


Introduction
Arboreal macrolichens represent interesting indicators of forest conditions as they are distributed across a wide range of ecological, chemical, moisture and climatic gradients, are relatively easy to identify, and many respond differently to environmental changes, hence offering a measure of forest resilience and environmental stress. They are used as indicators of air quality (Barkman Reference Barkman1969; LeBlanc & Rao Reference LeBlanc and Rao1973; Geiser & Neitlich Reference Geiser and Neitlich2007), forest integrity (McMullin et al. Reference McMullin, Ure, Smith, Clapp and Wiersma2017), old-growth forest and continuity (Rose Reference Rose, Brown, Hawksworth and Bailey1976; Arsenault & Goward Reference Arsenault and Goward2016; Dymytrova et al. Reference Dymytrova, Brändli, Ginzler and Scheidegger2018), sustainable forest management and land use (Pinho et al. Reference Pinho, Bergamini, Carvalho, Branquinho, Stofer, Scheidegger and Máguas2012; Brunialti et al. Reference Brunialti, Frati, Calderisi, Giorgolo, Bagella, Bertini, Chianucci, Fratini, Gottardini and Cutini2020).
One group of tree-dwelling lichens that has received significant attention as bioindicators of environmental change are the cyanolichens. Cyanolichens have been defined as the lichen symbiosis between a lichenized mycobiont and a cyanobacteria (Büdel & Rhiel Reference Büdel and Rhiel1987; Ahmadjian Reference Ahmadjian1989; Brodo et al. Reference Brodo, Sharnoff and Sharnoff2001). For simplicity, we also include what are referred to as cephalolichens (Gauslaa & Goward Reference Gauslaa and Goward2012), which are tripartite associations involving a lichenized mycobiont, a cyanobacterium contained in specialized structures referred to as cephalodia, and a green alga. Arboreal cyanolichens are very sensitive to air pollution such as SOx compounds that inhibit photosynthesis (LeBlanc & Rao Reference LeBlanc and Rao1973; Gunawardana et al. Reference Gunawardana, Wanigatunge, Wewalwela, Vithanage and Wijeyaratne2023) and Nr compounds which interfere with nitrogen fixation. They are often associated with humid old-growth forests and old trees (Goward Reference Goward1994; Arsenault & Goward Reference Arsenault and Goward2016), and many are considered rare or endangered. In Canada, half of the lichens assessed as being at risk are arboreal cyanolichens (Government of Canada 2025). They are also most often associated with circumneutral to basic substrata (Gauslaa Reference Gauslaa1985, Reference Gauslaa1995; Goward & Arsenault Reference Goward and Arsenault2000a).
Arboreal cyanolichens of the Lobarion community in western North America are often associated with old-growth conifers in coastal and inland temperate rainforests, while in eastern North America they are mostly associated with old broadleaf trees (Goward & Arsenault Reference Goward and Arsenault2000a). Exceptions to the former are situations where the canopy of Populus trees facilitates the establishment of cyanolichens on conifers in young forests through the phenomenon known as the dripzone effect (Arsenault & Goward Reference Arsenault, Goward and Darling2000; Goward & Arsenault Reference Goward and Arsenault2000a; Gauslaa & Goward Reference Gauslaa and Goward2012). Thus far, this dripzone phenomenon has been described only from western North America. Goward & Arsenault (Reference Goward and Arsenault2000b) postulated that the association of the Lobarion with old broadleaf trees in eastern North America and in Europe could be related to high historical levels of acid rain.
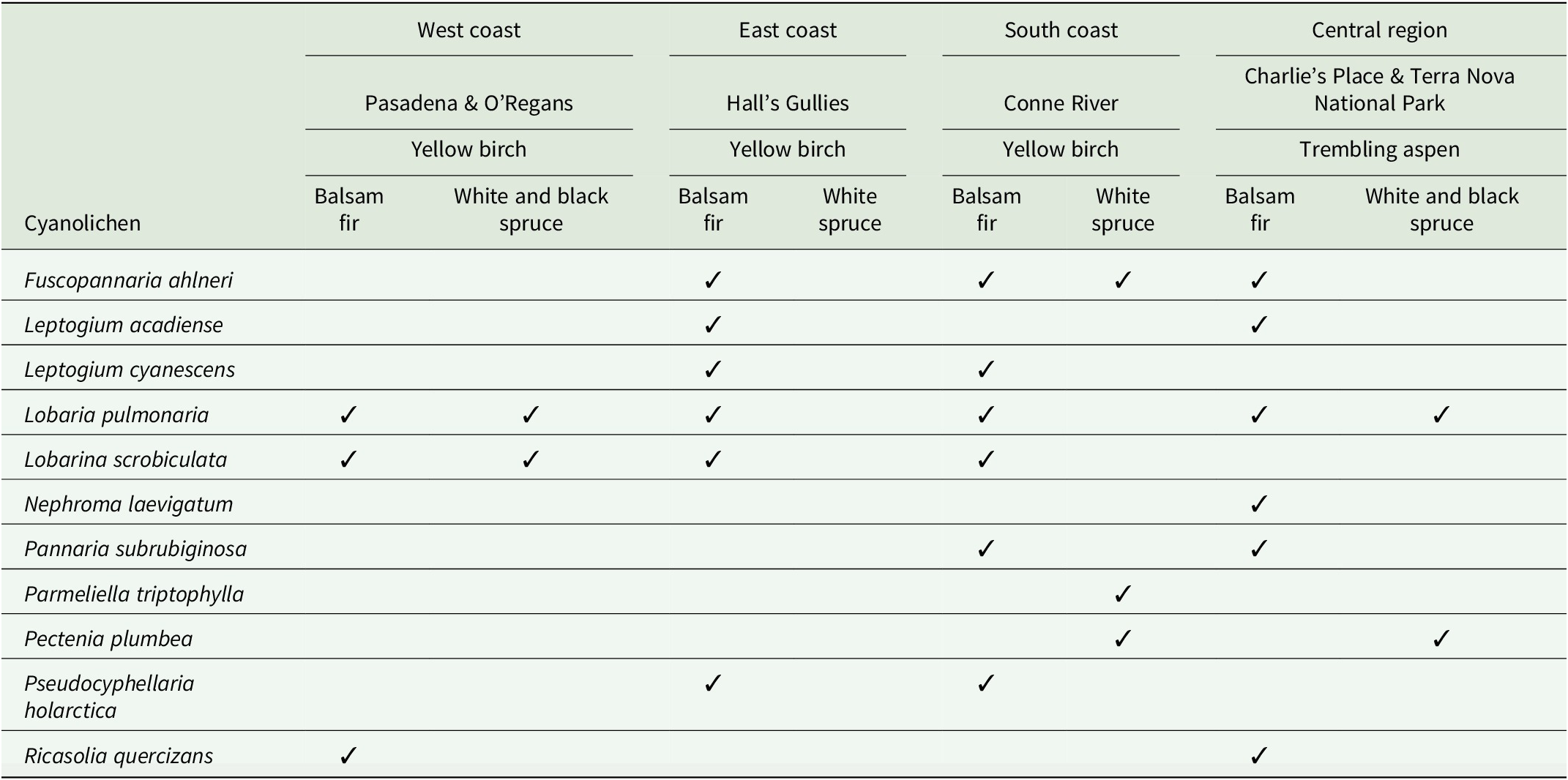
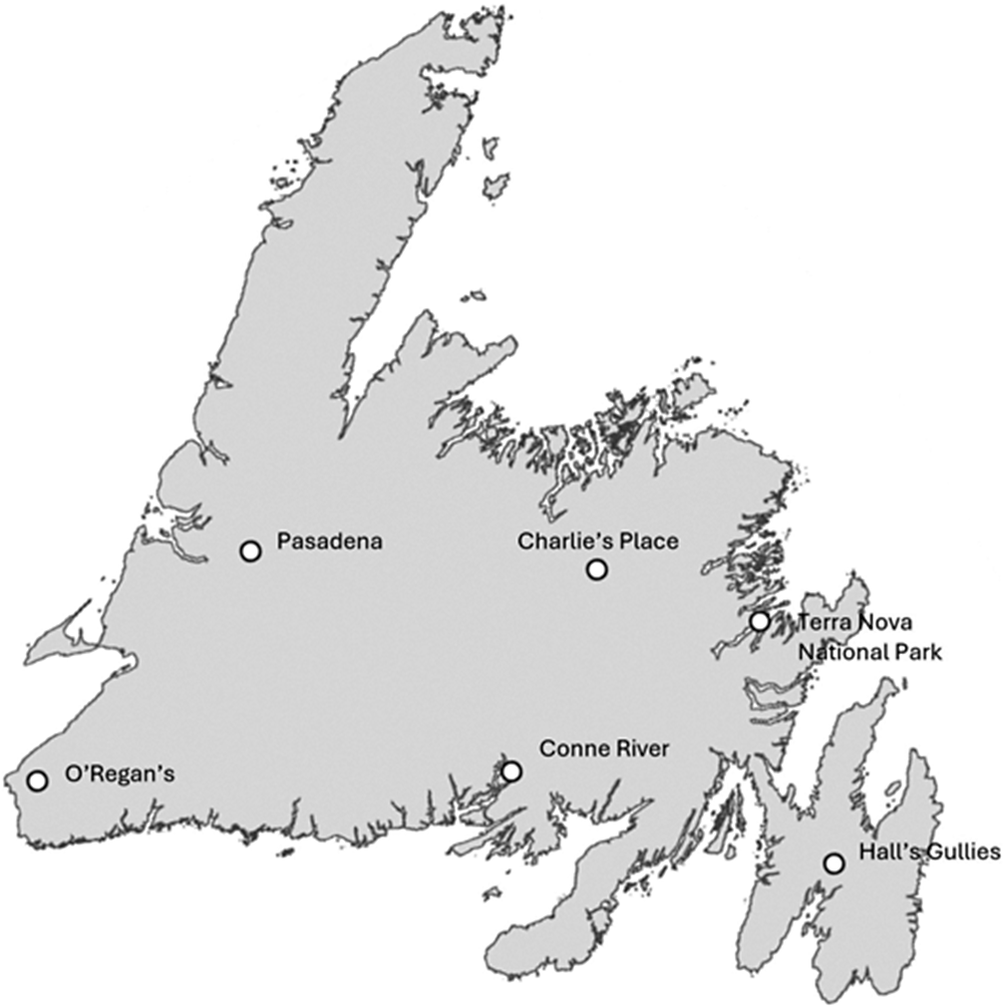
During surveys for lichen diversity with a focus on rare arboreal lichens along transects in Newfoundland forests, we observed cyanolichens growing mostly on Abies balsamea (balsam fir), and sporadically on Picea glauca (white spruce) and Picea mariana (black spruce), under the dripzone of old Betula alleghaniensis (yellow birch) and Populus tremuloides (trembling aspen), that were not present on conifers outside the dripzone. Eleven species of arboreal cyanolichens were found associated with this phenomenon in four different regions on the island portion of Newfoundland and Labrador (Table 1). These were represented by Fuscopannaria ahlneri (P. M. Jørg.) P. M. Jørg., Leptogium acadiense J. W. Hinds et al., Leptogium cyanescens (Ach.) Körb., Lobaria pulmonaria (L.) Hoffm., Lobarina scrobiculata (Scop.) Nyl., Nephroma laevigatum Ach., Pannaria subrubiginosa Brodo & McMullin, Parmeliella triptophylla (Ach.) Müll. Arg., Pectenia plumbea (Lightf.) P. M. Jørg. et al., Pseudocyphellaria holarctica McCune et al., and Ricasolia quercizans (Michx.) Stizenb. Pectenia plumbea, considered a species of conservation concern, was found on white spruce under the canopy of aspen in Central Newfoundland and under yellow birch on the south coast. It should be noted that in these areas all cyanolichens present in the dripzone were also present on either the yellow birch or trembling aspens above them. This is the first report of the dripzone phenomenon enhancing cyanolichen establishment on conifers outside western North America. Most of the observations (Table 1) are associated with yellow birch on the west coast near Pasadena and O’Regans, on the east coast in Hall’s Gullies (McMullin & Arsenault Reference McMullin and Arsenault2019), and on the south coast near Conne River (Fig. 1). In Central Newfoundland, there were two observations associated with trembling aspen (Table 1), one in Charlie’s Place (Gillingham et al. Reference Gillingham, Hodge, Skeard, Hanel, Wiersma, Arsenault, Driscoll, Paquette and McMullin2024) and one in Terra Nova National Park (Fig. 1). In addition to the dripzone effect, we have also observed, along our survey transects on the west, east and south coasts, cases where cyanolichens are present on conifers where no apparent enrichment is obvious. One example of this is from the west coast, where Lobaria pulmonaria can be abundant on white spruce trees while only very rarely seen on black spruce or balsam fir outside of yellow birch dripzones. A similar observation was made in central British Columbia by Gauslaa et al. (Reference Gauslaa, Goward and Pypker2020). Another example is from the east and south coast study areas, where there exists a cyanolichen community associated with Erioderma pedicellatum (Hue) P. M. Jørg. that can be abundant on balsam fir and regularly includes Coccocarpia palmicola (Spreng.) Arv. & D. J. Galloway, Parmeliella parvula P. M. Jørg. and Fuscopannaria alhneri. In these areas, however, the Lobarion community is predominantly found on balsam fir within yellow birch dripzones. These cases point to other factors influencing cyanolichen distribution and abundance on conifers in eastern North America, but they are beyond the scope of this paper.
Field observations of arboreal cyanolichens on conifers facilitated by the dripzone of broadleaf trees in four different regions of Newfoundland (Fig. 1). Overstorey broadleaf dripzone trees are in bold above the understorey conifers.
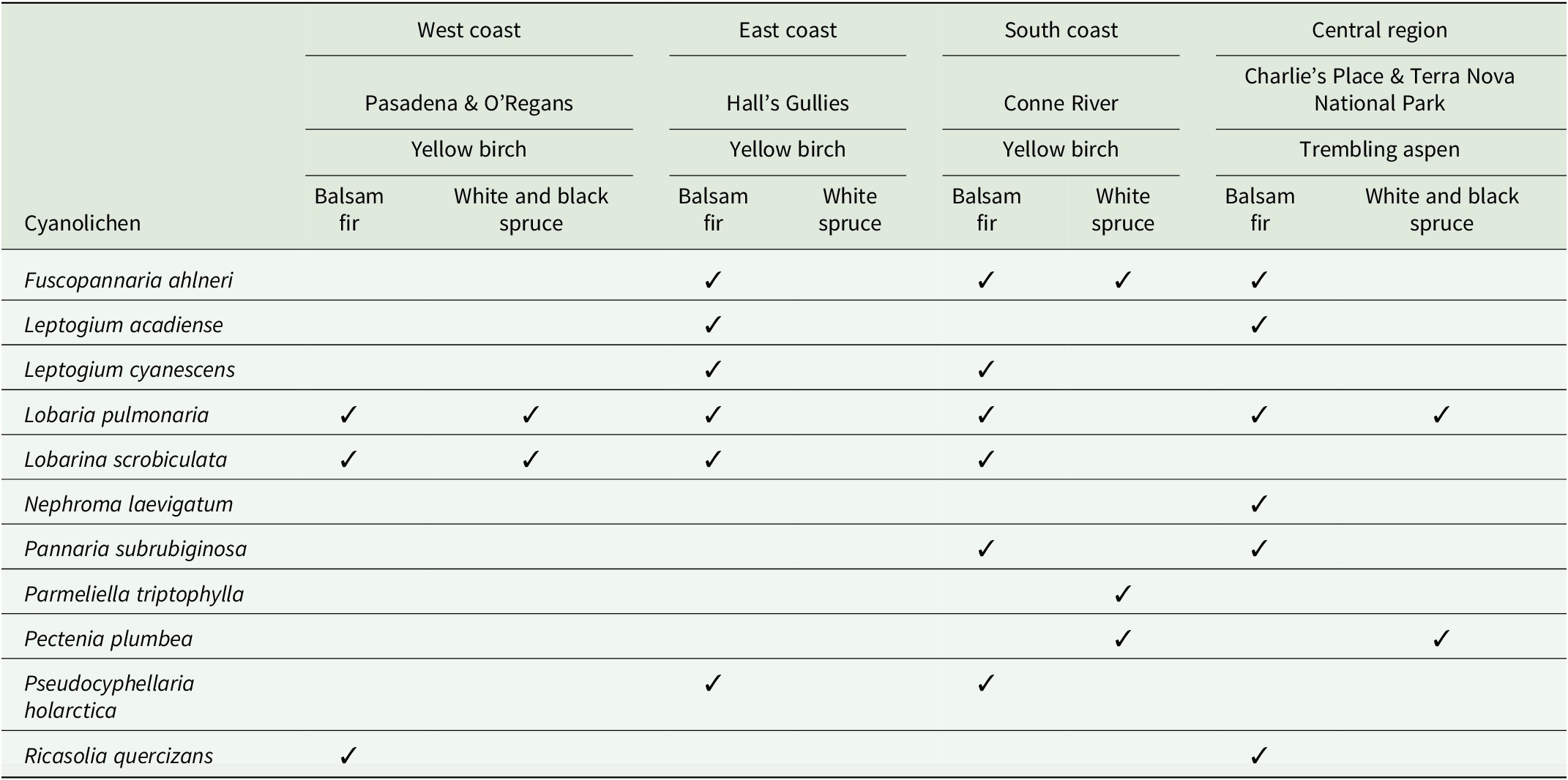
Table 1. Long description
Starting from the leftmost column, cyanolichen species are listed: Fuscopannaria ahlneri, Leptogium acadiense, Leptogium cyanescens, Lobaria pulmonaria, Lobarina scrobiculata, Nephroma laevigatum, Pannaria subrubiginosa, Parmeliella triptophylla, Pectenia plumbea, Pseudocyphellaria holarctica, Ricasolia quercizans. Moving right, the table is divided into West coast (Pasadena and O'Regans), East coast (Hall's Gullies), South coast (Conne River), and Central region (Charlie's Place and Terra Nova National Park). Each region is further split by overstorey broadleaf trees (Yellow birch or Trembling aspen, in bold) and understorey conifers (Balsam fir, White spruce, Black spruce). Presence of each cyanolichen species is indicated by a checkmark in the corresponding cell. Lobaria pulmonaria and Lobarina scrobiculata are present in multiple regions and conifer hosts. Fuscopannaria ahlneri, Leptogium acadiense, Leptogium cyanescens, Pannaria subrubiginosa, Pectenia plumbea, Pseudocyphellaria holarctica, and Ricasolia quercizans show regionally restricted distributions. Nephroma laevigatum is only found in the Central region. Parmeliella triptophylla and Pectenia plumbea are limited to the South coast and Central region. The spatial arrangement highlights the facilitation of cyanolichen presence by broadleaf dripzone trees above conifers.
Locations on the island portion of Newfoundland and Labrador where the dripzone effect has been observed. Pasadena and O’Regans are on the west coast, Hall’s Gullies is on the east coast, Conne River is on the south coast, and Charlie’s Place and Terra Nova National Park are in the Central region of Newfoundland.

Materials and Methods
In order to quantify the dripzone phenomenon observed above, we focused on balsam fir trees in and outside the understorey of yellow birch in three regions of Newfoundland. The Pasadena watershed is located on the west coast, Hall’s Gullies on the east coast, and Conne River on the south coast (Fig. 1). The forests from all three regions are considered wet boreal (Thompson et al. Reference Thompson, Larson and Montevecchi2003) but the west coast is colder and drier with an average temperature of 4.4 °C and 124 days of rain, while the south and east coasts are warmer and wetter with average annual temperatures of 4.9 °C and 5.5 °C and 153 and 178 days of rain, respectively. For this study, we selected wet forests or forests on toe positions next to wetlands. Balsam fir trees exhibiting the dripzone effect under individual mature yellow birch trees were sub-sampled along 1–2 km transects previously used for rare lichen surveys. Balsam fir trees immediately outside the downward projected canopy footprint of the same individual yellow birch trees were also selected. These were within 5–15 m from the ‘dripzone’ trees and were chosen to be comparable in size and age. Yellow birch trees ranged from c. 30–50 cm in diameter at breast height (dbh) and were 118–226 years old, while balsam fir trees varied between c. 6–30 cm in dbh and were 30–90 years old. We collected 18–19 balsam fir branches from each region at or below 2 m from the ground, both directly under and immediately outside the canopy-footprint of old yellow birch trees to minimize ecological differences between the branches collected. Branches were returned to the laboratory where all sections of each branch were measured, including twigs and branchlets (all subdivisions), to obtain a total sampled branch length. All lichen thalli were identified to species and counted for each branch, with individual thalli grouped into size categories based on thallus diameter: 1 = 0.1–0.99 cm, 2 = 1–4.99 cm, 3 = 5–9.99 cm, 4 = 10–14.99 cm and 5 = 15–30 cm. We report a standardized metric of thalli abundance by dividing the total number of thalli by the total length of all branch sections and branchlets that were examined for each individual branch. We also calculated an estimate for area of lichen thalli by assuming thalli to be circular and using the median of each size category as the diameter. We then added the area of each thallus by species on individual branches and divided by the total branch length to obtain standardized cumulative areas for each branch sampled. Branch pH was tested on a subset of 78 small, dead, lichen-free twig samples from each branch following the method described by Gauslaa & Holien (Reference Gauslaa and Holien1998). A two-sample t-test for unequal variance with one-tailed probability was used to compare bark pH with the abundance and cumulative area of chlorolichens and cyanolichens on branches inside and outside the dripzone. The units compared were individual branches for each region.
Results and Discussion
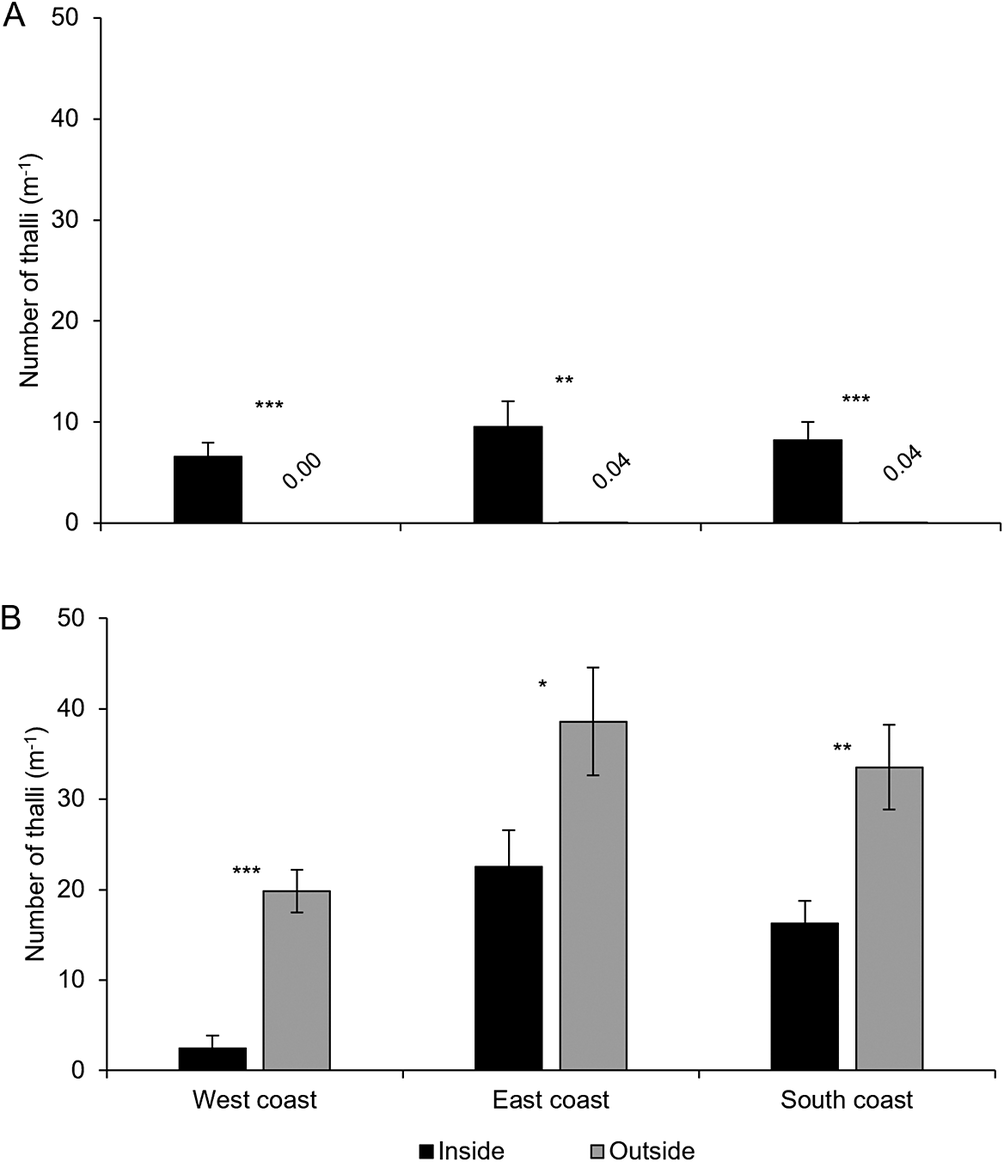
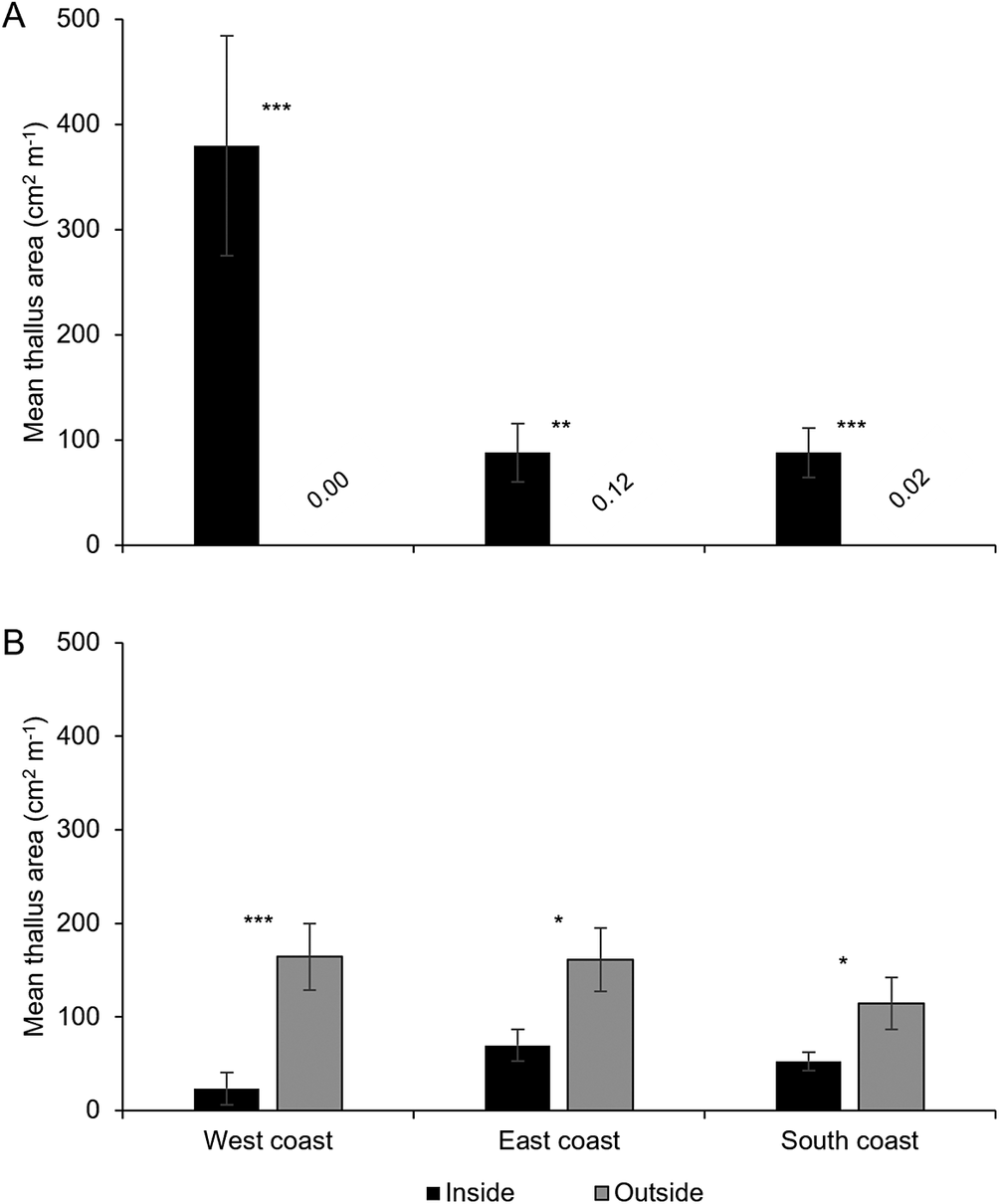
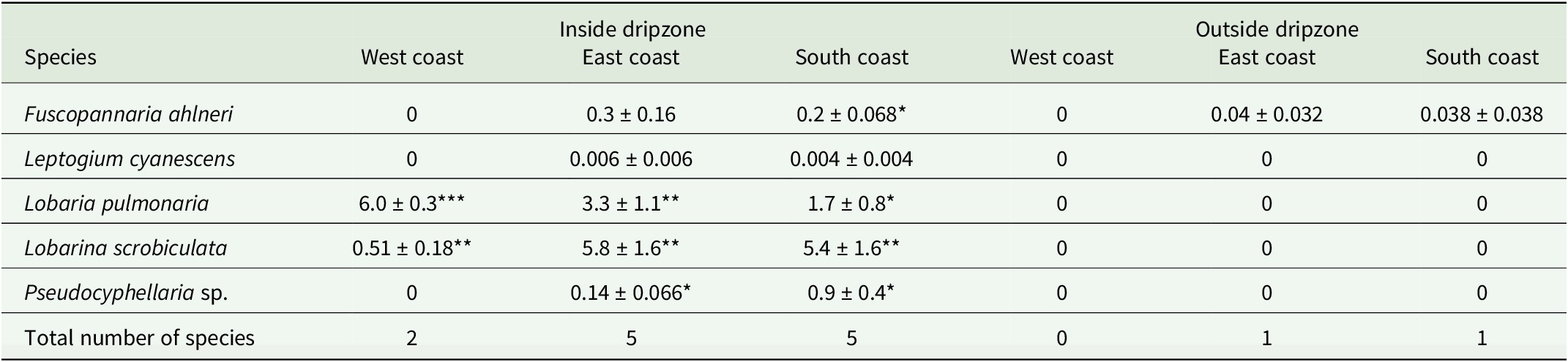
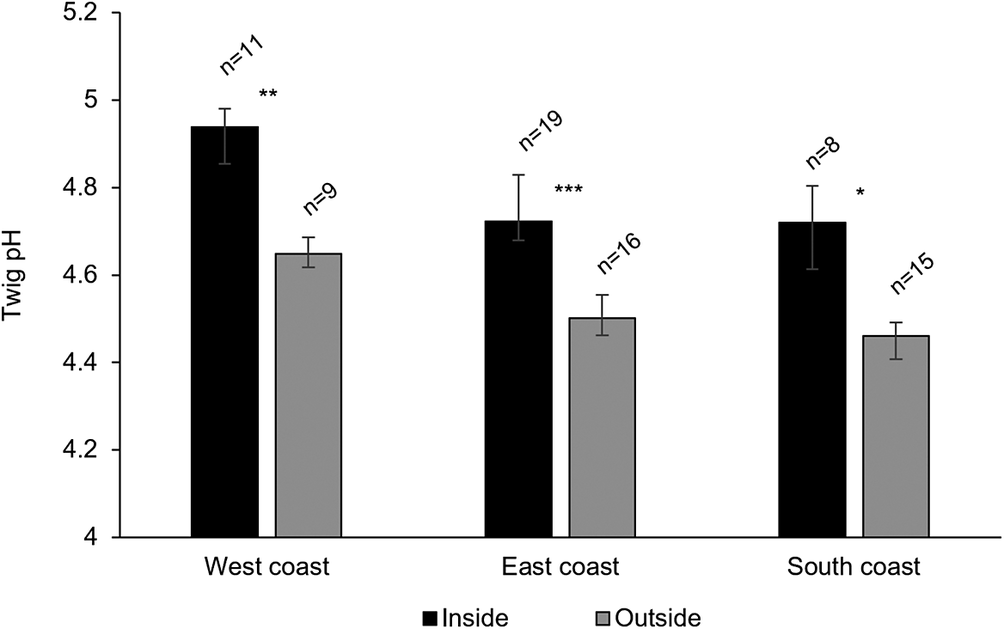
A total of 11 113 thalli were enumerated on 111 branches. Five cyanolichen species occurred inside the dripzone and only one occurred outside. The number of cyanolichen thalli was significantly higher under the dripzone of yellow birch compared to immediately outside (Fig. 2A), whereas the number of chlorolichen thalli was significantly higher outside of the dripzone (Fig. 2B). Goward & Arsenault (Reference Goward and Arsenault2003) also found that the abundance of a number of acidophilic chlorolichens was negatively affected by the Populus dripzone. The total area of cyanolichens and chlorolichens on branches followed the same trends as the number of thalli (Fig. 3A & B). Five species of cyanolichens occurred inside the dripzone of yellow birch trees while only one occurred outside of their canopy in all three regions studied (Table 2). Of these, Lobaria pulmonaria and Lobarina scrobiculata were the most abundant and showed the most significant difference within and outside of the dripzone across all three regions. The abundance of Pseudocyphellaria holarctica was slightly higher within compared to outside the dripzone, while the other two cyanolichens present on the east coast had low frequency, lacked statistical power and were not significant. On the south coast, Fuscopannaria alnheri and Pseudocyphellaria holarctica were significantly more abundant inside the dripzone but there was no significant difference for Leptogium cyanescens. Virtually no cyanolichens were found outside the yellow birch dripzone, except for Fuscopannaria alnheri on the east and south coast of Newfoundland (Table 2). The bark pH of balsam fir twigs was significantly higher under the dripzone of yellow birch trees in all three regions (Fig. 4), which is consistent with evidence that the canopy of this tree species enriches the pH of throughfall (Bouman et al. Reference Bouman, Mazzocca, Bennett and Oakes2020). A similar relatively narrow range in pH of conifer bark between Lobarion- versus Parmelion-dominated Picea branches was reported in Norway (Nilsson et al. Reference Nilsson, Solhaug and Gauslaa2022). The difference in conifer bark pH in our dripzone study is not as great as that found in the aspen dripzone studies from British Columbia, suggesting probable differences in the chemistry of canopies of different phorophyte species. Our study system involves balsam fir trees whose bark has been shown to immobilize manganese and thus limit the influence of that element on epiphytic lichen composition (Schmull & Hauck Reference Schmull and Hauck2003). Our results therefore challenge the suggestion made by Hauck & Spribille (Reference Hauck and Spribille2002) that the dripzone facilitating cyanolichen establishment on conifers is a reflection of the Mn/Ca ratio. Further investigation of bark and lichen chemistry will be required to clarify this in our study system.
Mean abundance per branch length (number of thalli m-1) ± SE of cyanolichens (A) and chlorolichens (B) within and outside the dripzone of old yellow birch trees in three regions of Newfoundland. Significance is denoted with * P < 0.05, **P < 0.01, *** P < 0.001.
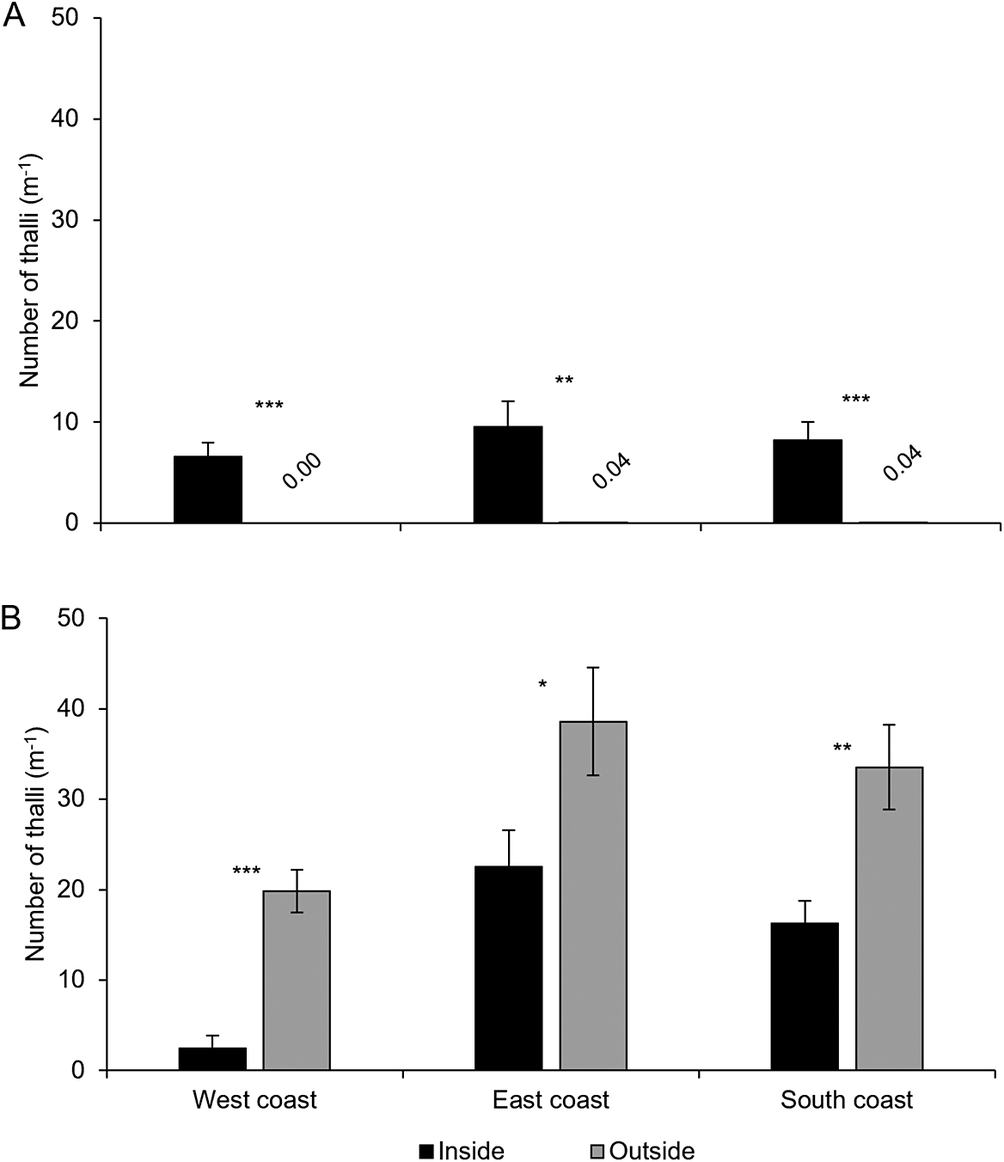
Figure 2. Long description
Panel A shows cyanolichen abundance per meter branch length on the y-axis, with regions west coast, east coast, and south coast on the x-axis. For each region, black bars (inside dripzone) are higher than white bars (outside), with outside values near zero. West coast inside is about 8, outside is 0.00, significance triple asterisk. East coast inside is about 12, outside is 0.04, double asterisk. South coast inside is about 10, outside is 0.04, triple asterisk. Panel B shows chlorolichen abundance. For each region, gray bars (outside) are higher than black bars (inside). West coast inside is about 4, outside is about 20, triple asterisk. East coast inside is about 20, outside is about 40, single asterisk. South coast inside is about 15, outside is about 30, double asterisk. Error bars indicate standard error. Significance is marked with asterisks: one for P less than 0.05, two for P less than 0.01, three for P less than 0.001.
Mean thallus area per branch length (m) ± SE of cyanolichens (A) and chlorolichens (B) within and outside the dripzone of old yellow birch trees in three regions of Newfoundland. Significance is denoted with * P < 0.05, ** P < 0.01, *** P < 0.001.
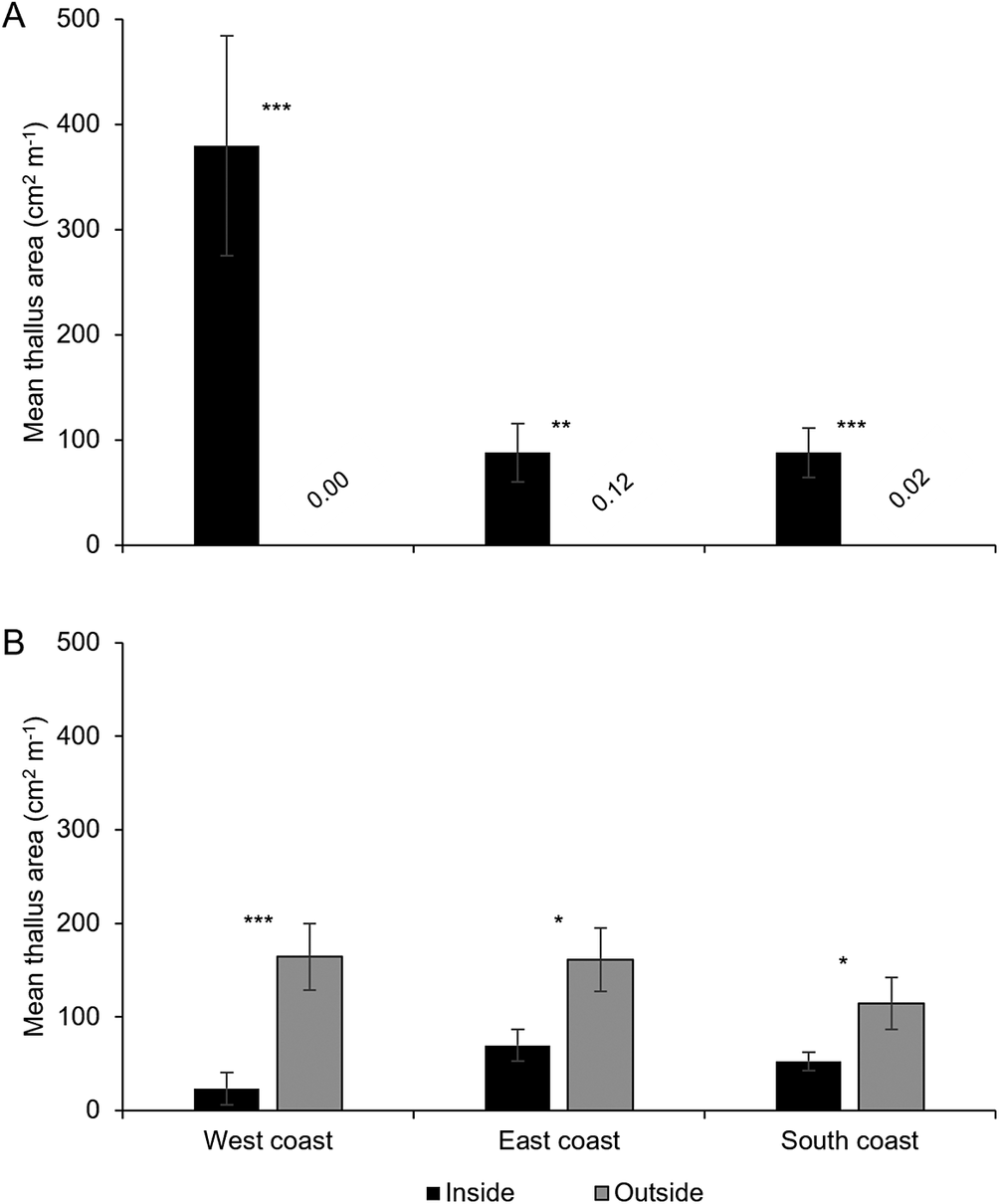
Figure 3. Long description
Panel A at the top shows mean thallus area in square centimeters per meter for cyanolichens. The y-axis ranges from 0 to 500. Three groups are shown left to right: inside the dripzone (black bar, about 400, triple asterisk), outside the dripzone (gray bar, about 100, double asterisk), and another outside group (gray bar, about 100, triple asterisk). Values below bars are 0.00, 0.12, and 0.02. Panel B below shows mean thallus area for chlorolichens with the same y-axis. Three regions are labeled: West coast, East coast, South coast. For each, black bars (inside) are lower than gray bars (outside). West coast: inside about 40, outside about 180, triple asterisk. East coast: inside about 70, outside about 140, single asterisk. South coast: inside about 50, outside about 120, single asterisk. Error bars indicate standard error. Significance is marked with asterisks above bars.
Mean abundance of cyanolichen species (number of thalli/metre) ± SE inside and outside the dripzone of yellow birch in three regions of Newfoundland. Significance is denoted with * P < 0.05, ** P < 0.01, *** P < 0.001.
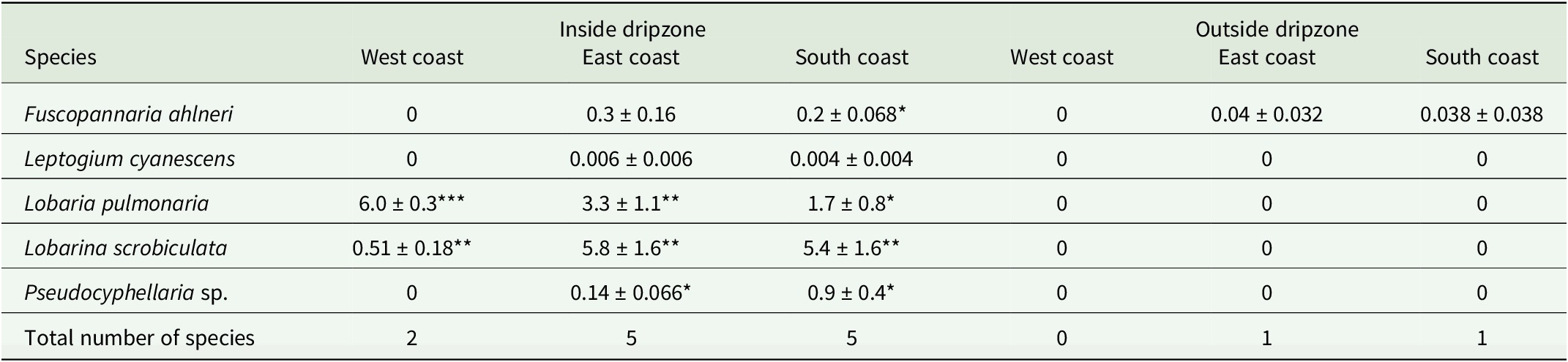
Table 2. Long description
Starting from the top row, species are listed vertically: Fuscopannaria ahlneri, Leptogium cyanescens, Lobaria pulmonaria, Lobarina scrobiculata, and Pseudocyphellaria sp. For each species, mean abundance (number of thalli per metre plus or minus standard error) is shown horizontally for West coast, East coast, and South coast, first inside the dripzone, then outside. Significant values are marked with asterisk symbols. Fuscopannaria ahlneri shows low abundance inside the dripzone (East coast 0.3 plus or minus 0.16, South coast 0.2 plus or minus 0.068, significant at P less than 0.05), and near zero outside. Leptogium cyanescens is nearly absent except for minimal presence inside the dripzone on East and South coasts. Lobaria pulmonaria is abundant inside the dripzone (West coast 6.0 plus or minus 0.3, P less than 0.001; East coast 3.3 plus or minus 1.1, P less than 0.01; South coast 1.7 plus or minus 0.8, P less than 0.05), but absent outside. Lobarina scrobiculata is present inside (West coast 0.51 plus or minus 0.18, East coast 5.8 plus or minus 1.6, South coast 5.4 plus or minus 1.6, all P less than 0.01), absent outside. Pseudocyphellaria sp. is only found inside the dripzone (East coast 0.14 plus or minus 0.066, South coast 0.9 plus or minus 0.4, both P less than 0.05). The total number of species is highest inside the dripzone (West coast 2, East coast 5, South coast 5), and lowest outside (West coast 0, East coast 1, South coast 1). Significant differences are concentrated inside the dripzone, especially for Lobaria pulmonaria and Lobarina scrobiculata.
The pH of twigs inside and outside the dripzone of old yellow birch trees in three regions of Newfoundland. Significance is denoted with * P < 0.05, ** P < 0.01, *** P < 0.001.
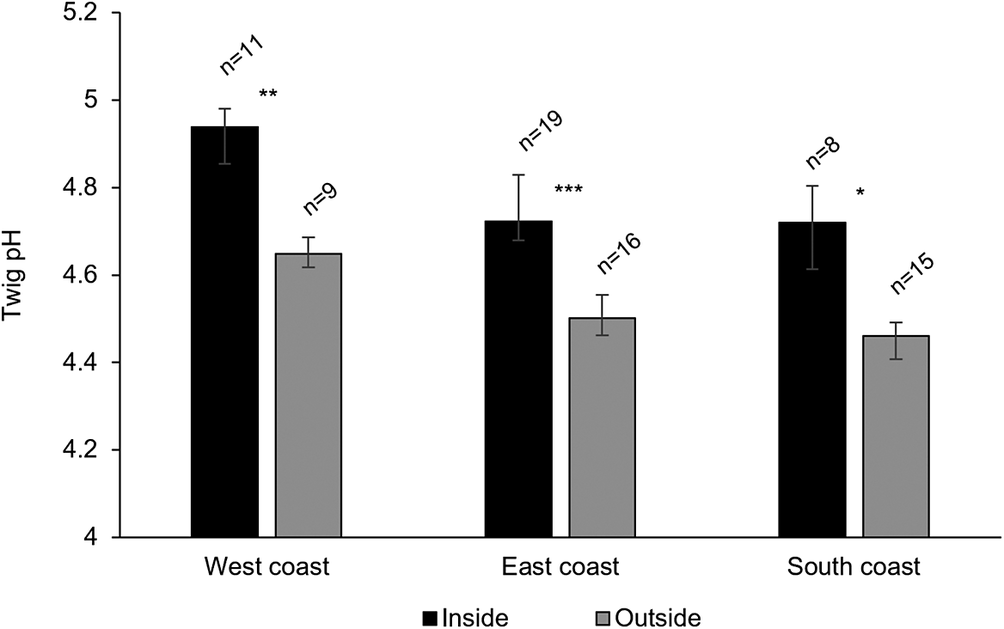
Figure 4. Long description
The x-axis lists West coast, East coast, and South coast from left to right. The y-axis is labeled Twig p H, ranging from 4 to 5.2. For each region, two bars are shown: black for Inside and gray for Outside the dripzone. On the West coast, Inside (n equals 11) has a mean p H just above 4.9, Outside (n equals 9) is about 4.65, with double asterisk indicating P less than 0.01. On the East coast, Inside (n equals 19) is about 4.8, Outside (n equals 16) is about 4.5, with triple asterisk for P less than 0.001. On the South coast, Inside (n equals 8) is about 4.75, Outside (n equals 15) is about 4.45, with single asterisk for P less than 0.05. Error bars are present for all means. The legend at the bottom identifies black as Inside and gray as Outside.
The dripzone pattern associated with yellow birch is similar to that observed by Goward & Arsenault (Reference Goward and Arsenault2000a) under the canopy of Populus trees in the Central Interior of British Columbia. However, probably due to the isolated patchiness of these old yellow birch trees in a conifer-dominated matrix, the dripzone phenomenon described here is spatially restricted to the area directly under the canopy-footprint of yellow birch, as opposed to a gradient of decreasing cyanolichens with distance from Populus trees described in Goward & Arsenault (Reference Goward and Arsenault2000a). The other key difference in our dripzone study compared to the original findings in British Columbia is that all cyanolichen species found on balsam fir under the dripzone were also found on the old overstorey yellow birch trees themselves, and in the case of Charlie’s Place and Terra Nova National Park on the overstorey mature aspen trees. This offers an expanded interpretation of how the dripzone effect works. We hypothesize that as broadleaf trees get old and harbour an abundant community of cyanolichen epiphytes that thrive on higher bark pH, they, along with their epiphytes, enhance the bark chemistry of conifers immediately under them, thereby enabling the establishment of cyanolichens. This is different from the original dripzone hypothesis in which Goward & Arsenault (Reference Goward and Arsenault2000a) did not consider cyanolichen epiphytes contributing to the primary aspen throughfall enrichment due to their scarcity or absence on Populus trees in young forests of BC’s Central Interior. However, they did suggest that once established on the understorey conifers, cyanolichens would then contribute to secondary enrichment, similar to what Gauslaa & Holien (Reference Gauslaa and Holien1998) had observed on spruce twigs in Norway. The results of the current study also suggest another difference from the original hypothesis, that the cyanolichen community on old aspens and yellow birch trees in Newfoundland contribute directly to the dispersal of propagules, namely broken cyanolichen fragments, soredia, isidia, mycobiont spores, and cyanobacteria including photobionts. This throughfall ‘cocktail’ of nutrients and cyanolichen propagules, in a more basic medium than unaltered precipitation, is a vector for the establishment of cyanolichens on understorey conifers (Fig. 5). Thus, the dripzone phenomenon is expressed along gradients of enrichment relating to factors including stand age, humidity, and available nutrients in the soil. This gradient is reflected in our own data showing higher cyanolichen diversity in the more humid forests of the east and south coasts compared to the Pasadena watershed on the west coast, and is similar to that described by Goward & Arsenault (Reference Goward, Arsenault and Darling2000c) for the intermontane region of British Columbia, where the most humid inland rainforests harbour the highest cyanolichen diversity and that of oceanic lichens (Goward Reference Goward1994).
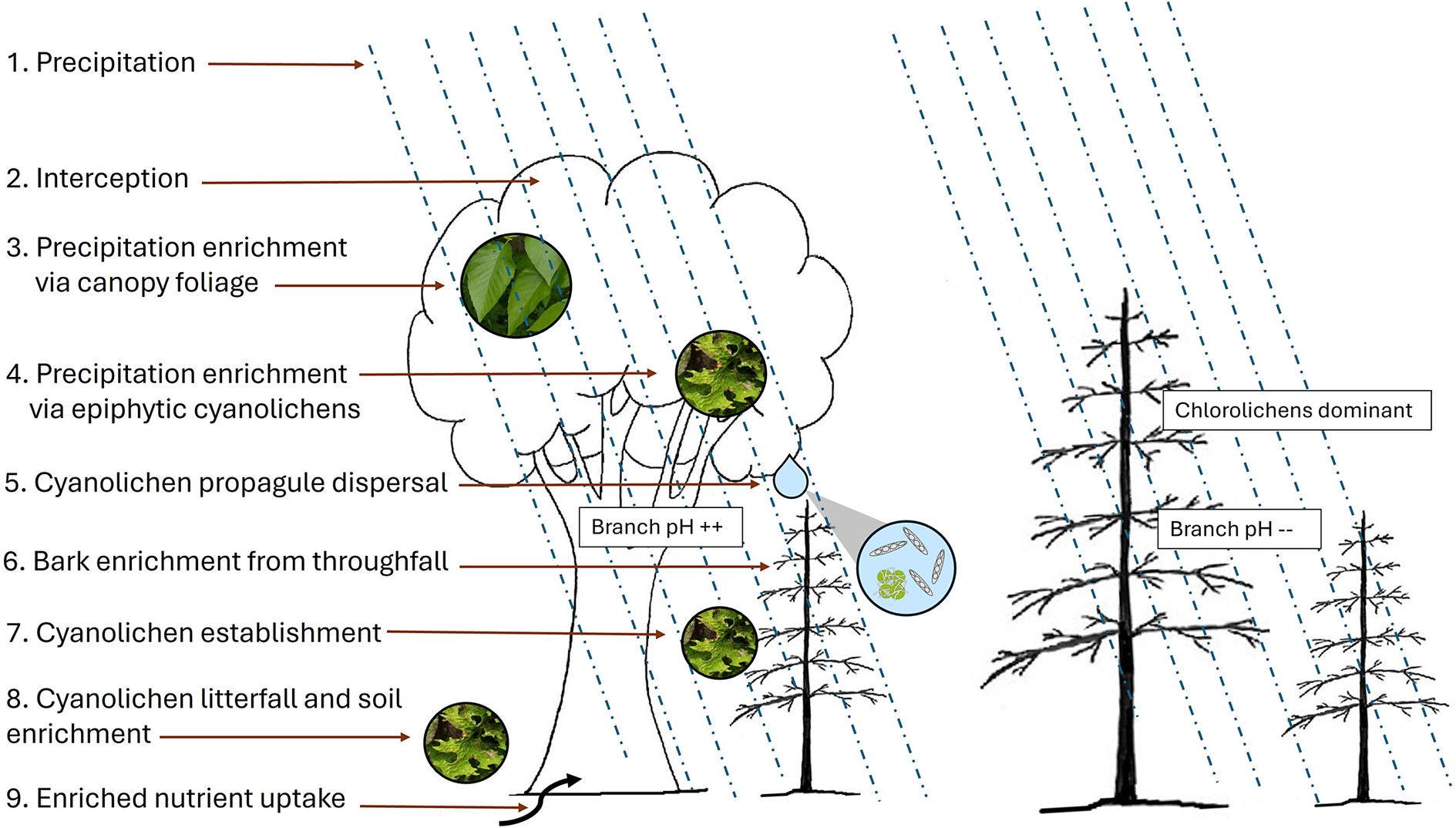
A conceptual representation of the dripzone effect of old yellow birch facilitating the establishment of cyanolichens on conifers under its canopy, modified from Arsenault & Goward (Reference Arsenault, Goward and Darling2000). In colour online.
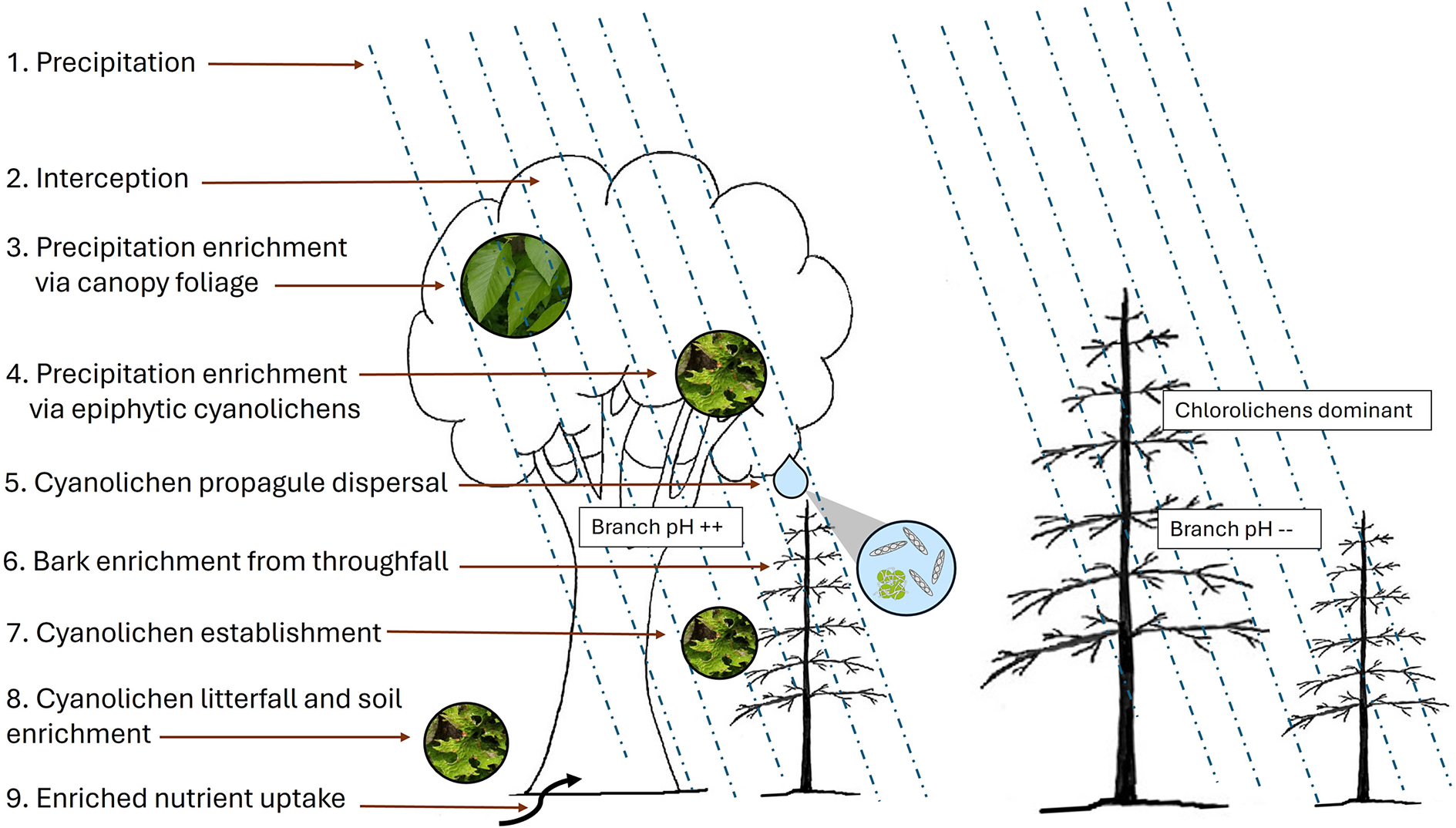
Figure 5. Long description
Starting at the top left, step 1 is precipitation, indicated by blue dashed lines falling vertically. Step 2 is interception by the yellow birch canopy. Step 3 shows precipitation enrichment via canopy foliage, with an arrow pointing to a leaf image. Step 4 details further enrichment via epiphytic cyanolichens, with an arrow to a lichen image. Step 5 is cyanolichen propagule dispersal, depicted by an arrow and a magnified inset showing spores and lichen. Step 6 is bark enrichment from throughfall, labeled ‘Branch p H plus plus.’ Step 7 is cyanolichen establishment, with an arrow to a branch. Step 8 is cyanolichen litterfall and soil enrichment, with an arrow to the ground and a lichen image. Step 9 is enriched nutrient uptake, with an arrow to the tree roots. To the right, conifers outside the birch canopy are labeled ‘Chlorolichens dominant’ and ‘Branch p H minus minus,’ indicating lower p H and absence of cyanolichen processes. The diagram visually contrasts the enriched microhabitat under yellow birch with the less enriched conifer zone.
Conclusion
It is uncertain how widespread this dripzone phenomenon is in eastern North America. We have not come across a record of the dripzone phenomenon in the Atlantic region of Canada in the scientific literature, even from papers specifically reporting on cyanolichen distribution and their habitats (i.e. Maass Reference Maass1980; Gowan & Brodo Reference Gowan and Brodo1988; Cameron & Richardson Reference Cameron and Richardson2006; Anderson Reference Anderson2007; Cameron & Neily Reference Cameron and Neily2008; McMullin et al. Reference McMullin, Duinker, Cameron, Richardson and Brodo2008; Clayden Reference Clayden, McAlpine and Smith2010). Although it is possible that the dripzone phenomenon occurs in other wet forests, especially along the Atlantic coast, it is probably rarer than in Newfoundland due to extensive logging of old broadleaf trees (Clayden Reference Clayden2014) and higher levels of sulphate deposition in previous decades (Meyer-Jacob et al. Reference Meyer-Jacob, Michelutti, Paterson, Cumming, Keller and Smol2019). Our study also points to the importance of old-growth forests and old broadleaf trees for conservation of biodiversity. We have observed on the east coast of Newfoundland that individual yellow birch trees retained in clearcut areas do not survive well. We suggest that conserving large patches of old broadleaf trees with their understorey intact may provide important biodiversity reservoirs in logged areas. In addition, given that the dripzones we observed are associated with wet forest types, it would be wise to connect these areas with riparian conservation corridors, ensuring that these are large enough not to be susceptible to secondary disturbances such as windthrow. Old broadleaf trees are becoming increasingly endangered in eastern North America and represent important keystone species for biodiversity (Clayden Reference Clayden2014) and Indigenous culture (Gagnon & Glynn Reference Gagnon and Glynn2009). Current Indigenous-led conservation initiatives (Government of Canada 2022; Gillingham et al. Reference Gillingham, Hodge, Skeard, Hanel, Wiersma, Arsenault, Driscoll, Paquette and McMullin2024), and a proposed protected area strategy (WERAC 2020), offer promise to the areas we examined during this study.
Acknowledgements
This study has evolved over the years from field observations associated with lichen ecology projects supported by the Canadian Forest Service, and with the Boreal Sentinels project supported by Natural Resources Canada, Parks Canada and Miawpukek First Nation. We are grateful to Miawpukek First Nation, including past and present Natural Resources Managers Gregory Jeddore and Angelina Francis, and the Land Guardians Andy Joe, David Jeddore, Raymond Jeddore and Craig Benoit for their insight and support in the field, particularly on the distribution of witch hazel trees (yellow birch). We also thank Frank Skeard, Dean Gillingham and Justin Hodge of Qualipu First Nation, and the Miawpukek First Nation Land Guardians, for inviting the first author, and other Foray NL faculty, to Charlies Place to survey lichens and fungi. Thanks to Tegan Padgett for sharing information on rare lichens in Terra Nova National Park and assisting with fieldwork. We are grateful to Robert LeBlanc, Randy Skinner, Candice Power and Katherine Flores for assistance in the field. Thanks to Doreen Churchill and Nick Lake for reviewing this paper.
Author ORCID
André Arsenault, 0000-0001-8165-7183.

 Open access
Open access