


Introduction
Nutrition is important for human health as it provides metabolites essential to a healthy life, with roles in, e.g., energy creation, signalling pathways or providing metabolites that cannot be synthesised in the human body(Reference Chen, Michalak and Agellon1). Dietary habits can cause an imbalance of key metabolites that are linked to diseases, such as anaemia and coagulation disorders(Reference Kiani, Dhuli and Donato2). Additionally, over- or under consumption of groups of metabolites or macronutrients, such as fibre or saturated fatty acids, are linked to non-communicable diseases, such as cancer, type 2 diabetes and chronic inflammation(Reference Adolph and Tilg3–Reference Gropper5). Conversely, nutrition has been used to treat, alleviate and prevent the same diseases that it can cause, showing the duality of nutrition in human health(Reference Kimokoti and Millen6,Reference Brandhorst and Longo7) .
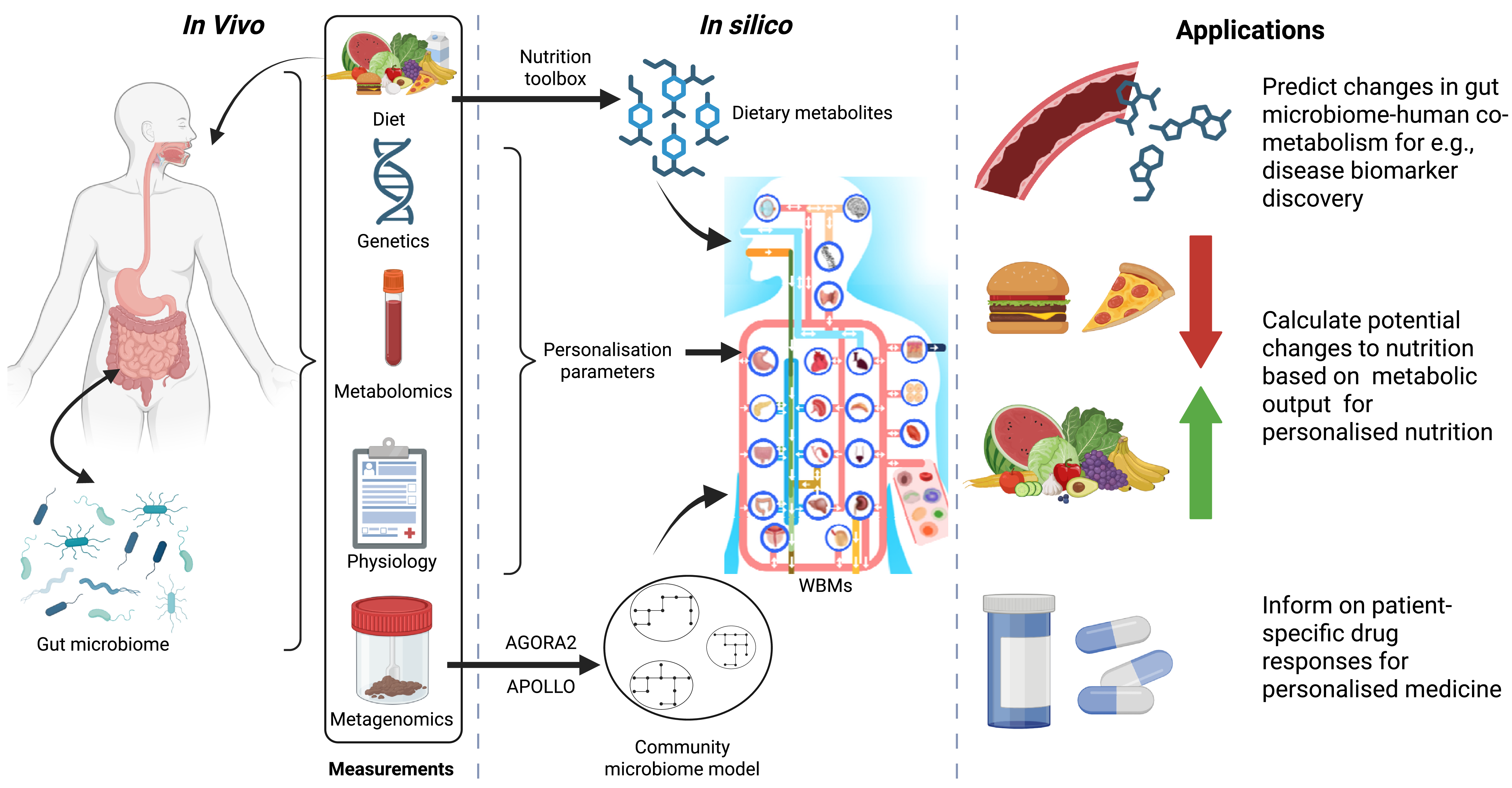
Investigating the links between nutrition and human health is usually done through analysing associations between various -omics data (e.g., metagenomics, genomics or metabolomics) and dietary questionnaires (e.g., food dairies or food frequency questionnaires)(Reference Norheim, Gjelstad and Hjorth8,Reference Ulaszewska, Weinert and Trimigno9) (Fig. 1). However, understanding these associations can be difficult as the human body is a complex system. To unravel this complexity, the effects of nutrition on human health can be investigated by studying how the nutrient composition of the diet interact with human metabolism.
Schematic overview of factors that can influence human metabolism, which can be measured and incorporated into the whole-body models (WBMs) and how the WBMs can be used in the context of human health. The figure was generated with Biorender.

Fig. 1. Long description
A schematic of the human body, in the box in vivo which shows the gut. microbiome. There is a box called measurements that include diet, genetics, metabolomics, physiology and metagenomics. Diet is pointed to the human schematic. Next to the in vivo box, there is a box called in silico. Here is a schematic overview of the whole-body metabolic models. An arrow points from the diet to dietary metabolites with the text nutrition toolbox. Dietary metabolites then point to the schematic of the WBM. Genetics, metabolomics and physiology are summed into personalisation parameters and also point towards the WBM schematic. Finally the metagenomics point to a schematic of the community microbiome models, with AGORA2 and APOLLO written alongside. The community microbiome model also points towards the WBM schematic. The last box is called application and had 3 text boxes. The first text box reads "Predict changes in gut microbiome-human co-metabolism for e.g., disease biomarker discovery" and is flanked by a schematic of a blood vessel and generic molecules. The second text box
Human metabolism is influenced by genetics and health status of the human body, nutrition and the microbiomes which exist on and in us (most notably the gut microbiome)(Reference Kastenmuller, Raffler and Gieger10,Reference Yadav, Verma and Chauhan11) . Nutrition can alter the composition of the gut microbiome(Reference Singh, Chang, Yan and Lee12–Reference David, Maurice and Carmody14), whereas the gut microbiome produces many bioactive compounds for the human host(Reference Varshney, Saini, Jha and Verma15,Reference Oliphant and Allen-Vercoe16) . Each individual has their own unique gut microbiome, which will react differently when people are given the same diet(Reference Simon, Sina and Ferrario17). The high variability in gut microbiome composition adds an additional layer of complexity when investigating nutrition and human health. A computational method that allows for disentangling the complex interactions between nutrition, the gut microbiome and the human host is genome-scale metabolic modelling(Reference Thiele, Sahoo and Heinken18) (Fig. 1).
This review outlines the general concepts of genome-scale metabolic modelling and the links between nutrition, the gut microbiome and human metabolism. Current methodologies are highlighted, with which the effect of nutrition on human metabolism through metabolic modelling can be examined. Finally, areas in the use and set-up of WBMs that require further research are discussed as well as potential applications of the mentioned methodologies.
Nutrition and human metabolism
Metabolism describes the collection of biochemical reactions in an organism. These reactions include breaking down (catabolism) or producing (anabolism) molecules. These biochemical reactions are usually catalysed by enzymes encoded in the genome and facilitate reaction rate by several orders of magnitude(Reference Sousa, Calixto and Ferreira19). In higher organisms, such as humans, metabolism is highly specific, with different organs and cell types exhibiting distinct metabolic capabilities(Reference Olander, Wisniewski and Artursson20–Reference Shroff, Aina and Maass22) and a spatial separation within the cells where enzymes are located in distinct subcellular locations(Reference Schmitt and An21).
Human metabolism starts at the digestive tract, which begins in the mouth where food is cut, ground and mixed, and enzymes are added that perform the initial breakdown of macromolecules(Reference Boland23). The food mixture is passed to the stomach where further enzymatic breakdown of the food occurs. The broken-down food enters the small intestine, where the majority of nutrients are taken up. The small intestine is also where bile is excreted, which contains bile acids that assist in lipid digestion and waste from the liver. The remainder of the metabolites not absorbed in the small intestine move to the large intestine, where additional nutrients are taken up(Reference Boland23). The large intestine is also the location in the human body with the largest community of microbes, known as the gut microbiome(Reference Perler, Friedman and Wu24,Reference Madhogaria, Bhowmik and Kundu25) . Finally, all the waste from the human body, gut microbiome and undigested metabolites are excreted in the faeces.
It is important to accurately measure which food in which quantities are ingested to understand which nutrients are available to the human body and the gut microbiome. Methods to measure diet composition to understand the foods people consume have been around for decades(Reference Todd, Hudes and Calloway26). Common methods include food frequency questionnaires, recall diets of various time intervals and more recently wearable camera technology(Reference Cuparencu, Diener and Wilson27). Although these methods give an insight into food consumption, there are limitations to what they can capture. Self-reporting methods are prone to participants over- or under-reporting consumption of food items(Reference Cuparencu, Diener and Wilson27–Reference Macdiarmid and Blundell29). The wearable camera methods could have difficulties obtaining individual ingredients and consumed amounts for highly mixed foods, such as soups or pies(Reference Cuparencu, Diener and Wilson27,Reference Rollo, Williams and Burrows30) .
The gut microbiome
The gut microbiome consists of billions of microbes, including archaea, bacteria, fungi and viruses(Reference Perler, Friedman and Wu24,Reference Madhogaria, Bhowmik and Kundu25) . The composition of the gut microbiome is unique to each individual(Reference Gilbert, Blaser and Caporaso31). The gut microbiome assists the digestive system and participates in metabolism by breaking down complex carbohydrates, such as resistant starch and fibre, and producing essential bioactive metabolites, such as short-chain fatty acids (i.e., acetate, propionate and butyrate), neurotransmitters, such as serotonin and dopamine, and vitamins(Reference Perler, Friedman and Wu24,Reference Durack and Lynch32) . Additionally, the gut microbiome modulates the human immune system and it protects the gut against pathogens(Reference Perler, Friedman and Wu24).
The composition of the gut microbiome is influenced by various factors such as socioeconomic status, geography, sex, age and diet. Of these, diet has shown it can rapidly alter the composition of the gut microbiome(Reference Singh, Chang, Yan and Lee12,Reference David, Maurice and Carmody14) . Nutrition can thus not only influences what the gut microbiome can produce but also which microbes are present. Influencing the presence and abundance of microbes is important as in many non-communicable diseases the gut microbiome composition is measurably changed(Reference Madhogaria, Bhowmik and Kundu25).
Methods to identify which microbes are present in the gut microbiome have changed and become cheaper and more comprehensive over the last decades(Reference Satam, Joshi and Mangrolia33,Reference Kuziel and Rakoff-Nahoum34) . The most common methods to discover which microbes are present in a sample are 16S rRNA and whole-genome shotgun sequencing(Reference Thomas, Clark and Dore35). The sequenced data is compared against a reference database containing the sequenced data of known microbes to find the closest related match. Each sequencing technique has its own advantages and disadvantages, 16S rRNA is cheaper but has a decreased sequencing depth, whereas whole-genome shotgun sequencing is more expensive but is able to distinguish microbial species in a microbiome sample(Reference Knight, Vrbanac and Taylor36). Typically, faecal samples are sequenced to understand which microbes are present in the gut microbiome. This approach assumes that the microbiome found in the faecal sample is representative of the entire large intestine(Reference Levitan, Ma and Giovannelli37,Reference Kong, Liang and Liu38) . This assumption has yielded valuable insights(Reference Madhogaria, Bhowmik and Kundu25,Reference Durack and Lynch32,Reference Yoshida, Yamashita and Hirata39) , but research has shown that the gut microbiome changes considerable depending on the location in the large intestine(Reference Kong, Liang and Liu38,Reference Tang, Jin, Wang and Liu40) . Other sampling techniques include biopsies, brushing and more recently capsules that capture the microbiome in specific areas of the colon(Reference Tang, Jin, Wang and Liu40,Reference Nejati, Sarnaik and Gopalakrishnan41) . These techniques offer specific microbiome composition in specific areas in the colon but are often invasive or not yet well established.
Once the microbes are identified, they can be characterised by metabolic function, ecological niches or pathogenicity. Additionally, metatranscriptomics(Reference Ojala, Hakkinen and Kankuri42) (expressed mRNA by the microbiome), metaproteomics (proteins expressed by the microbiome) and metabolomics(Reference Kong, Liang and Liu38,Reference Li, Chen, Gustafsson and Wang43) can further provide insights into the functional capabilities of the gut microbiome. These methods, however, often give microbiome-wide associations as opposed to species-specific interactions. Additionally, there are limitations the preparation of these omics, such extraction and yield, and reference databases can be incomplete(Reference Yang, Han and Lee44–Reference Valles-Colomer, Darzi and Vieira-Silva46). Another method that aims to elucidate, mechanistically, gut microbiome-human metabolism is genome-scale metabolic modelling.
What is genome-scale metabolic modelling
Genome-scale metabolic modelling is a mathematical approach to metabolism and is the core concept for the constraint-based reconstruction and analysis (COBRA) framework(Reference Heirendt, Arreckx and Pfau47). Converting metabolism of an organism to a mathematical representation is typically done by identifying the biochemical reactions based on its genome. Any missing pathways that have limited genomic annotation or consist of non-enzymatic reactions have to be added manually or through algorithms(Reference Thiele and Palsson48,Reference Marin de Mas, Herand and Carrasco49) . There are multiple tools available to automate the conversion of the genome of an organism to its corresponding mathematical representation(Reference Arkin, Cottingham and Henry50–Reference Zimmermann, Kaleta and Waschina54). Additionally, metabolic modelling can take information from the biological context to inform its predictions, e.g., consumption of metabolites to ensure maintenance of an organ or the intake of dietary nutrients.
Generally, metabolic modelling does not yet take into account enzyme kinetics or regulation of enzyme expression. Inclusion of these additional constraints requires information on all enzymes in the metabolic system which is often unknown for well-characterised bacteria, let alone for all possible human enzymes. However, there are tools available to add on additional constraints to the metabolic models based on transcriptomics or proteomics(Reference Agren, Bordel and Mardinoglu55–Reference Becker and Palsson57).
Current status of modelling human metabolism
Over the last two decades, metabolic modelling has been used to study bacteria(Reference Durot, Bourguignon and Schachter58), fungi(Reference Han, Tafur Rangel and Pomraning59), plants(Reference Baghalian, Hajirezaei and Schreiber60), animals(Reference Li, Chen, Gustafsson and Wang43) and humans(Reference Thiele, Sahoo and Heinken18,Reference Brunk, Sahoo and Zielinski61,Reference Robinson, Kocabas and Wang62) . In the context of diet-gut microbiome-human metabolism, metabolic models for the gut microbiome and the human are required. The latest microbiome collections, AGORA2 with over 7000 microbial models(Reference Heinken, Hertel and Acharya63) and APOLLO with over 230,000 microbial models(Reference Heinken, Hulshof and Nap64) can be used to create and simulate human microbiome metabolism. AGORA2 and APOLLO have been used to accurately predict changes in gut microbiome metabolism in different non-communicable diseases(Reference Heinken, Hertel and Thiele65–Reference Bhalla, Rengaswamy and Karunagaran69).
There are two mathematical models of generic human metabolism, Recond3D(Reference Brunk, Sahoo and Zielinski61) and Human1(Reference Robinson, Kocabas and Wang62), which have been used to simulate cell- and organ-specific metabolism. However, these models do not account for human physiology, organ-organ interactions and biofluid movements, leading to the creation of whole-body metabolic models (WBMs)(Reference Thiele, Sahoo and Heinken18). The WBMs are sex-specific and have over 20 organs and six cell types that are specific in their metabolic capacities and are connected through 13 different biofluids(Reference Thiele, Sahoo and Heinken18). WBMs representing infants have also recently been published, which allows for the metabolic modelling of infants from age day 0 to 180(Reference Zaunseder, Mutze and Okun70).
The WBMs can ‘consume’ a diet as long as the diet is defined by the metabolite constituents. There are eleven in silico diets available on the virtual metabolic human website (https://vmh.life) that represent generic diets(Reference Noronha, Modamio and Jarosz71), which allows for investigations into the influence of diet on human metabolism. However, creation of in silico diets is time-consuming when done manually and has limited the use of individual diets in WBM modelling, even when dietary information was available. Recently, the nutrition toolbox has been published, which allows for efficient and robust generation of in silico diets(Reference Nap, Weston and Brandt72). When dietary information is available, this toolbox can be used to ‘feed’ the WBM adjusted to the specific diet that an individual has actually consumed. Additionally, the nutrition algorithm(Reference Weston and Thiele73) has been adapted to be compatible with the WBMs. The nutrition algorithm calculates the smallest adjustment to an existing diet that is required to achieve the largest change in metabolism. The WBMs can also intake drugs, leading to the ability of predicting drug metabolism, paving the way for personalised medicine.
The WBMs can be further personalised by adjusting physiological parameters, such as weight or body fat percentage, based on an individual. Additionally, the gut microbiome models can be integrated with the large intestinal lumen of the WBMs to account for gut microbiome-host co-metabolism. The WBMs have been used to simulate metabolism in various disease contexts, such as inborn errors of metabolism(Reference Thiele, Sahoo and Heinken18), COVID-19(Reference Thiele and Fleming74), Parkinson disease(Reference Hensen and Thiele75), Alzheimer’s disease(Reference Martinelli, Heinken and Henning76) and metabolic syndrome(Reference Alessi, McCreery and Zomorrodi77).
Future prospects
The recent advances in genome-scale metabolic modelling outlined above have resulted in an attractive portfolio of computational modelling tools amenable towards the creation of digital metabolic twins (DMTs), i.e., digital replicas of an individual’s metabolism, for predictive decision-making along the nutrition-host-microbiome-health axis (Fig. 1).
The constituting resource are continuously updated to capture the growing knowledge content. The human reconstructions have grown from ∼3,000 reactions and metabolites (Recon1(Reference Duarte, Becker and Jamshidi78)) to now ∼80,000, including sex-, organ- and cell type-specific content in the WBMs(Reference Heinken, Magnusdottir and Fleming53). The microbiome efforts originated from reconstructing individual microbes(Reference Edwards and Palsson79) to entire resources now encompassing almost a quarter million microbes(Reference Heinken, Hulshof and Nap64). However, the current reconstructions primarily capture the metabolism of endogenous metabolites, i.e., further efforts are required to incorporate information on xenobiotics, including phytochemicals(Reference Rathaur and S80). Currently, in silico diets are created based on information derived from publicly available databases, such as the USDA FoodData Central(81) or FRIDA(82), which capture individual metabolite-level information for different food items. However, the metabolite coverage remains limited, at least in part due to current challenges in the required analytical measurement challenges in the field of foodomics(Reference Valdes, Alvarez-Rivera and Socas-Rodriguez83). Further, efforts are ongoing to specify the molecular composition of complex carbohydrates, such as starch or fibre, which are usually too broadly captured in current databases(Reference Suarez, Cheang and Larke84). Increasing the content of plant-specific metabolism both in current nutrition databases and in the genome-scale reconstructions will allow for more precise and accurate simulations with the WBMs, enabling better prediction of dietary supplementation from foods in the same food groups. Future developments will expand the scope of modelling from diet alone to a more holistic capture of health modulators, such as medication use, physical activity, lifestyle factors or environmental exposures, all of which influence metabolic function. Incorporating such information will allow models to capture diet–host–microbiome interactions in the broader context of modulators of human metabolism (e.g., environmental chemicals or lifestyle factors) and support nutrition strategies tailored to specific clinical conditions or health goals.
Current workflows have emerged primarily from a pre-clinical fundamental research context, with a focus on computational biology. Therefore, the specific tools and skills to generate COBRA-derived mechanism-informed hypotheses remain largely confined to a specialist computational modelling community. To enable in silico modelling-enabled decision support systems amenable to providing dietary advice, there is a need to collaborate with the intended end users, such as nutritionists, to create tools which align with their needs and expectations. Ongoing efforts to diversify the modelling user base include, e.g., the creation of the VMH (www.vmh.life) database(Reference Noronha, Modamio and Jarosz71), which hosts and interconnects the current genome-scale modelling resources through a user-friendly web interface.
Thus far, policymaking has relied on population-based statistics for deriving dietary guidelines. Therefore, while largely sound at a general population level, the guidelines may not align with the dietary requirements at an individual level, e.g., relating to the substantial variation in daily energy requirements(Reference Pontzer, Yamada and Sagayama85). It is further suggested that personalised approaches are more effective to catalyse changes in dietary behaviour than conventional approaches(Reference Celis-Morales, Livingstone and Marsaux86), i.e., population-level dietary guidelines may be ineffective at addressing ongoing challenges, such as tackling increasing obesity rates and associated co-morbidities(Reference Ahmed and Mohammed87). Genome-scale metabolic modelling may therefore be an attractive complement to evidence-based policymaking by enabling the creation of sub-population-derived models for hypothesis testing and validation, including in cases where the corresponding population data may not yet be available. For example, WBMs provide a straightforward solution for testing differences in males compared to females (by using the appropriate sex-specific models), which may help mitigate gender-related biases in large population surveys(Reference Weber, Gupta and Abdalla88). Notably, the WBMs can be personalised using different diets, i.e., dietary advice may be tested in silico to inform or validate the hypotheses derived from population-based evidence. Therefore, genome-scale metabolic modelling along the diet-host-microbiome-health axis may provide a powerful set of computational tools to inform future evidence-based policymaking efforts.
Overall, the recent advances in metabolic modelling provide a fruitful ground for developing DMTs to test dietary interventions in silico before implementation in real life. The current technology development trajectory supports decision-support systems for end users, such as clinicians and dietitians, enabling the comparison of alternative dietary scenarios and identifying interventions most likely to achieve a desired metabolic outcome. Further, integration with wearable devices and other digital health technologies may enable continuous data collection to support, e.g., personalised coaching for nutritional advice to be dynamically adjusted over time. Beyond the metabolic modelling as such, the successful implementation of DMT development will also rely on several enabling technology developments. The generation and availability of high-quality input data is essential, including accurate dietary assessment, expanded food composition databases capturing a comprehensive spectrum of food metabolites and high-resolution multi-omics methodologies to characterise host and microbiome function. Interoperability with established biomedical and nutrition databases, standardised data formats and robust analytical pipelines will be critical to ensure scalability and reproducibility. Ultimately, computational modelling approaches have the potential to empower individuals by providing transparent, biologically grounded explanations of how dietary choices may influence their metabolism and health. However, model predictions represent hypotheses derived from mathematical representations and must be validated and interpreted within the context of evidence-based nutritional practice. Close collaboration between modellers, nutrition professionals, clinicians and regulators will therefore be essential to translate metabolic modelling into safe, effective and sustainable precision nutrition.
Conclusion
Understanding how nutrition influences human metabolism has become more accessible with the rise of novel methodologies. However, that has also increased the complexity of linking the various data type together to formulate and test hypotheses to finally propose dietary advice. Genome-scale metabolic modelling with WBMs can incorporate various types of omics data and physiological parameters to accurately represent an individual’s metabolism leading the way to DMTs. Currently, human-microbiome co-metabolism can be modelled through the use of the AGORA2 and APOLLO resources, whereas creation of personalised in silico diets have been made more efficient and robust through the nutrition toolbox. Although advances in methodologies and applications for the WBMs have been in development, there are still areas that require further attention. The availability of metabolite content in food databases limits coverage the effects of nutrition on human metabolism simulations and there is of yet limited engagement with potential end users of the simulated results such as nutritionists and policy makers. However, the WBMs represent a crucial step to an efficient and reliable method to understand human metabolism and propose individual-specific dietary changes for health.
Acknowledgements
The authors acknowledge the Nutrition Society for the invitation to present this work at the Nutrition Society Conference 2026 in London.
Author contributions
B.N. gave the presentation, B.N. and C.T. drafted the manuscript, B.N, C.T. and I.T. revised the manuscript.
Financial support
This work was supported by the European Research Council (ERC) under the European Union’s Horizon 2020 Research and Innovation Programme [grant agreement no. 101125633], by the Science Foundation Ireland under Grant number 12/RC/2273-P2 and support of Taighde Éireann - Research Ireland and the Department of Agriculture, Food and Marine on behalf of the Government of Ireland underGrant Number [21/RC/10303_P2] - VistaMilk..
Competing interests
The authors declare none.

 Open access
Open access