


Introduction
Rats are considered one of the most important laboratory animals in research conducted in many fields, particularly biology and physiology. The development of different mutant and transgenic rat strains has created a global need for the preservation and distribution of wild-type strains. In this regard, the successful freezing of gametes and embryos is of great importance in the study of rat sperm biology by genome banking of transgenic and mutant rat strains. However, compared with many other species, the freezability of rat sperm is quite low. Rat sperm cells have a more sensitive structure compformulated based on the protocolared to the spermatozoa of other domestic animal species (Kim et al., Reference Kim, Agca and Agca2012). The head region of rat spermatozoa has a different structure compared with mammals; their tails are longer than those of other species, their plasma membrane has low water permeability, and they are extremely sensitive to factors such as cold shock and mechanical stress (e.g., centrifugation, mixing, and pipetting) during the cryopreservation process, negatively affecting sperm freezing success (Nakatsukasa et al., Reference Nakatsukasa, Inomata, Ikeda, Shino and Kashiwazaki2001; Nakata et al., Reference Nakata, Okuda, Yamashita, Nakauchi, Ito and Kashiwazaki2012; Nakagata et al., Reference Nakagata, Mikoda, Nakao, Nakatsukasa and Takeo2020; Dayanç and Yıldız, Reference Dayanç and Yıldız2025).
Increased stress conditions during cryopreservation can lead to overproduction of reactive oxygen species (ROS). The production of ROS at the physiological level is essential for normal cellular functions. However, excessive production can cause damage to the sperm plasma membrane and disrupt genome integrity. Therefore, antioxidant substances added to sperm freezing mediums play a significant role in reducing oxidative stress and freezing sperm cells in a healthy way. Controlling ROS levels is also critical for maintaining sperm motility and viability. Numerous studies have reported that various antioxidants have beneficial effects in the treatment of male infertility (Li et al., Reference Li, Lin, Liu, Xiao and Liu2010; Ranjan et al., Reference Ranjan, Kumar, Swain, Singh, Kharche and Chauhan2022; Sinclair Reference Sinclair2000). In this study, ascorbic acid (Vitamin C), a powerful antioxidant, and biotin, a vitamin that plays an indirect role in the antioxidant defence system, were added to the cryopreservation diluent to increase cryosurvival.
Ascorbic acid is a powerful non-enzymatic, water-soluble ROS scavenger (Silva Reference Silva2006). Due to this feature, it has been used as an additive in cryopreservation protocols in many studies and clinical trials (Asadpour et al., Reference Asadpour, Jafari and Tayefi-Nasrabad2011; Hu et al., Reference Hu, Tian, Zhao, Zan, Wang, Li and Xin2010; Li et al., Reference Li, Lin, Liu, Xiao and Liu2010; Paudel et al., Reference Paudel, Kumar, Meur and Kumaresan2010).
Biotin, also known as vitamin B7, is an essential micronutrient that is necessary for the normal growth and development of the body. Although the effect of biotin on sperm function is not fully elucidated, some studies show that it increases motility and decreases ROS levels (Kalthur et al., Reference Kalthur, Salian, Keyvanifard, Sreedharan, Thomas, Kumar and Adiga2012; Montazari et al Reference Montazari, Golshan-Iranpour, Dashti, Ishaqi and Dashti2021; Ranjan et al., Reference Ranjan, Kumar, Swain, Singh, Kharche and Chauhan2022).
This study aimed to examine the effects of adding Ascorbic acid (4 mg/mL), an antioxidant not previously investigated in rat models, and biotin (20 nM), a component that supports the antioxidant defence system, to the cryopreservation diluent during long-term cryopreservation of rat sperm. In this regard, sperm quality parameters such as motility, viability, plasma membrane integrity, and acrosome morphology ratios were analysed. In addition, the expression levels of Hsp40, Hsp60, and Hsp70 genes, which potentially play a role in sperm function and the heat shock response during the cryopreservation process, were examined. In conclusion, this study aims to establish a scientific basis for the development of innovative, effective, and reliable cryoprotectant formulations in the field of cryobiology.
Materials and method
Animals
In the study, 24 adult male Wistar Albino rats aged 10–16 weeks were used. The rats were kept at a room temperature of 24 ± 3°C, 60% relative humidity, and a 12-hour light/12-hour dark cycle. Their nutrition was provided by pellets fed ad libitum and tap water. All procedures were carried out in line with the approval of the Hatay Mustafa Kemal University Animal Experiments Local Ethics Committee (Ethics Approval No: 2024/ 11-01).
Chemicals
All chemical substances were obtained from Sigma Chemical Company (St. Louis, MO). Human Tubal Fluid (HTF) medium was formulated based on the protocol by Quinn et al. (Reference Quinn, Kerin and Warnes1985). The cryopreservation medium (CPA) was developed by adapting the protocol defined by Nakatsukasa et al. (Reference Nakatsukasa, Inomata, Ikeda, Shino and Kashiwazaki2001). In this process, 23% fresh hen egg yolk, 8% lactose monohydrate, 1 mg/mL streptomycin sulphate, and 1000 IU penicillin G potassium were added to pure distilled water and mixed for 5 min at a temperature of +4 °C. Then the mixture was centrifuged at 1600 × g for 15 min. After centrifugation, the upper phase was separated and collected, and 0.1% ATP (adenosine 5′-triphosphate) was added to this phase. The resulting diluent was divided to form two separate experimental groups. The Ascorbic acid group was prepared by adding 4 mg/mL Ascorbic acid to one group, and the Biotin group was prepared by adding 20 nM biotin to the other group. CPA without any additions was defined as the control group. Finally, the pH values of all experimental groups were adjusted to 7.4 using a pH metre (Thermo Scientific, USA) with a pre-prepared 10% Tris-amine and distilled water mixture.
Collection, freezing, and thawing of spermatozoa
The rats were sacrificed by cervical dislocation method after anaesthesia. After ensuring sterile conditions, both cauda epididymides were removed and placed in 35 mm diameter Petri dishes (Nunc™, Massachusetts, USA) containing 1 mL of CPA, which were kept at room temperature. The procedures for spermatozoa dilution and freezing were performed based on the protocol reported by Nakagata et al. (Reference Nakagata, Mikoda, Nakao, Nakatsukasa and Takeo2020), with some modifications. Cauda epididymides were fixed with blunt-tipped forceps under a stereo microscope (Euromex/Nexius Zoom, Netherlands). Then, 10–12 deep incisions were made in a Petri dish using sharp-tipped scissors. Thus, spermatozoa were transferred to the CPA medium. For the release and equilibration of spermatozoa from the cauda, Petri dishes were placed on a metal plate pre-positioned inside a styrofoam box and incubated for 10 min. After incubation, 30 µL HTF, 10 mm air gap, 150 µL sperm suspension, another 10 mm air gap, and 30 µL HTF were loaded into 0.25 mL straws (IMV, France) in that order. The straws were then pressed shut. For the cultivation process, the straws were kept on the metal sheet placed on the ice for 30 min. The equipment used during this process (syringe connector, straws, and HTF) was prepared to be at a temperature of 0°C. For the freezing process, the straws were placed in a styrofoam box measuring 270 × 220 × 270 mm, on a straw stand located 4 cm above the liquid nitrogen level, and kept in liquid nitrogen vapour for 10 min.
Then, the freezing process was completed by immersing the straws directly in liquid nitrogen. The thawing process was carried out by soaking the straws in a water bath at 37°C (decongelator, CITO 026897, IMV, France) for 30 s. The dissolved straw content was transferred to 1 mL of HTF in a carbon dioxide incubator operating at 37°C in a 5% CO2 atmosphere.
Sperm concentration
Sperm concentration was determined using a Makler counting chamber (Sefi-Medical Instruments, Haifa, Israel) in accordance with the World Health Organization (WHO, 1999) guidelines. Sperm samples were immobilized by diluting 1:10 with phosphate-buffered saline (PBS) solution containing 10 mM formaldehyde. Taking 10 μL of the prepared suspension, cell counts were performed in 10 different square areas on the counting chamber under a 20X objective on an (Olympus CX31, JP) microscope. Sperm concentration was calculated in millions/mL using the obtained data. Measurements were repeated three times for each sample, and the results were averaged.
Sperm motility
Sperm motility analysis was performed using a Makler counting chamber (Sefi-Medical Instruments, Haifa, Israel) in accordance with World Health Organization (WHO, 1999) criteria. For this purpose, 20 μL of HTF and sperm suspension were taken and placed on a Makler slide preheated to 37°C. The prepared samples were examined at 20X magnification using a temperature-controlled phase contrast microscope (Olympus CX31, JP). The number of spermatozoa showing forward linear movement was determined in 100 small squares on the micrometer slide, and the ratio of total motile and immotile cells was calculated. For each sample, the analysis was repeated three times. Motility results were expressed as a percentage (%) by taking the average of the obtained data.
Sperm viability
Sperm viability analysis was performed based on the eosin-nigrosin staining method described by Agarwal et al. (Reference Agarwal, Gupta and Sharma2016). 10 μL of sperm samples were taken and mixed with a total of 20 μL of 1% eosin and 10% nigrosine solutions. From the resulting mixture, smear preparations were prepared on slides and then dried on a surface at 60°C within 2–3 s. Viability assessment was performed using a phase contrast microscope (Olympus CX31, Japan) at 40X objective magnification. At least 300 spermatozoa were counted and analysed in each preparation. Spermatozoa with heads stained red or dark pink were considered dead, while those with unstained heads were considered alive. The viability rate was calculated as a percentage (%) over the total count.
Sperm plasma membrane integrity
To evaluate the plasma membrane integrity of spermatozoa, the Hypo-Osmotic Swelling Test (HOST) defined by Jeyendran et al. (Reference Jeyendran, Van der Ven, Perez-Pelaez, Crabo and Zaneveld1984) was applied with modifications. The solution used in the test was prepared by dissolving 1.1 g of fructose and 0.55 g of sodium citrate in 100 mL of distilled water, and the osmotic pressure was measured with an osmometer (Osmomat 3000, Gonotec, USA). 100 μL of HOST solution and 10 μL of sperm sample were taken and transferred to a 1.5 mL Eppendorf tube. The prepared mixture was placed in the incubator at 37°C and incubated for 45 min. At the end of the incubation period, a sample was taken and placed on a slide preheated to 37°C and examined under a 40X objective using a phase contrast microscope (Olympus CX31, Japan). Spermatozoa exhibiting swelling or coiling in the tail region were classified as HOST positive (+), while those showing no morphological changes were classified as HOST negative (−). In each sample, a total of 200 spermatozoa were counted, and the percentage (%) of HOST-positive spermatozoa was recorded.
Acrosome status
Abnormal acrosome assessment was performed based on the method reported by Somfai et al. (Reference Somfai, Bodo, Nagy, Gocza, Ivancsics and Kovács2002). Hancock’s solution (Hancock Reference Hancock1952) was used in the analysis. 500 μL of the final solution was taken into the Eppendorf tube, and 50 μL of the sperm sample was added to it. 50 μL of the resulting mixture was taken and dripped onto the slide, and covered with a coverslip. The prepared specimens were examined under a microscope using a 100X objective magnification with immersion oil (Günay et al., Reference Günay, Nur and Soylu2003). A total of 300 spermatozoa were evaluated in each sample, and the percentage (%) of cells with acrosome damage was recorded.
Live/Dead viability evaluation
Rat sperm were examined for viability using a Live/Dead™ sperm viability kit after cryopreservation (L7011Thermo Fisher Scientific, USA). Post-thaw sperm samples were diluted approximately 1:10 in a HEPES-buffered saline solution containing bovine serum albumin (10 mM HEPES, 150 mM NaCl, 10% BSA, pH 7.4) until a sperm count of 1 million per millilitre was achieved. One mL of diluted sperm samples was taken, and 5 μL of SYBR-14 stock solution (diluted 1:10 with DMSO) was added to it. The mixture was incubated at 36°C for 5 min. Then, 5µl of Propidium Iodide (PI) was added to achieve a final concentration of 12 µM, and incubated for 5 min. Sperm samples were evaluated under a fluorescent microscope (Eclipse Ni, Nikon) at 400 times magnification according to the colour of the head part (excitation wavelength of 490 nm, emission wavelength of 520 nm). Sperm heads showing red fluorescence were considered dead, and sperm heads showing green fluorescence were considered alive. A total of 400 spermatozoa were counted, and the results were expressed in percentage (%) (Garner and Johnson, Reference Garner and Johnson1995).
Total RNA isolation
Sperm suspension was washed with PBS and centrifuged at 150 × g for 10 min at 4°C. To eliminate somatic cell contamination, the pellets were treated with a somatic cell lysis buffer (0.05% sodium dodecyl sulphate and 0.25% Triton X-100 in distilled water) and incubated on ice for 60 min. Total RNA was extracted using TRIzol reagent (TRIzol® Reagent, Ambion) following the manufacturer’s protocol. Carefully dissected semen sample was submerged in 1 mL of TRIzol reagent and homogenized for 45 s with a homogenizer. The homogenate was incubated for 5 min and 200 μL chloroform was added; shaken vigorously. The mixture was centrifuged at 12,000 × g at 4°C for 15 min. Following this centrifugation step, the upper aqueous layer was transferred to another nuclease-free 1.5 mL tube and 500 μL of isopropanol was added; shaken vigorously. The sample was centrifuged at 12,000 × g at 4°C for 10 min to pellet the precipitated RNA. The RNA-containing pellet was washed twice with 500 μL of 70% ethanol and centrifuged at 7,500 × g at 4°C for 5 min. Final RNA pellet was washed with 500 μL of 99% ethanol and centrifuged at 7,500 × g at 4°C for 5 min. The RNA-containing pellet was air-dried and dissolved in 30 mL of nuclease-free water. The RNA samples were stored at −80°C for downstream application later. The concentration and purity of RNA were assessed by measuring absorbance at 260/280 nm using a NanoDrop™ 1000 spectrophotometer (Thermo Fisher Scientific, USA).
cDNA synthesis
Complementary DNA (cDNA) was synthesized from 1 μg of total RNA using the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, USA). The reaction included 1 μL of oligo(dT)18 primer and 1 μL of random hexamer primer, which were mixed with the RNA and incubated at 65°C for 5 min, followed by immediate cooling on ice. Subsequently, 8 μL of cDNA synthesis mix (comprising 5X reaction buffer, RiboLock RNase Inhibitor, dNTP mix, and RevertAid M-MuLV reverse transcriptase) was added. The reaction was incubated at 42°C for 60 min, followed by 25°C for 5 min, and terminated by heating at 70°C for 5 min.
Quantitation of gene expression by RT-qPCR
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) was performed using RealQ Plus 2X Master Mix Green (Ampliqon, Denmark). Gene-specific primers of Hsp40, Hsp60, and Hsp70, which belong to the heat shock protein family, were used (Table 1). A total of 100 ng of cDNA was used per 25 μL reaction, which included 150 nM of each primer and 12.5 μL of RealQ Plus Master Mix. Reactions were run in a CFX96 system (Bio-Rad) with the following thermal profile: initial denaturation at 95°C for 15 min, followed by 40 cycles of 95°C for 30 s, 60°C for 60 s, and 72°C for 30 s. Gene expression levels were normalized against housekeeping gene (GAPDH) using the 2ΔΔCt method (Livak and Schmittgen Reference Livak and Schmittgen2001). Each experimental group included eight animals (n = 8 per group) representing biological replicates. For the RT-qPCR analysis, each cDNA sample was analysed in duplicate as technical replicates; data were reported as fold-change using the 2ΔΔCt method.
Primer sets used in the study

Statistical analysis
The SPSS 22.0 software was used for statistical evaluations (IBM Corporation, Armonk, NY). In the evaluation of spermatological parameters, one-way analysis of variance (ANOVA) was used to compare the significance of the difference between the groups. The results were evaluated as mean ± SE, and those with p < 0.05 were considered statistically significant. The Duncan test was used to indicate the significance of the difference between the groups as a result of the analysis. For the gene expression analysis, a one-way ANOVA followed by a post hoc Tukey test was performed using GraphPad Prism 9.1.1 (GraphPad, San Diego, CA, USA). Statistical significance was considered at p < 0.05. The use of different statistical software and post hoc tests was based on the distinct data structures, variability, and analytical requirements of the spermatological and gene expression datasets, ensuring appropriate and reliable statistical evaluation.
Results
Spermatological parameters determined after freezing-thawing
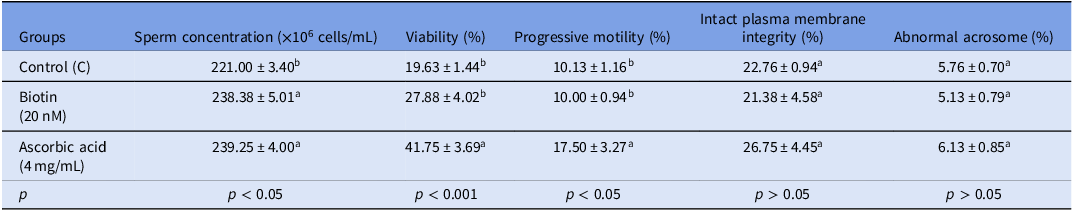
The sperm concentration, viability, progressive motility, intact plasma membrane integrity, and abnormal acrosome ratio in the study groups after post-thawing processes are presented in Table 2.
Spermatological findings were determined after freezing-thawing in the study groups (n = 8)

a–b Different letters in the same column indicate differences between groups (p < 0.05–0.001).
Spermatozoon concentration
Determination of sperm concentration was performed to verify the required cell count for analyses to be conducted after the freezing-thawing process and to standardize experimental procedures. In the analysis, statistically significant differences were found between the groups (p < 0.05). The highest sperm concentration was found in the Ascorbic acid (4 mg/mL) group (239.25 ± 4.00) and the biotin (20 nM) group (238.38 ± 5.01). There was no difference between the study groups (p > 0.05). Although the results show a statistical difference, they provide the optimum number of cells required for the study. In addition, this difference may be due to researcher-induced variations or qualitative factors.
Rate of viable spermatozoon
When the viability rates of sperm cells were evaluated, it was determined that there were statistically significant differences between the groups (p < 0.05). The highest viability rate was found in the Ascorbic acid group (41.75 ± 3.69). This rate was found to be significantly higher compared with the control group (19.63 ± 1.44) (p < 0.001). In the biotin-added group, the viability rate was found to be 27.88 ± 4.02, but no statistically significant difference was found compared with the control group (p > 0.05). These findings suggest that Ascorbic acid may increase sperm viability after freezing and thawing.
Motility values
The highest progressive motility rate was measured in the Ascorbic acid group (17.50 ± 3.27), and this rate was found to be statistically significantly higher compared with the control group (10.13 ± 1.16) (p < 0.05). In the biotin group, the progressive motility rate was found to be 10.00 ± 0.94, and no statistically significant difference was found compared with the control group (p > 0.05). These findings indicate that Ascorbic acid can significantly increase sperm progressive motility during the cryopreservation process. Although the effect of biotin on motility is not statistically significant, its potential to support the antioxidant defence system indicates that it may indirectly positively affect sperm function.
Plasma membrane integrity ratio
According to the results of the plasma membrane integrity test performed to evaluate sperm cell membrane integrity, no statistically significant differences were found between the groups (p > 0.05). The highest HOST positive rate was determined to be 26.75 ± 4.45 in the group supplemented with 4 mg/mL of Ascorbic acid. This rate was found to be significantly higher compared with the control group (22.76 ± 0.94) (p > 0.05). In the group supplemented with biotin at a concentration of 20 nM, this ratio was determined to be 21.38 ± 4.58; however, no significant difference was found between the control group and the other groups (p > 0.05).
Abnormal acrosome rate
There was no statistically significant difference between the groups in terms of abnormal acrosome rate (p > 0.05). The abnormal acrosome rate was determined to be 5.76 ± 0.70 in the control group, 6.13 ± 0.85 in the Ascorbic acid group, and 5.13 ± 0.79 in the biotin group.
Live/dead cell ratios
The viability rates of spermatozoa were evaluated using the fluorescent method and analysed using the SYBR-14/PI test (Figure 1). No statistically significant differences were found between the groups (p > 0.05). The highest viability value was measured in the Ascorbic acid group (32.75 ± 4.14%) and did not show a statistically significant difference compared with the control group (25.25 ± 6.70%) (p > 0.05). In the biotin-added group, the viability rate was found to be % 20.75 ± 4.01, and no statistically significant difference was found compared with the control group (p > 0.05).
Assessment of viability using the Live/Dead™ Viability Kit. It was evaluated with the SYBR-14/PI test. Spermatozoa stained green were classified as alive (a), while those stained red were classified as dead (b), and the viability index (%) of cells in each group was quantitatively determined (c). Data are shown as mean ± standard error (n = 8 in each group). The values indicated by the same superscripts did not differ statistically significantly (p > 0.05).

Effects of biotin and ascorbic acid supplementation on the expression of apoptosis-related genes in frozen–thawed sperm using real-time PCR
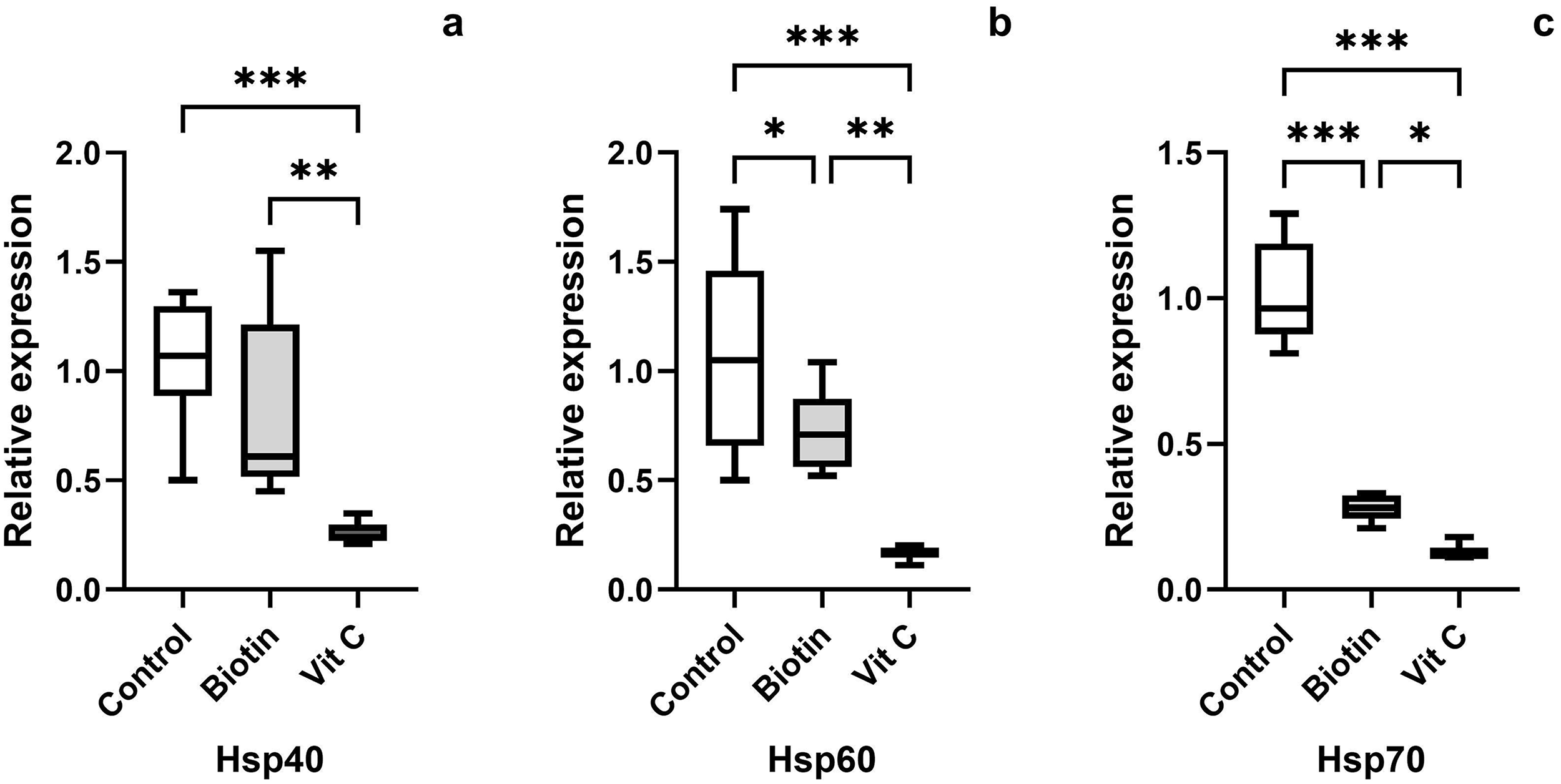
To investigate whether biotin and ascorbic acid can alleviate cryodamage in spermatozoa after frozen-thawed were analysed the expression of heat shock protein-related genes. Addition of 20 nM Biotin and 4 mg/mL Ascorbic acid to the cryoprotectant agent affected Hsp40, Hsp60, and Hsp70 mRNA levels compared with the control group (Figure 2). Hsp40 gene expression level was statistically significantly decreased in the ascorbic acid group compared with the control group (p < 0.001). While no significant difference was observed between the control group and the biotin group (p > 0.05), a difference was observed between the biotin and ascorbic acid groups (p < 0.01). Hsp60 gene expression level showed changes in biotin and ascorbic acid groups compared with the control group. The decreases in all groups were statistically significant. A markedly decline was observed in biotin (p < 0.05) and ascorbic acid (p < 0.001) groups in comparison to the control group. It was also observed that a statistically significant decrease was observed in the ascorbic acid group compared with the biotin group (p < 0.01). It was observed that the Hsp70 gene expression level decreased in the biotin (p < 0.001) and ascorbic acid (p < 0.001) groups compared with the control group. In addition, it was observed that the ascorbic acid group decreased statistically significantly compared with the biotin group (p < 0.05). These results suggest that freeze–thaw induces heat stress in rat sperm, and the addition of vitamin and micronutrient (Biotin) and antioxidant (vitamin C) to the cryoprotectant agent reduces heat shock after thawing.
Quantitative PCR analysis of the relative expression of Hsp40 (a), Hsp60 (b), and Hsp70 genes (c) on frozen-thawed epididymal rat sperm. All data were presented as mean ± SEM (n = 8 in each group). *p < 0.05, **p < 0.01, and ***p < 0.001 vs the other group.

Discussion
Cryopreservation of rat spermatozoa is of great importance for genome banking in the preservation of transgenic and mutant rat strains. However, rat spermatozoa are more sensitive to freezing and thawing processes compared with other mammalian species (Nakatsukasa et al., Reference Nakatsukasa, Inomata, Ikeda, Shino and Kashiwazaki2001; Seita et al., Reference Seita, Sugio, Ito and Kashiwazaki2009). Therefore, developing effective cryoprotective strategies for rat sperm is quite difficult due to existing challenges. This study demonstrated that the addition of 20 nM biotin and 4 mg/mL Ascorbic acid separately to the cryopreservation medium produced protective effects on sperm quality after cryopreservation. The dose of ascorbic acid used in this study was selected based on the unique structural characteristics of rat sperm plasma membranes. Rat spermatozoa contain a higher proportion of polyunsaturated fatty acids and exhibit distinctive membrane fluidity compared with other mammalian species, making them particularly susceptible to oxidative stress during cryopreservation. Consistent with the findings of Hu et al. (Reference Hu, Tian, Zhao, Zan, Wang, Li and Xin2010), supplementation with ascorbic acid at comparable or higher concentrations has been shown to enhance post-thaw sperm quality without evidence of pro-oxidant effects. In line with these observations, our results demonstrate that this concentration improved motility and viability, and numerically increased plasma membrane integrity, suggesting that under controlled cryopreservation conditions, ascorbic acid provides effective antioxidant protection without inducing pro-oxidant activity.
A significant difference was found between the groups in terms of sperm concentration (p < 0.05), with the biotin (238.38 ± 5.01) and Ascorbic acid (239.25 ± 4.00) groups showing higher values than the control group. However, it was assessed that these differences were not at a level that would affect experimental standardization. This observation is consistent with the findings of Kempinas and Lamano-Carvalho (Reference Kempinas and Lamano-Carvalho1988), who reported that sperm concentrations in the rat cauda epididymis can naturally vary over a relatively wide range even after enzymatic homogenization. In their study, sperm concentrations ranged from 152.5 to 230.0 × 107 spermatozoa/ml, with a mean of 187.7 (± 5.6 SEM) × 107 spermatozoa/ml. Such variability is considered normal and is mainly attributed to individual differences among animals and the uneven distribution of spermatozoa within the cauda epididymis. In the present study, since all animals were maintained under identical housing and feeding conditions, were of similar age and body weight, and samples were collected using the same standardized method by a single operator, the minor differences observed did not compromise the suitability of the samples for subsequent analyses.
In this study, sperm viability was determined both by eosin staining using a conventional light microscope and by the SYBR-14/PI fluorescent staining technique. Both methods provide significant information in assessing the damage caused by cryopreservation at the cellular level. According to eosin staining results, the highest viability rate after thawing was observed in the Ascorbic acid group (41.75 ± 3.69), and this value was found to be significantly higher compared with the control group (19.63 ± 1.44) (p < 0.05). Although the viability rate was higher in the biotin group (27.88 ± 4.02) than in the control group, no statistically significant difference was found (p > 0.05). These findings suggest that ascorbic acid has a strong cryoprotective and antioxidant effect on rat sperm, while biotin has a more limited but positive contribution. The protective effect of Ascorbic acid on sperm viability is also supported by studies conducted in different species. Mittal et al. (Reference Mittal, Anand, Madan, Yadav and Kumar2014) reported that the application of 5 mM Ascorbic acid in bull sperm significantly increased viability (82.06 ± 0.38), while Gangwar et al. (Reference Gangwar, Kharche, Ranjan, Kumar, Goel, Jindal and Agarwal2015) reported that 56.78 µM Ascorbic acid in ram sperm had positive effects on post-thaw viability (68.21 ± 1.37). Similarly, Wittayarat et al. (Reference Wittayarat, Kimura, Kodama, Namula, Chatdarong, Techakumphu and Otoi2012) reported that the addition of 0.5 mM Ascorbic acid to dog sperm resulted in high viability. Azawi and Hussein (Reference Azawi and Hussein2013) reported that in short-term storage of ram sperm (+5°C), ascorbic acid supplementation increased sperm viability percentage from 40.9 ± 1.8 to 59.7 ± 1.3. These findings indicate that Ascorbic acid supports sperm viability in different types by reducing cell damage through its antioxidant effect.
Although the viability rate was higher in the biotin group than in the control group, no statistically significant difference was found. However, it is considered that biotin may indirectly protect sperm function through cellular metabolism and by reducing lipid peroxidation. Bazpour et al. (Reference Bazpour, Vakili, Mirzadeh and Aghaei2025) demonstrated that 25 nM biotin improved viability and membrane integrity in turkey sperm during short-term storage, while Ranjan et al. (Reference Ranjan, Kumar, Swain, Singh, Kharche and Chauhan2022) showed that 3 µM biotin significantly increased viability (51.90 ± 2.28%) in ram sperm. In the analyses performed by the SYBR-14/PI staining method, the viability rates of the control, biotin, and Ascorbic acid groups after thawing were determined as 24.25 ± 6.70, 20.75 ± 4.01, and 32.75 ± 4.14, respectively. Although there was no statistically significant difference between the groups (p > 0.05), the highest rate (32.75 ± 4.14) was observed in the ascorbic acid group. This result suggests that Ascorbic acid supports cell functions after cryopreservation by maintaining membrane integrity.
The quantitative difference between the findings obtained in both methods may be due to the nature of the evaluation techniques. The eosin staining method provides a direct visual assessment based on cell morphology and membrane integrity, while the SYBR-14/PI fluorescent method provides a more precise measurement by distinguishing between live and dead cells based on membrane permeability. Since even minimal changes in membrane permeability can be classified as dead cells in fluorescence-based analyses, this method generally yields lower viability rates than manual evaluation. This difference was clearly observed in our study. In the literature, it is reported that SYBR-14/PI analysis has high sensitivity in determining sperm viability after cryopreservation and provides less subjective results than traditional staining methods. Giaretta et al. (Reference Giaretta, Estrada, Bucci, Spinaci, Rodríguez-Gil and Yeste2015) found that adding 100 μM ascorbic acid to boar sperm significantly increased post-thaw viability rates. Pandey (Reference Pandey2025) reported that 0.5 mmol ascorbic acid significantly increased the percentage of viable cells (p = 0.0009) in spotted dogfish (Anarhichas Minor) sperm, and 1.0 mmol ascorbic acid provided moderate improvement. Bodu et al. (Reference Bodu, Öztürk, Kılınç, Hatipoğlu, Ataman, Bucak and Kul2024) reported that the addition of 5 mM/mL ascorbic acid to boar sperm yielded positive results in SYBR-14/PI analysis. In the present study, the highest viability rate obtained with both methods in the Ascorbic acid group supports the antioxidant and membrane-stabilizing effects of this compound. The effect of biotin was more limited. Hence, it can be concluded that ascorbic acid is a more potent antioxidant and cryoprotectant agent compared with biotin in enhancing post-cryopreservation viability in rat sperm.
Sperm motility is considered an indicator of cell viability and functionality after freezing and thawing. In this study, the highest motility rate was determined in the Ascorbic acid group (17.50 ± 3.27) and found to be significantly higher compared with the control group (10.13 ± 1.16) (p < 0.05). Although the motility of the biotin group was similar to that of the control group, it was found to be higher than the motility value reported by Nakatsukasa et al. (Reference Nakatsukasa, Inomata, Ikeda, Shino and Kashiwazaki2001) (4.6 ± 0.1). This positive effect of Ascorbic acid on motility has also been reported in the sperm of bulls (Mittal et al., Reference Mittal, Anand, Madan, Yadav and Kumar2014), rams (Shehab-El-Deen et al., Reference Shehab-El-Deen, Ali and Al-Sharari2022), goats (Gangwar et al., Reference Gangwar, Kharche, Ranjan, Kumar, Goel, Jindal and Agarwal2015), dogs (Wittayarat et al., Reference Wittayarat, Kimura, Kodama, Namula, Chatdarong, Techakumphu and Otoi2012), and roosters (Amini et al., Reference Amini, Kohram, Zare Shahaneh, Zhandi, Sharideh and Nabi2015). The increase in motility (50.61 ± 1.62) reported by Hu et al. (Reference Hu, Tian, Zhao, Zan, Wang, Li and Xin2010) with the application of 4.5 mg/mL ascorbic acid in bull sperm is consistent with the results of our study. Similarly, Paudel et al. (Reference Paudel, Kumar, Meur and Kumaresan2010) demonstrated that 10 mM ascorbic acid improved sperm parameters in crossbred bull sperm, while Li et al. (Reference Li, Lin, Liu, Xiao and Liu2010) showed that 300 µM ascorbate provided the best cryoprotection in human sperm. Asadpour et al. (Reference Asadpour, Jafari and Tayefi-Nasrabad2011) reported that 2 mM Ascorbic acid increased motility in bull sperm. In addition, it has been reported that Ascorbic acid supplementation increases sperm motility from 35.9 ± 1.9% to 54.7 ± 1.1% in short-term storage, at +5°C (Azawi and Hussein Reference Azawi and Hussein2013). These studies confirm that Ascorbic acid promotes motility during cryopreservation and short-term storage in different species.
The plasma membrane integrity ratio was higher in the Ascorbic acid group (26.75 ± 4.45) compared with the control (22.76 ± 0.94) and biotin (21.38 ± 4.58) groups, but the difference was not statistically significant (p > 0.05). This result suggests that ascorbic acid may maintain membrane stability by reducing oxidative stress, but the effect may depend on dose and species differences. Similarly, biotin also contributed to membrane stability but did not make a significant difference compared with the control group. The findings are consistent with those reported by Franco et al. (Reference Franco, Chaveiro, Góis and da Silva2013), who found that ascorbic acid supplementation (0.45, 0.9, and 1.8 g/L) improved plasma membrane integrity in horse sperm, but this effect was not statistically significant (p > 0.05). On the other hand, Gangwar et al. (Reference Gangwar, Kharche, Ranjan, Kumar, Goel, Jindal and Agarwal2015) reported that plasma membrane integrity was higher in goat sperm at an ascorbic acid concentration of 56.78 μM compared with the control group. Bazpour et al. (Reference Bazpour, Vakili, Mirzadeh and Aghaei2025) reported that a biotin concentration of 25 nM significantly improved plasma membrane integrity and sperm quality (p < 0.05) in turkey sperm during short-term storage. Similarly, Ranjan et al. (Reference Ranjan, Kumar, Swain, Singh, Kharche and Chauhan2022) demonstrated that a 3 µM biotin concentration in frozen ram sperm significantly increased plasma membrane integrity (51.86 ± 1.96) (p < 0.05).
There was no statistically significant difference between the control (5.76 ± 0.70), biotin (5.13 ± 0.79), and ascorbic acid (6.130.85) groups in terms of abnormal acrosome rates (p > 0.05). However, the biotin and Ascorbic acid groups showed numerically lower rates of abnormal acrosomes compared with the control group. Numerically lower abnormal rates indicate the protective effect of biotin on the acrosomal structure.
Heat shock proteins (HSPs) are molecular chaperones that regulate protein folding under cellular stress conditions and provide protection against oxidative damage. The heat stress generated during the freeze–thaw process increases the expression of these proteins in spermatozoon, triggering cellular defence (Neuer et al., Reference Neuer, Spandorfer, Giraldo, Dieterle, Rosenwaks and Witkin2000). In the study, it was determined that adding biotin (20 nM) and Ascorbic acid (4 mg) to the cryoprotectant agent modulated the mRNA expression levels of the Hsp40, Hsp60, and Hsp70 genes and reduced cryodamage. Especially in the ascorbic acid group, a significant decrease in the expression of Hsp40, Hsp60, and Hsp70 was observed (p < 0.001). These findings suggest that antioxidant supplements such as ascorbic acid and biotin may reduce oxidative stress by scavenging ROS and limiting lipid peroxidation, thereby decreasing the need for a heat shock protein response (Manikandan et al., Reference Manikandan, Beulaja, Thiagarajan and Arumugam2011). In addition, the observed decrease in Hsp40, Hsp60, and Hsp70 expression may reflect a reduced requirement for stress response activation, rather than a compromise in the cellular defence mechanism. A previous study reported that HSPs are activated by oxidative stress and help protect the cell from oxidative damage (Wang et al., Reference Wang, Jia, Li, Xie, Zhang and Qu2017). Ascorbic acid suppressed the expression of Hsp40 and Hsp60 more strongly than biotin (p < 0.01), while a significant decrease in Hsp70 expression was observed in both groups compared with the control group (p < 0.001). This suggests that Ascorbic acid reduces oxidative stress and lowers the need for a heat shock response. Wang et al. (Reference Wang, Jia, Li, Xie, Zhang and Qu2017) reported that Hsps are activated by oxidative stress and protect the cell from oxidative damage.
The fact that biotin did not significantly alter Hsp40 expression (p > 0.05) suggests that this compound has a more limited effect than Ascorbic acid in modulating the stress response. However, decreases in Hsp60 and Hsp70 expression (p < 0.05 and p < 0.001, respectively) indicate that biotin also indirectly reduces stress by supporting cellular metabolism. Consistent with previous reports, effective antioxidant protection may reduce the necessity for elevated Hsp synthesis, as cells experience a lower overall stress burden (Bahaloo et al., Reference Bahaloo, Rezvani, Yazd, Mehrjerdi, Davari, Roohbakhsh and Mehrparvar2020; Partyka et al., Reference Partyka, Bajzert and Niżański2022). Therefore, the observed downregulation of Hsp-related mRNA levels in the treated groups is interpreted as being associated with reduced cellular stress during cryopreservation. The effects of antioxidants on Hsp expression may differ depending on the agent used and the experimental conditions. Deng et al. (Reference Deng, Sun, Yu, Wang, Zhang, Zhang and Liu2017) reported that the addition of 0.1 mM melatonin to bull sperm increased Hsp70 and Hsp90 gene expression in frozen-thawed sperm.
In our study, the highest motility rate (17.50 ± 3.27; p < 0.05) was observed in the Ascorbic acid group, along with a decrease in Hsp70 expression. This finding is in line with previous studies reporting that Hsp70 level is associated with sperm function (Pardede et al., Reference Pardede, Kusumawati, Pangestu and Purwantara2023; Zhang et al., Reference Zhang, Hong, Yan, Wang, Li and Hu2015). Zhang et al. (Reference Zhang, Hong, Yan, Wang, Li and Hu2015) reported a positive correlation (0.327–0.785) between Hsp70 expression and sperm motility in bull sperm, demonstrating that the freezing-thawing process reduces Hsp70 expression. According to the study, Hsp70 expression was reported to be highest in fresh sperm and lowest after freezing-thawing. Similar studies have also shown that Hsps play significant roles in combating apoptosis and oxidative stress during the cryopreservation process of proteins (Ikwegbue et al. Reference Ikwegbue, Masamba, Oyinloye and Kappo2017; Kampinga et al. Reference Kampinga, Hageman, Vos, Kubota, Tanguay, Bruford and Hightower2009). This study shows that biotin and ascorbic acid supplementation suppress Hsp40, Hsp60, and Hsp70 gene expressions by reducing oxidative stress during the freeze–thaw process in rat sperm, thereby modulating the heat shock response. These effects suggest that both substances have the potential to mitigate cryodamage in rat sperm.
In conclusion, the addition of 4 mg/mL Ascorbic acid and 20 nM biotin to the diluent contributed to the preservation of cell functions in rat sperm cryopreservation, improving motility, viability, and membrane integrity after freezing and thawing. In addition, it was determined that both agents reduce cryodamage by regulating the expression of the Hsp40, Hsp60, and Hsp70 genes. These findings suggest that Ascorbic acid and biotin can be considered as potential cryoprotectant agents in rat sperm. A limitation of the present study is the absence of protein-level validation, such as Western blot or immunofluorescence analyses. Due to budgetary limitations, protein-level analyses could not be performed within the scope of the present study. Nevertheless, we acknowledge that mRNA expression data alone may not always directly reflect protein expression levels, and therefore, the findings should be interpreted with this consideration in mind.
Data availability
The data used in this study were generated by the author(s) and are available from the corresponding author upon reasonable request.
Acknowledgements
We would like to express our gratitude to all researchers for their valuable contributions.
Author contributions
O.K.Y., F.Y.D. and A.E.Ş. designed the study, performed the experiments, performed the analyses, and prepared the first drafts of the manuscript based on the data obtained. Then, they made final revisions on the manuscript. All authors equally contributed to the interpretation of the results.
Funding
This study did not receive any financial support.
Competing interests
None of the authors has any conflicts of interest to declare that are either directly or indirectly related to the study design, execution or reporting of this work.
Ethical standards
Ethical approval for this study was obtained by the Animal Experiments Local Ethics Committee (HADYEK) of Hatay Mustafa Kemal University, Hatay, Türkiye (Ethical Approval Number 2024/11-01).



 Open access
Open access