


Introduction
Plant competition is one of the most powerful biological interactions shaping plant community structure and function. Its importance is evident in the substantial impact weeds have on crop survival, reproduction, and ultimately yield, an impact significant enough to have motivated the formation of our professional weed science societies. Given such importance, this review highlights our research and builds on the contributions of many individuals who have advanced the field of plant competition with the goal to deepening our understanding of the molecular and physiological mechanisms underlying plant competition and providing insight into how crop seedlings lose yield potential in the presence of neighboring weed seedlings.
Several questions challenge our current understanding of crop-weed competition. For example, how can crop seedlings lose yield potential simply from early exposure to neighboring weed seedlings, even when light, water, nutrients, and space are abundant? Also, are resource-based mechanisms sufficient to explain competitive interactions, or do additional processes remain to be identified? These questions are central to clarifying the still-elusive mechanisms underlying plant competition.
Within the plant competition literature, direct competition for resources is generally considered the primary evolutionary driver influencing competitive outcomes (Craine and Dybzinski Reference Craine and Dybzinski2013; Grime Reference Grime2006; Radosevich et al. Reference Radosevich, Holt and Ghersa2007; Zimdahl Reference Zimdahl2018). Resource competition can be defined as the capture of essential resources from a common, finite pool by neighboring individuals (Grime Reference Grime1979). Numerous studies in plant ecology and weed science have explored the effects of resource competition on plant community structure, growth, development, and seed yield (Crawley and May Reference Crawley and May1987; Ford Reference Ford2014; Page et al. Reference Page, Cerrudo, Westra, Loux, Smith, Foresman, Wright and Swanton2012; Tollenaar et al. Reference Tollenaar, Nissanka, Aguilera, Weise and Swanton1994). In agricultural systems, the role of resource limitation in crop-weed interactions is well established; insufficient access to light, water, nutrients, or space invariably reduces crop yield. Thus, resource availability remains a critical determinant of competitive outcomes.
The majority of plant competition studies involving resource limitation have traditionally measured morphological traits, such as root architecture, leaf appearance rate, leaf area development, total leaf number, and biomass partitioning, to explain yield loss in crops (Cathcart and Swanton Reference Cathcart and Swanton2003; Cerrudo et al. Reference Cerrudo, Page, Tollenaar, Stewart and Swanton2012; Chikoye et al. Reference Chikoye, Weise and Swanton1995; Mclachlan et al. Reference Mclachlan, Swanton, Weise and Tollenaar1993; Page et al. Reference Page, Cerrudo, Westra, Loux, Smith, Foresman, Wright and Swanton2012; Tollenaar et al. Reference Tollenaar, Nissanka, Aguilera, Weise and Swanton1994; Worthing et al. Reference Worthing, Mills and McKenzie-Gopsill2025). While these studies have provided valuable insights, the recorded traits are “descriptive outcomes” rather than “physiological or molecular mechanisms” (Rajcan and Swanton Reference Rajcan and Swanton2001; Trinder et al. Reference Trinder, Brooker and Robinson2013). If such morphological and partitioning changes are considered to be descriptive consequences of crop-weed interactions, what then are the mechanisms underlying competition?
Research Background
In order to address this fundamental question, we begin with our studies on the critical period for weed control (CPWC) and weed thresholds, because it was those studies that first prompted us to explore the potential of “resource independent plant competition”. In the early 1990s, we initiated a series of studies to determine the timing and duration of weed control required to prevent unacceptable yield loss. Knowledge of the CPWC is a key component of an integrated weed management strategy in field and vegetable cropping systems (Swanton and Murphy Reference Swanton and Murphy1996; Swanton and Valente Reference Swanton, Valente and RL2017; Swanton and Weise Reference Swanton and Weise1991; Swanton et al. Reference Swanton, Mahoney, Chandler and Gulden2008). The CPWC has two components; the first is the length of time weed control efforts must be maintained to prevent crop yield loss; the second is the length of time weeds can remain in the crop before they interfere with crop growth and ultimately reduce yield.
Studies conducted in field crops in Ontario found the critical period occurs between the second trifoliate and first flower stage of growth in white beans (Phaseolus vulgaris L.) (Wooley et al. 2017); from the 4-leaf to 14-leaf stage of growth in corn (Zea mays L.) (Hall et al. Reference Hall, Swanton and Anderson1992); and between the V2 and R3 stage of growth, approximately 9 to 38 d after soybean [Glycine max (L) Merr.] seedling emergence (Van Acker et al. Reference Van Acker, Swanton and Weise1993). A period of weed control lasting up to the fourth node growth stage of soybean, approximately 30 d after emergence, was adequate to prevent a yield loss of more than 2.5%. Similar studies continue to be published on a wide range of crops (Contreras et al. Reference Contreras, Leon, Post and Everman2025; Li et al. Reference Li, Zhu, Ma, Xu, Chen, Liu, Wang, Kan, Zheng and Chen2025; Mallick et al. Reference Mallick, Begum, Roy, Anwar, Rashid and Paul2025). Although the duration of the critical period can be affected by abiotic and biotic variables, such as weed species, planting pattern, and soil and environmental conditions, the results from these studies are helpful in determining when weed control is most critical.
One of the most interesting insights regarding plant competition obtained from these studies was the analysis of the second component of the CPWC; namely, the length of time weeds can remain in the crop before they interfere with crop growth and ultimately reduce yield. A short exposure to neighboring weed seedlings that had emerged with the crop seedling or within one to two leaf stages of crop seedling development resulted in a reduction in final crop yield (Hall et al. Reference Hall, Swanton and Anderson1992; Van Acker et al. Reference Van Acker, Swanton and Weise1993; Wooley et al. 2017). Under well-managed field conditions, how could a loss in yield potential occur due to early seedling exposure to weeds, despite plentiful resources? For example, the rate of corn yield loss following the onset of the CPWC ranged from 0.05 to 0.22 Mg ha−1 d− 1 (approximately 0.5% d−1 to 1.6% d−1) when corn was grown in the presence of neighboring weed seedlings from the V1 to V3 stage of growth (Page et al. Reference Page, Cerrudo, Westra, Loux, Smith, Foresman, Wright and Swanton2012). For the period from soybean emergence to the first flower stage of growth (approximately zero to 30 d after emergence), mean daily soybean yield loss per day due to weed competition was estimated to be 0.16% d−1 (Van Acker et al. Reference Van Acker, Swanton and Weise1993). In studies conducted in Argentina, early weed competition with soybean reduced final yield by 2.5% with only one day of competition and by 10% when weeds competed up to V2 (15 d after emergence) (Eyherabide and Cendoya Reference Eyherabide and Cendoya2009). This is a remarkable rate of yield loss, reflecting molecular, physiological, and morphological changes in crop seedlings that led to a rapid and irreversible yield loss despite plentiful light, water, nutrients, and space being available.
We continued our CPWC studies by introducing weed thresholds within the context of the critical period, as suggested by Swanton and Weise (Reference Swanton and Weise1991), to address whether weed species that emerge or escape control during the CPWC cause significant yield loss. It was our initial intent that our weed threshold studies would provide an action threshold, or useful information regarding the necessity and timing of herbicide treatments. Several researchers conducted threshold studies using corn (Bosnic and Swanton Reference Bosnic and Swanton1997; Cathcart and Swanton Reference Cathcart and Swanton2003; Knezevic et al. Reference Knezevic, Weise and Swanton1994), soybean (Cowan et al.Reference Cowan, Weaver and Swanton1998; Dieleman et al. Reference Dieleman, Hamill, Weise and Swanton1995), and white bean (Chikoye et al. Reference Chikoye, Weise and Swanton1995). A common ragweed density of 1.5 ragweed seedlings per meter of row emerging at the VE stage of white bean growth resulted in a yield loss of 10% to 22% compared to the weed-free treatment. Substantially less yield loss, of 4% to 9%, occurred when 1.5 ragweed seedlings per meter of row emerged at the V3 stage of white bean growth (Chikoye et al. Reference Chikoye, Weise and Swanton1995). Similarly, Bosnic and Swanton (Reference Bosnic and Swanton1997) reported that 30 barnyard grass seedlings per meter of row that emerged at the 3-leaf stage of corn development resulted in 14% yield loss compared with 4% yield loss when seedlings emerged at the 7-leaf stage.
All of these threshold studies conducted with different crops and weeds resulted in three important observations. First, the timing of weed emergence relative to the crop was critical in determining the magnitude of yield loss (see also Dew Reference Dew1972; O’Donovan et al. Reference O’Donovan, De St. Remy, O’Sullivan, Dew and Sharma1985). Second, depending on the time of weed emergence the outcome of competition varied dramatically across crop leaf stages, suggesting that the dynamics of the competitive interactions between crops and weeds were changing on a daily basis. Third, the presence of neighboring weed seedlings early in crop development caused a rapid and irreversible yield loss, despite resources being plentiful. As suggested by Horvath et al. (Reference Horvath, Clay, Swanton, Anderson and Chao2023), the fact that neighboring weed seedlings can affect crop yield early in the growing season even when resources are abundant, suggests that alternative mechanisms are in play other than direct resource competition. Such alternative mechanisms may include resource-independent processes such as the release of volatile or soil mobile compounds, which upon detection, alters signaling pathways including hormones and secondary metabolites, ultimately modifying crop growth and developmental trajectories (Aphalo et al. Reference Aphalo, Ballaré and Scopel1999; Kegge and Pierik Reference Kegge and Pierik2010; Kegge et al. Reference Kegge, Weldegergis, Soler, Eijk, Dicke, Voesenek and Pierik2013, Reference Kegge, Ninkovic, Glinwood, Welschen, Voesenek and Pierik2015; Pierik et al. Reference Pierik, Mommer and Voesenek2013).
If Not Resources, Then What?
Can differences in light spectral quality signals play a role in mediating resource-independent competition? In 1952, Borthwick et al. reported that red light promoted and far-red light inhibited the germination of lettuce seed (Lactuca sativa L.), suggesting the presence of a photo-reversible pigment. This photo-reversible pigment was isolated and identified as phytochrome, composed of two interconvertible forms: the red-absorbing Pr and the far-red absorbing Pfr (Borthwick and Hendricks Reference Borthwick and Hendricks1960; Butler et al.Reference Butler, Norris, Siegelman and Hendricks1959; Holmes and Smith Reference Holmes and Smith1975; Huq et al. Reference Huq, Lin and Quail2024). In darkness, phytochrome exists primarily as Pr, which is converted to the physiologically active Pfr form upon exposure to red light. The conversion of the inactive Pr to the physiologically active Pfr can regulate morphological developmental responses in plants based on changes in the light quality environment (Schmitt and Wulff Reference Schmitt and Wulff1993) (Figure 1).
A mechanistic model of red and far-red light regulation of phytochrome signaling and downstream growth-defense trade-offs. Under high R:FR light, phytochrome B (phyB) is maintained in its biologically active Pfr form, which translocates to the nucleus and promotes phosphorylation and proteasomal degradation of phytochrome-interacting factors (PIFs). This represses elongation-associated gene expression and supports salicylic acid (SA)-dependent and jasmonate (JA)-dependent defense signaling. Under low R:FR light, far-red enrichment converts phyB to the inactive Pr form, reducing nuclear phyB levels and allowing PIF accumulation. PIFs activate auxin and gibberellin biosynthesis and signaling pathways, promoting cell elongation and shade-avoidance responses. Concurrently, low R:FR attenuates SA and JA signaling, reflecting a resource allocation trade-off between growth and immune function.

Kasperbauer (Reference Kasperbauer1971) determined that plants exposed to far red (FR) light developed longer internodes, were light green in color, had thinner leaves, and exhibited no axillary branching compared to red-irradiated plants. Further studies determined that FR light, once sensed by phytochrome, altered the partitioning of photosynthate; plants developed longer stems, higher shoot-to-root ratios, reduced root biomass, thinner leaves, inhibition of branching, closure of stomata, leaf angle changes, early flowering, changes to chlorophyll a and b synthesis, and a reduction in root nodules, root biomass, and rooting depth (Gal et al. Reference Gal, Afifi, Lee, Lukens and Swanton2015; Kasperbauer and Hunt Reference Kasperbauer and Hunt1994; Kasperbauer et al. Reference Kasperbauer, Hunt and Sojka1984; Wang et al. Reference Wang, Jiao, Zhang, Shi, Wang, Zhou, Xu, Wang, Kong, Zhang and Li2025). These changes in growth and development have since been determined to be mediated by multiple photoreceptors, notably phytochrome B, and are now collectively described as the shade avoidance syndrome (SAS) (Ballaré and Pierik Reference Ballaré and Pierik2017; Smith Reference Smith2000; Smith and Whitelam Reference Smith and Whitelam2008). These morphological changes associated with SAS may, under certain conditions, optimize plant canopy performance, thereby increasing the potential for optimum canopy photosynthesis (Aphalo et al. Reference Aphalo, Ballaré and Scopel1999; Rajcan et al. Reference Rajcan, Chandler and Swanton2004; Zhang et al. Reference Zhang, Van Westreenen, He, Evers, Anten and Marcelis2021).
Are these temporary changes or do they persist? Chitwood et al. (Reference Chitwood, Kumar, Ranjan, Pelletier, Townsley, Ichihashi, Martinez, Zumstein, Harada, Maloof and Sinha2015) observed that the changes associated with SAS during early leaf development in tomato persisted as the plant grew, despite being transferred from a light quality treatment of the red to far-red ratio (R:FR) = 0.5 to 1.5. This observation suggested that once these changes occur, they may persist throughout the lifecycle of the plant, supporting the concept of a “morphological legacy effect” or carryover effect. A study by McKenzie-Gopsill et al. (Reference McKenzie-Gopsill, Amirsadeghi, Fillmore and Swanton2020) reported that different durations of weed presence affected photosynthetic efficiency and yield recovery in common bean. Although photosynthetic efficiency improved once weed competition ended, full recovery did not occur when weeds had been present for an extended period. Additionally, when photosynthetic metrics partially rebounded, yield did not fully recover if weeds had competed with crops beyond a certain critical period. These studies not only reinforce the concept of a critical weed-free period but also suggest a legacy effect, in which past physiological stresses due to the presence of neighboring weed seedlings continued to influence crop performance even after the weeds were removed. Research continues to support the concept that changes in the light quality, specifically the R:FR ratio detected by phytochrome, may result in a persistent alteration of plant growth and development (Horvath et al. Reference Horvath, Bruggeman, Moriles-Miller, Anderson, Dogramaci, Scheffler, Hernandez, Foley and Clay2018; Lazzarin et al. Reference Lazzarin, Dupont, van Ieperen, Marcelis and Driever2025; Reid et al. Reference Reid, Gonzalez, Sikkema, Lee, Lukens and Swanton2014; Ugarte et al. Reference Ugarte, Trupkin, Ghiglione, Slafer and Casal2010).
We interpret the detection of FR light emitted from neighboring weeds by crop plants not only as a warning of impending competition (Ballaré and Austin Reference Ballaré and Austin2019; Ballaré and Casal Reference Ballaré and Casal2000; Ballaré et al. Reference Ballaré, Sánchez, Scopel, Casal and Ghersa1987, Reference Ballaré, Scopel and Sanchez1990) but also as an environmental signal that can trigger molecular and physiological mechanisms that will result in a reproductive cost, thereby altering the yield potential of a crop plant. Throughout our experimental methodology, we used supplemental FR treatments combined with weed proximity to distinguish FR-specific effects from those caused by the biological presence of weeds, including species- and cultivar-specific volatile organic compounds (VOCs) (Effah et al. Reference Effah, Holopainen and McCormick2019; Kegge et al. Reference Kegge, Ninkovic, Glinwood, Welschen, Voesenek and Pierik2015; Kigathi et al. Reference Kigathi, Weisser, Reichelt, Gershenzon and Unsicker2019; Ninkovic et al. Reference Ninkovic, Rensing, Dahlin and Markovic2019). We acknowledge that VOCs, ranging from ethylene and methyl jasmonate to volatile terpenes and other green-leaf compounds, are emitted in response to biotic and abiotic stressors (Jin et al. Reference Jin, Zhao, Jing, Zhang, Lu, Yu, Wang, Guo, Pan, Hoffmann, Schwab and Song2023; Kessler et al. Reference Kessler, Mueller, Kalske and Chautá2023). Our experimental design revealed how early season light cues can initiate changes that ultimately reduce reproductive output. In all our laboratory and field experiments, both crop and weed seedlings were grown under non-limiting resource conditions. For experimental details see studies reported by Berardi et al. (Reference Berardi, Amirsadeghi and Swanton2024), Green-Tracewicz et al. (Reference Green-Tracewicz, Page and Swanton2012), Kramer et al. (Reference Kramer, Amirsadeghi, McKenzie-Gopsill and Swanton2025), McKenzie-Gopsill et al. (Reference McKenzie-Gopsill, Lee, Lukens and Swanton2016a), and Page et al. (Reference Page, Liu, Cerrudo, Lee and Swanton2011).
Results and Discussion
Weed-Driven Light Signals, Shade Avoidance, and Crop Morphological Plasticity
In natural ecosystems, the shade-avoidance response is considered an adaptive strategy, but in an agricultural system, this same response can be undesirable because reallocating biomass toward shoot and internode elongation can compromise yield (Ballenger et al. Reference Ballenger, Adjesiwor, Claypool and Kniss2024; Green-Tracewicz et al. Reference Green-Tracewicz, Page and Swanton2011; McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Amirsadeghi, Fillmore and Swanton2020; Page et al. Reference Page, Tollenaar, Lee, Lukens and Swanton2009, Reference Page, Tollenaar, Lee, Lukens and Swanton2010). For example, corn seedlings emerging into a canopy of redroot pigweed (Amaranthus retroflexus L.), which generates a consistent low R:FR of 0.3 to 0.7, exhibit increased plant height and delayed rate of leaf appearance compared with seedlings that emerge in a weed-free environment (Liu et al. Reference Liu, Mahoney, Sikkema and Swanton2009). Although internode elongation is the archetypal shade avoidance characteristic, increased plant height by corn under low R:FR conditions are often transient. In a glasshouse study, Page et al. (Reference Page, Tollenaar, Lee, Lukens and Swanton2009) observed increased seedling height 3 d after emergence, but this difference disappeared 6 d later, after which seedlings of weedy and weed-free treatments remained similar in height. Similarly, Liu et al. (Reference Liu, Mahoney, Sikkema and Swanton2009) reported a 2% to 10% increase in height from the V1 stage to the V9 stage when corn emerged into a canopy of redroot pigweed, yet as development progressed, height equalized such that no differences were detected at the V11 stage.
Indeed, many studies have noted that corn tends to be shorter in response to low R:FR light such as those created by high-density planting or severe shade (Fournier and Andrieu Reference Fournier and Andrieu2000; Jafari et al. Reference Jafari, Wang, Wang and Zou2024). Substantial genotypic variation exists in corn height response to low R:FR with select hybrids displaying zero or minimal elongation, whereas others display rapid height increases (Hallauer and Sears Reference Hallauer and Sears1969; Maddonni et al. Reference Maddonni, Otegui, Andrieu, Chelle and Casal2002; Ren et al. Reference Ren, Li, Dong, Liu, Zhao and Zhang2017; Stupar et al. Reference Stupar, Gardiner, Oldre, Haun, Chandler and Springer2008). Interestingly, corn height responses seem to be correlated with canopy architecture. Corn canopies with random leaf orientation exhibit minimal height response, while those that reorganize leaf angle away from low R:FR light exhibit greater elongation (Maddonni et al. Reference Maddonni, Otegui, Andrieu, Chelle and Casal2002; Zhou et al. Reference Zhou, Kusmec and Schnable2024). Elongation in response to low R:FR in corn is partially mediated by chloroplast ribosomal proteins such as DBB2, which modulates overall plant height via effects of mesocotyl, leaf, and leaf sheath elongation (Wang et al. Reference Wang, Jiao, Zhang, Shi, Wang, Zhou, Xu, Wang, Kong, Zhang and Li2025). Increased corn canopy height due to early season weed competition can influence lodging risk (Zhang P et al. Reference Zhang, Van Westreenen, He, Evers, Anten and Marcelis2021) and ear placement (Zhao et al. Reference Zhao, Zhang, Lv, Ning, Cao, Liao, Wang and Huang2022), highlighting the potential of gene targets such as DBB2 for improving stress tolerance in corn.
In contrast to corn, soybean is highly plastic and typically displays a strong shade avoidance response, with stem elongation occurring as early as emergence (McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Lee, Lukens and Swanton2016a). This indicates that light signals reflected from neighboring weeds can penetrate the upper layers of soil, be perceived by soybean phytochrome, and influence developmental morphology prior to emergence. Indeed, longer wavelengths such as FR light can pass through soil pores and trigger widespread transcriptomic changes in soybean (Benvenuti Reference Benvenuti1995; Bliss and Smith Reference Bliss and Smith1985; McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Lee, Lukens and Swanton2016a; Tester and Morris Reference Tester and Morris1987). As in corn, there exists considerable variation in shade avoidance responses across soybean genotypes including plant height (Campos et al. Reference Campos, Leite, Matos, Dobbss, Nicoli and Evaristo2024; Wu et al. Reference Wu, Yang, Gong, Shoaib, Fan, Wu, Yong, Liu, Shu, Liu, Du and Wang2017; Zhang et al. Reference Zhang, Hong, Liu, Wang, Zhang, Zhao, Yuan, Abdelghany, Zhang, Lamlom and Ren2025). Increased plant height early in development is not necessarily detrimental to soybean yield because this may allow for greater yield potential by increasing the number of nodes and branches available for pod formation (Chang et al. Reference Chang, Guo, Sun, Zhang, Wang, Kong, He, Sharmin and Zhao2018; Ravelombola et al. Reference Ravelombola, Qin, Shi, Song, Yuan, Wang, Chen, Yan, Feng, Zhao and Meng2021). However, branching is often suppressed in soybean under low R:FR conditions (Green-Tracewicz et al. Reference Green-Tracewicz, Page and Swanton2011; Liu et al. Reference Liu, Rahman, Song, Su, Yang, Yong, Wu, Zhang and Yang2017), compromising leaf formation and resulting in suboptimal canopy structure (Luo et al. Reference Luo, Yuan, Zhang, Fu, Lin, Li, Li, Pu, Wang, Yang and Yong2025). Because light interception over the growing season is highly correlated with yield potential in soybean (Gaspar and Conley Reference Gaspar and Conley2015; Koester et al. Reference Koester, Skoneczka, Cary, Diers and Ainsworth2014), a poorly developed canopy may reduce canopy-level photosynthesis and limit yield. The extent to which low R:FR reflected upward from neighboring weeds alters soybean canopy architecture and affects final yield remains poorly understood.
Shade Avoidance Is a Driver of Yield-Limiting Biomass Loss
A lasting consequence of weed-induced shade avoidance during the CPWC is a gradual and progressively greater reduction in both aboveground and belowground biomass accumulation (Cerrudo et al. Reference Cerrudo, Page, Tollenaar, Stewart and Swanton2012; Green-Tracewicz et al. Reference Green-Tracewicz, Page and Swanton2012; Liu et al. Reference Liu, Mahoney, Sikkema and Swanton2009; McKenzie-Gopsill et al. Reference McKenzie-Gopsill2016; Page et al. Reference Page, Tollenaar, Lee, Lukens and Swanton2010). For example, Liu et al. (Reference Liu, Mahoney, Sikkema and Swanton2009) reported a 5% decline in corn stem biomass and 22% decline in ear biomass when weed removal was delayed from the V3 stage to silking. Similarly, Page et al. (Reference Page, Tollenaar, Lee, Lukens and Swanton2009) and Cerrudo et al. (Reference Cerrudo, Page, Tollenaar, Stewart and Swanton2012) observed corn aboveground biomass declines of 32% to 82% when weed removal was delayed from the V1 stage to the V10 stage. Soybean displays comparable reductions in biomass in response to early and persistent low R:FR reflected from neighboring weeds. Green-Tracewicz et al. (Reference Green-Tracewicz, Page and Swanton2011) observed declines of 27% to 38% in shoot biomass and 40% to 48% in root biomass as weed removal was delayed from the first trifoliate to the pod-fill stage. Reduced biomass associated with shade avoidance cues from neighboring weeds has also been observed in sugar beet (Beta vulgaris L.). Schambow et al. (Reference Schambow, Adjesiwor, Lorent and Kniss2019) found that exposure to low R:FR reflected from neighboring weeds from emergence to harvest reduced sugar beet shoot biomass by 21% to 30% and root biomass by 70% to 72%, along with 10 to 14 fewer leaves and a 48% to 57% reduction in leaf biomass 90 d after planting. In all of these studies, crops and weeds were luxury fed and grown without physical contact between plants. This demonstrates that declines in crop biomass and shifts in biomass allocation patterns are not driven by early season resource competition, but rather by the expression of the shade avoidance phenotype itself.
Plant-to-Plant Variability and Yield Consequences of Shade Avoidance
In addition to changes in biomass accumulation, induction of shade avoidance in response to low R:FR reflected from neighboring weeds alters biomass allocation patterns and increases plant-to-plant variability, which can have lasting consequences for stress tolerance and final yield. Cerrudo et al. (Reference Cerrudo, Page, Tollenaar, Stewart and Swanton2012) and Page et al. (Reference Page, Liu, Cerrudo, Lee and Swanton2011) noted increased plant-to-plant variability among corn plants in aboveground biomass, creating size hierarchies that can be amplified by subsequent stress. Furthermore, increased allocation of biomass to shoots at the expense of roots in corn can shift the overall population below the critical biomass threshold required for kernel set, acting additively with additional stressors (Page et al. Reference Page, Liu, Cerrudo, Lee and Swanton2011). It is through the combined effects of multiple stressors post-expression of the shade avoidance response that can disrupt reproductive allometry and lead to yield loss. For instance, delayed weed control lengthens the anthesis-silking interval, slowing ear development and silk extrusion (Bassetti and Westgate Reference Bassetti and Westgate1994; Reid et al. Reference Reid, Gonzalez, Sikkema, Lee, Lukens and Swanton2014). A shorter anthesis-silking interval is associated with increased yield via improved fertilization success (Hall et al. Reference Hall, Vilella, Trapani and Chimenti1982). As such, the anthesis-silking interval has been a key target in corn breeding programs to improve tolerance to stressors such as drought and nutrient deficiency (Bolaños and Edmeades Reference Bolaños and Edmeades1996; Silva et al. Reference Silva, Sanchez, Opazo, Mardones and Acevedo2022) and may also be relevant to improving crop yield stability against weeds.
Similarly, sugar beet exhibits limited ability to adjust its growth trajectory following commitment to a shade avoidance phenotype. Adjesiwor et al. (Reference Adjesiwor, Ballenger, Weinig, Ewers and Kniss2021) found that weeds present at emergence of sugar beet and controlled at the second leaf stage, reduced leaf number, leaf area, and leaf biomass by 7%, 22%, and 25%, respectively. These reductions were similar regardless of whether weeds were removed at the second leaf stage or allowed to grow for the remainder of the growing season. However, when sugar beet was maintained weed-free until the second-leaf stage and then exposed to weeds, the negative effects on leaf traits became progressively more severe (Adjesiwor et al. Reference Adjesiwor, Ballenger, Weinig, Ewers and Kniss2021). This indicates that early exposure to low R:FR cues triggers a shade avoidance response that commits sugar beet to a growth strategy focused on light competition. Although adaptive, this strategy reduces the plant’s ability to reallocate resources in response to later stress cues (Weinig and Delph Reference Weinig and Delph2001).
An increase in plant-to-plant variability associated with induction of the shade avoidance response has also been reported in soybean with the potential to influence stress tolerance (Green-Tracewicz et al. Reference Green-Tracewicz, Page and Swanton2011). Unlike corn and sugar beet, however, this does not appear to increase susceptibility to stress because soybean exhibits substantial vegetative and reproductive plasticity, enabling resource reallocation in response to additional stresses (McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Lukens, Lee and Swanton2016b). As a result, soybean can often recover from early season weed competition when resources remain sufficient throughout the remainder of the growing season. In contrast, other studies have noted an increase in shoot and root biomass in response to declining R:FR suggesting the context- and species-dependent nature of FR responsiveness and cross-talk between shoot and root developmental responses (Li et al. Reference Li, Hamani, Si, Liang, Gao and Duan2020; van Gelderen et al. Reference van Gelderen, Kang, Paalman, Keuskamp, Hayes and Pierik2018; Wang et al. Reference Wang, Mao, Ji, Wang and Fu2024). Taken together, these findings suggest that crop perception of low R:FR reflected from neighboring weeds elicits a shade avoidance response that alters biomass partitioning, breaks reproductive allometry, and reduces stress tolerance, contributing to decreased yield with variability across species. The extent to which this response can be mitigated or regulated across a wide range of crop species remains poorly understood and represents a promising target for genetic manipulation and selection in breeding programs to improve crop competitive ability against weeds without sacrificing stress tolerance and yield.
Canopy Architecture, Light Capture, and Photosynthetic Responses Under Competition
Changes to biomass allocation patterns associated with the shade avoidance response during the CPWC can have lasting consequences on biomass accumulation and final yield. These changes often are linked to reductions in leaf- and canopy-level photosynthesis. Studies examining the effects of FR light and the shade avoidance response on the photosynthetic apparatus show that responses vary by species, cultivar, and light intensity (Cagnola et al. Reference Cagnola, Ploschuk, Benech-Arnold, Finlayson and Casal2012; Huber et al. Reference Huber, de Boer, Romanowski, van Veen, Buti, Kahlon, van der Meijden, Koch and Pierik2024; Murchie and Horton Reference Murchie and Horton1997; Shibuya et al. Reference Shibuya, Endo, Hayashi and Kitaya2012; Xu et al. Reference Xu, Wang, Wang, Li, Liu and Fu2022; Zhou et al. Reference Zhou, Kusmec and Schnable2024; Zienkiewicz et al. Reference Zienkiewicz, Drożak, Wasilewska, Bacławska, Przedpełska-Wąsowicz and Romanowska2015) with direct implications for understanding weed-induced yield loss during the CPWC.
FR light alone contributes minimally to quantum yield of photosynthesis (Zhen and Bugbee Reference Zhen and Bugbee2020; Zhen and van Iersel Reference Zhen and van Iersel2017; Zhen et al. Reference Zhen, Haidekker and van Iersel2019). However, FR light can synergistically enhance photosynthesis in combination with other wavelengths via the Emerson effect (Emerson Reference Emerson1958). FR photons preferentially excite photosystem I (PS I), allowing for remaining photons to over-excite PS II, leading to increased photosynthetic rate (Huber et al. Reference Huber, de Boer, Romanowski, van Veen, Buti, Kahlon, van der Meijden, Koch and Pierik2024; Laisk et al. Reference Laisk, Oja, Eichelmann and Dall’Osto2014; Yang et al. Reference Yang, Feng, Liu, Wu, Fan, Raza, Cheng, Chen, Wang, Yong and Liu2018, Reference Yang, Liu, Cheng, Feng, Wu, Fan, Raza, Wang, Yong, Liu and Liu2020; Zhen and Bugbee Reference Zhen and Bugbee2020; Zhen and van Iersel Reference Zhen and van Iersel2017). Long-term exposure to FR light shifts photosynthetic stoichiometry, favoring PS I excitation, leading to an increase in the relative abundance of PS II (Feild et al. Reference Feild, Nedbal and Ort1998; Melis and Harvey Reference Melis and Harvey1981; Smith et al. Reference Smith, Samson and Fork1993). This shift, though, can decrease the capacity of a plant to acclimate to higher light intensity later in development (Morelli et al. Reference Morelli, Paulišić, Qin, Iglesias-Sanchez, Roig-Villanova, Florez-Sarasa, Rodriguez-Concepcion and Martinez-Garcia2021). Plants acclimated to FR light in general display a reduction in net photosynthesis (Gómez-Ocampo et al. Reference Gómez-Ocampo, Cascales, Medina-Fraga, Ploschuk, Mantese, Crocco, Matsusaka, Sánchez and Botto2023; Hirth et al. Reference Hirth, Dietzel, Steiner, Ludwig, Weidenbach and Pfannschmidt2013; Maliakal et al. Reference Maliakal, McDonnell, Dudley and Schmitt1999; Sleeman and Dudley Reference Sleeman and Dudley2001; Xu et al. Reference Xu, Wang, Wang, Li, Liu and Fu2022; Zienkiewicz et al. Reference Zienkiewicz, Drożak, Wasilewska, Bacławska, Przedpełska-Wąsowicz and Romanowska2015). Altered R:FR ratios have been shown to strongly influence the expression of photosynthesis-related genes, reducing overall photosynthetic capacity with variability across plant species (Cagnola et al. Reference Cagnola, Ploschuk, Benech-Arnold, Finlayson and Casal2012; Devlin et al. Reference Devlin, Yanovsky and Kay2003; Geisler et al. Reference Geisler, Gibson, Lindsey, Millar and Wood2012; Gómez-Ocampo et al. Reference Gómez-Ocampo, Cascales, Medina-Fraga, Ploschuk, Mantese, Crocco, Matsusaka, Sánchez and Botto2023; Horvath et al. Reference Horvath, Gulden and Clay2006, Reference Horvath, Hansen, Moriles-Miller, Pierik, Yan, Clay, Scheffler and Clay2015, Reference Horvath, Clay, Bruggeman, Anderson, Chao and Yeater2019; Moriles et al. Reference Moriles, Hansen, Horvath, Reicks, Clay and Clay2012).
Reductions in CO2 assimilation rate have been observed in several plant species during interspecific competition including spring wheat (Iqbal and Wright Reference Iqbal and Wright1999), cotton (Ratnayaka et al. Reference Ratnayaka, Molin and Sterling2003; Niu et al. Reference Niu, Yuan, Zhang, Liu, Zhang, Huang and Wan2005; Reynolds et al. Reference Reynolds, Simpson, Thevathasan and Gordon2007), and soybean (McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Amirsadeghi, Earl, Jones, Lukens, Lee and Swanton2019, Reference McKenzie-Gopsill, Amirsadeghi, Fillmore and Swanton2020). Interestingly, these reductions in CO2 assimilation were not attributed to changes in stomatal conductance (Iqbal and Wright Reference Iqbal and Wright1999; McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Amirsadeghi, Earl, Jones, Lukens, Lee and Swanton2019, Reference McKenzie-Gopsill, Amirsadeghi, Fillmore and Swanton2020; Niu et al. Reference Niu, Yuan, Zhang, Liu, Zhang, Huang and Wan2005), light interception (McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Amirsadeghi, Earl, Jones, Lukens, Lee and Swanton2019; Reynolds et al. Reference Reynolds, Simpson, Thevathasan and Gordon2007), or water or nutrient availability (Iqbal and Wright Reference Iqbal and Wright1999; McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Amirsadeghi, Earl, Jones, Lukens, Lee and Swanton2019; Reynolds et al. Reference Reynolds, Simpson, Thevathasan and Gordon2007; Robinson et al. Reference Robinson, Wagner, Bell and Swanton2001). Rather, low R:FR reflected from neighboring weeds has been associated with several anatomical and biochemical acclimation responses in the photosynthetic machinery. Long-term exposure to shade light enriched in FR drives photosynthetic acclimation responses, including changes to leaf structure (Gómez-Ocampo et al. Reference Gómez-Ocampo, Cascales, Medina-Fraga, Ploschuk, Mantese, Crocco, Matsusaka, Sánchez and Botto2023) and downregulation of genes associated with photosynthetic pigments and light harvesting (Cagnola et al. Reference Cagnola, Ploschuk, Benech-Arnold, Finlayson and Casal2012; Huber et al. Reference Huber, de Boer, Romanowski, van Veen, Buti, Kahlon, van der Meijden, Koch and Pierik2024). For example, Gómez-Ocampo et al. (Reference Gómez-Ocampo, Cascales, Medina-Fraga, Ploschuk, Mantese, Crocco, Matsusaka, Sánchez and Botto2023) observed that potato leaves (Solanum tuberosum L.) exposed to shade light had a thinner mesophyll cell layer yet a thicker epidermis cell layer with no overall change in leaf thickness. The thicker epidermis layer in potato leaves increased mesophyll conductance and suppressed the CO2 assimilation rate (Gómez-Ocampo et al. Reference Gómez-Ocampo, Cascales, Medina-Fraga, Ploschuk, Mantese, Crocco, Matsusaka, Sánchez and Botto2023). In contrast, Huber et al. (Reference Huber, de Boer, Romanowski, van Veen, Buti, Kahlon, van der Meijden, Koch and Pierik2024) exposed seedlings of seven cultivars of rice (Oryza sativa L.) to supplemental FR light and observed an increase in leaf area, yet leaf dry matter decreased, suggesting thinner leaves. Huber et al. (Reference Huber, de Boer, Romanowski, van Veen, Buti, Kahlon, van der Meijden, Koch and Pierik2024) observed an overall decline in total chlorophyll content in rice, yet no effect on photosynthetic efficiency across irradiance levels. Interestingly, there was an instantaneous rise in CO2 assimilation in rice seedlings pretreated with a pulse of FR light. Pretreatment of rice seedlings with supplemental FR light resulted in a doubling of this rate as compared with control plants and overall growth promotion. This may point to a priming response and a mechanism for improving crop stress tolerance in response to weed competition and alleviating the impact of weeds on crop photosynthesis.
The effects of low R:FR ratio and the shade avoidance response on photosynthetic parameters varies widely across crops and within cultivars. It is well documented that modern corn hybrids tolerate a higher density planting than historic hybrids. This has been attributed to improved canopy architecture and light capture in modern corn hybrids that allows the maintenance of photosynthetic capacity at high density (Huang et al. Reference Huang, Guo, Tan, Zhang, Li, Zhou and Duan2024; Yan et al. Reference Yan, Duan, Li, Zhao, Hou, Zhao, Li, Wang, Dai and Zhou2024; Ye et al. Reference Ye, Chen, Yu, Gao, Muneer, Fan, Wu and Wu2025; Zhou et al. Reference Zhou, Kusmec and Schnable2024). Furthermore, canopy architecture in corn is understood to be mediated by FR light with partial overlap in genes involved in shade avoidance responses. Screening a wide panel of corn hybrids, Zhou et al. (Reference Zhou, Kusmec and Schnable2024) found that the expression of ligule development genes was essential for repositioning of corn leaves perpendicular to the row to maximize light interception. Rajcan et al. (Reference Rajcan, Chandler and Swanton2004) demonstrated a similar phenomenon in corn leaves in response to low R:FR reflected from neighboring weeds. Corn leaves shifted away from weeds and low R:FR and toward empty space to maximize light interception through twisting of internodes (Girardin and Tollenaar Reference Girardin and Tollenaar1994; Rajcan et al. Reference Rajcan, Chandler and Swanton2004). Yet, there is great variability in canopy responses to inter- and intra-specific competition. Marin and Weiner (Reference Marín and Weiner2014) and later, Marin (Reference Marín2021), observed that corn hybrids with the greatest variability in leaf angles were the least competitive yet had minimal response in grain yield to increased plant density. Therefore, stability in canopy architecture and accompanying maintenance of photosynthetic activity may be essential to improvements in corn and other crops response to low R:FR from both intra- and inter-specific competition (Perez et al. Reference Perez, Fournier, Cabrera-Bosquet, Artzet, Pradal, Brichet, Chen, Chapuis, Welcker and Tardieu2019).
Biochemical Constraints on Photosynthesis in Response to Weed Competition
In addition to the anatomical and architectural changes associated with shade avoidance, biochemical limitations have also been implicated in the downregulation of photosynthetic activity. In tomato (Solanum lycopersicum L.), low R:FR triggers downregulation of genes involved in the Calvin cycle and isoprenoid, chlorophyll, and carotenoid biosynthesis pathways (Cagnola et al. Reference Cagnola, Ploschuk, Benech-Arnold, Finlayson and Casal2012). This coincided with a decline in CO2 assimilation, with a larger reduction observed in stem tissue than leaf tissue. This was attributed to an energetic trade-off by tomato seedlings to prioritize stem elongation over photosynthetic capacity (Cagnola et al. Reference Cagnola, Ploschuk, Benech-Arnold, Finlayson and Casal2012). Similarly, Sales et al. (Reference Sales, Ribeiro, Marchiori, Kromdijk and Machado2023) documented metabolic limitations on the Calvin cycle suppressing photosynthetic capacity in sugarcane in response to low R:FR. The authors noted an increase in the number of closed PS II reaction centers in sugarcane exposed to low R:FR, indicating they were reduced and unavailable for photochemical quenching. This was accompanied by a decline in the maximum carboxylation rate (V cmax ), which measures the speed of ribulose-1,5-bisphosphate (RuBP) carboxylation, and no impact on stomatal conductance. These findings suggest that biochemical limitations, rather than anatomical or stomatal limitations, primarily affect crop photosynthetic responses to weeds.
Suppression of photosynthesis by low R:FR is not universal. There is wide phenotypic variability in shade avoidance responses across species. While many crop plants display a classic shade avoidance phenotype to low R:FR, others including several members of the Brassicaceae family, such as hairy bittercress (Cardamine hirsute L.) or watercress (Nasturtium officinale W. T. Aiton), display a shade-tolerant phenotype. Morelli et al. (Reference Morelli, Paulišić, Qin, Iglesias-Sanchez, Roig-Villanova, Florez-Sarasa, Rodriguez-Concepcion and Martinez-Garcia2021) found that CO2 assimilation was similar in low and high light in all five of the shade-tolerant Brassica species they evaluated. In contrast, shade-avoiding species, such as shepherd’s purse [Capsella bursa-pastoris (L.) Medik.] and pink shepherd’s purse (Capsella rubella Reut.), had compromised CO2 assimilation when moved from low to high light. The chloroplast ultrastructure was modified in these shade-avoiding species with larger grana stacks and fewer, smaller plastoglobules. Morelli et al. (Reference Morelli, Paulišić, Qin, Iglesias-Sanchez, Roig-Villanova, Florez-Sarasa, Rodriguez-Concepcion and Martinez-Garcia2021) suggested these alterations precondition shade-avoiding plants to low light conditions. Yet, when light conditions change these species have reduced capacity to remodel their chloroplast ultrastructure and suffer reduced photosynthetic capacity, demonstrating wide phenotypic variability in shade avoidance responses.
Persistence Photosynthetic Penalties from Early Weed Presence
Recent studies in our laboratory have shown that early season weed competition during the CPWC can have a lasting impact on photosynthetic rate and efficiency. McKenzie-Gopsill et al. (Reference McKenzie-Gopsill, Amirsadeghi, Earl, Jones, Lukens, Lee and Swanton2019) observed a 12% and 11% decrease in the chlorophyll precursor chlorophyllide a and total chlorophyll, respectively, which was exaggerated following a 6-h dark incubation. In addition, we reported an increase in the ratio of protochlorophyllide a to chlorophyllide a (48%) and chlorophyll (58%), along with a decline in leaf carotenoids (39%). This suggests that an early consequence of low R:FR reflected from neighboring weeds is decreased photoconversion of protochlorophyllide a to chlorophyllide a and chlorophyll. Protochlorophyllide is a potent photosensitizer and forms toxic free radicals upon exposure to light (Pattanayak and Tripathy Reference Pattanayak and Tripathy2011), thereby compromising photosynthetic potential. Indeed, this response was accompanied by a decline in leaf-level CO2 assimilation despite increased leaf internal CO2 and stomatal conductance (McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Amirsadeghi, Earl, Jones, Lukens, Lee and Swanton2019). Similarly, McKenzie-Gopsill et al. (Reference McKenzie-Gopsill, Amirsadeghi, Fillmore and Swanton2020) demonstrated that weed competition suppressed CO2 assimilation in common bean throughout early growth stages in the absence of direct shading. Removal of weeds prior to the R1 stage allowed for recovery of CO2 assimilation rate. Persistent weed competition into reproductive stages, however, led to consistent and irreversible declines in CO2 assimilation rate and photosynthetic efficiency. These declines were associated with reduced water use efficiency and crop yield. Importantly, limitations on photosynthetic efficiency were primarily related to reduced carboxylation rate of Rubisco (V cmax ) rather than reduced light interception or stomatal conductance. In contrast, other studies have noted declines in CO2 assimilation were accompanied by restricted stomatal conductance, highlighting the species-dependent nature of this response (Li et al. Reference Li, Hamani, Si, Liang, Gao and Duan2020; Vysotskaya et al. Reference Vysotskaya, Wilkinson, Davies, Arkhipova and Kudoyarova2011). Taken together, these studies demonstrate that early season weed competition during the CPWC can induce persistent biochemical limitations on photosynthesis, linking induction of the shade avoidance response to the CPWC and yield loss.
Intra-Specific Versus Inter-Specific Light Cues and Photosynthetic Adjustments
The identity of neighboring plants has a strong influence on its photosynthetic performance. Recent studies have begun to unravel how plants respond to different neighbors, adjusting leaf- and canopy-level photosynthesis in both competitive and facilitative interactions. Using plant simulation models, Zhang et al. (Reference Zhang, Van Westreenen, He, Evers, Anten and Marcelis2021) showed that upper shoots of roses (Rosa hybrida Vill.) respond to low R:FR reflected upward from their lower leaves by increasing leaf inclination angle and maximizing light interception and canopy-level photosynthesis. Notably, this response was observed only when R:FR was reflected off a plant’s own leaf surface and not that of a neighbor. In contrast, low R:FR reflected from a neighbor suppressed photosynthesis. Zhang et al. (Reference Zhang, Van Westreenen, He, Evers, Anten and Marcelis2021) proposed this to be a self-signaling response inducing canopy reorganization to minimize self-competition. Canopy reorganization to minimize self-competition has also been documented in the other species including sunflower (Helianthus annuus L.), Arabidopsis, and soybean (Crepy and Casal Reference Crepy and Casal2015; López Pereira et al. Reference López Pereira, Sadras, Batista, Casal and Hall2017; Murphy et al. Reference Murphy, Van Acker, Rajcan and Swanton2017). For example, López Pereira et al. (Reference López Pereira, Sadras, Batista, Casal and Hall2017) showed that in high-density sunflower stands, taller “pioneer” plants create a low R:FR gradient toward their neighbors, triggering adjacent plants to self-direct toward higher R:FR. This population-level reorganization of the canopy optimized canopy structure and improved light interception, leading to an increase of 19% to 47% in seed oil yield. Canopy reorganization was not induced by blue light, suggesting that it was not a phototropic response but rather one mediated by FR light. It remains to be understood how crops can distinguish between intraspecific versus interspecific neighbors and the possible interactions between light and chemical signals to optimize leaf- and canopy-level photosynthesis. To date, this continues to be an unexplored area of improving crop productivity, tolerance to high-density planting, and improved competitive ability against weeds.
Low R:FR Light Alters Reactive Oxygen Species Homeostasis
Recent studies on soybean and Arabidopsis (Berardi et al. Reference Berardi, Amirsadeghi and Swanton2024; McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Amirsadeghi, Earl, Jones, Lukens, Lee and Swanton2019) indicate that alterations in reactive oxygen species (ROS) homeostasis are an early consequence of the perception of low R:FR light reflected from neighboring weeds. Increased production of H2O2 and singlet oxygen was also reported in soybean seedlings exposed to low R:FR light generated by neighboring weeds (McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Amirsadeghi, Earl, Jones, Lukens, Lee and Swanton2019). Low R:FR light generated artificially by FR light-emitting diodes or neighboring weeds can trigger the accumulation of singlet oxygen in Arabidopsis leaves (Berardi et al.Reference Berardi, Amirsadeghi and Swanton2024). This increased leaf production of singlet oxygen under resource-independent weed competition does not cause cell death but may act as a signaling molecule. Singlet oxygen production may activate transcription of genes that encode negative regulators of jasmonic acid level to suppress defense and promote the shade-avoidance response in response to neighboring weeds. The underlying mechanism of singlet oxygen accumulation under low R:FR light, however, remains unclear (Berardi et al. Reference Berardi, Amirsadeghi and Swanton2024). Earlier studies showed that exposure of isolated chloroplasts to FR light can lead to damage of PS I through superoxide generation, and PS II through both superoxide and hydrogen peroxide (Tjus et al. Reference Tjus, Scheller, Andersson and Møller2001). Under FR light, protochlorophyllide accumulation in Arabidopsis leaves can lead to singlet oxygen generation upon transfer to white light (Page et al. Reference Page, McCormac, Smith and Terry2017). To date, studies quantifying ROS under low R:FR light conditions provide no clear consensus on the extent or direction of ROS changes across plant species. Contrary to the expectation that shade might reduce oxidative by-products, a recent study found increases in ROS and nitric oxide accumulation in Arabidopsis hypocotyls under a canopy-like shade condition (Iglesias et al. Reference Iglesias, Costigliolo Rojas, Bianchimano, Legris, Schön, Gergoff Grozeff, Bartoli, Blázquez, Alabadí, Zurbriggen and Casal2024). These findings highlight the complexity of ROS-mediated regulation of shade avoidance in crop plants exposed to early season weed competition.
Effects of Low R:FR Light on Enzymatic Antioxidants Are Species-Specific and Condition-Dependent
Recent studies show that impacts on enzymatic antioxidants associated with the shade-avoidance response are inconsistent across crop species. For example, we demonstrated that low R:FR reflected from neighboring weeds decreased superoxide dismutase (SOD) activity, increased ascorbate peroxidase (APx) and glutathione peroxidase (GPx) activity, yet did not impact catalase (Cat) activity in soybean seedlings at the V1 growth stage (McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Amirsadeghi, Earl, Jones, Lukens, Lee and Swanton2019). In contrast, low R:FR light was associated with increased SOD activity, reduced Cat activity, and unchanged peroxidase (POD) activity in tomato (Cao et al. Reference Cao, Yu, Xu, Ai, Bao and Zou2018). Decreased SOD activity and increased Cat and POD activity has been reported in lettuce plants subjected to low R:FR light; however, this may be cultivar dependent (Jeong et al. Reference Jeong, Zhang, Niu and Zhen2024; Ju et al. Reference Ju, Zhang, Hu, Zhang, He, Li, Liu and Liu2024; Lee et al. Reference Lee, Xu, Wang and Rajashekar2019; Meng et al. Reference Meng, Song, Ni and Kamaruddin2024). It should also be noted that the transcript levels of SOD, POD, and Cat decreased in the leaves but increased in the roots of the C4 bermudagrass [Cynodon dactylon (L.) Pers.] subjected to shade (Wang et al. Reference Wang, Mao, Ji, Wang and Fu2024). Increases in SOD, glutathione reductase, and monodehydroascorbate reductase activities in tea (Camellia sinensis L.) leaves following transfer from shade to high light may help protect against oxidative damage under high light (Sano et al. Reference Sano, Takemoto, Ogihara, Suzuki, Masumura, Satoh, Takano, Mimura and Morita2020). Furthermore, proteomic analysis of corn leaves under shade tents providing 60% shade revealed increased expression of proteins involved in stress, defense, and detoxification (Gao et al. Reference Gao, Liu, Zhao, Liu and Zhang2020). In the latter two studies, however, the effect of shade treatments on the R:FR ratio of light under experimental conditions was not clearly specified. Overall, the effects of low R:FR and resource-independent competition on enzymatic antioxidants vary across plant species, preventing a clear consensus.
Effects of Low R:FR Light on Nonenzymatic Antioxidants Are Not Consistent Across Plant Species
In addition to impacts on enzymatic antioxidants, several recent studies have demonstrated a link between shade avoidance and plant redox regulation. Decreases in total ascorbic acid and increases in oxidized ascorbate dehyroascorbate has been reported in soybean subjected to low R:FR light reflected from neighboring weeds (McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Amirsadeghi, Earl, Jones, Lukens, Lee and Swanton2019). Low R:FR light led to a decrease in ascorbic acid content in bean leaves and was associated with reduced activity of ascorbic acid biosynthetic enzymes and glutathione level indicating a coordinated redox adjustment that results in lower antioxidant capacity (Bartoli et al. Reference Bartoli, Tambussi, Diego and Foyer2009). These responses may be specific to bean, since under low R:FR light, reduced and oxidized glutathione levels have been reported to be unchanged in Arabidopsis yet results are inconsistent in wheat and lettuce (Gasperl et al. 201; Toldi et al. Reference Toldi, Gyugos, Darkó, Szalai, Gulyás, Gierczik, Székely, Boldizsár, Gilaba, Müller and Simon-Sarkadi2019; Meng et al. Reference Meng, Song, Ni and Kamaruddin2024; Dou et al. Reference Dou, Li, Li, Song, Yang and Yan2024; Zou et al. Reference Zou, Fanourakis, Tsaniklidis, Woltering, Cheng and Li2023; Ju et al. Reference Ju, Zhang, Hu, Zhang, He, Li, Liu and Liu2024). These discrepancies highlight the differential responses in non-enzymatic antioxidants associated with the shade avoidance response across crop species and warrants further study for their potential for reducing crop stress.
Low R:FR Light Enhances Plant Susceptibility to Herbivory
A growing body of evidence indicates that low R:FR light can reduce plant resistance to herbivory, yet there remain limited examples linking early season crop-weed competition, the shade-avoidance response, and herbivory. Several studies, however, have demonstrated that low R:FR and induction of the shade-avoidance response is prioritized over plant insect- and disease-resistance responses. For example, exposure of Nicotiana longiflora Cav. to reflected FR light enhanced herbivory of tobacco hornworm (Manduca sexta L.) and suppressed the accumulation of phenolic compounds typically triggered by herbivory (Izaguirre et al. Reference Izaguirre, Mazza, Biondini, Baldwin and Ballaré2006). Similarly, induction of the shade-avoidance response in tomato plants lacking PhyB1-PhyB2 increased susceptibility to herbivory by southern armyworm (Spodoptera eridania Stoll) and bean thrip (Caliothrips pahseoli Hood), resulting in greater insect growth and higher levels of naturally chewing insect damage (Izaguirre et al. Reference Izaguirre, Mazza, Biondini, Baldwin and Ballaré2006). Cucumber seedlings (Cucumis sativus L.) expressing the shade-avoidance response displayed 93% more herbivory by striped cucumber beetle (Acalymma vittatum Fabricius) compared with the control supporting the prioritization of shade-avoidance response over defense against herbivory (McGuire and Agrawal Reference McGuire and Agrawal2005). Under simulated shade conditions, common lambsquarters (Chenopodium album L.) were taller and showed reduced resistance to the armyworm caterpillar (Spodoptera exigua Hübner) compared with plants grown in control light. Herbivory, however, led to a reduction in plant height across all light treatments, suggesting that the combined effects of shade and herbivore damage may constrain the subsequent development of adaptive shade-avoidance traits (Kurashige and Agrawal Reference Kurashige and Agrawal2005). Despite clear linkages between low R:FR, shade avoidance, and herbivory, there remains a gap in our understanding of how early season weed competition may influence crop tolerance to insect feeding. Anecdotal evidence indicates that the accumulation of phenolic compounds in soybean leaves is suppressed in response to low R:FR reflected from neighboring weeds, suggesting that early season weed competition may compromise soybean defense against herbivory (unpublished data). On the other hand, transcriptomic studies have indicated an upregulation of genes associated with salicylic acid-mediated and jasmonic acid-mediated defense signaling in corn leaves exposed to neighboring weeds under field conditions (Horvath et al. Reference Horvath, Bruggeman, Moriles-Miller, Anderson, Dogramaci, Scheffler, Hernandez, Foley and Clay2018). Further studies, however, are needed to determine whether a decrease in leaf phenolic content translates into increased insect feeding.
Low R:FR Light Compromises Pest and Disease Resistance Via Suppression of Salicylic Acid- and Jasmonic Acid-Mediated Defense Responses
Under low R:FR light conditions generated by neighboring weeds, the drive to capture light causes the benefits of shade avoidance to far exceed the drawbacks of compromised immunity to bacterial and fungal pathogens. This is exemplified in studies, where exposure to low R:FR light did not suppress the SAS response in Arabidopsis plants even after inoculation with either a biotrophic pathogen (Pseudomonas syringae van Hall.) or a necrotrophic pathogen (Botrytis cinerea Pers.). Instead, low R:FR light conditions enhanced susceptibility to both pathogens and significantly reduced the salicylic acid-dependent and jasmonic acid-dependent transcriptional defense responses in a FR light-dependent but phytochrome-independent manner (de Wit et al. Reference de Wit, Spoel, Sanchez-Perez, Gommers, Pieterse, Voesenek and Pierik2013). Exposure to low R:FR light significantly reduced Arabidopsis resistance to necrotrophic fungus grey mold (Botrytis cinerea Pers.). This reduced resistance was linked to a suppression of several jasmonic acid-mediated defense responses and accumulation of defense-related phenolic compounds (Cerrudo et al. Reference Cerrudo, Page, Tollenaar, Stewart and Swanton2012). It has been shown that the phytochrome signaling pathway is indispensable for triggering systemic acquired resistance, increasing salicylic acid levels, and activating salicylic acid-dependent defense responses in Arabidopsis against Pseudomonas syringae pv. maculicola (Griebel and Zeier Reference Griebel and Zeier2008).
Plants may use overlapping signaling components to respond to both neighboring plants and pathogens. In this context, the constitutive shade avoidance phenotype observed in the Arabidopsis csa1 mutant carrying a mutation in a TIR-NBS-LRR-type gene (Toll/Interleukin1 receptor-nucleotide binding site-leucine-rich repeat) was rescued by overexpressing RPS4, the closest homolog of CSA1. RPS4 encodes a resistance protein that provides defense against Pseudomonas syringae pv tomato strain DC3000 (Faigón-Soverna et al. Reference Faigón-Soverna, Harmon, Storani, Karayekov, Staneloni, Gassmann, Más, Casal, Kay and Yanovsky2006). These findings provide strong evidence that a TIR-NBS-LRR signaling protein can modulate both shade avoidance and bacterial pathogen defense, highlighting a shared or convergent signaling architecture between these two biological processes.
Integration of Light Signaling with Various Stresses Optimizes Plant Stress Responses
Collectively, the evidence demonstrates that the shade avoidance response induced by low R:FR and neighboring weeds, act as primary integrators of environmental information, interfacing with ROS dynamics, redox buffering capacity, and hormone cross-talk. The resulting phenotype reflects a coordinated reallocation of metabolic and signaling resources toward shade-avoidance growth over all else, underscoring the central role of light quality in dictating the balance between vegetative expansion and immunity. Despite growing evidence that competition cues affect ROS homeostasis, major knowledge gaps remain in understanding how early season weed competition affects crop plant herbivory and susceptibility to bacterial and fungal pathogens. These include an incomplete understanding of species- and context-dependent variation in nonenzymatic and enzymatic antioxidant activities as well as limited knowledge of the ecological consequences of competition-induced ROS shifts for plant fitness, survival, and disease resistance under realistic competitive environments. Resolving these knowledge gaps will provide a more mechanistic framework for understanding how competition cues during the CPWC modulate ROS metabolism and downstream signaling networks. Such insights will be essential in order to reduce the impact of weeds on crop productivity and yield.
Crop-Weed Competition Influences Nitrogen Metabolism
Changes in nitrogen uptake have been the focus of several competition studies (Ashton et al. Reference Ashton, Miller, Bowman and Suding2008; Dunbabin Reference Dunbabin2007; Miller et al. Reference Miller, Bowman and Suding2007; Paulding et al. Reference Paulding, Baker and Warren2010; Simon Reference Simon2023; Trinder et al. Reference Trinder, Brooker, Davidson and Robinson2021). Fewer studies however, have investigated nitrogen metabolism under competition. For example, transcriptomic analysis of corn leaves under weed stress revealed altered expression of genes involved in nitrate uptake, transport, and assimilation in plants grown under field conditions (Horvath et al. Reference Horvath, Bruggeman, Moriles-Miller, Anderson, Dogramaci, Scheffler, Hernandez, Foley and Clay2018). We have not detected changes in several key enzymes of the nitrogen metabolism pathway in either corn or soybean in response to competition cues and plentiful resources, including nitrate reductase (NR), nitrite reductase, and glutamine synthetase (McKenzie-Gopsill Reference McKenzie-Gopsill2016; Kramer et al. Reference Kramer, Amirsadeghi, McKenzie-Gopsill and Swanton2025). In contrast, interspecific competition between redroot pigweed and soybean under different nitrogen fertilizer treatments led to reduced levels of nitrate and ammonium, as well as decreased activities of NR, glutamine synthetase, and glutamate dehydrogenase (Jiang et al. Reference Jiang, Zhou, Lu, Li, Yang, Feike, Zhang, Guan, Zhao and Liu2020). In addition, the decreased performance of Arabidopsis under competition at high density was accompanied by upregulation of genes involved in ion transport in the roots and downregulation of genes involved in nitrate assimilation in the leaves. Such responses may contribute to mechanisms underlying nutrient competition under high-density conditions (Masclaux et al. Reference Masclaux, Bruessow, Schweizer, Gouhier-Darimont, Keller and Reymond2012). Additionally, a study investigating the relationship between NR activity in rice cultivars and their competitive ability found significant differences in NR activity depending on genotype, nitrogen source, and developmental stage (Ouko Reference Ouko2003). In this study, a competitive index was defined as the ratio of dry matter accumulation under monoculture to that under mixed culture. Interestingly, at higher external nitrate concentrations (e.g., 40 parts per million), NR activity was higher, yet the competitive index was lower.
Under controlled environmental conditions, supplemental FR light has been shown to increase leaf nitrate levels in spinach (Spinacia oleracea L.), corn, and soybean, while ammonium levels remained constant (Johnson et al. Reference Johnson, Langhans, Albright, Combs, Welch, Heller, Glahn, Wheeler and Goins1999; Kramer et al. Reference Kramer, Amirsadeghi, McKenzie-Gopsill and Swanton2025). Mechanistically, nitrate accumulation in corn was linked to a decrease in ferredoxin-dependent glutamate synthase, whereas the activities of other nitrate assimilation enzymes were unchanged. A similar increase in leaf nitrate levels was also observed in soybean seedlings exposed to surrounding six soybean seedlings of the same variety (kin plants), even when direct root contact was prevented (Kramer et al. Reference Kramer, Amirsadeghi, McKenzie-Gopsill and Swanton2025). These studies indicate that resource-dependent and resource-independent experimental conditions can differentially affect nitrate metabolism in plants under competitive stress. The latter study found several differentially expressed genes (DEGs) involved in nitrate metabolism and transport, highlighting the intricacies of corn responses to reflected low R:FR light from neighboring weeds under conditions without resource competition. No evidence, however, supported the involvement of tonoplast-localized nitrate transporter genes in nitrate accumulation in leaf tissue. Therefore, the authors of Kramer et al. (Reference Kramer, Amirsadeghi, McKenzie-Gopsill and Swanton2025) suggested that the effect of low R:FR light was more closely related to altered nitrate assimilation rather than to transporter upregulation or downregulation. The low R:FR light-mediated increase in leaf nitrate levels under resource-independent competition may not be limited to corn and soybean leaves, as our preliminary analyses indicate that similar increases also occur in their root tissues (unpublished data). Therefore, the role of nitrate transporters in such complex responses cannot be ruled out.
The regulation of nitrate transporters occurs through transcriptional and post-translational processes, as well as interactions with other proteins (Aluko et al. Reference Aluko, Kant, Adedire, Li, Yuan, Liu and Wang2023; Dölfors et al. Reference Dölfors, Ilbäck, Bejai, Fogelqvist and Dixelius2024; Jacquot et al. Reference Jacquot, Li, Gojon, Schulze and Lejay2017). Nitrate transporters are increasingly recognized as playing roles in both abiotic and biotic stress responses. Our literature search, however, found no published studies that explicitly examine how plant-plant competition or the effects plant neighbors influence nitrate transporter gene expression. Additionally, we found no studies that have investigated this under resource-independent competition, where low R:FR light is the major competitive cue. This research gap highlights the novelty and potential contribution of such studies that link interspecific competition with nitrate transporter gene regulation. Furthermore, research on competition typically focuses on resource capture and growth outcomes, overlooking potential shifts in nitrate metabolism at the enzymatic level. A direct link between competition-induced signaling and the coordinated regulation of nitrate transporters and assimilation enzymes remains largely unexplored. Addressing this gap would advance our understanding of how plants integrate environmental competition cues with internal nitrogen metabolism to optimize resource use under crowded conditions.
Competition Cues Affect Root Traits and Nitrogen Uptake
Aboveground plant competition cues shape root development, nitrogen acquisition, and metabolic regulation in plants. These cues, such as low R:FR light, anticipate shading and can modify root growth, nutrient allocation, and nitrate assimilation. Collectively, these responses illustrate how plants integrate light cues and nutrient metabolism to optimize nitrogen use and sustain performance under competitive environments. Competing species often modify their root architecture and adjust their preference for nitrogen forms, such as nitrate or ammonium, to reduce resource overlap and maintain uptake efficiency (Ashton et al. Reference Ashton, Miller, Bowman and Suding2008; Bueno et al. Reference Bueno, Greenfield, Pritsch, Schmidt and Simon2019; Feng et al. Reference Feng, Huang, Sun, Sun, Guan, Qi, Liu, Qu and Feng2025; Liu and Zhao Reference Liu and Zhao2023; Miller et al. Reference Miller, Bowman and Suding2007; Paulding et al. Reference Paulding, Baker and Warren2010; Raza et al. Reference Raza, Din, Zhiqi, Gul, Ur Rehman, Bukhari, Haider, Rahman, Liang, Luo, El Sabagh, Qin and Zhongming2023; Schofield et al. Reference Schofield, Rowntree, Paterson, Brewer, Price, Brearley and Brooker2019; Tahir et al. Reference Tahir, Wei, Liu, Li, Zhou, Kang, Jiang and Yan2023; Zhang et al. Reference Zhang, Yang, Zhang, Bezemer, Liang, Li and Li2023; Zheng et al. Reference Zheng, Zhou, Chen, Zhang, Du, Yang, Wang, Yang, Xiao, Li, Yang and Yong2022).
Other studies have noted that competition cues and low R:FR may also affect a plant’s preference for a nitrogen source. For example, an application of uncomposted cattle manure improved the competitive ability of a range of annual and perennial cover crops over weeds compared with application of synthetic fertilizer, leading to greater weed suppression. This effect was attributed to the different forms and delayed release of nitrogen from manure relative to synthetic fertilizer. This change in fertilizer source suppressed the emergence of nitrophilous species while enhancing the growth rate of cover crops (McKenzie-Gopsill et al. Reference McKenzie-Gopsill, Nyiraneza and Fillmore2024). In excised roots of two hygrophytic plant species, Carex thunbergii Steud. and Polygonum criopolitanum, competition for nitrate (NO3 −), the preferred form of nitrogen in monoculture, increased with neighbor density. While the fast-growing C. thunbergii maintained its nitrate preference, P. criopolitanum shifted toward greater uptake of ammonium (NH4 +) and glycine as neighbor density increased. This shift in preference for alternative nitrogen forms may serve as a mechanism facilitating plant coexistence under varying competitive conditions (Jia et al. Reference Jia, Huangfu and Hui2020). We must be cautious not to generalize these results to broader plant communities, because the use of excised roots in hydroponic assays may not fully reflect in situ nitrogen uptake dynamics (Jia et al. Reference Jia, Huangfu and Hui2020). This shift in nitrogen preference may also be an important factor influencing the invasion success of three invasive weed species, Xanthium strumarium L., Ambrosia trifida L., and Bidens frondosa L. against their respective phylogenetically related native species [Xanthium sibiricum Patrin ex Widder, Sigesbeckia glabrescens (Makino) Makino, and Bidens biternata (Lour.) Merr. & Sherff.]. The invasive species showed a stronger growth response to nitrate than to ammonium under mixed cultures, indicating a preference for nitrate, whereas the native species exhibited the opposite pattern. These findings suggest that more efficient use of soil nitrogen may explain the greater growth advantage of the invasive species over related native species when grown in mixed culture compared to monoculture (Feng et al. Reference Feng, Huang, Sun, Sun, Guan, Qi, Liu, Qu and Feng2025).
Changes to nitrogen uptake by crops in response to competition cues, including low R:FR light, may be due to overall changes in root morphology and development. In a study of eight grain crops, root traits associated with spatial efficiency (e.g., geotropism index, deflection index, and root branch angle) or nutrient uptake efficiency (e.g., phosphate uptake efficiency) were prioritized under weed-free environments. In contrast, under weedy environments, grain crops emphasized root branch spacing and the growth rate of root axes. These results suggest that grain crops optimize root traits based on the presence of neighboring weeds and that these traits could be used to improve weed competitiveness and yield in grain crops (Dunbabin Reference Dunbabin2007). A recent meta-analysis of plant responses to population density revealed that the negative effects of shading on plants outweigh those of nutrient depletion (Postma et al. Reference Postma, Hecht, Hikosaka, Nord, Pons and Poorter2021). Increases in corn planting density resulted in a corresponding increase in specific root length (total root length per root dry weight) at the flowering stage. This change in root morphology was a major factor contributing to resource competition, influencing both the relative intensity of competition and soil carbon and nitrogen turnover (Sun et al. Reference Sun, Zang, Splettstößer, Kumar, Xu, Kuzyakov and Pausch2021). An assessment of various factors affecting competition between two herbaceous perennials, orchardgrass (Dactylis glomerata L.) and narrowleaf plantain (Plantago lanceolata L.), indicated that focal species identity, nitrogen ion form, and the interaction between nitrogen ion form and neighbor nitrogen uptake consistently influenced both competitive nitrogen uptake and biomass production. In the absence of a competitor, there was a positive relationship between nitrogen uptake and biomass, whereas in the presence of a competitor, nitrogen uptake was significantly decreased, and this relationship was disrupted. Overall, the findings suggest that competitive outcomes may be shaped by a narrower set of mechanistic drivers than is often assumed, and that direct measures of resource capture may better disentangle the drivers of competition than biomass alone, which serves only as a proxy endpoint for competition outcomes (Trinder et al. Reference Trinder, Brooker, Davidson and Robinson2021).
Summary
Overall, we have highlighted that an intricate array of responses to weed competition cues affects the early development of crop plants during the CPWC (Figure 2). Maladaptive phenotypic responses, disrupted carbon allocation, and lasting biochemical constraints on CO2 assimilation, rapid alterations in ROS homeostasis and reduced crop immunity, and disruptions in nitrate assimilation, among other factors, disrupt crop metabolism and development. These accumulated effects alter a crop plant’s growth trajectory, ultimately reducing yield potential. It should be noted that responses to competition cues or low R:FR light are not controlled solely by the classical shade-avoidance pathway involving phytochrome, phytochrome-interacting factors, auxin, and growth-related genes, but also by epigenetic and RNA-mediated layers of regulation. In this regard, long noncoding RNAs (lncRNAs) have emerged as functional regulators of SAS (García-López et al. Reference García-López, Vélez-Ramírez, Gillmor and Fernandez-Valverde2024; Mammarella et al. Reference Mammarella, Lucero, Hussain, Muñoz-Lopez, Huang, Ferrero, Fernandez-Milmanda, Manavella, Benhamed, Crespi, Ballare, Gutiérrez Marcos, Cubas and Ariel2023), indicating that chromatin remodeling by noncoding transcription is part of the light-signaling and photomorphogenic network in plants. This epigenetic modulation of SAS by lncRNAs may provide a mechanism for plants to remember past light-exposure environments, potentially explaining variability in SAS across environments and over time. In this context, changes in lncRNAs during SAS may be disadvantageous to plants, especially in agricultural settings. Because lncRNAs fine-tune hormonal, transcriptional, and chromatin-level responses during SAS, maladaptive or exaggerated changes in their expression may have unfavorable consequences. These include unnecessary elongation growth and yield penalties associated with the critical period of weed competition, impaired recovery when weeds are controlled and canopy light conditions change, weakened immunity to diseases and insects, amplified plant-to-plant variability, and increased metabolic costs. Lastly, because lncRNAs link light cues to hormone regulation, chromatin remodeling, and downstream developmental programs, studies of plant competition and SAS should expand beyond conventional factors (e.g., transcription factors and hormone pathways) to include noncoding RNAs.
Far-red light reprograms plant physiology with persistent developmental consequences. Low R:FR light triggers the stress-avoidance syndrome (SAS) and reshapes multiple plant processes. Phytochrome-mediated hormonal rebalancing and transcriptional reprogramming redirect development toward elongation-driven competitive growth. When SAS is established by early emerging weeds during the critical period of weed control (CPWC), these effects may persist even after weed removal, potentially imposing yield penalties. It may also establish persistent transcriptional and metabolic states, generating developmental memory effects that further constrain yield potential later in the life cycle. Abbreviations: lncRNAs, long noncoding RNAs; R:FR: red:far red; ROS, reactive oxygen species.

Funding
This work was supported by the Natural Sciences and Engineering Research Council of Canada (file RGPIN-2022-03454; C. Swanton) and the University of Guelph’s Food from Thought research program, which is funded by the Canada First Research Excellence Fund.
Competing Interests
Authors declare they have no competing interests.
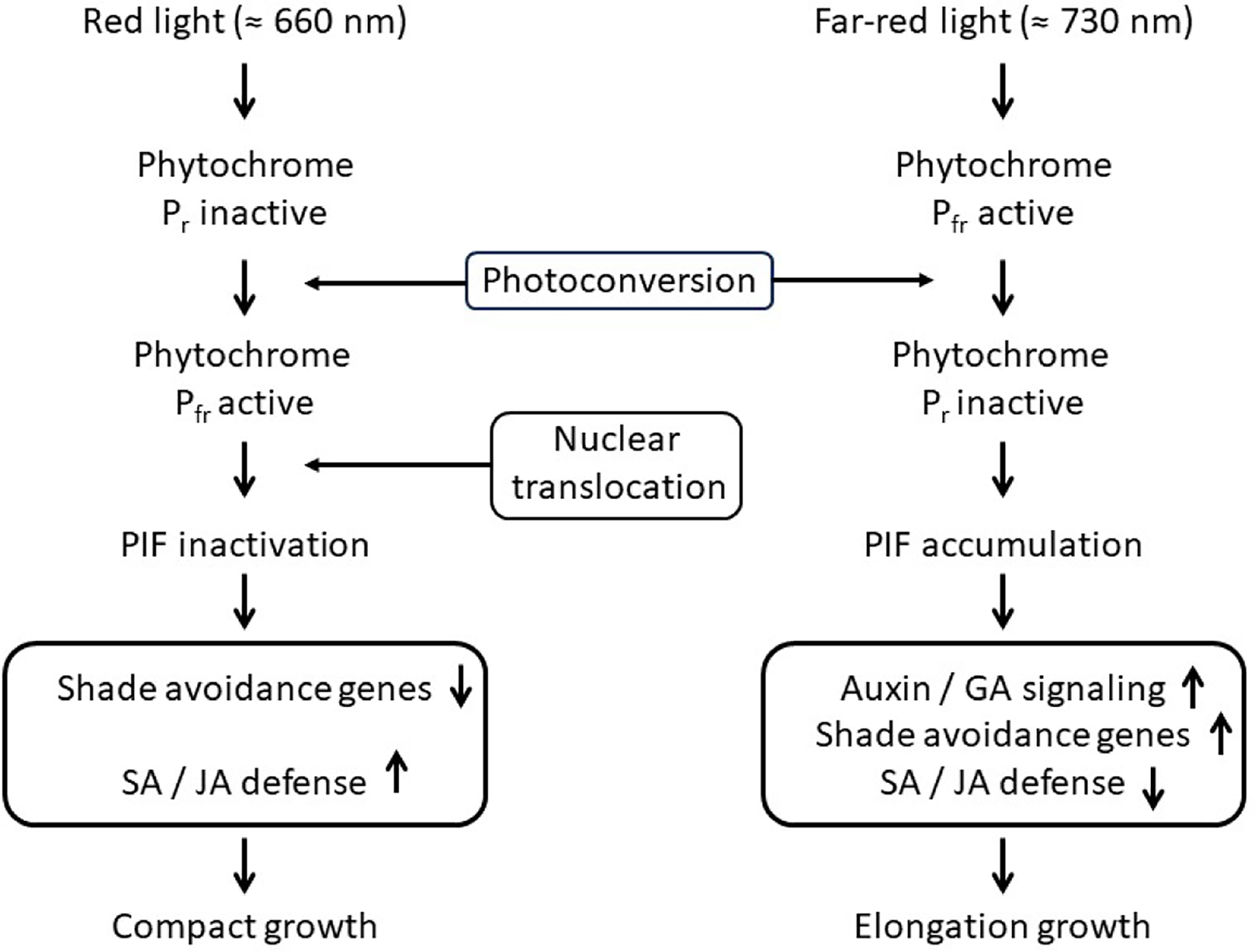
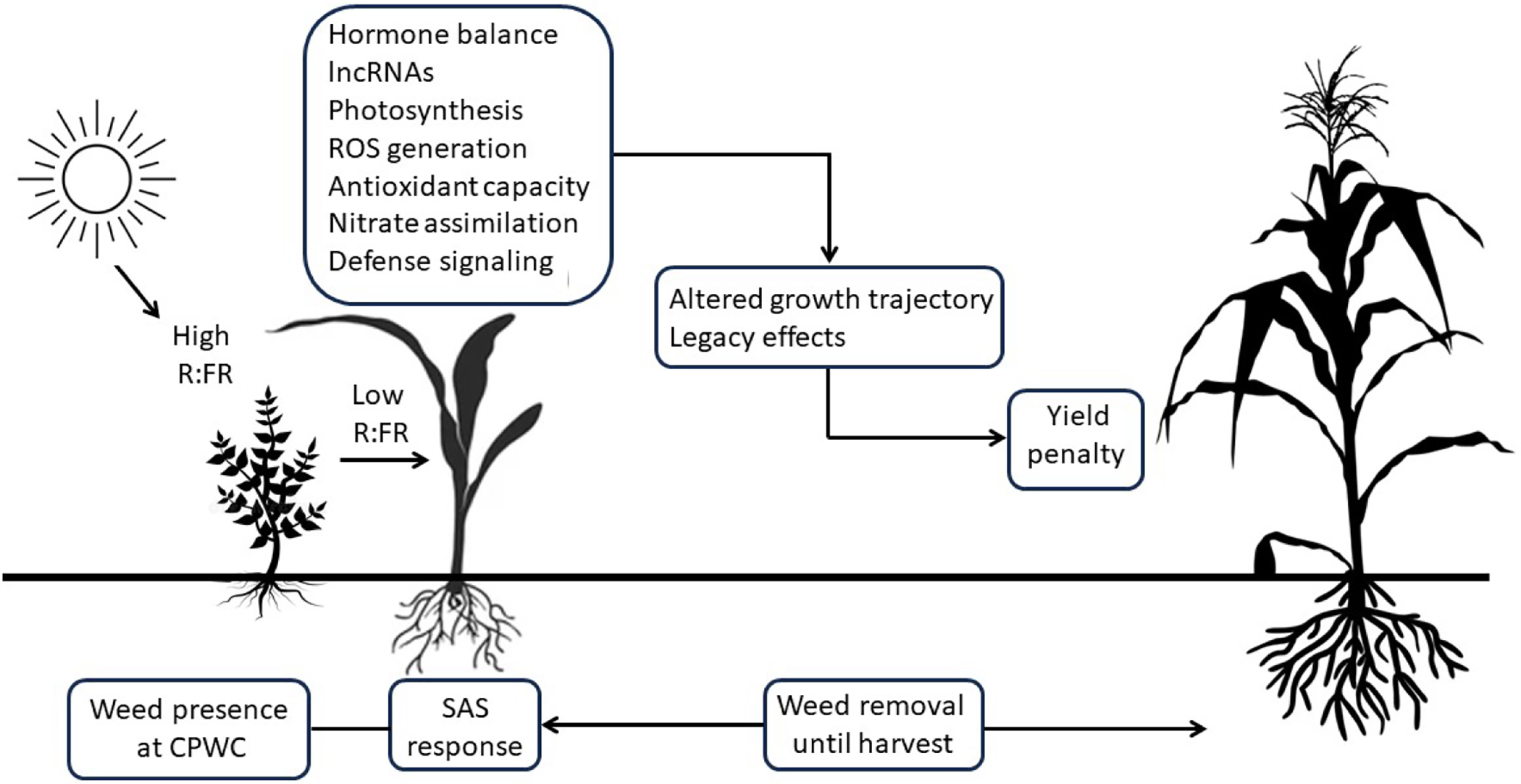

 Open access
Open access