Non-technical Summary
Cambrian trilobites have been known from southern Arizona since 1904 when reported by Charles Walcott and subsequently reported by other geologists and paleontologists. However, they have rarely been illustrated, except by Stoyanow in 1936 and subsequently a few specimens by Muhlbauer and colleagues in 2025. Illustrated and discussed here are 19 species from the Abrigo Formation of the Santa Catalina Mountains. These exposures consist of sandy, shallow-water rocks that differ markedly from the deeper-water deposits farther to the south. The contained trilobites indicate a traditional middle-late Cambrian age.
Introduction
Recent work that integrates biostratigraphy with chemostratigraphy and tandem U-Pb detrital zircon geochronology has helped refine our understanding of rates of evolution, extinction, and environmental change in Cambrian systems (e.g., Sundberg et al., Reference Sundberg, Karlstrom, Geyer, Foster, Hagadorn, Mohr, Schmitz, Dehler and Crossey2020; Dehler et al., Reference Dehler, Sundberg, Karlstrom, Crossey, Schmitz, Rowland and Hagadorn2024). Nowhere have these revisions been more pronounced than along the southwestern margin of Laurentia (modern-day coordinates), where global transgressions and durations of deposition are shown to be rapid and short-lived (e.g., Karlstrom et al., Reference Karlstrom, Mohr, Schmitz, Sundberg and Rowland2020; Sundberg et al., Reference Sundberg, Cothren and Dehler2024); cratonic syndepositional tectonics appears to be more influential than previously thought (e.g., Karlstrom et al., Reference Karlstrom, Hagadorn, Gehrels, Matthews and Schmitz2018); and the durations of global changes in seawater chemistry (e.g., Howley and Jiang, Reference Howley and Jiang2010; Cothren et al., Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022; Rowland et al., Reference Rowland, Korolev, Hagadorn and Ghosh2023; Farrell et al., Reference Farrell, Cothren, Sundberg, Schmitz, Dehler, Landing, Karlstrom, Crossey and Hagadorn2025) and environments are tied to biological and biostratigraphic changes (e.g., Lin et al., Reference Lin, Sundberg, Jiang, Montañez and Wotte2019; Sundberg et al., Reference Sundberg, Karlstrom, Geyer, Foster, Hagadorn, Mohr, Schmitz, Dehler and Crossey2020). A unifying element of these advances is the integration of clastic and carbonate records with intensive fossil collecting, particularly in successions in which fossils are scarce (see synthesis by Sundberg et al., Reference Sundberg, Foster, Webster, Miller and Hagadorn2026). With this context in mind, we augment historic (Gilluly, Reference Gilluly1956; Hayes, Reference Hayes1975) and emerging work (Łabaj, Reference Łabaj2015; Łabaj and Pratt, Reference Łabaj and Pratt2016; Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025) on the more fossiliferous, mixed siliciclastic-carbonate succession of southern Arizona by reporting new fossils from the Santa Catalina Mountains that allow further biostratigraphic zonation of the Cambrian strata of southern Arizona, and more accurate integration of these strata in to the global record of Cambrian Earth history.
Geologic context
The middle-late Cambrian Abrigo Formation (~200 m thick) is a mixed siliciclastic-carbonate succession of shallow-water, low-gradient, coastal sediments that were deposited along the present-day southern margin of Laurentia, at ~10° paleosouth (Fig. 1.1; see overviews by Hayes, Reference Hayes1975, Reference Hayes, Callender, Witt, Clemons and James1978; Page et al., Reference Page, Harris, Repetski, Derby, Fritz, Longacre, Morgan and Sternbach2012; Sundberg and Cuen-Romero, Reference Sundberg and Cuen-Romero2021). The Abrigo Formation is conformably underlain by the Bolsa Quartzite and is unconformably overlain by the Devonian Martin Formation. The Bolsa-Abrigo succession was deposited during the Sauk transgression, recording the transition from predominantly continental cratonic environments to inner-shelf marine environments, with the Abrigo Formation indicating deposition in lower offshore to upper shoreface settings (Łabaj and Pratt, Reference Łabaj and Pratt2016; Sundberg and Cuen-Romero, Reference Sundberg and Cuen-Romero2021). Recent CA-ID-TIMS U-Pb detrital zircon geochronology from Bolsa Quartzite and Abrigo Formation outcrops to the south at French Joe Canyon, in the Whetstone Mountains of Arizona (Fig. 1.2) indicates that the lower two members of the Abrigo Formation are ≤ 502.40 ± 0.55 Ma and part of the upper member is ≤ 488.34 ± 0.59 Ma (Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025). Previous work reports trilobites from the Bolaspidella through Aphelaspis biozones (Łabaj, Reference Łabaj2015; Łabaj and Pratt, Reference Łabaj and Pratt2016; Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025). Correlative carbon isotope values from this same section are consistent with DICE and TOCE records, but the SPICE record is absent due to erosion at the Sauk II-III unconformity (Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025 and references therein).
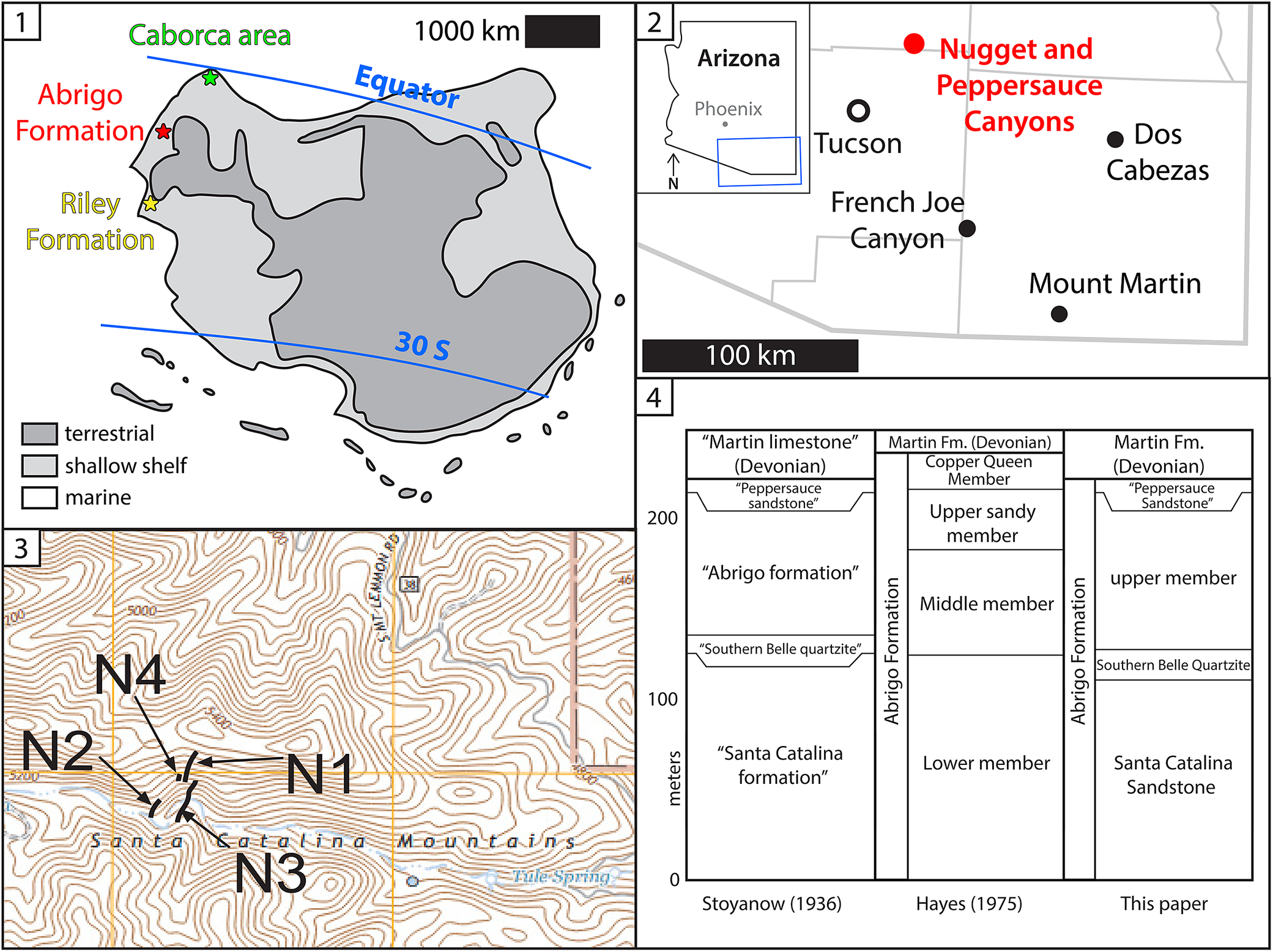
Geologic background for the Abrigo Formation and Nugget Canyon section. (1) Paleogeographic map of Laurentia during the Miaolingian (~500 Ma), indicating the subequatorial setting in which the Abrigo Formation and comparable units were deposited. (2) Simplified regional map with county lines adapted from Łabaj (Reference Łabaj2015), with the locations of Nugget Canyon and Peppersauce Canyon (red dot) and other relevant localities (black dots). Blue inset depicts the location of the map in Arizona, USA. (3) Topographic map (7.5’ Campo Bonito, Arizona, 2021) illustrating the location of the individual sections N1, N2, N3, and N4 (see Appendix) measured for this study. (4) Comparisons of terminology for the Abrigo Formation (Fm.) and its members, for clarification.

Figure 1. Long description
Panel 1 is a paleogeographic map of Laurentia during the Miaolingian. The landmass is centered between the Equator and 30 degrees South. A legend indicates terrestrial areas in dark gray, shallow shelf in light gray, and marine areas in white. Three stars on the Northwest coast mark the Caborca area in green, Abrigo Formation in red, and Riley Formation in yellow. A 1000 kilometer scale bar is at the top right.
Panel 2 is a regional map of Arizona with a 100 kilometer scale bar. A red dot in the North marks Nugget and Peppersauce Canyons. Black dots mark Tucson and French Joe Canyon to the Southwest, and Dos Cabezas and Mount Martin to the Southeast. An inset shows the map's location in Southeast Arizona.
Panel 3 is a topographic map of the Santa Catalina Mountains. Four black labels with arrows point to specific measured sections: N 1 and N 3 are located on the East side of a ridge, while N 2 and N 4 are on the West side.
Panel 4 is a stratigraphic chart with a vertical axis in meters from 0 to over 200. It compares three terminologies.
* Stoyanow Reference Stoyanow1936: Santa Catalina formation at the base, followed by Southern Belle quartzite, Abrigo formation, Peppersauce sandstone, and Martin limestone.
* Hayes Reference Hayes1975: Abrigo Formation divided into Lower, Middle, Upper sandy, and Copper Queen members, topped by Martin Fm. Devonian.
* This paper: Santa Catalina Sandstone at the base, followed by Southern Belle Quartzite, Abrigo Formation upper member, Peppersauce Sandstone, and Martin Fm. Devonian.
This report focuses on exposures of the Abrigo Formation along the sides of Nugget Canyon in the Santa Catalina Mountains. Unlike other sections in southern Arizona and Sonora, this area contains mostly siliciclastic strata and fewer carbonates; as such, it has unique member-level stratigraphic terminology and history that we briefly review here.
The first trilobites reported from the Abrigo Formation were from Mount Martin (Walcott in Ransome, Reference Ransome1904) and suggest a middle Cambrian age and correlation with Texas strata. The first described and figured trilobites from the unit were by Stoyanow (Reference Stoyanow1936), who named the trilobites Hesperaspis ransomei Stoyanow, Reference Stoyanow1936 and H. butleri Stoyanow, Reference Stoyanow1936 from exposures in Peppersauce Canyon. Nugget Canyon, which is the adjacent canyon to Peppersauce Canyon, is the focus of this contribution. Trilobites have also been reported (but not figured) from more southern exposures of the Abrigo Formation at French Joe Canyon in the Whetstone Mountains and at Bisbee in the Mule Mountains and at Ajax Hill near Tombstone, south of Black Diamond Peak in the Dragoon Mountains and near Seven Dash Ranch in the Little Dragoon Mountains and the Whitehead Ranch in the Swisshelm Mountains (Palmer in Gilluly, Reference Gilluly1956). Palmer (in Cooper and Silver, Reference Cooper and Silver1964) reported trilobites from Rattlesnake Ridge, Johnny Lyon Hills, and the Little Dragoon Mountains that might possibly represent the Ehmaniella and Cedaria, Crepicephalus, Aphelaspis, Elvinia, and Ptychaspis biozones. Taylor (in Hayes, Reference Hayes1975) also identified trilobites from other exposures of the Abrigo Formation in southern Arizona, including the Swisshelm Mountains, Whetstone Mountains, Waterman Mountains, Slate Mountains, and at Mount Martin and Picacho de Calera as well as from strata in New Mexico and Texas. These fossils represent the Bolaspidella to Crepicephalus biozones, as well as post-Elvinia biozones.
These earlier contributions employed a different stratigraphic nomenclature than is used today. For example, Stoyanow (Reference Stoyanow1936) used the term Troy Quartzite to describe what is now the Bolsa Quartzite. Overlying that unit, he reported the Santa Catalina Sandstone (= lower Abrigo Formation), Southern Bell Quartzite (= middle Abrigo Formation), Abrigo Formation (= upper Abrigo Formation), and Peppersauce Sandstone (= uppermost Abrigo Formation). Later, Hayes (Reference Hayes1972, Reference Hayes1975) redefined the Abrigo Formation as having lower, middle, and upper sandy members capped by the dolomitic sandstones of the Copper Queen Member (= Peppersauce Sandstone of Stoyanow, Reference Stoyanow1936). Here, we follow Muhlbauer et al. (Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025) in employing a hybrid approach to defining the succession in the Santa Catalina Mountains. We use the terms Bolsa Quartzite and Abrigo Formation, and in the latter recognize the following members: Santa Catalina Sandstone, Southern Bell Quartzite, upper member, and Peppersauce Sandstone (Fig. 1.4).
The Copper Queen Member as used by Hayes (Reference Hayes1972, Reference Hayes1975) in Nugget Canyon is not used herein. Trilobites from the base to the top of the Copper Queen Member at Mount Martin near the southern Arizona border (Palmer in Gilluly, Reference Gilluly1956; Palmer in Cooper and Silver, Reference Cooper and Silver1964; Taylor in Hayes, Reference Hayes1975) include: Drumaspis sp. indet., Saratogia (Idahoia Walcott, Reference Walcott1924) sp. indet., Maladia sp. indet., Wilbernia sp. indet., and Ptychaspis sp. indet. These taxa occur in the Ellipsocephaloides Biozone (Longacre, Reference Longacre1970; Westrop, Reference Westrop1986; Hegna, Reference Hegna2006; Sundberg et al., Reference Sundberg, Cothren and Dehler2024). Stage 10 strata are present in the Copper Queen Member, consistent with a detrital zircon MDA at French Joe Canyon that constrains the upper two meters of Abrigo Formation to < 488.34 ± 0.59 Ma, within Stage 10 (Farrell et al., Reference Farrell, Cothren, Sundberg, Schmitz, Dehler, Landing, Karlstrom, Crossey and Hagadorn2025; Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025).
Preservation
The majority of the trilobite specimens are preserved as molds and casts in medium- to fine-grained sandstone. They are generally preserved with hematite or limonite coatings, and external ornamentation is rarely preserved in detail. In addition, several specimens have been compressed and show tectonic deformation. Due to the absence of surface ornamentation and some associated sclerites, along with tectonic deformation and compression, specific identification is not possible for several specimens.
Methods
The majority of fossils reported here were collected during the winter of 1996/1997 from Nugget Canyon by the lead author and are reposited at the Sam Noble Museum at the University of Oklahoma (OU). Additional material, collected by Alexander Stoyanow from the adjacent Peppersauce Canyon north of Nugget Canyon, was examined for this study and is reposited at the Los Angeles County Natural History Museum (LACNHM). All specimens were mechanically prepared, blackened with colloidal graphite (unless otherwise noted), whitened with ammonium chloride sublimate, and photographed with a Nikon D70S camera and AF NIKKOR 50 mm ƒ/1.4D lens with macrophotography bellows.
Repositories and institutional abbreviations
OU, Sam Noble Museum, University of Oklahoma, Norman, USA; LACMIP, Natural History of Los Angeles County Museum, Invertebrate Paleontology, Los Angeles, California, USA; USNM, National Museum of Natural History [United States National Museum], Smithsonian Institution, Washington, DC, USA.
Systematic paleontology
Order Agnostida Salter, Reference Salter1864
Family Agnostidae M’Coy, Reference M’Coy1849
Subfamily Agnostinae M’Coy, Reference M’Coy1849
Agnostidae gen. indet. sp. indet.
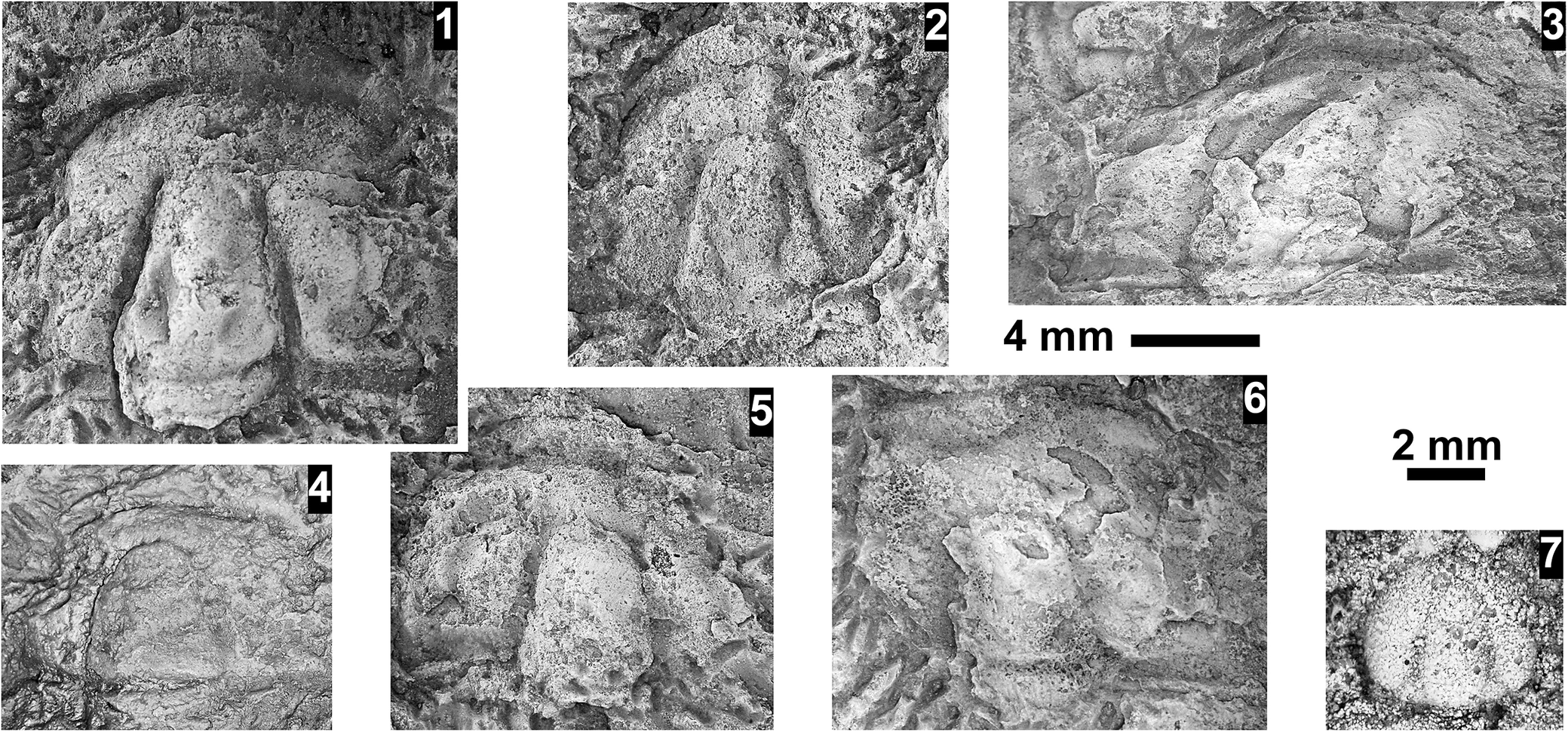
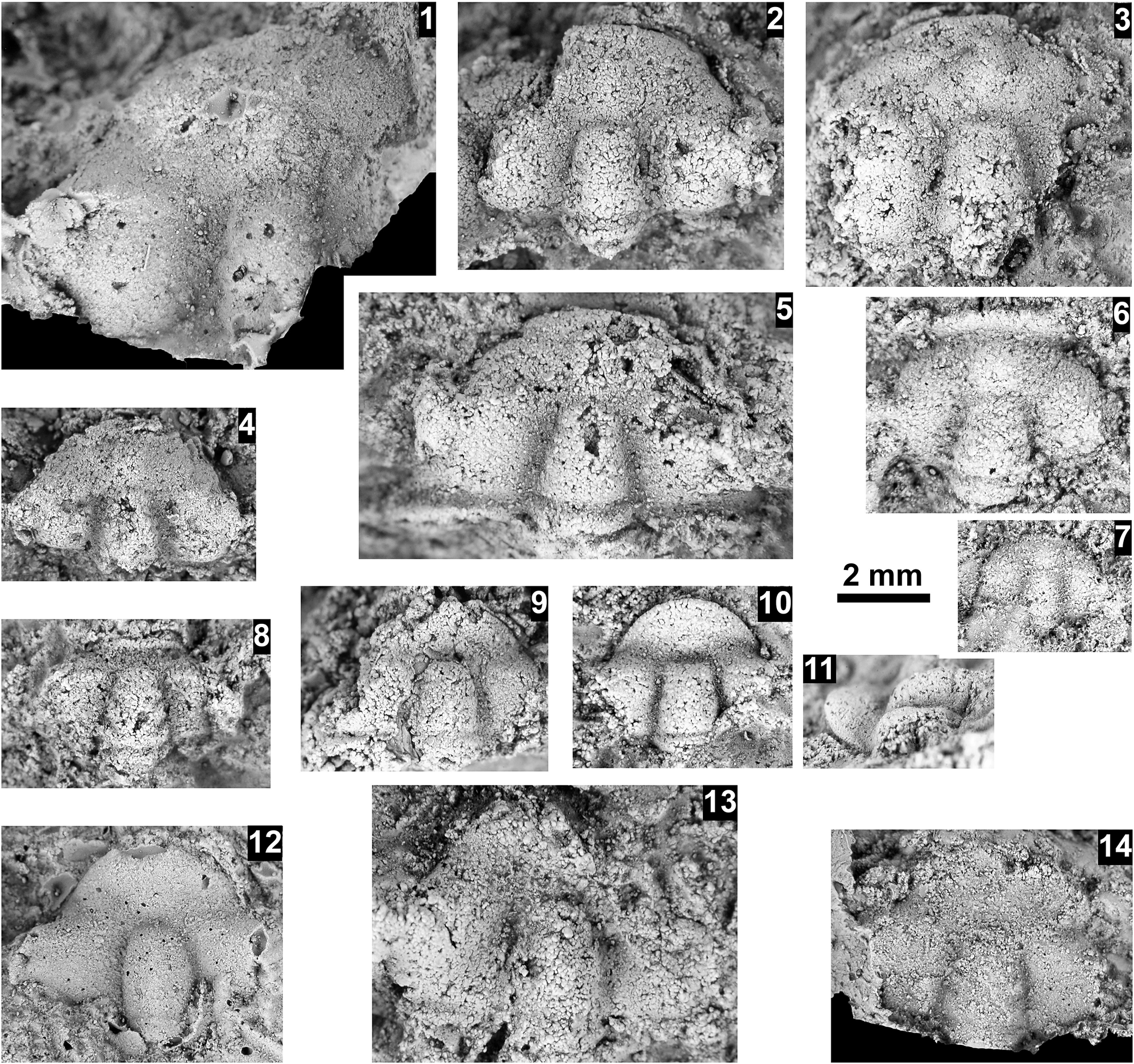
Ehmaniella sp. indet. and Agnostidae gen. indet. sp. indet. from the Abrigo Formation, Nugget Canyon, Arizona, USA. (1–6) Ehmaniella sp. indet., all preserved in sandstone, tectonically distorted, from OU loc. IP 01804, Santa Catalina Member: (1) partial cranidium OU 239040; (2) partial cranidium OU 239041; (3) partial cranidium OU 239042; (4) partial cranidium OU 239043; (5) partial cranidium OU 239044; (6) partial cranidium OU 239045. (7) Agnostidae gen. indet. sp. indet., cephalon OU 239046 from OU loc. IP 01821, upper Abrigo Formation. Scale bars = 4 mm (1–6); 2 mm (7).

Figure 2. Long description
A multi-panel plate containing seven grayscale photographs of fossilized trilobite parts embedded in textured sandstone.
* Panel 1 at the top left shows a partial cranidium with a distinct central glabella and a curved anterior border.
* Panel 2 to the right of 1 shows a smaller, more weathered cranidium.
* Panel 3 at the top right displays a wide, distorted cranidium fragment.
* Below these, a 4 millimeter scale bar is positioned horizontally.
* Panel 4 at the bottom left shows a cranidium with a smooth, rounded glabella.
* Panel 5 in the lower middle shows a cranidium with a prominent, elongated central lobe.
* Panel 6 to the right of 5 shows a larger, flattened cranidium fragment with visible surface pitting.
* Panel 7 at the bottom right is a much smaller, circular cephalon specimen.
* A 2 millimeter scale bar is located above panel 7.
All specimens in panels 1 through 6 exhibit tectonic distortion and are preserved in a coarse-grained matrix.
Occurrence
Cedaria Biozone, upper Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01821).
Remarks
A single cephalon with the glabella with basal glabellar lobes, rounded frontal lobe that does not reach the anterior border, moderate fibular furrow, and without preglabellar median furrow similar to Baltagnostus Lochman in Lochman and Duncan, Reference Lochman and Duncan1944; Itagnostus Öpik, Reference Öpik1979; and Peronopsis Corda in Hawle and Condra, Reference Hawle and Corda1847. However, without a pygidium, generic identification is not possible.
Class Trilobita Walch, Reference Walch1771
Order Ptychopariida Swinnerton, Reference Swinnerton1915
Family Alokistocaridae Resser, Reference Resser1939
Subfamily Alokistocarinae Hupé, Reference Hupé1955
Genus Ehmaniella Resser, Reference Resser1937
Type species
Crepicephalus (Loganellus Devine, Reference Devine1863) quadrans Hall and Whitfield, Reference Hall and Whitfield1877, Ute Formation (by original designation).
Ehmaniella sp. indet.
Occurrence
Ehmaniella? to Bolaspidella biozones, Santa Catalina Member, Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01803, 01804).
Remarks
The specimens from Nugget Canyon are preserved in quartzite and are tectonically deformed and flattened, which makes their identification questionable. However, given their tapering glabella with a truncated frontal lobe and their relatively long palpebral lobes, wide fixigena, and wide, narrow posterior area of the fixigena, they resemble Ehmaniella (e.g., Ehmaniella waptaensis Rasetti, Reference Rasetti1951; see Sundberg, Reference Sundberg1994, fig. 37). The absence of pygidia and well-preserved librigena prevents specific identification.
Genus Neoblairella Özdikmen, Reference Özdikmen2006
Type species
Blairella crassimarginata Rasetti, Reference Rasetti1965a, Pleasant Hill Formation, Pennsylvania, USA.
Remarks
The genus name Blairella Rasetti, Reference Rasetti1965a for trilobites from the Pleasant Hill Formation, Pennsylvania was preoccupied by the bivalve named by Miller and Gurley (Reference Miller and Gurley1896). Özdikmen (Reference Özdikmen2006, p. 186) replaced the occupied name with Neoblairella assigning the two species of trilobites named to Blairella by Rasetti (Reference Rasetti1965a) to this genus.
Neoblairella sp. indet.
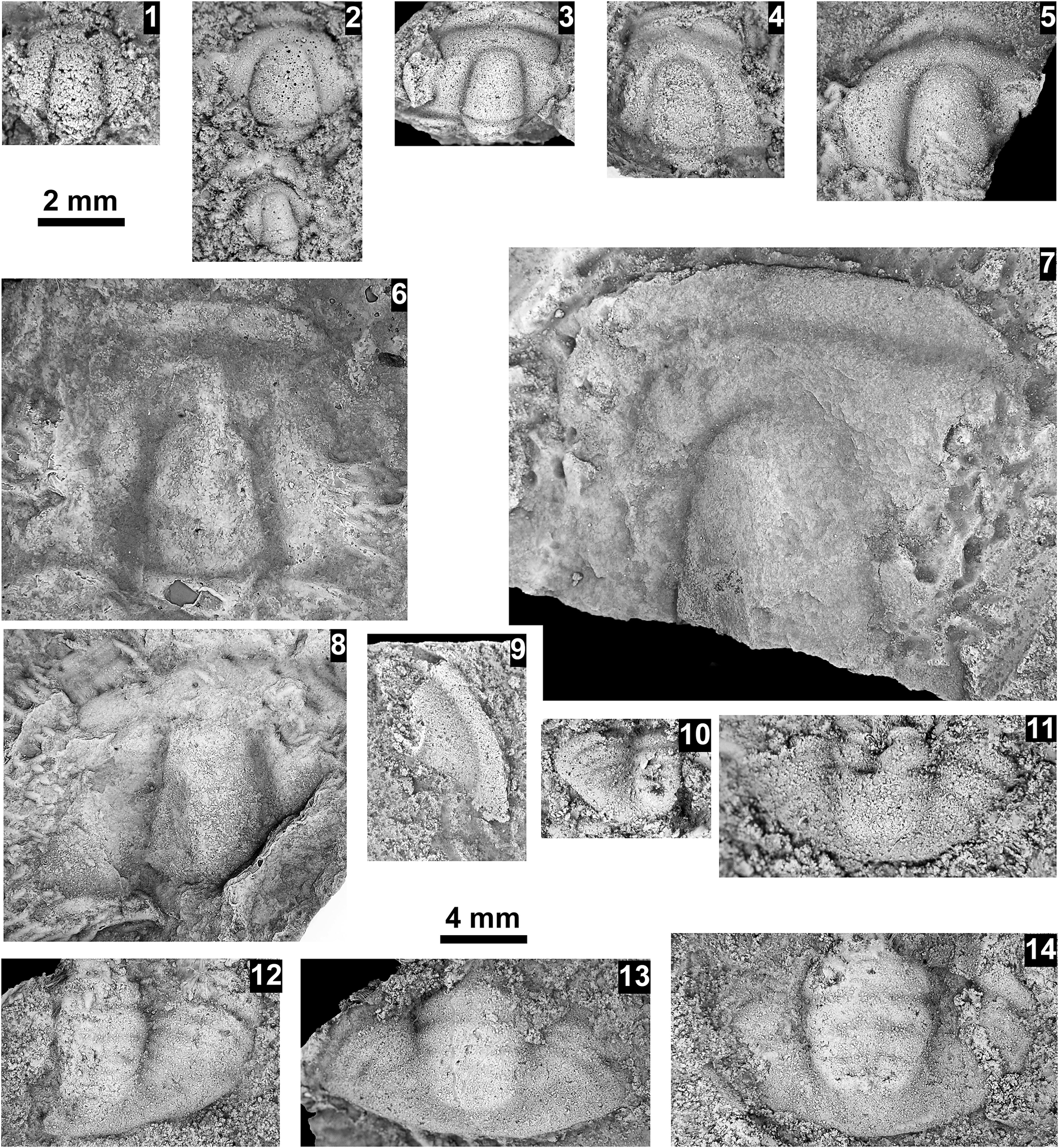
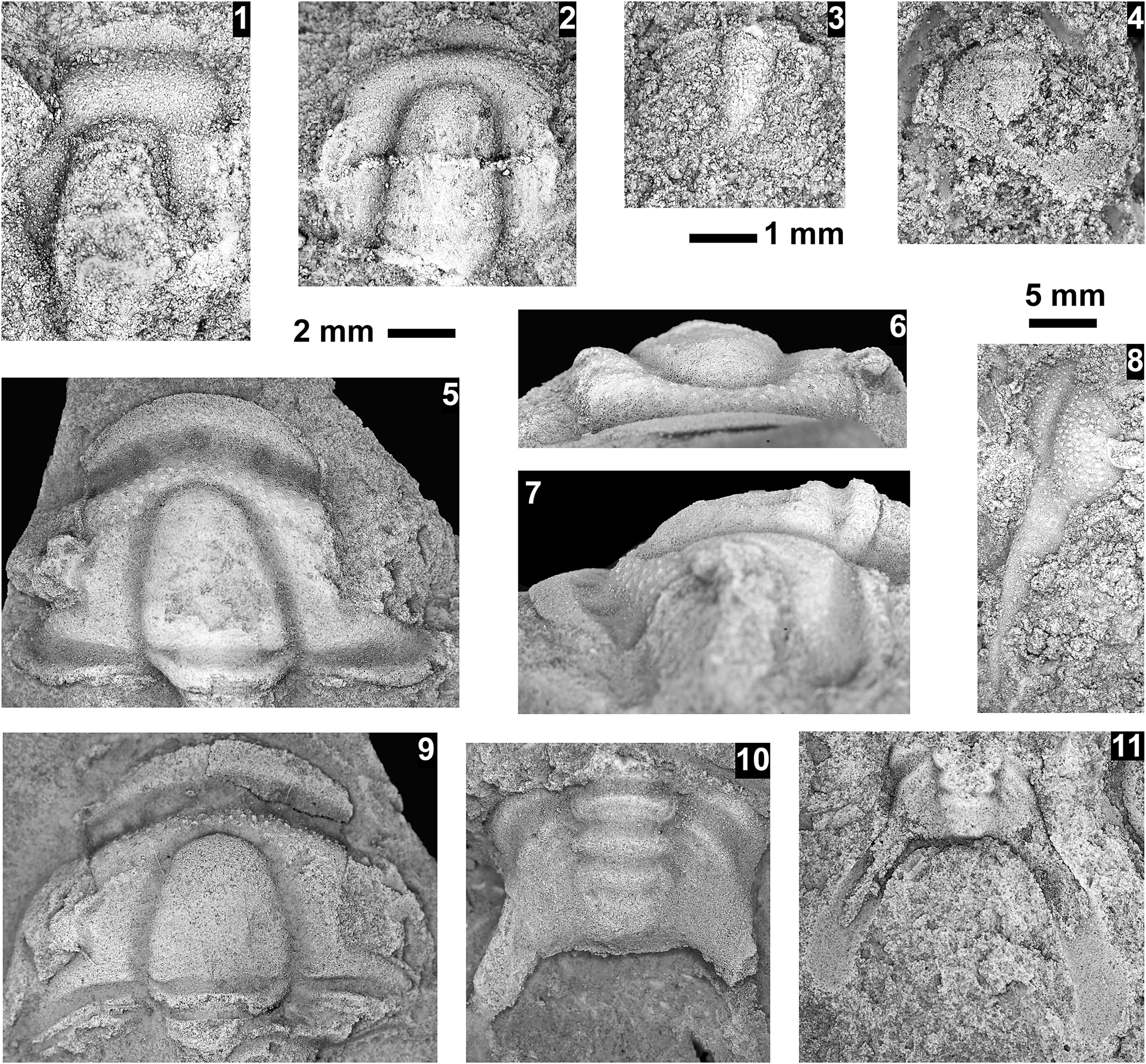
Neoblairella sp. indet., Santa Catalina Member, Abrigo Formation, Nugget Canyon, Arizona, USA; all specimens preserved in sandstone: (1) partial small cranidium OU 239047, internal mold from OU loc. IP 01806; (2) partial cranidia OU 239048 (upper), 239049 (lower), latex mold, from OU loc. IP 01810; (3) mostly complete cranidium OU 239050, internal mold from OU loc. IP 01821; (4) distorted, partial cranidium OU 239051, latex mold, from OU loc. IP 01808; (5) partial cranidium OU 239052, latex mold, OU loc. IP 01805; (6) partial, flattened cranidium OU 239053, latex mold, OU loc. IP 01808; (7) partial cranidium OU 239054, internal mold, from OU loc. IP 01812; (8) partial, flattened cranidium OU 239055, latex mold, OU loc. IP 01821; (9) librigena OU 239056, internal mold from OU loc. IP01812; (10) partial pygidium OU 239057, latex mold, from OU loc. IP 01814; (11) partial pygidium OU 239058, latex mold, from OU loc. IP 01808; (12) partial pygidium OU 239059, latex mold, from OU loc. IP 01808; (13) pygidium OU 239060, latex mold, from OU loc. IP 01808; (14) partial pygidium OU 239061, latex mold, from OU loc. IP 01808. Scale bars = 4 mm (2–9, 12–14); 2 mm (1, 10, 11).

Figure 3. Long description
A collection of 14 fossil specimens of Neoblairella sp. indet. arranged in a grid.
Top row (1 to 5):
1. A small, grainy cranidium.
2. Two cranidia stacked vertically, the upper one more distinct.
3. A wide, mostly complete cranidium showing a central glabella.
4. A distorted cranidium with a rounded anterior.
5. A partial cranidium showing the right side and central glabella.
A 2 mm scale bar is located below panel 1.
Middle section (6 to 9):
6. A large, flattened cranidium with a prominent central glabella.
7. A highly detailed, large partial cranidium showing the texture of the glabella and fixed cheeks.
8. A partial, flattened cranidium with visible surface grain.
9. A curved librigena (free cheek) fragment.
Bottom section (10 to 14):
10. A small, partial pygidium (tail piece).
11. A wider, partial pygidium.
12. A partial pygidium showing segmented lobes.
13. A wide, nearly complete pygidium with a distinct central axis.
14. A partial pygidium showing clear horizontal segmentation.
A 4 mm scale bar is located above panel 13.
All specimens are preserved in sandstone, appearing as light gray textured shapes against a darker, grainy matrix.
Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025 Blairella sp., Muhlbauer et al., fig. 3b.
Occurrence
Bolaspidella to Cedaria biozones, Santa Catalina Sandstone, Abrigo Formation, Nugget Canyon, Arizona, USA (OU locs. IP 01805–01812, 01814).
Remarks
The cranidia assigned here to Neoblairella sp. indet. are similar in glabellar outline and overall cranidial shape to the new species recognized by Łabaj (Reference Łabaj2015). The cranidia differs from the type species, N. crassimarginata (Rasetti, Reference Rasetti1965a), in having a shorter, narrower, less convex anterior border; an exception is the specimen in Figure 3.5, which has a stronger, curved anterior border. The cranidia from Nugget Canyon differ from N. triangularis (Rasetti, Reference Rasetti1965a) in having a rounded posterior margin of the more-level occipital ring, and a narrower interocular area of the fixigena.
Rasetti (Reference Rasetti1965a) named the two known species based only on cranidia. Łabaj (Reference Łabaj2015) illustrated pygidia and librigena associated with the cranidia. The librigena at Nugget Canyon is similar to those illustrated by Łabaj (Reference Łabaj2015) in possessing a relatively short genal spine and wide genal area (Fig. 3.9). The pygidia reported here are also similar to those illustrated by Łabaj (Reference Łabaj2015) in their transverse width, relatively wide axis, pleural bands and furrows joining near the anterior border, and anterior lateral margins located near the axis midlength. It should be noted that these pygidia are similar to some species of Modocia Walcott, Reference Walcott1924 (see Rasetti, Reference Rasetti1965a, b) and Ehmaniella (see Sundberg, Reference Sundberg1994), suggesting a close phylogenetic relationship with Marjumiidae and Alokistocaridae.
Subfamily Altiocculinae Sundberg, Reference Sundberg1994
Genus Altiocculus Sundberg, Reference Sundberg1994
Type species
Altiocculus drumensis Sundberg, Reference Sundberg1994, Swasey Limestone, Utah, USA.
Altiocculus sp. indet.
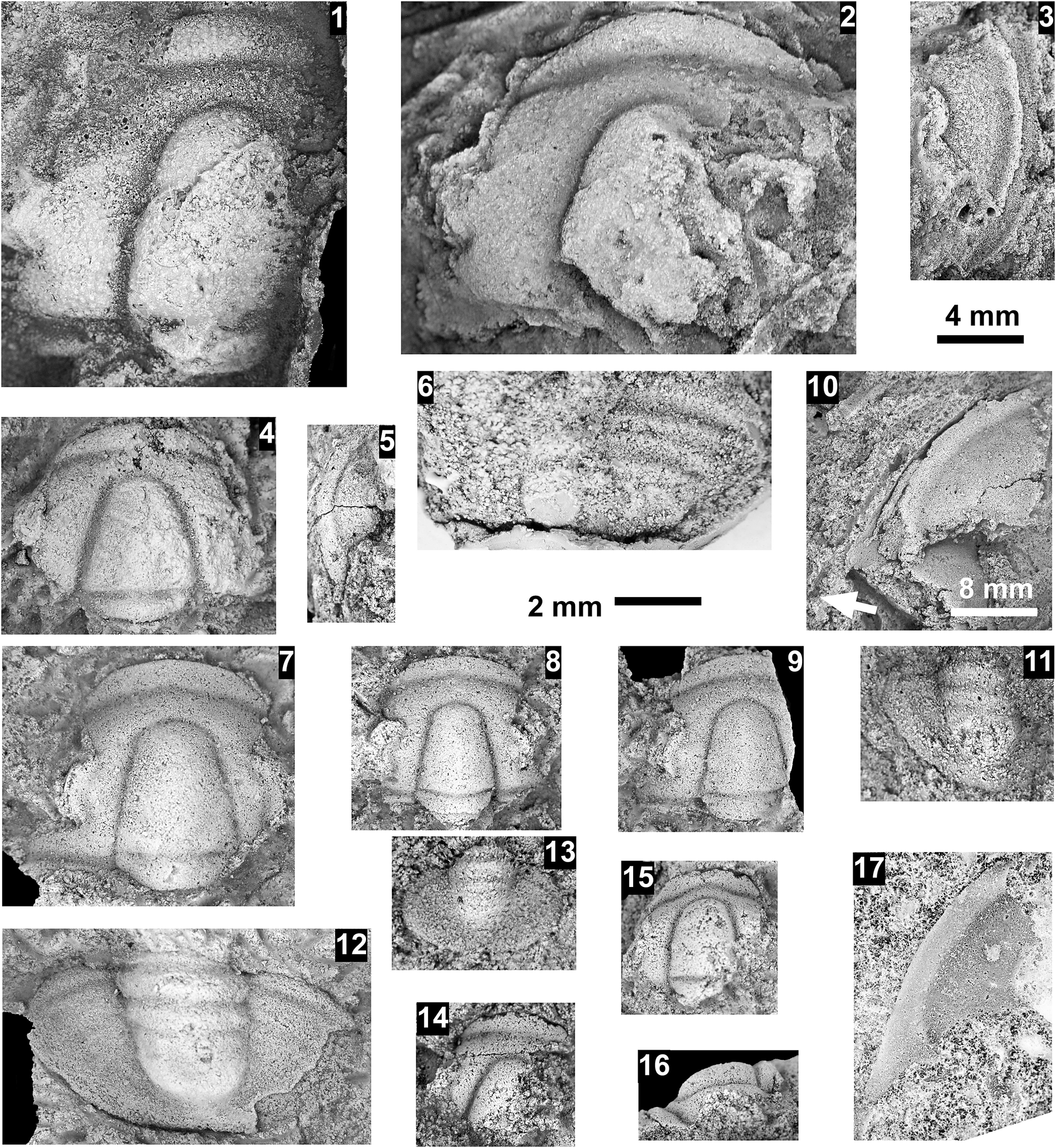
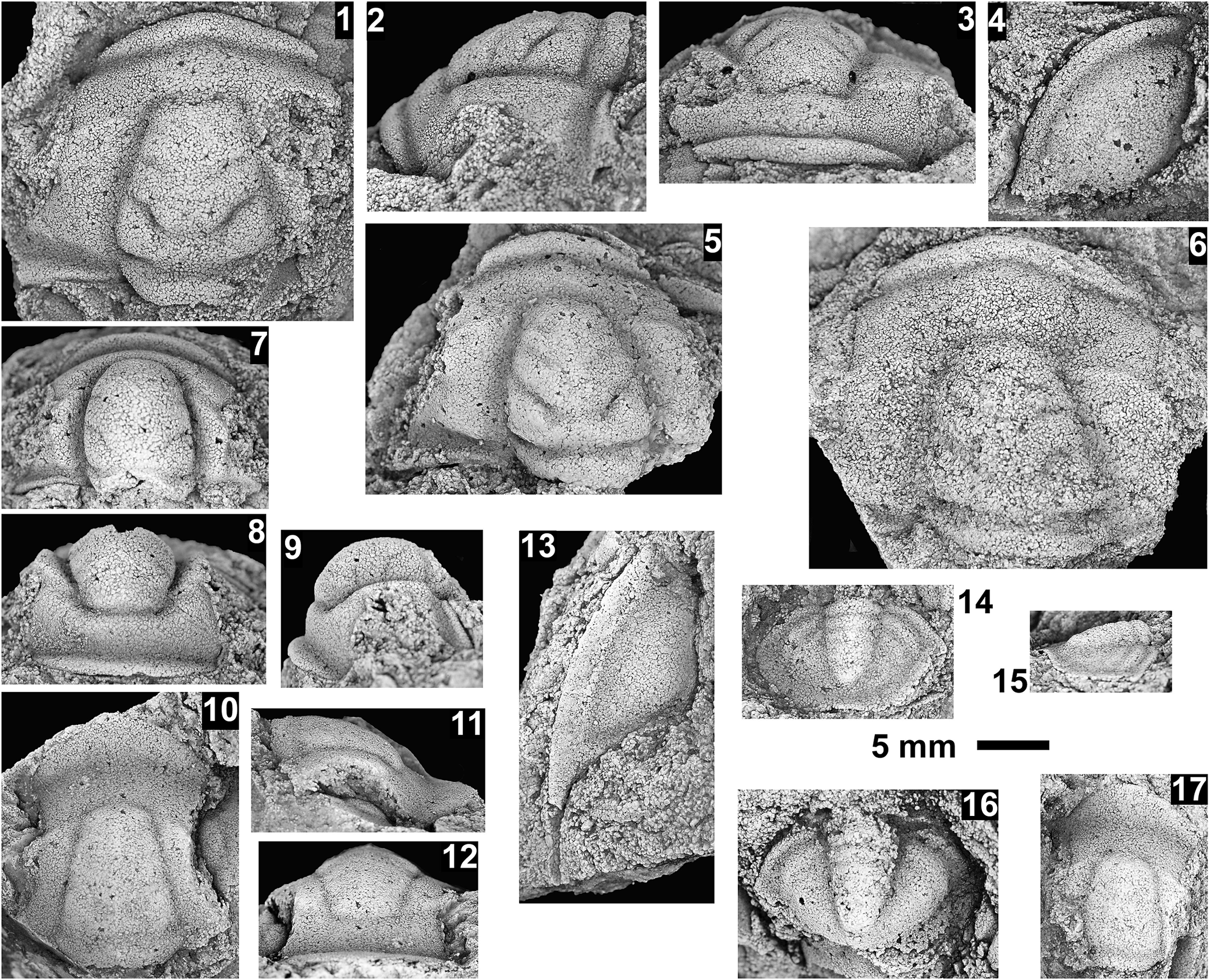
Altiocculus sp. indet., Santa Catalina Member, Abrigo Formation, Nugget Canyon, Arizona, USA: (1–3) nearly complete cranidium OU 239066, internal mold in dorsal, anterior, and lateral views, from OU loc. IP 01816; (4) complete librigena OU 239062, internal mold, from OU loc. IP 01805; (5) partial cranidium OU 239063, latex mold, tectonically deformed, from OU loc. IP 01805; (6) partial cranidium OU 239064, internal mold, from OU loc. IP 01805; (7) incomplete cranidium OU 239065, internal mold, from OU loc. IP 01805.

Figure 4. Long description
A collection of seven fossil photographs labeled 1 through 7. A scale bar in the upper center indicates 4 m m.
* Panel 1 at the top left shows a dorsal view of a nearly complete cranidium O U 239066, an internal mold with a granular texture and a distinct central glabella.
* Panel 2 at the top right shows the same specimen in an anterior view, highlighting the convex curvature of the head shield.
* Panel 3, below panel 2, shows the same specimen in a lateral view, revealing the profile of the glabella and fixed cheeks.
* Panel 4 on the far left is a vertical rectangular panel showing a complete librigena O U 239062, an internal mold with a triangular shape and a long genal spine.
* Panel 5 in the middle right shows a partial cranidium O U 239063, a latex mold that appears tectonically deformed with a stretched glabella.
* Panel 6 at the bottom left shows a partial cranidium O U 239064, an internal mold with a well-defined glabella and frontal area.
* Panel 7 at the bottom right shows an incomplete cranidium O U 239065, an internal mold with a prominent, rounded glabella and surrounding fixigenae.
Occurrence
Bolaspidella to Cedaria biozones, Santa Catalina Sandstone, Abrigo Formation, Nugget Canyon, Arizona, USA (OU locs. IP 01805, 01809, 01812, 01816).
Remarks
This species is known only from cranidia and one librigena from the Nugget Canyon section. The specimens are commonly distorted and preserved in sandstone that does not preserve surface textures; however, one specimen (Fig. 4.1–4.3) illustrates the wide fixigena, the elevated palpebral lobes, upturning to almost vertical palpebral lobes, upsloping fixigena, and the long frontal area characteristic of Altiocculus (see Sundberg, Reference Sundberg1994). The specimens are similar to Altiocculus sinuatum (Rasetti, Reference Rasetti1951) with its convex anterior border and more posteriorly placed palpebral lobes, but differs in the less-tapered glabella. Nomenclature is left open due to the poor preservation.
Family Marjumiidae Kobayashi, Reference Kobayashi1935
Genus Modocia Walcott, Reference Walcott1924
Type species
Arionellus (Crepicephalus Owen, Reference Owen1852) oweni Meek and Hayden, Reference Meek and Hayden1861, Deadwood Formation, Wyoming, USA (by original designation).
Modocia centralis (Whitfield, Reference Whitfield1877)
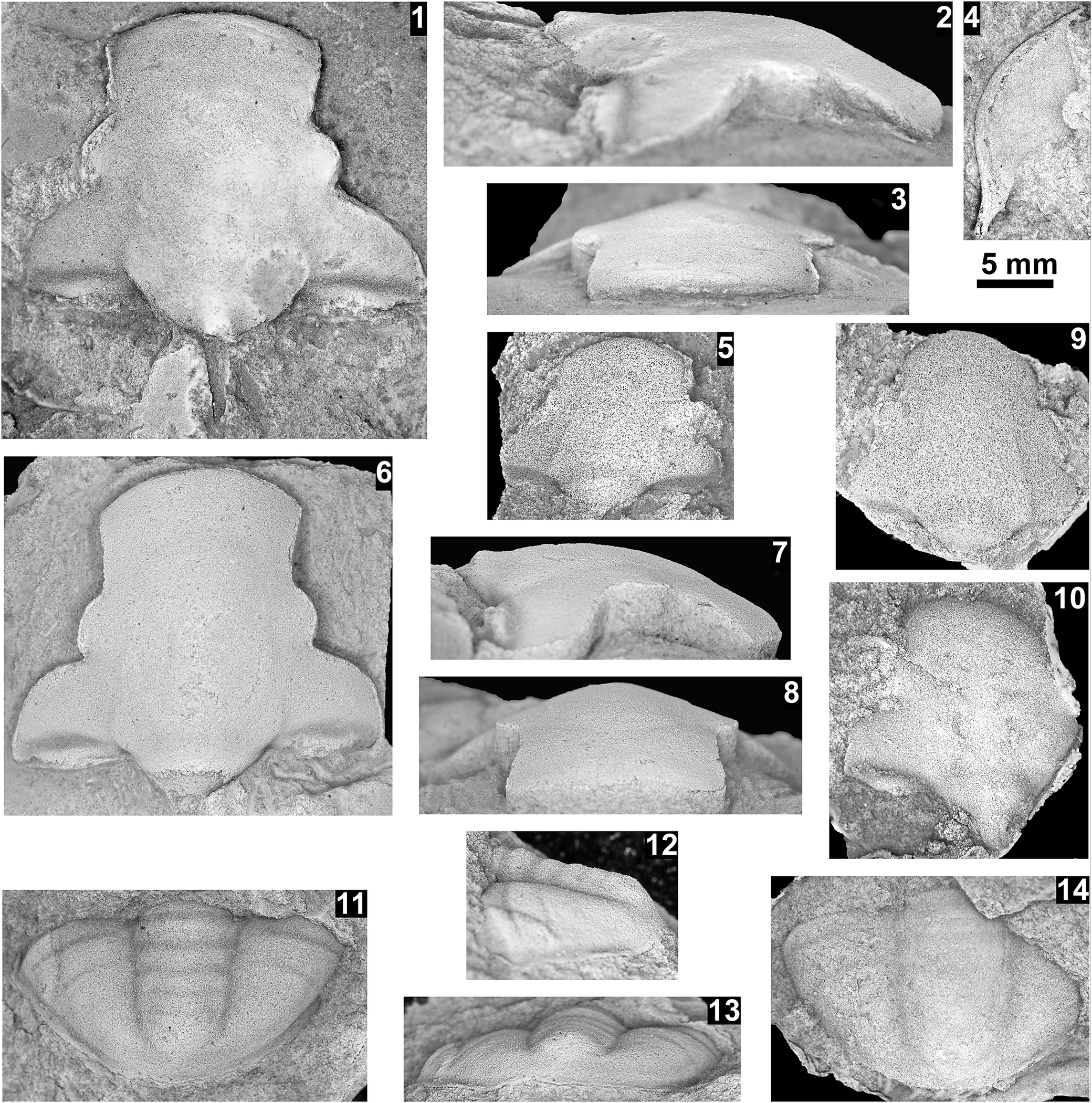
Modocia spp., Abrigo Formation, Nugget Canyon, Arizona, USA. (1–6) Modocia centralis (Whitfield, Reference Whitfield1877), in limonitic dolostone above the Southern Belle Quartzite, from OU loc. IP 01818 unless otherwise noted: (1) partial cranidium OU 239067, silicon mold; (2) partial cranidium OU 239068, internal mold; (3) librigena OU 239069, internal mold; (4) partial cranidium OU 239070, internal mold; (5) librigena OU 239071, internal mold; (6) partial pygidium OU 239072, latex mold, questionably assigned to the species, from OU loc. IP 01819. (7–13) Modocia oweni? (Meek and Hayden, Reference Meek and Hayden1861), in sandstones from the Santa Catalina Sandstone, from OU loc. IP 01816 unless otherwise noted: (7) cranidium OU 239073, internal mold; (8) cranidium OU 239074, internal mold; (9) partial cranidium OU 239075, internal mold; (10) partial librigena OU 239076 with impression of spine (arrow), internal mold; (11) partial pygidium OU 239077, latex mold; (12) distorted pygidium OU 239078, internal mold, from OU loc. IP 01813; (13) pygidium OU 239079, latex mold, from OU loc. IP 01815. (14–17) Modocia cf. M. crassimarginata Rasetti Reference Rasetti1965b, Santa Catalina Sandstone, Abrigo Limestone, from OU loc. IP 01812: (14) partial cranidium OU 239080, internal mold; (15, 16) partial cranidium OU 239082, internal mold, in dorsal and lateral views; (17) librigena OU 239081, exterior mold, inverted color and flipped horizontally. Scale bars = 8 mm (10); 4 mm (1–5, 7–9, 11–17); 2 mm (6).

Figure 5. Long description
The photo contains 17 numbered panels showing fossil molds and casts.
* Panels 1 through 6 show Modocia centralis. Panel 1 is a partial cranidium silicon mold. Panel 2 is a partial cranidium internal mold. Panel 3 is a librigena internal mold. Panel 4 is a partial cranidium internal mold. Panel 5 is a librigena internal mold. Panel 6 is a partial pygidium latex mold.
* Panels 7 through 13 show Modocia oweni. Panels 7 and 8 are full cranidia internal molds. Panel 9 is a partial cranidium internal mold. Panel 10 is a partial librigena with a white arrow pointing to a spine impression. Panel 11 is a partial pygidium latex mold. Panel 12 is a distorted pygidium internal mold. Panel 13 is a pygidium latex mold.
* Panels 14 through 17 show Modocia cf. M. crassimarginata. Panel 14 is a partial cranidium internal mold. Panels 15 and 16 show a partial cranidium internal mold in dorsal and lateral views respectively. Panel 17 is a librigena exterior mold.
Three scale bars are provided: an 8 mm bar for panel 10, a 4 mm bar for panels 1 through 5 and 7 through 9 and 11 through 17, and a 2 mm bar for panel 6.
Reference Whitfield1877 Crepicephalus (Loganellus) centralis Whitfield, p. 10.
Reference Shaw1956 Modocia centralis; Shaw, p. 141 (see for synonymy).
Reference Deland and Shaw1956 Modocia centralis; Deland and Shaw, p. 358, pl. 64, fig. 10.
Reference Lochman and Hu1961 Modocia centralis; Lochman and Hu, p. 136, pl. 29, figs. 1–39.
Reference Stitt1998 Modocia centralis; Stitt, p. 1038, fig. 6.15–6.20, 6.23.
Holotype
Cranidium USNM 24581, Deadwood Formation, South Dakota, USA (Whitfield, Reference Whitfield1877, p. 10; Walcott, Reference Walcott1925, pl. 16, figs. 1, 2; see Shaw, Reference Shaw1956).
Occurrence
Cedaria Biozone, upper Abrigo Formation, Nugget Canyon, Arizona, USA (OU locs. IP 01818–01820, 01822). Cedaria dakotaensis Biozone, Deadwood Formation, South Dakota, USA (Whitfield, Reference Whitfield1877; Walcott, Reference Walcott1925; Stitt, Reference Stitt1998). Cedaria Biozone, DuNoir Limestone, Wind River Mountains (Lochman and Hu, Reference Lochman and Hu1961) and Park Shale (Deland and Shaw, Reference Deland and Shaw1956), Wyoming, USA.
Remarks
The specimens from Nugget Canyon occur a few meters above the Southern Bell Quartzite in soft, friable limonite beds. Some specimens illustrate the granular surface of the cranidium and librigena, as well as the relatively wide fixigena typical of the specimens illustrated by Stitt (Reference Stitt1998, fig. 6.15–6.19, 6.23) and Łabaj (Reference Łabaj2015, pl. 9, figs. 5–11). The single fragmentary pygidium questionably assigned to this species appears to be more transverse than those illustrated by these two authors and might belong to a different taxon.
Modocia cf. M. crassimarginata Rasetti, Reference Rasetti1965b
cf. 1965b Modocia crassimarginata Rasetti, p. 109, pl. 2, figs. 1–9.
Occurrence
Cedaria Biozone, Santa Catalina Sandstone, Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01812).
Remarks
The species from Nugget Canyon is known from a few cranidia and librigena. These specimens are assigned based on the narrow preglabellar region and relatively high profile of the cranidium illustrated by Rasetti (Reference Rasetti1965a, pl. 2, figs. 1–5, 8) and Łabaj (Reference Łabaj2015, pl. 8, figs. 14–18).
Modocia oweni ? (Meek and Hayden, Reference Meek and Hayden1861)
Reference Meek and Hayden1861 Arionellus oweni Meek and Hayden, p. 436.
Reference Walcott1924 Modocia oweni; Walcott, p. 59, pl. 12, fig. 7.
Reference Walcott1925 Modocia oweni; Walcott, p. 106, pl. 16, figs. 1–3 (see for synonymy).
Reference Palmer1954b Modocia cf. M. oweni; Palmer, p. 763, pl. 87, figs. 3, 4, 6.
Reference Stitt1998 Modocia oweni; Stitt, p. 1038, fig. 6.21, 6.22, 6.24–6.26.
Reference Jell2021 Modocia oweni; Jell, fig. 1A, B.
Holotype
Cranidium USNM 1180 from Deadwood Formation, Wyoming, USA (Meek and Hayden, Reference Meek and Hayden1861, p. 436; Walcott, Reference Walcott1925, pl. 16, fig. 3).
Occurrence
Cedaria Biozone, Santa Catalina Sandstone, Abrigo Formation, Nugget Canyon, Arizona, USA (OU locs. IP 01813, 01815, 01816). Cedaria dakotaensis Biozone, Deadwood Formation, Wyoming, USA (Meek and Hayden, Reference Meek and Hayden1861; Walcott, Reference Walcott1925; Stitt, Reference Stitt1998).
Remarks
The species from Nugget Canyon is known from cranidia, librigena, and pygidia. The cranidia are similar to those illustrated by Stitt (Reference Stitt1998, fig. 6.24) and Łabaj (Reference Łabaj2015, pl. 9, figs. 3, 4) in possessing a narrower fixigena and evenly cured anterior border. However, the most examples of cranidia from Nugget Canyon have a more truncated glabella (e.g., Fig. 5.7, 5.8) than that illustrated by Stitt (Reference Stitt1998, fig. 6.24). The pygidia, however, differ from those illustrated by Stitt (Reference Stitt1998, fig. 6.21, 6.22) in being more oval, similar to the pygidia illustrated as Modocia cf. M. crassimarginata by Łabaj (Reference Łabaj2015, pl. 8, figs. 19–23), with the exception of the dorsal arch of the anterior margin. The difference in pygidial shape is the reason the species identification is questionable.
Family Asaphiscidae Raymond, Reference Raymond1924
Subfamily Kingstoniinae Kobayashi, Reference Kobayashi1933
Genus Brachyaspidion Miller, Reference Miller1936a
Type species
Brachyaspis rynchina Miller, Reference Miller1936b, Gros Ventre Formation, Wyoming, USA.
Brachyaspidion sp. indet.
Brachyaspidion sp. indet., Santa Catalina Member, Abrigo Formation, Nugget Canyon, Arizona, USA, from OU loc. IP 01803, in sandstone and internal molds: (1) poorly preserved cranidium OU 239083; (2) small cranidium OU 239084; (3) partial cranidium OU 239085; (4) partial cranidium OU 239086.

Occurrence
Ehmaniella? to Bolaspidella biozones, Santa Catalina Member, Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01803).
Remarks
The specimens from Nugget Canyon are poorly preserved as internal molds in a medium-grained sandstone. They have the characteristic anterior border, anteriorly set palpebral lobes, and subtriangular cranidial shape similar to specimens of Brachyaspidion sulcatum Robison, Reference Robison1964 (pl. 86, figs. 4–6) or Brachyaspidion rynchina (Miller, Reference Miller1936b). Łabaj (Reference Łabaj2015, p. 170) considered these two species as synonyms. The poor preservation is the reason for leaving them in open nomenclature.
Family Menomoniidae Walcott, Reference Walcott1916
Genus Bolaspidella Resser, Reference Resser1937
Type species
Ptychoparia housensis Walcott, Reference Walcott1886, Wheeler Formation, Utah, USA (by original designation).
Bolaspidella sp. indet.
Bolaspididae and Menomoniidae representatives from the Santa Catalina Sandstone, Abrigo Formation, Nugget Canyon, Arizona, USA, all preserved in sandstone. (1–5) Eldoradia linnarssoni (Walcott, Reference Walcott1884) from OU loc. IP 01817: (1) partial cranidium OU 239087, latex mold; (2) partial cranidium OU 239088, internal mold; (3) partial cranidium OU 239089, internal mold; (4) partial cranidium OU 239090, internal mold; (5) nearly complete cranidium OU 239091, internal mold. (6, 7) Eldoradia prospectensis (Walcott, Reference Walcott1884) from OU loc. IP 01809: (6) nearly complete cranidium OU 239092, latex mold; (7) nearly complete cranidium OU 239093, latex mold. (8) Bolaspidella sp. indet. from OU loc. IP 01816, nearly complete cranidium OU 239094. (9–14) Bolaspididae gen. indet. sp. indet. from OU loc. IP 01817: (9) partial cranidium OU 239095, latex mold; (10, 11) partial cranidium OU 239099, internal mold, dorsal and lateral views; (12) partial cranidium OU 239096, latex mold; (13) poorly preserved cranidium OU 239097, internal mold; (14) partial cranidium OU 239098, latex mold.

Figure 7. Long description
A collection of 14 fossil specimens labeled with numbers in the top-right corner of each panel. A 2 m m scale bar is located near the center-right.
* Panel 1: A large, partial cranidium of Eldoradia linnarssoni showing a textured latex mold surface.
* Panel 2: A smaller, centered internal mold of an E. linnarssoni cranidium with a distinct glabella.
* Panel 3: A partial E. linnarssoni cranidium with a grainy, weathered texture.
* Panel 4: A small, wide internal mold of E. linnarssoni.
* Panel 5: A nearly complete, wide cranidium of E. linnarssoni showing the full lateral extent.
* Panel 6: A nearly complete cranidium of Eldoradia prospectensis with a smooth, rounded glabella.
* Panel 7: A smaller specimen of E. prospectensis showing similar rounded features.
* Panel 8: A cranidium of Bolaspidella species indeterminate, showing three distinct lobes.
* Panel 9: A partial cranidium of Bolaspididae genus and species indeterminate with a high-relief glabella.
* Panel 10: A dorsal view of a Bolaspididae specimen showing a semi-circular anterior margin.
* Panel 11: A lateral view of the same specimen from panel 10, showing the vertical profile and curvature.
* Panel 12: A latex mold of a Bolaspididae cranidium with a very smooth, elongated central glabella.
* Panel 13: A poorly preserved, highly textured internal mold of a Bolaspididae cranidium.
* Panel 14: A wide, partial latex mold of a Bolaspididae cranidium with visible granular surface details.
Occurrence
Bolaspidella Biozone, Santa Catalina Member, Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01806).
Remarks
Bolaspidella sp. indet. is represented by a single internal mold of a cranidium in sandstone. The small specimen has a nearly straight anterior border furrow and upturned convex border, palpebral area upsloping and wide, and long palpebral lobes typical of the genus Bolaspidella (see Robison, Reference Robison1964). This specimen is similar to a new species recognized by Łabaj (Reference Łabaj2015, pl. 16, figs. 1–12) with a reduced preglabellar area, subrectangular glabella, nearly straight anterior border, and wide fixigenae.
Family Bolaspididae Howell in Moore, Reference Moore1959
Genus Eldoradia Resser, Reference Resser1935
Type species
Ptychoparia? linnarssoni Walcott, Reference Walcott1884 (by original designation).
Eldoradia prospectensis (Walcott, Reference Walcott1884)
Reference Walcott1884 Ptychoparia? prospectensis Walcott, p. 46, pl. 9, fig. 20.
Reference Walcott1916 Alokistocare prospectense; Walcott, p. 186, pl. 25, fig. 8.
Reference Resser1935 Eldoradia prospectensis ; Resser, p. 27.
Reference Palmer1954a Eldoradia prospectensis; Palmer, p. 77, pl. 16, fig. 8.
Holotype
Cranidium USNM 15441 from the Secret Canyon Shale?, Nevada, USA (Walcott, Reference Walcott1884, pl. 9, fig. 20; Palmer, Reference Palmer1954a, pl. 16, fig. 8).
Occurrence
Cedaria Biozone, Santa Catalina Member, Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01809). ‘Middle Cambrian,’ Secret Canyon Shale?, Nevada, USA (Walcott, Reference Walcott1884; Palmer, Reference Palmer1954a).
Remarks
The specimens of Eldoradia from Nugget Canyon display subquadrate cranidia, prominent glabella, with relatively small, forward tapering and truncate frontal lobe, shallow or absent glabellar furrows, long frontal area with large median node extending from the frontal lobe to anterior border, wide fixigena, and relatively small palpebral lobes position next to anterior third of the glabella (see Palmer, Reference Palmer1954a, p. 76). Eldoradia prospectensis is characterized by a weakly defined preglabellar boss and a better-defined anterior border (Palmer, Reference Palmer1954a; Łabaj, Reference Łabaj2015). Only a few specimens of this species are in the Nugget Canyon section, where they occur as internal molds in sandstones.
Eldoradia linnarssoni (Walcott, Reference Walcott1884)
Reference Walcott1884 Ptychoparia? linnarssoni Walcott, p. 47, pl. 9, fig. 18, 18a.
Reference Walcott1916 Alokistocare linnarssoni; Walcott, p. 185, pl. 25, fig. 7, 7a.
Reference Resser1935 Eldoradia linnarssoni; Resser, p. 26.
Reference Resser1935 Eldoradia lata Resser, p. 27.
Reference Palmer1954a Eldoradia linnarssoni; Palmer, p. 77, pl. 16, figs. 9, 10.
Holotype
Cranidium USNM 24611 from the Secret Canyon Shale, Nevada, USA (Walcott, Reference Walcott1884, pl. 9, fig. 18, 18a; Palmer, Reference Palmer1954a, pl. 16, fig. 10).
Occurrence
Cedaria Biozone, Santa Catalina Member, Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01817). ‘Middle Cambrian,’ Secret Canyon Shale, Nevada, USA (Walcott, Reference Walcott1884; Palmer, Reference Palmer1954a).
Remarks
Eldoradia linnarssoni is characterized by a well-defined preglabellar boss and poorly defined anterior border. The material from Nugget Canyon is generally preserved as internal molds in sandstone and the preglabellar boss is commonly poorly defined.
Bolaspididae gen. indet. sp. indet.
Occurrence
Bolaspidella Biozone, Santa Catalina Member, Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01817).
Remarks
Cranidia of Bolaspidid gen. indet. sp. indet. are distinctive in a large rounded, upturned anterior border and lack of a preglabellar area and medial swelling. The specimens are similar to Eldoradia (see Palmer, Reference Palmer1954a; Peel, Reference Peel2021) with its wider interocular region, glabellar shape and length, and laterally projected ocular ridges. However, the frontal area lacks any apparent swelling typical of Eldoradia and other representative of Bolaspididae. The cranidia are also similar to that of Bolaspidella burnetensis (Walcott, Reference Walcott1890) (see Palmer, Reference Palmer1954b, pl. 83, fig. 10), but the latter has a shorter anterior border, narrower fixigena, and a longer preglabellar area. Additional better and more complete material needs to be found before generic or familial assignments can be made.
Family Cedariidae Raymond, Reference Raymond1937
Genus Cedaria Walcott, Reference Walcott1924
Type species
Cedaria prolifica Walcott, Reference Walcott1924, Conasauga Formation, Alabama, USA (by original designation).
Cedaria sp. indet.
Trilobites from the upper Abrigo Formation, Nugget and Peppersauce canyons, Arizona, USA, all preserved in sandstone. (1, 2) Cedaria sp. indet. from OU loc. IP 01821 float: (1) partial cranidium OU 239100, external mold, latex mold; (2) partial cranidium OU 239101, external mold, latex mold. (3, 4) Crepicephalus sp. indet., OU 239104, partial and poorly preserved pygidium, from OU loc. IP 01825: (3) exterior mold (latex mold); (4) internal mold. (5–11) Tricrepicephalus texanus (Shumard, Reference Shumard1861): (5–7) Stoyanow specimen, cranidium LACMIP 15026, in dorsal, anterior, and lateral views, from LACMIP loc. 29165; (8) librigena OU 239102, latex mold, from OU loc. IP 01821; (9) Stoyanow specimen, cranidium LACMIP 15027, from LACMIP loc. 29165; (10) Stoyanow specimen, pygidium LACMIP 15028, from LACMIP loc. 29165; (11) Stoyanow specimen, pygidium with flattened spines LACMIP 15029, from LACMIP loc. 29165. Scale bars = 5 mm (5–11); 2 mm (1, 2); 1 mm (3, 4).

Figure 8. Long description
A collection of eleven fossil specimens numbered 1 through 11.
* Panels 1 and 2 at the top left show partial cranidia of Cedaria species indet. preserved as latex molds of external molds. A 2 m m scale bar is positioned between panels 2 and 5.
* Panels 3 and 4 at the top right show a partial, poorly preserved pygidium of Crepicephalus species indet. Panel 3 is a latex mold of an exterior mold and panel 4 is an internal mold. A 1 m m scale bar is located below panel 3.
* Panels 5 through 11 show Tricrepicephalus texanus specimens. A 5 m m scale bar is located above panel 8.
* Panels 5, 6, and 7 show a single cranidium (L A C M I P 15026) in dorsal, anterior, and lateral views respectively. The dorsal view in panel 5 shows a rounded glabella and wide fixed cheeks.
* Panel 8 shows a long, curved librigena preserved as a latex mold.
* Panel 9 shows a dorsal view of a second cranidium (L A C M I P 15027).
* Panel 10 shows a pygidium (L A C M I P 15028) with a distinct segmented axis.
* Panel 11 shows a pygidium (L A C M I P 15029) featuring two long, flattened spines extending posteriorly from the lateral margins.
Occurrence
Cedaria Biozone, upper Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01821).
Remarks
Two cranidia have the flat anterior border, tapered glabella, relatively long palpebral lobes, and narrow interocular area similar to those of Cedaria eurycheilos Palmer, Reference Palmer1954b as illustrated by Łabaj (Reference Łabaj2015). However, the specimens are distorted, have a narrower preglabellar area, and lack associated pygidia to confirm species identification.
Family Tricrepicephalidae Palmer, Reference Palmer1954b
Genus Tricrepicephalus Kobayashi, Reference Kobayashi1935
Type species
Arionellus (Bathyurus Billings, Reference Billings1859) texanus Shumard, Reference Shumard1861, Riley Formation, Texas, USA (by original designation).
Tricrepicephalus texanus (Shumard, Reference Shumard1861)
Reference Shumard1861 Arionellus (Bathyurus) texanus Shumard, p. 218.
Reference Palmer1954b Tricrepicephalus texanus: Palmer, p. 755, pl. 81, fig. 9 [see for synonymy].
not Reference Pratt1992 Tricrepicephalus texanus: Pratt, p. 62, pl. 21, figs. 1–7 [= T. coria (Walcott, Reference Walcott1916)].
not Reference Sundberg and Cuen-Romero2021 Tricrepicephalus texanus: Sundberg and Cuen-Romero, p. 7, fig. 5a–g [= T. coria].
Reference Tortello2023 Tricrepicephalus texanus: Tortello, p. 8, fig. 4.9–4.13 [not fig. 4.1–4.6 = T. coria; fig. 4.14–4.20 = specific placement unsure, see Remarks].
not Reference Cuen-Romero, Ochoa-Granillo, Noriega-Ruiz, Tortello, Monreal and Ozua-Ibarra2024 Tricrepicephalus texanus: Cuen-Romero et al., p. 3, fig. 3A–E [= T. coria].
Neotype
Types from the Riley Formation, Texas were destroyed and a neotype has yet to be designated (see Pratt, Reference Pratt1992, p. 62).
Occurrence
Cedaria to Crepicephalus? biozones, upper Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01821, 01824; LACMIP loc. 29165). Lower Coosella Biozone (= upper Cedaria Biozone), Riley Formation, Texas, USA (Palmer, Reference Palmer1954b). Pilgrim Limestone, Montana, USA (Resser, Reference Resser1937; Lochman, Reference Lochman1940; Lochman and Duncan, Reference Lochman and Duncan1944). ‘Upper Cambrian’ Nolinchucky Formation, Tennessee, Alabama, USA (Resser, Reference Resser1938). Cedaria Biozone, Bonneterre Dolostone, Missouri, USA (Lochman, Reference Lochman1940). Cedaria Biozone, Warrior Formation, Pennsylvania, USA (Tasch, Reference Tasch1951). Crepicephalus Biozone Western Newfoundland, Canada, (Kindle, Reference Kindle1948, Reference Kindle1982). Cedaria prolifica Biozone, La Cruz Olistoliths, San Isidro area, western Argentina (Pratt, Reference Pratt1992; Tortello, Reference Tortello2023).
Remarks
There are several similarities between Tricrepicephalus coria (Walcott, Reference Walcott1916) and T. texanus. Palmer (Reference Palmer1954b) separated the two species based on the latter having unequal granulation and absence on the glabella except in the anterior portion. Palmer (Reference Palmer1954b) also considered the two species as possibly representing a ‘species group.’ Palmer (Reference Palmer1954b, p. 755) also mentioned that isolated pygidia cannot be assigned to an induvial species. Pratt (Reference Pratt1992), Sundberg and Cuen-Romero (Reference Sundberg and Cuen-Romero2021), Tortello (Reference Tortello2023), and Cuen-Romero et al. (Reference Cuen-Romero, Ochoa-Granillo, Noriega-Ruiz, Tortello, Monreal and Ozua-Ibarra2024) regarded them as synonyms; a more detailed study is needed to justify grouping the two taxa into a single species. The specimens from Nugget and Peppersauce canyons lack the granulation on the glabella (Fig. 8.5–8.7, 8.9) and are thus assigned to T. texanus.
One large librigena and one poorly preserved cranidium occur at loc. IP 01821 and IP 01824, respectively. Stoyanow specimens from Peppersauce Canyon (Fig. 8.5–8.7, 8.9–8.11) are a few meters above the Southern Bell Quartzite. The librigena and cranidia are exfoliated but display the coarse and relatively dense granules on the preglabellar area and the fixigena, however, the glabella appears to lack the granules, typical of Tricrepicephalus texanus. The pygidium of Stoyanow’s specimen (Fig. 8.11) has broader, flattened terminal ends to the pygidial spine, which is similar to those of T. rex Lochman, Reference Lochman1940 that was assigned to T. texanus by Palmer (Reference Palmer1954b). A poorly preserved cranidium from the Crepicephalus Biozone illustrates the three pits in the anterior border furrow typical of the genus and is questionably assigned to the species.
Family Crepicephalidae Kobayashi, Reference Kobayashi1935
Genus Crepicephalus Owen, Reference Owen1852
Type species
Dikelocephalus? iowensis Owen, Reference Owen1852, Eau Claire Formation, Minnesota, USA (designated by Walcott, Reference Walcott1886, p. 206).
Crepicephalus sp. indet.
Occurrence
Crepicephalus Biozone, upper Abrigo Formation, Nugget Canyon, Arizona, USA (OU loc. IP 01825).
Remarks
A single, poorly preserved, small pygidium similar to Crepicephalus micrans Resser, Reference Resser1938 occurs in the upper Abrigo Formation. The specimen appears to have two pygidial spines and a curved posterior margin.
Family Elviniidae Kobayashi, Reference Kobayashi1935
Subfamily Elviniinae Kobayashi, Reference Kobayashi1935
Genus Elburgia Palmer, Reference Palmer1960
Type species
Crepicephalus (Loganellus) granulosus Hall and Whitfield, Reference Hall and Whitfield1877 (by original designation).
Elburgia sp. indet.
Trilobites from the upper Dunderbergia Biozone from the Peppersauce Sandstone, upper Abrigo Formation, Peppersauce Canyon, Arizona, USA; all originally collected by Stoyanow, preserved in sandstone and exfoliated. (1–6) Elburgia sp. indet.: (1–3) cranidium LACMIP 15030, in dorsal, lateral, and anterior views, from LACMIP loc. 29164; (4) librigena LACMIP 15031, from LACMIP loc. 29166; (5) cranidium LACMIP 15032, from LACMIP loc. 29164; (6) cranidium LACMIP 15033, from LACMIP loc. 29164. (7–9) Iddingsia sp. indet., cranidium LACMIP 15034, dorsal, anterior, and lateral views, from LACMIP loc. 29164. (10–17) Strigambitus cf. S. transversus Palmer, Reference Palmer1965: (10–12) cranidium LACMIP 15035, in dorsal, lateral, and anterior views, from LACMIP loc. 29166; (13) librigena LACMIP 15036, from LACMIP loc. 29166; (14, 15) pygidium LACMIP 15037, in dorsal and lateral views, from LACMIP loc. 29166; (16) pygidium LACMIP 15038, from LACMIP loc. 29166; (17) cranidium LACMIP 15039, from LACMIP loc. 29166.

Figure 9. Long description
The plate contains 17 black and white photographs of trilobite fossils with a 5 millimeter scale bar located near the bottom right.
* Panels 1 through 6 show Elburgia sp. indet. 1, 2, and 3 are dorsal, lateral, and anterior views of a cranidium. 4 is a librigena. 5 and 6 are additional cranidia in dorsal view.
* Panels 7 through 9 show Iddingsia sp. indet. cranidium LACMIP 15034 in dorsal, anterior, and lateral views respectively.
* Panels 10 through 17 show Strigambitus cf. S. transversus. 10, 11, and 12 are dorsal, lateral, and anterior views of a cranidium. 13 is a librigena. 14 and 15 are dorsal and lateral views of a pygidium. 16 is a second pygidium in dorsal view. 17 is a final cranidium in dorsal view.
The fossils exhibit a granular, exfoliated texture and are embedded in a coarse sandstone matrix. The cranidia typically feature a prominent central glabella and distinct ocular ridges.
Occurrence
Dunderbergia Biozone, Peppersauce Sandstone, Abrigo Formation, Peppersauce Canyon, Arizona, USA (LACMIP locs. 29164, 29166).
Remarks
The specimens here reported were originally collected by Stoyanow from the adjacent Peppersauce Canyon. Three cranidia and two librigena are present in the collection. All specimens are exfoliated and internal molds are composed of medium-grained sandstone. The specimens have features typical of Elburgia illustrated by Palmer (Reference Palmer1960, Reference Palmer1965) in the glabellar shape and furrows, placement of the palpebral lobes, width of librigena, frontal area length, and curvature of the anterior border. Species within the genus are recognized on surface ornamentation, which is lacking in these specimens; thus, the specimens are left in open nomenclature.
Subfamily Dokimocephalinae Kobayashi, Reference Kobayashi1935
Genus Iddingsia Walcott, Reference Walcott1924
Type species
Ptychoparia similis Walcott, Reference Walcott1884, Dunderberg Formation, Nevada, USA (by original designation).
Iddingsia sp. indet.
Occurrence
Dunderbergia Biozone, Peppersauce Sandstone, Abrigo Formation, Peppersauce Canyon, Arizona, USA (LACMIP loc. 29164).
Remarks
The specimen here was originally collected by Stoyanow from the adjacent Peppersauce Canyon. A single cranidium is present in the collection and is exfoliated. The specimen is most similar to the specimen of Iddingsia cf. I. intermedia Palmer, Reference Palmer1965 illustrated by Sundberg et al. (Reference Sundberg, Cothren and Dehler2024, fig. 13) in its high glabellar and cranidium convexity with a strongly downsloping frontal area and level anterior border. The lack of an exoskeleton and the moderately coarse sandstone matrix in the internal mold prevents the specimen’s assignment to a specific species, thus it is left in open nomenclature.
Family Pterocephaliidae Kobayashi Reference Kobayashi1935
Subfamily Pterocephaliinae Kobayashi, Reference Kobayashi1935
Genus Strigambitus Palmer, Reference Palmer1965
Type species
Strigambitus transversus Palmer, Reference Palmer1965, Hicks Formation, Utah, USA (by original designation).
Strigambitus cf. S. transversus Palmer, Reference Palmer1965
cf. Reference Palmer1965 Strigambitus transversus Palmer, p. 77, pl. 16, figs. 6–10.
Occurrence
Dunderbergia Biozone, Peppersauce Sandstone, Abrigo Formation, Peppersauce Canyon, Arizona, USA (LACMIP loc. 29164).
Remarks
The specimens here reported were originally collected by Stoyanow from the adjacent Peppersauce Canyon. Two cranidia, one librigena, and two pygidia are present in the collection and are exfoliated. The specimens are most similar to the specimens of Strigambitus transversus illustrated by Palmer (Reference Palmer1965, pl. 16, figs. 6–8) in having cranidia with tapered glabella, long frontal area, narrow fixigena, arched anterior border, librigena with unfurrowed lateral border, and pygidia with a more rounded outline, two well defined pleural bands, and five axial rings. The specimens differ from Strigambitus transversus in having a cranidium that has a more arched anterior border with a shallower furrow and shallower glabellar axial and lateral furrows, pygidia with shallower pleural furrows, and smaller transverse width (e.g., Palmer, Reference Palmer1965, pl. 16, fig. 9). The absence of the exoskeletons in the Stoyanow specimens prevent confirmation of the specimens to Strigambitus transversus or as a new species; thus, the specimens are left in open nomenclature.
Family Uncertain
Genus Arapahoia Miller, Reference Miller1936b
Type species
Arapahoia typa Miller Reference Miller1936b, Depass Formation, Wyoming, USA (by original designation).
Arapahoia ransomei (Stoyanow, Reference Stoyanow1936)
Arapahoia ransomei (Stoyanow, Reference Stoyanow1936) from the upper Abrigo Formation, Nugget and Peppersauce canyons, Arizona, USA; all preserved in limonitic sandstone: (1–3) Stoyanow’s lectotype of Hesperaspis ransomei, exfoliated cranidium LACMIP 7804, in dorsal, lateral, and anterior views, from LACMIP loc. 16249; (4) librigena OU 239109, exterior mold, latex mold, from OU loc. IP 01822; (5) exfoliated cranidium OU 239105, from OU loc. IP 01821 float; (6–8) Stoyanow’s holotype of Hesperaspis butleri, exfoliated cranidium LACMIP 8095, in dorsal, lateral, and anterior views, from LACMIP loc. 16249; (9) exfoliated cranidium OU 239106, from OU loc. IP 01821 float; (10) cranidium with partial occipital spine OU 239107, external mold, latex mold, from OU loc. IP 01821; (11–13) Stoyanow’s lectoparatype of Hesperaspis ransomei, exfoliated pygidium LACMIP 8096, in dorsal, lateral, and posterior views, from LACMIP loc. 16249; (14) partial pygidium OU 239108, external mold, latex mold, from OU loc. IP 01823.

Figure 10. Long description
The plate contains 14 numbered panels showing various fossil fragments.
Panels 1 through 3 show the lectotype cranidium L A C M I P 7804 in dorsal, lateral, and anterior views, featuring a smooth, broad glabella and wide fixed cheeks.
Panel 4 shows a librigena O U 239109 as a latex mold, displaying a curved outer margin.
Panel 5 shows an exfoliated cranidium O U 239105 in dorsal view.
Panels 6 through 8 show the holotype cranidium L A C M I P 8095 in dorsal, lateral, and anterior views, similar in structure to the lectotype but with slightly different preservation.
Panel 9 shows an exfoliated cranidium O U 239106.
Panel 10 shows a cranidium O U 239107 with a partial occipital spine extending from the posterior margin.
Panels 11 through 13 show the lectoparatype pygidium L A C M I P 8096 in dorsal, lateral, and posterior views, revealing a semi-circular shape with a segmented axis.
Panel 14 shows a partial pygidium O U 239108.
A 5 mm scale bar is located in the top right quadrant near panel 4.
Reference Stoyanow1936 Hesperaspis ransomei Stoyanow, p. 468, pl. 1, figs. 6, 7.
Reference Stoyanow1936 Hesperaspis butleri Stoyanow, p. 469, pl. 1, fig. 8.
Reference Resser1937 Arapahoia ransomei; Resser, p. 2.
Reference Resser1937 Arapahoia butleri; Resser, p. 2.
Type specimens
Lectotype, cranidium LACMIP 7804 (Fig. 10.1–10.3), LACMIP loc. 16249, Upper Abrigo Formation, Peppersauce Canyon, Arizona, USA. Paralectotype, pygidium LACMIP 7804 (Fig. 10.11–10.13), same location as lectotype.
Occurrence
Cedaria Biozone, upper Abrigo Formation, Nugget and Peppersauce canyons, Arizona, USA (OU loc. IP 01821–01823; LACMIP loc. 16249).
Remarks
Several specimens are preserved in fine-grained sandstone and express the shallow furrows on the cranidium and librigena typical of the species. In some instances, cranidia have a broad and relatively long occipital ring spine (Fig. 10.10). Also illustrated here are Stoyanow’s (Reference Stoyanow1936) type specimens of Hesperaspis ransomei (Figs. 10.1–10.3, 10.11–10.13) and H. butleri (Fig. 10.6–10.8). The two species were separated primarily based on the occipital spine, absent in H. butleri and present in H. ransomei. However, the holotype of H. butleri has a broken posterior end of the occipital ring as do other specimens collected by Stoyanow and in this project. The variability in the occipital ring preservation suggests that the two co-occurring species should be synonymized. Arapahoia ransomei is used here due to the better-preserved holotype cranidium with an associated pygidium.
Biostratigraphy
The biostratigraphy of the Abrigo Formation ranges from the Ehmaniella to Ellipsocephaloides biozones (Palmer in Gilluly, Reference Gilluly1956; Taylor in Hayes, Reference Hayes1975; Łabaj, Reference Łabaj2015; Łabaj and Pratt, Reference Łabaj and Pratt2016; Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025; see above). In Peppersauce and Nugget canyons, only five biozones are recognized, including the Ehmaniella, Bolaspidella, Cedaria, Crepicephalus, and Dunderbergia biozones (Fig. 11). There is no record of the Aphelaspis, Dicanthopyge, and Prehousia biozones that were expected to occur between the Crepicephalus and Dunderbergia biozones in the study area. In addition, the carbon isotope values from the upper Abrigo to Peppersauce Sandstone members (Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025) are not comparable to δ13C values in strata spanning the uppermost Crepicephalus to Elvinia biozones at the global reference section at Smithfield Canyon, Utah, USA (Cothren et al., Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022; Sundberg et al., Reference Sundberg, Cothren and Dehler2024).
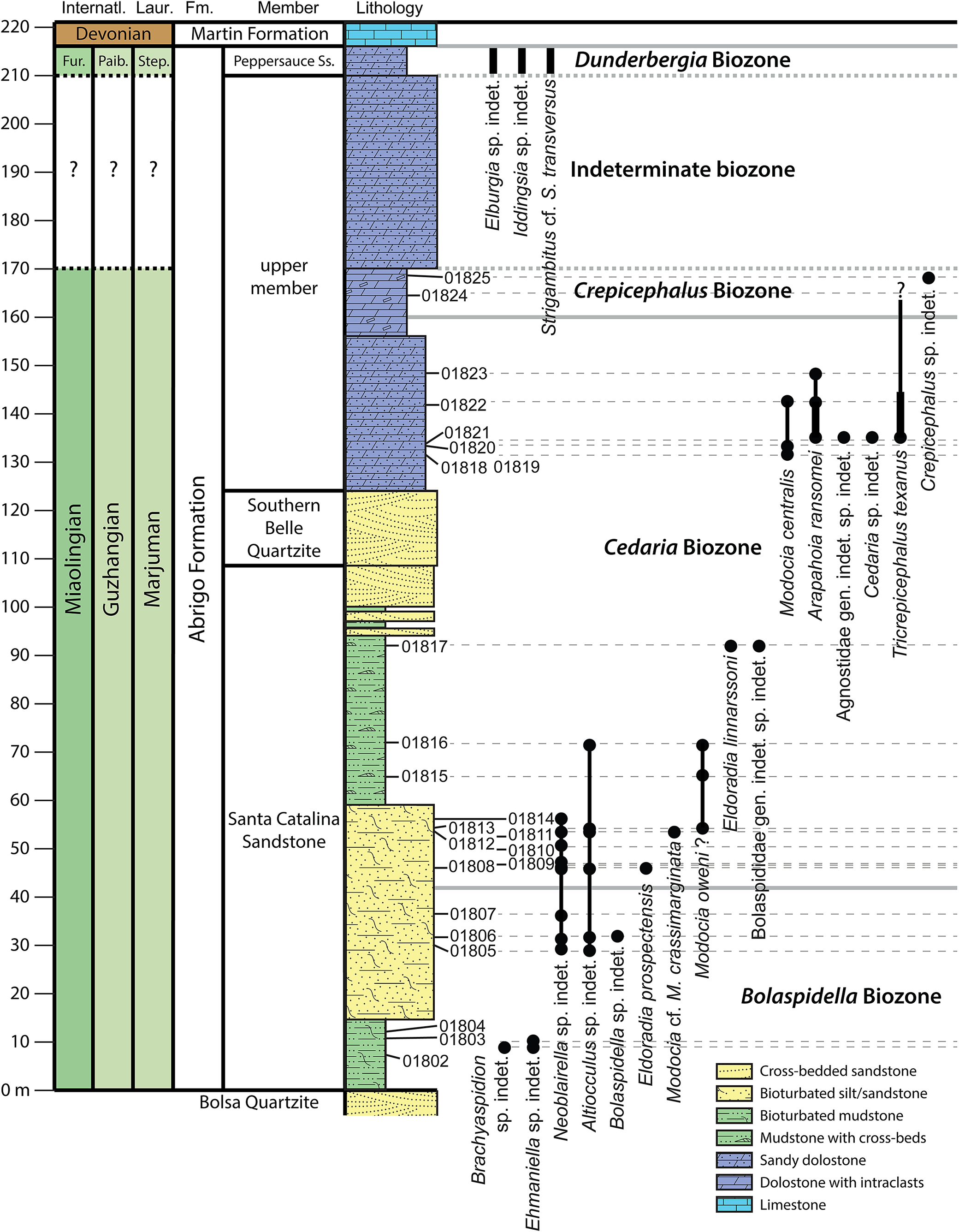
Biostratigraphic range chart of the Abrigo Formation, Nugget Canyon section, Santa Catalina Mountains, Arizona, USA. Stratigraphic thickness of the Abrigo Formation is 216 m. Sample numbers marked alongside lithostratigraphic column. Inferred biozone boundaries in bold gray lines. Black range bars are the approximate position of Stoyanow’s specimens from Peppersauce Canyon. Fm., Formation; Fur., Furongian; Internatl, International; Laur., Laurentian; Paib., Paibian; Step., Steptoean.

Figure 11. Long description
The chart is organized into vertical columns.
1. Chronostratigraphy: The leftmost columns list International and Laurentian stages. From bottom to top, these include Miaolingian (Guzhangian) and Marjuman, followed by Furongian (Paibian and Steptoean) and Devonian.
2. Lithostratigraphy: The Abrigo Formation is divided into the Santa Catalina Sandstone (0 to 108 meters), Southern Belle Quartzite (108 to 124 meters), and an upper member (124 to 210 meters), capped by the Peppersauce Sandstone and Martin Formation.
3. Lithology and Samples: A central column uses color-coded patterns.
- Green: Bioturbated mudstone and mudstone with cross-beds (0 to 95 meters).
- Yellow: Bioturbated silt/sandstone and cross-bedded sandstone (found in Santa Catalina and Southern Belle units).
- Blue: Sandy dolostone and dolostone with intraclasts (124 to 210 meters).
- Light Blue: Limestone at the top.
Sample numbers (01802 to 01825) are marked at specific meter intervals.
4. Fossil Ranges and Biozones: Vertical black bars with circular endpoints indicate fossil occurrences.
- Bolaspidella Biozone (0 to 92 meters): Includes Brachyaspidion sp. indet., Ehmaniella sp. indet., Neoblairella sp. indet., Altiocculus sp. indet., Bolaspidella sp. indet., Eldoradia prospectensis, Modocia cf. M. crassimarginata, and Modocia oweni.
- Cedaria Biozone (92 to 160 meters): Includes Eldoradia linnarssoni, Bolaspididae gen. indet. sp. indet., Modocia centralis, Arapahoia ransomei, Agnostidae gen. indet. sp. indet., and Cedaria sp. indet.
- Crepicephalus Biozone (160 to 170 meters): Includes Tricrepicephalus texanus and Crepicephalus sp. indet.
- Indeterminate biozone (170 to 210 meters): Includes Elburgia sp. indet., Iddingsia sp. indet., and Strigambitus cf. S. transversus.
- Dunderbergia Biozone: Located at the top of the section above 210 meters.
However, the Aphelaspis Biozone occurs at sections to the south at Ajax Hill, Johnny Lyon Hills, French Joe Canyon, Dragoon Mountains, Little Dragoon Mountains, and Picacho de Calera (Palmer in Gilluly, Reference Gilluly1956; Taylor in Hayes, Reference Hayes1975; Łabaj, Reference Łabaj2015; Łabaj and Pratt, Reference Łabaj and Pratt2016; Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025). At French Joe Canyon, the Aphelaspis Biozone is overlain by Stage 10 strata across a karsted surface, and the Dunderbergia, Elvinia, and Taenicephalus biozones are absent (Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025).
The Nugget Canyon section also does not contain trilobite biozones above the Dunderbergia Biozone, with the Devonian Martin Formation sitting directly and unconformably on the Peppersauce Sandstone. The Elvina Biozone, which directly overlies the Crepicephalus or Aphelaspis biozones, is known from the Swisshelm Mountains, Rattlesnake Ridge, and Ajax Hill (Taylor in Hayes, Reference Hayes1975; Łabaj, Reference Łabaj2015; Łabaj and Pratt, Reference Łabaj and Pratt2016), suggesting that an unconformity exists between Sauk II and Sauk III at these locations (Łabaj, Reference Łabaj2015; Łabaj and Pratt, Reference Łabaj and Pratt2016).
Ehmaniella and Bolaspidella biozones
The lower part of the Santa Catalina Sandstone member of the Abrigo Limestone contains taxa that could represent the Ehmaniella Biozone: Ehmaniella sp. indet. and Altiocculus sp. indet. Ehmaniella is typical of the Ehmaniella subzone of the Ehmaniella Biozone (Sundberg, Reference Sundberg1994). However, Ehmaniella ranges into the Altiocculus subzone, Ehmaniella Biozone in Utah (Sundberg, Reference Sundberg1994) and ranges into the Bolaspidella Biozone in other regions (Robison and Babcock, Reference Robison and Babcock2011). A key point is that the Altiocculus subzone in Utah and Nevada marks a major change in biofacies and lithofacies and is overlain by the deeper-water deposits of the Wheeler Shale and lithologic equivalents (Sundberg, Reference Sundberg, Cooper and Stevens1991). The Ptychagnostus gibbus Biozone overlies the Altiocculus subzone and is considered to be partly equivalent. This biozone assemblage is dominated by agnostids, generally lacks ptychopariids, and represents a significant biofacies change. The genus Altiocculus ranges from the upper Ehmaniella Biozone (Sundberg, Reference Sundberg1994) to the Bolaspidella Biozone—i.e., Altiocculus harrisi (Robison, Reference Robison1971)—in the Drum Mountains and House Range, Utah. Altiocculus americanum (Walcott, Reference Walcott1916) was reported from the Abrigo Formation by Łabaj (Reference Łabaj2015; as Alokistocare Lorenz, Reference Lorenz1906; Sundberg, Reference Sundberg1999 and Esteve et al., Reference Esteve, Sundberg, Zamora and Gozalo2012 assigned this species to Altiocculus) from the upper Bolaspidella Biozone and by Schwimmer (Reference Schwimmer1989) from the Bolaspidella Biozone in the Conasauga Formation, Georgia. Altiocculus sp. indet. occurs in Nugget Canyon in the upper Bolaspidella to lower Cedaria biozones.
Taxa from the Bolaspidella Biozone include: Brachyaspidion sp. indet., Bolaspidella sp. indet., and Neoblairella sp. indet. Robison (Reference Robison1964) and Łabaj (Reference Łabaj2015) recognized Brachyaspidion from the lower portion of the biozone (Bathyuriscus fimbriatus Subzone of Robison, Reference Robison1964). Bolaspidella sp. indet. is similar to a species recognized by Łabaj (Reference Łabaj2015) in the upper part of the biozone; however, the genus ranges through the entire Bolaspidella Biozone (Robison, Reference Robison1964). Neoblairella was also recognized by Łabaj (Reference Łabaj2015) in the upper part of the biozone.
Cedaria Biozone
The upper portion of the Santa Catalina Sandstone member and the lower dolostone facies above the Southern Belle Quartzite contain taxa belonging to the Cedaria Biozone, including: Cedaria sp. indet., Eldoradia linnarssoni, Eldoradia prospectensis, Modocia centralis, Modocia cf. M. crassimarginata, Arapahoia ransomei, and Tricrepicephalus texanus. The two Eldoradia species and Modocia cf. M. crassimarginata were recognized by Łabaj (Reference Łabaj2015) in the middle part of the Cedaria Biozone. Modocia centralis and Arapahoia ransomei were recognized by Łabaj (Reference Łabaj2015) as occurring in the upper Cedaria Biozone.
In Nugget Canyon, a few specimens are identified as Modocia oweni? in what has been assigned as the lower Cedaria Biozone. Stitt (Reference Stitt1998) reported this species from the Cedaria dakotaensis Biozone in the lower part of the Deadwood Formation of South Dakota; however, Łabaj (Reference Łabaj2015) reported it from the Bolaspidella Biozone of the Abrigo Limestone. The new occurrence could suggest an extended range of the taxon if its identification can be verified. Tricrepicephalus texanus was recognized by Łabaj (Reference Łabaj2015) as coming from the upper Cedaria Biozone.
Crepicephalus Biozone
The only biozone-diagnostic taxon from the Crepicephalus Biozone is a single, small, poorly preserved pygidium of Crepicephalus. As mentioned above, Tricrepicephalus texanus is also known from the Crepicephalus Biozone and occurs just below the Crepicephalus sp. indet. specimen (Fig. 11).
Aphelaspis, Dicanthopyge, and Prehousia biozones
These biozones are not documented in the Nugget Canyon section, but might be represented in the unfossiliferous upper 40 m of the upper member.
Dunderbergia Biozone
Material collected by Stoyanow from the Peppersauce Sandstone in Peppersauce Canyon contains trilobites of the Dunderbergia Biozone. Although specific identifications cannot be made due to the lack of exoskeletons in these samples, similar taxa occur in the Dunderbergia Biozone of the Great Basin (Palmer, Reference Palmer1960, Reference Palmer1965). Present in the fauna from the Peppersauce Sandstone are: Elburgia sp. indet., Iddingsia sp. indet, and Strigambitus cf. S. transversus.
Palmer (Reference Palmer1965) reported species of Elburgia and Strigambitus transversus from the Dunderbergia Biozone and species of Iddingsia from the upper Dunderbergia to Elvina biozones of Nevada and Utah. Sundberg et al. (Reference Sundberg, Cothren and Dehler2024) reported Iddingsia cf. I. intermedia from the middle part of the Dunderbergia Biozone.
Elvinia, Taenicephalus, and Ellipsocephaloides biozones
These biozones are not documented in the Nugget Canyon section.
Discussion
Paleontology
Abrigo Formation trilobites are predominantly forms that are widespread along the Laurentian margin, with taxa known from localities ranging from Sonora, Mexico to British Columbia, Canada, and from Georgia to western Newfoundland, Canada (Lochman, Reference Lochman1938; Schwimmer, Reference Schwimmer1989; Pratt, Reference Pratt1992; Sundberg and Cuen-Romero, Reference Sundberg and Cuen-Romero2021). As Palmer (in Gilluly, Reference Gilluly1956) noted for other Abrigo Formation localities, the Nugget Canyon assemblage seems most similar to that of the Riley Formation of Texas (Palmer, Reference Palmer1954b). Described trilobites were transported, but the assemblage is dominated by ptychopariids, which are thought to be epibenthic forms that lived on or near the seafloor. There are also geographically cosmopolitan forms like Bolaspidella and rare occurrences of agnostoids, which might have inhabited the open ocean realm. The taxa are typical of environments characterized by normal marine salinity, which is consistent with the hypothesized habitats represented by other invertebrate fossils in the succession, e.g., hyolithids, brachiopods, and echinoderms. The trilobites figured here are known from a variety of peritidal to deep subtidal settings elsewhere, which is internally consistent with the environments thought to be represented by the Abrigo Formation across southern Arizona (Łabaj and Pratt, Reference Łabaj and Pratt2016).
Diachroneity of lithostratigraphic units
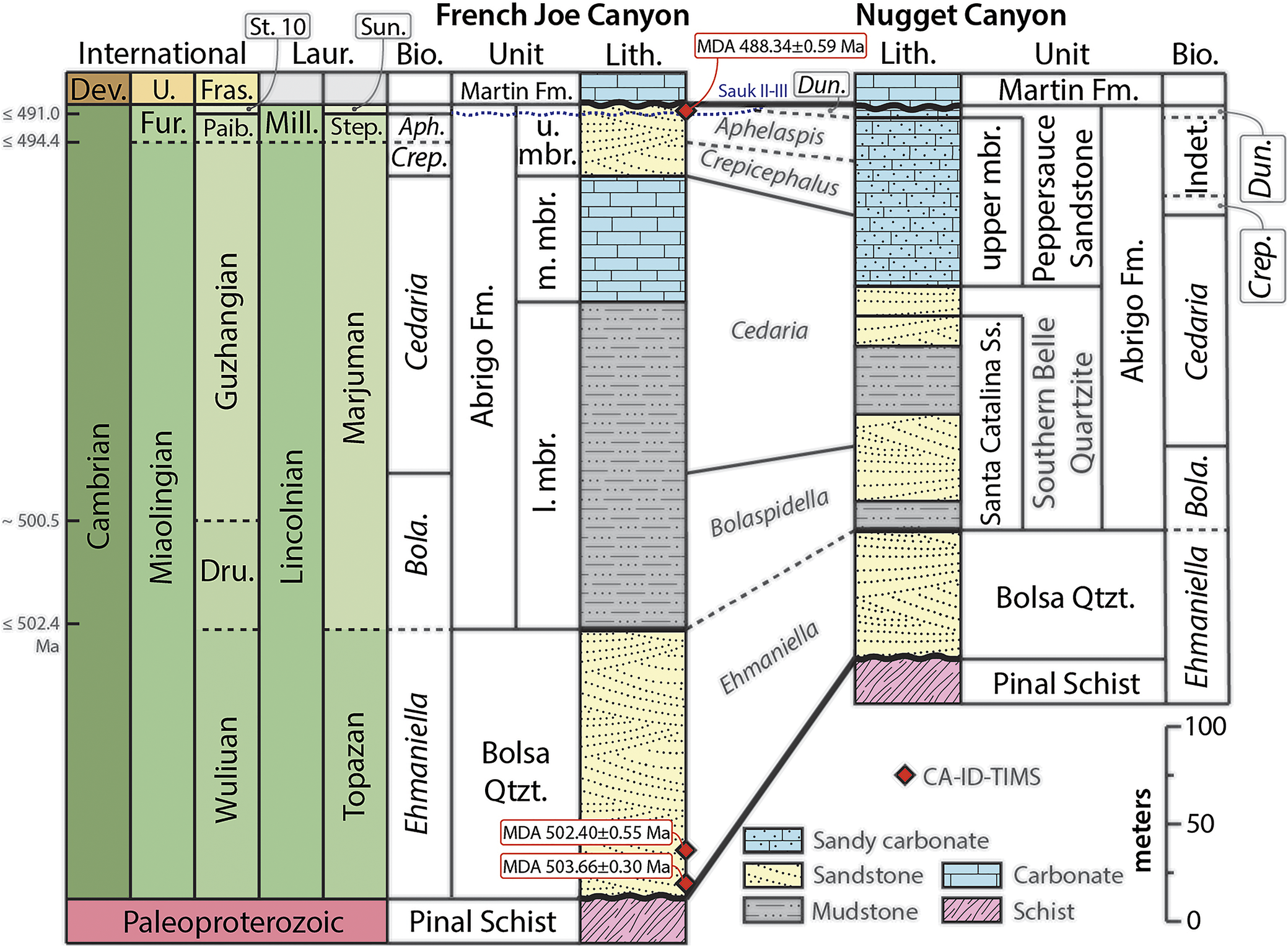
The Bolsa Quartzite and Abrigo Formation are part of an onlapping and time-transgressive sedimentary package deposited as a rising Cambrian Sea advanced cratonward toward the Laurentian interior. These relationships make it difficult to generalize about the age of the units, their stratigraphic correlation, and comparing these units across large geographic areas, whether those be to other exposures in Arizona or to hypothesized coeval strata in New Mexico and Texas (Sabins, Reference Sabins1957; Hayes, Reference Hayes1975, Reference Hayes, Callender, Witt, Clemons and James1978). For example, at French Joe Canyon, the Bolsa Quartzite is entirely Ehmaniella Biozone (Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025), whereas in the Dos Cabezas area and Blue Mountains to the east, the unit spans the Cedaria to Aphelaspis biozones (Wilson and Flower in Sabins, Reference Sabins1957). Strikingly, to the south and west of these exposures, these same biostratigraphic zones are present in the upper Abrigo Formation, not in the underlying Bolsa Quartzite (Łabaj, Reference Łabaj2015; Łabaj and Pratt, Reference Łabaj and Pratt2016; Muhlbauer et al., Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025). Moreover, the base of the Abrigo Formation at French Joe Canyon and vicinity spans the Bolaspidella Biozone (Łabaj, Reference Łabaj2015; Łabaj and Pratt, Reference Łabaj and Pratt2016) whereas to the east at Dos Cabezas and Blue Mountain, it is Aphelaspis Biozone or younger (Wilson in Sabins, Reference Sabins1957). These relationships highlight the need to be cautious when blending data from multiple mountain ranges in southern Arizona and vicinity or extending this biostratigraphy farther afield (Fig. 12).
Correlation of simplified measured sections for French Joe and Nugget canyons, compiled from Hayes (Reference Hayes, Callender, Witt, Clemons and James1978), Łabaj and Pratt (Reference Łabaj and Pratt2016), Muhlbauer et al. (Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025), and this study, annotated with new biostratigraphic controls. Unit names, trilobite biozones, and the geologic time scale are provided for reference (Cohen et al., Reference Cohen, Finney, Gibbard and Fan2013; Peng et al., Reference Peng, Babcock, Ahlberg and Gradstein2020). Maximum depositional ages (MDA) from chemical abrasion isotope-dilution thermal-ionization mass spectrometry (CA-ID TIMS) are provided for detrital zircon samples from French Joe Canyon. Detrital zircon geochronology and geochemistry were detailed by Muhlbauer et al. (Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025). The numerical date for the base of the Drumian is updated based on the results of Muhlbauer et al. (Reference Muhlbauer, Schmitz, Sundberg, Dehler, Karlstrom and Hagadorn2025); the base of the Paibian is updated using the MDA of Cothren et al. (Reference Cothren, Farrell, Sundberg, Dehler and Schmitz2022). Aph., Aphelaspis; Bio., biostratigraphy; Bola., Bolaspidella; Crep., Crepicephalus; Dev., Devonian; Dru., Drumian; Dun., Dunbergeria; Fm., Formation; Fras., Frasnian; Fur., Furongian; indet., indeterminate; l., lower; Laur., Laurentian; Lith., lithostratigraphy; m., middle; mbr., member; Mill., Millardan; Paib., Paibian; Qtzt., quartzite; Ss., sandstone; St. 10, Stage 10; Step., Steptoean; Sun., Sunwaptan; U./u., upper.

Figure 12. Long description
The diagram is organized into vertical columns. On the far left, the International time scale shows the Cambrian Period divided into Miaolingian and Furongian epochs, with stages including Wuliuan, Drumian, Guzhangian, and Paibian. Numerical dates are marked at 502.4 M a, 500.5 M a, 494.4 M a, and 491.0 M a. To the right, the Laurentian scale lists Millardan and Steptoean stages.
In the center, the French Joe Canyon section begins at the base with Pinal Schist, followed by the Bolsa Quartzite. Above this is the Abrigo Formation, divided into lower, middle, and upper members. The lithology transitions from yellow sandstone at the base to grey mudstone and blue carbonate layers. Three red diamonds indicate C A dash I D dash T I M S maximum depositional ages: 503.66 plus or minus 0.30 M a and 502.40 plus or minus 0.55 M a near the base, and 488.34 plus or minus 0.59 M a at the top of the Abrigo Formation.
The Nugget Canyon section on the right correlates with French Joe Canyon. It shows the Pinal Schist at the base, followed by Bolsa Quartzite, then the Santa Catalina Sandstone and Southern Belle Quartzite, which are equivalent to the Abrigo Formation. The top of both sections is capped by the Martin Formation.
Trilobite biozones are mapped between the sections, including Ehmaniella, Bolaspidella, Cedaria, Crepicephalus, and Aphelaspis. A legend at the bottom right identifies lithology symbols: sandy carbonate (blue with dots), sandstone (yellow with dots), mudstone (grey with dashes), carbonate (solid blue), and schist (pink with diagonal lines). A vertical scale bar on the right indicates height in meters from 0 to 100.
Acknowledgments
We are grateful to the National Science Foundation (grants EAR-1955115, EAR-1954634) and the patrons of the Denver Museum of Nature and Science for funding, to K. Kirschner for help in the field, and K. Karlstrom, M. Schmitz, L. Crossey, J. Foster, and B. Pratt for insight and advice. Thanks to the Bureau of Land Management for permits to collect on public land, as well as to L. Farrar and A. Hendy for assistance with reposited collections. We are also grateful for the constructive comments from the reviewers N.M. Handkamer and F.M. Tortello.
Competing interests
The authors declare none.
Appendix
Locality data. All material was collected from the Abrigo Limestone from Nugget Canyon, Santa Catalina Mountains, Arizona (~ 32°31’29” N, 110°42’34”W). Sections were measured on the south facing slope of the ridge that separates Peppersauce Canyon from Nugget Canyon, ~ 0.8 km (0.5 mi) west of Tule Springs. Stratigraphic levels of collections are from the top of the Bolsa Quartzite with the level from base of the measured section and/or the field locality numbers are in parentheses.
Section N1. Measured and collected 15 November 1996 and 22 February 1997. Base of section is the top of the Southern Belle Quartzite member.
Section N2. Measured and collected 30 November 1996. Base of section is the top of the Bolsa Quartzite and represents the 113 m of Santa Catalina Sandstone to the base of the Southern Bell Quartzite (22 m thick).
Section N3. Measured and collected 1 December 1996. Base of section 97 m is below the base of the Southern Bell Quartzite (31 m thick) and represents the upper portion of the Santa Catalina Sandstone that was poorly exposed in section N2.
Section N4. Measured and collected 22 February 1997. Base of section top of the Southern Bell Quartzite and represents the upper portion of the Santa Catalina Sandstone that was poorly exposed in section N2. Section starts about 30 m above wash.
OU IP01802 Brown sandstone at 7.5 m (N2-1). Missing from collection.
OU IP01803 Brown sandstone at 11 m (N2-2). Brachyaspidion sp. indet., Ehmaniella sp. indet.
OU IP01804 Brown sandstone at 12 m (N2-3). Ehmaniella sp. indet.
OU IP01805 Yellow brown sandstone at 30 m (N2-4). Neoblairella sp. indet., Altiocculus sp. indet.
OU IP 01806 Yellow brown sandstone at 40.5 m (24.5m, N3-1). Neoblairella sp. indet., Bolaspidella sp. indet.
OU IP 01807 Yellow brown sandstone at 36.5 m (20.5 m, N3-2). Neoblairella sp. indet.
OU IP01808 Yellow brown sandstone at 47 m (N2-5). Neoblairella sp. indet.
OU IP01809 Yellow brown sandstone at 53.5 m (37.5 m, N3-3). Altiocculus sp. indet., Eldoradia prospectensis, Neoblairella sp. indet.
OU IP01810 Yellow brown sandstone at 59 m (43 m, N3-4). Neoblairella sp. indet.
OU IP01811 Yellow brown sandstone at 62 m (46 m, N3-5). Neoblairella sp. indet.
OU IP01812 Yellow brown sandstone at 53 m (N2-6). Neoblairella sp. indet., Altiocculus sp. indet., Modocia cf. M. crassimarginata
OU IP01813 Yellow brown sandstone at 53.5 m (N2-7). Modocia oweni?
OU IP01814 Yellow brown sandstone at 66 m (50 m, N3-6). Neoblairella sp. indet.
OU IP01815 Yellow brown sandstone at 66 m (N2-8). Modocia oweni?
OU IP01816 Yellow brown sandstone at 72 m (N2-9). Altiocculus sp. indet., Modocia oweni?
OU IP01817 Yellow brown sandstone at 101 m float (85 m, N3-7). Bolaspididae gen. indet. sp. indet., Eldoradia linnarsonella
OU IP01818 Yellow limonitic dolostone at 147 m (3 m, N4-1a). Modocia centralis
OU IP01819 Same horizon as above, but collected along strike. Yellow limonitic dolostone at 147 m (3 m, N4-1b). Modocia centralis
OU IP01820 Yellow limonitic dolostone at 148 m (4 m, N4-2). Modocia centralis
OU IP01821 Calcareous yellow brown sandstone at 151 m (7 m, N1-1). Agnostidae gen. indet. sp. indet., Cedaria sp. indet., Tricrepicephalus texanus, Arapahoia ransomei
OU IP01822 Sandy dolostone at 155 m (13 m, N1-2) in float. OU lot #238683. Arapahoia ransomei, Modocia centralis
OU IP01823 Sandy dolostone at 164 m (20 m, N1-3). Arapahoia ransomei
OU IP01824 Sandy dolostone at 154 m (~10 m, N1-4). OU lot #238685. Tricrepicephalus texanus?
OU IP01825 Sandy dolostone at 168.5 m (~14.5 m, N1-5). OU lot #238686. Crepicephalus sp. indet.
OU IP01826 Sandy dolostone at 199 m (55 m, N1-6) in float. Articulate Brachiopoda, no trilobites.
Stoyanow’s localities. Material collected by Stoyanow from Peppersauce Canyon are listed only by formations in his unpublished text. We here assign them to the members of the Abrigo Formation, but the exact horizons that the specimens were collected are unknown.
LACMIP loc. 16249. Yellow limonitic dolostone from a few meters above the Southern Bell Quartzite. Arapahoia ransomei
LACMIP loc. 29164. Sandstone from the Peppersauce Sandstone, upper Abrigo Formation. Elburgia sp. indet., Iddingsia sp. indet., Strigambitus cf. S. transversus
LACMIP loc. 29165. Yellow limonitic dolostone from a few meters above the Southern Bell Quartzite. Tricrepicephalus texanus
LACMIP loc. 29166. Sandstone from the Peppersauce Sandstone, upper Abrigo Formation. Elburgia sp. indet., Strigambitus cf. S. transversus