


Introduction
Foliicolous lichens are an ecologically defined group of lichenized fungi that show strong preferences for living leaves as substrata (Santesson Reference Santesson1952; Sérusiaux Reference Sérusiaux1989, Reference Sérusiaux1996; Lücking Reference Lücking2001, Reference Lücking2008). Their highest diversity occurs in tropical and subtropical regions (Lücking Reference Lücking2003). While the distribution of foliicolous lichens is pantropical, they have been studied most extensively in neotropical rainforests (Lücking Reference Lücking2008). Foliicolous lichens exhibit relatively high diversity, with nearly 1000 species known worldwide (Lücking & Martínez-Colín Reference Lücking and Martínez-Colín2004; Lücking Reference Lücking2008; the present paper). However, recent studies show that lichen diversity has historically been underestimated and that further studies are needed to better understand the taxonomy of these organisms (Sipman & Aptroot Reference Sipman and Aptroot2001; Cáceres et al. Reference Cáceres, Lücking and Rambold2008). Pioneering molecular studies have shown a striking degree of hidden diversity in foliicolous lichens, such as in Ectolechiaceae, Gomphillaceae and Strigulaceae (Wang et al. Reference Wang, van den Boom, Sangvichien and Wei2020; Jiang et al. Reference Jiang, Lücking, Liu, Wei, Xavier-Leite, Portilla, Ren and Wei2022; Xavier-Leite et al. Reference Xavier-Leite, Cáceres, Aptroot, Moncada, Lücking and Goto2022; Lebreton et al. Reference Lebreton, Ertz, Lücking, Aptroot, Carriconde, Ah-Peng, Huang, Chen, Stenger and Cáceres2025). In addition to contributing to biodiversity assessment, foliicolous lichens can provide an insight into broader environmental patterns, such as altitudinal zonation, microclimatic conditions, and disturbance patterns of tropical ecosystems (Lücking Reference Lücking1995, Reference Lücking1997; Martínez-Colín et al. Reference Martínez-Colín, Lücking and Herrera-Campos2021).
Lichenological studies have been undertaken to a variable degree in most South-East Asian countries, including India, Thailand, Vietnam, Malaysia, Singapore, Indonesia, the Philippines, southern China and Hong Kong, Taiwan, and Papua New Guinea (Vězda Reference Vězda1977; Aptroot et al. Reference Aptroot, Diederich, Sérusiaux and Sipman1997; Aptroot & Seaward Reference Aptroot and Seaward1999; Wolseley et al. Reference Wolseley, Aguirre-Hudson and McCarthy2002; Aptroot & Sparrius Reference Aptroot and Sparrius2006; Singh & Sinha Reference Singh and Sinha2010; Sipman Reference Sipman2010; Schumm & Aptroot Reference Schumm and Aptroot2012a, Reference Schumm and Aptrootb ; Buaruang et al. Reference Buaruang, Boonpragob, Mongkolsuk, Sangvichien, Vongshewarat, Polyiam, Rangsiruji, Saipunkaew, Naksuwankul and Kalb2017; Joshi et al. Reference Joshi, Upreti and Hur2017; Hardini et al. Reference Hardini, Kasiamdari and Santosa2018; Sinha et al. Reference Sinha, Nayaka and Joseph2018; Jatnika et al. Reference Jatnika, Weerakoon, Arachchige, Noer, Voytsekhovich and Lücking2019; Kistenich et al. Reference Kistenich, Bendiksby, Vairappan, Weerakoon, Wijesundara, Wolseley and Timdal2019; Jannah et al. Reference Jannah, Hariri, Kasiamdari and Handayani2021). However, the lichen biota of Brunei Darussalam, a small country located in the north of the island of Borneo, is understudied. According to the Recent Literature on Lichens (RLL) database (https://nhm2.uio.no/botanisk/lav/RLL/RLL.HTM), there is only one known set of lichen collections from the country, most of these representing foliicolous lichens, with 27 species excluding the lichenicolous taxa (Aptroot et al. Reference Aptroot, Ferraro, Lai, Sipman and Sparrius2003). The Global Biodiversity Information Facility (GBIF) results show c. 40 records, mostly the same foliicolous lichens reported by Aptroot et al. (Reference Aptroot, Ferraro, Lai, Sipman and Sparrius2003), except Coenogonium hypophyllum, and almost all held at Meise Botanic Garden Herbarium (BR). There are no lichen collections in any of the three registered herbaria in Brunei (BRUN, IBER, UBDH), which limits further study of this group of organisms in the country. To improve this dire situation, we performed broad sampling of foliicolous and also corticolous lichens across the country. In this first contribution, we focus on the foliicolous lichen biota of Brunei Darussalam, presenting a detailed inventory across the country and its common habitat types.
Material and Methods
Study area
The country of Brunei Darussalam experiences a largely aseasonal tropical climate, with an average rainfall of 3386 mm y−1 and an average temperature of 27.7 °C (1991–2020 at 50 km × 50 km resolution) (Harris et al. Reference Harris, Osborn, Jones and Lister2020). The climate supports a high diversity and abundance of potential leaf substrata for foliicolous lichens, with 1407 tree species recorded within the country (BGCI 2025). The high tree diversity is found across the seven primary forest types that are recognized within Brunei Darussalam: the mixed dipterocarp, heath, mangrove, peat swamp, montane, beach-type, and freshwater swamp forests. These forests cover 72.1% of the land area according to Brunei Darussalam’s report to the United Nations Framework Convention on Climate Change (Brunei Darussalam Reference Brunei2022). To capture the foliicolous lichen diversity across the diverse landscapes for this study, we sampled 18 sites across all four geographical districts (Table 1, Fig. 1).
District, locality, forest type, latitude and longitude of all 18 lichen sampling sites within Brunei Darussalam, sorted by district, sampling strategy and site number. Opp = opportunistic; Qua = quantitative.
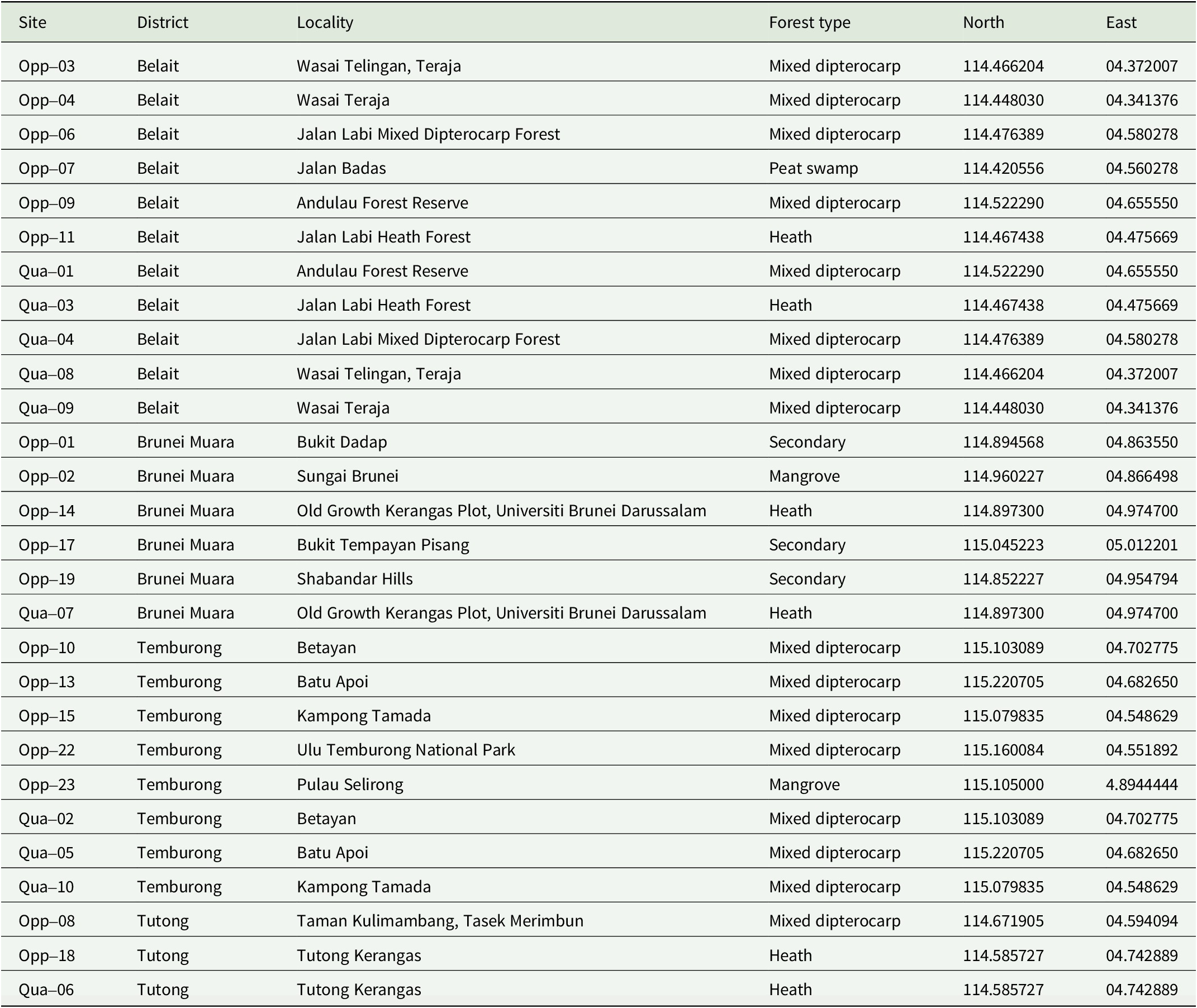
Map of the sampling sites within Brunei Darussalam. Each collection locality is indicated by a coloured shape corresponding to the four districts. The map was made using ArcGIS Pro 3.1. In colour online.
The inventory was performed using a combination of opportunistic and quantitative sampling to capture the highest possible species richness (Cáceres et al. Reference Cáceres, Lücking and Rambold2008). Sampling and identification work aimed to include rare, inconspicuous, sterile, and cryptic species, which are difficult to capture, yet important for accurately representing species richness (Cáceres et al. Reference Cáceres, Lücking and Rambold2008).
Taxonomic work
We separated and examined a total of 1763 specimens of foliicolous lichens from the collected material. In most cases, leaves were cut into smaller pieces which were curated separately to highlight particular taxa present on each piece. In selected cases, we kept entire leaves with lists of all species occurring on them, to document high levels of small-scale diversity. Duplicate sets are deposited at the Universiti Brunei Darussalam herbarium (UBDH) and the Herbarium Berolinense (B) of the Botanischer Garten und Botanisches Museum of the Freie Universität Berlin.
Specimens were identified by DAW and RL, with the assistance of BM, during a 4-week visit by DAW to the Botanischer Garten Berlin. For the assessment of morphological features, we used a LEICA Zoom 2000 stereomicroscope. Thin hand-cut sections were studied under a Zeiss Axioskop compound microscope, and all measurements were made on material mounted in water. For microchemical tests, we used Lugol’s solution (I), 10% KOH (K), commercial bleach (C) and paraphenylene-diamine crystals dissolved in ethanol (P). Macroscopic images were captured with a Nikon D5300 digital SLR camera using a Nikon AF-S DX Micro NIKKOR 40 mm macro lens with a Raynox digital camera macroscopic lens ×4 attached, and a Cullmann CUlight V 220DL LED light source. Microscopic images were taken under ×400 and ×1000 magnification with a Samsung Galaxy A71 mobile phone attached to the Pl ×10/20 ocular of the Zeiss Axioskop compound microscope.
For taxonomic identifications, we used keys published in various works (Santesson Reference Santesson1952; Sérusiaux Reference Sérusiaux1985, Reference Sérusiaux1996; Farkas & Vězda Reference Farkas and Vězda1987; Vězda Reference Vězda1987; Kalb & Vězda Reference Kalb and Vězda1994; Lücking & Vězda Reference Lücking and Vězda1998; Kalb et al. Reference Kalb, Lücking and Sérusiaux2000; Ferraro et al. Reference Ferraro, Lücking and Sérusiaux2001; Sérusiaux & Lücking Reference Sérusiaux and Lücking2003, Reference Sérusiaux and Lücking2007; Lücking Reference Lücking2004, Reference Lücking2008; Sparrius Reference Sparrius2004; Lücking et al. Reference Lücking, Thor, Aptroot, Kalb and Elix2006; Sérusiaux et al. Reference Sérusiaux, Lücking and Lumbsch2008; McCarthy Reference McCarthy2010; Flakus & Farkas Reference Flakus and Farkas2013; Neuwirth & Stocker-Wörgötter Reference Neuwirth and Stocker-Wörgötter2017; Naksuwankul & Lücking Reference Naksuwankul and Lücking2019; Jiang et al. Reference Jiang, Lücking, Xavier-Leite, Cáceres, Aptroot, Portilla and Wei2020, Reference Jiang, Lücking, Liu, Wei, Xavier-Leite, Portilla, Ren and Wei2022; Wang et al. Reference Wang, Sangvichien, Buaruang, Jiang, T-Z and Wei2021), as well as unpublished keys by RL. Where necessary, specimens were compared to photographed type material, either in the database of RL or on JSTOR Global Plants (https://plants.jstor.org).
Biogeographic analysis
To place the foliicolous lichen biota of Brunei into context, we established an updated global occurrence database of over 1000 foliicolous lichen species using Lücking (Reference Lücking2003) as a base, expanding it with published checklists and inventories of foliicolous lichens from 35 selected countries in the Neotropics, the African Paleotropics, and the eastern Paleotropics (Lücking Reference Lücking1999; Santesson & Lücking Reference Santesson and Lücking1999; Kalb et al. Reference Kalb, Lücking and Sérusiaux2000; Lücking & Kalb Reference Lücking and Kalb2000, Reference Lücking and Kalb2001; Ferraro et al. Reference Ferraro, Lücking and Sérusiaux2001; Lücking et al. Reference Lücking, Streimann and Elix2001; Ferraro Reference Ferraro2002; Ferraro & Lücking Reference Ferraro and Lücking2003; Aptroot et al. Reference Aptroot, Ferraro, Lai, Sipman and Sparrius2003; Farkas Reference Farkas2004, Reference Farkas2014, Reference Farkas2015a; Herrera-Campos et al. Reference Herrera-Campos, Lücking, Pérez-Pérez, Campos, Colín and Barcenas-Peña2004; Pinokiyo & Singh Reference Pinokiyo and Singh2006; Neuwirth Reference Neuwirth2007; Vězda Reference Vězda2007; Flakus & Lücking Reference Flakus and Lücking2008; Singh & Pinokiyo Reference Singh and Pinokiyo2008; Mateus et al. Reference Mateus, Lücking and Aguirre-C2011; Breuss Reference Breuss2012; Farkas et al. Reference Farkas, Elix and Flakus2012; Flakus Reference Flakus2013; Flakus & Farkas Reference Flakus and Farkas2013; Van den Broeck et al. Reference Van den Broeck, Lücking and Ertz2014; Farkas & Flakus Reference Farkas and Flakus2015; Gupta & Sinha Reference Gupta and Sinha2016; Jiang et al. Reference Jiang, Wei and Wei2017a, Reference Jiang, Wei and Weib , Reference Jiang, Lücking, Xavier-Leite, Cáceres, Aptroot, Portilla and Wei2020, Reference Jiang, Lücking, Liu, Wei, Xavier-Leite, Portilla, Ren and Wei2022).
The individual country biota was ordinated through a presence/absence data matrix using Principal Coordinate Analysis (PCoA) and cluster analysis. Jaccard’s distance measure was used to quantify dissimilarities with two dimensions retained (k = 2). Eigenvalues were extracted and plotted to evaluate the proportion of variation explained by each axis. The same Jaccard dissimilarity matrix was used for cluster analysis with the complete linkage method. Country biota were grouped by shape and colour into broad biogeographical regions (red circles represent Afrotropics, green triangles represent Asia, yellow squares represent Neotropics, and purple diamonds represent Oceania). The analyses were performed in R Studio v. 4.4.2 using the vegan package for ordination methods for community ecology, and the ggplot2, ggrepel and dendextend packages for visualization (Galili Reference Galili2015; Wickham Reference Wickham2016; Slowikowski Reference Slowikowski2024; Oksanen et al. Reference Oksanen, Simpson, Blanchet, Kindt, Legendre, Minchin, O’Hara, Solymos, Stevens and Szoecs2022; R Core Team 2024).
Results and Discussion
We identified a total of 193 species in 55 genera and 17 families in the material studied (Table 2; Supplementary Material File S1). This includes 21 species described below as new to science, in the genera Arthonia, Byssolecania, Byssoloma, Chroodiscus, Echinoplaca, Eremothecella, Fellhanera, Mazosia, Ocellularia, Opegrapha, Phylloporis, Porina, Spinomyces and Trichothelium, and three new continental records: Bapalmuia pallescens (Paleotropics), Lyromma palmae (Paleotropics) and Tapellaria major (Paleotropics).
List of species of foliicolous lichens found in the studied material from Brunei Darussalam. Taxa in bold are newly described in the present paper.

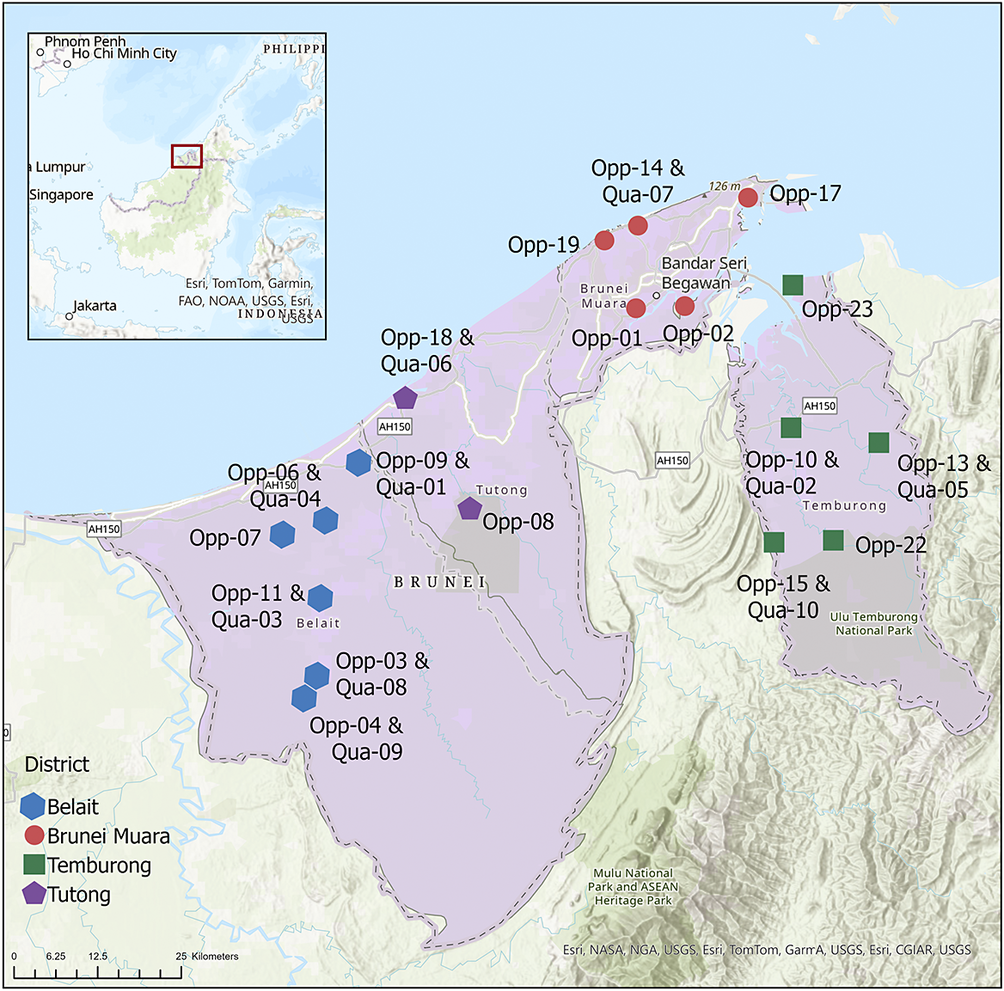
Of the 27 taxa reported by Aptroot et al. (Reference Aptroot, Ferraro, Lai, Sipman and Sparrius2003), the following six were not found in the present material: Chroodiscus mirificus (Kremp.) R. Sant., C. aff. mirificus (with larger ascospores), Loflammia epiphylla (Fée) Lücking & Vězda, Opegrapha cf. lambinonii Sérus., Porina aff. lucida R. Sant. (with longer, 13-septate ascospores) and Spinomyces albostrigosus (R. Sant.) Xavier-Leite et al.. The total number of foliicolous species known from Brunei is thus raised from 27 to 199, an increase of 172 taxa. Higher numbers are known only from Brazil (455 species), Costa Rica (435), Ecuador (365), Mexico (340), Guyana (296), Australia (238), Bolivia and Colombia (228 each), and Papua New Guinea (213). However, due to its small size, Brunei currently has the highest species density value relative to absolute area, with over 34 species per km2, four times higher than Costa Rica, and the sixth highest based on log(area) (Table 3).
List of the 15 countries with the highest known richness of foliicolous lichens.
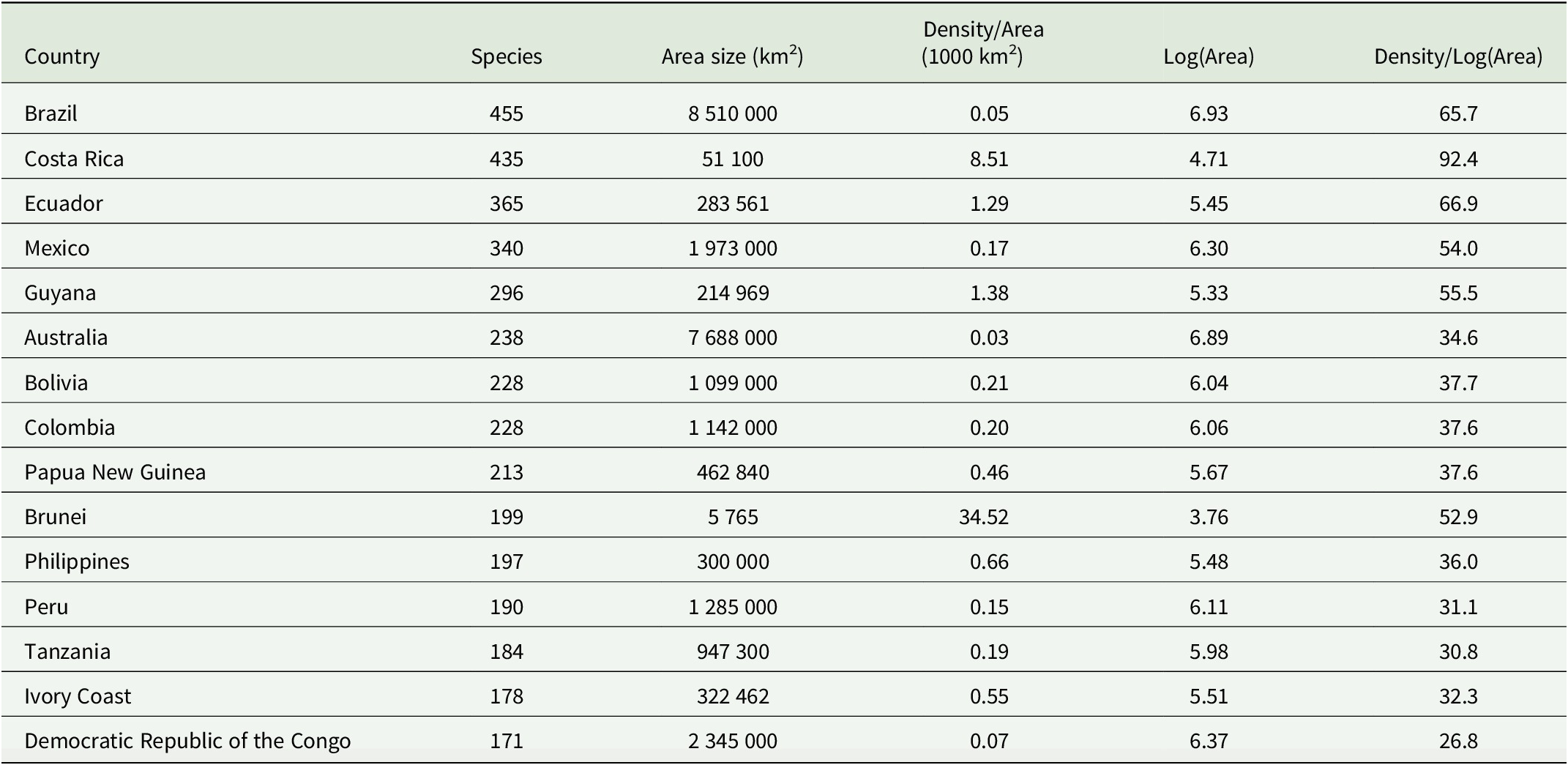
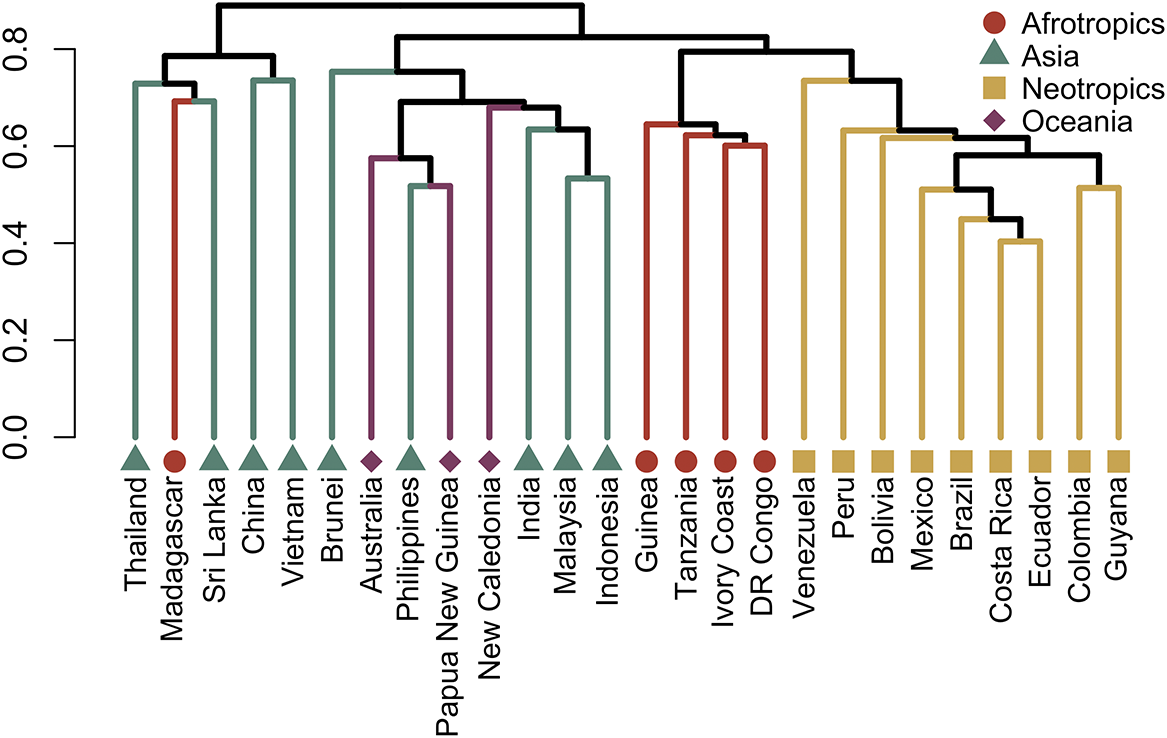
The foliicolous lichen biota of Brunei has a strong affinity with other paleotropical regions, clustering within a group of countries representing insular South-East Asia and Australasia (Oceania), specifically the Philippines, Indonesia, Papua New Guinea and Australia, with somewhat less affinity to continental South-East Asia (Figs 2 & 3). With the exception of Madagascar, the foliicolous lichen biota of selected tropical countries form three distinct clusters that represent the Neotropics, Afrotropics and eastern Paleotropics (Asia and Oceania) (Fig. 3). Despite its geographical proximity to Africa, Madagascar clustered with Sri Lanka and Vietnam, which is a pattern that was similarly found in tropical tree genera and may be driven by shared climates (Slik et al. Reference Slik, Arroyo-Rodríguez, Aiba, Alvarez-Loayza, Alves, Ashton, Balvanera, Bastian, Bellingham and van den Berg2015). Vietnam did not cluster tightly with Indonesia, Malaysia and Thailand, despite being geographically close. This separation may be related to Vietnam’s slightly differing climate and substrata, which are related to the geological history and location of the country. These differences can be summarized by Vietnam’s mostly Indochinese floristic signature with temperate elements, in comparison to the Malesian-dominated affinities of Thailand, Malaysia, and Indonesia.
PCoA ordination of selected tropical countries based on similarities in their foliicolous lichen biota. In colour online.
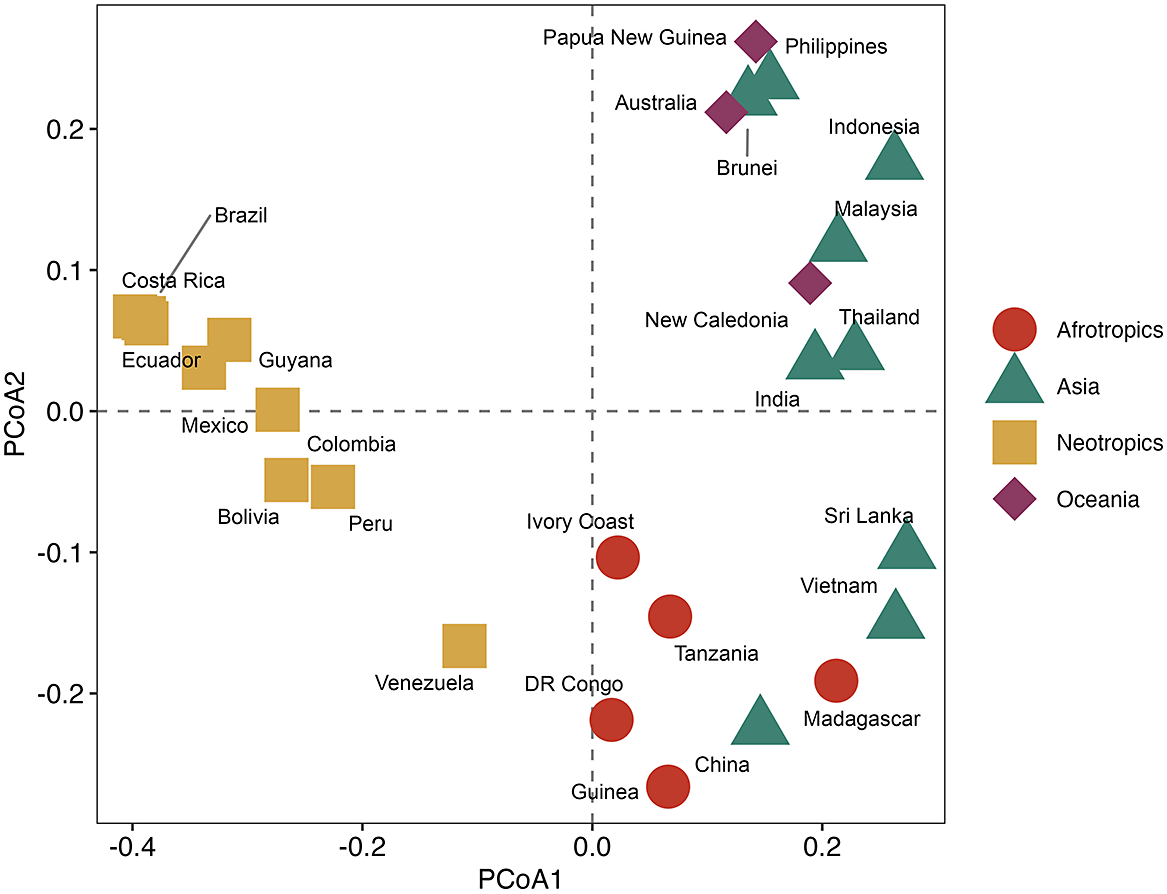
Cluster dendrogram of selected tropical countries based on similarities in their foliicolous lichen biota. The Neotropics, Afrotropics, and Eastern Paleotropics (Asia, Oceania) form three separate clusters, with the exception of Madagascar, clustering with Sri Lanka and Vietnam. Brunei forms part of a cluster including mostly insular South-East Asia and Oceania. In colour online.
Taxa identified as characteristic elements of the Paleotropics include, among others: Arthonia ramosii, Badimia cateilea and B. elegans, Byssoloma annuum, Chroodiscus argillaceus, C. defectus, C. homchantarae and C. verrucosus, Echinoplaca incrustaticiliata, Eremothecella macrocephala, E. macrosperma and E. variratae, Eugeniella micrommata, Fellhanera albidocincta and F. mastothallina, Gallaicolichen pacificus, Lasioloma phycophilum and L. phycophorum, Mazosia aptrootii, Musaespora coccinea and M. epiphylla, Phyllobathelium nigrum, Phylloblastia dolichospora, Phyllocraterina nuda, Porina applanata, P. cupreola, P. monocarpa, P. multiloculata, P. vanuatuensis and P. virescens, Raciborskiella talaumae, Rubrotricha helminthospora, Semigyalecta paradoxa, Tricharia novoguineensis and Trichothelium assurgens.
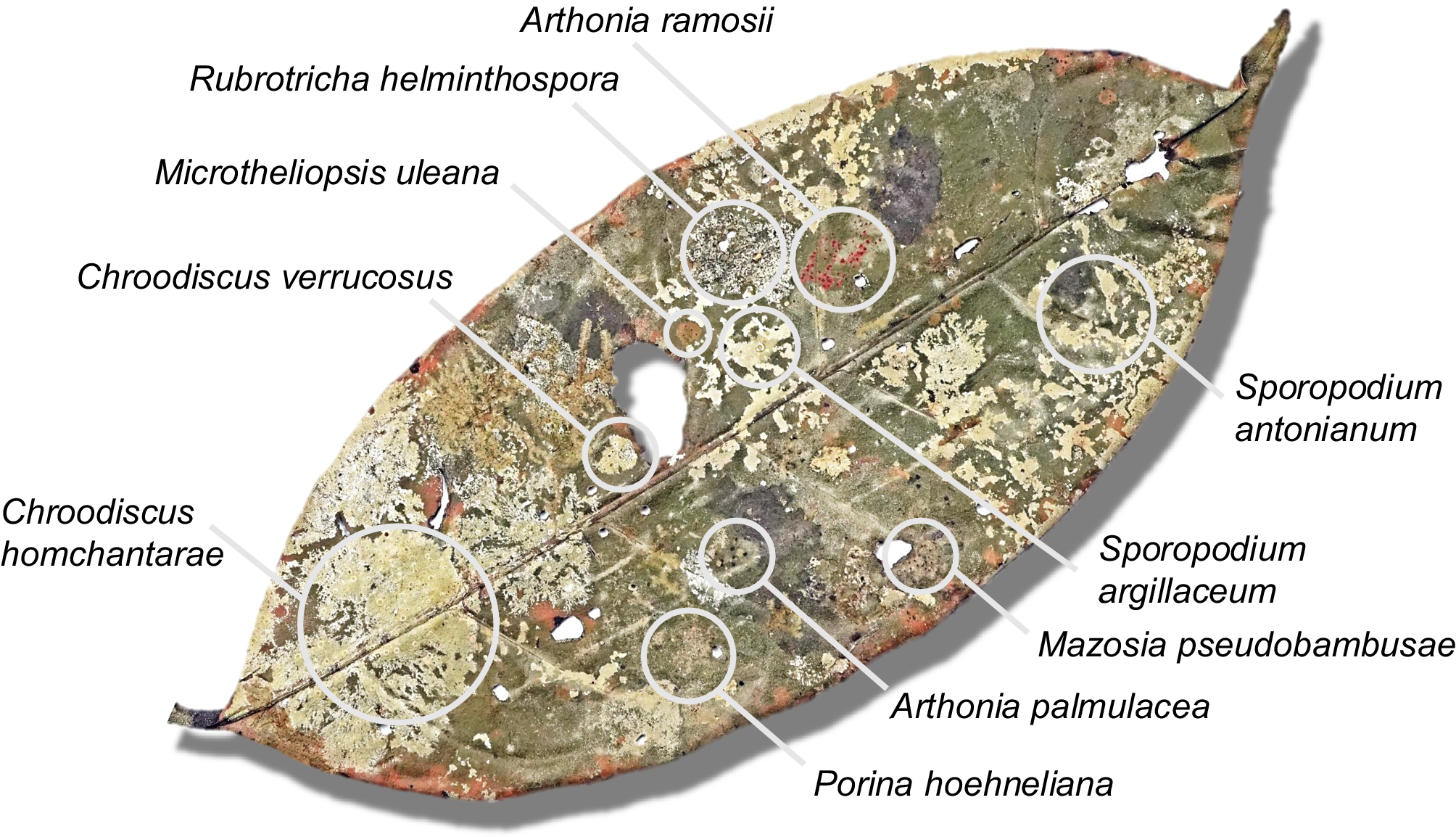
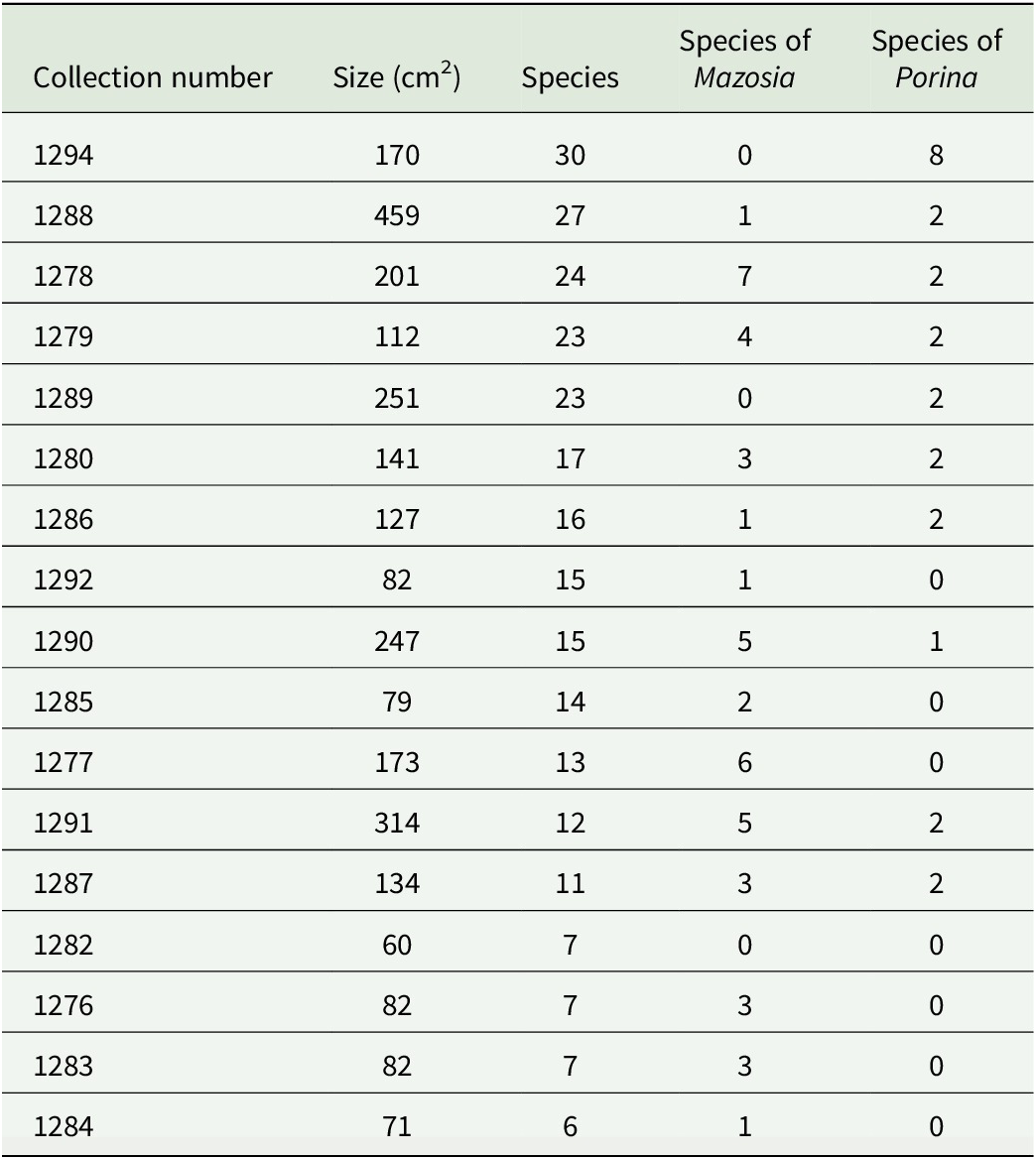
Selected individual leaves revealed between six and 30 species per leaf, with leaf sizes ranging from 60 to 459 cm2 (Fig. 4, Table 4). While not as high as the current records of comparably sized leaves (125 and 98 cm2) from Costa Rica and Ecuador, with 49 and 46 species, respectively (Lücking & Matzer Reference Lücking and Matzer2001), such high small-scale diversity is still remarkable. Our data also support the notion that closely related species (of the same genus) frequently grow side-by-side, demonstrating a high level of ecological equivalence. In the present material, this was observed particularly for species in the genera Mazosia, with up to seven species (out of a total of 24) on a single leaf (1278); in the case of the overall much more speciose genus Porina, only one leaf (1294) was found with a high number of species, namely eight out of 30 (Table 4).
Selected entire leaf (1292; see Table 4) from the material studied from Brunei Darussalam with 15 different foliicolous lichen species, some of them mapped. In colour online.
Number of foliicolous species found on selected leaves in the material studied from Brunei Darussalam.
Taxonomic Treatment
New or otherwise noteworthy taxa
Arthonia concentrica D. A. Ward, B. Moncada & Lücking sp. nov.
Index Fungorum No.: IF 905094
Differing from Arthonia trilocularis in the circular, slightly glossy thallus with concentrically arranged, thinly pruinose ascomata.
Type: Brunei Darussalam, Belait, Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 September 2021, D. A. Ward 274 (UBDH—holotype!; B—isotype!).
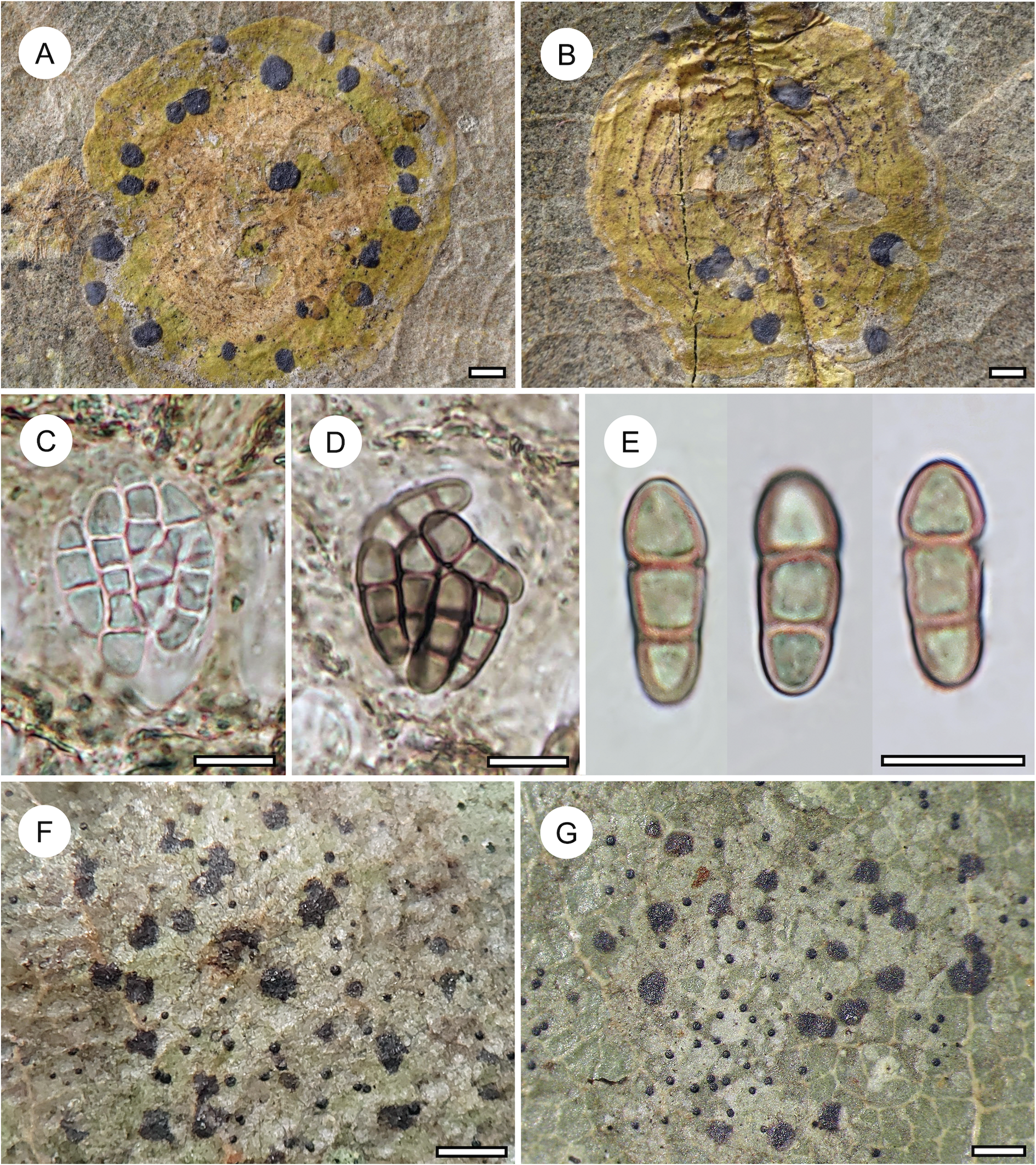
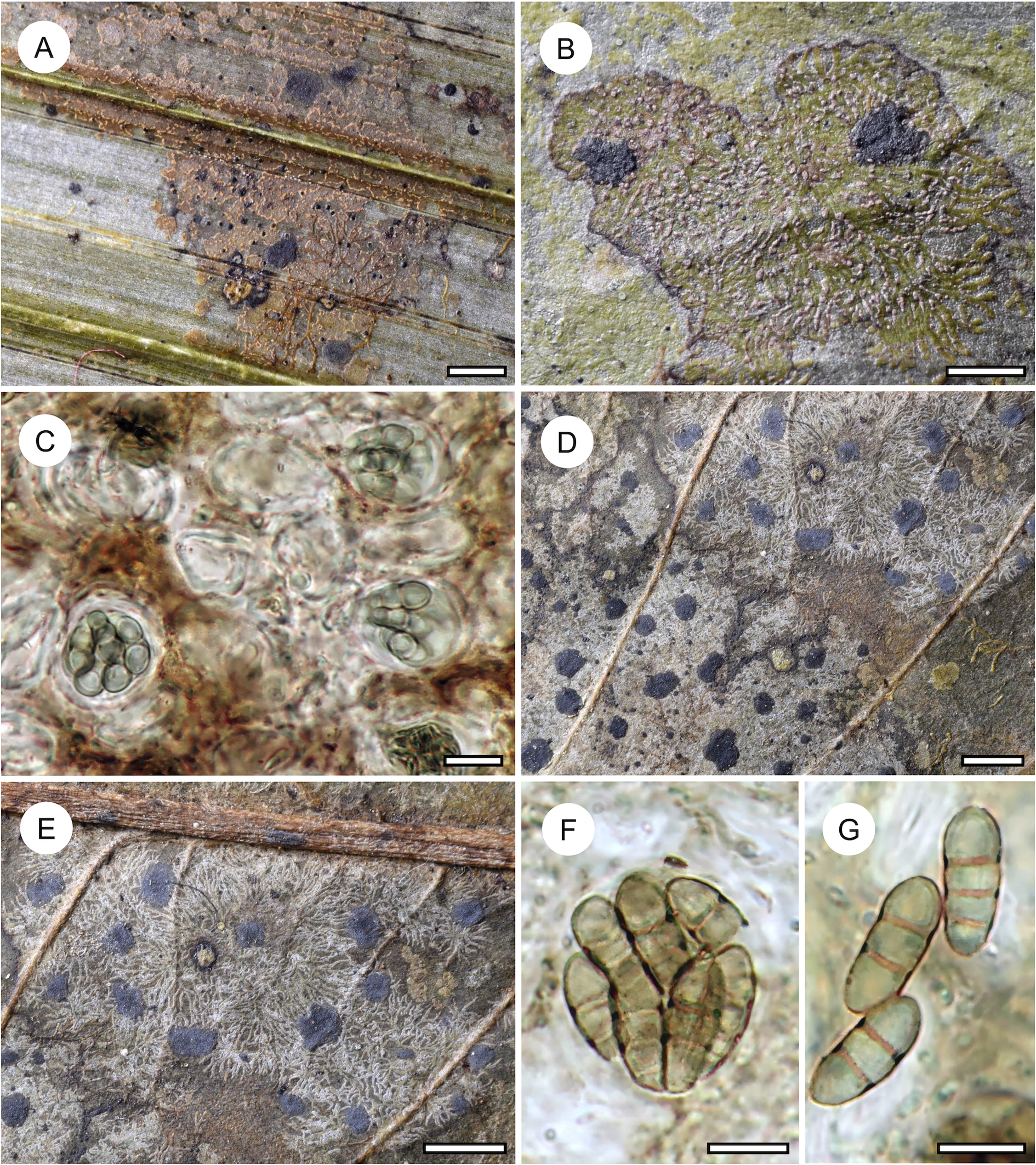
Habit and characteristics of newly described species. A–E, Arthonia concentrica (A & C–E, holotype; B, isotype). A & B, thalli with apothecia. C & D, asci with immature and mature ascospores. E, ascospores. F & G, Arthonia trilocularis for comparison (F, Brunei, Ward 620a; G, Seychelles, Norkett 17868A). Scales: A, B, F & G = 1 mm; C–E = 10 μm. In colour online.
Thallus continuous, very regularly rounded, smooth, 7–15 mm across, very thin (15–20 μm), ecorticate, yellowish brown to olive-brown, slightly glossy. Photobiont cells rectangular, 10–18 × 5–7 μm, in closed, regularly radiating plates.
Ascomata not or hardly raised above thallus level, conspicuously arranged in concentric fashion towards the periphery of the thallus, rounded to slightly irregular, 0.5–1.0 mm diam. and 30–40 μm high, greyish brown, appearing thinly purplish to brownish pruinose. Hypothecium inconspicuous. Epithecium formed by dense layer of parallel, periclinal hyphae, 3–5 μm high, colourless to pale brown, without any cover that would explain the pruinose appearance, K+ olive. Hymenium 30–40 μm high, colourless, I+ orange-red, KI+ pale bluish. Paraphyses branched and anastomosing. Asci broadly clavate to subglobose, 25–30 × 12–20 μm, I−, KI+ blue in inner parts of tholus. Ascospores 8 per ascus, drop-shaped, 2-septate, with the terminal cell enlarged, 12–15 × 4–5(–6) μm, grey-brown, I−.
Mature pycnidia not observed but one thallus had concentric rings of small, dark brown, hemispherical warts 0.05–0.07 mm diam., with a central pore visible under the microscope, that probably represent young pycnidia; the warts in section reacting K+ red.
Secondary chemistry
Not tested by TLC due to the very small thalli; standard spot tests on the thallus (C, K, P, UV) all negative; epithecium K+ olive, thallus warts (empty pycnidia?) K+ red.
Etymology
The epithet refers to the concentrically arranged ascomata.
Distribution and ecology
Known from a single locality of mixed dipterocarp forest in Brunei Darussalam.
Remarks
The genus Arthonia Ach. is a diverse element of foliicolous lichen communities, with 25 species currently accepted (Lücking Reference Lücking2008). Among these, there is a group of taxa with predominantly 2-septate, grey-brown ascospores, including A. flavoverrucosa, A. fuscocyanea U. Becker & Lücking, and A. trilocularis (Becker & Lücking Reference Becker, Lücking, Daniels, Schulz and Peine1995; Lücking Reference Lücking2008).
Arthonia concentrica is a further species with 2-septate, grey-brown ascospores. It is overall most similar to A. trilocularis (Lücking Reference Lücking2008), with which it shares the same ascospore type (and size range), but it differs in the larger, regularly rounded, continuous, brownish yellow, slightly glossy thallus patches, with the ascomata and pycnidia formed in concentric fashion towards the thallus periphery and producing a thin pruina. Arthonia trilocularis is abundantly present in the material studied here (Fig. 5F) and the specimens come close to the type material from Madagascar. This taxon has an irregularly dispersed, rather opaque thallus, with the individual patches usually below 1 mm diam. and the entire thallus not exceeding 5(–10) mm. The ascomata and pycnidia are dispersed over the entire thallus. The ascomata are brownish black with an irregular outline and lacking a pruina, except sometimes for the margins of the ascomata, and the pycnidia are flattened and black and do not display a K+ red reaction (see also Lücking Reference Lücking2008). We also compared numerous specimens of A. trilocularis from different regions of the Neotropics, Madagascar (the type), the Seychelles (Fig. 5G), Thailand, Australia, and New Caledonia, and could not find a morphology similar to A. concentrica. Arthonia trilocularis itself is probably a collective species that requires further study (Lücking Reference Lücking2008). Externally more similar to the new species is Arthonia accolens Stirt. (Lücking Reference Lücking2008), which often has a slightly glossy thallus but which differs in the lighter brown, non-pruinose ascomata and the colourless ascospores.
Arthonia flavirugosa D. A. Ward, B. Moncada & Lücking sp. nov.
Index Fungorum No.: IF 905095
Differing from Arthonia flavoverrucosa in the delicate, light yellowish ridges and the smaller ascospores.
Type: Brunei Darussalam, Belait, Jalan Labi Heath Forest, 4°28′32.4084″N, 114°28′2.7768″E, 51 m, tropical moist broadleaf forest ecoregion, heath forest, on palm leaves, 19 September 2021, D. A. Ward 765 (UBDH—holotype!; B—isotype!).
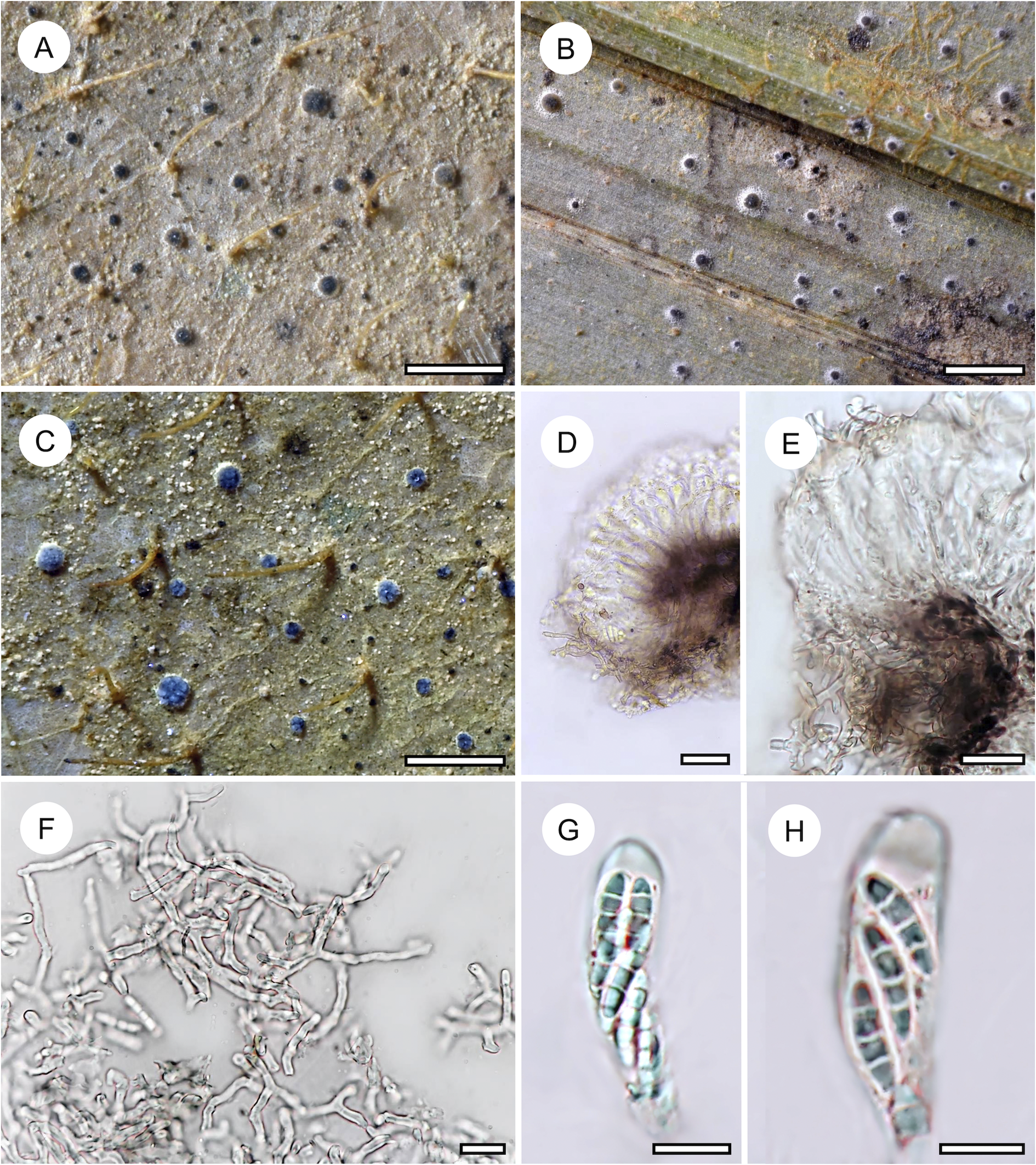
Habit and characteristics of newly described species. A–C, Arthonia flavirugosa (A & C, holotype; B, Ward 859). A & B, thalli with apothecia. C, asci with ascospores. D–G, Arthonia tenuilobata (holotype). D & E, thallus with apothecia. F, ascus with ascospores. G, ascospores. Scales: A, B, D & E = 1 mm; C, F & G = 10 μm. In colour online.
Thallus irregularly dispersed but individual patches connected by a translucent hypothallus delimited by a narrow, dark brown prothallus, finely rugose, 8–20 mm across, very thin (15–20 μm), ecorticate, light brown with a reddish tinge, opaque to slightly glossy; thallus patches furnished with delicate, irregular to radiating short ridges, often bordering the individual patches in concentric fashion, 0.1–0.3(–0.5) mm long and 50–80 μm wide, light yellowish, UV+ orange, in microscopic view filled with small, brownish crystals that quickly dissolve in K producing a light yellow cloud and retaining an orange colour. Photobiont cells oblong to rounded, 7–12 × 4–6 μm, in irregular plates leaving small interspaces.
Ascomata not raised above thallus level, rounded to elongate or irregularly angular, 0.4–0.6(–0.9) mm across and 40–45 μm high, dark greyish brown to brownish black, epruinose or thinly greyish pruinose. Hypothecium 5–7 μm high, colourless to brownish, K+ olive. Epithecium formed by a thin layer of parallel, periclinal hyphae, 3–5 μm high, brownish granulose, I−, the brownish granules dissolving in K forming a light yellow to greenish cloud, KI+ slightly olive. Hymenium 35–40 μm high, colourless, I+ orange, KI−. Paraphyses branched and anastomosing. Asci obovate to subglobose, 20–25 × 12–15 μm, I−, KI−. Ascospores 8 per ascus, drop-shaped, 2-septate, with the terminal cell enlarged, 10–12 × 3.5–4.5 μm, initially colourless but soon becoming grey-brown, I−.
Pycnidia rather frequent, applanately hemispherical, 0.1–0.15 mm diam., black, wall composed of dark brown, radiating hyphae. Conidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli; standard spot tests (C, K, P, UV) on the thallus negative, except for the ridges containing a brownish, crystalline pigment, with K+ light yellow efflux and retaining an orange colour, in surface view UV+ orange.
Etymology
The epithet refers to the light yellowish thallus ridges.
Distribution and ecology
Known from two localities of heath forest and mixed dipterocarp forest in Brunei Darussalam. The type collection on palm leaves is particularly plentiful.
Remarks
Arthonia flavirugosa is a second new species of foliicolous Arthonia with 2-septate, greyish brown ascospores discovered in material from Brunei, demonstrating a remarkable diversity of phenotypes in this group of species not recognized before. It differs from all other foliicolous species of Arthonia in the minutely rugose thallus with irregular to radiating or concentrically arranged, light yellowish ridges. This morphology is not entirely novel, as another species with thallus ornamentation, Arthonia flavoverrucosa, has previously been described from Côte d’Ivoire (Becker & Lücking Reference Becker, Lücking, Daniels, Schulz and Peine1995). Notably, that species was also found among the material from Brunei; it features pale yellow to orange-brown verrucae, much wider than the narrow ridges of A. flavirugosa. Its ascospores are also larger (12–16 × 4–6 μm) than those of the new species. A morphologically similar species with thallus ornamentation is Eremothecella rugosa, described below, which differs in the yellow-green thallus and the much larger, multiseptate, colourless ascospores.
We carefully evaluated the possibility that these cases could represent lichenicolous species of Arthonia inhabiting other lichen thalli, as such ornamentation is not known from corticolous species. If that were the case, the only candidates for potential host thalli having ornamentation and a Phycopeltis photobiont are in the genera Mazosia and Porina. However, no known species in these two genera has thalli with an ornamentation comparable to the three species mentioned above, including the material from Brunei studied here, so one would have to assume unknown host lichens. Also, lichenicolous species tend to have much smaller ascomata. If harmful to the host, they are usually limited to certain parts of the thallus which show signs of damage; if non-harmful parasymbionts, they are regularly dispersed on the host thallus and intermingled with the ascomata of the host lichen (Matzer Reference Matzer1996; Grube & Lücking Reference Grube and Lücking2002). Since none of these criteria apply to the material of the ornamented species of Arthonia and Eremothecella, there is no reason to assume they could represent lichenicolous species.
Additional specimen examined
Brunei Darussalam: Belait: Jalan Labi Mixed Dipterocarp Forest, 4°34′49″N, 114°28′35″E, 81 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 10 ix 2021, D. A. Ward 859 (B).
Arthonia tenuilobata D. A. Ward, B. Moncada & Lücking sp. nov.
Index Fungorum No.: IF 905096
Differing from Arthonia lobulicarpa in the more delicate lobes, pruinose ascomata and the brown ascospores.
Type: Brunei Darussalam, Belait, Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 September 2021, D. A. Ward 277a (UBDH—holotype!; B—isotype!).
Thallus continuous but finely dissected into thin lobes, smooth, 5–15 mm across, very thin (15–20 μm), ecorticate, yellowish brown, opaque to slightly glossy; lobes irregularly branched to radiating, 0.05–0.08 mm wide, in microscopic view composed of radiating and proliferating plates of photobiont cells connected by hyaline fungal hyphae. Photobiont cells rectangular, 8–15 × 3–5 μm, in closed, regularly radiating plates.
Ascomata not or hardly raised above thallus level, rounded, 0.3–0.6 mm diam. and 40–45 μm high, greyish brown, thinly brownish to greyish pruinose, giving the ascomata a somewhat bluish to purplish tinge. Hypothecium 5–7 μm high, brownish, I−, KI+ olive. Epithecium formed by a thin layer of parallel, periclinal hyphae, 3–5 μm high, colourless, I−, KI+ slightly olive. Hymenium 35–40 μm high, colourless, I+ light orange, KI+ light blue. Paraphyses branched and anastomosing. Asci obovate to subglobose, 25–30 × 12–15 μm, I−, KI−. Ascospores 8 per ascus, drop-shaped, 2-septate, with the terminal cell enlarged, 12–15 × 4–5 μm, for some time colourless but becoming grey-brown when mature, I−.
Pycnidia applanately conical, 0.15–0.2 mm diam., greyish black, wall composed of dark brown, parallel to radiating hyphae. Conidia narrowly oblong to fusiform with acute ends, non-septate, 10–15 × 2.5–3 μm, colourless.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) all negative or at best darkening.
Etymology
The epithet refers to the thin thallus lobes.
Distribution and ecology
Known from several localities of mixed dipterocarp (and riparian) forest in Brunei Darussalam.
Remarks
Arthonia tenuilobata represents a third new species with 2-septate, grey-brown ascospores. All hitherto known species in this group, including A. flavoverrucosa, A. fuscocyanea, A. trilocularis and the two new species described above, have continuous to dispersed thalli with rounded patches not dissected into lobes, with the exception of some specimens of A. trilocularis in which the thallus is partly dissected into irregular lobes (Lücking Reference Lücking2008). However, the apothecia of A. trilocularis lack a pruina giving them a bluish tinge, and the type of conidia found in the new species is not known from the latter. Arthonia fuscocyanea features ascomata with a pruina giving them a bluish tinge, but they have an angular shape and the thallus morphology is different.
Lobate thalli are otherwise found in Arthonia lobulicarpa U. Becker & Lücking, as in A. fuscocyanea described from tropical Africa (Becker & Lücking Reference Becker, Lücking, Daniels, Schulz and Peine1995). However, in that species the ascospores are colourless, the ascomata are non-pruinose and the thallus lobes are wider and shorter. Rather thin and often radiating thallus lobes may also be formed in Arthonia orbignyae (H. B. P. Upadhyay) Matzer (Lücking Reference Lücking2008), but that species has distinctly lirellate ascomata and the ascospores are also colourless. The ascomata of A. tenuilobata resemble those of A. cyanea Müll. Arg. in the bluish tinge, but the latter species has a different thallus morphology and its ascospores are colourless.
The thin and radiating lobes of Arthonia tenuilobata may resemble those of narrowly laciniate forms of Puigariella nemathora. In close-up, however, there are fundamental differences. The thallus of the latter grows beneath the leaf cuticle, with Cephaleuros as photobiont, and has convex lobules bearing numerous small, white papillae, whereas the thallus of Arthonia tenuilobata grows above the cuticle, with a Phycopeltis photobiont and completely flattened, thin lobes.
Additional specimen examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 262 (UBDH); Wasai Telingan, Teraja, 4°22′19.2252″N, 114°27′58.3344″E, 149 m, tropical moist broadleaf forest ecoregion, riparian forest/mixed dipterocarp forest, on leaves, 30 x 2021, D. A. Ward (with D. Cicuzza) 1102 (B, UBDH). Temburong: Kampong Tamada, 4°32′55.0644″N, 115°4′47.406″E, 48 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 13 xi 2021, D. A. Ward 592a (B).
Byssolecania fumosocaerulea D. A. Ward, B. Moncada & Lücking sp. nov.
Index Fungorum No.: IF 905097
Differing from Byssolecania fumosonigricans in greyish blue outer fringe of the apothecia.
Key to foliicolous species of Arthonia with 2(–3)-septate, grey-brown ascospores

Type: Brunei Darussalam, Belait, Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 September 2021, D. A. Ward 267 (UBDH—holotype!; B—isotype!).
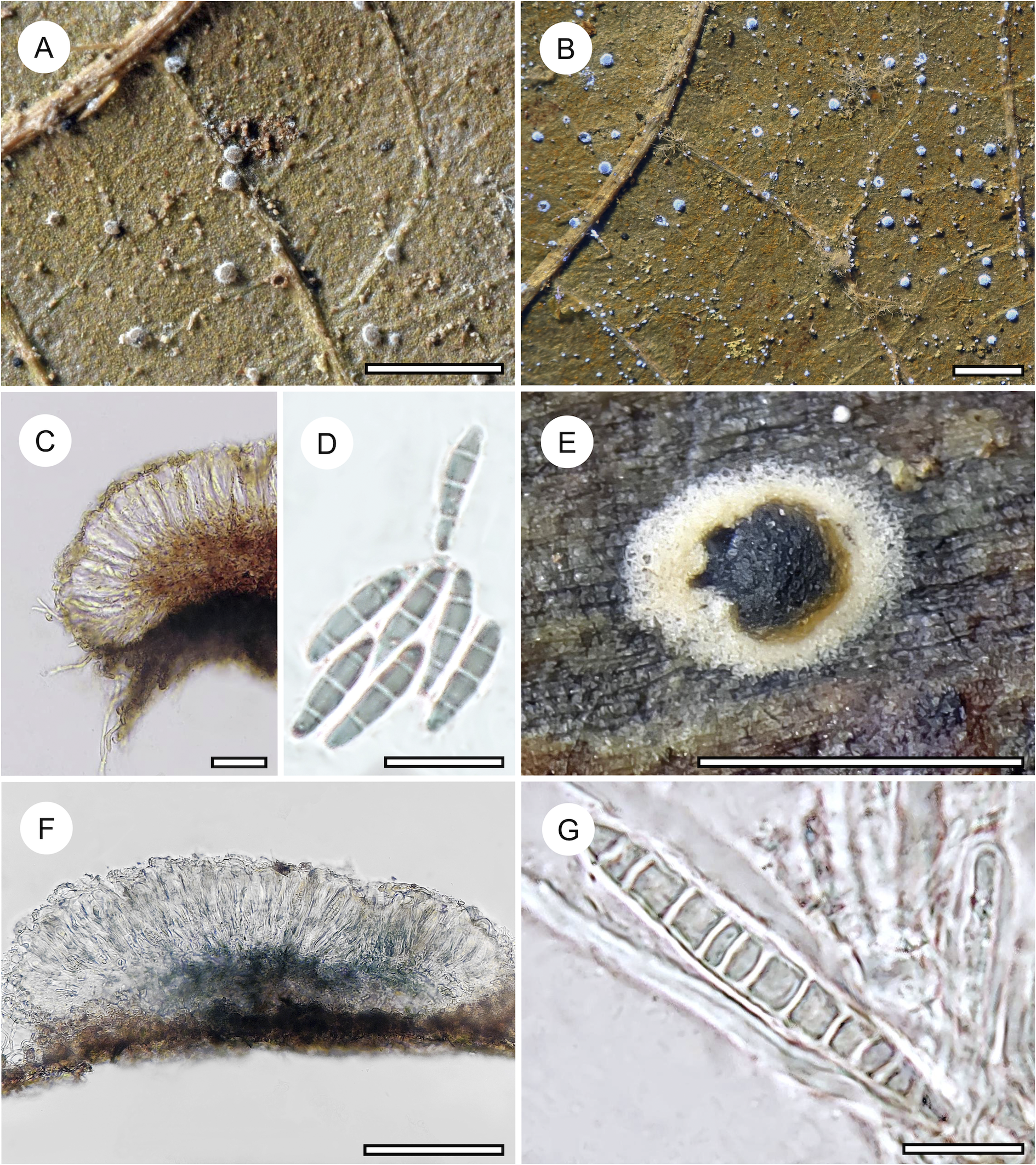
Habit and characteristics of newly described species. A–E, Byssolecania fumosocaerulea (holotype). A & B, thallus with apothecia. C & D, UV+ bright blue reaction of the apothecia. E, paraphyses seen from above. F, part of hymenium with ascus and ascospores. G, ascospore. Scales: A–D = 1 mm; E–G = 10 μm. In colour online.

Thallus continuous, irregular in outline, smooth, 10–30 mm across, very thin (15–25 μm), ecorticate, dark brown, opaque to slightly glossy. Photobiont cells bright green, rounded, 7–14 μm diam.
Ascomata not or hardly raised above thallus level, rounded to slightly irregular, 0.5–1.5 mm diam. and 30–40 μm high, light greyish brown in the centre but with a conspicuous, greyish blue rim around the centre, bordered by a thin, whitish margin, the entire ascoma UV+ bright blue. Hypothecium 5–10 μm high, dark aeruginous, I−, KI−. Epithecium indistinct. Hymenium 30–35 μm high, colourless, I+ blue, KI+ blue. Paraphyses unbranched, straight, rather thick (up to 3 μm) and densely arranged, embedded in a copious gelatinous matrix. Asci clavate, 28–33 × 8–10 μm, I+ blue, KI+ blue, with darker blue tubular structure in the tholus. Ascospores 8 per ascus, oblong-ellipsoid (to almost fusiform), sometimes with the upper median cell somewhat enlarged, 3-septate, without constrictions at the septa, 10–13 × 3–4 μm, colourless, I−, with irregular hyphae.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests on the thallus (C, K, P, UV) all negative. Apothecial disc UV+ bright blue.
Etymology
The epithet refers to the bluish hue of parts of the apothecial disc.
Distribution and ecology
Known from several localities of mixed dipterocarp forest in Brunei Darussalam.
Remarks
The genus Byssolecania is characterized by its flat (to slightly convex) apothecia resembling those of Arthonia but fundamentally differing from the latter in its internal anatomy, revealing it to be a member of the family Ectolechiaceae within the order Lecanorales (Lücking Reference Lücking2008). Santesson (Reference Santesson1952) originally distinguished only two species, based on ascospore septation alone. However, discrete differences in apothecial shape and colour have led to the separation of additional taxa (Lücking & Kalb Reference Lücking and Kalb2000; Herrera-Campos et al. Reference Herrera-Campos, Lücking, Pérez-Pérez, Campos, Colín and Barcenas-Peña2004; Breuss & Neuwirth Reference Breuss and Neuwirth2007; Lücking Reference Lücking2008). Thus far, seven species are recognized in this genus, differing mostly in the colour (and shape) of the apothecia and in ascospore septation. Among these, two species are characterized by completely flat apothecia and 3-septate ascospores, namely B. fumosonigricans (Müll. Arg.) R. Sant., with dark grey-brown apothecia and narrow whitish rim, and B. hymenocarpa (Vain.) Kalb et al., with light brown apothecia and wide whitish rim, typically partly detached from the thallus (Lücking Reference Lücking2008).
The new taxon comes close to B. fumosonigricans in the rather dark apothecia with narrow whitish rim, which rarely become partly detached from the thallus. However, it differs in the presence of a rather wide, greyish blue outer fringe of the apothecia, between the inner, greyish brown centre and the thin whitish margin. The colour variation is caused by the development of the apothecia starting centripetally from the margin. The thin whitish margin is caused by a hyphal excipulum composed of rather thick hyphae with rounded tips. These hyphae then transition into vertical paraphyses which grow vertically towards the centre, after which the asci are formed. The central portion containing asci is then seen as a grey-brown, somewhat irregular spot from above, whereas the fringe containing only paraphyses appears greyish blue, probably an effect of light reflection combined with the dark aeruginous hypothecium. We compared numerous specimens of B. fumosonigricans and never observed such a colour pattern.
The UV+ bright blue reaction of the apothecia came as a surprise. Upon checking, we found that other species of Byssolecania also display UV-fluorescence of the apothecia, usually bluish grey to greyish blue, but none as bright blue as in the new species.
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 466b (UBDH), D. A. Ward 470a (B); Wasai Teraja, 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 5 ix 2021, D. A. Ward (with D. Cicuzza and B. Zoletto) 179a (UBDH). Temburong: Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 ix 2021, D. A. Ward 934 (B, UBDH).
Byssoloma inconspicuum D. A. Ward, B. Moncada & Lücking sp. nov.
Index Fungorum No.: IF 905098
Differing from Byssoloma microcarpum in the dark brown apothecial disc and the excipulum lacking crystals.
Type: Brunei Darussalam, Belait, Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 September 2021, D. A. Ward 240a (UBDH—holotype!).
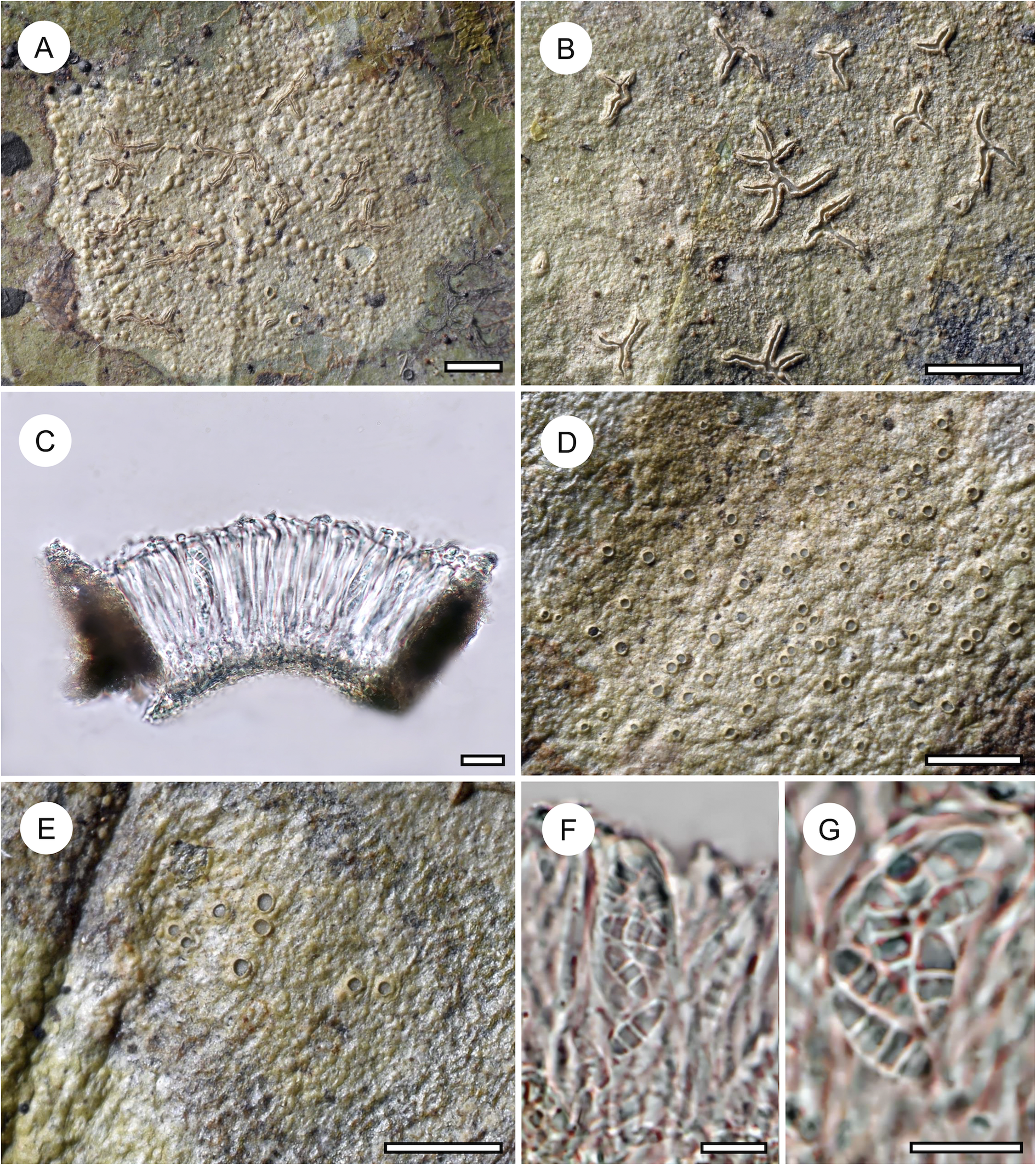
Habit and characteristics of newly described species. A–H, Byssoloma inconspicuum (A, C, D–H, holotype; B, Ward 771). A–C, thalli with apothecia, in C showing UV+ bluish reaction of the apothecial disc. D & E, section through apothecium. F, excipular hyphae. G & H, asci with ascospores. Scales: A–C = 1 mm; D & E = 20 μm; F–H = 10 μm. In colour online.
Thallus continuous, irregular in outline and overall inconspicuous or sometimes seemingly absent, in parts minutely farinose, 5–10 mm across, very thin (10–15 μm), ecorticate, light greyish to greyish brown, opaque, UV+ weakly light yellow; prothallus sometimes developed as thin, dark brown line. Photobiont cells green, rounded, 5–7 μm diam.
Ascomata biatorine with byssoid margin, sessile, rounded, 0.15–0.25 mm diam. and 120–150 μm high; disc dark brown to brownish black, UV+ bluish, margin cream-coloured to whitish, thin, not spreading laterally over the thallus surface; excipular hyphae hyaline, lacking crystals, 1.5–2 μm thick. Hypothecium 20–30 μm high, dark brown, K+ darker. Epithecium indistinct. Hymenium 50–60 μm high, colourless, I+ blue, KI+ blue. Paraphyses branched and somewhat anastomosing. Asci clavate, 40–50 × 8–12 μm, I+ blue, KI+ blue, with darker blue tubular structure in the tholus. Ascospores 8 per ascus, oblong, 3-septate, without constrictions at the septa, 10–13 × 2.5–3 μm, colourless, I−.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli; most standard spot tests on the thallus (C, K, P) all negative except a weak UV+ yellow reaction; hypothecium K+ darkening and apothecial disc UV+ bluish.
Etymology
The epithet refers to the inconspicuous thallus and apothecia.
Distribution and ecology
Known from several localities of mixed dipterocarp forest and heath forest in Brunei Darussalam; most abundantly found and best developed on hairy leaves.
Remarks
Byssoloma is one of the most speciose genera in Ectolechiaceae, with close to 70 species currently recognized (Lücking Reference Lücking2008; Wang et al. Reference Wang, van den Boom, Sangvichien and Wei2020; Miyazawa & Ohmura Reference Miyazawa and Ohmura2023). Delimitation of species, particularly among taxa with 3-septate ascospores and otherwise lacking readily diagnostic features, remains a challenge, especially in the B. leucoblepharum complex, but the limited molecular data available indicate that a high level of near-cryptic diversity may be present (Wang et al. Reference Wang, van den Boom, Sangvichien and Wei2020).
The new species is characterized by rather small apothecia with a thin, byssoid margin not spreading laterally over the thallus and lacking crystal inclusions. It could perhaps be considered a depauperate form of B. leucoblepharum or B. chlorinum s. lat. (Lücking Reference Lücking2008). However, the abundant material is very uniform and differs in several other characters from the latter two taxa: B. leucoblepharum has a smooth, dark green to bluish green thallus, whereas in B. chlorinum, the apothecial disc is typically almost black, creating a strong contrast with the whitish apothecial margin, and the thallus is well developed and light green. Other species that have small apothecia with a dark disc include B. diederichii Sérus., known from Macaronesia and western Europe (Sérusiaux Reference Sérusiaux1998), and B. microcarpum Kalb & Vězda, known from Malaysia and Papua New Guinea (Kalb & Vězda Reference Kalb and Vězda1994; Paukov et al. Reference Paukov, Sipman, Kukwa, Repin and Teptina2017). The first has even smaller apothecia with a bluish tinge and often produces bluish pycnidia with oval conidia, whereas the second has the byssoid excipulum encrusted with colourless crystals and the apothecial disc is blackish. It agrees with the new species in the bluish UV-reaction of the apothecial disc, but the thallus is much better developed and UV+ lemon yellow (see B. incrustatimurinum below for further discussion of UV-fluorescence in Byssoloma).
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 246 (B), 351 (B, UBDH); Jalan Labi Heath Forest, 4°28′32.4084″N, 114°28′2.7768″E, 51 m, tropical moist broadleaf forest ecoregion, heath forest, on leaves, 19 ix 2021, D. A. Ward 771 (UBDH), 1283e (B, filed under Mazosia melanophthalma); Wasai Teraja, 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 6 xi 2021, D. A. Ward (with D. Cicuzza) 1189 (B).
Byssoloma incrustatimurinum D. A. Ward, B. Moncada & Lücking sp. nov.
Index Fungorum No.: IF 905099
Differing from Byssoloma murinum in the shorter, non-tapering ascospores and the crystals in the excipulum.
Type: Brunei Darussalam, Tutong, Taman Kulimambang, Tasek Merimbun, 4°35′38.7384″N, 114°40′18.858″E, 31 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves (hypophyllous), 9 September 2021, D. A. Ward 221a (UBDH—holotype!; B—isotype!).
Habit and characteristics of newly described species. A–D, Byssoloma incrustatimurinum (holotype). A & B, thallus with apothecia, in B showing UV+ bright neon blue reaction of the apothecia. C, section through apothecium. D, ascospores. E–G, Byssoloma subaeruginosum (holotype). E, part of thallus with apothecium. F, section through apothecium showing aeruginous to smoky blue hypothecium. G, ascospore. Scales: A & B = 1 mm; C = 20 μm; D & G = 10 μm; E = 0.5 mm; F = 100 μm. In colour online.
Thallus hypophyllous, continuous, irregular in outline and overall inconspicuous and difficult to delimit, 5–10 mm across, very thin (5–15 μm), ecorticate, not distinguishable in colour from the lower leaf surface, opaque; prothallus absent or inconspicuous. Photobiont cells bright green, rounded, 8–13 μm diam.
Ascomata biatorine with byssoid margin, sessile, rounded, 0.15–0.2 mm diam. and 60–70 μm high; disc grey-brown, thinly whitish pruinose, margin whitish, thin, not spreading laterally over the thallus surface; margin and pruina of the disc UV+ very brightly neon blue. Hypothecium 20–30 μm high, brown to dark brown, K+ darker (blackish) brown, without reddish tinge. Epithecium indistinct, with scattered, colourless crystal granules, not dissolving in K. Hymenium 30–40 μm high, colourless, I+ blue, KI+ blue. Paraphyses branched and somewhat anastomosing. Asci clavate, 25–30 × 8–10 μm, I+ blue, KI+ blue, with darker blue tubular structure in the tholus. Ascospores 8 per ascus, oblong-ellipsoid, 3-septate, without constrictions at the septa, 9–12(–14) × 2–2.5 μm, colourless, I−.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) on the thallus all negative; apothecial margin and disc pruina with an unidentified, UV+ bright neon blue substance.
Etymology
The epithet refers to the excipulum encrusted with crystals, as compared to Byssoloma murinum Vězda.
Distribution and ecology
Growing hypophyllous on leaves of eudicots in the understorey of mixed dipterocarp forest; known from three localities in Brunei Darussalam.
Remarks
Byssoloma incrustatimurinum appears to be one of few genuinely hypophyllous species in this genus, together with B. farkasiae Sipman and B. hypophyllum Lücking & Kalb, both known from the Neotropics (Sipman Reference Sipman1992; Lücking & Kalb 2000; Lücking Reference Lücking2008), as well as B. murinum Vězda, described from tropical Africa (Vězda Reference Vězda1987). The first two differ in the persistently 1-septate ascospores and the larger apothecia, which are distinctly pruinose in B. farkasiae and non-pruinose in B. hypophyllum. Byssoloma murinum is morphologically and anatomically very similar to the material studied here and at first we considered both conspecific. However, the ascospores of the new taxon are consistently shorter (9–11(–13) μm vs (11–)13–16(–18) μm) and not tapering (as illustrated for B. murinum; Vězda Reference Vězda1987: fig. 1.1a), and the small crystals encrusting the hyphae of the excipulum were not reported or illustrated for B. murinum (Vězda Reference Vězda1987: fig. 1.1a). In the present taxon, the crystals are quite obvious under high magnification and it is unlikely that Vězda overlooked these in B. murinum, since he described other species in this genus containing excipular crystals. In addition, the reddish tinge of the hypothecium in B. murinum, becoming more distinctly reddish in K, was not observed in the new taxon.
The crystals of the excipulum and the pruina probably cause the UV+ bright neon blue fluorescence in B. incrustatimurinum. Specimens of B. farkasiae tested did not exhibit UV-fluorescence. In B. hypophyllum, the non-pruinose, orange-brown apothecia exhibit a distinct, rather bright white UV-fluorescence. We also tested other species of Byssoloma for UV-fluorescence; most species did not react but typical specimens of B. leucoblepharum had the same bluish UV-fluorescence of the apothecial disc as found in Byssolecania. Specimens of B. microcarpum Kalb & Vězda show a combination of blue fluorescence of the apothecial disc and lemon yellow fluorescence of the farinose thallus, whereas in B. subdiscordans (Nyl.) P. James, the whitish thallus reacts UV+ light yellowish. The orange-brown apothecia of B. minutissimum Kalb & Vězda have the same UV+ white fluorescence as B. hypophyllum. Clearly, this character provides a very useful toolbox in the taxonomy of foliicolous lichens, which are often too small to perform TLC or HPLC, and we are grateful to one of the reviewers of this manuscript for pointing out this possibility.
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves (hypophyllous), 1 ix 2021, D. A. Ward 691 (B, UBDH). Temburong: Betayan, 4°42′9.99″N, 115°6′11.1204″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves (hypophyllous), 17 ix 2021, D. A. Ward 737 (B).
Byssoloma aff. multipunctatum Lücking
Remarks
This material is characterized by a brownish thallus furnished with numerous dark, wart-shaped pycnidia and scattered apothecia with a dark brown disc, a blackish brown hypothecium and a cream-coloured to light brownish, rather thin but distinctly byssoid margin lacking crystals. The ascospores are 3-septate and c. 10 × 3 μm in size. It comes close to Byssoloma multipunctatum, described from Costa Rica (Lücking Reference Lücking2008), but lacks the small black dots characteristic of the latter. Given that there is only a single specimen with few apothecia, we refrain from recognizing this material formally as a new species.
Specimen examined
Brunei Darussalam: Temburong: Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 ix 2021, D. A. Ward 521a (B).
Byssoloma subaeruginosum D. A. Ward, B. Moncada & Lücking sp. nov.
Index Fungorum No.: IF 905100
Differing from Byssoloma usambarense in the dark aeruginous hypothecium and the (olive-)brown thallus.
Type: Brunei Darussalam, Belait, Wasai Teraja, 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 5 September 2021, D. A. Ward (with D. Cicuzza and B. Zoletto) 183a (UBDH—holotype!).
Thallus continuous, irregular in outline, smooth, 10–20 mm across, very thin (10–15 μm), ecorticate, dark brown to dark olive, opaque, sometimes hypophyllous; prothallus sometimes developed as thin, dark brown line. Photobiont cells bright green, rounded, 5–8 μm diam.
Ascomata biatorine with byssoid margin, adnate to prominent, round, 0.3–0.5 mm diam. and 90–110 μm high; disc dark brown to blackish brown (or greyish when hypophyllous), slightly convex, UV+ bluish, margin cream-coloured to whitish, thin but more or less spreading laterally over the thallus surface. Hypothecium 40–50 μm high, very intensely dark aeruginous to smoky blue, in thicker sections appearing blackish, I−, KI−; apothecial base blackish brown. Epithecium indistinct. Hymenium 45–55 μm high, colourless, I+ blue, KI+ blue. Paraphyses branched and somewhat anastomosing. Asci clavate, 45–50 × 10–13 μm, I+ blue, KI+ blue, with I+ darker blue tubular structure in the tholus. Ascospores 8 per ascus, arranged in a bundle, oblong-fusiform, (5–)7(–11)-septate, without constrictions at the septa, 27–35(–40) × 3–4 μm, colourless, I−.
Pycnidia wart-shaped, 0.04–0.07 mm diam., black in surface view, dark aeruginous under the microscope, wall composed of aeruginous, irregular hyphae. Conidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P) all negative; apothecia with a UV+ bluish reaction.
Etymology
The epithet refers to the aeruginous hypothecium.
Distribution and ecology
Known from several localities of mixed dipterocarp forest in Brunei Darussalam.
Remarks
Species with multiseptate ascospores are comparatively rare in the genus Byssoloma. Closest in septation and size of the ascospores are B. vanderystii and B. usambarense Vězda, both described from Africa (Sérusiaux Reference Sérusiaux1979; Vězda Reference Vězda1987), with predominantly 7-septate ascospores c. 25–30 × 2.5–3.5 μm in size. However, B. vanderystii has strongly convex, reddish brown apothecia with a brown hypothecium, whereas B. usambarense differs in the (light) brown apothecia with reddish brown hypothecium, both lacking an aeruginous to smoky blue tinge in the subhymenial layers. Both also differ in the lighter greenish thallus.
The UV+ bluish reaction of the apothecial disc appears to be the same as observed in several Byssolecania species and in Byssoloma leucoblepharum (see above under Byssolecania fumosocaerulea and Byssoloma incrustatimurinum), although it is much less intense than in Byssolecania fumosocaerulea.
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 245 (B), 701a (B); Wasai Teraja, 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 5 ix 2021, D. A. Ward (with D. Cicuzza and B. Zoletto) 185 (UBDH), 196b (B, filed under Phylloporis verrucosula); ibid., 6 xi 2021, D. A. Ward (with D. Cicuzza) 1183b (UBDH). Temburong: Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 ix 2021, D. A. Ward 935 (B, UBDH), 936a (UBDH); ibid., on dicot leaves, 26 ix 2021, D. A. Ward 1291a (B).
Chroodiscus lirellicarpus D. A. Ward, B. Moncada & Lücking sp. nov.
Index Fungorum No.: IF 905101
Differing from Chroodiscus argillaceus in the lirellate apothecia.
Type: Brunei Darussalam, Temburong, Kampong Tamada, 4°32′55.0644″N, 115°4′47.406″E, 48 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 13 November 2021, D. A. Ward 1224 (UBDH—holotype!; B—isotype!).
Habit and characteristics of newly described species. A–C, Chroodiscus lirellicarpus (A & C, holotype; B, Ward 1262). A & B, thalli with apothecia. C, section through apothecium. D–G, Chroodiscus minutus (D, F & G, holotype; E, Ward 316). D & E, thallus with apothecia. F & G, asci with ascospores. Scales: A, B, D & E = 1 mm; C, F & G = 10 μm. In colour online.
Thallus continuous, rounded to irregular in outline, uneven to irregularly verrucose, 5–10 mm across, thin (20–30 μm), with a cartilaginous corticiform layer, pale (yellowish) grey, slightly nitidous; verrucae 0.05–0.15(–0.2) mm diam., pale (yellowish) grey, in section filled with large clusters of calcium oxalate crystals. Photobiont cells oblong to almost rounded, 7–14 × 4–7 μm, in irregular plates leaving small to larger interspaces.
Apothecia immersed-erumpent, distinctly lirelliform, unbranched to branched or sometimes stellate, (0.3–)0.5–1(–1.5) mm long, 0.15–0.2 mm wide, and 50–60 μm high; disc slit-like, barely visible, plane, light greyish brown, K+ yellow then orange; thalline margin prominent, pale (yellowish) grey, K+ yellow then orange. Excipulum 5–8 μm wide, colourless to dirty yellowish or appearing brownish in thicker sections, K+ orange. Hypothecium 10–20 μm high, colourless to pale yellowish. Epithecium indistinct. Hymenium 35–40 μm high, colourless. Paraphyses unbranched. Asci clavate, 35–40 × 7–8 μm, I−, KI−. Ascospores ellipsoid-fusiform, (1–)3-septate, without constrictions at septum, 8–12 × 2.5–3 μm, colourless.
Pycnidia not observed.
Secondary chemistry
Stictic acid; not tested by TLC due to the very small thalli, but thallus and apothecial sections with K+ persistently light yellow efflux and apothecia P+ orange; other spot tests (C, UV) negative.
Etymology
The epithet refers to the lirellate apothecia.
Distribution and ecology
Known from a single locality of mixed dipterocarp forest in Brunei Darussalam.
Remarks
Chroodiscus (Müll. Arg.) Müll. Arg. is a small genus of exclusively foliicolous lichens in the Graphidaceae. Species either have an orange-red, K+ purple disc or lack pigment (but then invariably produce stictic acid). Within that latter group, all known species have rounded, rather conspicuous apothecia, which sets the new species apart rather easily. In ascospore type, it is most similar to C. argillaceus (Müll. Arg.) Lücking & Papong (Papong et al. Reference Papong, Lücking, Thammathaworn and Boonpragob2009) and the newly described C. minutus (see below). The only other species with lobate-lirellate apothecia in Chroodiscus is C. graphideus Lücking & Kalb (Lücking Reference Lücking2008), which has larger apothecia representing the group with bright orange-red discs; in C. graphideus, the apothecia are more irregularly branched and wider.
The larger verrucae on the thallus of the new species are reminiscent of perithecia in the genus Porina, which initially led us to think that this species grows initially on thalli of that genus, as has been reported for several other species of Chroodiscus (Lücking & Grube Reference Lücking and Grube2002). However, all verrucae checked contained crystals only.
Chroodiscus lirellicarpus is yet another example of species with disparate ascoma morphology being closely related within a single genus. Similar cases among foliicolous taxa include the aforementioned, lirellate Arthonia orbignyae and the lirellate Aulaxina opegraphina versus the mostly rounded to angular ascomata in other foliicolous species of these genera. Another prominent example is the corticolous genus Glyphis in Graphidaceae. The new species somewhat resembles a species of Platythecium in Graphidaceae, but the latter genus differs fundamentally in thallus structure, including a distinct, true cortex, and in ascospore type, featuring graphidean ascospores with thickened, amyloid walls and lens-shaped to rounded lumina.
Additional specimen examined
Brunei Darussalam: Temburong: Kampong Tamada, 4°32′55.0644″N, 115°4′47.406″E, 48 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 13 xi 2021, D. A. Ward 1262 (B).
Chroodiscus minutus D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905102
Differing from Chroodiscus argillaceus in the very small apothecia.
Type: Brunei Darussalam, Belait, Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 September 2021, D. A. Ward 693 (UBDH—holotype!; B—isotype!).
Thallus continuous, rounded to irregular in outline, uneven to irregularly verrucose, 4–6 mm across, thin (20–30 μm), with a cartilaginous corticiform layer, (yellowish) grey, slightly nitidous; verrucae 0.03–0.1(–0.15) mm diam., (yellowish) grey, in section filled with large clusters of calcium oxalate crystals. Photobiont cells oblong to almost rounded, 6–15 × 4–7 μm, in irregular plates leaving small to larger interspaces.
Apothecia immersed-erumpent, regularly rounded, 0.15–0.2(–0.25) mm diam., 50–60 μm high; disc largely exposed, plane, light greyish brown, K+ yellow then orange; thalline margin prominent, (yellowish) grey, K+ yellow then orange. Excipulum 3–6 μm wide, colourless to pale yellowish. Hypothecium 5–10 μm high, colourless to pale yellowish. Epithecium indistinct. Hymenium 35–40 μm high, colourless. Paraphyses unbranched. Asci clavate, 35–40 × 7–8 μm, I−, KI−. Ascospores ellipsoid-fusiform, (1–)3-septate, without constrictions at septa, 7–10 × 2.5–3 μm, colourless.
Pycnidia not observed.
Secondary chemistry
Stictic acid; not tested by TLC due to the very small thalli, but thallus and apothecial sections with K+ persistently light yellow efflux and apothecia P+ orange; other spot tests (C, UV) negative.
Etymology
The epithet refers to the small thallus and apothecia.
Distribution and ecology
Known from two areas of mixed dipterocarp forest in Brunei Darussalam.
Remarks
Chroodiscus minutus is another new species in the C. argillaceus group, characterized by a non-pigmented apothecial disc, an uneven-verrucose thallus with irregular photobiont, and stictic acid as secondary compound. The round apothecia and small, 3-septate ascospores, together with the absence of (disc-shaped) isidia, place it in the vicinity of C. argillaceus (Papong et al. Reference Papong, Lücking, Thammathaworn and Boonpragob2009). The latter differs in the much larger apothecia (up to 0.5 mm diam.) and the overall lighter thallus and light grey to yellowish apothecial disc. Chroodiscus argillaceus was found frequently in the present material and the morphological differences are constant and quite obvious.
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 316a (B), 317a (UBDH). Temburong: Kampong Tamada, 4°32′55.0644″N, 115°4′47.406″E, 48 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 13 xi 2021, D. A. Ward 1204 (UBDH), 1209 (B, UBDH).
Echinoplaca aff. hispida Sipman
(Fig. 11A)
Habit and characteristics of newly described or otherwise interesting species. A, Echinoplaca aff. hispida (Ward 169a), thallus with sterile setae. B–F, Echinoplaca septemseptata (holotype). B–D, thallus with apothecia. E & F, asci with ascospores. Scales: A–D = 1 mm; E & F = 10 μm. In colour online.
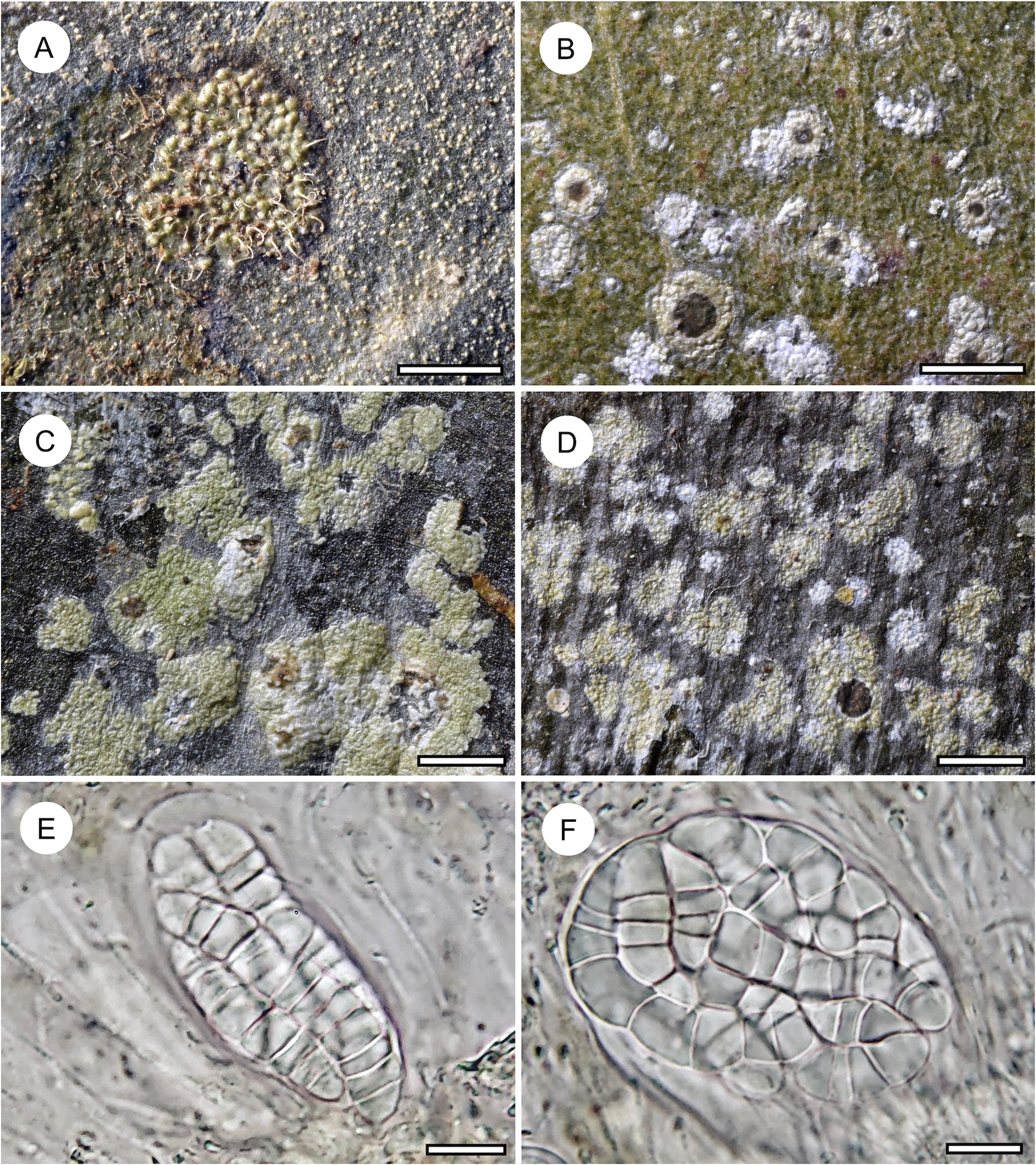
Remarks
This unidentified taxon was represented only by a single, small specimen, characterized by a densely verrucose-bullate thallus furnished with whitish setae. This thallus type is rare among Gomphillaceae and not yet known in combination with sterile setae. Echinoplaca hispida Sipman, which was also found in the material, is somewhat similar, but the thallus verrucae are more irregular and separated by thinner thallus parts (Aptroot & Sipman Reference Aptroot and Sipman1993). Another similar species is Gyalectidium microcarpum (Vězda) Lücking et al. (Ferraro et al. Reference Ferraro, Lücking and Sérusiaux2001), which produces rather large, regular verrucae, but in that species the sterile setae are more robust.
Specimen examined
Brunei Darussalam: Belait: Wasai Telingan, Teraja, 4°22′19.2252″N, 114°27′58.3344″E, 149 m, tropical moist broadleaf forest ecoregion, riparian forest/mixed dipterocarp forest, on leaves, 5 ix 2021, D. A. Ward (with D. Cicuzza and B. Zoletto) 169a (B).
Echinoplaca septemseptata D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905103
Differing from Echinoplaca tetrapla in the 7-septate ascospores.
Type: Brunei Darussalam, Temburong, Betayan, 4°42′9.99″N, 115°6′11.1204″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 17 September 2021, D. A. Ward 494a (UBDH—holotype!).
Thallus dispersed, with individual patches becoming confluent, up to 10 mm across, irregular in outline but individual patches rounded, up to 1 mm diam., connected through a translucent hypothallus, 15–25 μm thick, with a cartilaginous corticiform layer, their surface finely verrucose, pale greenish grey to whitish, slightly nitidous; verrucae 0.05–0.1 mm diam., whitish, in section filled with clusters of calcium oxalate crystals. Photobiont cells rounded to somewhat angular, 4–5 μm diam.
Apothecia adnate and spot-like, emarginate, rounded, 0.2–0.5 mm diam., 50–60 μm high; disc plane, greyish brown. Excipulum composed of branched, radiating hyphae embedded in gelatinous matrix, reduced and spreading over thallus surface, 30–50 μm wide, colourless. Hypothecium 5–10 μm high, pale yellowish. Epithecium 3–5 μm high, yellowish granulose. Hymenium 45–50 μm high, colourless. Paraphyses branched and anastomosing. Asci clavate to ellipsoid, 40–45 × 12–16 μm, I−, KI−. Ascospores oblong, 5–7-septate, with slight constrictions at the septa, 20–25 × 5–6 μm, colourless.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) all negative.
Etymology
The epithet refers to the 7-septate ascospores.
Distribution and ecology
Known from several localities of mixed dipterocarp (and riparian) forest in Brunei Darussalam.
Remarks
This new species belongs to a group of species with dark, brownish apothecia and small, transversely septate ascospores. Previously included in a broadly defined Echinoplaca pellicula (Santesson Reference Santesson1952), forms with dark apothecia were later separated under the name E. tetrapla (Zahlbr.) Lücking (Lücking Reference Lücking2001, Reference Lücking2008). However, this still appears to represent a species complex. Genuine E. tetrapla, described from China (Zahlbruckner Reference Zahlbruckner and Handel-Mazetti1930), is characterized by a dispersed to confluent thallus and rather small (12–14 × 5–6 μm), 3-septate ascospores. Neotropical material identified with this name has a more or less continuous, marginally somewhat diffuse thallus, the apothecia are quite dark, and the ascospores are predominantly 5-septate and 18–26 × 5–7 μm in size (Lücking Reference Lücking2008). The present material combines the external morphology of authentic material of E. tetrapla with the much larger ascospore size of the neotropical material; however, it differs from both in the mostly 7-septate ascospores, thus justifying its consideration as a separate taxon. As a consequence, the neotropical material will also have to be recognized as a separate taxon.
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 340 (UBDH); Wasai Telingan, Teraja, 4°22′19.2252″N, 114°27′58.3344″E, 149 m, tropical moist broadleaf forest ecoregion, riparian forest/mixed dipterocarp forest, on leaves, 5 ix 2021, D. A. Ward (with D. Cicuzza and B. Zoletto) 170 (B).
Eremothecella cyaneoides Lücking
Habit and characteristics of newly described or otherwise interesting species. A–D, Eremothecella cyaneoides (A–C, Ward 257; D, Ward 204). A & D, thalli with apothecia. B & C, asci with ascospores. E & F, Eremothecella helicella (Ward 260), conidia. Scales: A & D = 1 mm; B, C, E & F = 10 μm. In colour online.
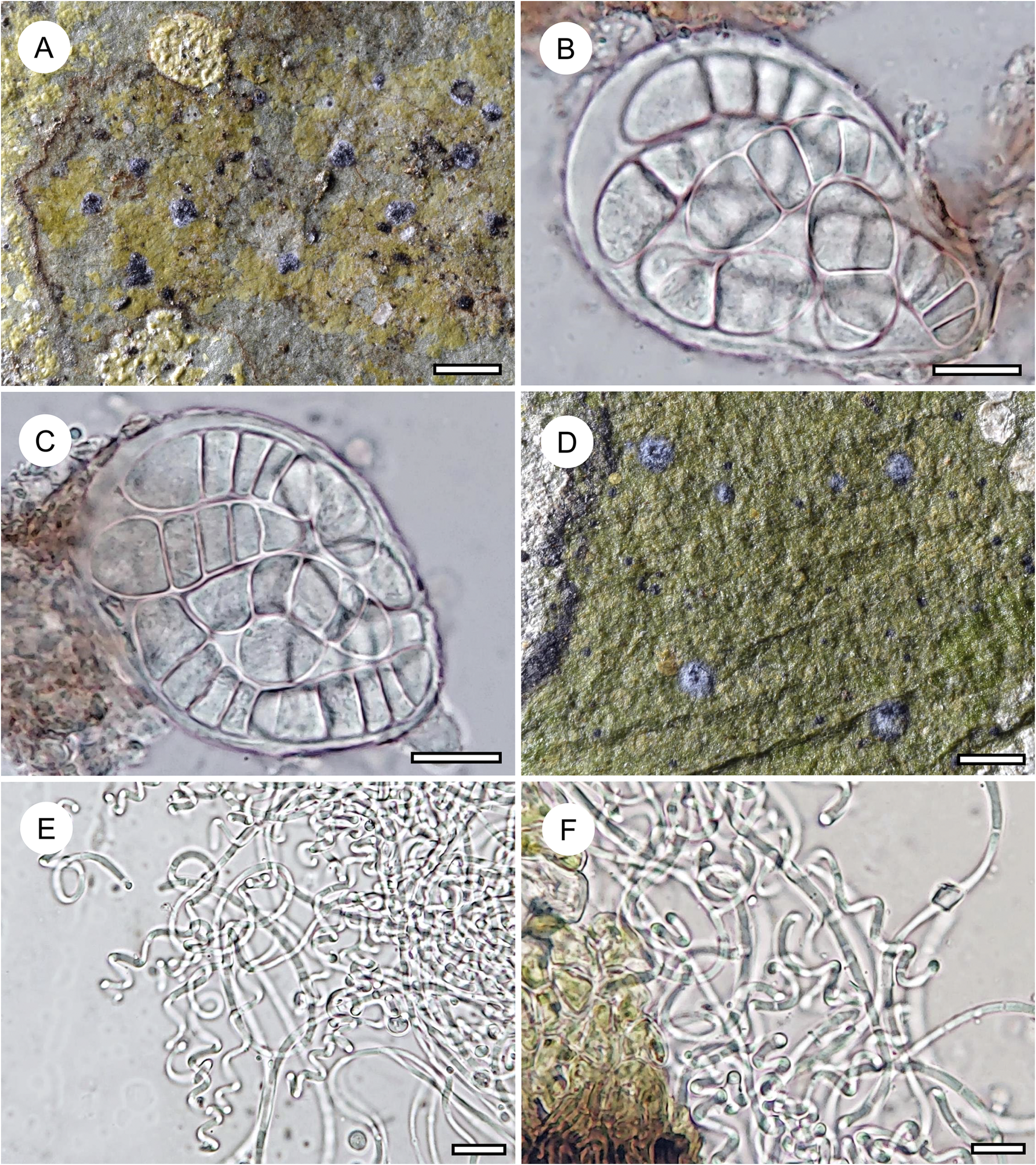
Distribution and ecology
Foliicolous in the understorey of tropical rainforests. A new record for tropical Asia.
Remarks
This species was so far known only from Australia, based on rather limited material (Lücking et al. Reference Lücking2001). The material from Brunei has mostly 5(–7)-septate ascospores, whereas in the type collection from Australia they are (3–)5-septate, but since the size is about the same (30–35 × 7–9 μm vs 25–35 × 8–10 μm) we consider the material from Brunei conspecific. Pycnidia with conidia were not found in the present material. The species resembles Arthonia cyanea Müll. Arg. (Lücking Reference Lücking2008), Eremothecella ajaysinghii Jagad. Ram & G. P. Sinha (Jagadeesh Ram & Sinha Reference Jagadeesh Ram and Sinha2019) and E. macrocephala (Thor et al. Reference Thor, Lücking and Matsumoto2000) in the pruinose ascomata with a bluish tinge, but is intermediate in ascospore dimensions and septation between the Arthonia and the other two Eremothecella species.
Specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on dicot leaves, 1 ix 2021, D. A. Ward 257 (B, UBDH), 304 (B), 1280m (B; filed under Raciborskiella talaumae); ibid., on fern leaves, 1 ix 2021, D. A. Ward 710a (UBDH).
Eremothecella helicella Aptroot & M. Cáceres
Distribution and ecology
Corticolous and foliicolous in the understorey of tropical rainforests. A new record for tropical Asia.
Remarks
Eremothecella helicella was described as a species based on pycnidia only, with the conidia having the shape of a spiral (Cáceres et al. Reference Cáceres, Aptroot and Ertz2014). Two foliicolous specimens with pycnidia only, producing spirally twisted conidia, were found in the present material and are identified with this name, although we suspect that possibly more than one species, including still undescribed taxa, might produce such conidia.
Specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 260 (B, UBDH), 266 (UBDH).
Eremothecella rugosa D. A. Ward, B. Moncada & Lücking sp. nov.
Index Fungorum No.: IF 905104
Differing from Eremothecella calamicola in the ridged thallus and the much longer conidia.
Type: Brunei Darussalam, Belait, Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 September 2021, D. A. Ward 255 (UBDH—holotype!; B—isotype!).
Habit and characteristics of newly described species. A–F, Eremothecella rugosa (A–E, holotype; F, Ward 272). A & B, thallus with apothecia. C, ascus with ascospores. D & E, ascospores. F, thallus with pycnidia. Scales: A, B & F = 1 mm; C–E = 10 μm. In colour online.

Thallus dispersed into irregular patches, rugose, 5–15 mm across, very thin (7–10 μm), ecorticate, yellowish green, slightly glossy, with flattened irregular to elongate warts and ridges, up to 30 μm thick, filled with colourless crystals that dissolve in K. Photobiont cells angular-elongate, 5–10 × 4–6 μm, in irregular to partly radiating plates.
Ascomata hardly raised above thallus level, rounded, 0.4–0.8 mm diam. and 50–60 μm high, brownish black. Hypothecium 3–5 μm high, colourless, I−, KI−. Epithecium formed by dense layer of parallel, periclinal hyphae, 5–8 μm high, reddish brown with purplish tinge, K+ olive to military green, I+ brownish, KI+ greenish grey. Hymenium 45–50 μm high, colourless, I−, KI−. Paraphyses branched and anastomosing. Asci obovate to globose, 35–40 × 30–35 μm, I−, KI−. Ascospores 8 per ascus, clavate, 5–6-septate, with the terminal cell enlarged, 30–35 × 7–8 μm, colourless, I−.
Pycnidia applanate, elongate, 0.5–0.6 mm long and 0.15–0.2 mm wide, brownish black in surface view, dark brown under the microscope, wall composed of dark brown, parallel to radiating hyphae. Conidia long filiform with tapering ends, 50–60-septate, 300–400 × 2–2.5 μm, colourless.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) all negative except for the crystals in the warts and ridges dissolving in K.
Etymology
The epithet refers to the rugose thallus.
Distribution and ecology
Known from several localities of mixed dipterocarp forest in Brunei Darussalam; particularly abundant in Andulau Forest Reserve.
Remarks
Eremothecella is a comparatively small genus of foliicolous lichens comprising ten species, separated from Arthonia by the long-filiform conidia and from foliicolous species of Arthonia by the longer, multiseptate ascospores (Lücking Reference Lücking2008; Jagadeesh Ram & Sinha Reference Jagadeesh Ram and Sinha2019). Within this genus, species are distinguished by ascospore septation and size, as well as the presence or absence of a variously coloured pruina on the ascomata. Eremothecella rugosa is the first species in the genus producing a thallus ornamented with warts and ridges. Its non-pruinose ascomata, with comparatively small ascospores, place it otherwise near E. calamicola, which differs in the entirely smooth thallus (Lücking Reference Lücking2008). In its thallus morphology, the new species resembles Arthonia flavoverrucosa and A. flavirugosa (see above), but both differ clearly in the small, 2-septate, grey-brown ascospores. The conidia of the new species appear to be by far the longest known in the genus.
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 252 (UBDH), 253 (B, UBDH), 272a (B), 275 (B, UBDH), 338 (B), 408 (B), 460a (UBDH), 690a (B, UBDH); Jalan Labi Heath Forest, 4°28′32.4084″N, 114°28′2.7768″E, 51 m, tropical moist broadleaf forest ecoregion, heath forest, on leaves, 19 ix 2021, D. A. Ward 1283f (B, filed under Mazosia melanophthalma); Jalan Labi Mixed Dipterocarp Forest, 4°34′49″N, 114°28′35″E, 81 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 10 ix 2021, D. A. Ward 1285g (B, filed under Semigyalecta paradoxa); Wasai Teraja, 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 6 xi 2021, D. A. Ward (with D. Cicuzza) 1186 (UBDH); ibid., on dicot leaves, 6 xi 2021, D. A. Ward (with D. Cicuzza) 1290h (B; filed under Mazosia melanophthalma). Temburong: Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 ix 2021, D. A. Ward 1292l (B, filed under Microtheliopsis uleana); Kampong Tamada, 4°32′55.0644″N, 115°4′47.406″E, 48 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 13 xi 2021, D. A. Ward 1196 (B).
Eremothecella verrucispora D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905105
Differing from Eremothecella calamicola in the brown, ornamented ascospores with more numerous septa.
Type: Brunei Darussalam, Belait, Wasai Teraja; 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 5 September 2021, D. A. Ward (with D. Cicuzza and B. Zoletto) 175 (UBDH—holotype!).
Habit and characteristics of newly described or otherwise interesting species. A–D, Eremothecella verrucispora (holotype). A, thallus with apothecia. B–D, asci with immature and mature ascospores. E & F, Fellhanera aff. bouteillei (Ward 605), thallus with apothecia, in F showing UV+ yellow fluorescence. Scales: A, E & F = 1 mm; B–D = 10 μm. In colour online.
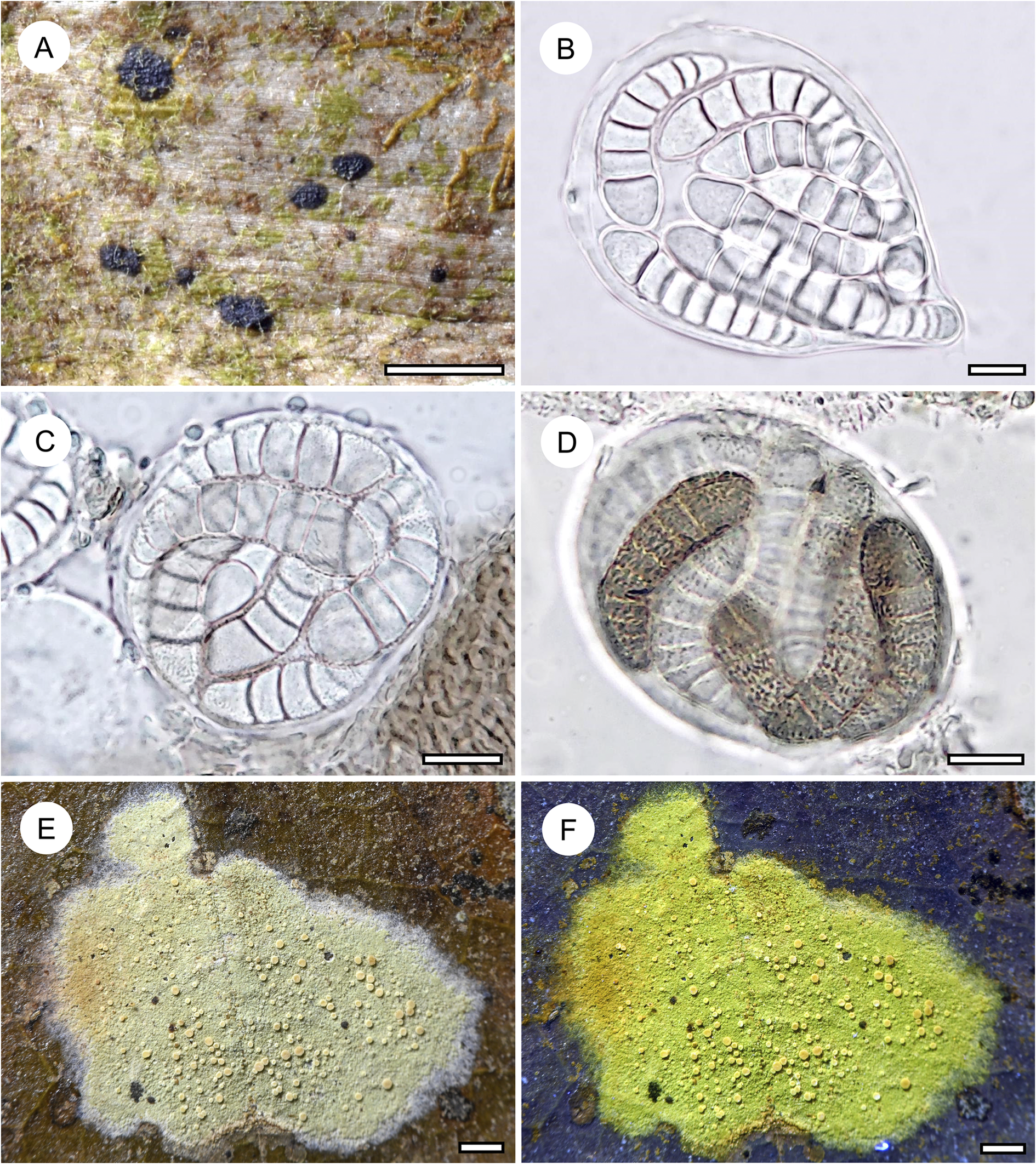
Thallus dispersed into irregular patches, smooth, 5–10 mm across, very thin (7–10 μm), ecorticate, greenish, opaque. Photobiont cells angular-elongate, 5–10 × 4–6 μm, in irregular to partly radiating plates.
Ascomata hardly raised above thallus level, rounded to somewhat irregular in outline, 0.4–0.7 mm diam. and 50–60 μm high, dark brown to blackish brown. Hypothecium 3–5 μm high, colourless, I−, KI−. Epithecium formed by dense layer of parallel, periclinal hyphae, 5–8 μm high, brownish, K− to K+ sordid olive green, KI+ sordid olive green. Hymenium 45–50 μm high, colourless, I−, KI−. Paraphyses branched and anastomosing. Asci obovate to globose, 45–50 × 35–40 μm, I−, KI−. Ascospores 8 per ascus, clavate, (7–)9-septate, with the terminal cell enlarged, 35–40 × 9–10 μm, at first colourless and with smooth walls but soon walls developing minute verrucae and ascospores finally becoming brown, I−.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) all negative.
Etymology
The epithet refers to the minutely verrucose ascospores.
Distribution and ecology
Known only from the type collection from a mixed dipterocarp forest in Brunei Darussalam.
Remarks
Although pycnidia with filiform conidia, characteristic of the genus Eremothecella, were not observed in the material, the new species is placed in this genus given the multiseptate, worm-like ascospores (Lücking Reference Lücking2008; Jagadeesh Ram & Sinha Reference Jagadeesh Ram and Sinha2019). Eremothecella verrucispora is the first species in this genus known to produce brown, verruculose ascospores, such ascospores otherwise being found in some corticolous species of Arthonia and Opegrapha s. lat. Within Eremothecella, E. ajaysinghii and E. microcephalica Sipman have been described with ascospores becoming brown and wrinkled (Sipman Reference Sipman2008; Jagadeesh Ram & Sinha Reference Jagadeesh Ram and Sinha2019), but this appears to be an effect of overaging, and no verrucae are produced. In external appearance and ascospore size, the new species comes close to E. calamicola Syd., but in the number of septa it is intermediate between E. calamicola and E. macrosperma (Jagadeesh Ram & Sinha Reference Jagadeesh Ram and Sinha2019)
Fellhanera aff. bouteillei (Desm.) Vězda
Distribution and ecology
A presumably subcosmopolitan species with a wide ecological amplitude, ranging from tropical to temperate climates and found on leaves, needles and bark.
Remarks
Fellhanera bouteillei, though not usually abundant, is presumed to be the most widespread species commonly found on leaves, and also known from bark and rock (Fig. 15). Its wide ecological amplitude challenges its status as a single taxon, but so far attempts to apply a more detailed species concept have failed, although molecular data support a multispecies complex (Santesson Reference Santesson1952; Lücking Reference Lücking2008; Wang et al. Reference Wang, van den Boom, Sangvichien and Wei2020). The only species thus far described as a separate taxon within this complex is F. parvula (Vězda) Vězda (Vězda Reference Vězda1986), also identified in the present material, differing in the lack of secondary compounds, the somewhat translucent apothecia, and the smaller ascospores. In the present material, we found several well-developed individuals that could be identified as F. bouteillei s. lat., due to the 1-septate ascospores and the yellowish apothecia. However, they consistently differ from typical F. bouteillei in the light yellowish green (vs pale bluish grey), UV+ bright yellow thallus and the small (0.1–0.2 mm diam.), light yellow (vs larger, orange-yellow) apothecia, as well as the smaller ascospores (10–12 × 3–4 μm vs 13–17 × 4–6 μm). In the size of the apothecia and the ascospores, the material comes close to F. parvula, but the apothecia are not translucent and the distinctive thallus colour and UV-reaction suggests the presence of various secondary compounds. Unfortunately, due to the small thalli, we have not yet been able to assess the secondary chemistry of this taxon. We did check all synonyms listed under Fellhanera bouteillei, including three from the Philippines and one from Tanzania (see Santesson Reference Santesson1952), but none fits this material. The fact that all specimens with this morphology were found in a single locality supports the notion of a potentially undescribed, recently evolved species.
Presumed world distribution of Fellhanera bouteillei as documented by occurrence records in the GBIF platform (GBIF.org (09 February 2026); GBIF Occurrence Download https://doi.org/10.15468/dl.ktzap8). In colour online.
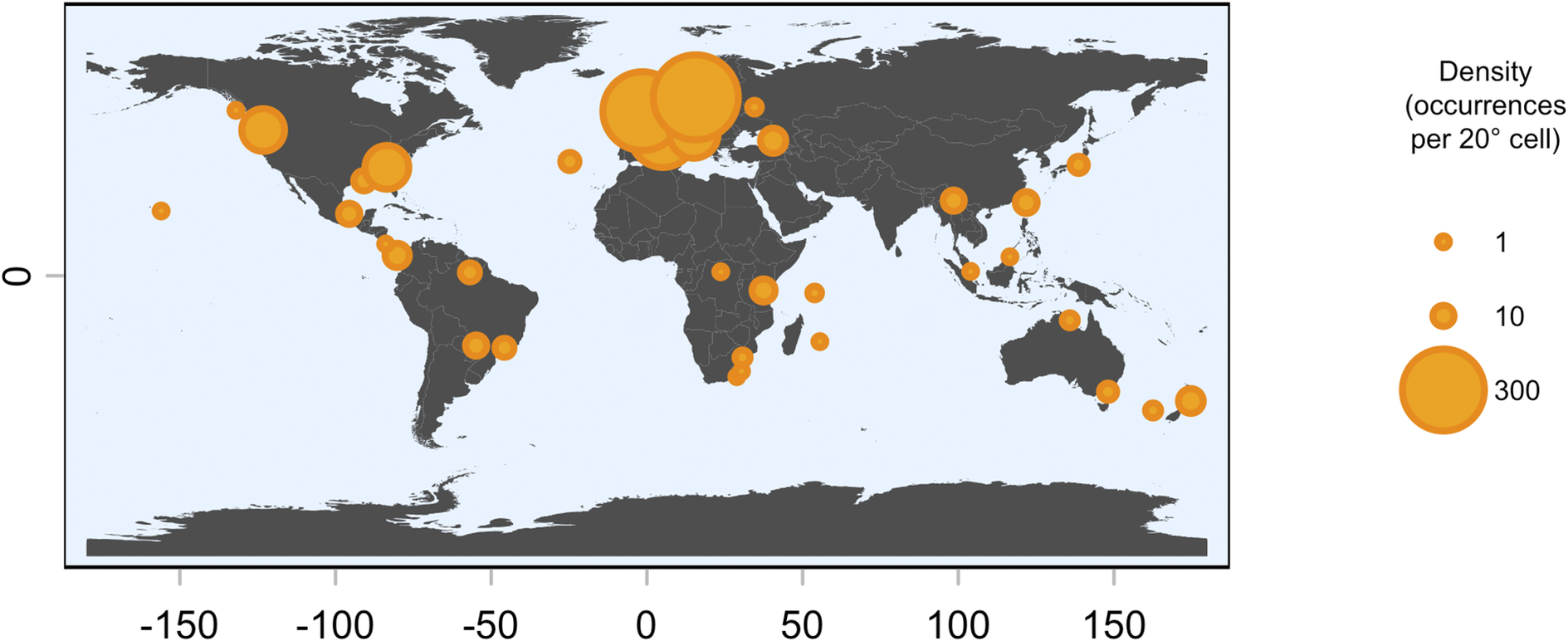
Specimens examined
Brunei Darussalam: Brunei Muara: Bukit Tempayan Pisang, 5°0′43.9236″N, 115°2′42.8028″E, 56 m, tropical moist broadleaf forest ecoregion, secondary forest, on leaves, 6 xii 2021, D. A. Ward (with J. Wong) 605 (B), 606b (B; filed under Coenogonium subluteum), 610b (UBDH; filed under Mazosia melanophthalma), 612b (UBDH).
Fellhanera tutongensis D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905106
Differing from Fellhanera microdiscus in the irregularly 3–7-septate, more elongate and narrower ascospores.
Type: Brunei Darussalam, Tutong, Tutong Kerangas, 4°44′34.4004″N, 114°35′8.6172″E, 25 m, tropical moist broadleaf forest ecoregion, heath forest, on leaves, 18 October 2021, D. A. Ward 986 (UBDH—holotype!).
Habit and characteristics of newly described or otherwise interesting species. A–E, Fellhanera tutongensis (holotype). A & B, thallus with apothecia. C & D, section through apothecium. E, ascospores. F, Graphis pinicola (Ward 953a), thallus with lirellae. Scales: A, B & F = 1 mm; C & D = 20 μm; E = 10 μm. In colour online.

Thallus continuous, 5–10 mm across and 8–15 μm thick, very minutely farinose, light brown with an orange tinge, UV+ faintly orange.
Apothecia round, 0.2–0.3 mm diam. and 80–100 μm high; disc plane, dark brown; margin thin but distinct, chamois-coloured. Excipulum paraplectenchymatous, 15–25 μm wide, partly brownish, encrusted with ochraceous crystals dissolving in K forming a spurious, yellowish cloud. Hypothecium 10–15 μm high, orange-brown, K+ blackish brown; apothecial base blackish, K+ black. Epithecium indistinct, 3–5 μm high, pale yellowish. Hymenium 35–40 μm high, colourless. Asci 30–35 × 7–8 μm. Ascospores 8 per ascus, cylindrical with the distal end slightly tapering, irregularly 3–7-septate, without constrictions at the septa, 18–25 × 2–2.5 μm, 9–10 times as long as wide.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but most standard spot tests (C, K, P) negative except for the ochraceous, K+ dissolving crystals in the excipulum and the faint UV+ orange reaction of the thallus, suggesting the presence of xanthones.
Etymology
The epithet refers to the type region.
Distribution and ecology
Known from two localities of heath forest and mixed dipterocarp forest in Brunei Darussalam.
Remarks
Fellhanera is one of the most diverse genera segregated from the collective genus Bacidia (Vězda Reference Vězda1986), with well over 100 species known (Sérusiaux Reference Sérusiaux1996; Lücking Reference Lücking2008; Farkas Reference Farkas2015b; van den Boom Reference van den Boom2021). This new species belongs in a diverse morphogroup of species characterized by 3–7-septate ascospores and brownish apothecia. Within this group, it is characterized by the K+ blackening hypothecium, in combination with rather narrow ascospores, and the orange-brown thallus. Most similar are F. endopurpurea Hafellner & Vězda, F. rhapidophylli (Rehm) Vězda, and F. wirthii (Vězda) Vězda (Vězda Reference Vězda1986; Vězda & Hafellner Reference Vězda and Hafellner1991; Lücking Reference Lücking2008), all with regularly 3-septate ascospores and a greyish to greenish thallus. Fellhanera endopurpurea and F. wirthii also differ in the larger apothecia with a reddish tinge and K+ purplish hypothecium; F. endopurpurea has wider ascospores (18–25 × 3.5–4 μm, 5–7 times as long as wide), whereas those of F. wirthii are much shorter (8–13 × 2–2.5 μm, 4–5 times as long as wide). Fellhanera rhapidophylli differs in the aeruginous apothecial base and in the shorter and wider ascospores (12–16 × 3–5 μm, 3–5 times as long as wide). Fellhanera microdiscus (Vain.) Vězda (Santesson Reference Santesson1952; Vězda Reference Vězda1986) is morphologically similar to the new species in the often somewhat brownish thallus and the small apothecia, as well as the irregularly 3–5-septate ascospores, but in that species the ascospores are shorter and wider (14–21 × 3.5–5 μm, 5–7 times as long as wide) and the thallus lacks an orange tinge.
The orange-brown colour of the thallus of the new species is not an artifact of preservation, since the entire material was carefully collected and preserved and the leaves harbouring the new species also have many other species present, with typically developed thalli.
Additional specimens examined
Brunei Darussalam: Belait: Jalan Labi Mixed Dipterocarp Forest, 4°34′49″N, 114°28′35″E, 81 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 10 ix 2021, D. A. Ward 813b (B). Temburong: Kampong Tamada, 4°32′55.0644″N, 115°4′47.406″E, 48 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 13 xi 2021, D. A. Ward 594a (B).
Graphis pinicola Zahlbr.
(Fig. 16F)
Remarks
Species of Graphis are only accidentally found on leaves and then almost invariably on thick, often sclerophyllous leaves, where they penetrate the cuticle. We checked several specimens from the Neotropics and they all seem to represent the same species.
Specimen examined
Brunei Darussalam: Tutong: Tutong Kerangas, 4°44′34.4004″N, 114°35′8.6172″E, 25 m, tropical moist broadleaf forest ecoregion, heath forest, on dicot leaves, 18 x 2021, D. A. Ward 953a (B).
Gyalideopsis sp.
Habit and characteristics of newly described or otherwise interesting species. A–F, Gyalideopsis sp. (Ward 995a). A, thallus with hyphophores. B & C, diahyphal ball. D–F, diahyphae. Scales: A = 1 mm; B & C = 20 μm; D–F = 10 μm. In colour online.
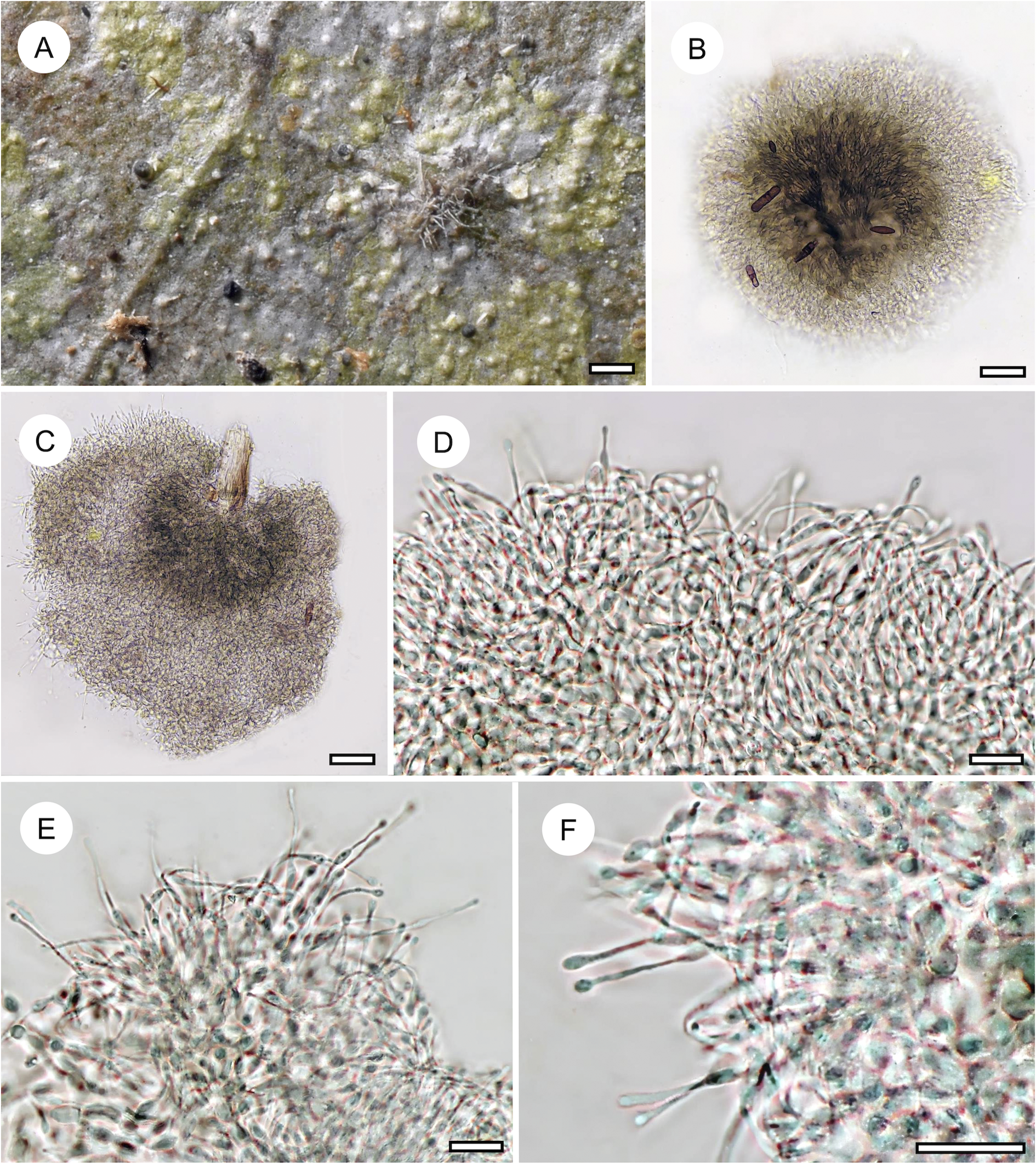
Remarks
This material is represented by two thalli lacking apothecia but bearing hyphophores, with a short, whitish stipe, c. 100 × 15 μm, and an apical, subglobose to bell-shaped, dark grey hyphal bunch projecting laterally from the bent tip of the stipe. The diahyphae are densely packed and moniliform with spermatozoid end cells. No species is currently known with this combination of characters, but in the absence of apothecia we refrain from recognizing this taxon formally. The spermatozoid end cells of the diahyphae are reminiscent of the genera Actinoplaca and Vezdaemyces (Xavier-Leite et al. Reference Xavier-Leite, Cáceres, Aptroot, Moncada, Lücking and Goto2022, Reference Xavier-Leite, Tomio-Goto, Lücking and Cáceres2023; Lebreton et al. Reference Lebreton, Ertz, Lücking, Aptroot, Carriconde, Ah-Peng, Huang, Chen, Stenger and Cáceres2025).
Specimen examined
Brunei Darussalam: Tutong: Tutong Kerangas, 4°44′34.4004″N, 114°35′8.6172″E, 25 m, tropical moist broadleaf forest ecoregion, heath forest, on dicot leaves, 18 x 2021, D. A. Ward 995a (B).
Mazosia bambusiradiata D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905107
Differing from Mazosia bambusae in the radiately arranged thallus verrucae.
Type: Brunei Darussalam, Temburong, Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 September 2021, D. A. Ward 908a (UBDH—holotype!; B—isotype!).
Habit and characteristics of newly described or otherwise interesting species. A–D, Mazosia bambusiradiata (A & B, holotype; C, Ward 430; D, Ward 505a). A–D, thalli with apothecia, in part showing the somewhat fuzzy, dark prothallus; in D a rather wide, dark prothallus. E & F, Mazosia bambusae (E, Ward 908c: F, Ward 463), thallus with apothecia). Scales = 1 mm. In colour online.
Thallus continuous to marginally dispersed, finely verrucose, 5–25 mm across, thin (10–15 μm), brownish grey to olive green, matt to slightly shiny, usually with dark brown prothallus (sometimes fuzzy or only partially developed); verrucae 0.02–0.03 mm diam., dark brown to blackish brown, arranged in distinct (to sometimes diffuse), often radiating lines, especially near the apothecia. Photobiont cells rectangular, 10–15 × 4–5 μm, in regularly radiating plates.
Ascomata round, 0.4–0.6 mm diam. and 70–80 μm high; disc dark grey to black, translucent when moistened; margin gently sloping outwards, usually darker than the thallus. Excipulum 20–35 μm thick, formed by a layer of blackish brown hyphae, externally covered by a thin layer of greyish crystals, a single photobiont layer and a thin layer of colourless hyphae. Hypothecium 10–15 μm high, pale yellowish brown. Hymenium 60–70 μm high. Asci 50–60 × 10–14 μm. Ascospores fusiform, 3-septate, 18–25 × 4–5 μm, upper median cell slightly enlarged, colourless.
Pycnidia low conical, 0.1–0.15 mm diam. Macroconidia not observed. Microconidia ellipsoid, non-septate, 2.5–3.5 × 1–1.3 μm.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) on the thallus all negative.
Etymology
The epithet refers to the similarity with Mazosia bambusae (Vain.) R. Sant. and the radiately arranged thallus verrucae.
Distribution and ecology
Known from several localities of mixed dipterocarp and heath forest in Brunei Darussalam; particularly abundant in Andulau Forest Reserve.
Remarks
This new species is similar to Mazosia bambusae (Santesson Reference Santesson1952; Lücking Reference Lücking2008; see Fig. 18E & F) in most aspects, in particular the thin, brownish olive thalli with dark prothallus and very small, dark verrucae. However, in the latter species, which often occurs sympatrically in the present material, the verrucae are always dispersed over the thallus and not arranged in (radiating) lines. Indeed, the arrangement of the verrucae gives the new species a characteristic appearance. It also provides a morphology that is intermediate between the mostly verrucose species of Mazosia and M. rotula (Mont.) A. Massal., which feature radiating ridges (Santesson Reference Santesson1952; Lücking Reference Lücking2008).
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 374 (B, UBDH), 378 (B), 381 (UBDH), 430 (B), 435b (UBDH; filed under Porina rufula), 467 (B), 699b (B; filed under Badimia elegans), 1277d (B; filed under Coenogonium interplexum), Ward 1278c (B; filed under Bapalmuia pallescens); Jalan Labi Heath Forest, 4°28′32.4084″N, 114°28′2.7768″E, 51 m, tropical moist broadleaf forest ecoregion, heath forest, on leaves, 19 ix 2021, D. A. Ward 505a (B); Jalan Labi Mixed Dipterocarp Forest, 4°34′49″N, 114°28′35″E, 81 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 10 ix 2021, D. A. Ward 861a (B); Wasai Teraja, 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 5 September 2021, D. A. Ward (with D. Cicuzza and B. Zoletto) 191 (UBDH). Temburong: Betayan, 4°42′9.99″N, 115°6′11.1204″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 17 ix 2021, D. A. Ward 739a (B, UBDH); Kampong Tamada, 4°32′55.0644″N, 115°4′47.406″E, 48 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 13 xi 2021, D. A. Ward 1217b (B, UBDH), 1236b (UBDH; filed under Mazosia bambusae). Tutong: Tutong Kerangas, 4°44′34.4004″N, 114°35′8.6172″E, 25 m, tropical moist broadleaf forest ecoregion, heath forest, on leaves, 18 x 2021, D. A. Ward 964 (UBDH), 979a (UBDH).
Mazosia melanophthalma (Müll. Arg.) R. Sant.
Habit and characteristics of newly described or otherwise interesting species. A–D, Mazosia melanophthalma (A, Ward 506; B, Ward 743; C, Ward 774; D, Ward 796), thalli with apothecia. Scales = 1 mm. In colour online.
Distribution and ecology
A presumably pantropical species, usually growing in the shaded forest understorey.
Remarks
Mazosia melanophthalma is a presumably widespread species, found in all tropical regions (Santesson Reference Santesson1952; Lücking Reference Lücking2008; Aptroot et al. Reference Aptroot, Menezes, Xavier-Leite, Santos, Alves and Cáceres2014). However, molecular data suggest some degree of hidden diversity in this taxon (Yao et al. Reference Yao, S-H and Jia2021). To some extent, there are also distinct morphodemes mixed into this species. In the present material, two morphodemes could be distinguished, one with fine, distinct verrucae corresponding to what is typically understood as M. melanophthalma, and one with wider, shallower, confluent verrucae. For the latter, the name Metasphaeria incompleta Rehm is theoretically available, described from the Philippines (Santesson Reference Santesson1952) and agreeing in the shallow, confluent verrucae. However, a more thorough study is needed on ample material of this complex from all tropical regions before applying a more refined species concept.
Mazosia rugosa D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905108
Differing from Mazosia pseudobambusae in the rugose instead of verrucose thallus surface.
Type: Brunei Darussalam, Belait, Jalan Labi Mixed Dipterocarp Forest, 4°34′49″N, 114°28′35″E, 81 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 10 September 2021, D. A. Ward 853 (UBDH—holotype!; B—isotype!).
Habit and characteristics of newly described or otherwise interesting species. A–F, Mazosia rugosa (A, B, E & F, holotype; C, Ward 852; D, Ward 778). A–D, thalli with apothecia. E & F, asci with ascospores. Scales: A–D = 1 mm; E & F = 10 μm. In colour online.
Thallus continuous but algiferous patches irregularly dispersed over a darkened hypothallus, finely rugose, 10–30 mm across, thin (10–20 μm), algiferous patches light brownish grey to cream-coloured and slightly shiny, UV+ patchily bluish white, hypothallus brown to dark brown, opaque; thallus delimited by a dark brown prothallus; ridges formed by confluent verrucae, covering the algiferous patches, 0.03–0.05 mm thick, cream-coloured, forming a minutely reticulate pattern leaving tiny depressions. Photobiont cells rectangular to oblong, 9–15 × 4–5 μm, in irregular plates leaving small interspaces.
Ascomata round, 0.3–0.4 mm diam. and 70–80 μm high; disc black, translucent when moistened; margin irregular, gently to sometimes steeply sloping outwards, the light grey to cream-coloured thalline cover often disrupted to expose the blackish excipulum. Excipulum 10–25 μm thick, formed by a layer of blackish brown hyphae, externally partly covered by a thin layer of greyish crystals, a single photobiont layer and a thin layer of colourless hyphae. Hypothecium 10–15 μm high, pale yellowish brown to brown. Hymenium 60–70 μm high. Asci 50–60 × 12–15 μm. Ascospores fusiform, 3-septate, 15–20 × 4–6 μm, upper median cell very slightly enlarged, colourless.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) on the thallus all negative.
Etymology
The epithet refers to the rugose thallus.
Distribution and ecology
Known from several localities of mixed dipterocarp and heath forest in Brunei Darussalam; particularly abundant in Andulau Forest Reserve.
Remarks
Mazosia rugosa at first glance resembles M. pseudobambusae Kalb & Vězda (see Lücking Reference Lücking2008), due to its rather light thallus bordered by a dark prothallus and the apparently pale verrucae. However, on closer inspection, the thallus does not produce discrete verrucae; instead, these are strongly confluent to form irregular, reticulate ridges, giving the thallus a rugose appearance. Such a morphology is thus far unique within the genus Mazosia. All other species with a verrucose thallus have discrete verrucae, including M. tumidula (Müll. Arg.) Zahlbr. and the corticolous M. ocellata (Nyl.) R. C. Harris, in which the verrucae are quite large (Lücking Reference Lücking2008), hence identification with any of these species is not possible. The apothecia are also smaller than in most other species and the thalline cover of the excipulum is usually incomplete or in patches, another unique feature of this new species. The algiferous thallus patches are irregular in outline and are dispersed over a dark hypothallus, and they feature a non-radiating photobiont, a combination of characters likewise unknown in other species of the genus.
Mazosia rugosa was found quite abundantly at various localities, with very little variation in its morphology, supporting its interpretation as a new and possibly regionally endemic species.
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 356 (UBDH), 358 (B, UBDH), 376 (B, UBDH), 382 (B, UBDH); Jalan Labi Heath Forest, 4°28′32.4084″N, 114°28′2.7768″E, 51 m, tropical moist broadleaf forest ecoregion, heath forest, on leaves, 19 ix 2021, D. A. Ward 778a (UBDH); Jalan Labi Mixed Dipterocarp Forest, 4°34′49″N, 114°28′35″E, 81 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 10 ix 2021, D. A. Ward 852b (B; filed under Mazosia dispersa). Temburong: Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 ix 2021, D. A. Ward 868a (UBDH).
Ocellularia andulauensis D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905109
Differing from Ocellularia daniana in the more delicate, foliicolous thallus and the much smaller ascospores.
Type: Brunei Darussalam, Belait, Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 September 2021, D. A. Ward 434 (UBDH—holotype!).
Habit and characteristics of newly described species. A–D, Ocellularia andulauensis (holotype). A, thallus with apothecia. B, section through apothecium. C, section through hymenium enlarged. D, asci with ascospores. E & F, Opegrapha albomarginata (E, holotype; F, Ward 414) E, thallus with ascomata. F, thallus with goniocystangia. Scales: A, E & F = 1 mm; B–D = 20 μm. In colour online.
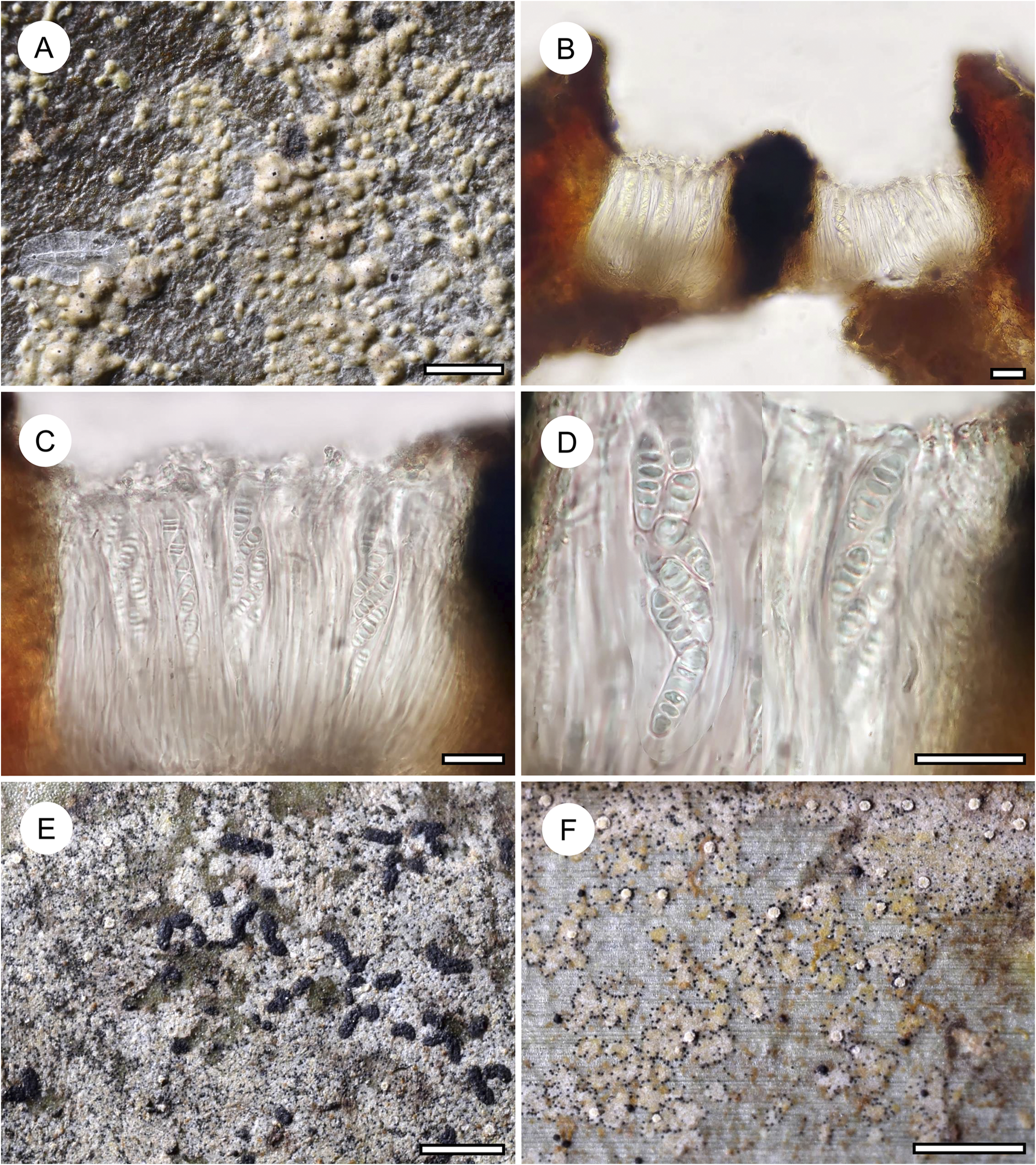
Thallus continuous, coarsely verrucose, up to 30 mm across, moderately thick (50–150 μm), light greenish yellow to light green, opaque to slightly shiny, prothallus absent; verrucae prominent, 0.1–0.2 mm diam. Photobiont cells oblong to irregular, 7–12 × 5–7 μm, in irregular groups or short threads. Thallus in section with dense, prosoplectenchymatous cortex, irregular photobiont layer, and irregular, white medulla; verrucae strongly encrusted with clusters of calcium oxalate crystals and medulla nubilous with numerous small, grey crystals.
Ascomata round, erumpent to prominent, with complete thalline margin, 0.25–0.4 mm diam., 0.15–0.2 mm high; disc covered by 0.05–0.1 mm wide pore, filled by black-tipped columella; proper margin indistinct, sometimes visible as thin whitish rim around the pore; thalline margin entire, smooth, light greenish yellow to light green. Excipulum entire, light orange-brown, 20–30 μm wide, laterally fused with thalline margin; laterally covered by algiferous, corticate thallus layer with thick medulla containing numerous small, grey crystals; columella present, finger-like, up to 50 μm wide and up to 100 μm high, carbonized. Hypothecium prosoplectenchymatous, 10–15 μm high, yellowish. Hymenium 70–80 μm high, hyaline, clear. Epithecium indistinct. Paraphyses unbranched, apically smooth; periphysoids absent. Asci cylindrical to clavate, 70–75 × 12–13 μm. Ascospores 8 per ascus, irregularly arranged, oblong-ellipsoid, 5-septate, 13–15 × 4.5–5 μm, c. 3 times as long as wide, hyaline, distoseptate with lens-shaped lumina, I+ violet-blue.
Secondary chemistry
Probably protocetraric acid and satellite substances; medulla C−, K− to K+ faintly yellowish, P+ deep orange(-red); thallus, apothecia and medulla UV−.
Etymology
The epithet refers to the type locality.
Distribution and ecology
Growing epiphyllous in the understorey of tropical rainforests; thus far known only from the type collection.
Remarks
While Graphidaceae is the most diverse and abundant family of tropical lichens (Lücking et al. Reference Lücking, Johnston, Aptroot, Kraichak, Lendemer, Boonpragob, Cáceres, Ertz, Ferraro and Jia2014), its members very rarely grow on living leaves, with the exception of the genus Chroodiscus (Santesson Reference Santesson1952; Papong et al. Reference Papong, Lücking, Thammathaworn and Boonpragob2009). Typically corticolous species may grow opportunistically foliicolous, although mostly at higher elevations on leathery, long-lived leaves. The material described above is apparently the first species of Ocellularia G. Mey. and related taxa reported from living leaves, Myriotrema foliicola (Hale) Hale not actually being a foliicolous species (Hale Reference Hale1975, Reference Hale1980; Lücking et al. Reference Lücking, Mangold and Lumbsch2016). Due to its chemistry and the presence of a columella, the material belongs in the O. perforata (Leight.) Müll. Arg. morphodeme, which was traditionally circumscribed as a single, variable taxon but represents many, often not directly related species differing in various phenotypic characters (Lücking Reference Lücking2014). Ocellularia andulauensis is in most aspects closest to the recently described O. daniana Lücking from Cuba (Lücking Reference Lücking2015), including the coarsely verrucose thallus, the non-carbonized, orange excipulum combined with a carbonized, finger-like columella and the 5-septate ascospores, in addition to the identical chemistry. However, overall it is much more delicate and the ascospores are distinctly smaller (20–25 × 7–8 μm). Given that the ascospores are mature, with a well-developed, amyloid endospore, the possibility that this foliicolous material is a juvenile form of O. daniana can be rejected. The genus Ocellularia is otherwise quite diverse in Brunei, with numerous species detected on bark (D. H. Ward et al., unpublished data), but none of these exhibit characters in agreement with this foliicolous collection. We therefore consider it justified to formally describe this taxon.
Opegrapha albomarginata D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905110
Differing from all other species of Opegrapha with goniocystangia, in the white margin of the goniocystangia.
Type: Brunei Darussalam, Belait, Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 September 2021, D. A. Ward 411 (UBDH—holotype!).
Thallus dispersed into minute, irregular patches becoming confluent and leaving irregular interspaces, entire thallus bordered by a dark brown prothallus, 5–10 mm across, smooth, thin (8–12 μm), whitish to very pale greenish grey, UV+ faintly pale yellow, with numerous scattered, minute (20–35 μm diam.) black dots originally bordering each patch but eventually irregularly dispersed, opaque. Photobiont cells oblong to irregular, 8–13 × 4–6 μm, in irregular plates. Goniocystangia abundant, rounded, 0.08–0.1 mm diam., with white margins, contents (goniocysts) pale greenish, UV+ pale yellow.
Ascomata adnate, shortly elongate to lirellate, unbranched, 0.2–0.5 mm long and 0.1–0.15 mm wide, 70–90 μm high; disc slit-like; margin thick, black. Excipuloid tissue 20–30 μm thick, basally up to 40 μm wide, absent below hypothecium. Hypothecium 7–15 μm high, colourless. Hymenium in all sectioned ascomata degenerated, without asci and ascospores.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but most standard spot tests (C, K, P) on the thallus negative except for the faint, pale yellow UV-reaction of the thallus and goniocysts.
Etymology
The epithet refers to the white margin around the goniocystangia.
Distribution and ecology
Known from several locations in mixed dipterocarp forest in the Belait area.
Remarks
Opegrapha is one of the largest genera of lichenized fungi, considered to contain c. 300 species (Lücking et al. Reference Lücking, Hodkinson and Leavitt2017); however, molecular studies have shown it to be a collective genus, with several smaller groups already segregated (Ertz & Tehler Reference Ertz and Tehler2011; Ertz et al. Reference Ertz, Tehler, Irestedt, Frisch, Thor and van den Boom2015; Ertz Reference Ertz2018). Around a dozen species are foliicolous, some of them now recognized in the genus Fouragea (Frisch et al. Reference Frisch, Thor, Ertz and Grube2014; Ertz Reference Ertz2020). A small group of five species is known to produce so-called goniocystangia, soralia-like structures with a defined border, namely O. dibbenii, O. lambinonii Sérus., O. luzonensis Sérus., O. santessonii Sérus. and O. serusiauxii Lücking (Sérusiaux Reference Sérusiaux1978, Reference Sérusiaux1985; Lücking Reference Lücking2008). All produce goniocystangia with black margins and, with the exception of O. luzonensis, have a photobiont with rectangular cells in radiating rows, resulting in a continuous, rounded thallus with a characteristic appearance. The goniocystangia with white margins differentiates the new species from all others with those structures; it agrees with O. luzonensis in the irregular photobiont but, apart from the black margins of the goniocystangia, this species has a brown thallus with scattered or indistinct black dots. Sorediate species are otherwise rare in Opegrapha; one paleotropical taxon is Zwackhia sorediifera (P. James) Ertz (Ertz Reference Ertz2009), but here the soralia are diffuse and not delimited in regularly circular, well-delimited structures. The degenerated hymenia suggest that the species is mainly dispersed by its goniocysts.
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 414 (B), 686b (B; filed under Badimia polillensis); Jalan Labi Mixed Dipterocarp Forest, 4°34′49″N, 114°28′35″E, 81 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 10 ix 2021, D. A. Ward 1286e (B; filed under Musaespora epiphylla); Wasai Teraja, 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on dicot leaves, 6 xi 2021, D. A. Ward (with D. Cicuzza) 1290f (B; filed under Mazosia melanophthalma).
Phylloporis verrucosula D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905111
Differing from Phylloporis palmae in the small perithecia and comparatively narrow ascospores.
Type: Brunei Darussalam, Belait, Wasai Telingan, Teraja, 4°22′19.2252″N, 114°27′58.3344″E, 149 m, tropical moist broadleaf forest ecoregion, riparian forest/mixed dipterocarp forest, on leaves, 30 October 2021, D. A. Ward (with D. Cicuzza) 1061 (UBDH—holotype!; B—isotype!).
Habit and characteristics of newly described species. A–G, Phylloporis verrucosula (holotype). A, thallus with perithecia. B & C, asci with ascospores. D–G, ascospores. H & I, Porina riparia (holotype), thallus with disc-shaped isidia. Scales: A, H & I = 1 mm; B–G = 10 μm. In colour online.
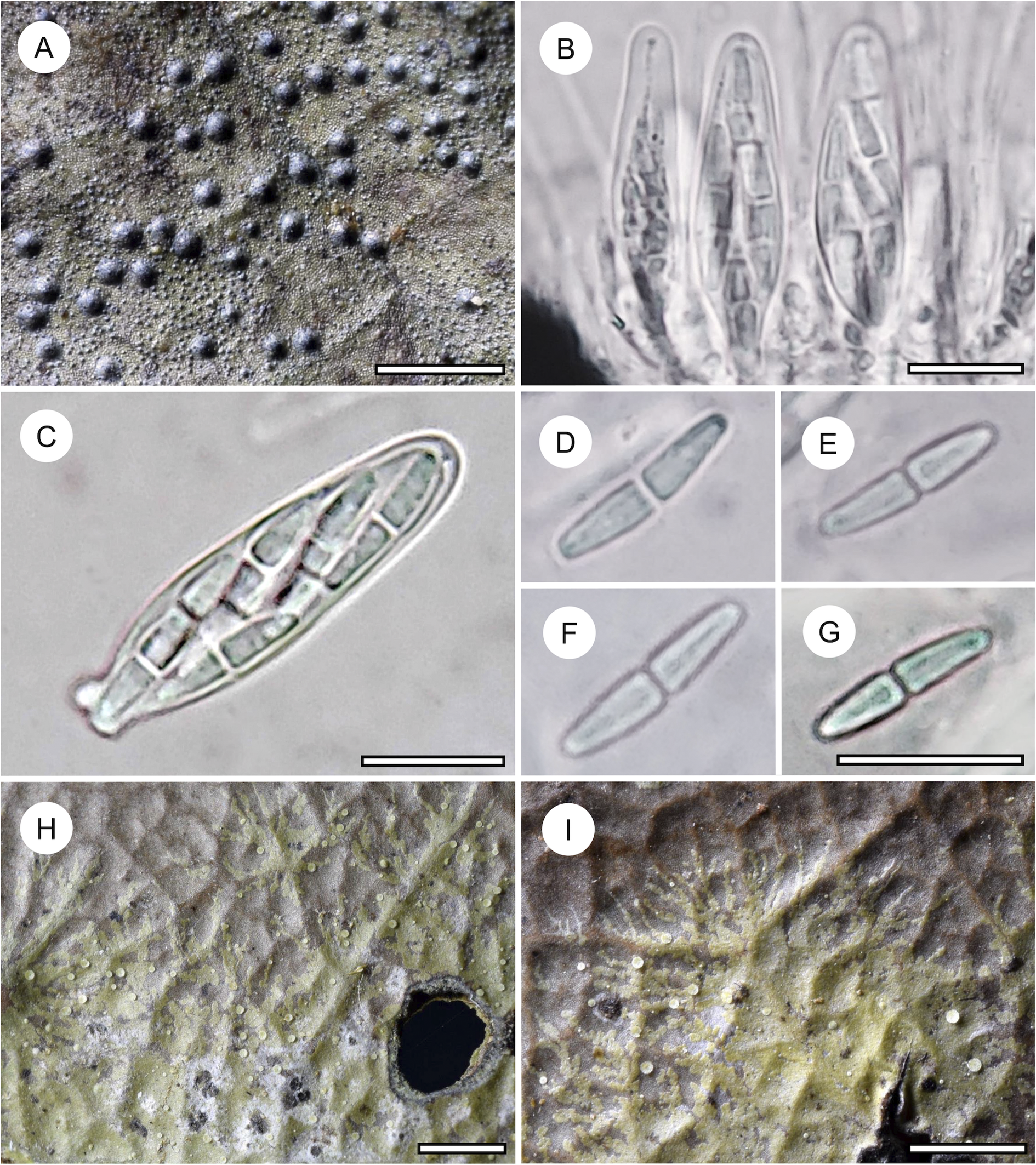
Thallus supracuticular, continuous, 5–15 mm across and 5–10 μm thick, very minutely verrucose, grey, slightly shiny, often bordered by a narrow, dark brown prothallus; verrucae wart-shaped, 0.02–0.03 mm diam., blackish. Photobiont a species of Phycopeltis, cells rectangular to oblong, 8–13 × 3–5 μm, forming nets with distinct interspaces.
Perithecia covered by a thin thallus layer up to ostiole, wart-shaped to almost conical, 0.15–0.25 mm diam. and 70–100 μm high, greyish black. Excipulum prosoplectenchymatous, 7–10 μm thick, colourless to brownish. Involucrellum carbonized, 15–25 μm thick, black. Paraphyses unbranched. Asci obclavate (widest near the base), 25–30 × 6–7 μm. Ascospores irregularly biseriate, oblong, 1-septate, not constricted at the septum, 10–12 × 1.8–2.3 μm, 5–6 times as long as wide.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) on the thallus all negative.
Etymology
The epithet refers to the minutely verrucose thallus.
Distribution and ecology
Known from several localities of mixed dipterocarp (and riparian) forest in Brunei Darussalam; particularly abundant in Andulau Forest Reserve and in Teraja.
Remarks
Phylloporis is a small genus of ten species, recently resurrected to accommodate species with a supracuticular thallus, all closely related and with little variation in perithecial anatomy and ascospore size (Jiang et al. Reference Jiang, Lücking, Xavier-Leite, Cáceres, Aptroot, Portilla and Wei2020). So far two species are known with a minutely verrucose thallus, P. cinefaciens (Nyl.) S. H. Jiang et al. [= P. multipunctata (G. Merr. ex R. Sant.) Vězda] and P. palmae (Cavalc. & A. A. Silva) S. H. Jiang et al. Both differ in ascospore size (12–16 × 3.5–4.5 vs 8.5–10 × 2–3 μm; Jiang et al. Reference Jiang, Lücking, Xavier-Leite, Cáceres, Aptroot, Portilla and Wei2020). The new species described here is distinguished by its very small perithecia, not exceeding 0.3 mm in size, whereas in the other two species they are 0.4–0.7 mm in diameter. The ascospores are intermediate in length between the other two species but much narrower, with a length-to-width ratio of c. 5–6 vs 3.5–4.5 in P. cinefaciens and P. palmae. Phylloporis verrucosula may be more widespread in tropical Asia but might have been confused with P. cinefaciens (= P. multipunctata).
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 416 (UBDH), 419 (UBDH), 421b (UBDH; filed under Phylloblastia dolichospora), 424 (B, UBDH), 425 (B, UBDH); Wasai Telingan, Teraja, 4°22′19.2252″N, 114°27′58.3344″E, 149 m, tropical moist broadleaf forest ecoregion, riparian forest/mixed dipterocarp forest, on leaves, 5 ix 2021, D. A. Ward (with D. Cicuzza and B. Zoletto) 160a (B, UBDH), 196a (B); ibid., 30 x 2021, D. A. Ward (with D. Cicuzza) 1052b (B; filed under Calenia phyllogena), 1103a (B), 1113b (UBDH; filed under Sporopodium argillaceum). Temburong: Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 ix 2021, D. A. Ward 531c (UBDH; filed under Calenia phyllogena), 556 (B, UBDH), 880 (B), 893a (UBDH), 900a (B, UBDH), 947a (B, UBDH), 948a (UBDH).
Porina riparia D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905112
Differing from Porina pseudoapplanata [= Phyllophiale viridis] in the finely laciniate thallus with non-radiating photobiont.
Type: Brunei Darussalam, Temburong, Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 September 2021, D. A. Ward 943a (UBDH—holotype!; B—isotype!).
Thallus continuous but starting out as radiating laciniae and remaining irregularly laciniate towards the margins, often leaving irregular interspaces, smooth, 5–15 mm across and 8–10 μm thick, pale greenish grey, opaque to slightly shiny. Isidia of the Phyllophiale-type always present, usually scattered, disc-shaped, flattened to slightly concave, 0.1–0.15 mm diam., lighter than the thallus to whitish, under the microscope composed of a plate of regularly radiating, colourless hyphae with a cluster of algal cells in the central part. Photobiont cells angular-rounded to oblong, 8–12 × 4–6 μm, in irregular plates.
Perithecia not observed.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) on the thallus all negative.
Etymology
The epithet refers to the occurrence in a riverine forest.
Distribution and ecology
Known from various localities on leaves in mixed dipterocarp forest, partly near streams.
Remarks
The genus Phyllophiale was originally established by Santesson (Reference Santesson1952) for a single, sterile species producing regularly disc-shaped isidia. Much later, Santesson (in Lücking Reference Lücking1991) described a second species, P. fusca R. Sant., and a third species, P. viridis Lücking & M. Cáceres, was subsequently discovered (Lücking & Cáceres Reference Lücking and Cáceres1999). It was eventually found that Phyllophiale-type isidia were associated with Porina-type perithecia (Aptroot & Sipman Reference Aptroot and Sipman1993; Lücking & Vězda Reference Lücking and Vězda1998), and the original species was recombined into Porina, as P. alba (R. Sant.) Lücking, whereas Phyllophiale fusca and P. viridis were established as synonyms of Porina fusca Lücking and P. pseudoapplanata Lücking & M. Cáceres, respectively (Lücking Reference Lücking2004).
The new species is most similar to P. pseudoapplanata and P. weghiana Van den Broeck et al. in having a smooth, pale greenish grey thallus lacking crystals, but differs from both species in the finely laciniate thallus and from P. pseudoapplanata also in the non-radiating photobiont; from P. alba it differs in the absence of calcium oxalate crystals in the thallus (see key below). In his description of Phyllophiale alba, Santesson (Reference Santesson1952) gave the thallus as rough to smooth and the photobiont with radiating or irregular plates, so in the material he cited from across the tropics, he probably mixed specimens now separated as P. pseudoapplanata, P. weghiana and P. riparia within a broadly defined Phyllophiale alba. Despite a careful search, we have not seen any Porina with a similar thallus that could be the fertile counterpart to the isidiate specimens in the material.
Additional specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 666a (B), 671a (B), 706a (B, UBDH), 1278b (B; filed under Bapalmuia pallescens), 1279u (B; filed under Aspidothelium fugiens); Jalan Labi Mixed Dipterocarp Forest, 4°34′49″N, 114°28′35″E, 81 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 10 ix 2021, D. A. Ward 863 (UBDH), 1286k (B; filed under Musaespora epiphylla). Temburong: Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 ix 2021, D. A. Ward 569 (B), 1294g (B; filed under Bapalmuia pallescens); ibid., on dicot leaves, 26 ix 2021, D. A. Ward 1291b (B; filed under Byssoloma subaeruginosum); Kampong Tamada, 4°32′55.0644″N, 115°4′47.406″E, 48 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 13 xi 2021, D. A. Ward 1200 (UBDH), 1266 (UBDH).
Key to foliicolous species of Porina with Phyllophiale-type isidia
Spinomyces rubrotrichoides D. A. Ward, B. Moncada & Lücking sp. nov.
Index Fungorum No.: IF 905113
Differing from Rubrotricha helminthospora, with which it agrees in ascospore type, in the entirely whitish, more delicate sterile thallus setae.

Type: Brunei Darussalam, Temburong, Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 September 2021, D. A. Ward 513a (UBDH—holotype!).
Habit and characteristics of newly described species. A & B, Spinomyces rubrotrichoides (holotype). A, thallus with setae and apothecia. B, ascospores. C, Sporopodium sp. 1 (Ward 680), thallus with apothecia. D, Sporopodium sp. 2 (Ward 810), thallus with apothecia. Scales: A, C &D = 1 mm; B = 10 μm. In colour online.

Thallus continuous, smooth, 5–10 mm across, very thin (8–15 μm), with a cartilaginous corticiform layer, light greenish grey, slightly glossy, furnished with whitish, apically somewhat bent, 0.5–0.7 mm long and 30–40 μm wide, sterile setae, concentrated around the apothecia; no conspicuous prothallus. Photobiont cells bright green, rounded, 5–9 μm diam.
Apothecia biatorine, sessile, regularly rounded, 0.15–0.2 mm diam. and 0.15–0.2 μm high; disc deeply immersed, beige, slightly translucent; proper margin strongly prominent, entire, thin, pale beige to almost white, slightly translucent. Hypothecium 20–30 μm high, colourless, I−, KI−. Epithecium inconspicuous. Hymenium 120–150 μm high, colourless, I−, KI−. Paraphyses unbranched, embedded in a gelatinous matrix. Asci clavate to oblong, 75–110 × 25–45 μm, I−, KI−. Ascospores (1–)2(–4) per ascus, cylindrical to worm-shaped, (7–)9–11-septate, (50–)60–80 × 10–13(–15) μm, with constrictions at the septa, colourless, I−.
Hyphophores not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) all negative.
Etymology
The epithet refers to the similarity of the ascospores to those of Rubrotricha helminthospora.
Distribution and ecology
Known from several localities of mixed dipterocarp (and riparian) forest in Brunei Darussalam.
Remarks
On account of its smooth thallus with white setae and its sessile apothecia with thin but distinct, prominent margin, this new species is placed in the recently reinstated genus Spinomyces Bat. & Peres ex Xavier-Leite et al. (Xavier-Leite et al. Reference Xavier-Leite, Tomio-Goto, Lücking and Cáceres2023). It somewhat resembles Spinomyces aggregatus (Lücking) Xavier-Leite et al., in particular in the white sterile setae clustered around the apothecia, but the latter is overall larger, with yellow apothecia, and the ascospores are muriform. Indeed, no species of Spinomyces with long, multiseptate ascospores is so far known, and such spores are generally rare in Gomphillaceae. Among those with sterile setae formerly classified in Tricharia s. lat., only two feature such ascospores, namely Rubrotricha helminthospora and R. subhelminthospora Lücking (Santesson Reference Santesson1952; Lücking et al. Reference Lücking, Sérusiaux and Vězda2005; Lücking Reference Lücking2008). In both species, the ascospores and apothecia are similar to the new taxon described here but the sterile setae are longer (up to 2 mm) and more robust, and for most of their length reddish brown, with only the apical portion pale.
Additional specimens examined
Brunei Darussalam: Belait: Wasai Telingan, Teraja, 4°22′19.2252″N, 114°27′58.3344″E, 149 m, tropical moist broadleaf forest ecoregion, riparian forest/mixed dipterocarp forest, on leaves, 30 x 2021, D. A. Ward (with D. Cicuzza) 1024 (UBDH); Wasai Teraja, 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 6 xi 2021, D. A. Ward (with D. Cicuzza) 1149 (B), 1164 (B), 1289p (B; filed under Raciborskiella talaumae).
Sporopodium aff. antoninianum Elix, Lumbsch & Lücking
Remarks
The present material is characterized by a pale greenish, finely verrucose thallus with the central portions furnished with densely arranged, pale hairs. It resembles Sporopodium antoninianum, which differs in producing the hairs on an algal-free prothallus (Elix et al. Reference Elix, Lumbsch and Lücking1995; Lücking Reference Lücking2008). Due to the sparse material, the taxonomic status of this form cannot be assessed further.
Specimen examined
Brunei Darussalam: Belait: Wasai Teraja, 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 5 ix 2021, D. A. Ward (with D. Cicuzza and B. Zoletto) 202 (B).
Sporopodium cupuliferum (Nyl.) Vain.
Remarks
This name is reinstated here for material similar to Sporopodium leprieurii Mont. but featuring apothecia with a dark brown apothecial margin.
Specimen examined
Brunei Darussalam: Belait: Wasai Teraja, 4°20′28.9536″N, 114°26′52.908″E, 173 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 5 ix 2021, D. A. Ward (with D. Cicuzza and B. Zoletto) 205 (UBDH).
Sporopodium aff. podosphaera Lücking & R. Sant.
Remarks
This material consists of two well-developed specimens with a pale yellowish grey, smooth thallus and grey, pruinose campylidia lacking a distinct socle. It differs from other species of the genus in the UV+ yellow (not orange) reaction, indicating the presence of lichexanthone instead of, or in addition to, chlorinated xanthones (Elix et al. Reference Elix, Lumbsch and Lücking1995).
Specimen examined
Brunei Darussalam: Brunei Muara: Bukit Tempayan Pisang, 5°0′43.9236″N, 115°2′42.8028″E, 56 m, tropical moist broadleaf forest ecoregion, secondary forest, on leaves, 6 xii 2021, D. A. Ward (with J. Wong) 611 (B, UBDH).
Sporopodium sp. 1
(Fig. 23C)
Remarks
The two collections feature a taxon with an irregularly laciniate thallus leaving large interspaces, with a pale bluish grey colour and minute, whitish verrucae, and a single apothecium with vividly orange disc and cream-coloured, hairy margin. There is no known species of Sporopodium that corresponds with this material (Sérusiaux et al. Reference Sérusiaux, Lücking and Lumbsch2008), but unfortunately it is too scanty to describe formally as a new taxon.
Specimens examined
Brunei Darussalam: Belait: Andulau Forest Reserve, 4°39′19.98″N, 114°31′20.244″E, 72 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 1 ix 2021, D. A. Ward 680a (B, UBDH). Tutong: Tutong Kerangas, 4°44′34.4004″N, 114°35′8.6172″E, 25 m, tropical moist broadleaf forest ecoregion, heath forest, on leaves, 26 xii 2021, D. A. Ward 616 (B, UBDH).
Sporopodium sp. 2
(Fig. 23D)
Remarks
This material is similar to Sporopodium sp. 1, but the thallus is smooth, lacking verrucae, and the apothecial disc is brown rather than orange. Here also, no corresponding species is known (Sérusiaux et al. Reference Sérusiaux, Lücking and Lumbsch2008).
Specimen examined
Brunei Darussalam: Belait: Jalan Labi Mixed Dipterocarp Forest, 4°34′49″N, 114°28′35″E, 81 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 10 ix 2021, D. A. Ward 810 (B).
Trichothelium subrobinsonii D. A. Ward & Lücking sp. nov.
Index Fungorum No.: IF 905114
Differing from Trichothelium robinsonii in the consistently 7-septate ascospores.
Type: Brunei Darussalam, Temburong, Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 September 2021, D. A. Ward 514 (UBDH—holotype!; B—isotype!).
Habit and characteristics of newly described species. A–F, Trichothelium subrobinsonii (holotype). A & B, thallus with perithecia. C, ascus with ascospores. D–F, ascospores. Scales: A & B = 1 mm; C–F = 10 μm. In colour online.
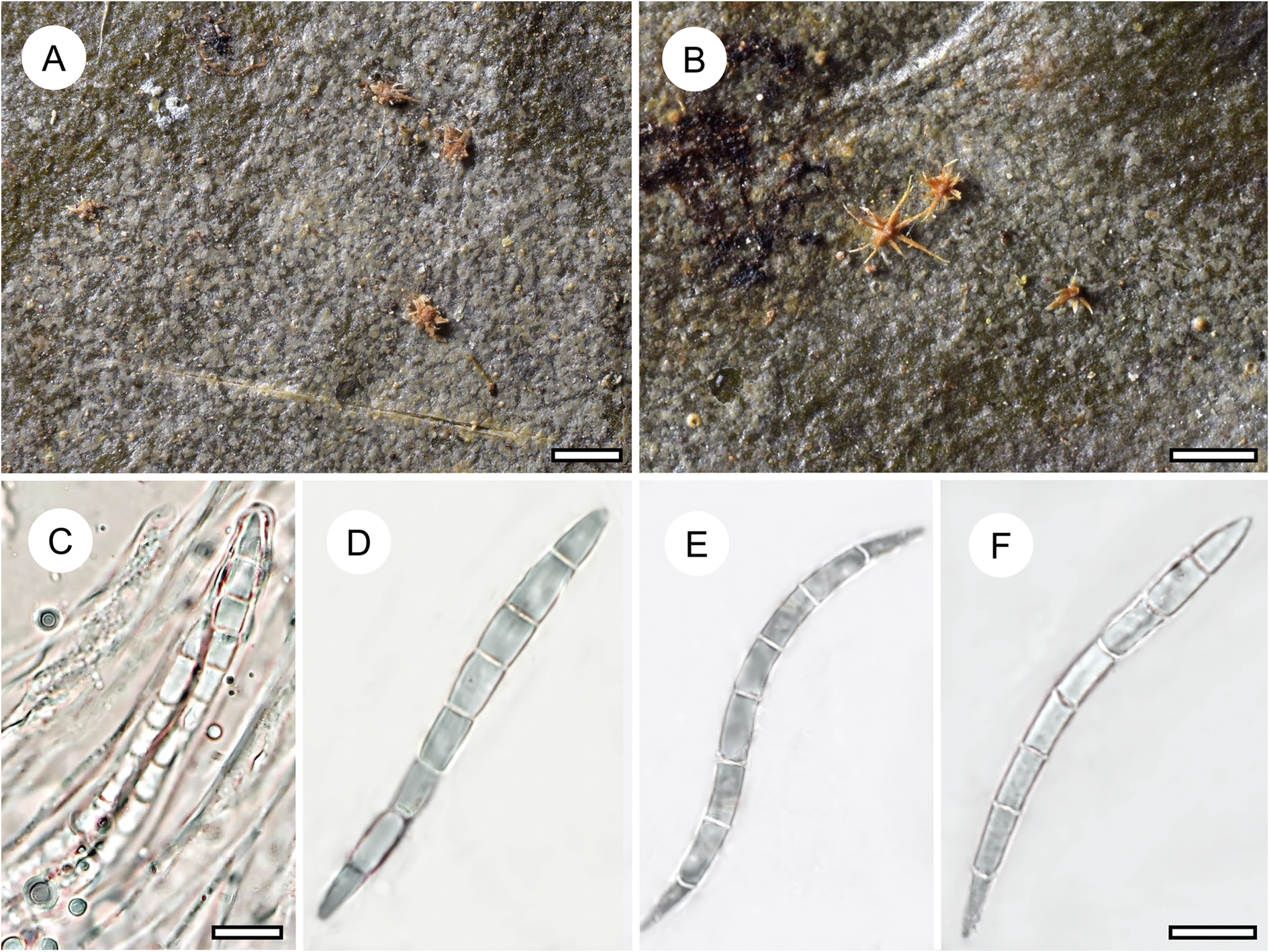
Thallus continuous but finely lacerate-reticulate, leaving small interspaces, smooth, 5–10 mm across and 5–10 μm thick, light grey, slightly shiny. Photobiont cells angular-rounded to oblong, 6–10 × 3–6 μm, in irregular plates leaving small interspaces.
Perithecia sessile, subglobose, 0.2–0.3 mm diam. and 250–300 μm high, apically setose, brownish orange, matt; setae 7–10, acute to bristle-like, projecting horizontally, 0.3–0.5 mm long and 30–80 μm wide at base, pale orange to beige. Excipulum prosoplectenchymatous, 5–15 μm thick, pale yellowish, K+ orange. Involucrellum not distinctly separated from the excipulum, paraplectenchymatous, 25–35 μm thick, yellowish to orange, K+ orange-red, externally covered by 5–10 μm thick, yet indistinct, algiferous thallus layer. Paraphyses unbranched, flexuose. Asci narrowly fusiform, 130–150 × 10–12 μm. Ascospores oblong-bacillar to narrowly fusiform, with the distal end somewhat tapering and sometimes slightly bent, 7-septate, without or with very slight constrictions at septa, 50–65(–80) × 4–5(–6) μm, 9–12(–15) times as long as wide, colourless.
Pycnidia not observed.
Secondary chemistry
Not tested by TLC due to the very small thalli, but standard spot tests (C, K, P, UV) on the thallus all negative; perithecial wall containing Porina yellow, K+ orange-red.
Etymology
The epithet refers to the similarity with Trichothelium robinsonii Vain.
Distribution and ecology
Known only from the type locality but represented by several collections; growing foliicolous in mixed dipterocarp forest.
Remarks
This new species is externally indistinguishable from Trichothelium robinsonii, described from the Philippines (Vainio Reference Vainio1921). Both share the continuous, yet finely lacerate, light greyish thallus and the orange perithecia with distinct setae containing Porina yellow in their walls. Vainio (Reference Vainio1921: 322) gave no ascospore details for his new species, describing T. robinsonii with the remark: ‘Tantum unum apothecium lectum et eam ob causam microscopio haud examinatum.’ However, Lücking (Reference Lücking2008) examined the type material and found the ascospores to be 9–13(–15)-septate, similar to those of T. croceum Sérus., another species described from Papua New Guinea. Lücking (Reference Lücking2008) considered the two taxa potentially conspecific, but T. croceum differs in the rounded thallus patches with the photobiont forming rectangular, regularly radiating cells, and in the white setae contrasting with the orange perithecial wall; the ascospores are also somewhat wider. Trichothelium subrobinsonii agrees with T. robinsonii in external morphology and ascospore type, but the ascospores are always only 7-septate and hence have very elongate cells.
Additional specimens examined
Brunei Darussalam: Temburong: Batu Apoi, 4°40′57.54″N, 115°13′14.538″E, 120 m, tropical moist broadleaf forest ecoregion, mixed dipterocarp forest, on leaves, 26 ix 2021, D. A. Ward 517a (UBDH), 1294z (B; filed under Bapalmuia pallescens).
Supplementary Material
The Supplementary Material for this article can be found at http://doi.org/10.1017/S0024282926101406.
Acknowledgements
We acknowledge the Forestry Department at the Ministry of Primary Resources and Tourism of Brunei Darussalam for providing the export permit number JPH/HOB/TAD/51–180. We also thank Universiti Brunei Darussalam for facilitating the Research Collaboration Agreement reference number UBD/AVC-R/1.21.1 [a]. We thank Dr Siti Salwa Abd Khalid, Dr Norhayati binti Hj Ahmad, and Rodzay bin Hj Abd Wahab for kindly assisting in acquiring the permits. We deeply appreciate field assistance from Jerry Wong, Bianca Zoletto and Boon Yu. The Verein der Freunde des Botanischen Gartens und Botanischen Museums Berlin-Dahlem e.V. provided financial support enabling the first author to visit the BGBM for four weeks for the taxonomic study of the material. The first author gratefully acknowledges financial support for this study from the Fulbright US Student Program, sponsored by the US Department of State. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the Fulbright Program or the Government of the United States. We also thank two anonymous reviewers for valuable comments on an earlier version of the manuscript, in particular the recommendation to check UV-fluorescence in the different species. Mounes Bakhshi (Royal Botanic Gardens Kew, Curator Index Fungorum) is thanked for assistance with bulk registration of the new species names.
Author ORCIDs
Danielle A. Ward, 0009-0000-0595-643X; Robert Lücking, 0000-0002-3431-4636; Bibiana Moncada, 0000-0001-9984-2918.
Competing Interests
The authors declare none.
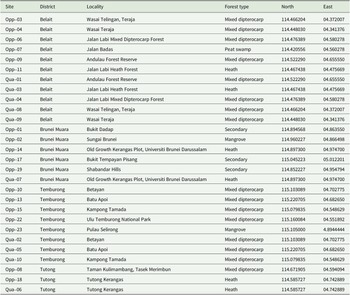
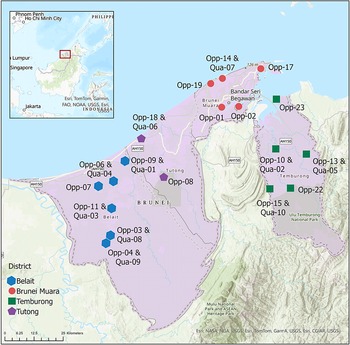

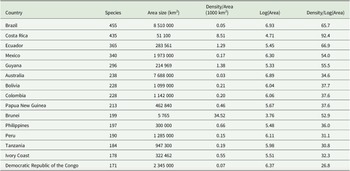
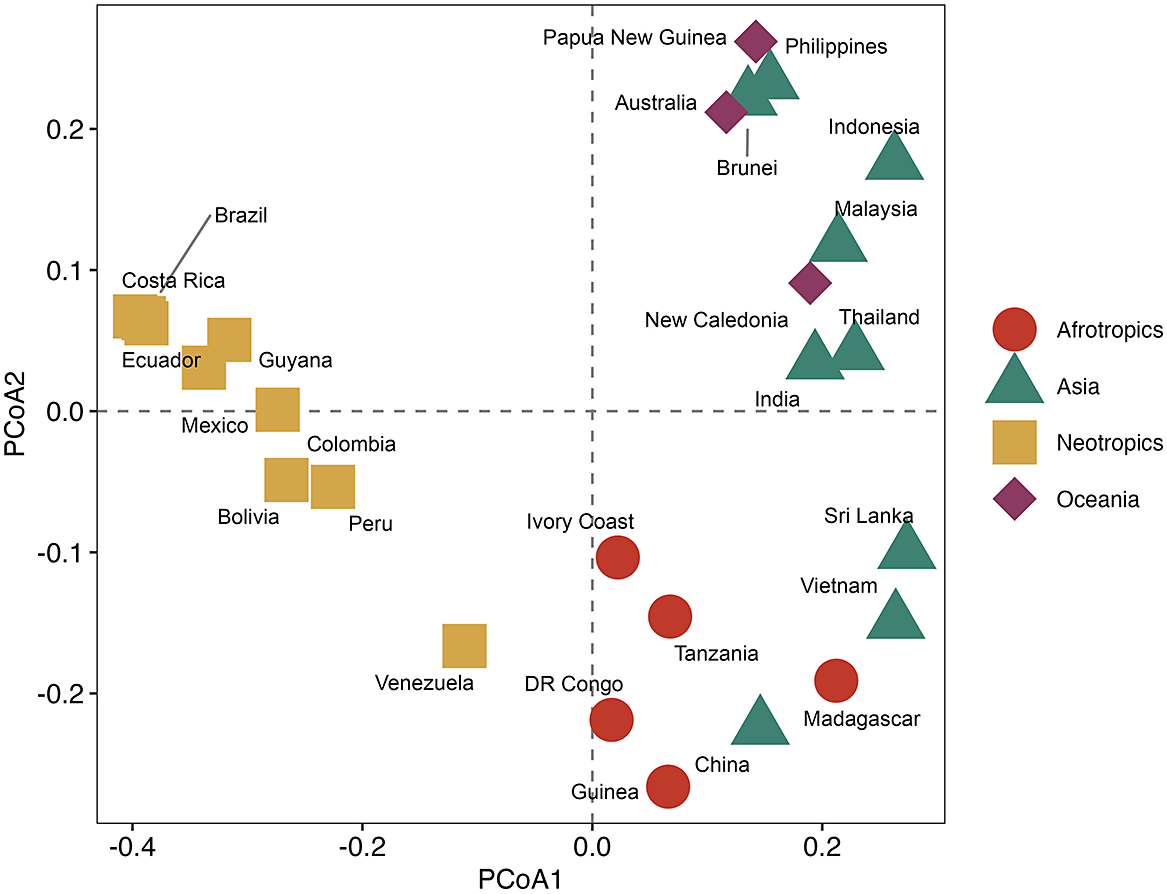
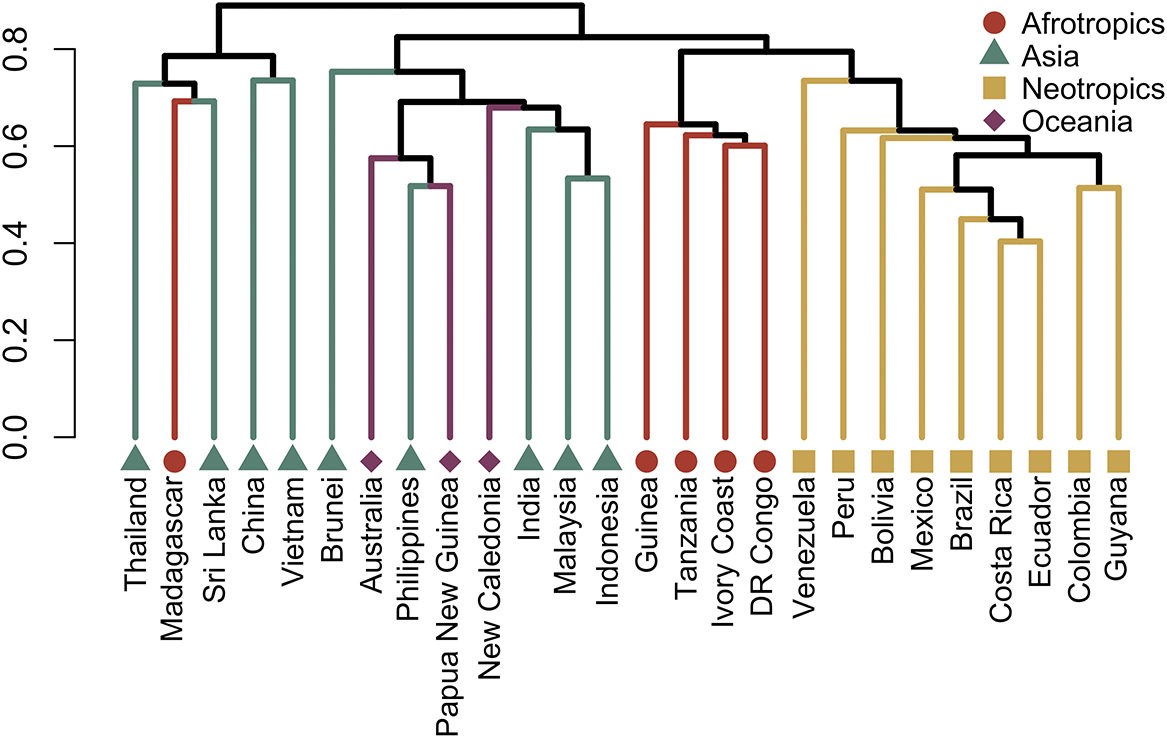
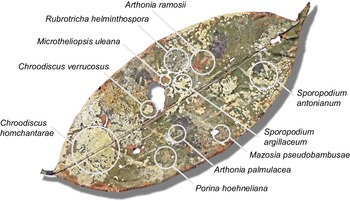
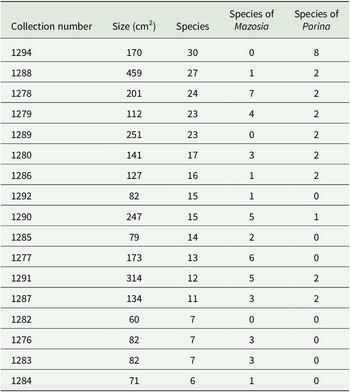
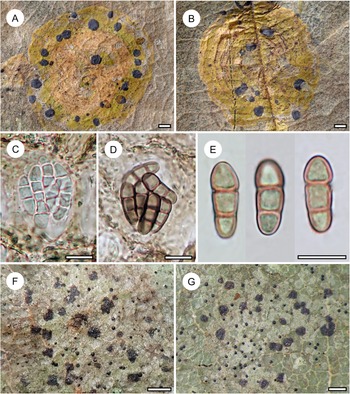

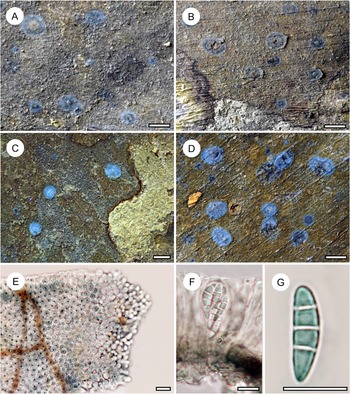
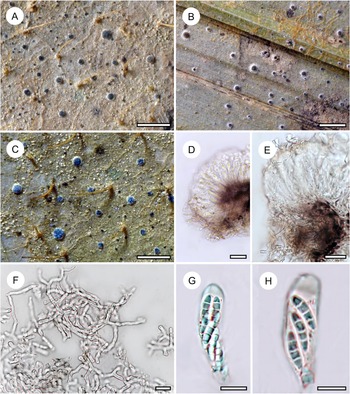
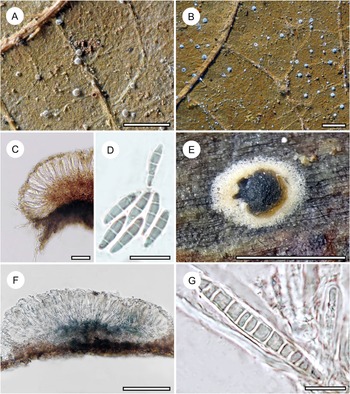
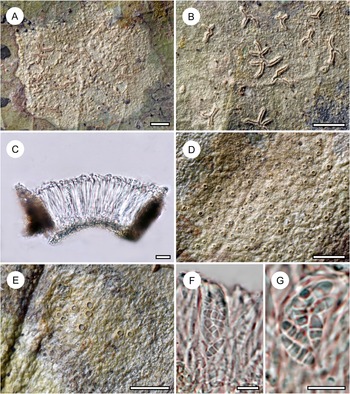
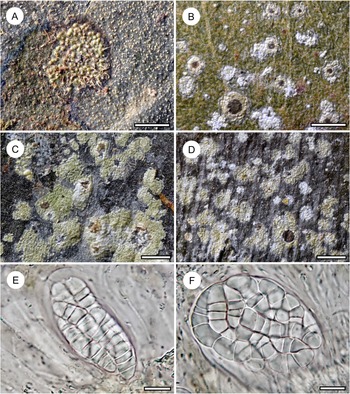
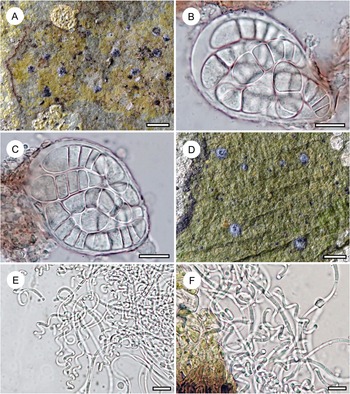

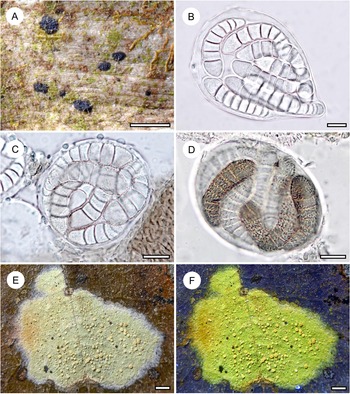
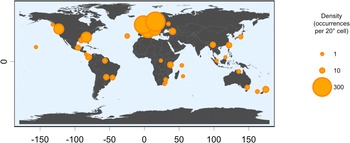
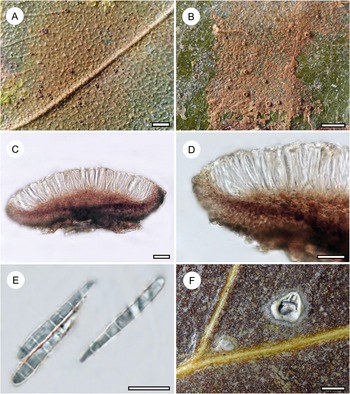
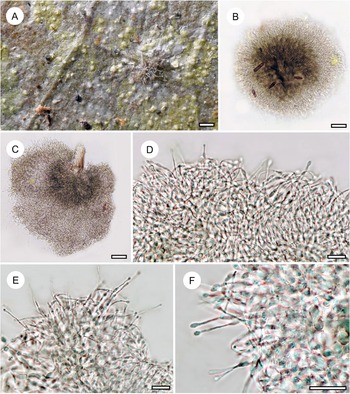
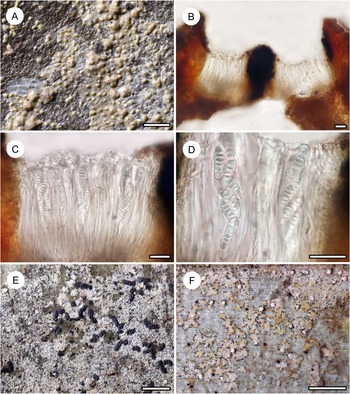
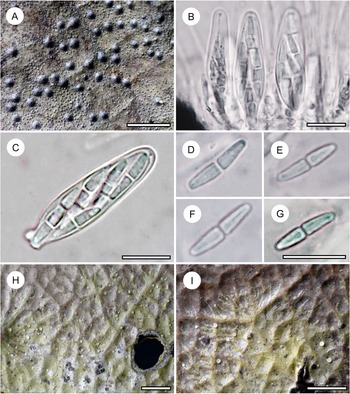

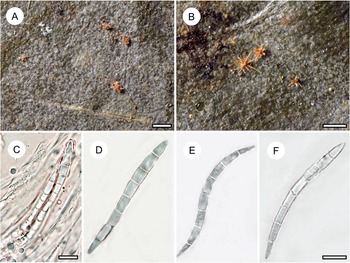

 Open access
Open access