


Introduction
Disorders characterized by difficulties regulating mood, such as anxiety and depression, are highly prevalent and recurring (Gotlib et al., Reference Gotlib, Goodman and Humphreys2020; Kessler et al., Reference Kessler, Petukhova, Sampson, Zaslavsky and Wittchen2012; Scholten et al., Reference Scholten, Have, Geel, Balkom, Graaf and Batelaan2023). In fact, recent research indicates that mood difficulties have increased during and since the COVID-19 pandemic, with mothers being disproportionately affected (Racine et al., Reference Racine, Hetherington, McArthur, McDonald, Edwards, Tough and Madigan2021). This carries important implications not only for mothers’ own well-being but also for how they interact with their offspring (MacNeill et al., Reference MacNeill, Fu, Buss and Pérez-Edgar2022; Rao et al., Reference Rao, Barker, O’Farrelly and Ramchandani2021). Children exposed to more severe and frequent maternal mood difficulties are at increased risk for experiencing a range of negative developmental outcomes (Barker et al., Reference Barker, Jaffee, Uher and Maughan2011; Behrendt et al., Reference Behrendt, Wade, Bayet, Iii and Enlow2020; Gotlib et al., Reference Gotlib, Buthmann and Miller2023; M. M. Weissman et al., Reference Weissman, Berry, Warner, Gameroff, Skipper, Talati, Pilowsky and Wickramaratne2016). For example, maternal anxiety has been associated with increased risk for children’s social-emotional problems (Behrendt et al., Reference Behrendt, Wade, Bayet, Iii and Enlow2020), reduced cognitive and language development (Keim et al., Reference Keim, Daniels, Dole, Herring, Siega-Riz and Scheidt2011,) and more severe symptoms of psychopathology (Barker et al., Reference Barker, Jaffee, Uher and Maughan2011).
Consistent with the focus of developmental psychopathology on multiple levels of analysis (Cicchetti & Toth, Reference Cicchetti and Toth2009), recent research has examined maternal mood in the context of child neurodevelopmental outcomes, including alterations in brain structure (Donnici et al., Reference Donnici, Long, Dewey, Letourneau, Landman, Huo and Lebel2021; Gotlib et al., Reference Gotlib, Buthmann and Miller2023; Qiu et al., Reference Qiu, Rifkin-Graboi, Chen, Chong, Kwek, Gluckman, Fortier and Meaney2013; Rifkin-Graboi et al., Reference Rifkin-Graboi, Bai, Chen, Hameed, Sim, Tint, Leutscher-Broekman, Chong, Gluckman, Fortier, Meaney and Qiu2013, Reference Rifkin-Graboi, Meaney, Chen, Bai, Hameed, Tint, Broekman, Chong, Gluckman, Fortier and Qiu2015). In particular, potential associations of difficulties in maternal mood with children’s hippocampal and amygdala volume have garnered significant attention (Gotlib et al., Reference Gotlib, Buthmann and Miller2023; Qiu et al., Reference Qiu, Rifkin-Graboi, Chen, Chong, Kwek, Gluckman, Fortier and Meaney2013; Rifkin-Graboi et al., Reference Rifkin-Graboi, Bai, Chen, Hameed, Sim, Tint, Leutscher-Broekman, Chong, Gluckman, Fortier, Meaney and Qiu2013). These brain regions are often conceptualized as “stress sensitive,” with abundant glucocorticoid receptors that are important for regulating children’s response to environmental stressors (McEwen et al., Reference McEwen, Nasca and Gray2016). In fact, researchers have proposed that early adverse psychosocial experiences, including maternal mood difficulties, are biologically embedded in the structural development of these regions (Berens et al., Reference Berens, Jensen and Nelson2017). However, prior studies have yielded inconsistent findings regarding the nature of the relations between maternal mental health and children’s subcortical volumes. Some studies have reported that maternal mood difficulties are associated with reduced hippocampal or amygdala volume in offspring (Acosta et al., Reference Acosta, Tuulari, Scheinin, Hashempour, Rajasilta, Lavonius, Pelto, Saunavaara, Parkkola, Lähdesmäki, Karlsson and Karlsson2020; Chen et al., Reference Chen, Hamilton and Gotlib2010; Moog et al., Reference Moog, Nolvi, Kleih, Styner, Gilmore, Rasmussen, Heim, Entringer, Wadhwa and Buss2021), findings that align with other research indicating that exposure to adversity, broadly defined, is associated with reduced subcortical volumes, particularly the hippocampus (Calem et al., Reference Calem, Bromis, McGuire, Morgan and Kempton2017). Conversely, other studies have found larger amygdala or hippocampal volumes in offspring exposed to maternal mood difficulties (Donnici et al., Reference Donnici, Long, Dewey, Letourneau, Landman, Huo and Lebel2021; Lupien et al., Reference Lupien, Parent, Evans, Tremblay, Zelazo, Corbo, Pruessner and Séguin2011) or have not documented statistically significant associations with either region (Lupien et al., Reference Lupien, Parent, Evans, Tremblay, Zelazo, Corbo, Pruessner and Séguin2011; Wylie et al., Reference Wylie, Short, Fry, Mills-Koonce and Propper2024). It is also important to note that most of these studies have focused on maternal depression, especially during the perinatal period. Fewer studies have considered maternal anxiety during early childhood (Kemp et al., Reference Kemp, Bernier, Lebel and Kopala-Sibley2022), despite the fact that this is a period during which offspring may be particularly sensitive to the adverse effects of maternal anxiety (Gjerde et al., Reference Gjerde, Eilertsen, Eley, McAdams, Reichborn-Kjennerud, Røysamb and Ystrom2020). One significant challenge within the field is establishing how maternal mood dysfunction is best conceptualized to test associations with offspring neurodevelopment. Researchers have historically focused on average or total levels of symptom severity, frequency, or number of symptoms based on maternal responses to questionnaires. Although there are well-established relations between traditional assessments of maternal mood symptom levels and child adjustment, it is unclear whether summing symptom scores is the optimal approach for considering maternal mood in relation to offspring brain development. Assessing mood based on the sum symptom scores ignores information about variability in mood. In fact, researchers have proposed that intraindividual inconsistencies in caregiver mood states reflect unpredictable, fragmented signals that shape offspring brain development (Davis & Glynn, Reference Davis and Glynn2024; Glynn & Baram, Reference Glynn and Baram2019). This perspective is consistent with models that conceptualize unpredictability, or fluctuation in environmental conditions, as a core dimension of early life adversity, with neurodevelopment evolving to be sensitive to the statistical structure of the environment (Ellis et al., Reference Ellis, Sheridan, Belsky and McLaughlin2022). Interestingly, in rodent offspring, fragmented maternal signals have been linked to impoverished dendritic trees in the hippocampus (Brunson et al., Reference Brunson, Kramár, Lin, Chen, Colgin, Yanagihara, Lynch and Baram2005) and to alterations in hippocampus-dependent memory (Davis et al., Reference Davis, Stout, Molet, Vegetabile, Glynn, Sandman, Heins, Stern and Baram2017). Further, unpredictable maternal signals have been associated with altered expression of stress response genes in the amygdala of rodent offspring (Demaestri, Reference Demaestri2023) and with altered structural connectivity between the amygdala and prefrontal cortex in human offspring (Granger et al., Reference Granger, Glynn, Sandman, Small, Obenaus, Keator, Baram, Stern, Yassa and Davis2021). Together, these findings with rodent models provide a foundation for considering predictable versus unpredictable aspects of maternal mood and hippocampal and amygdala development in humans as a cross-species comparison.
Recently, researchers have used a novel approach to assess this aspect of maternal mood dysfunction, drawing on Shannon’s entropy, which measures the “randomness or unpredictability of the probability distribution of a random variable” (Howland et al., Reference Howland, Sandman, Davis, Stern, Phelan, Baram and Glynn2021, p. 490). Thus, mood entropy reflects the randomness or unpredictability of an individual’s responses to mood questionnaire items. For example, if a participant endorsed always feeling rested and never feeling nervous or restless on a questionnaire assessing anxiety, their item responses would be considered predictable and consistent and would yield a low entropy score. Conversely, a participant who completed the questionnaire at random would be considered highly unpredictable and, thus, obtain a high entropy score. Importantly, mood entropy based on questionnaire measures has shown to have convergent validity with variability in daily affect measured by ecological momentary assessments (Glynn et al., Reference Glynn, Howland, Sandman, Davis, Phelan, Baram and Stern2018; Ugarte, Reference Ugarte2023). Thus, entropy provides a sensitive index of intraindividual inconsistencies in mood states as assessed by questionnaire items. Although there are other approaches to quantifying unpredictability (Ellis et al., Reference Ellis, Sheridan, Belsky and McLaughlin2022; Ugarte & Hastings, Reference Ugarte and Hastings2023), entropy aligns with our guiding framework (Davis & Glynn, Reference Davis and Glynn2024) and our aim to assess unpredictability specifically in maternal mood (Glynn et al., Reference Glynn, Howland, Sandman, Davis, Phelan, Baram and Stern2018; Howland et al., Reference Howland, Sandman, Davis, Stern, Phelan, Baram and Glynn2021). Entropy also quantifies unpredictability in a manner consistent with information theory, rather than defining it simply as variability (within-person standard deviation) or as deviations from model predictions (residual approach), neither of which directly indexes the unpredictability of mood symptoms.
A small body of research suggests that maternal mood entropy has associations with child outcomes that are unique from maternal mood levels (Glynn et al., Reference Glynn, Howland, Sandman, Davis, Phelan, Baram and Stern2018; Howland et al., Reference Howland, Sandman, Davis, Stern, Phelan, Baram and Glynn2021; Jirsaraie et al., Reference Jirsaraie, Palma, Small, Sandman, Davis, Baram, Stern, Glynn and Yassa2024). For example, Glynn and colleagues (Reference Glynn, Howland, Sandman, Davis, Phelan, Baram and Stern2018) found that maternal mood entropy was a unique predictor of maternal reports of negative affectivity in infancy and childhood and of child and adolescent self-reports of anxiety and depressive symptoms. Only one prior study, however, has focused on maternal mood entropy and the neurodevelopment of offspring: Jisaraie and colleagues (Reference Jirsaraie, Palma, Small, Sandman, Davis, Baram, Stern, Glynn and Yassa2024) found that prenatal maternal mood entropy, but not mood symptom level, was associated with salience network integrity in children. Whether maternal mood entropy is associated with the structural development of children’s hippocampus and amygdala in a way that suggests biological embedding, and whether these associations are unique from maternal mood levels, is not yet known.
Another important limitation of the literature examining maternal mood and child neurodevelopment is that most studies have conducted a neuroimaging assessment at only a single point in time. However, developmental psychopathology frameworks posit that early risk factors, such as maternal mood difficulties, trigger developmental cascades that unfold over time and influence subsequent neurodevelopment (Masten & Cicchetti, Reference Masten and Cicchetti2010). Thus, longitudinal data with repeated neuroimaging assessments are essential for examining how the effects of early life experiences, such as exposure to maternal mood difficulties, unfold across childhood. Developmental psychologists have formulated models for testing two competing perspectives regarding how early experience shapes adaptation over time (Haltigan et al., Reference Haltigan, Roisman and Fraley2013; Magro et al., Reference Magro, Fraley and Roisman2020; Raby et al., Reference Raby, Roisman, Fraley and Simpson2015), but have not applied these models to longitudinal neuroimaging data. The revisionist perspective posits that early experiences are related to early developmental outcomes, but that their direct effects fade over time and are increasingly accounted for by the stability in the outcome (Fraley et al., Reference Fraley, Roisman and Haltigan2013; Fraley & Roisman, Reference Fraley and Roisman2015). Within this framework, this pattern is called a “transient” effect, referring specifically to the direct association fading over time. From this perspective, exposure to maternal mood difficulties in early childhood may have a transient direct association with brain structure in early childhood while still exerting indirect effects at later assessments via rank-order stability of brain development over time (i.e., consistently high, moderate, or low ranking relative to others). The formal model for testing this perspective would demonstrate that maternal mood difficulties are associated directly with children’s early brain structure but only indirectly with brain structure at subsequent assessments through stability in brain development. Thus, the revisionist model suggests that maternal mood plays a transient but potentially foundational role in early neurodevelopment. It is important to note that the term “transient” in this context does not imply that the effect is developmentally unimportant; it refers specifically to the fading over time of the direct association. Because transient direct effects may be carried forward indirectly through the stability of brain development, they may nonetheless have lasting implications.
In contrast, the enduring perspective suggests that some early experiences continue to directly shape adaptation across development, not only during early childhood. An example of the enduring model comes from the proposition of attachment theory that early relationships with caregivers organize early development and that this history continues to play an important role in shaping development over time (Sroufe et al., Reference Sroufe, Egeland and Kreutzer1990). From this perspective, early life interpersonal experiences are both foundational for early child development and serve as a direct scaffold for future development (Fraley et al., Reference Fraley, Roisman and Haltigan2013; Fraley & Roisman, Reference Fraley and Roisman2015). Thus, the enduring effects perspective would posit that early maternal mood difficulties have lasting and direct predictive significance for subcortical volumes across childhood (over and above the stability of these outcomes).
The current study had two aims. First, we examined whether maternal mood levels (sum of item responses) and entropy (unpredictability of item responses) are independently associated with children’s hippocampal and amygdala volumes. Given that risk factors have frequently been associated with smaller hippocampal volume in children (Humphreys et al., Reference Humphreys, King, Sacchet, Camacho, Colich, Ordaz, Ho and Gotlib2019; Miller et al., Reference Miller, Gluckman, Fortier, Chong, Meaney, Tan and Gotlib2024; D. G. Weissman et al., Reference Weissman, Lambert, Rodman, Peverill, Sheridan and McLaughlin2020a), we expected to observe smaller hippocampal volumes in the context of maternal mood difficulties. We did not have a priori hypotheses, however, regarding whether this association would be specific to maternal mood levels or entropy. Given inconsistent findings in the literature (Calem et al., Reference Calem, Bromis, McGuire, Morgan and Kempton2017), we also did not have specific hypotheses about whether maternal mood would be related to smaller or larger amygdala volume. Second, we explored whether early patterns of maternal mood have transient associations with offspring neurodevelopment in early childhood or enduring effects that extend into late childhood. We tested models of transient versus enduring effects that have been developed in other areas of research (Fraley et al., Reference Fraley, Roisman and Haltigan2013; Fraley & Roisman, Reference Fraley and Roisman2015; Haltigan et al., Reference Haltigan, Roisman and Fraley2013; Raby et al., Reference Raby, Roisman, Fraley and Simpson2015) but have yet to be applied to longitudinal neuroimaging data.
Methods
Participants and study design
Participants were drawn from the Growing Up in Singapore Towards Healthy Outcomes (GUSTO) birth cohort study. The original GUSTO study design, including details regarding procedures and study exclusion criteria, is described in Soh et al. (Reference Soh, Lee, Hoon, Tan, Goh, Lee, Shek, Teoh, Kwek, Saw, Godfrey, Chong, Gluckman and van Bever2012). Participants were included in our analyses if they provided data on maternal mood symptoms at the age 3 months, years 1, 2, 3, and 4.5 assessments, and/or usable T1-weighted neuroimaging scans at any of the age 4.5, 6, 7.5, or 10.5 year assessments (n = 1,425 total; n = 674 children with neuroimaging; 47.8% female, Chinese 54.8%, Malay 26.5%, and Indian 18.7%). All demographic data were provided by mothers at 26 weeks’ gestation. GUSTO was approved by the National Healthcare Group Domain Specific Review Board and the SingHealth Centralized Institutional Review Board in Singapore. Parents provided informed consent for participation and received compensation.
Measures
Maternal mood level and entropy
Mothers completed the State-Trait Anxiety Inventory (STAI; Spielberger, Reference Spielberger1983) at 3 months and 1, 2, 3, and 4.5 years after the child’s birth. The STAI measures current anxiety and “general” feelings of anxiety and has been used in prior studies comparing maternal mood levels and entropy (Howland et al., Reference Howland, Sandman, Davis, Stern, Phelan, Baram and Glynn2021). The STAI consists of 40 questions assessing severity and frequency of symptoms on a 1–4 scale. Following the scoring protocol for the STAI, 17 questions were reverse-scored (Bee Seok et al., Reference Bee Seok, Abd Hamid, Mutang and Ismail2018). To compute maternal mood level, item responses were summed, yielding a score between 40 and 160. The sum of responses to these 40 questions represents the level of maternal mood. These maternal mood level scores were averaged across assessment waves to create a composite score representing maternal mood level.
Maternal mood variability was calculated using Shannon’s entropy formula. Consistent with Glynn et al. (Reference Glynn, Howland, Sandman, Davis, Phelan, Baram and Stern2018), we applied Shannon’s entropy formula to STAI responses.
This yields probability distributions reflecting a given participant’s propensity to respond to STAI items in a consistent versus inconsistent manner, thus creating a measure of the unpredictability of mood states by entropic item responses. Entropy scores were calculated in R Studio (2023) utilizing code provided by Kelm, Reference Kelm2023, which can be found at https://contecenter.uci.edu/measuring-maternal-mood/. Mood entropy and mood level scores were auto-correlated across assessments (all rs > .29, ps < .001 for mood entropy, and all rs > .48, ps < .001 for mood levels). These correlations are presented in Supplemental Table S3. An overall maternal mood entropy score was calculated in the same manner as maternal mood level, averaging entropy scores across assessment waves.
Magnetic resonance imaging at ages 4.5, 6.0, 7.5, and 10.5 years
Participants completed magnetic resonance imaging (MRI) using a 3 T Siemens Skyra scanner at ages 4.5 and 6 years and using a 3 T Siemens Magnetom Prisma scanner at age 7.5 and 10.5 years. As reported in prior in prior analyses of GUSTO data (Miller et al., Reference Miller, Gluckman, Fortier, Chong, Meaney, Tan and Gotlib2024), high-resolution T1-weighted structural scans were collected using a Magnetization-Prepared Rapid Gradient-Echo (MP-RAGE) sequence with the following parameters: 192 slices, repetition time = 2000 ms, echo time = 2.08 ms, 1 mm isotropic voxels, and matrix size = 192 × 192. Data harmonization using longitudinal ComBat v.0.0.0.90 (Beer et al., Reference Beer, Tustison, Cook, Davatzikos, Sheline, Shinohara and Linn2020) was done with two batches – (1) ages 4.5 and 6 years and (2) age 7.5 years.
Segmentation
Segmentation was previously described in Miller et al. (Reference Miller, Gluckman, Fortier, Chong, Meaney, Tan and Gotlib2024). Briefly, MP-RAGE images underwent the standard FeeSurfer recon-all stream (v.7.0, http://surfer.nmr.mgh.harvard.edu). FreeSurfer outputs were inspected for registration between the structural images and atlas. Images showing poor registration were manually edited and reprocessed to ensure good correspondence with the atlas. Segmented hippocampal and amygdala masks were then used to estimate the volume measurements at each scan assessment. Quality control of scan data was performed at the raw MP-RAGE data and FreeSurfer recon-all output stages. For raw data, this involved a scoring system of pass (no or minimal visible motion lines, clear WM/GM distinction), good (visible motion lines that may be persistent with clear WM/GM distinction), questionable (visible motion lines that may be persistent with poor WM/GM distinction), or fail (ripples of motion lines that are prominent and persistent – integrity of WM/GM distinction are almost lost). Following our quality check, the number of scans that failed were 98 at age 4.5, 67 at age 6, 16 at age 7, and 3 at age 10.5. The numbers of children with 0, 1, 2, 3, and 4 scans are presented in Supplemental Table S2.
Data analysis
Brain volume data within each neuroimaging assessment timepoint were winsorized such that values higher and lower than the 95% and 5% quantile were replaced by those values, respectively. Bivariate correlations were conducted to assess the relations among maternal mood level, maternal mood entropy, and brain volumes. For hippocampal or amygdala volumes that showed significant associations with maternal mood levels or entropy, we then conducted a path analysis to formally investigate whether maternal mood variables showed transient associations specific to the region of interest in early childhood (i.e., significant direct effects at ages 4.5 and 6 years) or enduring associations with the region that continued across childhood (i.e., significant direct effects at all ages from 4.5 through 10.5 years). We tested separate models for the left and right hippocampus and amygdala volumes. After evaluating whether associations conformed more to a transient or enduring model, we tested whether the associations between maternal mood and brain volumes persisted after adjusting for covariates. Given that socioeconomic status is related to both maternal mental health and child neurodevelopment (Hair et al., Reference Hair, Hanson, Wolfe and Pollak2015; Masarik & Conger, Reference Masarik and Conger2017), covariates included a latent factor score for socioeconomic status based on maternal education level and household income, both of which were assessed during a prenatal assessment. In addition, covariate-adjusted models included biological sex assigned at birth, intracranial volume to isolate region-specific variability, and both maternal mood levels and mood entropy as predictors to test their unique effects. This approach is consistent with prior studies that examined maternal mood levels and entropy in the same model (Glynn et al., Reference Glynn, Howland, Sandman, Davis, Phelan, Baram and Stern2018; Howland et al., Reference Howland, Sandman, Davis, Stern, Phelan, Baram and Glynn2021). Within the adjusted models, these covariates were included for every path between maternal mood and brain volume across the four neuroimaging assessments. All statistical models were conducted using the lavaan package in R (2023). We conducted Little’s MCAR (missing completely at random) test to assess systematic missingness. The test failed to reject the null hypothesis (p = .163), indicating that the data are consistent with the assumption of MCAR. We used full information maximum likelihood (FIML) to handle missing data and to estimate model parameters. As a result, model parameters are informed by different numbers of participants depending on patterns of missingness across variables. To assess model fit, we used χ2, the Comparative Fit Index (CFI), and the root mean square error of approximation (RMSEA) with a 90% confidence interval (CI).
Results
Descriptive statistics and correlations
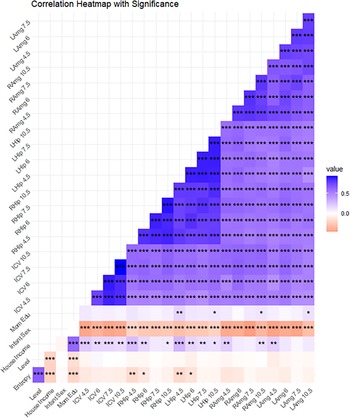
Table 1 presents descriptive statistics, and Figure 1 presents a plot of the zero-order correlations among our variables of interest (see Table S1 for descriptive statistics using non-winsorized data). Maternal mood level and entropy were positively correlated with each other (r = .66, p < .001, 95% CI = 0.624, 0.695). Maternal mood entropy was negatively correlated with left and right hippocampal volume at ages 4.5 and 6 (all rs > −.12, ps < .04), but not at ages 7.5 and 10.5 (all rs < −.07, all ps > .153). Maternal mood level was not significantly correlated with any brain volume metric (all rs < −.09, all ps > .185). All brain volume metrics were positively associated with each other across regions, hemispheres, and timepoints (all rs > .49, ps < .001). Each measurement of brain volume showed strong rank-order stability over time (all rs > .76, all ps < .001). Females had smaller brain volumes across regions and ICV compared to males (all rs > −.38, all ps < .001). Household income was significantly positively correlated with most measurements of brain volumes (all rs > .10, all ps < .044), except for right and left amygdala at ages 6 and 7.5, left amygdala at age 10.5, and right hippocampal volume at age 7.5 (all rs < .10, all ps > .075). Maternal education was significantly positively associated with left and right amygdala and left hippocampal volumes at 10.5 and left hippocampal volume at 4.5 (all rs > .10, all ps < .038), but not any other brain metric (all rs < .10, all ps > .066). A table of all correlations and their respective p-values is presented in Supplemental Table S4.
Correlations. Note. Squares in blue represent positive correlations, squares in red represent negative correlations. Stronger correlations are represented by darker squares. Mom Edu = maternal education; Sex coded as 0 = male and 1 = female. ***p <.001, **p <.01, *p <.05.

Figure 1 Long description
A heat map titled Correlation Heatmap with Significance. The heat map represents the correlation between various factors and brain regions, with significance levels indicated by stars. The heat map has a grid layout with rows and columns labeled with different factors and brain regions. The x-axis and y-axis both list factors such as LAmg 7.5, RAmg 6, LHip 10.5, RHip 4.5, ICV 6, Mom Edu, Infant Sex, House Income, Level, and Entropy. The color scale ranges from light pink to dark blue, indicating the strength of the correlation, with darker colors representing stronger correlations. Positive correlations are shown in blue, and negative correlations are shown in red. Stars indicate significance levels, with *** representing p <.001, ** representing p <.01, and * representing p <.05. Notable regions include clusters of dark blue squares along the diagonal, indicating strong positive correlations among certain brain regions. There are also areas with red squares, indicating negative correlations, particularly in the lower left corner. Overall, the heat map shows a mix of positive and negative correlations with varying levels of significance.
Descriptive statistics

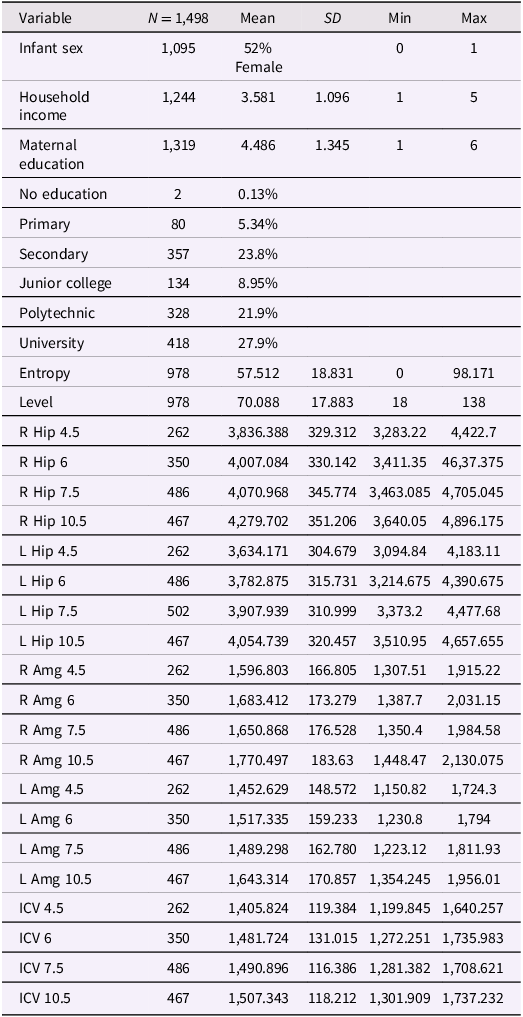
Table 1 Long description
A table with 1498 rows and 30 columns presenting descriptive statistics of various variables. The table includes columns for variable names, sample size (N), mean, standard deviation (SD), minimum (Min), and maximum (Max) values. The variables include infant sex, household income, maternal education, entropy, level, and various brain volume metrics measured at different ages and regions. Maternal education is further broken down into categories such as no education, primary, secondary, junior college, polytechnic, and university. The brain volume metrics include measurements of the right and left hippocampus, right and left amygdala, and intracranial volume (ICV) at ages 4.5, 6, 7.5, and 10.5. Each row provides specific values for these metrics.
Note. All brain metric units are in mm3; ICV = intracranial volume (scaled by dividing ICV values by 1,000); Hip = hippocampus; Amg = amygdala; Entropy = mood entropy; Level = mood level; R = right; L = left. Household Income reported in Singapore dollars per month: $0–$999, $1,000–$1999, $2000–$3999, $4000–$5999, more than $6000. For Maternal Education, 1 = No education, 2 = Primary, 3 = Secondary, 4 = Junior college or Institute of Technical Education, 5 = Polytechnic, 6 = University (Bachelor’s, Graduate degree).
Path analysis models of maternal mood entropy and hippocampal volume
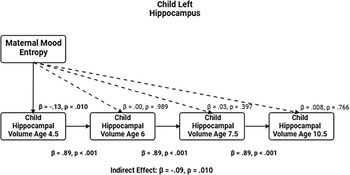
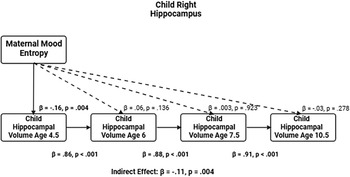
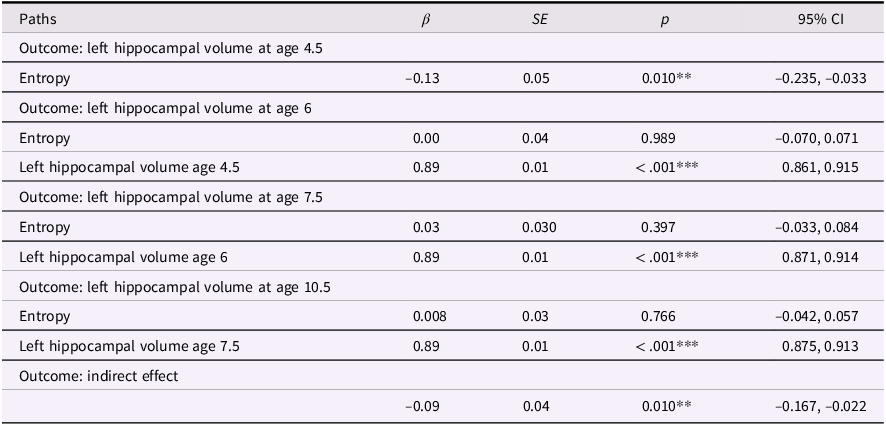
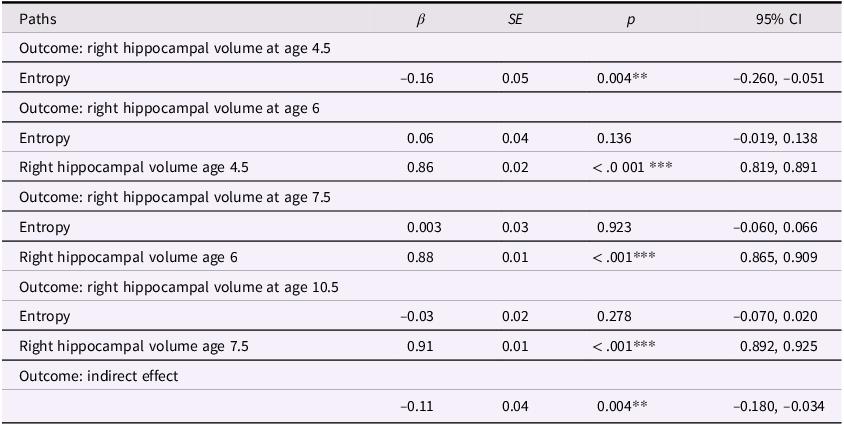
The correlations showed that maternal mood entropy, but not level, was negatively associated with hippocampal volumes at ages 4.5 and 6. Therefore, we conducted path analysis models focused on maternal mood entropy as a predictor of hippocampal volumes, controlling for rank-order stability over time, to examine the degree to which this association was transient versus enduring. The left and right hippocampal volume models (not adjusted for covariates) are presented in Figures 2 and 3, respectively (see Tables 2 and 3 for parameter estimates).
Entropy and left hippocampal volume model. Note. Solid lines are paths significant at p <.05.

Entropy and right hippocampal volume model. Note. Solid lines are paths significant at p <.05.

Entropy and left hippocampal volume model
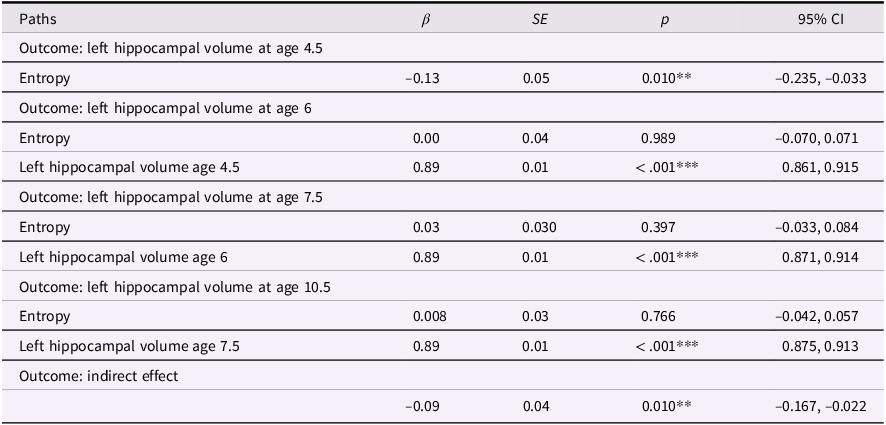
Table 2 Long description
The table presents data on the relationship between entropy and left hippocampal volume at different ages. It has five rows and five columns. The columns are labeled Paths, Beta, SE, p, and 95% CI. The rows are grouped by different outcomes: left hippocampal volume at ages 4.5, 6, 7.5, 10.5, and indirect effect. Each row provides specific values for these columns. For example, the first row under the outcome of left hippocampal volume at age 4.5 shows a beta value of -0.13, a standard error of 0.05, a p-value of 0.010, and a 95% confidence interval of -0.235 to -0.033. The table captures the statistical relationships and significance levels of these variables.
Note. ***p < .001, **p < .01, *p < .05. β = standardized beta; SE = standard error; CI = confidence interval.
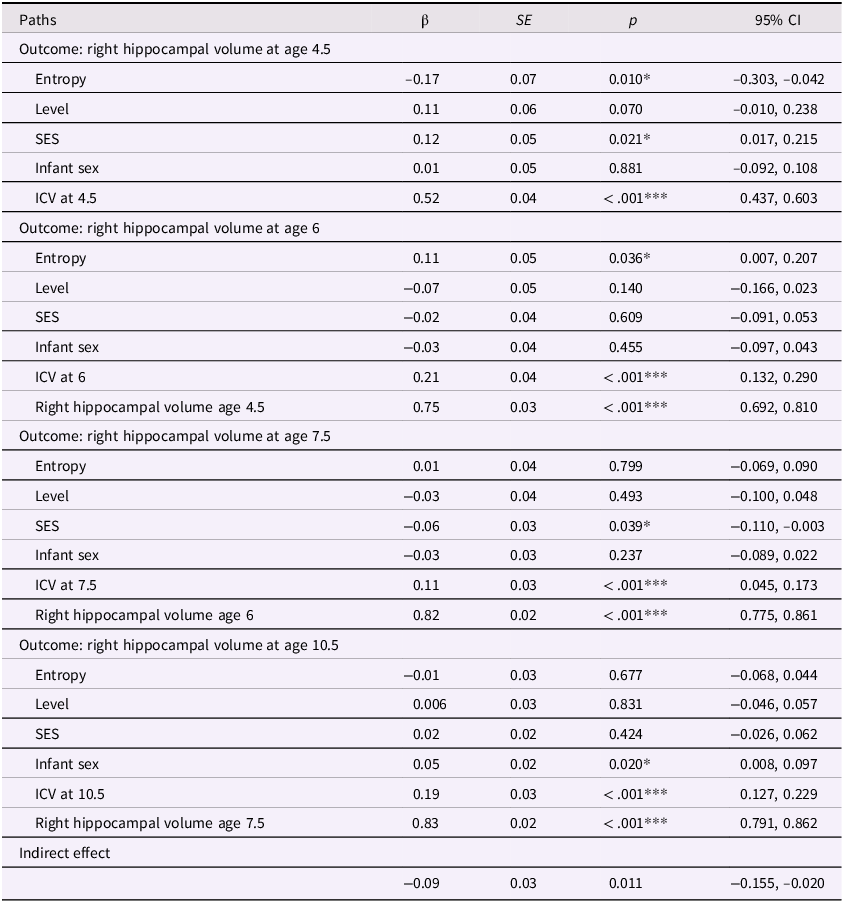
Entropy and right hippocampal volume model
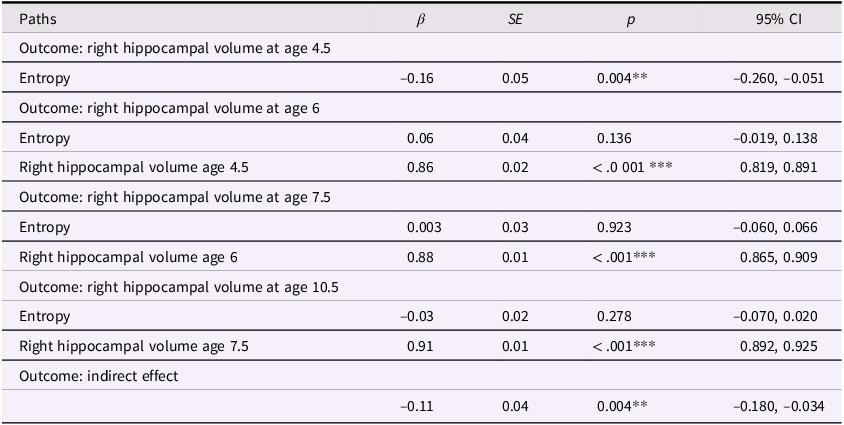
Table 3 Long description
A table with five rows and five columns. The columns are labeled Paths, beta, SE, p, and 95% CI. The table presents data on the correlation between maternal mood entropy and right hippocampal volume at ages 4.5, 6, 7.5, and 10.5. Row 1: Outcome: right hippocampal volume at age 4.5, Path: Entropy, beta: -0.16, SE: 0.05, p: 0.004**, 95% CI: -0.260, -0.051. Row 2: Outcome: right hippocampal volume at age 6, Path: Entropy, beta: 0.06, SE: 0.04, p: 0.136, 95% CI: -0.019, 0.138. Row 3: Outcome: right hippocampal volume at age 6, Path: Right hippocampal volume age 4.5, beta: 0.86, SE: 0.02, p: <.001***, 95% CI: 0.819, 0.891. Row 4: Outcome: right hippocampal volume at age 7.5, Path: Entropy, beta: 0.003, SE: 0.03, p: 0.923, 95% CI: -0.060, 0.066. Row 5: Outcome: right hippocampal volume at age 7.5, Path: Right hippocampal volume age 6, beta: 0.88, SE: 0.01, p: <.001***, 95% CI: 0.865, 0.909. Row 6: Outcome: right hippocampal volume at age 10.5, Path: Entropy, beta: -0.03, SE: 0.02, p: 0.278, 95% CI: -0.070, 0.020. Row 7: Outcome: right hippocampal volume at age 10.5, Path: Right hippocampal volume age 7.5, beta: 0.91, SE: 0.01, p: <.001***, 95% CI: 0.892, 0.925. Row 8: Outcome: indirect effect, Path: , beta: -0.11, SE: 0.04, p: 0.004**, 95% CI: -0.180, -0.034.
Note. ***p < .001, **p < .01, *p < .05. β = standardized beta; SE = standard error; CI = confidence interval.
These models showed acceptable fit to the data (left hippocampus model: χ2(13) = 174.134, p < .001; CFI = .924; RMSEA = .094, CI [0.082, 0.106]; right hippocampus model: χ2(13) = 140.861, p < .001; CFI = .938; RMSEA = .083, CI [0.071, 0.096]). Our results were more consistent with a transient effect than enduring effect model. Specifically, maternal mood entropy was negatively associated with child left and right hippocampal volume at age 4.5 years (β = −.13, p = .010 and β = −.16, p = .004); however, after controlling for rank-order stability of hippocampal volume over time, maternal mood entropy was no longer associated with left or right hippocampal volume at age 6 years (both βs < .07, ps > .136). Maternal mood entropy was also not associated with hippocampal volumes at 7.5 and 10.5 years (all βs < .04, ps > .278). Additionally, maternal mood entropy was indirectly associated with reduced hippocampal volume at 10.5 years through rank-order stability of hippocampal volume over time (serial indirect β = −.09, p = .010, and serial indirect β = −.11, p = .004 for left and right hippocampal volumes, respectively).
Path analysis with covariates
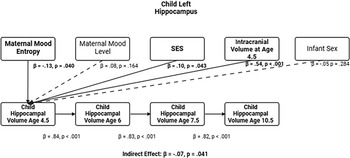
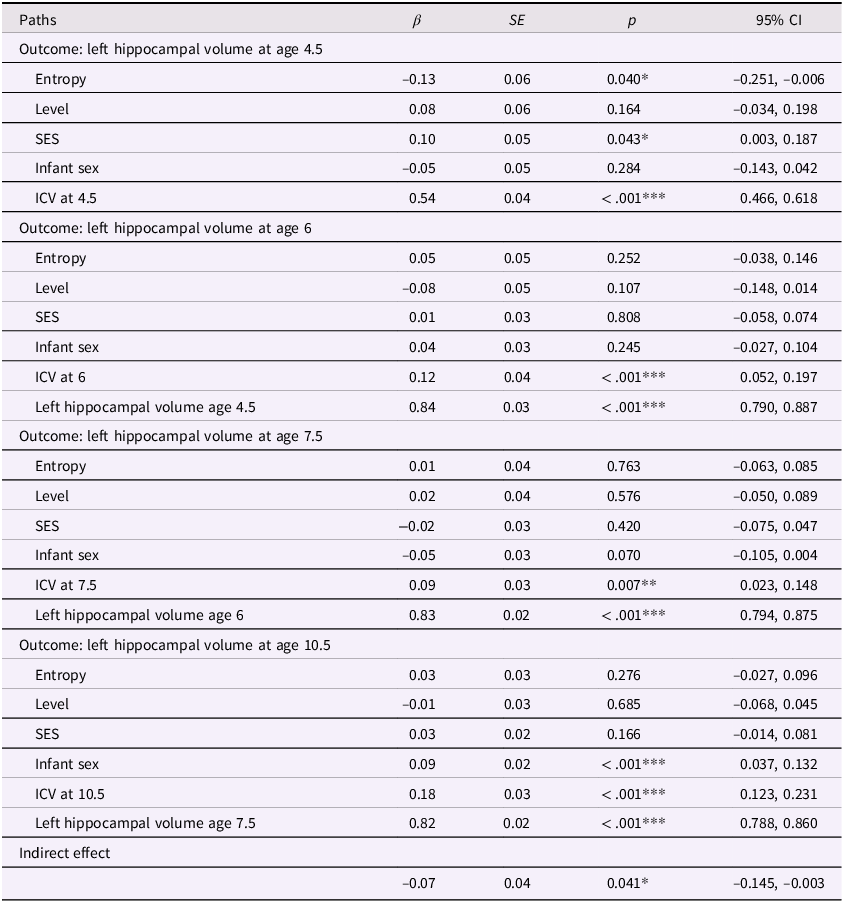
In our previous models, we determined that maternal mood entropy was specifically associated with child hippocampal volume. After adjusting for covariates (see Figures 4 and 5 and Tables 4 and 5), the direct associations of maternal mood entropy with hippocampal volumes at age 4.5 remained statistically significant (both ps < .040 for left and right hippocampal volume). These associations were present over and above the effects of socioeconomic status (both βs < .12, ps < .021) and intracranial volume (both βs < .54, ps < .001) and were specific to maternal mood entropy (not levels). The indirect effect of maternal mood entropy on left and right hippocampal volume at 10.5 was also still significant in a covariate-adjusted model.
Entropy and left hippocampal model after adjusting for covariates. Note. Solid lines are paths significant at p <.05. Maternal mood level, SES, ICV, and sex were included as covariates at all time points but are omitted from the plot for clarity. Further statistical details regarding these results are presented in Table 4.

Entropy and right hippocampal model after adjusting for covariates. Note. Solid lines are paths significant at p <.05. Maternal mood level, SES, ICV, and sex were included as covariates at all time points but are omitted from the plot for clarity. Further statistical details regarding these results are presented in Table 5.

Entropy and left hippocampal volume model with covariates
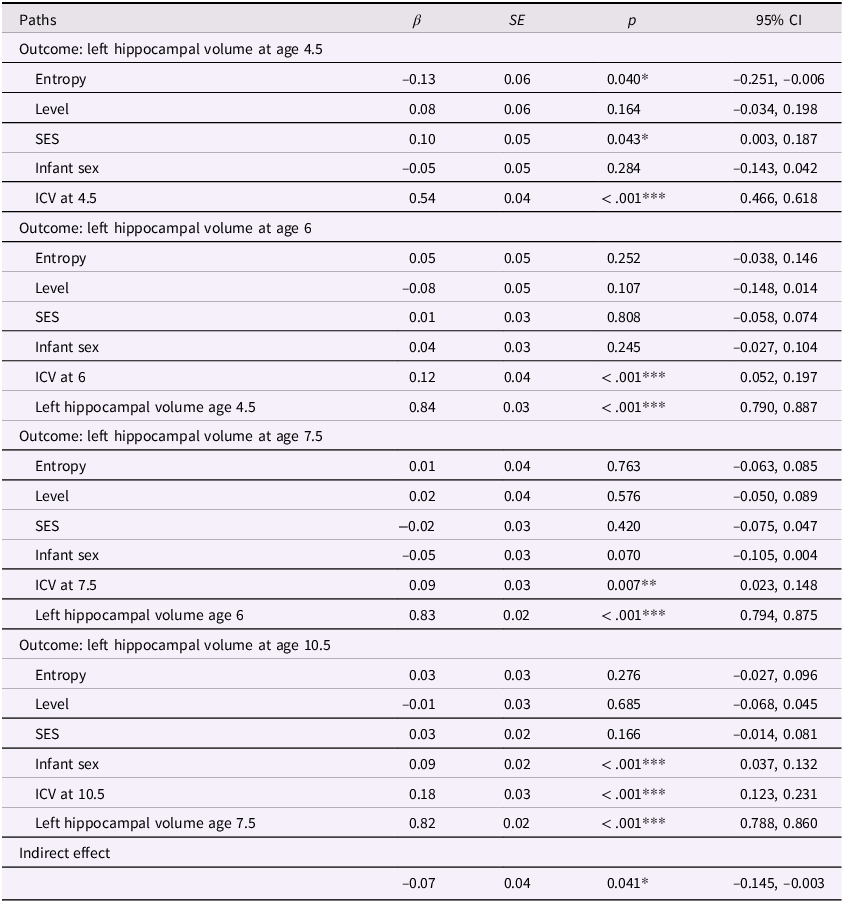
Table 4 Long description
The table presents data on the relationship between various factors and left hippocampal volume at ages 4.5, 6, 7.5, and 10.5. It includes columns for paths, beta coefficients, standard errors, p-values, and 95% confidence intervals. The table is divided into four sections, each corresponding to a different age outcome. Each section lists the factors: Entropy, Level, SES, Infant sex, ICV at the respective age, and previous hippocampal volume measurements. For example, the first section shows the impact of these factors on left hippocampal volume at age 4.5, with Entropy having a beta coefficient of -0.13 and a p-value of 0.040. The subsequent sections follow a similar format, detailing the impact of the same factors on hippocampal volume at ages 6, 7.5, and 10.5. Notable trends include significant p-values for ICV at each age and the previous hippocampal volume measurements having high beta coefficients and significant p-values.
Note. ***p < .001, **p < .01, *p < .05. β = standardized beta; SE = standard error; CI = confidence interval; ICV = intracranial volume (scaled by dividing ICV values by 1,000); Sex coded as 0 = male and 1 = female.
Entropy and right hippocampal model with covariates
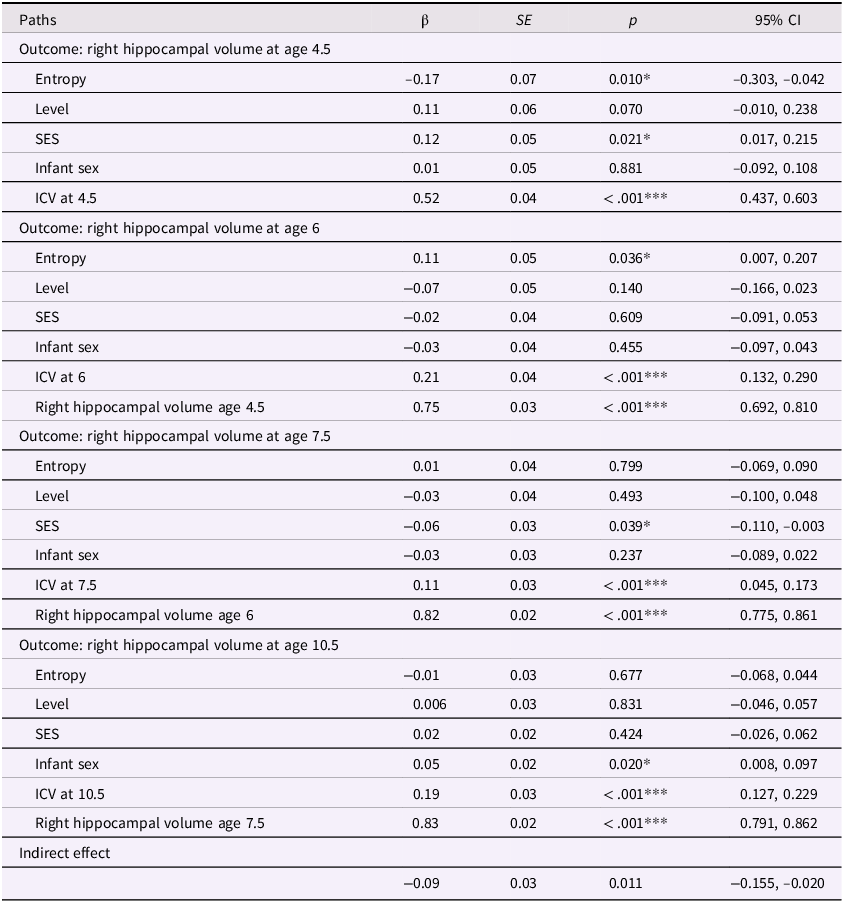
Table 5 Long description
The table presents data on the relationship between various factors and right hippocampal volume at ages 4.5, 6, 7.5, and 10.5. It includes columns for paths, beta coefficients, standard errors, p-values, and 95% confidence intervals. The table is divided into four sections, each corresponding to a different age outcome. Each section lists the factors: Entropy, Level, SES, Infant sex, ICV at the respective age, and previous right hippocampal volume where applicable. For example, the first section shows the impact of these factors on right hippocampal volume at age 4.5, with Entropy having a beta coefficient of -0.17 and a p-value of 0.010. The second section shows the impact on right hippocampal volume at age 6, with Entropy having a beta coefficient of 0.11 and a p-value of 0.036. The third section shows the impact on right hippocampal volume at age 7.5, with Entropy having a beta coefficient of 0.01 and a p-value of 0.799. The fourth section shows the impact on right hippocampal volume at age 10.5, with Entropy having a beta coefficient of -0.01 and a p-value of 0.677. The table also includes an indirect effect row with a beta coefficient of -0.09 and a p-value of 0.011.
Note. ***p < .001, **p < .01, *p < .05. β = standardized beta; SE = standard error; CI = confidence interval; ICV = intracranial volume (scaled by dividing ICV values by 1,000); Sex coded as 0 = male and 1 = female.
In the covariate-adjusted model for the right hippocampus, maternal mood entropy was significantly and positively associated with right hippocampal volume at age 6 (β=.11, p=.036). Given that the zero-order correlations between maternal mood entropy and right hippocampal volume at ages 4.5 and 6 were negative and significant, it is possible that the positive association at age 6 in the covariate-adjusted model is a suppression effect; therefore, we did not interpret this effect. See Tables S5 and S6 for results using non-winsorized data.
Amygdala model
In models controlling for rank-order stability (i.e., autocorrelation) and covariates, maternal mood entropy was not significantly associated with amygdala volume at any timepoint for either hemisphere (all absolute βs < .10, ps < .110).
Testing a latent factor model of maternal mood dysregulation
The significant correlation between maternal mood entropy and mood levels raises the possibility that these measures are manifestations of a broader construct of mood dysregulation that may be implicated in children’s neurodevelopment. Therefore, we tested whether a latent factor based on mood entropy and mood levels was associated with children’s hippocampal and amygdala volumes. We did not find that this latent factor was significantly associated with left or right volumes at any time point (all βs < 0.05, ps > .332). Further details on these results for the hippocampus are presented in the supplemental materials, Tables S7 and S8.
Testing sex differences
Prior work suggests that there are sex-specific subcortical alterations in offspring exposed to difficulties in maternal mood (Wen et al., Reference Wen, Poh, Ni, Chong, Chen, Kwek, Shek, Gluckman, Fortier, Meaney and Qiu2017). To examine whether similar patterns of findings are found in the context of maternal mood entropy, we tested whether the effects of maternal mood entropy (and mood level) on subcortical volumes differed by sex. Specifically, we added interaction terms of sex and maternal mood entropy, as well as sex and maternal mood level, to our path analysis models. One model yielded significant sex moderation for the association of maternal mood entropy with right hippocampal volume (β = −.07, p = .045) at age 10.5 years. To probe these effects, we reran our models separately in males and females. For both right hippocampal and left amygdala volume at age 10.5, the direct effect of maternal mood entropy was not statistically significant for either females or males (all βs < −.11, ps > .115). Thus, although the associations of maternal mood entropy with right hippocampal and left amygdala volumes at 10.5 differed significantly for males and females, they did not differ from zero for either sex. Consequently, we do not interpret these interactions further. We present additional details about these results in Supplemental Tables S9 and S10. We also provide in Figure S1 a visualization of average changes in hippocampal and amygdala volumes across time.
Further sensitivity analyses
Analyses incorporating a measure of maternal depression (Beck Depression Inventory II; BDI-II) did not yield significant associations with offspring neurodevelopment. We considered BDI-based measures of entropy and levels and an aggregate measures of BDI with STAI. Maternal mood entropy and mood levels based on BDI were not associated with hippocampal volumes (all βs < .13 ps > .128), with the exception of entropy predicting left hippocampal volume at age 10.5 (β = .12, p = .008). Although an aggregate measure of BDI entropy and STAI entropy had small but significant zero-order correlations with both left and right hippocampal volumes at age 4.5 (both rs > −.14, both ps < .023), these associations were no longer statistically significant after adjusting for covariates (both rs < −.05, both ps > .466).
We also conducted analyses testing our covariate-adjusted models but limited to the subset of participants who had at least one neuroimaging timepoint (n = 674). These analyses yielded the same pattern of findings as models based on the full sample. Specifically, mood entropy, but not mood level, was associated with right hippocampal volume at age 4.5 (β = −.16, p = .010), and this association was carried forward indirectly to right hippocampal volume at age 10.5 via the rank-order stability of hippocampal volume over time (β = −.08, p = .011). Our left hippocampal model showed the same pattern of results. Model estimates are presented in Supplemental Tables S11 and S12.
Discussion
This study had two primary aims. First, to determine whether early maternal mood symptoms are implicated in offspring hippocampal and amygdala volume development, testing whether these associations were specific to maternal mood symptom levels or entropy. Second, we explored two competing perspectives regarding how the effects of early experiences on subcortical development unfold across time. Our results demonstrate that early patterns of maternal mood dysregulation are implicated in early childhood brain structure, laying the foundation for subsequent neurodevelopment. Early maternal mood entropy, which may reflect intraindividual patterns of mood instability or variability, is implicated in the underdevelopment of offspring’s hippocampi that predisposes them to continue showing this pattern in the future. Our findings both contribute to a developing body of research that entropy is an independent and meaningful component of mood dysregulation (Glynn et al., Reference Glynn, Howland, Sandman, Davis, Phelan, Baram and Stern2018; Howland et al., Reference Howland, Sandman, Davis, Stern, Phelan, Baram and Glynn2021; Jirsaraie et al., Reference Jirsaraie, Palma, Small, Sandman, Davis, Baram, Stern, Glynn and Yassa2024) and suggest a specific developmental process that comprises the changing association between early maternal mood entropy and children’s hippocampal volume across childhood. Consistent with prior research on maternal mood entropy in relation to other child outcomes, our results indicate that maternal mood entropy is uniquely implicated in child hippocampal volume over and above mood level. Our prediction that maternal mood abnormalities are negatively associated with hippocampal development was also confirmed. We did not find effects of maternal mood entropy or levels on children’s amygdala volume, which fits with the inconsistencies reported in previous studies of early life adversity more broadly (Calem et al., Reference Calem, Bromis, McGuire, Morgan and Kempton2017). Lastly, we found inconsistent evidence for sex differences within our models. Maternal mood entropy only significantly interacted with sex to directly predict hippocampal volume at age 10.5. However, the effects of mood entropy were not significantly different from zero for either males or females.
Our finding that maternal mood entropy was uniquely associated with child hippocampal development highlights the importance of considering intraindividual inconsistencies in mood in understanding associations between maternal functioning and child neurodevelopment. Research examining alterations in maternal mood in relation to offspring neurodevelopment may benefit from focusing on entropy, which is not captured by traditional approaches that focus on frequency and severity of mood difficulties. Our findings are consistent with the perspective that unpredictable maternal signals shape offspring neurodevelopment (Davis & Glynn, Reference Davis and Glynn2024) and with findings from recent studies with rodents suggesting that offspring hippocampal structure is sensitive to these signals (Davis et al., Reference Davis, Stout, Molet, Vegetabile, Glynn, Sandman, Heins, Stern and Baram2017). Further, our findings support broader models that frame environmental unpredictability as an evolutionarily meaningful dimension of early life adversity (Ellis et al., Reference Ellis, Sheridan, Belsky and McLaughlin2022). The current study provides the first evidence in humans that early patterns of maternal mood entropy are associated with altered hippocampal volume and development in offspring. Importantly, reduced hippocampal volume has been observed in the context of other early life adversities (Humphreys et al., Reference Humphreys, King, Sacchet, Camacho, Colich, Ordaz, Ho and Gotlib2019; Miller et al., Reference Miller, Gluckman, Fortier, Chong, Meaney, Tan and Gotlib2024) and has been associated with children’s heightened stress sensitivity and psychopathology (Durham et al., Reference Durham, Jeong, Moore, Dupont, Cardenas-Iniguez, Cui, Stone, Berman, Lahey and Kaczkurkin2021; D. G. Weissman et al., Reference Weissman, Lambert, Rodman, Peverill, Sheridan and McLaughlin2020b). These relations, in combination with the current findings, suggest that reduced hippocampal volumes are one neurodevelopmental mechanism by which exposure to maternal mood entropy increases children’s risk for mental health difficulties.
Our findings also contribute to our understanding of the legacy of early childhood experiences for neurodevelopment across childhood. We used a developmental modeling approach, with repeated neuroimaging observations, to formally test whether the association between maternal mood patterns and offspring neurodevelopment changes over time. A key statistical distinction between enduring and revisionist models is whether early life experiences are associated directly with future development (i.e., Enduring) or indirectly through the stability of development over time (i.e., Revisionist) (Fraley et al., Reference Fraley, Roisman and Haltigan2013; Fraley & Roisman, Reference Fraley and Roisman2015). Our findings provide support for a revisionist model of biological embedding. That is, exposure to early maternal mood entropy appears to have a significant but transient direct association with reduced hippocampal volume in early childhood. Although maternal mood entropy did not have enduring direct predictive significance, an indirect effect was carried forward through rank-order stability of hippocampal volumes, leading to underdevelopment of the hippocampus in late childhood. Taken together, maternal mood entropy serves as a foundational experience that is directly associated with early child hippocampal volume (age 4.5) and continues indirectly throughout childhood (out to age 10.5). Many researchers endorse the biological embedding perspective, which posits that early life experiences organize neurobiological structure and function (Berens et al., Reference Berens, Jensen and Nelson2017; Shonkoff, Reference Shonkoff2012). It is important to note, however, that most studies have used cross-sectional designs and retrospective reporting of early life experiences or have focused on associations with future neural outcomes assessed at a single timepoint. Fewer studies have utilized longitudinal modeling to characterize biological embedding of early life experience as a developmental process that shapes neurodevelopment across time. Early maternal mood entropy appears to predispose children to develop smaller hippocampal volumes, which leads to reduced volumes in the future, a finding that would have gone unnoticed using an approach that focused on a prospective association with future hippocampal volumes at a single timepoint. We believe that our work serves as a call for more longitudinal neuroimaging data given that these study designs are necessary for modeling developmental processes (Roisman & Fraley, Reference Roisman and Fraley2013).
Although it was not an aim of our study, we also found that family SES was positively associated with hippocampal volume in early childhood, suggesting that higher SES serves as a compensatory resilience factor that offsets the effects of maternal mood entropy on hippocampal development. Conversely, lower SES and higher maternal mood entropy appear to have additive effects on children’s reduced hippocampal volume. Collectively, these findings are consistent with prior research suggesting that family SES is related to variability in neuroanatomy (Colich et al., Reference Colich, Rosen, Williams and McLaughlin2020; Hair et al., Reference Hair, Hanson, Wolfe and Pollak2015; Miller et al., Reference Miller, López, Buthmann, Garcia and Gotlib2022) and promotes resilient developmental outcomes in the context of various risk factors (Fergus & Zimmerman, Reference Fergus and Zimmerman2005).
It is also worth noting that our findings were specific to maternal anxiety; using a measure of maternal depression (BDI-II) in our analyses did not replicate our findings. It is unclear whether this is due to differences between the constructs of anxiety and depression and/or to differences between the STAI and BDI-II. For example, there was more variability in responses to the STAI than to the BDI-II, which may be related to the fact that the BDI-II items include relatively extreme and less prevalent symptoms (e.g., suicidal ideation). As a result, participants may have been less likely to use the full range of response options on the BDI-II items. The BDI-II is also shorter than the STAI, potentially further limiting opportunities to show unpredictable responding. These features of the BDI-II and/or depression as a construct may have constrained our ability to uncover associations with offspring neurodevelopment. Although prior studies of mood entropy included the STAI and BDI, they combined these measures with other questionnaires (Glynn et al., Reference Glynn, Howland, Sandman, Davis, Phelan, Baram and Stern2018; Howland et al., Reference Howland, Sandman, Davis, Stern, Phelan, Baram and Glynn2021). Further research is needed to determine if integrating multiple measures of symptoms of psychopathology yields a more robust index of entropy or whether associations with entropy are driven by specific measures.
We should note four limitations of this study. First, GUSTO is a birth cohort study in Singapore, and the generalizability of the current findings to other populations and sociocultural contexts is unclear. Second, our earliest measurement of child neuroimaging data was at age 4.5 years. In our longitudinal models, we treated the hippocampal and amygdala volume data at this timepoint differently than we did at subsequent assessments, which included paths to control for rank-order stability. It is possible that had we been able to model hippocampal volume at age 4.5 years as being predicted by volumes at earlier ages, we would not have observed an association with maternal mood entropy at this timepoint. In this context, however, it is important to note that maternal mood entropy was associated specifically with hippocampal volume at age 4.5 (and age 6) in our zero-order correlation analyses, which is consistent with the path analyses. Nonetheless, it is possible that maternal mood entropy is directly related to offspring structural neurodevelopment at even earlier ages. Third, there are alternative methods for quantifying maternal mood, each of which captures a different aspect of unpredictability. Our findings are specific to mood entropy, and operationalizing unpredictability in other ways may yield different results. Future research should examine and compare systematically the strengths and limitations of various quantitative approaches, as well as the operationalization of unpredictability reflected by each approach. Finally, our models did not include potential mechanisms linking maternal mood entropy to hippocampal volumes. Although it is not clear precisely how exposure to mood entropy might be related to reduced hippocampal volume, theoretical and empirical work points to fragmented sensory signals as one possibility (Davis et al., Reference Davis, Stout, Molet, Vegetabile, Glynn, Sandman, Heins, Stern and Baram2017; Davis & Glynn, Reference Davis and Glynn2024). Fluctuations in maternal emotions and moods, corresponding to changes in emotion-specific behaviors (Keltner & Gross, Reference Keltner and Gross1999), may lead to increased inconsistencies in auditory, tactile, and visual input to the child from the caregiver which, in turn, may be a more proximal factor influencing maturation of the developing brain.
Despite these limitations, our data show that maternal mood entropy is uniquely associated with reduced hippocampal volume in early childhood, which sets the stage for continued structural underdevelopment of the hippocampus across childhood. This study is the first to link early maternal mood entropy to structural development of the hippocampus in humans. It will be important for future longitudinal neuroimaging research to consider other measures of maternal mood and caregiving unpredictability in order to build on our findings and advance our understanding of the biological embedding of early life experiences in neurodevelopment.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S0954579426101291.
Data availability statement
The data for this paper are derived from the GUSTO longitudinal birth cohort study. Restrictions apply to data availability. However, the data can be requested at (https://gustodatavault.sg/), which will require a project description and scientific reasoning.
Acknowledgements
We thank the GUSTO team and all the families who participated in this study.
Funding statement
This work was supported by the Singapore National Research Foundation (NMRC) under the Open Fund Large Collaborative Grant administered by the Singapore Ministry of Health’s National Medical Research Council. Additional funding was provided by the Agency for Science, Technology and Research (A*STAR) Strategic Research Program Brain-Body Initiative, the Hope for Depression Research Foundation (M.J.M.), the NMRC Transition Award (MOH-001273-00; A.P.T.), and the National Institute of Mental Health (grant R37MH101495; I.H.G.).
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Pre-registration statement
These analyses were largely exploratory and were not pre-registered.
AI statement
ChatGPT was used to assist with troubleshooting and developing R code for analyses and to edit grammar and punctuation while drafting the manuscript.




 Open access
Open access