


Introduction
Seeds serve as both the starting point and conclusion of an annual plant’s life cycle (Sajeev et al. Reference Sajeev, Koornneef and Bentsink2024). Because of the critical role seeds play, especially for annual species, plants have evolved complex regulatory systems that control when seeds germinate or remain dormant (Franklin and Whitelam Reference Franklin and Whitelam2005; Graeber et al. Reference Graeber, Nakabayashi, Miatton, Leubner-Metzger and Soppe2012). These adaptations ensure that seedlings establish under favorable conditions while allowing seeds to persist across heterogeneous environments and time periods (Tuan et al. Reference Tuan, Sun, Nguyen, Park, Ayele, Feng, Nemzer and DeVries2018). In weed species, these mechanisms are particularly important, because dormancy and germination timing directly influence seedbank persistence, emergence patterns, crop yield loss potential, and management outcomes in agroecosystems (Fu et al. Reference Fu, Ma, Li, Hou, Zeng, Zhang, Liu, Bi, Tan, Yu, Bi and Luo2024; Sajeev et al. 2024; Schwerzel and Thomas Reference Schwerzel and Thomas1971).
Light is a key environmental signal that regulates seed germination, through responses mediated by specialized photoreceptors (Jiang et al. Reference Jiang, Xu, Jing, Tang and Lin2016; Shinomura Reference Shinomura1997; Tripathi et al. Reference Tripathi, Hoang, Han and Kim2019). These photoreceptors allow seeds to integrate environmental information and determine suitable conditions for germination (Paik and Huq Reference Paik and Huq2019). Photoperiod, light intensity, and spectral composition play particularly important roles in regulating plant responses (Møller et al. Reference Møller, Ingles and Whitelam2002; Montgomery and Lagarias Reference Montgomery and Lagarias2002). Phytochromes are the primary photoreceptors involved in sensing light quality and activating metabolic pathways necessary for germination (Chen et al. Reference Chen, Wang, Guo, Li, Tian, Han, Tang and Liu2023; Hu et al. Reference Hu, Li, Yang, Yuan, Yang, Fu, Zhang, Li, Shen and Jiang2024; Simlat et al. Reference Simlat, Ślęzak, Moś, Warchoł, Skrzypek and Ptak2016; Wei et al. Reference Wei, Wang and Yu2023). Phytochrome B (phyB) primarily mediates low-fluence responses to red light, whereas phytochrome A (phyA) governs sensitivity to very low fluence and continuous far-red light, producing very low fluence and high-irradiance responses (VLFR and HIR) (Shichijo et al. Reference Shichijo, Katada, Tanaka and Hashimoto2001; Takaki Reference Takaki2001).
Light quality is particularly important for detecting competition from neighboring vegetation (Franklin and Whitelam Reference Franklin and Whitelam2005). Green leaves absorb most red wavelengths while transmitting or reflecting far-red light, thereby lowering the red:far-red (R:FR) ratio beneath plant canopies. This shift in spectral composition is a reliable indicator of impending shading and competition for light. Through phytochrome-mediated perception, seeds effectively “sense” the surrounding environment and adjust germination accordingly (Schmitt and Wulff Reference Schmitt and Wulff1993; Uyehara et al. Reference Uyehara, Bechinger, Jordan and van Kleunen2024). Germination is often suppressed under far red–enriched conditions, preventing seedling emergence in shaded microsites, where competitive pressure is high and establishment success is reduced (Lee and Lopez-Molina Reference Lee and Lopez-Molina2012).
Changes in light quality also underlie the suppressive effects of surface residues and cover crop canopies on weed emergence. As cover crops develop, they decrease red light, thereby altering the R:FR ratio at the soil surface, creating spectral conditions that inhibit germination of photoblastic weed seeds (Camargo Silva and Bagavathiannan Reference Camargo Silva and Bagavathiannan2023; Horvath et al. Reference Horvath, Bruggeman, Moriles-Miller, Anderson, Dogramaci, Scheffler, Hernandez, Foley and Clay2018; Page et al. Reference Page, Tollenaar, Lee, Lukens and Swanton2009). Many weed species, especially those with smaller seeds, rely on red light as a cue for germination, and exposure to low R:FR ratios associated with canopy shade can maintain dormancy or induce secondary dormancy (Benech-Arnold et al. Reference Benech-Arnold, Sánchez, Forcella, Kruk and Ghersa2000). These responses illustrate how light quality functions as a key ecological signal in regulating weed emergence.
Seed dormancy and germination behavior strongly influence the effectiveness of weed management strategies that depend on emergence timing, including the use of cover crops and residue-based systems (Nakabayashi and Leubner-Metzger Reference Nakabayashi and Leubner-Metzger2021; Sias et al. Reference Sias, Wolters, Reiter and Flessner2021; Travlos et al. Reference Travlos, Gazoulis, Kanatas, Tsekoura, Zannopoulos and Papastylianou2020). Preplant burndown herbicides and soil-residual herbicides are also dependent on weed emergence timing (Owen et al. Reference Owen, Steckel, Koger, Main and Mueller2009; Sbatella and Wilson Reference Sbatella and Wilson2010). Kochia [Bassia scoparia (L.) A.J. Scott] and common lambsquarters (Chenopodium album L.) are widespread and problematic weed species that exhibit contrasting germination and dormancy characteristics. Although both species are known to differ in their sensitivity to environmental cues, substantial intraspecific variation in light-dependent germination has been reported, particularly in C. album (Eslami Reference Eslami2011; Friesen et al. Reference Friesen, Beckie, Warwick and Van Acker2009). However, the extent to which primary dormancy, far-red light–enforced inhibition, and seed size interact to regulate germination responses across germplasm sources within these species remains unclear.
This study aimed to quantify the germination responses of B. scoparia and C. album under contrasting light-quality signals, determine whether primary dormancy is coordinated with far-red light–enforced inhibition, and evaluate whether seed size is associated with light-dependent dormancy responses. Specifically, we asked: (1) Do populations of B. scoparia and accessions of C. album respond similarly to light quality with respect to germination? (2) Is primary dormancy associated with far-red light–enforced inhibition? (3) Is seed size related to variation in light-dependent dormancy responses?
Materials and Methods
Plant Materials and Seed Increase
All research was conducted in controlled environments between 2022 and 2025 at the University of Wyoming Laramie Research and Extension Center (LREC) greenhouse and the seed laboratory of the Department of Plant Sciences, Laramie, WY. Forty-nine accessions of C. album were obtained from the USDA Germplasm Resources Information Network (USDA-GRIN) and propagated under greenhouse conditions at LREC. In addition, 70 populations of B. scoparia were collected from Colorado State University and from agricultural fields in Wyoming, Nebraska, and Montana. Seed from each germplasm source was increased under a common maternal environment to generate sufficient seed quantity and ensure uniform seed stock for germination experiments.
To increase the seed of each germplasm source, plants were grown in 8.9-cm by 8.9-cm square pots 3.5-inch by 3.5-inch square pots filled with BM 7 professional growing mix (Berger Peat Moss, Saint-Modeste, QC, Canada) and fertilized biweekly with a water-soluble 15:15:15 (N–P–K) fertilizer. Greenhouse temperatures were maintained at approximately 24 C during the day and 21 C at night under a 16-h photoperiod. Plants were allowed to produce seeds under pollination bags to prevent gene flow and were left to dry naturally on greenhouse benches for approximately 1 mo before seed harvest. Seeds were subsequently cleaned using a gravity separator. Once seed was cleaned, it was either used immediately in a light-quality germination experiment or placed into storage for later studies.
Germination Studies
Germination experiments were conducted on freshly harvested seed and then again after 2 yr of storage. The first germination assay for B. scoparia was conducted approximately 1 mo after seed harvest. Due to a necessary afterripening requirement, the first germination assay for C. album was conducted after 6 mo of dry storage followed by 2 wk of cold stratification at 4 C. Seeds not used in the initial germination experiments were stored at room temperature (C. album) or in a refrigerator at 4 C (B. scoparia).
Germination experiments employed a factorial treatment arrangement of light environment (3) and germplasm source (49 C. album accessions or 70 B. scoparia populations). The experiment was conducted as a single controlled-environment run with replication at the petri dish level (three replicates per treatment combination). Environmental conditions were tightly controlled to minimize variability. Light environments were generated using light-emitting diode panels under a 16-h photoperiod, and included:
-
1. very low fluence white light (VLF; hereafter referred to as “dark”): R:FR = 4.2; 0.007 to 0.094 μmol m⁻2 s⁻1, simulating VLFR primarily mediated by phyA;
-
2. low-fluence red light: R:FR = 5.3; 157 to 184 μmol m⁻2 s⁻1, inducing low-fluence responses (LFR) typically mediated by phyB; and
-
3. high-irradiance far-red light: R:FR = 0.0003; 29 to 32 μmol m⁻2 s⁻1, eliciting HIR mediated by phyA.
These classifications follow established phytochrome response definitions (Shichijo et al. Reference Shichijo, Katada, Tanaka and Hashimoto2001; Takaki Reference Takaki2001).
For each germplasm source and light environment, three replicate 10-cm-diameter polystyrene petri dishes (VWR International, Radnor, PA, USA), were prepared, each containing 30 seeds placed on two moistened sheets of circular blue Anchor germination paper (Anchor Paper, St Paul, MN, USA). Individual petri dishes served as experimental units and were randomly assigned to light treatments. Germination was recorded daily for 14 d, and germinated seeds were removed after counting. The initial germination experiment on freshly harvested seed established baseline germination responses of both species under contrasting light environments.
Far-red suppression of germination was calculated for each accession (Equation 1) using germination responses under red light (Treatment 2) and far-red light (Treatment 3). Specifically, far-red suppression was calculated by dividing the proportion of germinated seeds under far-red light (germfr) by the proportion of germinated seeds under red light (germr). The dark treatment (Treatment 1) was not included in this calculation, because it does not provide a directional phytochrome signal for comparison between active (Pfr) and inactive (Pr) states. If germination for a germplasm source is similar between red and far-red light environments, then far-red suppression is near 0. If germination is completely inhibited under far-red light, then far-red suppression approaches 1.
 $${\text{Far-red suppression}}\;{\rm{ = 1 - (germ}_{fr}}{\rm{/}}\;{\rm{ger}}{{\rm{m}}_{\rm{r}})}$$
$${\text{Far-red suppression}}\;{\rm{ = 1 - (germ}_{fr}}{\rm{/}}\;{\rm{ger}}{{\rm{m}}_{\rm{r}})}$$
For C. album, the germination study was repeated after 2 yr of dry storage using the same methods as were used for the freshly harvested seed, but with only the red light and far-red light treatments. This allowed us to compare two physiological states (freshly harvested vs. stored seed) for each germplasm source and estimate primary dormancy. Primary dormancy was estimated using a dormancy index (ΔRed), defined as the difference in final germination of stored seed (germstored) and freshly harvested seed (germfresh) when exposed to red light (Equation 2). We used germination under red light because it was the light environment most conducive to stimulation of C. album germination. If the proportion of seeds that germinate after storage is substantially greater than the proportion of seeds that germinate immediately after harvest, then ΔRed will be large, suggesting higher levels of primary dormancy. Lower values of ΔRed indicate lower levels of primary dormancy.
 $$\Delta {\rm{Red = Red \; germ}_{{\rm{stored}}}}{\rm{ - Red \; germ}_{\rm{fresh}}}$$
$$\Delta {\rm{Red = Red \; germ}_{{\rm{stored}}}}{\rm{ - Red \; germ}_{\rm{fresh}}}$$
To assess whether seed size influenced dormancy or light sensitivity, seeds from each C. album accession were subsampled using a pie-slice method to obtain representative samples (Copeland and McDonald Reference Copeland and McDonald2001). Subsamples were weighed using an analytical balance, and mean seed weight per accession was calculated from three replicates.
Statistical Analysis
All statistical analyses were conducted using R software (R Core Team 2024). The final germination proportion after 14 d was analyzed using a generalized linear mixed-effects model that included the effects of light environment and accession or population as fixed effects, and petri dish as a random effect. A binomial error distribution was used to account for the binomial nature of germination data. Type III ANOVA was conducted using Satterthwaite’s approximation for degrees of freedom, and estimated marginal means were obtained using the emmeans package (Lenth et al. Reference Lenth, Singmann, Love, Buerkner and Herve2018). Although germination dynamics over time can be analyzed using nonlinear regression models, our objective was to compare final germination responses among treatments; therefore, analyses focused on final germination proportion.
To examine whether primary dormancy was associated with sensitivity to far-red light, we used Spearman’s rank correlation analysis on the ΔRed and far-red suppression metrics across all accessions. Similarly, relationships between mean seed weight and dormancy indices (ΔRed) and far-red suppression were evaluated using Spearman’s rank correlation analysis. All visualizations were produced using ggplot2 (Wickham et al. Reference Wickham, Navarro and Pedersen2023), and statistical significance was evaluated at α = 0.05.
Results and Discussion
Germination Response of Bassia scoparia to Light Quality
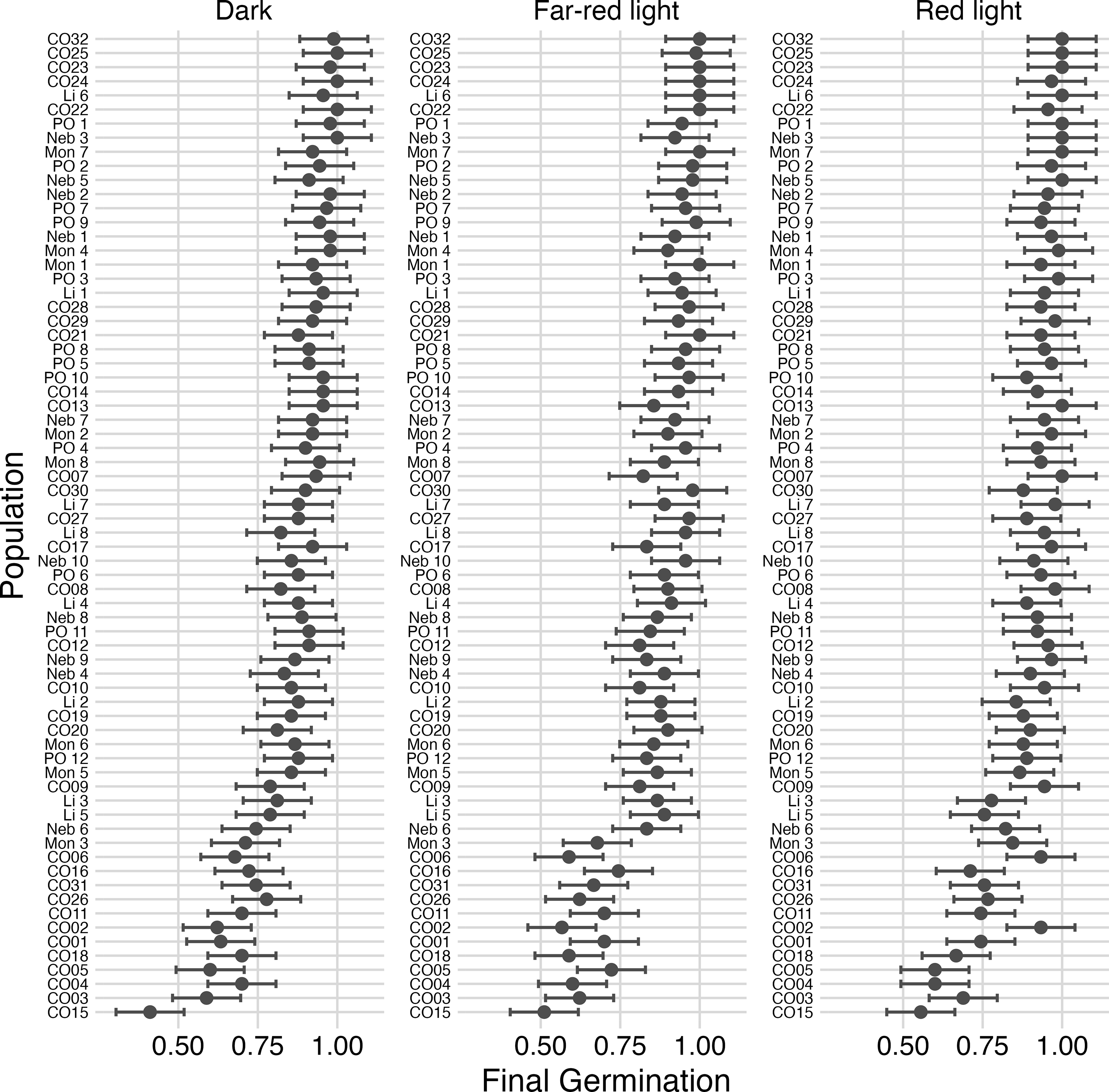
Germination of B. scoparia populations differed among light environments (P < 0.0001) and among populations (P < 0.0001), but the population by light interaction was not significant (P = 0.67; Figure 1). Across populations, mean final germination was 90.0% under red light, compared with 86.2% under dark conditions and 86.7% under far-red light. Germination under dark and far-red light did not differ, indicating that the response was not intermediate but rather characterized by a modest (∼3% to 4%) increase under red light. Despite these statistically detectable differences, germination remained uniformly high across all light environments, consistent with weak photoblastic regulation at the species level.
Germination proportion of Bassia scoparia populations under three light environments: far-red light, red light, and dark. Each point represents the mean final germination (± SE) for a population. Populations are ordered within each panel by final germination to facilitate visual comparison. Germination differed significantly among light environments (P < 0.0001) and among populations (P < 0.0001), whereas the population by light interaction was not significant (P = 0.67).

These results are consistent with previous reports showing that B. scoparia exhibits minimal light-dependent dormancy and readily germinates across a wide range of environmental conditions (Everitt et al. Reference Everitt, Alaniz and Lee1983; Maguire and Overland Reference Maguire and Overland1959). Bassia scoparia seeds are known to possess little primary dormancy and can germinate rapidly following dispersal, often within 24 h across a broad temperature range (Mohler et al. Reference Mohler, Teasdale and DiTommaso2019). The weak response to light quality observed here likely reflects an ecological strategy favoring rapid colonization of disturbed environments, where immediate germination provides a competitive advantage. Additionally, the relatively short seed longevity reported for B. scoparia may further reduce reliance on light-quality cues, as rapid germination may be favored over prolonged persistence in the soil seedbank (Mohler et al. Reference Mohler, Teasdale and DiTommaso2019). Collectively, these findings support the conclusion that light quality plays a limited role in regulating B. scoparia germination relative to other environmental cues.
Germination Response of Chenopodium album to Light Quality
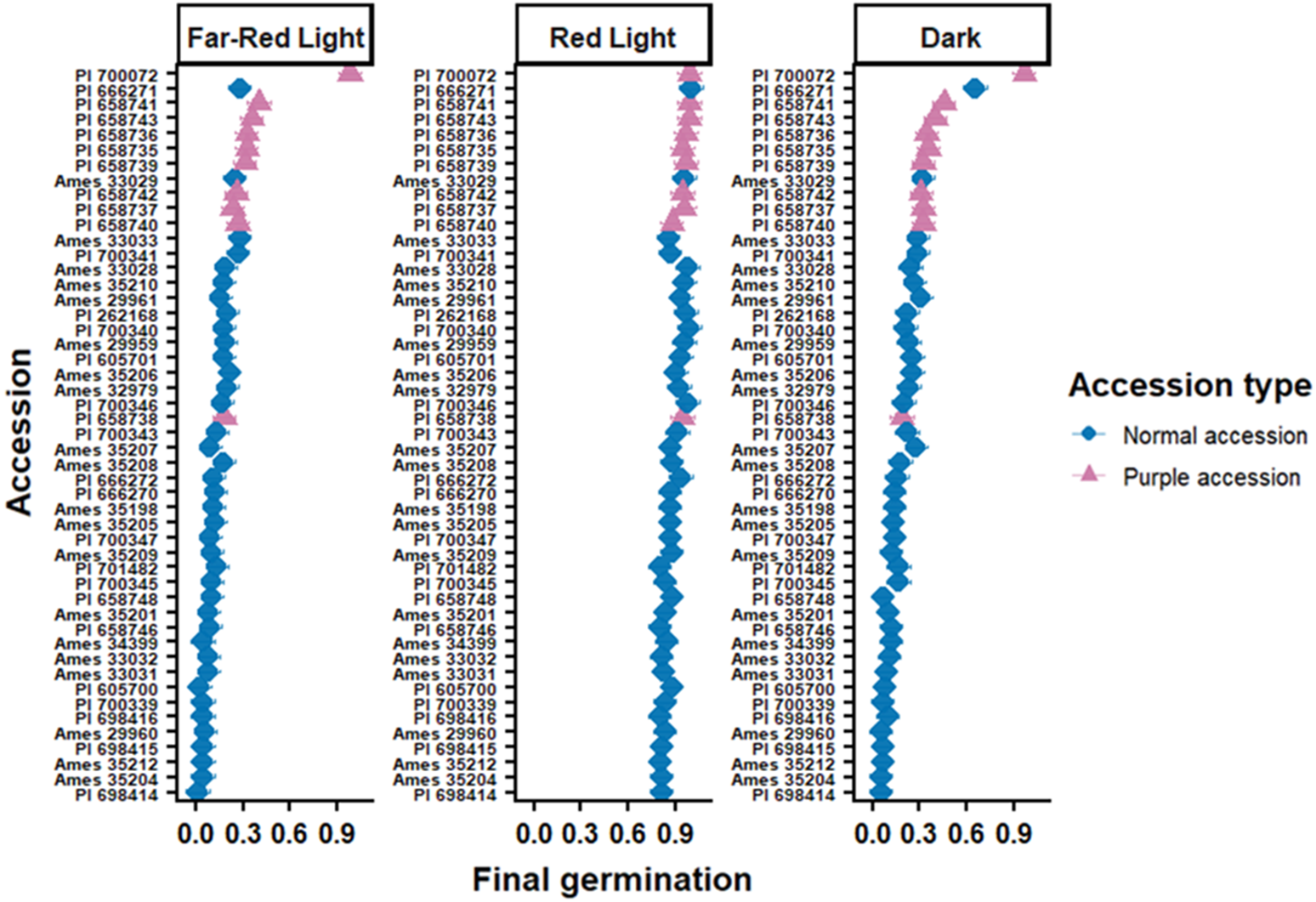
In contrast to B. scoparia, germination of C. album differed among accessions and light environments, with a significant accession by light interaction effect (P < 0.0001; Figure 2). Accessions exhibited a wide range of responses, from near-complete germination under red light with strong inhibition under far-red light to consistently low germination across all light environments. Accessions were further categorized based on vegetative pigmentation, with point color indicating leaf pigmentation: purple points represent accessions that produce purple-pigmented leaves, a trait associated with anthocyanin accumulation, whereas green points represent accessions with normal green leaves. This heterogeneity indicates substantial variation among accessions in light-dependent germination behavior. Maternal environmental conditions during seed development may also influence progeny germination responses to light quality, as demonstrated in other species (Causin Reference Causin2004). However, in this study, seed increase was conducted under a common greenhouse environment to minimize maternal environmental variation among accessions, although subtle effects cannot be completely excluded.
Germination proportion of Chenopodium album accessions under three light environments: red light, far-red light, and dark. Each point represents the accession-level mean final germination proportion (±SE) calculated across three replicate petri dishes (30 seeds per replicate). Accessions are ordered within each panel from lowest to highest germination to facilitate comparison of accession-specific responses. Point colors indicate leaf pigmentation phenotype (purple vs. non-purple) for each accession. Germination was significantly affected by light environment (P < 0.0001) and by the accession by light interaction (P < 0.0001), indicating substantial genetic variation in light-dependent germination responses.

These patterns are consistent with well-established phytochrome-mediated control of germination in small-seeded, photoblastic species. Seeds of Chenopodium and related genera preferentially germinate under high R:FR ratios characteristic of open soil surfaces, whereas far red–enriched light associated with canopy shade maintains dormancy (Botto et al. Reference Botto, Sánchez, Whitelam and Casal1996; Taylorson Reference Taylorson2008). Similar responses have been reported in Palmer amaranth (Amaranthus palmeri S. Watson), where afterripened seeds require red or natural light for germination and are inhibited by far-red light or darkness (Jha et al. Reference Jha, Norsworthy, Riley and Bridges2010). Previous work has also demonstrated that in C. album, red light promotes germination and interacts positively with temperature, stratification, and nitrate availability (Tang et al. Reference Tang, Guo, Yin, Ding, Xu, Wang, Yang, Xiong, Zhong, Tao and Sun2022). Notably, accession PI 700072 deviated strongly from this general pattern, exhibiting consistently high germination (>80%) across all light environments, including far-red light and dark conditions. This lack of light-dependent inhibition distinguishes PI 700072 as an outlier within the germplasm and indicates a markedly reduced dependence on phytochrome-mediated light cues for germination. Chenopodium album is known to produce polymorphic seeds differing in seed coat color (brown and black), and this somatic polymorphism has been shown to influence dormancy depth and responsiveness to environmental cues, including light and temperature (Williams and Harper Reference Williams and Harper1965; Yao et al. Reference Yao, Lan and Zhang2010). Seed polymorphism was observed only in accession PI 700072. The light-insensitive germination behavior of PI 700072 may therefore reflect reduced dormancy depth or altered dormancy heterogeneity. However, seed morph composition was not quantified in this study, and its contribution to germination responses could not be assessed.
Together, these results indicate that while C. album generally exhibits strong light-dependent germination, individual accessions such as PI 700072 can display near-complete release from light regulation, highlighting substantial genetic and physiological diversity in dormancy control that may influence emergence under shaded, residue-covered, or buried field conditions.
Relationship between Primary Dormancy and Far-Red Light–enforced Inhibition
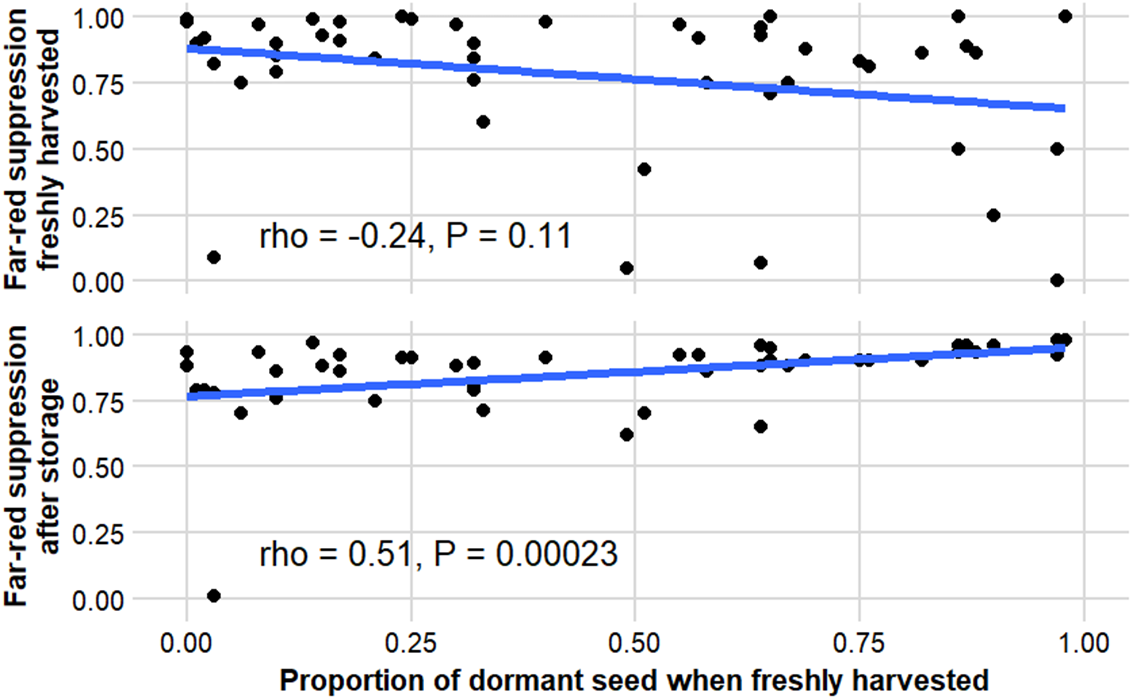
Primary dormancy varied widely among C. album accessions, as indicated by the dormancy index (ΔRed), which ranged from near zero to >0.6 (Figure 3). Accessions with low initial germination under red light exhibited the greatest increase after storage, confirming that dry afterripening effectively alleviated primary dormancy. In contrast, accessions that germinated readily when freshly harvested showed little change following storage, indicating minimal dormancy at dispersal.
Relationship between primary dormancy and far-red light suppression in Chenopodium album accessions. Primary dormancy was quantified as ΔRed (Redstored − Redfresh). Far-red suppression was calculated as 1 − (FR/Red). The upper panel shows freshly harvested seeds, and the lower panel shows seeds after afterripening. Each point represents an accession; lines indicate fitted trends.

The relationship between primary dormancy and far-red suppression depended on seed physiological status. In freshly harvested seeds, the association was weak and negative, whereas it became positive after storage, indicating that accessions with stronger primary dormancy also exhibited greater sensitivity to far-red inhibition following afterripening (Figure 4). This coordination suggests shared phytochrome-mediated regulation of dormancy depth and light sensitivity (Crepy and Casal Reference Crepy and Casal2015; Takaki Reference Takaki2001). Dormant seeds require higher proportions of the active phytochrome form (Pfr) to initiate germination and therefore respond primarily to light with high R:FR ratios typical of exposed soil surfaces (Benech-Arnold et al. Reference Benech-Arnold, Sánchez, Forcella, Kruk and Ghersa2000). Far red–enriched light characteristic of canopy shade or deeper burial favors the inactive Pr form, maintaining dormancy (Botto et al. Reference Botto, Sánchez, Whitelam and Casal1996).
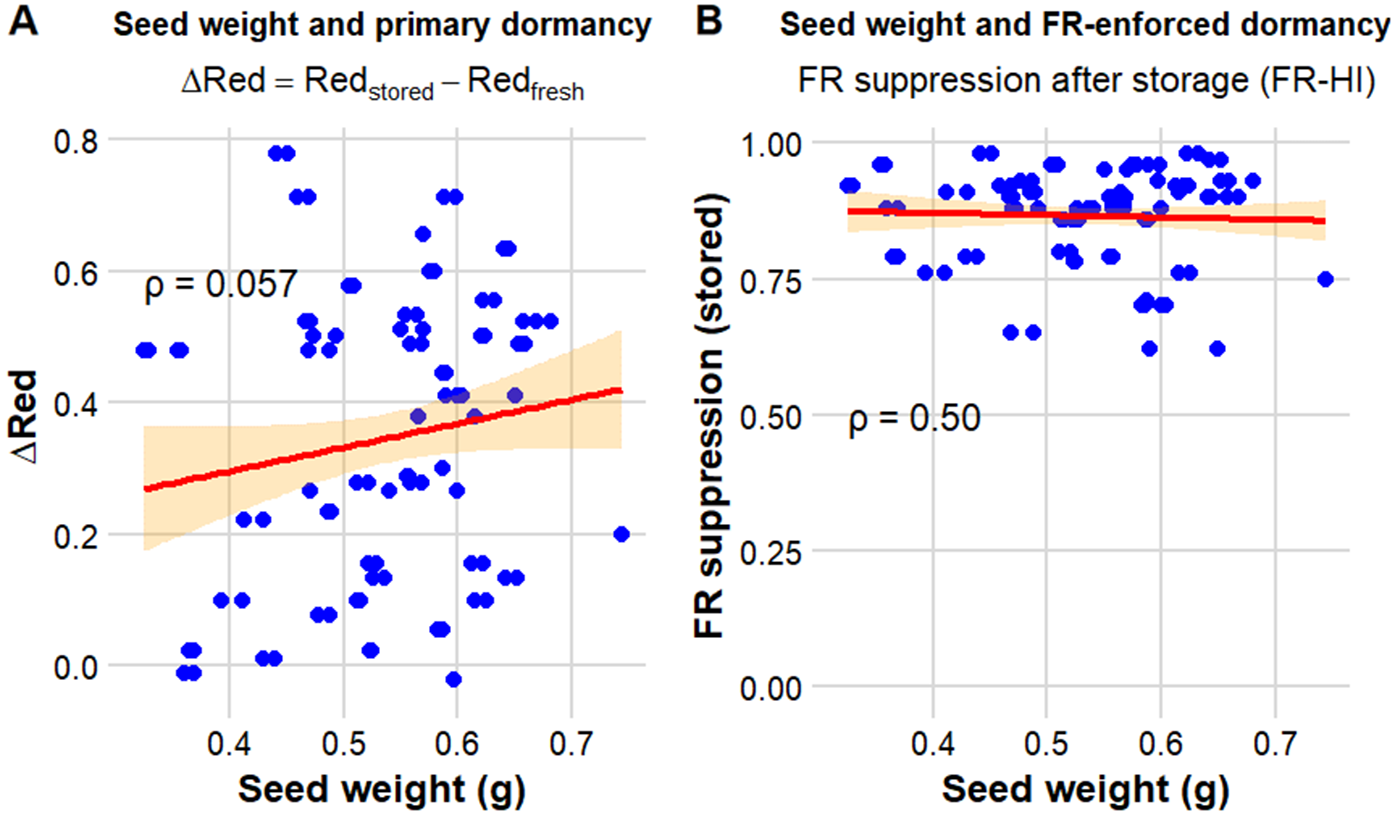
Relationships between seed weight and dormancy traits in Chenopodium album. (A) Relationship between seed weight and primary dormancy, expressed as ΔRed (Redstored − Redfresh). (B) Relationship between seed weight and far-red suppression after storage, calculated as 1 − (FR/Red). Each point represents an accession; red lines indicate fitted trends, and orange-shaded areas represent 95% confidence intervals.

From an ecological perspective, the coordination between primary dormancy and far-red sensitivity represents a bet-hedging strategy that promotes persistence in variable soil seedbanks (Vleeshouwers et al. Reference Vleeshouwers, Bouwmeester and Karssen1995). Accessions with strong dormancy and high far-red inhibition remain quiescent under shaded or buried conditions, whereas with weaker dormancy germinate rapidly when favorable microsites become available. Such variation likely contributes to the success of C. album across heterogeneous agricultural environments.
Relationship between Seed Size, Primary Dormancy, and Far-Red Inhibition
Seed weight was not associated with dormancy-related traits in C. album (Figure 4). The correlation between seed weight and primary dormancy (ΔRed) was not significant (Spearman ρ = 0.057, P = 0.11). Likewise, seed weight was not related to far-red suppression after storage (ρ = 0.50, P = 0.06), indicating no consistent size-dependent pattern in these responses.
These results indicate that seed size alone is a poor predictor of either primary dormancy or far-red–enforced inhibition in C. album. Although larger seeds may theoretically sustain dormancy for longer periods due to greater carbohydrate reserves, seed weight was not significantly associated with dormancy indices or far-red suppression in this study. Instead, the wide range of dormancy responses observed among seeds of similar mass suggests that dormancy regulation in C. album is governed primarily by physiological and genetic factors rather than by morphological provisioning alone (Rees Reference Rees1996; Yao et al. Reference Yao, Lan and Zhang2010). Similar conclusions have been reported across diverse herbaceous floras, where seed weight did not reliably predict germination responses to R:FR ratios (Jankowska-Blaszczuk and Daws Reference Jankowska-Blaszczuk and Daws2007). At the genetic level, natural variation in phytochrome-mediated light perception has been shown to underlie differences in dormancy expression among ecotypes (Taylor et al. Reference Taylor, Cooper, Sellamuthu, Braun, Migneault, Browning, Perry and Schmitt2017). This study provides additional evidence that variation in light-dependent dormancy and germination in C. album is likely driven more by regulatory mechanisms than by seed size.
Bassia scoparia populations exhibited a consistent germination response across light environments, indicating weak photoblastic regulation and minimal influence of light quality on dormancy expression in this species. In contrast, C. album accessions showed pronounced population-level variation in light-dependent germination, reflecting substantial genetic and physiological diversity in dormancy and light sensitivity across accessions originating from diverse geographic regions (Supplementary Table S1). The positive association between primary dormancy and far-red light–enforced inhibition after afterripening suggests coordinated phytochrome-mediated regulation of dormancy depth and light responsiveness.
Seed weight was not significantly associated with either primary dormancy or far-red light–enforced inhibition, indicating that seed size alone does not reliably predict dormancy expression or light-mediated germination responses in C. album. Instead, variation in germination behavior appears to be governed primarily by regulatory mechanisms controlling light perception and dormancy cycling. These findings highlight the role of light quality and dormancy diversity in shaping emergence patterns of C. album, contributing to its persistence across variable canopy and soil environments in agricultural systems.
A limitation of this study is the lack of temporal replication across independent experimental runs. Although environmental conditions were tightly controlled, additional replication across time would strengthen inference regarding environmental variability.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/wsc.2026.10121
Acknowledgments
The Chenopodium album seed was acquired from the USDA Germplasm Resources Information Network (USDA-GRIN). We thank Todd Gaines (Colorado State University) for providing Bassia scoparia populations collected in Colorado. The authors used ChatGPT (OpenAI) to assist with language editing and manuscript clarity. All scientific content, data analysis, interpretation, and conclusions were developed, verified, and approved by the authors.
Funding statement
This research was supported by Hatch Research Capacity Fund WYO-631-22 from the USDA National Institute of Food and Agriculture.
Competing interests
The authors declare no competing interests.



 Open access
Open access