


Introduction
Herbicide resistance represents one of the clearest and most rapidly replicated examples of contemporary evolution in managed ecosystems, with Palmer amaranth (Amaranthus palmeri S. Watson) standing out as one of the most evolutionarily enabled and concerning species due to its interference potential (yield and/or harvest quality loss), fecundity, outcrossing nature, and extensive exposure to selection (Baucom Reference Baucom2019; MacRae et al. Reference MacRae, Webster, Sosnoskie, Culpepper and Kichler2013; Morgan et al. Reference Morgan, Baumann and Chandler2001; Oerke Reference Oerke2006; Powles and Yu Reference Powles and Yu2010; Rowland et al. Reference Rowland, Murray and Verhalen1999; Smith et al. Reference Smith, Baker and Steele2000). Resistance in A. palmeri continues to accelerate and diversify, often masking differences among management programs. Recent reports document emerging resistance to newer cotton (Gossypium hirsutum L.) herbicides, including dicamba and glufosinate (Foster and Steckel Reference Foster and Steckel2022; Heap Reference Heap2025; Priess et al. Reference Priess, Norsworthy, Godara, Mauromoustakos, Butts, Roberts and Barber2022). Mechanistically, resistance frequently arises from minor nucleotide alterations, although large structural variants, such as extrachromosomal circular DNA, have been identified, amplifying target-site loci and conferring co-resistance to key chemistries like glyphosate and glufosinate, thereby complicating stewardship and program durability (Carvalho-Moore et al. Reference Carvalho-Moore, Borgato, Cutti, Porri, Meiners, Lerchl, Norsworthy and Patterson2025). Despite these issues, many weed management programs prioritize short-term, in-season suppression over long-term population and seedbank dynamics (Chauhan and Johnson Reference Chauhan and Johnson2010), which can obscure the underlying processes driving resistance.
Integrated weed management (IWM) is widely recognized as a durable approach, because it distributes mortality across weed life stages and reduces reliance on any single herbicide mechanism (Liebman and Gallandt Reference Liebman, Gallandt and Jackson1997; Mortensen et al. Reference Mortensen, Egan, Maxwell, Ryan and Smith2012; Swanton and Weise Reference Swanton and Weise1991). However, adoption is inconsistent, partly because tactics that mitigate long-term risk, such as preventing seed rain, diversifying rotations, and incorporating nonchemical methods, impose immediate costs and complexity not offset by typical commodity margins (Beckie and Harker Reference Beckie and Harker2017; Riar et al. Reference Riar, Norsworthy, Steckel, Stephenson, Eubank, Bond and Scott2013). Broad syntheses indicate that maintaining efficacy depends more on systems-level designs that limit evolutionary opportunities and tactic dependence than on herbicides with consistently high efficacy (Bagavathiannan and Davis Reference Bagavathiannan and Davis2018; Baucom Reference Baucom2019; MacLaren et al. Reference MacLaren, Storkey, Menegat, Metcalfe and Dehnen-Schmutz2020; Mortensen et al. Reference Mortensen, Egan, Maxwell, Ryan and Smith2012; Norsworthy et al. Reference Norsworthy, Ward, Shaw, Llewellyn, Nichols, Webster, Bradley, Frisvold, Powles and Burgos2012; Powles et al. Reference Powles, Preston, Bryan and Jutsum1997).
A critical yet underappreciated factor in resistance evolution is the role of preemergence herbicide escapes. Preemergence herbicide escapes represent the cohort exposed to postemergence herbicides, serving as the substrate for resistance evolution (Neve et al. Reference Neve, Norsworthy, Smith and Zelaya2011; Powles and Yu Reference Powles and Yu2010). Even when postemergence control seems effective for yield protection, repeated exposure of a persistent cohort can hasten resistance if survivors disproportionately replenish the seedbank (Evans et al. Reference Evans, Tranel, Hager and Schutte2016; Gressel and Segel Reference Gressel and Segel1978). Thus, programs minimizing preemergence escape numbers and duration may better mitigate evolutionary risk than those focused solely on end-of-season density. While preemergence herbicide resistance has been documented in A. palmeri and other species, it remains rare compared with postemergence resistance, owing to diffuse exposure, lower selection intensity, and reduced survival of tolerant individuals under soil-applied residuals (Heap Reference Heap2025; Powles and Yu Reference Powles and Yu2010). In contrast, postemergence herbicides impose intense, repeated selection on emerged cohorts under near-lethal conditions across broad areas, accelerating adaptation (Beckie and Reboud Reference Beckie and Reboud2009; Neve et al. Reference Neve, Norsworthy, Smith and Zelaya2011). Therefore, while preemergence herbicides are effective, the survivors exposed to postemergence applications constitute a key, often overlooked determinant of resistance risk.
Several IWM tactics influence preemergence escape dynamics via distinct mechanisms. Preventing seed rain directly limits seedbank replenishment and accelerates population decline when enforced consistently (Schwartz et al. Reference Schwartz, Norsworthy, Young, Bradley, Kruger, Davis, Steckel, Loux and Conley2016). Given the extreme fecundity (Bond and Oliver Reference Bond and Oliver2006; Horak and Loughin Reference Horak and Loughin2000; Sellers et al. Reference Sellers, Smeda, Johnson, Kendig and Ellersieck2003) and rapid rebound capacity of A. palmeri, this tactic serves as a primary lever for long-term suppression and resistance mitigation. Field studies show significant seed retention at harvest, supporting near-zero seed strategies such as harvest interception or zero-tolerance management (Barber et al. Reference Barber, Smith, Scott, Norsworthy and Vanglider2015), which reduce the system’s evolutionary memory (Norsworthy et al. Reference Norsworthy, Griffith, Griffin, Bagavathiannan and Gbur2014; Schwartz et al. Reference Schwartz, Norsworthy, Young, Bradley, Kruger, Davis, Steckel, Loux and Conley2016). Deep inversion tillage, dependent on existing seedbank structure, redistributes seeds vertically, curbing emergence from buried cohorts and enhancing decay (Aulakh et al. Reference Aulakh, Price, Enloe, van Santen, Wehtje and Patterson2012; Jha and Norsworthy Reference Jha and Norsworthy2009; Mohler Reference Mohler, Liebman, Mohler and Staver2001; Yenish et al. Reference Yenish, Doll and Buhler1992). Cover crops like cereal rye (Secale cereale L.) suppress early emergence through mulch effects, microclimate changes, and allelopathy (DeVore et al. Reference DeVore, Norsworthy and Brye2012; Mirsky et al. Reference Mirsky, Ryan, Teasdale, Curran, Reberg-Horton, Spargo, Wells, Keene and Moyer2013; Teasdale and Mohler Reference Teasdale and Mohler2000). Dicamba-inclusive programs improve control and add herbicide diversity (Inman et al. Reference Inman, Jordan, York, Jennings, Monks, Everman, Bollman, Fowler, Cole and Soteres2016; Johnson et al. Reference Johnson, Young, Matthews, Marquardt, Slack, Bradley, York, Culpepper, Hager, Al-Khatib, Steckel, Moechnig, Loux, Bernards and Smeda2010), although resistance to synthetic auxins in A. palmeri is a growing concern (Foster and Steckel Reference Foster and Steckel2022; Heap Reference Heap2025; Priess et al. Reference Priess, Norsworthy, Godara, Mauromoustakos, Butts, Roberts and Barber2022). Notably, the evolutionary value of these tactics lies in regulating the cohort subjected to postemergence selection; programs reducing preemergence escapes, even if densities converge later, can alter cumulative evolutionary pressure in postemergence-reliant systems.
Despite extensive research on individual IWM components, their multiyear interactive effects on both demographic trajectories and cumulative selection pressure remain insufficiently resolved, representing a critical gap in our understanding of long‑term weed population dynamics. Stacked tactics may not yield strictly additive effects, as low densities impose biological limits leading to diminishing returns (MacLaren et al. Reference MacLaren, Storkey, Menegat, Metcalfe and Dehnen-Schmutz2020; Mortensen et al. Reference Mortensen, Egan, Maxwell, Ryan and Smith2012).
This study addresses these gaps by quantifying (1) how distinct IWM components influence temporal trajectories of preemergence escapes in A. palmeri over 5 yr, (2) cumulative preemergence escape pressure as a proxy for long-term postemergence selection exposure, and (3) legacy effects of IWM systems on weed emergence after management withdrawal. A functional description of each IWM component and its hypothesized ecological role is provided in Table 1. By linking temporal suppression, cumulative exposure, and seedbank outcomes, this work offers mechanistic insights into IWM, where mortality is distributed across independent bottlenecks to enhance complementarity and system resilience.
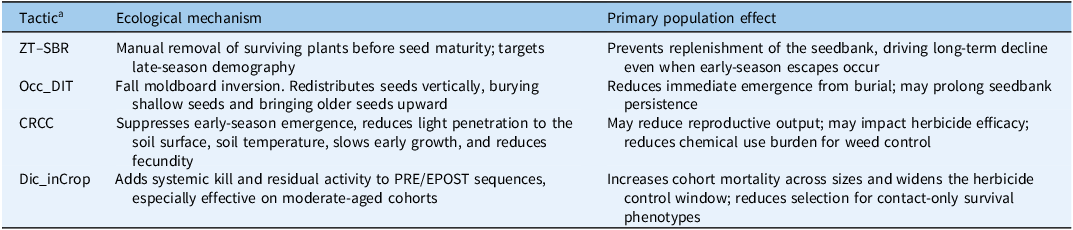
Generalized functional description of integrated weed management (IWM) tactics included in this study.

Table 1 Long description
A table comparing integrated weed management tactics, their ecological mechanisms, and primary population effects. The table has three columns: Tactic, Ecological mechanism, and Primary population effect. It contains four rows of data. Row 1: Tactic, ZT-SBR; Ecological mechanism, Manual removal of surviving plants before seed maturity; targets late-season demography; Primary population effect, Prevents replenishment of the seedbank, driving long-term decline even when early-season escapes occur. Row 2: Tactic, Occ_DIT; Ecological mechanism, Fall moldboard inversion. Redistributes seeds vertically, burying shallow seeds and bringing older seeds upward; Primary population effect, Reduces immediate emergence from burial; may prolong seedbank persistence. Row 3: Tactic, CRCC; Ecological mechanism, Suppresses early-season emergence, reduces light penetration to the soil surface, soil temperature, slows early growth, and reduces fecundity; Primary population effect, May reduce reproductive output; may impact herbicide efficacy; reduces chemical use burden for weed control. Row 4: Tactic, Dic_inCrop; Ecological mechanism, Adds systemic kill and residual activity to PRE/EPOST sequences, especially effective on moderate-aged cohorts; Primary population effect, Increases cohort mortality across sizes and widens the herbicide control window; reduces selection for contact-only survival phenotypes.
a The descriptions provided are generalized based on established ecological principles to frame the hypotheses and provide context for the design of the management systems analyzed in this manuscript. The observed field data may reflect the complex interactions of these mechanisms. ZT–SBR, zero tolerance for seedbank replenishment; OCC_DIT, occasional deep inversion tillage; CRCC, cereal rye cover crop; Dic_inCrop; dicamba-inclusive herbicide program; PRE, preemergence; EPOST, early postemergence.
Materials and Methods
Experimental Design and Site
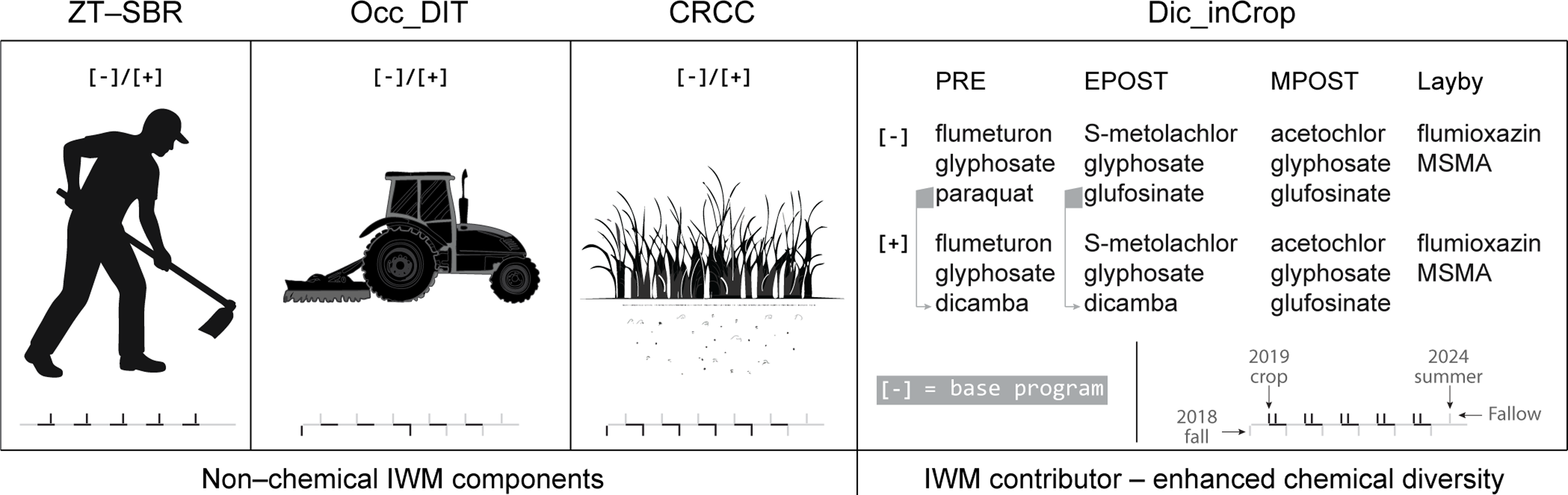
A large-plot, multiyear cotton field experiment was initiated in fall 2018 at the Lon Mann Cotton Research Station near Marianna, AR (34.73°N, 90.74°W) on a Convent silt loam soil (organic matter and texture reported in Farr et al. [Reference Farr, Norsworthy, Kouame, Barber, Butts and Roberts2022]). The study was arranged as a randomized complete block design with four replications in a split–split–split–split plot structure. The whole-plot factor was zero tolerance for A. palmeri seedbank replenishment (ZT–SBR; present/absent), the subplot factor was occasional deep inversion tillage (Occ_DIT; present/absent), the sub-subplot factor was cereal rye cover crop (CRCC; present/absent), and the sub-sub-subplot factor was the in-crop herbicide program (dicamba-inclusive vs. non-dicamba). Individual sub-sub-subplots measured approximately 37 m by 8 m, accommodating eight cotton rows on 1-m centers. All treatments were maintained on the same plots for the duration of the experiment (2019 to 2023), with temporal implementation of components shown in Figure 1.
Schematic representation of the 5-yr integrated weed management (IWM) experiment in cotton (2019–2023) followed by a legacy year (2024). Four management components were factorially combined: zero tolerance for Amaranthus palmeri seedbank replenishment (ZT–SBR), occasional deep inversion tillage (Occ_DIT; fall 2018 and fall 2021), cereal rye cover crop (CRCC), and dicamba-based herbicide program (Dic_inCrop). Herbicide applications consisted of layered preemergence and postemergence mixtures applied at preemergence, early postemergence (early POST), mid-postemergence (mid-POST), and layby timings. The base program (all factors absent) is indicated for reference. Nonchemical IWM components (ZT–SBR, Occ_DIT, CRCC) and enhanced chemical diversity via Dic_inCrop are expected to distinctly structure demography and act as evolutionary filters.

Treatment Implementation
Occ_DIT was implemented via moldboard inversion to 20 to 25 cm, a depth selected to reposition near‑surface seed into zones of reduced germination potential. Inversions were carried out at study initiation in fall 2018 and again in fall 2021, after which plots were bedded and managed as no‑till between inversion events.
The CRCC treatment consisted of ‘Wrens Abruzzi’ cereal rye drill-seeded in the fall at 84 kg ha−1, then chemically terminated approximately 21 d before cotton planting using glyphosate (1,260 g ae ha−1) plus dicamba (560 g ae ha−1), consistent with Farr et al. (Reference Farr, Norsworthy, Kouame, Barber, Butts and Roberts2022).
The two herbicide programs each consisted of a preplant burndown, followed by preemergence at planting, early postemergence at ∼21 d after planting (DAP), mid-postemergence at ∼42 DAP, and a layby post-directed application at ∼63 DAP (specific products and rates listed in treatment tables in Farr et al. [Reference Farr, Norsworthy, Kouame, Barber, Butts and Roberts2022] and the present paper’s treatment summary). Burndown, preemergence, and early postemergence applications were applied at 140 L ha−1 using a self-propelled sprayer (Bowman Manufacturing, Newport, AR, USA); mid-postemergence was applied using a tractor-mounted hooded sprayer, and layby applications were applied using a tractor-mounted post-directed sprayer (also at 140 L ha−1). Applications containing dicamba used TTI 11006 nozzles, layby applications used XR 11006E nozzles, and other applications used AIXR 11006 nozzles (TeeJet® Technologies, Wheaton, IL, USA).
ZT–SBR was operationalized as a single, targeted hand-removal event after layby, conducted approximately 14 d after the layby application, to eliminate surviving A. palmeri individuals before seed return (timing and execution consistent with Farr et al. [Reference Farr, Norsworthy, Kouame, Barber, Butts and Roberts2022]).
Data Collection
Weed emergence was quantified using four fixed 1-m2 quadrats per plot, with A. palmeri counts collected at key points aligned with herbicide passes (e.g., before early postemergence, mid-postemergence, layby, and before the ZT–SBR hand-removal event), consistent with Farr et al. (Reference Farr, Norsworthy, Kouame, Barber, Butts and Roberts2022). In the present study, preemergence escapes were defined as emerged A. palmeri individuals that survived the residual (“preemergence-layer”) component of the preceding pass and were therefore present immediately before the next application; counts within the four quadrats were summed to obtain plot-level estimates for each pass.
In Year 6 (2024), no crop or weed management practices were imposed to isolate residual seedbank effects from contemporary control. Natural emergence was monitored by counting weeds within the same four fixed 1-m2 quadrats per plot in mid-May, June, and July.
Statistical Analysis
All statistical analyses were conducted using R statistical software (v. 4.5.1; R Core Team 2025) within the RStudio integrated development environment (RStudio Team). Generalized linear mixed-effects models (GLMMs) (Stroup Reference Stroup2015) and nonlinear regression were employed using the following primary packages: glmmTMB 1.1.9 for GLMMs (Brooks et al. Reference Brooks, Kristensen, van Benthem, Magnusson, Berg, Nielsen, Skaug, Mächler and Bolker2017), DHARMa 0.4.7 for model diagnostics, multcomp 1.4-26 (Hothorn et al. Reference Hothorn, Bretz and Westfall2008), sjPlot 2.8.16 (Lüdecke Reference Lüdecke2024), emmeans 1.10.4 for post hoc comparisons (Lenth Reference Lenth2022), and randomForest 4.7-1.1 for variable importance. The significance level for all statistical tests was set at α = 0.05.
Season-total plot-level A. palmeri preemergence escape totals (2019 to 2023) were expressed as relative density (plot total as a percentage of the year-specific base program mean) to normalize annual variability and facilitate interpretation of long-term suppression. Treatment effects on relative density were analyzed using GLMMs fit with a Tweedie (compound Poisson–gamma) distribution and log link, which accommodates nonnegative, right-skewed responses and allows for strong suppression producing near-zero values. Replication and plot were incorporated as random effects, and a first-order autoregressive correlation structure [AR(1)] was specified within plots across years where supported by model convergence and diagnostics.
Competing temporal models were compared using information criteria (Akaike information criterion [AIC]/Bayes information criterion [BIC]) and predictive error (root mean-square error [RMSE]), contrasting a full factorial model (Year × ZT–SBR × Occ_DIT × CRCC × Dic_inCrop) against a parsimonious model focused on the dominant temporal driver (Year × Occ_DIT). The pooled Occ_DIT model was retained for primary inference, because it provided improved generalization and predictive performance while preserving the key biological contrast of interest. To evaluate nonlinear depletion dynamics consistent with seedbank-driven population responses, a quadratic temporal model was fit using orthogonal polynomial terms for year [poly(Year, 2)] and their interaction with Occ_DIT under a Tweedie (log) GLMM framework.
For cumulative preemergence escape pressure across the five managed years, plot-level totals were calculated by summing yearly relative densities (% of yearly base-program mean), and treatment effects were evaluated using a four-way Tweedie (log) GLMM (ZT–SBR × Occ_DIT × CRCC × Dic_inCrop), with replication as a random effect.
For the legacy year (2024), total A. palmeri counts were analyzed using separate GLMMs on the count scale to quantify residual effects of the multiyear IWM history. A factorial model tested ZT–SBR × Occ_DIT × CRCC × Dic_inCrop, with replication and nested random effects reflecting the split-plot structure. In parallel, random forest models were used to provide a nonparametric ranking of component importance using %IncMSE and node purity metrics.
Model assumptions were evaluated using simulation-based residual diagnostics (DHARMa). Estimated marginal means (EMMs) (Searle et al. Reference Searle, Speed and Milliken1980) were computed on the response scale from fitted models, and mean separation was summarized using compact letter displays (CLDs) derived from EMM contrasts. Unless otherwise noted, CLDs were generated without multiplicity adjustment to provide descriptive grouping, while primary inference was based on fixed-effect tests from the fitted GLMMs.
Results and Discussion
Results are presented in three parts: (1) temporal dynamics of preemergence escape trajectories during active management (2019 to 2023), (2) cumulative preemergence escape pressure as an integrated proxy for postemergence selection exposure, and (3) unmanaged legacy emergence in 2024 as the ecological endpoint of each system.
Temporal Dynamics of Preemergence Escapes
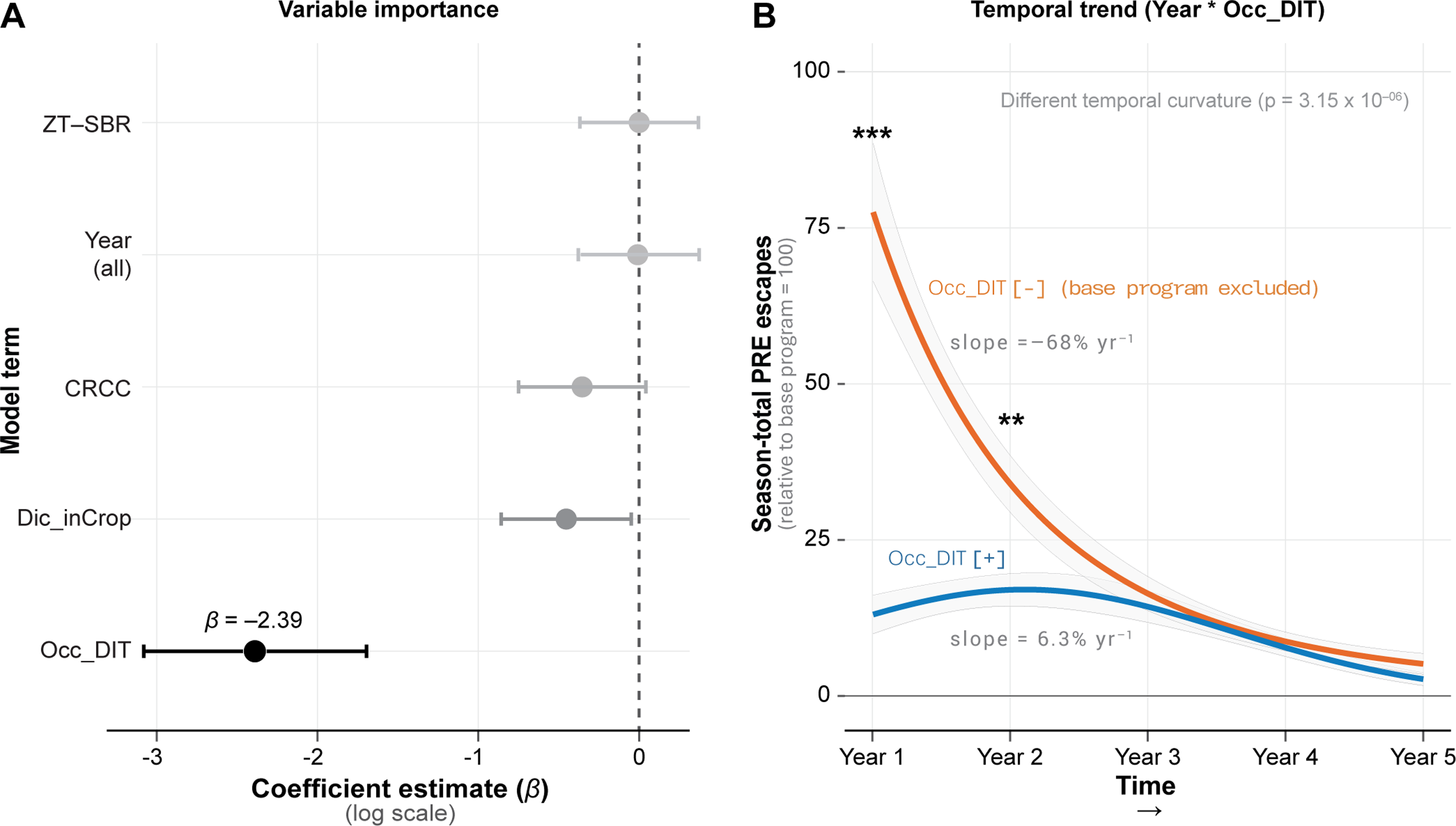
Model comparison for annual preemergence escape totals strongly favored a reduced model in which Occ_DIT was the only management factor allowed to interact with year (ΔBIC = 111.3; RMSE reduced from 8.74 to 6.76) over the full four-factor model, indicating that Occ_DIT largely structured population trajectories through time. Among the four IWM components, Occ_DIT was also the most influential main effect, with the largest negative coefficient on the log scale (β = −2.39; Figure 2A).
Temporal dynamics of season-total Amaranthus palmeri preemergence (PRE) escapes (relative to the base program) from 2019 to 2023. (A) Fixed-effect coefficient estimates (β, log scale) from the full generalized linear mixed-effects model (GLMM) highlighting the dominant influence of Occ_DIT. (B) Modeled (from temporal Occ_DIT only model) trajectories with 95% confidence band (relative to the base program) for systems without Occ_DIT (orange line) and with Occ_DIT (blue line). Strong Year × Occ_DIT interaction (P = 3.4 × 10⁻⁶) indicates fundamentally different temporal curvature. Slopes shown represent instantaneous rates of decline derived from the fitted model. CRCC, cereal rye cover crop; Dic_inCrop, dicamba-based herbicide program; Occ_DIT, occasional deep inversion tillage in fall 2018 and fall 2021; ZT–SBR, zero tolerance for seedbank replenishment.

The quadratic reduced model showed a highly significant Year × Occ_DIT interaction (P = 3.4 × 10−⁶), revealing divergent temporal pathways between inversion tillage regimes (Figure 2B). Systems without Occ_DIT began with high preemergence escape densities (∼75% of the base program in 2019) and required repeated annual suppression to achieve steep declines (instantaneous slope = −68% in 2019). In contrast, Occ_DIT systems exhibited an immediate reduction in preemergence escapes (12% of the base program in 2019) and rapidly approached a low-density equilibrium, with rates stabilizing in later years. Although both systems converged toward low preemergence escape densities by 2023, their routes differed markedly: Occ_DIT delivered a strong front-loaded reduction rather than a gradual depletion trajectory.
The dominant role of Occ_DIT aligns with its function as a seedbank redistribution tactic rather than a continuous suppression tool. Deep inversion tillage buries surface-resident seeds to depths where germination is constrained and mortality increases (Mohler Reference Mohler, Liebman, Mohler and Staver2001; Yenish et al. Reference Yenish, Doll and Buhler1992), effectively resetting the near-surface seedbank early in the IWM sequence. This temporal advantage also has evolutionary consequences. While non–Occ_DIT systems achieved comparable suppression by Year 3, they sustained much higher densities during the first 2 yr, increasing the cumulative number of individuals exposed to postemergence selection. Because resistance evolution depends on repeated exposure across generations, early suppression via Occ_DIT represents a disproportionate reduction in long-term selection pressure, even when final-year control appears similar. Thus, differences in the rate of decline matter: systems without Occ_DIT carried a larger cumulative burden of preemergence survivors—the substrate for resistance evolution. Despite this demonstrated efficacy, adoption of deep tillage in Midsouth cotton production remains limited, with primary barriers including the high capital cost of moldboard plowing equipment; concerns about soil structural disruption and erosion risk on silty soils; and the perception that performance benefits accrue over multiple seasons, a timescale poorly aligned with typical annual farm decision-making cycles (Aulakh et al. Reference Aulakh, Price, Enloe, van Santen, Wehtje and Patterson2012; DeVore et al. Reference DeVore, Norsworthy and Brye2012).
Five-Year Cumulative Preemergence Escape Pressure
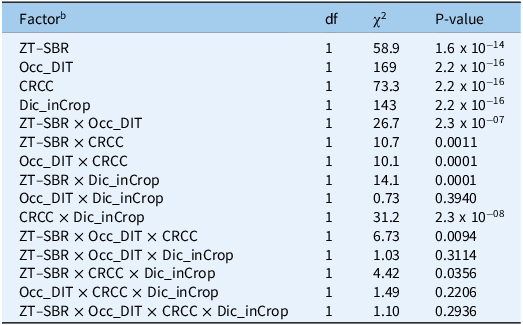
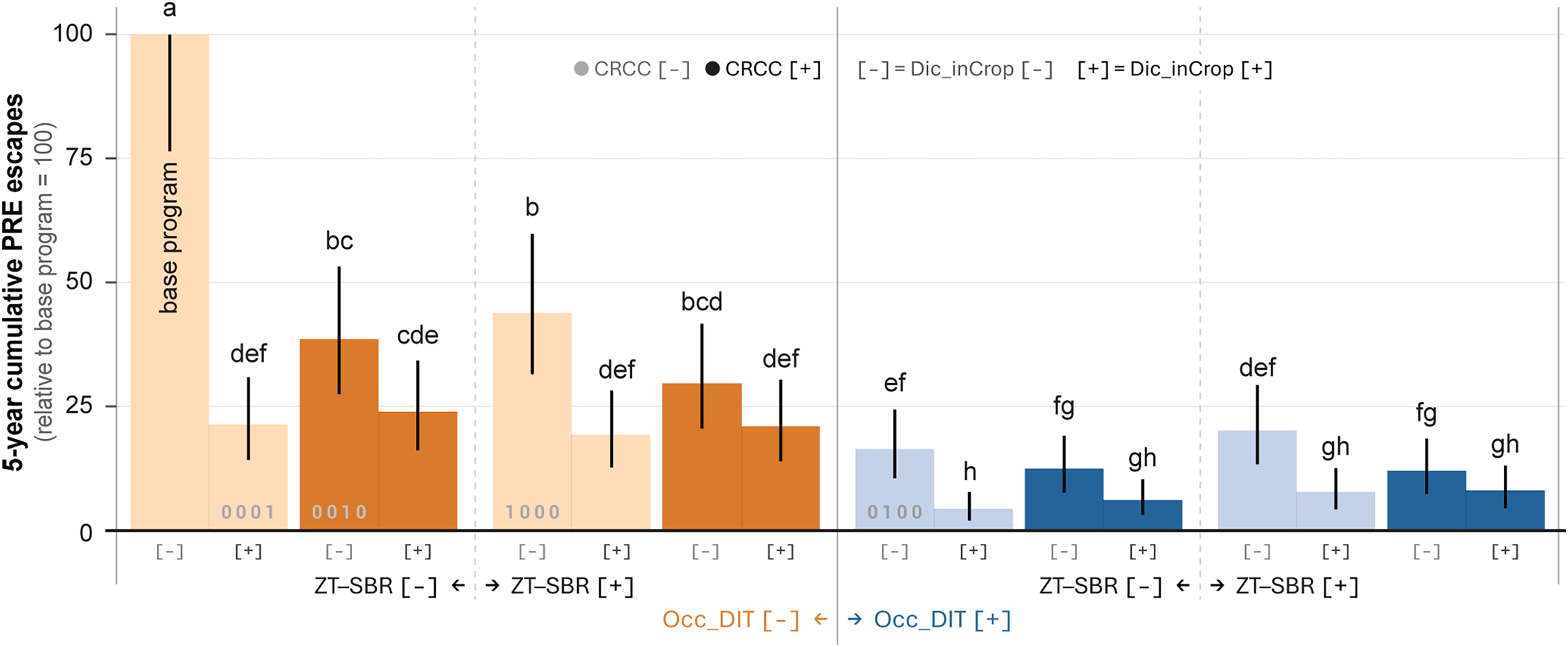
Cumulative preemergence escape pressure over the 5-yr imposition phase (2019 to 2023) quantifies the total “evolutionary load” placed on the postemergence herbicide program. This metric (sum of relative preemergence escapes across years) provides a direct measure of the population repeatedly entering the postemergence selection funnel. The 5-yr total was strongly affected by all four IWM components (Table 2). Occ_DIT exerted the largest main effect (χ2 = 169, P < 2.2 × 10−1⁶), followed by Dic_inCrop (χ2 = 143, P < 2.2 × 10−1⁶), CRCC (χ2 = 73, P = 2.2 × 10−1⁶), and ZT–SBR (χ2 = 58, P = 1.6 × 10−1⁴). Two-way interactions were also significant, particularly ZT–SBR × Occ_DIT (P = 2.3 × 10−⁷) and CRCC × Dic_inCrop (P = 2.3 × 10−⁸), indicating that cumulative exposure depended on tactic combinations rather than additive effects alone.
Type III Wald χ2 tests from a Tweedie(log) GLMM for cumulative Amaranthus palmeri preemergence escapes summed across 5 yr (2019–2023) under continuous IWM imposition in cotton near Marianna, AR. a

Table 2 Long description
A table with five rows and nine columns. The columns are labeled Factor, df, chi-squared, and P-value. The rows list different factors and their interactions, along with their degrees of freedom, chi-squared values, and P-values. The factors include ZT-SBR, Occ_DIT, CRCC, Dic_inCrop, and various interactions among them. Notable interactions include ZT-SBR x Occ_DIT and CRCC x Dic_inCrop, which show significant effects on cumulative preemergence escape pressure.
a Abbreviations: df, degrees of freedom; GLMM, generalized linear mix-effects model; IWM, integrated weed management; PRE, preemergence herbicide (residual) application.
b ZT–SBR, zero tolerance for seedbank replenishment; OCC_DIT, occasional deep inversion tillage; CRCC, cereal rye cover crop; Dic_inCrop; dicamba-inclusive herbicide program.
Four-way mean separation showed that systems containing Occ_DIT consistently achieved the lowest cumulative escape loads, regardless of ZT–SBR or Dic_inCrop status (Figure 3). The strongest combinations, typically Occ_DIT paired with Dic_inCrop, reduced the total number of weeds exposed to postemergence selection by >92% to 95% across 5 yr. While CRCC alone was highly effective, the CRCC × Dic_inCrop interaction further reduced preemergence escape load. These patterns were also visually apparent at mid-postemergence in Year 5: sub-sub-subplot aerial imagery captured June 19, 2023, showed distinct weed cover between the base program versus CRCC-only, Dic_inCrop-only, and CRCC + Dic_inCrop treatments (Figure 4).
Five-year cumulative preemergence herbicide (residual) application (PRE) escape pressure (sum of annual relative escapes) across all combinations of ZT–SBR, Occ_DIT, CRCC, and Dic_inCrop. Values are expressed relative to the annual base program mean and summed across 5 yr. Bars represent estimated marginal means (EMMs) from a four fixed factor generalized linear mixed-effects model (GLMM), with compact letter display (CLD) groupings indicating statistical separation (α = 0.05). Treatments are ordered to illustrate the suppression gradient. This cumulative metric serves as a proxy for accumulated postemergence selection pressure. CRCC, cereal rye cover crop; Dic_inCrop, dicamba-based herbicide program; Occ_DIT, occasional deep inversion tillage in fall 2018 and fall 2021; ZT–SBR, zero tolerance for seedbank replenishment; 1000, ZT–SBR only; 0100, Occ_DIT only; 0010, CRCC only; 0001, Dic_inCrop only.

Aerial imagery illustrating spatial arrangement of subplots within a representative block under ZT–SBR⁻ and Occ_DIT⁻ conditions. Sub-sub-subplots depict base program, CRCC only, Dic_inCrop only, and CRCC + Dic_inCrop treatments. The image highlights the accumulated multiyear effects evident in Year 5 of single or two-way integrated weed management (IWM) components on early-season weed population (June 19, 2023, at the time of mid-postemergence). CRCC, cereal rye cover crop; Dic_inCrop, dicamba-based herbicide program; Occ_DIT, occasional deep inversion tillage in fall 2018 and fall 2021; ZT–SBR, zero tolerance for seedbank replenishment.

Figure 4 Long description
Panel A: A line graph showing the spatial arrangement of subplots within Block 3. The horizontal axis represents distance in meters ranging from 0 to 36.6 meters. The vertical axis represents distance in meters ranging from 0 to 7.6 meters. The graph includes four lines representing different treatments: No IWM, base program + Dic_inCrop, base program + CRCC, and base program + CRCC + Dic_inCrop. Panel B: A line graph showing the spatial arrangement of subplots within Block 4. The horizontal axis represents distance in meters ranging from 0 to 36.6 meters. The vertical axis represents distance in meters ranging from 0 to 7.6 meters. The graph includes four lines representing different treatments: No IWM, base program + Dic_inCrop, base program + CRCC, and base program + CRCC + Dic_inCrop.
This cumulative metric provides a mechanistic distinction from the legacy analysis (see following section). The dominance of Occ_DIT over ZT–SBR in the cumulative count (χ2 = 169 vs. 58) reflects the timescale required for each mechanism to reshape the system. Occ_DIT provides an immediate demographic reset by burying the existing seedbank, shielding postemergence chemistry from the large emergence cohorts in Years 1 and 2. ZT–SBR is a long-term depletion strategy: it prevents seedbank recharge but requires multiple years to deplete the existing seed pool (Buhler et al. Reference Buhler, Hartzler and Forcella2001; Jha et al. Reference Jha, Norsworthy and Garcia2014). From a resistance-management perspective, the implication is substantial. Achieving >90% control of individuals repeatedly exposed to postemergence herbicides equates to an approximately 10‑fold reduction in the cohort of survivors within a single growing season. When sustained over multiple years, this level of control exerts a compounding effect, progressively diminishing the number of individuals subjected to recurrent herbicide selection. Such cumulative suppression substantially limits the opportunities for rare resistance alleles to be expressed and enriched, thereby contributing to the long‑term preservation of herbicide utility.
Legacy Emergence in the Sixth Year
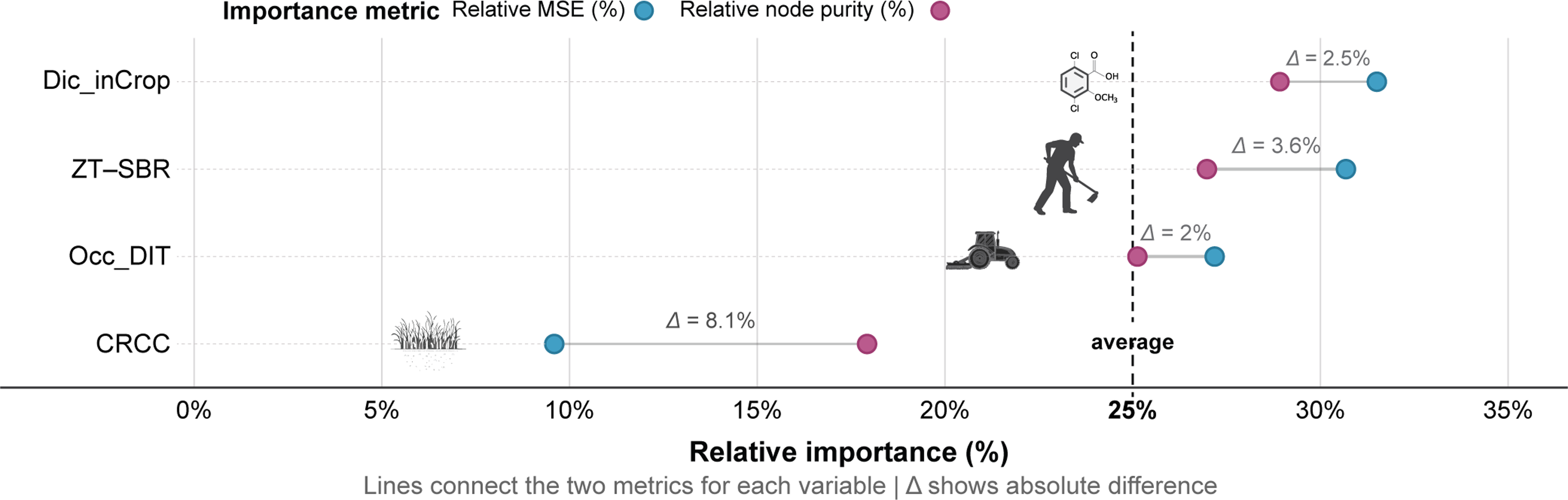
The unmanaged 2024 emergence phase measured the ecological endpoint of 5 yr of management by quantifying the residual seedbank after cessation of IWM. Random forest analysis corroborated the dominance of Dic_inCrop, ZT–SBR, and Occ_DIT as primary drivers of legacy emergence, with consistent rankings across metrics (Figure 5).
Relative importance of integrated weed management (IWM) components for predicting Amaranthus palmeri emergence counts following 5-yr continuous implementation in 2024 under fallow conditions, assessed using two different random forest importance metrics: % increase in mean squared error (%IncMSE) and increase in node purity. CRCC, cereal rye cover crop; Dic_inCrop, dicamba-based herbicide program; Occ_DIT, occasional deep inversion tillage in fall 2018 and fall 2021; ZT–SBR, zero tolerance for seedbank replenishment.

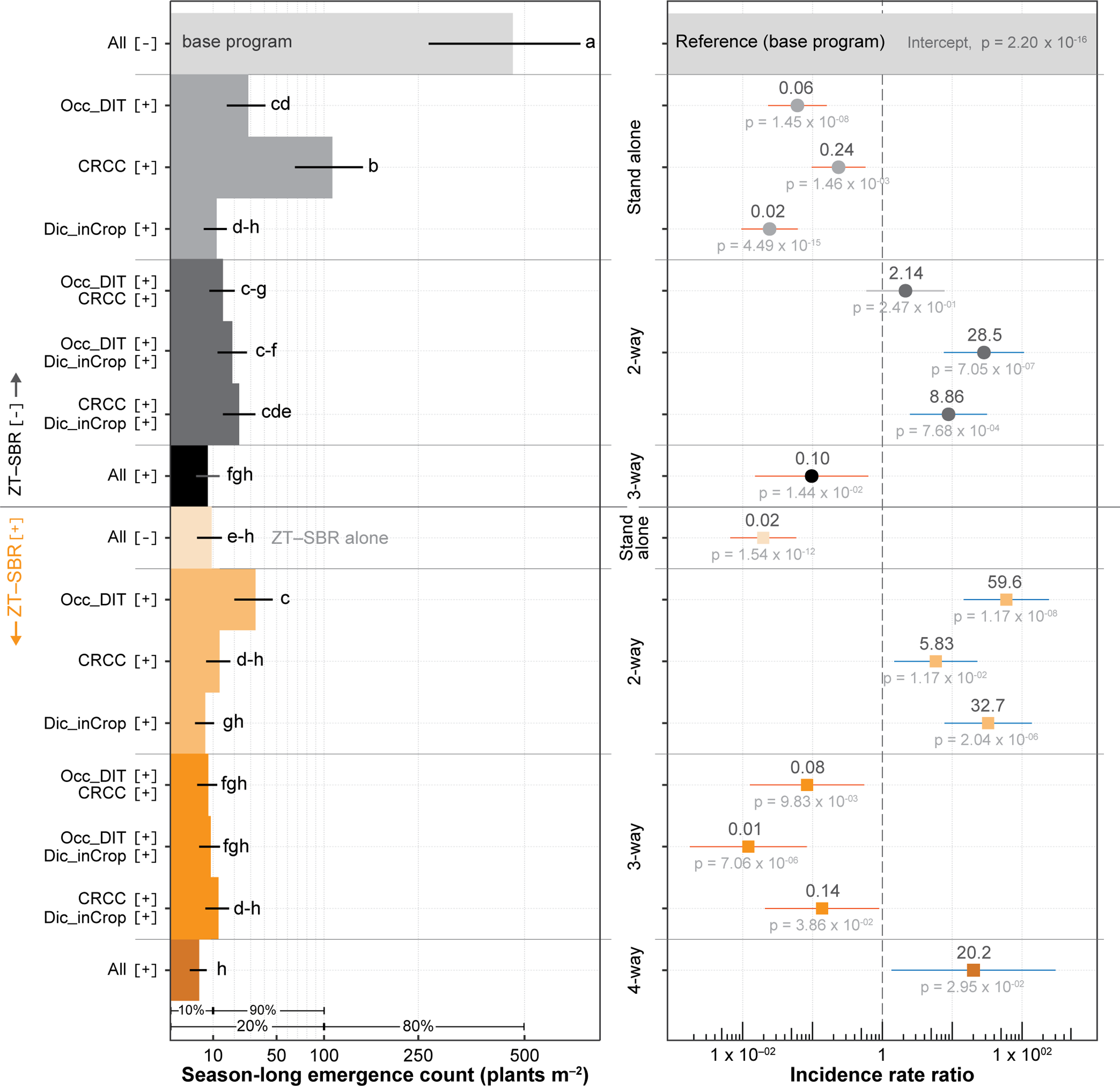
The GLMM indicated significant main effects of all four IWM components (P < 0.001; Figure 6). Coefficients are most interpretable on the exponential scale as incidence rate ratios (IRR): IRR < 1 indicates suppression relative to the reference condition, whereas IRR > 1 for interaction terms reflects departures from strict additivity on the log scale. Among main effects, ZT–SBR (IRR = 0.02, P = 1.5 × 10−1 2) and Dic_inCrop (IRR = 0.024, P < 5 × 10−1⁵) were the most suppressive components. Occ_DIT (IRR = 0.06, P = 1.4 × 10−⁸) and CRCC (IRR = 0.24, P = 0.0015) also exerted strong suppression, although of smaller magnitude. As single tactics, ZT–SBR reduced emergence to 9.2 plants m−2 (∼50-fold below the base), and Dic_inCrop reduced emergence to 11.37 plants m−2. Dunnett contrasts showed that every IWM combination, including all single tactics, significantly reduced emergence compared with the base program (P < 0.0001). The fully integrated four-component system resulted in the lowest emergence (∼5 plants m−2), representing a >99% reduction.
Legacy emergence of Amaranthus palmeri in 2024 under unmanaged conditions following 5 yr of integrated weed management (IWM). Left, estimated marginal means (EMMs) with compact letter display (CLD) groupings (plants m⁻2). Right, corresponding incidence rate ratios (IRRs) from the four-way generalized linear mixed-effects model (GLMM), displayed on a logarithmic scale. IRR < 1 indicates suppression relative to the base program. Positive-interaction IRRs reflect diminishing marginal returns as populations approach a biological floor, rather than loss of efficacy. Collectively, this figure demonstrates how apparent antagonism in interaction terms arises from repartitioning losses across the same finite population. CRCC, cereal rye cover crop; Dic_inCrop, dicamba-based herbicide program; Occ_DIT, occasional deep inversion tillage in fall 2018 and fall 2021; ZT–SBR, zero tolerance for seedbank replenishment.

Figure 6 Long description
Panel A: A vertical bar graph shows the season-long emergence count of Amaranthus palmeri plants per square meter under various management strategies. The horizontal axis lists different combinations of strategies, including base program, cover crop, herbicide program, tillage, and seedbank management. The vertical axis represents the emergence count. The bars are grouped and labeled with letters indicating statistical significance. Panel B: A scatter plot displays the incidence rate ratios on a logarithmic scale for different management strategies. The horizontal axis represents the incidence rate ratio, and the vertical axis categorizes the strategies into stand alone, 2-way, 3-way, and 4-way combinations. Each point is labeled with a numerical value and a p-value indicating statistical significance. The reference base program is marked with a dashed vertical line at an incidence rate ratio of 1.
Several two-way interaction terms were positive on the IRR scale, including ZT–SBR × Occ_DIT (IRR = 59.6, P = 1.17 × 10−⁸) and Occ_DIT × Dic_inCrop (IRR = 28.5, P = 7.05 × 10−⁷). These coefficients do not indicate biological antagonism; they reflect diminishing marginal returns as multiple tactics act on the same finite pool of potential survivors. Once emergence is driven toward a biological minimum by one or two highly effective components, additional tactics cannot reduce counts proportionally, resulting in non-additivity on the multiplicative scale.
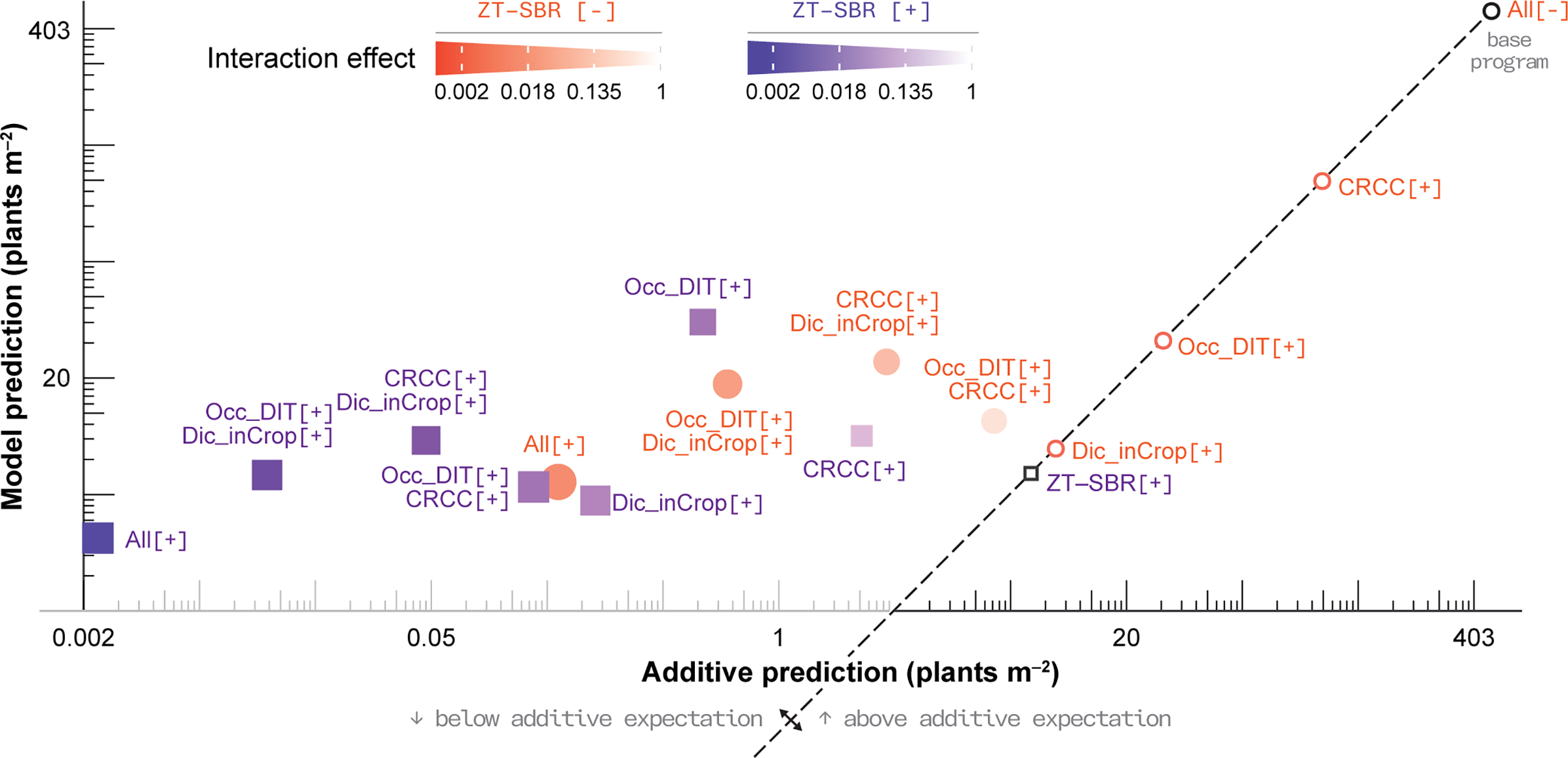
This diminishing-returns pattern is evident in the interaction surface (Figure 7): multi-tactic systems fall below the 1:1 line expected under purely additive suppression. In effect, losses are repartitioned across a shrinking pool of potential escapes rather than compounded indefinitely. These results matter for optimization: maximal suppression and maximal economic efficiency may not coincide once systems operate near the ecological floor of emergence. Economic analyses from companion work indicate that stacked tactics can return value primarily through sustained cotton yield and reduced herbicide reliance over 5 yr (Godar et al. Reference Godar, Norsworthy, Barber, Farr and Smith2025).
Comparison of additive predictions vs. full generalized linear mixed-effects model (GLMM) model predictions for legacy Amaranthus palmeri emergence. Each point represents an integrated weed management (IWM) combination; the dashed 1:1 line denotes additive expectation based on main effects alone. Points falling below the line indicate stronger-than-additive suppression, whereas points above indicate diminishing marginal returns, illustrating how strong demographic bottlenecks constrain further absolute reductions once emergence approaches a biological minimum and/or when mechanism act on overlapping life stages. CRCC, cereal rye cover crop; Dic_inCrop, dicamba-based herbicide program; Occ_DIT, occasional deep inversion tillage in fall 2018 and fall 2021; ZT–SBR, zero tolerance for seedbank replenishment.

Preemergence escapes have direct implications for resistance evolution, because they define the cohort repeatedly exposed to postemergence selection. postemergence resistance has historically outpaced preemergence resistance (Neve et al. Reference Neve, Norsworthy, Smith and Zelaya2011; Powles and Yu Reference Powles and Yu2010), making preemergence escapes a recurring substrate upon which postemergence resistance can be built. Systems that suppress early emergence, shift emergence timing, or eliminate reproductive survivors should dilute directional selection and reduce the consistency with which resistance mechanisms confer advantage across years.
Legacy-year responses provide a clear quantification of how 5 yr of IWM reshaped the A. palmeri seedbank and the evolutionary context governing future control. Across analyses, ZT–SBR and Dic_inCrop were strong determinants of long-term suppression, consistent with the principle that durable control depends on preventing new seed input while eliminating survivors before reproduction (Sosnoskie et al. Reference Sosnoskie, Webster and Culpepper2013). CRCC also contributed to reducing legacy emergence, and this legacy signal was most apparent early in the season, when residue-mediated suppression constrained recruitment. The near weed-free CRCC plot at 2 d after early postemergence in Year 3 (Figure 8) provides visual context for the Year 3 dynamics, consistent with CRCC acting as an early-season recruitment filter that reduces preemergence escape pressure and cumulative seedbank inputs.
Sub-sub-subplots 2 d after early postemergence (EPOST) application (Year 3; June 18, 2021) comparing base program + CRCC (left) and base program + Dic_inCrop (right) treatments. The images illustrate contrasting mechanisms of early-season physical suppression via surface residue (CRCC) vs. chemical control via enhanced herbicide diversity (Dic_inCrop), providing visual context for the quantitative differences observed in preemergence escape counts. The CRCC plot is nearly weed-free, whereas emerged weeds are still present at the time of EPOST in the Dic_inCrop plot. CRCC, cereal rye cover crop; Dic_inCrop, dicamba-based herbicide program; Occ_DIT, occasional deep inversion tillage in fall 2018 and fall 2021; ZT–SBR, zero tolerance for seedbank replenishment.

The effectiveness of ZT–SBR here depended on the base program maintaining low in-season densities; under widespread control failure, strict zero tolerance would become operationally infeasible. A similar context dependence applies to Occ_DIT, the effects of which depend on the preexisting vertical distribution of viable seed. Where seed is concentrated near the surface, inversion tillage can sharply reduce emergence by burial beyond the germination zone (Mohler Reference MacRae, Webster, Sosnoskie, Culpepper and Kichler2001; Yenish et al. Reference Yenish, Doll and Buhler1992); where viable seed exists at depth, tillage can redistribute dormant seed upward and weaken suppression (Farmer et al. Reference Farmer, Bradley, Young, Steckel, Johnson, Norsworthy, Davis and Loux2017; Korres et al. Reference Korres, Norsworthy, Young, Reynolds, Johnson and Conley2018). These constraints reinforce that neither ZT–SBR nor Occ_DIT is universally deployable as a stand-alone solution; their value emerges most clearly within diversified systems that regulate both seedbank size and seedbank structure. Mechanized tactics such as harvest weed seed control (HWSC) could complement ZT–SBR by intercepting A. palmeri seed at combine harvest and reducing seedbank replenishment without the labor demands of hand removal; however, its integration into cotton harvest systems remains limited by mechanical constraints and warrants further evaluation. Regarding the influence of seed burial, previous research has demonstrated that A. palmeri seeds buried to depths ≥10 cm experience substantially reduced germination due to light exclusion and altered thermal fluctuations (Jha and Norsworthy Reference Jha and Norsworthy2009; Mohler Reference MacRae, Webster, Sosnoskie, Culpepper and Kichler2001; Yenish et al. Reference Yenish, Doll and Buhler1992), with prolonged burial also increasing the probability of decay. However, as already noted, where viable seed already exists at depth before inversion, tillage can redistribute dormant seed upward and temporarily weaken suppression (Farmer et al. Reference Farmer, Bradley, Young, Steckel, Johnson, Norsworthy, Davis and Loux2017; Korres et al. Reference Korres, Norsworthy, Young, Reynolds, Johnson and Conley2018), a constraint reinforcing that Occ_DIT is most effective when near-surface seedbank densities are high.
Overall, the sustainability of herbicide-based weed management depends not only on achieving low end-of-season densities but on minimizing cumulative postemergence selection exposure over time. Even when systems converge to similar low-density endpoints, their evolutionary risk profiles can differ substantially, because the dominant process at low density is not additive suppression but the repartitioning of mortality across independent demographic bottlenecks (seedbank redistribution, recruitment filtering, and reproductive elimination). Thus, the value of diversified IWM lies less in pushing emergence marginally closer to zero and more in distributing control across mechanisms that reduce the consistency of selection on any single pathway, thereby extending the durability of chemical tools and stabilizing long-term control.
The findings reported here carry implications beyond cotton production systems. In cropping situations where selective weed control has become heavily reliant on preemergence herbicides or where glyphosate-resistant A. palmeri populations have proliferated, the concept of a biological reset via DIT offers a compelling strategic option. A single DIT event buries the existing near-surface seedbank, often dominated by resistance-enriched biotypes, into zones of reduced germination potential, creating a demographic window during which alternative herbicide chemistries, including diversified preemergence programs, can operate against a substantially weakened emergence cohort. The broader IWM framework demonstrated here, which distributes mortality across independent demographic bottlenecks to minimize cumulative postemergence selection exposure, is directly transferable to soybean [Glycine max (L.) Merr.], corn (Zea mays L.), and grain sorghum [Sorghum bicolor (L.) Moench] systems facing similar resistance trajectories. Critically, the value of this approach lies not only in simultaneous tactic stacking but in strategic sequencing: deploying Occ_DIT to rapidly reset near-surface seedbanks when resistant populations are first detected, followed by ZT–SBR to prevent seedbank recharge, may compress the resistance reversal timeline and extend the durability of existing herbicide programs.
Acknowledgments
This publication is a contribution of the University of Arkansas System Division of Agriculture.
Funding statement
Cotton Incorporated® provided funding for his research.
Competing interests
The authors declare no conflicts of interest.



 Open access
Open access