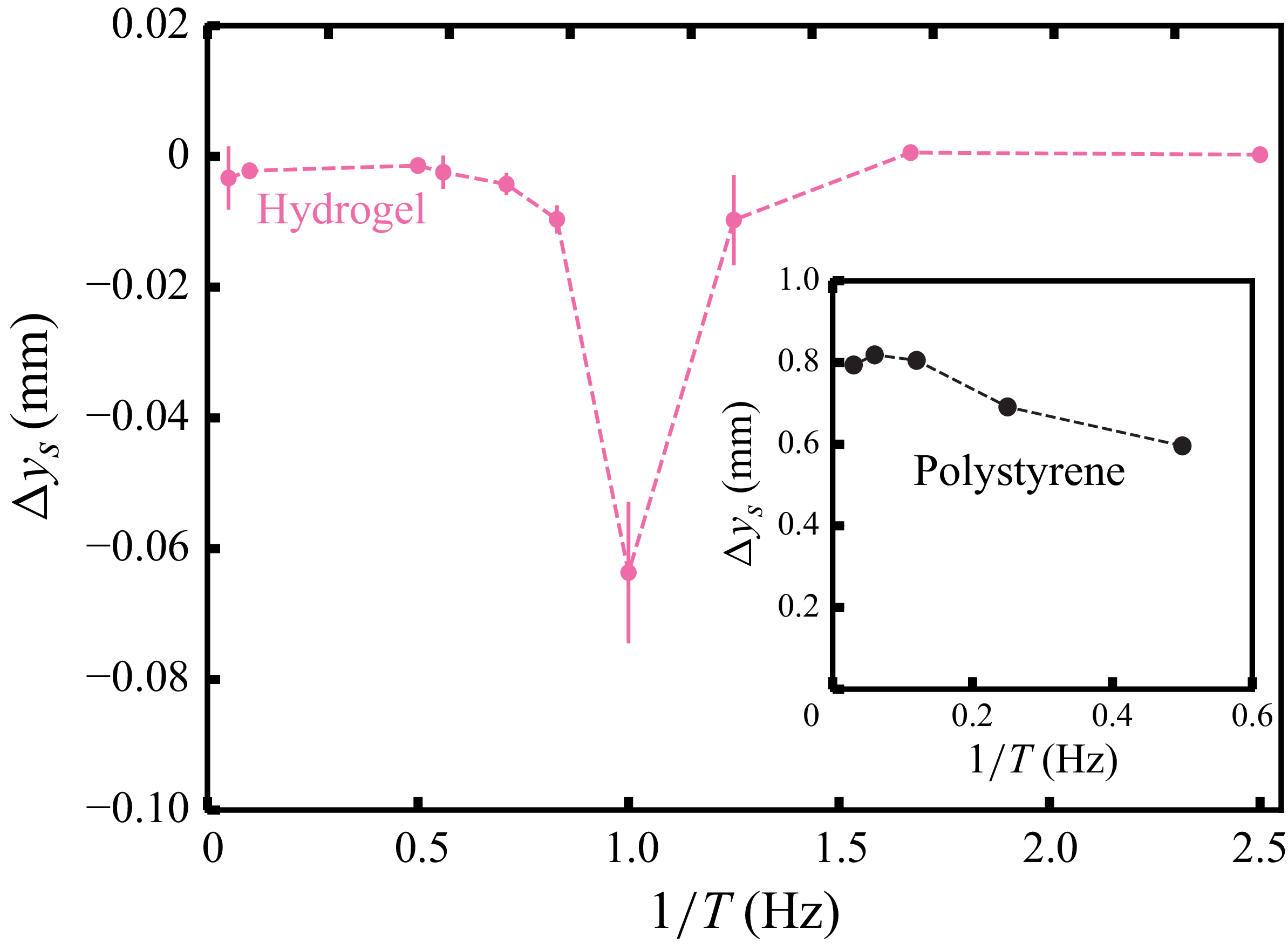
Figure 3 shows the net displacement of the swimmer per cycle,
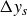 $\Delta y_s$
, as a function of the flapping frequency. These values are averages of the slope of
$\Delta y_s$
, as a function of the flapping frequency. These values are averages of the slope of
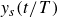 $y_s(t/T)$
at the end of each cycle (symbols in figure 2), quantified by linear fits over at least five successive data points in the linear regime after the initial relaxation (
$y_s(t/T)$
at the end of each cycle (symbols in figure 2), quantified by linear fits over at least five successive data points in the linear regime after the initial relaxation (
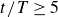 $t/T\ge 5$
). We obtain a sharp peak in magnitude at
$t/T\ge 5$
). We obtain a sharp peak in magnitude at
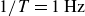 $1/T=1\,\text{Hz}$
, while for both
$1/T=1\,\text{Hz}$
, while for both
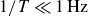 $1/T\ll 1\,\text{Hz}$
and
$1/T\ll 1\,\text{Hz}$
and
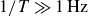 $1/T\gg 1\,\text{Hz}$
, the motion ceases. For comparison, the inset of figure 3 shows the same data for a swimmer moving in a polystyrene granulate. Although we cannot scan the same frequency range as for hydrogel particles due to limitations of the apparatus, we see that the net displacement per cycle is strictly positive and decreases slightly with increasing frequency
$1/T\gg 1\,\text{Hz}$
, the motion ceases. For comparison, the inset of figure 3 shows the same data for a swimmer moving in a polystyrene granulate. Although we cannot scan the same frequency range as for hydrogel particles due to limitations of the apparatus, we see that the net displacement per cycle is strictly positive and decreases slightly with increasing frequency
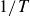 $1/T$
. This behaviour resembles the rate independence reported for other locomotion problems in plastically deforming granular media (Maladen et al. Reference Maladen, Ding, Li and Goldman2009; Hatton et al. Reference Hatton, Ding, Choset and Goldman2013). Whereas the persistent forward motion in stiff, frictional and non-cohesive granular media is associated with particle jamming in the vicinity of the swimmer (Xiao et al. Reference Xiao, Torres, Sack and Pöschel2025), the backward propulsion observed in hydrogel particles arises from a different mechanism, as discussed below.
$1/T$
. This behaviour resembles the rate independence reported for other locomotion problems in plastically deforming granular media (Maladen et al. Reference Maladen, Ding, Li and Goldman2009; Hatton et al. Reference Hatton, Ding, Choset and Goldman2013). Whereas the persistent forward motion in stiff, frictional and non-cohesive granular media is associated with particle jamming in the vicinity of the swimmer (Xiao et al. Reference Xiao, Torres, Sack and Pöschel2025), the backward propulsion observed in hydrogel particles arises from a different mechanism, as discussed below.
Net swimmer displacement per cycle as a function of the flapping frequency in hydrogel particles. Error bars indicate the differences between two experimental repetitions. The inset shows the data for a swimmer moving in polystyrene particles; here, the error bars are very small.

While figure 3 presents the average swimmer’s net displacement per cycle, figure 4 shows the sub-cycle phase-resolved velocity,
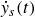 $\dot {y}_s(t)$
, together with the corresponding driving force,
$\dot {y}_s(t)$
, together with the corresponding driving force,
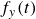 $f_y(t)$
, over a period
$f_y(t)$
, over a period
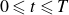 $0 \leqslant t \leqslant T$
for the flapping frequency
$0 \leqslant t \leqslant T$
for the flapping frequency
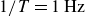 $1/T=1\,\text{Hz}$
corresponding to the peak average locomotion velocity. At the start of each stroke,
$1/T=1\,\text{Hz}$
corresponding to the peak average locomotion velocity. At the start of each stroke,
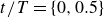 $t/T=\{0,0.5\}$
, the swimmer initially continues to move in the direction set by the preceding stroke, despite the reversal of the motion of the wings. During this period, the work done by the driving force is negative (
$t/T=\{0,0.5\}$
, the swimmer initially continues to move in the direction set by the preceding stroke, despite the reversal of the motion of the wings. During this period, the work done by the driving force is negative (
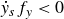 $\dot {y}_sf_y\lt 0$
), which decelerates the swimmer over a duration
$\dot {y}_sf_y\lt 0$
), which decelerates the swimmer over a duration
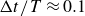 $\Delta t/T\approx 0.1$
, until
$\Delta t/T\approx 0.1$
, until
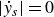 $|\dot {y}_s|=0$
. This behaviour suggests that the swimmer’s inertia cannot be neglected. If inertia were negligible, the swimmer would instantly follow the direction of wing motion and its velocity would change sign immediately at
$|\dot {y}_s|=0$
. This behaviour suggests that the swimmer’s inertia cannot be neglected. If inertia were negligible, the swimmer would instantly follow the direction of wing motion and its velocity would change sign immediately at
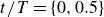 $t/T = \{0, 0.5\}$
.
$t/T = \{0, 0.5\}$
.
Phase-resolved swimmer velocity (black dashed line, right axis) and driving force (red solid line, left axis) at a flapping frequency of
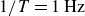 $1/T=1\,\text{Hz}$
in hydrogel particles. The vertical dotted line separates the opening and closing half-cycles, sketched in the insets.
$1/T=1\,\text{Hz}$
in hydrogel particles. The vertical dotted line separates the opening and closing half-cycles, sketched in the insets.

The swimming motion observed in our experiment occurs when the space-fixed swimmer moves relative to the frictionless container filled with hydrogel granulate. The effective mass associated with this motion is, therefore, that of the container and the hydrogel granulate, rather than the mass of the swimmer itself. Consequently, our experimental set-up corresponds to a free swimmer (neglecting rotation) whose density exceeds that of the ambient fluid by orders of magnitude. The high density should only affect the swimmer’s inertia, not its buoyancy. Therefore, a free-swimmer experiment that corresponds to our fixed-swimmer set-up should be carried out in the absence of gravity.
A closely related system was investigated by Gonzalez-Rodriguez & Lauga (Reference Gonzalez-Rodriguez and Lauga2009) and Hubert et al. (Reference Hubert, Trosman, Collard, Sukhov, Harting, Vandewalle and Smith2021), who found that reciprocal swimming is possible in a Newtonian fluid within the Stokes regime, provided the swimmer’s density is much larger than that of the fluid,
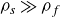 $\rho _{{s}}\gg \rho _{{f}}$
. In this case, the swimmer-based Reynolds number,
$\rho _{{s}}\gg \rho _{{f}}$
. In this case, the swimmer-based Reynolds number,
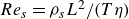 ${Re}_{{s}}=\rho _{{s}}L^2/(T\eta )$
, is finite, whereas the Reynolds number of the fluid,
${Re}_{{s}}=\rho _{{s}}L^2/(T\eta )$
, is finite, whereas the Reynolds number of the fluid,
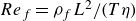 ${Re}_{{f}}=\rho _{{f}}L^2/(T\eta )$
, is negligible. Here,
${Re}_{{f}}=\rho _{{f}}L^2/(T\eta )$
, is negligible. Here,
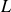 $L$
denotes the characteristic length of the swimmer and
$L$
denotes the characteristic length of the swimmer and
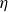 $\eta$
the viscosity of the fluid. The analytical result by Gonzalez-Rodriguez & Lauga (Reference Gonzalez-Rodriguez and Lauga2009) then predicts a cycle-averaged net velocity in the
$\eta$
the viscosity of the fluid. The analytical result by Gonzalez-Rodriguez & Lauga (Reference Gonzalez-Rodriguez and Lauga2009) then predicts a cycle-averaged net velocity in the
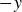 $-y$
direction that increases with the flapping frequency,
$-y$
direction that increases with the flapping frequency,
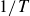 $1/T$
.
$1/T$
.
Although we acknowledge the substantial differences between the system examined by Gonzalez-Rodriguez & Lauga (Reference Gonzalez-Rodriguez and Lauga2009) and our own, we note that the direction of motion (along
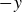 $-y$
) and the frequency dependence are similar, at least for low frequencies up to
$-y$
) and the frequency dependence are similar, at least for low frequencies up to
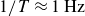 $1/T\approx 1\,\text{Hz}$
, cf. figure 3. At higher frequencies, however, the viscoelastic properties of the hydrogel become significant, and local density variations within the granulate arise due to the rapid motion of the wings. These effects will be discussed in detail in the following sections.
$1/T\approx 1\,\text{Hz}$
, cf. figure 3. At higher frequencies, however, the viscoelastic properties of the hydrogel become significant, and local density variations within the granulate arise due to the rapid motion of the wings. These effects will be discussed in detail in the following sections.
4. Dynamics of swimming in hydrogels
4.1. Inertia
The dynamics of the system is determined by the accelerated mass, the flapping period
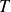 $T$
and the viscoelastic properties of the granulate, which relate the swimmer to the granulate-filled container. In our experiment, the swimmer is fixed in space, therefore, the relevant mass is the mass of the movable granulate-filled container. For a very low frequency,
$T$
and the viscoelastic properties of the granulate, which relate the swimmer to the granulate-filled container. In our experiment, the swimmer is fixed in space, therefore, the relevant mass is the mass of the movable granulate-filled container. For a very low frequency,
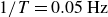 $1/T=0.05\,\text{Hz}$
, the container follows the flapping wings approximately, with no noticeable net displacement over one period, as evidenced by a symmetric displacement profile for the opening and closing half-periods, see figure 5. The phase-resolved displacement was obtained by averaging over all periods, disregarding the initial transient. Note that the net transport per cycle, shown in figure 2, is much smaller than the typical peak displacement. With its accelerated mass and the restoring force from the granulate, the container may thus be regarded as a driven nonlinear oscillator, oscillating about the swimmer’s position with a superimposed slow drift. For slow flapping,
$1/T=0.05\,\text{Hz}$
, the container follows the flapping wings approximately, with no noticeable net displacement over one period, as evidenced by a symmetric displacement profile for the opening and closing half-periods, see figure 5. The phase-resolved displacement was obtained by averaging over all periods, disregarding the initial transient. Note that the net transport per cycle, shown in figure 2, is much smaller than the typical peak displacement. With its accelerated mass and the restoring force from the granulate, the container may thus be regarded as a driven nonlinear oscillator, oscillating about the swimmer’s position with a superimposed slow drift. For slow flapping,
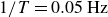 $1/T=0.05\,\text{Hz}$
, the force acting on the container is small, and the container follows the driving flaps without appreciable deformation of the granular particles. Consequently, the trajectory
$1/T=0.05\,\text{Hz}$
, the force acting on the container is small, and the container follows the driving flaps without appreciable deformation of the granular particles. Consequently, the trajectory
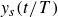 $y_s (t/T)$
is symmetric between the two half-cycles. For fast flapping (
$y_s (t/T)$
is symmetric between the two half-cycles. For fast flapping (
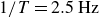 $1/T=2.5\,\text{Hz}$
), the container cannot follow the driving motion owing to its inertia. The wings strongly deform the granular particles, but their low elastic modulus prevents sufficient force transfer to accelerate the container appreciably, thus,
$1/T=2.5\,\text{Hz}$
), the container cannot follow the driving motion owing to its inertia. The wings strongly deform the granular particles, but their low elastic modulus prevents sufficient force transfer to accelerate the container appreciably, thus,
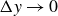 $\Delta y\to 0$
.
$\Delta y\to 0$
.
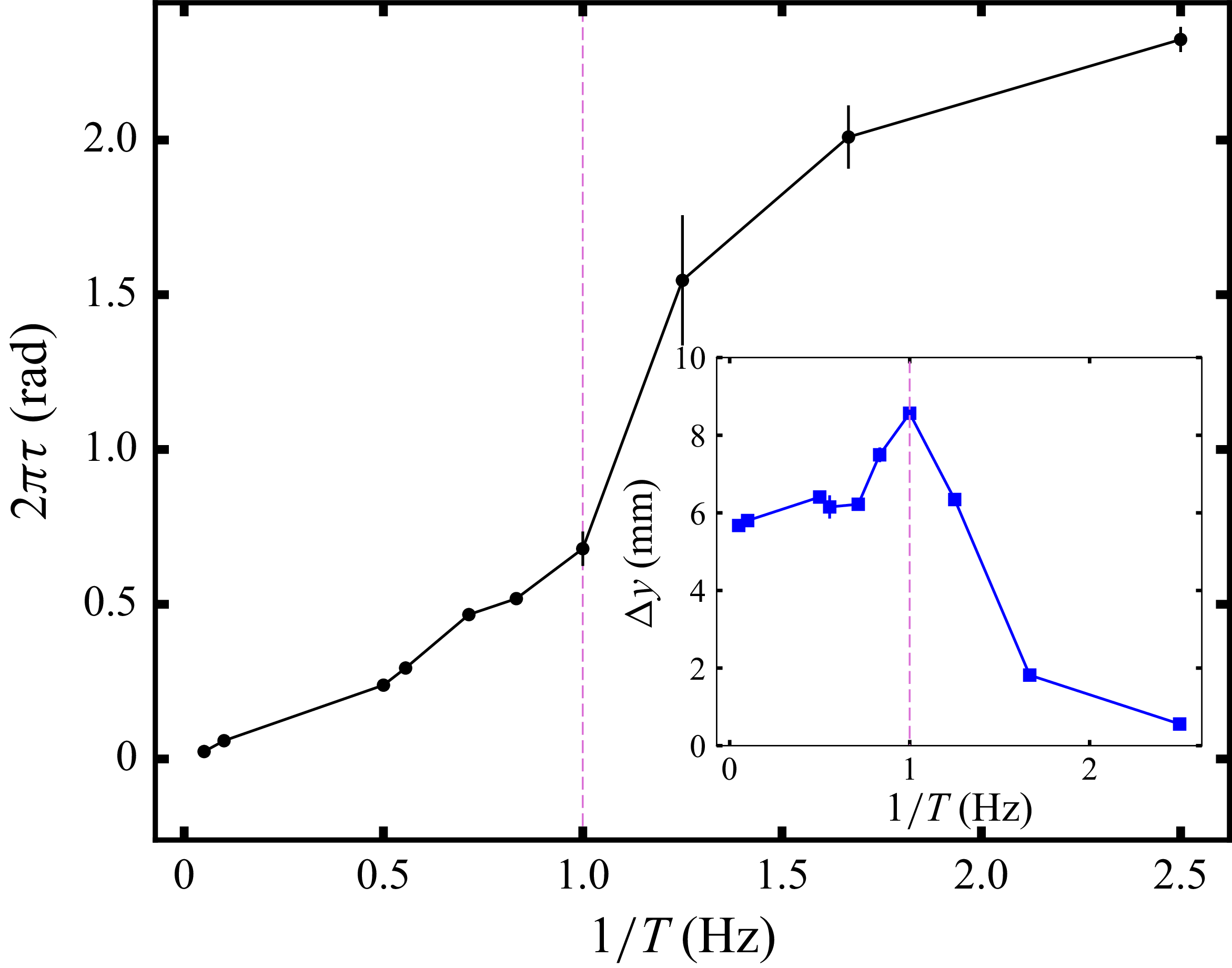
Averaged relative displacement of the swimmer as a function of the phase,
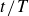 $t/T$
, for various flapping frequencies in hydrogel particles. For each case, the average was taken over at least five individual cycles beyond the initial transients. The vertical dotted line separates the opening and closing half-cycles. The figure also defines the (positive) peak location,
$t/T$
, for various flapping frequencies in hydrogel particles. For each case, the average was taken over at least five individual cycles beyond the initial transients. The vertical dotted line separates the opening and closing half-cycles. The figure also defines the (positive) peak location,
 $\tau$
, and the peak-to-peak amplitude,
$\tau$
, and the peak-to-peak amplitude,
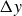 $\Delta y$
, that depend on the flapping frequency, see figure 6.
$\Delta y$
, that depend on the flapping frequency, see figure 6.

The inertia-based mechanism described above introduces a phase shift
 $\tau$
between the wing actuation and the relative displacement of the swimmer and container. While the phase shift
$\tau$
between the wing actuation and the relative displacement of the swimmer and container. While the phase shift
 $\tau$
increases with flapping frequency, the amplitude of the induced oscillation,
$\tau$
increases with flapping frequency, the amplitude of the induced oscillation,
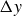 $\Delta y$
(defined in figure 5), varies non-monotonically. In the limit
$\Delta y$
(defined in figure 5), varies non-monotonically. In the limit
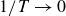 $1/T \to 0$
, the container follows the driving motion and
$1/T \to 0$
, the container follows the driving motion and
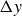 $\Delta y$
approaches a constant value,
$\Delta y$
approaches a constant value,
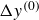 $\Delta y^{(0)}$
. In the opposite limit
$\Delta y^{(0)}$
. In the opposite limit
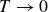 $T \to 0$
,
$T \to 0$
,
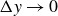 $\Delta y \to 0$
owing to the low elastic modulus of the medium. At intermediate frequency,
$\Delta y \to 0$
owing to the low elastic modulus of the medium. At intermediate frequency,
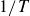 $1/T$
, we observe amplitudes
$1/T$
, we observe amplitudes
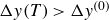 $\Delta y(T) \gt \Delta y^{(0)}$
, resembling resonant oscillations.
$\Delta y(T) \gt \Delta y^{(0)}$
, resembling resonant oscillations.
This analysis is supported by figure 6, which shows the phase shift
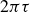 $2\pi \tau$
and the peak-to-peak amplitude
$2\pi \tau$
and the peak-to-peak amplitude
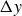 $\Delta y$
as functions of the flapping frequency
$\Delta y$
as functions of the flapping frequency
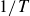 $1/T$
. The dotted lines reveal that the frequency at which
$1/T$
. The dotted lines reveal that the frequency at which
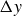 $\Delta y$
reaches its maximum coincides with the frequency of fastest locomotion, while the phase shift experiences a rapid increase from
$\Delta y$
reaches its maximum coincides with the frequency of fastest locomotion, while the phase shift experiences a rapid increase from
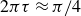 $2\pi \tau \approx \pi /4$
to
$2\pi \tau \approx \pi /4$
to
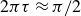 $2\pi \tau \approx \pi /2$
. These trends in
$2\pi \tau \approx \pi /2$
. These trends in
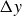 $\Delta y$
and
$\Delta y$
and
 $\tau$
resemble those of a periodically driven damped oscillator, with resonance occurring near
$\tau$
resemble those of a periodically driven damped oscillator, with resonance occurring near
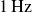 $1\,\text{Hz}$
, which is the frequency of maximum locomotion velocity plotted in figure 3.
$1\,\text{Hz}$
, which is the frequency of maximum locomotion velocity plotted in figure 3.
Phase shift
 $\tau$
and peak-to-peak amplitude
$\tau$
and peak-to-peak amplitude
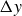 $\Delta y$
(inset) as functions of the flapping frequency in hydrogel particles. The vertical dashed lines mark the frequency of maximal locomotion (see figure 2). Error bars indicate the differences between two experimental repetitions, which are very small for
$\Delta y$
(inset) as functions of the flapping frequency in hydrogel particles. The vertical dashed lines mark the frequency of maximal locomotion (see figure 2). Error bars indicate the differences between two experimental repetitions, which are very small for
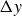 $\Delta y$
. The standard errors (not shown) in the cycle-averaged
$\Delta y$
. The standard errors (not shown) in the cycle-averaged
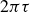 $2\pi \tau$
and
$2\pi \tau$
and
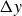 $\Delta y$
are also negligible. Lines guide the eye.
$\Delta y$
are also negligible. Lines guide the eye.

To clarify this coincidence, we examine the nature of the forces exerted on the swimmer by the hydrogel medium.
4.2. Viscoelastic relaxation in the hydrogel
Depending on the deformation rate, hydrogels exhibit both viscous and elastic behaviours. The hydrogel’s response determines the interaction force and thus the swimmer’s dynamics. We examined the viscoelastic response in relaxation experiments. In case (a), with the air bearings blocked to prevent reservoir displacement, the swimmer’s wings were rapidly opened from
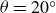 $\theta =20^\circ$
to
$\theta =20^\circ$
to
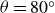 $\theta =80^\circ$
at
$\theta =80^\circ$
at
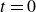 $t=0$
, and the subsequent relaxation of the force (magnitude) on the swimmer,
$t=0$
, and the subsequent relaxation of the force (magnitude) on the swimmer,
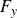 $F_y$
, was recorded. Case (b) was analogous, except that the wings were rapidly closed from
$F_y$
, was recorded. Case (b) was analogous, except that the wings were rapidly closed from
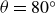 $\theta =80^\circ$
to
$\theta =80^\circ$
to
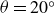 $\theta =20^\circ$
at
$\theta =20^\circ$
at
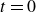 $t=0$
.
$t=0$
.
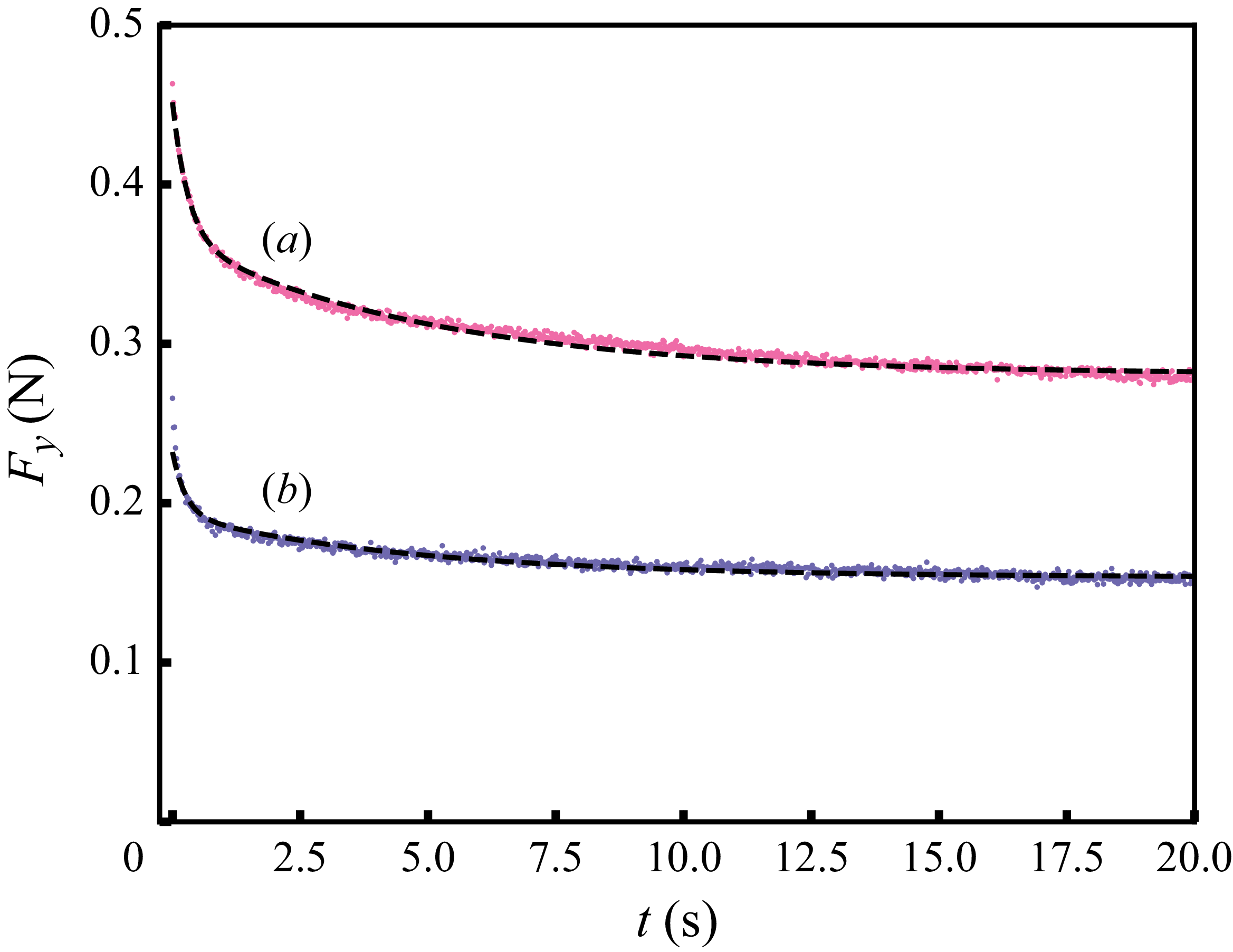
Figure 7 shows the measured forces, which exhibit a rapid decay within the first few seconds followed by a slower relaxation toward a long-term residual value, reflecting the combined viscous and elastic response of the medium.
Relaxation of the swimming force following a rapid stroke in hydrogel particles. For opening (a), the data are fitted to (4.1) (dashed curve) with
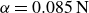 $\alpha =0.085\,\text{N}$
,
$\alpha =0.085\,\text{N}$
,
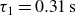 $\tau _1=0.31\,\text{s}$
,
$\tau _1=0.31\,\text{s}$
,
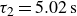 $\tau _2=5.02\,\text{s}$
and
$\tau _2=5.02\,\text{s}$
and
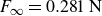 $F_\infty =0.281\,\text{N}$
. For closing (b), we obtain
$F_\infty =0.281\,\text{N}$
. For closing (b), we obtain
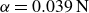 $\alpha =0.039\,\text{N}$
,
$\alpha =0.039\,\text{N}$
,
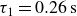 $\tau _1=0.26\,\text{s}$
,
$\tau _1=0.26\,\text{s}$
,
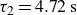 $\tau _2=4.72\,\text{s}$
and
$\tau _2=4.72\,\text{s}$
and
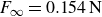 $F_\infty =0.154\,\text{N}$
.
$F_\infty =0.154\,\text{N}$
.

We fitted the function
 \begin{equation} F_y = \alpha \left (e^{-t/\tau _1}+e^{-t/\tau _2}\right ) + F_\infty, \end{equation}
\begin{equation} F_y = \alpha \left (e^{-t/\tau _1}+e^{-t/\tau _2}\right ) + F_\infty, \end{equation}
to the measured forces over
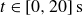 $t\in [0, 20]\,\text{s}$
to obtain the relaxation times:
$t\in [0, 20]\,\text{s}$
to obtain the relaxation times:
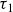 $\tau _1$
possibly associated with the responses at the individual particle level, including the dissipation within the hydrogel material and the inter-particle capillary bridges (Farmani et al. Reference Farmani, Wang, Stannarius and Dijksman2025), and
$\tau _1$
possibly associated with the responses at the individual particle level, including the dissipation within the hydrogel material and the inter-particle capillary bridges (Farmani et al. Reference Farmani, Wang, Stannarius and Dijksman2025), and
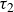 $\tau _2$
, associated with particle rearrangements, with
$\tau _2$
, associated with particle rearrangements, with
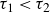 $\tau _1 \lt \tau _2$
. Averaging over opening and closing strokes gives
$\tau _1 \lt \tau _2$
. Averaging over opening and closing strokes gives
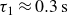 $\tau _1\approx 0.3\,\text{s}$
and
$\tau _1\approx 0.3\,\text{s}$
and
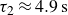 $\tau _2\approx 4.9\,\text{s}$
. Remarkably, the peak in backward locomotion at
$\tau _2\approx 4.9\,\text{s}$
. Remarkably, the peak in backward locomotion at
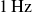 $1\,\text{Hz}$
(see figure 2) lies between
$1\,\text{Hz}$
(see figure 2) lies between
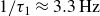 $1/\tau _1 \approx 3.3\,\text{Hz}$
and
$1/\tau _1 \approx 3.3\,\text{Hz}$
and
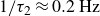 $1/\tau _2 \approx 0.2\,\text{Hz}$
. The corresponding Deborah numbers,
$1/\tau _2 \approx 0.2\,\text{Hz}$
. The corresponding Deborah numbers,
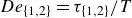 ${De}_{\{1,2\}}=\tau _{\{1,2\}}/T$
, are of the order of unity, indicating that both elastic and viscous effects contribute significantly to the swimmer’s propulsive force. Further increasing the swimming frequency may result in enhanced elastic effects. The corresponding decrease in the swimmer’s net displacement may be similar to that observed in Keim et al. (Reference Keim, Garcia and Arratia2012), where the propulsion speed of an oscillating dumbbell in a viscoelastic fluid reaches a peak at
${De}_{\{1,2\}}=\tau _{\{1,2\}}/T$
, are of the order of unity, indicating that both elastic and viscous effects contribute significantly to the swimmer’s propulsive force. Further increasing the swimming frequency may result in enhanced elastic effects. The corresponding decrease in the swimmer’s net displacement may be similar to that observed in Keim et al. (Reference Keim, Garcia and Arratia2012), where the propulsion speed of an oscillating dumbbell in a viscoelastic fluid reaches a peak at
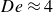 ${De}\approx 4$
. Note that the residual force
${De}\approx 4$
. Note that the residual force
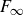 $F_\infty$
also contributes to the initial force. On the experimental time scale,
$F_\infty$
also contributes to the initial force. On the experimental time scale,
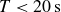 $T\lt 20\,\text{s}$
, it is effectively elastic.
$T\lt 20\,\text{s}$
, it is effectively elastic.
4.3. Variation of density in hydrogel media
Rearrangement of the hydrogel particles, characterised by the relaxation time
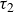 $\tau _2$
, occurs on a slower time scale than the wing motion, leading to densification of the medium in front of the moving wing and dilatation behind it. Whereas densification is limited by the material density of the hydrogel particles, dilatation is not, since air-filled voids may form between the granular particles. Consequently, the average density of the hydrogel granular medium over a flapping period is smaller in the volume swept by the wings than in regions far from the wings. Particle reordering counteracts this effect. Therefore, the difference in density is more pronounced for faster flapping, evidenced by figure 8, showing X-ray radiographs of the swimmer and the hydrogel granulate in the
$\tau _2$
, occurs on a slower time scale than the wing motion, leading to densification of the medium in front of the moving wing and dilatation behind it. Whereas densification is limited by the material density of the hydrogel particles, dilatation is not, since air-filled voids may form between the granular particles. Consequently, the average density of the hydrogel granular medium over a flapping period is smaller in the volume swept by the wings than in regions far from the wings. Particle reordering counteracts this effect. Therefore, the difference in density is more pronounced for faster flapping, evidenced by figure 8, showing X-ray radiographs of the swimmer and the hydrogel granulate in the
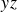 $y{}z$
plane (see figure 1). The radiographs (
$y{}z$
plane (see figure 1). The radiographs (
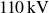 $110\,\text{kV}$
,
$110\,\text{kV}$
,
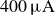 $400\,\unicode{x03BC} \text{A}$
) were recorded at
$400\,\unicode{x03BC} \text{A}$
) were recorded at
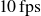 $10\, \text{fps}$
and averaged over all frames acquired during at least five flapping cycles.
$10\, \text{fps}$
and averaged over all frames acquired during at least five flapping cycles.
Time-averaged X-ray radiographs of the swimmer flapping with frequency
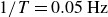 $1/T=0.05\,\text{Hz}$
(a) and
$1/T=0.05\,\text{Hz}$
(a) and
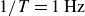 $1/T=1\,\text{Hz}$
(b) in hydrogel particles. In each case, the average was taken over entire swimming cycles for at least five cycles. Yellow boxes mark the regions swept by the wings, and red boxes indicate the position of the mounting rod. A higher intensity
$1/T=1\,\text{Hz}$
(b) in hydrogel particles. In each case, the average was taken over entire swimming cycles for at least five cycles. Yellow boxes mark the regions swept by the wings, and red boxes indicate the position of the mounting rod. A higher intensity
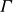 $\varGamma$
corresponds to a lower density in the hydrogel medium.
$\varGamma$
corresponds to a lower density in the hydrogel medium.

Comparison of the X-ray images in panels (a) and (b) shows that the granulate density in the region swept by the wings (yellow box) is lower during fast flapping (
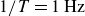 $1/T=1\,\text{Hz}$
) than during slow flapping (
$1/T=1\,\text{Hz}$
) than during slow flapping (
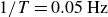 $1/T=0.05\,\text{Hz}$
). (Because of speed limitations in the X-ray imaging, radiographs could not be obtained for faster wing rotations.) Thus, owing to the relaxation time
$1/T=0.05\,\text{Hz}$
). (Because of speed limitations in the X-ray imaging, radiographs could not be obtained for faster wing rotations.) Thus, owing to the relaxation time
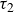 $\tau _2$
, voids form in the vicinity of the wings due to the viscoelasticity of the hydrogel granulate. For
$\tau _2$
, voids form in the vicinity of the wings due to the viscoelasticity of the hydrogel granulate. For
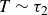 $T \sim \tau _2$
, the voids can be at least partially refilled during each stroke by particle rearrangements, leading to hysteresis in the swimming cycle despite the reciprocal wing motion. For
$T \sim \tau _2$
, the voids can be at least partially refilled during each stroke by particle rearrangements, leading to hysteresis in the swimming cycle despite the reciprocal wing motion. For
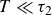 $T \ll \tau _2$
, particle reordering cannot keep pace with the wing motion, therefore voids persist during both opening and closing strokes, and symmetry remains unbroken. This is reminiscent of the propulsion loss observed in bacteria moving through dense polymer solutions, where a nanometre-sized flagellum is believed to deplete nearby polymer coils (Martinez et al. Reference Martinez, Schwarz-Linek, Reufer, Wilson, Morozov and Poon2014). In our study, the use of air as the interstitial fluid provides high contrast against the particles, enabling direct X-ray visualisation of this non-continuum effect.
$T \ll \tau _2$
, particle reordering cannot keep pace with the wing motion, therefore voids persist during both opening and closing strokes, and symmetry remains unbroken. This is reminiscent of the propulsion loss observed in bacteria moving through dense polymer solutions, where a nanometre-sized flagellum is believed to deplete nearby polymer coils (Martinez et al. Reference Martinez, Schwarz-Linek, Reufer, Wilson, Morozov and Poon2014). In our study, the use of air as the interstitial fluid provides high contrast against the particles, enabling direct X-ray visualisation of this non-continuum effect.
4.4. Implications for locomotion
From the examination of inertial and viscoelastic effects in §§ 4.1, 4.2 and 4.3, a consistent picture emerges for the propagation of a swimmer in a hydrogel granular medium, as shown in figure 2. To support the subsequent discussion, we introduce an idealised dynamical equation for the swimmer. Although the precise mathematical form of each term is not guaranteed, the equation should at least capture the essential features of the dynamics
 \begin{equation} m\frac {d^2y_s}{{\rm d}t^2}+\gamma (\theta )\frac {{\rm d}y_s}{{\rm d}t}+\kappa (\theta )(y_s-y_0(t))=f_{{w\textit{ing}}}(\theta ). \end{equation}
\begin{equation} m\frac {d^2y_s}{{\rm d}t^2}+\gamma (\theta )\frac {{\rm d}y_s}{{\rm d}t}+\kappa (\theta )(y_s-y_0(t))=f_{{w\textit{ing}}}(\theta ). \end{equation}
Here,
 $m$
is the mass of the movable container,
$m$
is the mass of the movable container,
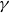 $\gamma$
is the drag coefficient,
$\gamma$
is the drag coefficient,
 $\kappa$
an elastic constant and
$\kappa$
an elastic constant and
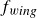 $f_{{w\textit{ing}}}$
the driving force generated by the wing’s motion. All terms depend on the instantaneous wing angle
$f_{{w\textit{ing}}}$
the driving force generated by the wing’s motion. All terms depend on the instantaneous wing angle
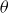 $\theta$
. In addition, the coefficients
$\theta$
. In addition, the coefficients
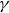 $\gamma$
and
$\gamma$
and
 $\kappa$
, as well as the driving force
$\kappa$
, as well as the driving force
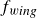 $f_{{w\textit{ing}}}$
, depend on the density of the granular medium in a complex manner that is not addressed here. In the stationary state, the mean position
$f_{{w\textit{ing}}}$
, depend on the density of the granular medium in a complex manner that is not addressed here. In the stationary state, the mean position
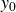 $y_0$
drifts at the rate
$y_0$
drifts at the rate
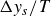 $\Delta y_s/T$
, as plotted in figure 3. We now discuss the system in three characteristic flapping regimes:
$\Delta y_s/T$
, as plotted in figure 3. We now discuss the system in three characteristic flapping regimes:
-
(i) For
 $1/T\gg 1\,\text{Hz}$
viscous effects cease as
$T\ll \tau _1 \lt \tau _2$
. Here,
$\gamma \approx 0$
, so the swimmer effectively flaps in an elastic medium while generating long-lasting voids. Owing to the absence of relaxation,
$y_0(t) \approx 0$
, and (4.2) therefore describes harmonic oscillations without locomotion.
$1/T\gg 1\,\text{Hz}$
viscous effects cease as
$T\ll \tau _1 \lt \tau _2$
. Here,
$\gamma \approx 0$
, so the swimmer effectively flaps in an elastic medium while generating long-lasting voids. Owing to the absence of relaxation,
$y_0(t) \approx 0$
, and (4.2) therefore describes harmonic oscillations without locomotion. -
(ii) For
$1/T\ll 1\,\text{Hz}$
, inertia does not influence swimming, as evidenced by the negligible phase shift in figure 6. In this regime, the inertial term
$m\,d^2y_s/{\rm d}t^2$
is obsolete, and (4.2) depends only on the wing angle
$\theta$
. At very low frequencies, particle rearrangements in the hydrogel granulate are fast enough to refill the voids during each stroke, making (4.2) effectively density independent. This situation resembles the original scallop theorem, which prohibits locomotion despite strong elastic effects, as seen in figure 7. -
(iii) For
$1/T\approx 1\,\text{Hz}$
, viscous and elastic effects coexist, and inertia becomes significant. In this regime, all terms in (4.2) contribute, with the microscopic origin linked to hysteresis in the granular density caused by partial refilling of voids on the time scale
$\tau _2 \sim T$
. While the interplay between inertia and viscous effects results in the net swimmer displacement per cycle, it is likely that the interplay between elastic and inertial effects amplifies the net displacement magnitude via a behaviour that resembles resonance, as shown in figure 6. This facilitates the energy transfer between the swimmer and the medium due to the phase shift (figures 4 and 6), allowing
$\gamma {\rm d}y_s/{\rm d}t$
and
$f_{{w\textit{ing}}}$
to act in the same direction after a switch of the wing rotation direction. This eventually results in a maximum net swimmer displacement at
$1/T\approx 1\,$
Hz.
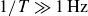
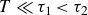
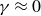
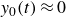
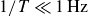
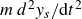
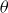
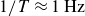
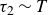
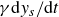
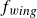
5. Summary
In summary, a scallop-like swimmer with reciprocally flapping wings in a granular bed of hydrogel spheres can achieve backward locomotion within a certain range of flapping frequencies. This behaviour arises from the combined effects of inertia and viscoelasticity of the soft granular hydrogel medium. The locomotion and its underlying mechanisms differ fundamentally from previous observations in idealised rigid granular media, such as plastic beads (Xiao et al. Reference Xiao, Torres, Sack and Pöschel2025).
Acknowledgements
R.S. and J.W. acknowledge funding by DLR within project 50WK2348. The authors would like to thank A. Nazemi for fruitful discussions.
Declaration of interests
The authors report no conflicts of interest.
Author contributions
H.X. and J.W. have contributed equally to this work.
































 Open access
Open access