Obesity, a metabolic disease with a global impact, is associated with extensive accumulation of white adipocytes(Reference Lee, Wu and Fried1). Adipocytes play key roles in bioenergy release and storage. However, excessive adipocyte accumulation is associated with hypoxia, enhanced inflammation and hormone dysregulation(Reference Crewe, An and Scherer2). H2O2 accumulates in adipocytes in response to hypoxia. The pro-inflammatory response is linked to the production of oxidative stress. Inflammation in adipocytes alters cytokine profiling, leading to a pro-inflammatory state characterised by the section of pro-inflammatory cytokines, such as IL-6 and TNF-α (Reference Chandel, Maltepe and Goldwasser3). Oxidative stress leads to excessive reactive oxygen species (ROS) production, which is associated with inflammation and the development of chronic diseases, particularly metabolic disorders(Reference Lumeng, Bodzin and Saltiel4–Reference McLaughlin, Ackerman and Shen6).
Excessive ROS production and diminished antioxidant capacity are linked to DNA damage, the activation of repair mechanisms, cell-cycle arrest and associated signalling pathways(Reference Al-Aubaidy and Jelinek7). The DNA repair response system plays a crucial role in repairing ROS-induced DNA damage, ensuring genomic stability and maintaining DNA integrity(Reference Al-Aubaidy and Jelinek7). While antioxidants are thought to contribute to the repair of ROS-induced DNA damage, the specific mechanisms by which phytonutrients regulate DNA damage under oxidative stress in individuals with obesity remain unclear.
Chronic hypoxia and excessive ROS production affect adipocyte adaptation, decrease translation, alter the transcription of DNA damage and repair response genes(Reference Bar, Levis and Rechler8,Reference Lin, Lee and Yun9) . Ataxia telangiectasia mutated and ataxia telangiectasia and Rad3-related checkpoints are bypassed in adipocytes with chronic excessive ROS accumulation and DNA repair signalling, including homologous recombination (HR), mismatch repair and base excision repair, is reduced(Reference Ching, Spears and Armon10–Reference Lans, Hoeijmakers and Vermeulen12). Reduced DNA repair in adipocytes leads to adipocyte accumulation, increased genomic instability, residual DNA damage, defective DNA repair, chromosomal aberrations, fragility and aneuploidy. Ataxia telangiectasia mutated regulates checkpoint kinase 1/2 (CHK1/CHK2, an inducer of DNA repair) genes related to downstream signalling in adipose tissue and adipocyte differentiation. Moreover, insulin resistance is impaired in ataxia telangiectasia mutated knockout cells(Reference Takagi, Uno and Nishi13).
We previously reported the therapeutic role of curcumin in cancer, involving the DNA repair system(Reference Choi and Park14). We found that curcumin inhibits Rad 51 foci formation and causes the accumulation of γ phosphorylated form of the histone H2AX (γH2AX) in MDA-MB-231 breast cancer cells and suggested that co-treatment with curcumin and a poly (ADP-ribose)-polymerase inhibitor can be considered a combinational chemotherapy, as a DNA repair agent, for breast cancer. Moreover, curcumin has anti-inflammatory properties against obesity-associated inflammation and related pathologies(Reference Weisberg, Leibel and Tortoriello15–Reference Hassan, Awadalla and Abd El-Kader19). Its anti-inflammatory effects reduce NF-κB activity in tissues, which is linked to a decrease in the expression of pro-inflammatory molecules such as TNFα (Reference Weisberg, Leibel and Tortoriello15–Reference Hassan, Awadalla and Abd El-Kader19).
Although the effects of curcumin on DNA damage and repair signalling have been studied, its relationship with obesity remains unclear. Therefore, this study investigated the role of curcumin, a natural antioxidant compound, in DNA repair signalling and maintenance of adipocyte-derived mesenchymal stem cell (ADMSC) populations in a mouse model of HFD-induced obesity.
Materials and methods
Chemicals and reagents
Standard chow (SC) and HFD were provided by Purina Mills and Research Diet (D12492), respectively. Curcumin, glucose and insulin were purchased from Sigma-Aldrich. Enzymatic assay kits for cholesterol and Triglyceride (TG) and ELISA kits were obtained from Wako Chemicals and ALPCO, respectively. Total RNA was isolated using TRIzol reagent from Invitrogen. Quantitative real-time PCR reagents were obtained from Roche Diagnostics and Thermo Fisher Scientific. For the flow cytometry experiment, type I collagenase was obtained from Worthington Biochemical Co. RIPA and polyvinylidene difluoride membranes were provided by Millipore.
Animals and experimental design
Four-week-old male C57BL/6 mice (18–21 g) were obtained from ORIENT BIO, Inc. (Seongnam, Gyeonggi-do, Republic of Korea) and adapted for 1 week. Animals were provided with ad libitum access to food and tap water. They were housed in a pathogen-free animal facility at a density of five mice per cage. The animal facility was maintained at a temperature of 20 ± 1°C and a humidity of 50 ± 5 %, with a 12:12 h light-dark cycle (lights on at 08.00). At 5 weeks of age, a total of forty mice were randomly assigned to four groups as follows: standard chow (SC + CON, n 10), SC treated with curcumin (SC + CUR, n 10), high-fat diet (HFD + CON, n 10) and HFD treated with curcumin (HFD + CUR, n 10) (Figure 1). Seven mice per group provided at least 90 % statistical power to examine between-group differences (HFD + CON v. HFD + CUR). With a 20 % attrition rate owing to dietary intake, we met the sample size required for the desired level of statistical power (n 10/group). Mice were fed SC comprising 10 % of total calories from fat or HFD comprising 60 % of total calories from fat for 8 weeks. Curcumin (2·5 g/kg diet) was thoroughly mixed with powdered SC or HFD to achieve a final concentration of 0·25 % (w/w, dry weight), and the mixture was subsequently reconstituted into pellets. The concentration of 0·25 % was selected based on previous reports indicating that dietary curcumin at 0·1–0·4 % is effective in reducing HFD-induced adiposity(Reference Feng, Zou and Su16,Reference Shao, Yu and Chiang20,Reference Li, You and Wang21) . Mice were weighed weekly.
Illustration of the experimental design of this study.

At the end of the experiment, all mice performed both glucose tolerance test (GTT) and insulin tolerance test (ITT). After killing, sixteen mice (four mice per group) were used for the histology and twenty-four mice (6–7 mice per group) were used for the other experimental analyses.
All animal experiments were approved by the Institutional Animal Use and Care Committee of Hannam University (HNU 2016-6). The animal study was performed in compliance with the ARRIVE guidelines.
Glucose tolerance test and insulin tolerance test
GTT and ITT were conducted at the end of the 8-week dietary intervention. GTT was measured blood glucose after intraperitoneally (i.p.) injection of 1·5 g/kg body weight of glucose into mice after fasting for 16 h. ITT was performed 5 days after GTT. After fasting for 4 h, the mice were injected (1 U/kg body weight i.p.) with insulin and blood glucose was checked. The levels of glucose were examined using a One Touch II glucose meter (Lifescan, Johnson & Johnson). The areas under the curve (arbitrary units, a.u.) for GTT and ITT were calculated using the linear trapezoidal method.
Tissue and serum samples
At the end of the experiments, the mice were overnight fasting and killed after being anesthetised with a mixture of Zoletil and Rompun. Blood and tissue samples were collected. Epididymal fat tissue weight was measured. All serum and tissue samples were immediately stored at –70°C until analysis.
Histology
The liver and epididymal adipose tissues were fixed in 10 % formalin for 24 h, embedded in paraffin, sectioned (5 μm) at midline using a microtome (CM3050S, Leica Microsystems) and stained with haematoxylin and eosin (H&E). Images were observed using a Leica DMLS light microscope (Leica Microsystems). To determine adipocyte diameter, three visual fields were randomly selected from each section using a Leica DMLS light microscope and counted using ImageJ software (National Institutes of Health). NAFLD activity score (NAS) was evaluated according to the criteria of the NASH Clinical Research Network Pathology Committee, including steatosis, lobular inflammation and hepatocellular ballooning(Reference Kleiner, Brunt and Natta22). The experiment was conducted by experts in a blind manner.
Blood analyses
Mouse blood was separated by centrifugation (1500 × g, 10 min) and stored at −80°C. Serum insulin and leptin (LEP) levels were quantified using commercially available ELISA kits (ALPCO) according to the manufacturer’s instructions. Serum total cholesterol levels were determined by using an enzymatic assay kit.
Liver lipid isolation and quantification
Total lipids in the liver were extracted using a slightly modified version of the Folch method(Reference Folch, Lees and Sloane Stanley23). Liver TG levels were measured using an enzymatic kit.
Flow cytometry
Mouse ADMSC were isolated as previously described(Reference Cho and Park24). Mouse epididymal fat tissues were dissected, finely minced in 5 ml of PBS and dissolved in 0·2 % type I collagenase for 45 min in a warm water bath at 37°C. Digestion was inhibited with Hanks’ balanced salt solution supplemented with 20 % fetal bovine serum, and the tissue was centrifuged at 450 × g for 5 min and passed through a 70-μm cell strainer. To block non-specific Fc receptor-mediated binding, digested adipocytes were incubated with anti-mouse CD16/CD32 antibodies for 10 min on ice. The single adipocyte suspension was incubated with a fluorescent-conjugated antibody cocktail (600 ng/106 cells with anti-Sca-1-PE antibody and 300 ng/106 cells with anti-CD45-APC antibody) diluted with PBS containing 2 % fetal bovine serum on ice for 1 h. Digested adipocytes were washed twice with PBS containing 2 % fetal bovine serum. Fluorescence-activated cell sorting was performed using fluorescence-activated cell sorting analysis software on a Beckman system. The information regarding the antibodies used in this study is provided in online Supplementary Table S1.
Primary human adipocyte culture
Human adipose stromal cells were provided from the American Type Culture Collection and cultured in α-MEM supplemented with 10 % fetal bovine serum, 100 units/ml penicillin and 100 µg/ml streptomycin. The stromal cells treated with curcumin were cultured overnight and harvested for RNA extraction. In the adipocyte differentiation experiment, 2-day post-confluence cells (day 0) were differentiated in an adipogenic differentiation induction cocktail (DMEM/F12, 15 mM HEPES, 25 mM NaHCO3, 33 μM d-biotin, 17 μM pantothenate, 100 nM dexamethasone, 100 nM insulin, 1 μM rosiglitazone, 0·5 mM 3-isobutyl-1-methylxanthine, 10 μg/ml transferrin and 2 nM T3). After 7 d of differentiation, cells were placed in a maintenance medium (DMEM/F12, 10 nM insulin and 10 nM dexamethasone). All incubations were performed in a 5 % CO2 atmosphere at 37°C.
Western blotting
Human adipose stromal cells were lysed using RIPA buffer containing a protease/phosphatase inhibitor cocktail (Cell Signaling Technology). Protein samples were resolved on SDS-PAGE and transferred onto polyvinylidene difluoride membranes. The membranes were blocked with 5 % skim milk in PBS (0·1 % Tween-20) for 1 h at room temperature and then incubated with the first antibodies at 4°C overnight. After incubation of the second antibody suitable for each species’ origin, development was performed using the HRP-ECL luminescence method. Band intensities were quantified using Image J software (National Institutes of Health). The information regarding the primary antibodies used in western blotting is provided in Supplementary Table S1.
Quantitative real-time PCR
Total RNA was isolated from adipocytes using TRIzol reagent and reversely transcribed using a Transcriptor First Strand cDNA Synthesis Kit. Quantitative real-time PCR was performed on a Light Cycler 480 (Roche Diagnostics) using Taqman probes as shown in online Supplementary Table S2. Target gene expression was normalised by peptidyl-prolyl cis/trans isomerase a (PPIA) expression. The mRNA expression levels were calculated using the 2−ΔΔCt method. All experiments were performed in triplicates.
Antioxidant assay by 2,2-diphenyl-1-picrylhydrazyl (DPPH)
The assay mixture contains 1·5 ml of 0·1mM DPPH methanolic solution, dimethyl sulfoxide (DMSO) solutions of various concentrations of the material and methanol in a total volume of 3 ml. Blanks (1·5 ml of methanol and 1·5 ml of DPPH solution) and controls (0·5 ml of curcumin solution in DMSO and 2·5 ml of methanol) were also taken(Reference Acharya and Ghaskadbi25). The mixture was incubated at 37°C for 30 min. The reduction of absorbance was measured spectrophotometrically at 516 nm, and the result is summarised.
The free radical scavenging activity is expressed as SC50 values, the concentration of the sample required for 50 % of the free radical to be scavenged. It is calculated according to the following equation (Ac = absorbance of control, At = absorbance of test solution.)
% Scavenging = Ac – At × 100 Ac
Statistical analysis
All data were analysed using SPSS Statistics (Version 25.0, IBM Corp.) and are presented as mean (standard deviation (sd)). In animal studies, ANOVA with a Tukey’s post hoc test was conducted to assess the significance of intergroup differences. For in vitro studies, statistical differences between groups were determined using Student’s t test. Statistical significance was defined as P < 0·05.
Results
Effects of curcumin on body weight and adipocyte diameter in mice with obesity induced by high-fat diet
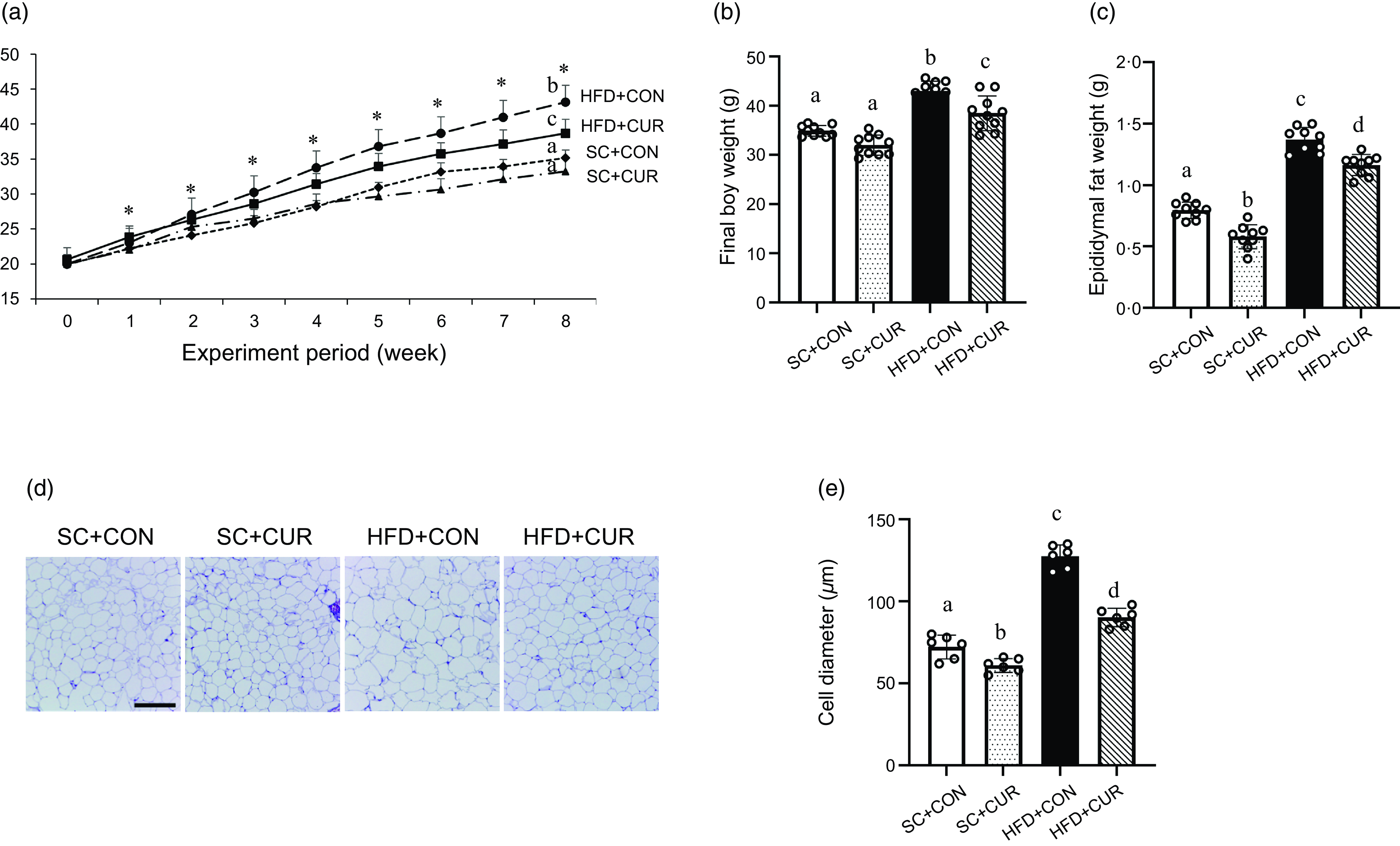
The HFD groups showed a rapid increase in body weight over the 8 weeks compared with the SC groups (Figure 2(a) and online Supplementary Table S3). At the end of the dietary period, the HFD + CON group exhibited significantly increased final body weight (mean 43·1 g v. 35·2 g of HFD + CON v. SC + CON group, respectively, P < 0·001) and epididymal fat mass (mean 1·38 g v. 0·7 g of HFD + CON v. SC + CON, respectively, P < 0·001) compared with the SC + CON group (Figure 2(a)–(c)). In addition, the HFD + CON group exhibited a significantly increased adipocyte diameter compared with the SC + CON group (mean 130·8 µm v. 75·0 µm of HFD + CON v. SC + CON, respectively, P < 0·001) (Figure 2(d)–(e)). Although food intake was similar, the HFD + CUR group showed significantly reduced HFD-induced body weight (mean 38·7 g v. 43·1 g of HFD + CUR v. HFD + CON group, respectively, P = 0·032, online Supplementary Table S3), epididymal fat mass (mean 1·16 g v. 1·38 g of HFD + CUR v. HFD + CON group, respectively, P = 0·007) and adipocyte diameter (mean 90·8 µm v. 130·8 µm of HFD + CUR v. HFD + CON group, respectively, P = 0·007) (Figure 2).
Curcumin reduces the body weight, epididymal fat weight and adipocyte size in mice with HFD-induced obesity. (a) Changes in body weight for 8 weeks; (b) final body weight; (c) epididymal fat mass; (d) representative H&E staining of the epididymal fat tissue sections; (e) adipocyte diameter. Data are mean (sd) (n 10 in all groups). *P < 0·05 among the four groups of body weight. Mean values with different letters represent significantly different. SC + CON, standard chow; SC + CUR, standard chow with curcumin; HFD + CON, high-fat diet; HFD + CUR, high-fat diet with curcumin. Scale bars are 200 μm.

Effects of curcumin on insulin resistance in mice with obesity induced by high-fat diet
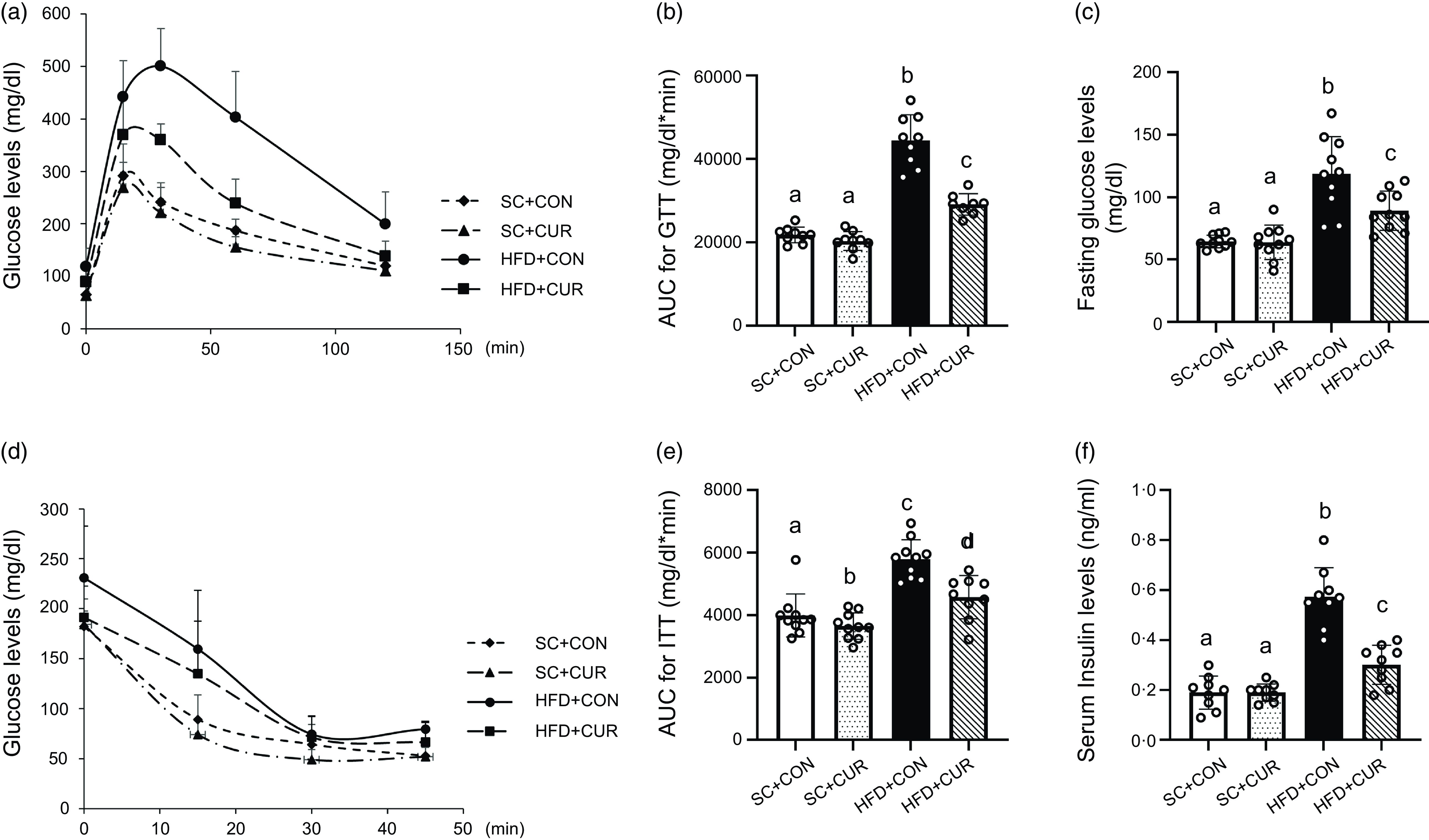
After 8-week of dietary treatment, the HFD + CON group exhibited impaired glucose tolerance compared with the SC + CON group (mean 41 203·5 mg/dl*min v. 25 580·0 mg/dl*min of HFD + CON v. SC + CON group, respectively, P < 0·001) (Figure 3(a)–(b)). Fasting serum glucose levels were higher in the HFD + CON group than in the SC + CON group (mean 118·5 mg/dl v. 63·8 mg/dl of HFD + CON v. SC + CON group, respectively, P < 0·001) (Figure 3(c)), and insulin sensitivity deteriorated in the HFD group (mean 6219·6 mg/dl*min v. 4065·0 mg/dl*min of HFD + CON v. SC + CON group, respectively, P < 0·001) (Figure 3(d)–(e)). Fasting insulin levels were significantly higher in the HFD + CON group than in the SC + CON group (mean 0·57 ng/ml v. 0·21 ng/ml of HFD + CON v. SC + CON group, respectively, P < 0·001) (Figure 3(f )). However, the HFD + CUR group showed significantly suppressed HFD-induced impairment of glucose tolerance (mean 29 142·7 mg/dl*min v. 41 203·5 mg/dl*min of HFD + CUR v. HFD + CON group, respectively, P < 0·001) and insulin sensitivity (mean 4995·0 mg/dl*min v. 6219·6 mg/dl*min of HFD + CUR v. HFD + CON group, respectively, P = 0·041), along with reduction of circulating fasting blood glucose (mean 89·3 mg/dl v. 118·5 mg/dl of HFD + CUR v. HFD + CON group, respectively, P < 0·001) and insulin levels (mean 0·35 ng/ml v. 0·57 ng/ml of HFD + CUR v. HFD + CON group, respectively, P = 0·002) (Figure 3(a)–(f )).
Curcumin improves insulin resistance in mice with HFD-induced obesity. (a) GTT; (b) AUC for GTT; (c) serum fasting glucose concentration. (d) ITT; (e) AUC for ITT; (f) serum fasting insulin concentration. Data are mean (sd) (n 10 in all groups). Mean values with different letters represent significantly different. SC + CON, standard chow; SC + CUR, standard chow with curcumin; HFD + CON, high-fat diet; HFD + CUR, high-fat diet with curcumin; GTT, glucose tolerance test; ITT, insulin tolerance test.

Effects of curcumin on hepatic steatosis and lipid profile in mice with obesity induced by high-fat diet
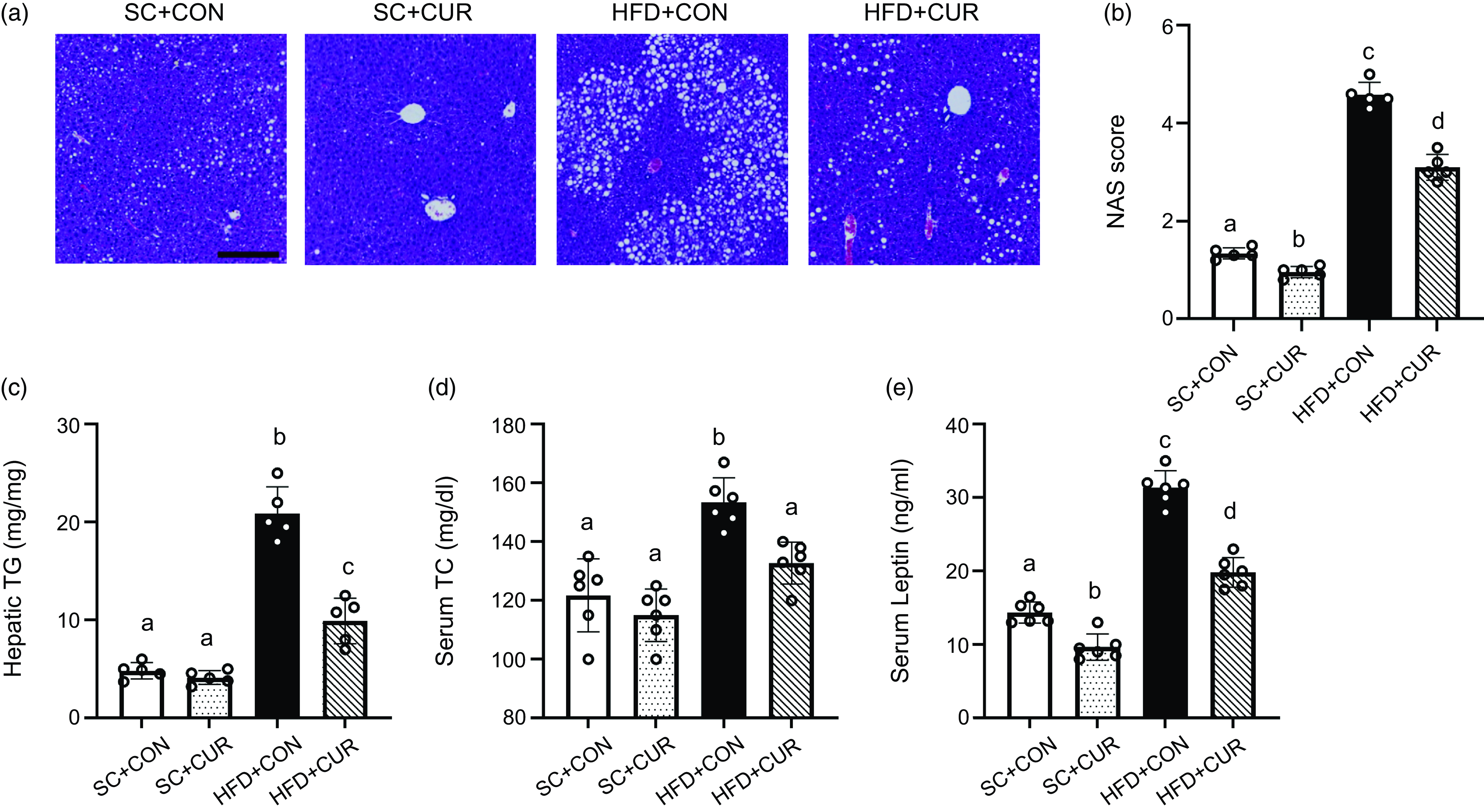
Liver H&E staining revealed significant steatosis in the HFD + CON group, including the formation of microvascular and macrovascular areas. The HFD + CON group showed significantly higher NAS score (mean 4·58 v. 1·34 of HFD + CON v. SC + CON group, respectively, P < 0·001), hepatic TG (mean 20·90 mg/mg v. 4·82 mg/mg of HFD + CON v. SC + CON group, respectively, P < 0·001) and serum total cholesterol levels (mean 159·2 mg/dl v. 129·4 mg/dl of HFD + CON v. SC + CON group, respectively, P = 0·019) than the SC + CON group (Figure 4(c), (d)). However, the HFD + CUR group showed significantly improved hepatic macrovascular and microvascular areas, along with reduced NAS score (mean 3·10 v. 4·58 of HFD + CUR v. HFD + CON group, respectively, P = 0·021), hepatic TG (mean 9·92 mg/mg v. 20·90 mg/mg of HFD + CUR v. HFD + CON group, respectively, P < 0·001) and serum total cholesterol levels (mean 130·8 mg/dl v. 159·2 mg/dl of HFD + CUR v. HFD + CON group, respectively, P = 0·024) (Figure 4(a)–(d)). Compared with the SC + CON group, the HFD + CON group showed significantly increased serum LEP levels (mean 14·9 ng/ml v. 30·9 ng/ml of SC + CON v. HFD + CON group, respectively, P < 0·001), which were mainly produced by white adipose tissue; however, the HFD + CUR group suppressed the increase in serum LEP levels (mean 19·5 ng/ml v. 30·9 ng/ml of HFD + CUR v. HFD + CON group, respectively, P < 0·001) (Figure 4(e)).
Effects of curcumin on HFD-related hepatic phenotype and lipid profiles. (a) Representative H&E staining images of the mouse liver tissues, (b) NAS score, (c) hepatic TG contents, (d) serum total cholesterol level and (e) serum LEP level. Data are mean (sd) (n 10 in all groups). Mean values with different letters represent significantly different. SC + CON, standard chow; SC + CUR, standard chow with curcumin; HFD + CON, high-fat diet; HFD + CUR, high-fat diet with curcumin. Scale bars are 200 μm.

Effects of curcumin on mouse adipocyte-derived mesenchymal stem cell population and DNA repair regulation
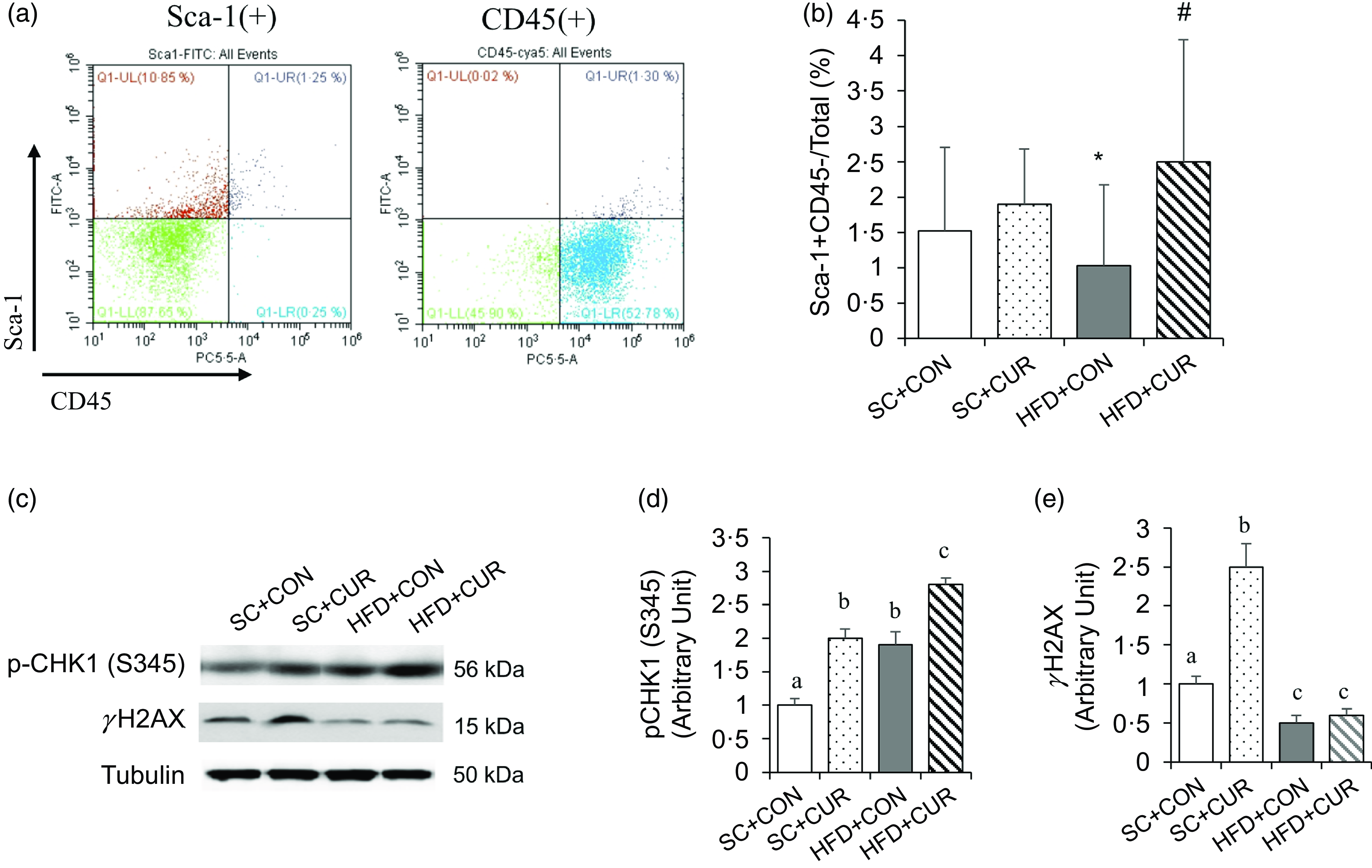
Fluorescence-activated cell sorting analysis was conducted to determine the mouse ADMSC populations stained with antibodies against Sca-1 and CD34. The results showed that the proportion of ADMSC (Sca1 + CD34-) in the HFD + CON group was lower than that in the SC + CON group (mean 0·9 % v. 1·5 % of the HFD + CON v. SC + CON group, respectively, P < 0·05), which was significantly reversed in the HFD + CUR group (mean 2·6 % v. 0·9 % of the HFD + CUR v. HFD + CON group, respectively, P < 0·05) (Figure 5(a), (b)). Western blot was used to examine the expression levels of pCHK (S345), as an inducer of DNA repair, and γH2AX, as a hallmark of DNA damage product, in the HFD-induced adipose tissue instead of ADMSCs due to the small amount to analyse protein expression. The pCHK1(S345) expression was significantly increased, while γH2AX expression was reduced in the HFD + CON group compared with the SC + CON group (Figure 5(c)–(e)). However, although the HFD + CUR group showed a significantly further increase in pCHK1(S345) expression compared with the HFD + CON group, there was no significant difference in γH2AX expression between the HFD + CUR and HFD + CON groups (Figure 5(d), (e)).
Curcumin increases the adipocyte-derived mesenchymal stem cell (ADMSC) population in mice with HFD-induced obesity. (a) The representative FACS data show the gating strategy of the mouse fat cells using antibodies against Sca-1 and CD45. (b) The HFD + CUR group shows a higher ADMSC population than the HFD + CON group. Mouse ADMSC are calculated from the cell population stained Sca-1 + CD45- antibodies. In this experiment, each group had 10 C57BL/6 male mice. Statistical significance was considered at P < 0·05. (c–e) Visceral fat tissues were obtained from HFD-induced mice treated with curcumin at the end of the experiment, and tissue lysates were analysed by immunoblotting with the indicated antibodies. (c) Representative western blotting to analyse pCHK1, total CHK1, γH2AX and tubulin levels in HFD-induced fat tissues. (d, e) Quantitative analysis of pCHK1, total CHK1, γH2AX and tubulin levels. Data are mean (sd) (n 6 in all groups). Mean values with different letters represent significantly different. SC + CON, standard chow; SC + CUR, standard chow with curcumin; HFD + CON, high-fat diet; HFD + CUR, high-fat diet with curcumin.

Effects of curcumin on DNA damage induced by H2O2 in human preadipocytes
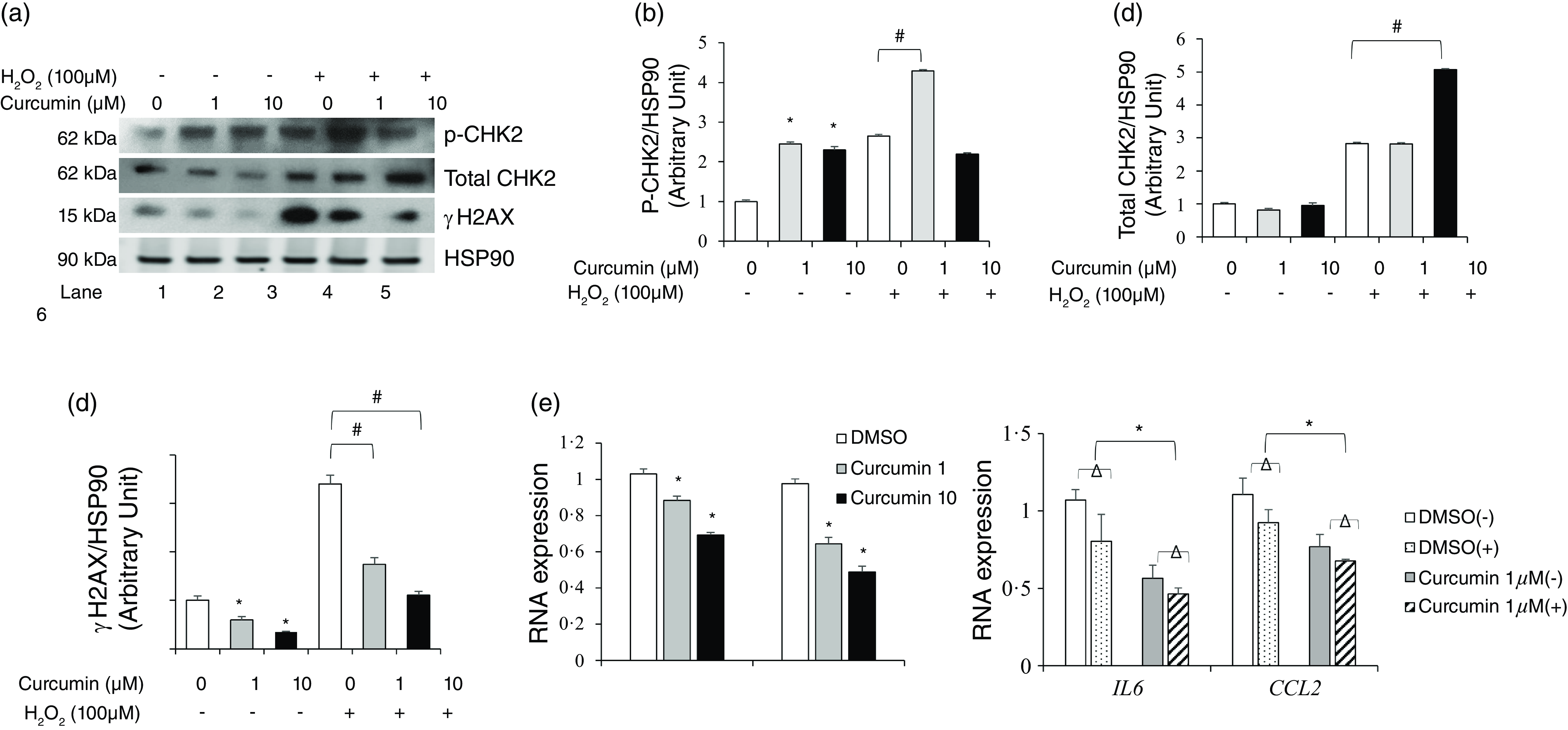
Although curcumin is a well-known activator of DNA repair signaling, its possible function in adipocytes remains unknown. Therefore, we assessed curcumin function as an activator of DNA repair signalling in H2O2-induced DNA damage in preadipocytes in vitro. We examined DNA damage/repair-associated protein expression in human preadipocytes treated with 1 or 10 µM curcumin for 12 h, followed by treatment with 100 µM H2O2, a DNA damaging agent. The DNA damage induced by 100 μM H2O2 increased the pCHK2 and total CHK2 protein levels. Curcumin pretreatment effectively suppressed this increase in pCHK2 and total CHK2 protein levels (Figure 6(a)–(d)). We also examined the protein expression levels of γH2AX in preadipocytes treated with 100 μM H2O2 following curcumin pretreatment. While γH2AX expression was drastically increased by H2O2 treatment, curcumin significantly decreased γH2AX protein expression in preadipocytes (Figure 6(a), (d)). Curcumin pretreatment suppressed H2O2-induced DNA damage by activating DNA repair signalling in preadipocytes.
Curcumin reduced DNA damage and inflammatory response in primary human preadipocytes. Human preadipocytes were treated with or without curcumin (1 and 10 µM) for 12 h, and then DNA damage was induced using 100 µM H2O2 for 3 h. Lysates were harvested after H2O2 treatment and analysed by immunoblotting and real-time PCR. (a) Western blot analysis of pCHK2, total CHK2, γH2AX and HSP90 protein expression. (b, c) Quantitative analysis of the pCHK2, total CHK2, γH2AX and HSP90 expression levels. Data are shown as mean (sd) of three independent experiments. (d) IL-6 and CCL2 mRNA levels were measured in preadipocytes using real-time PCR and found to be reduced in response to curcumin treatment. Data are mean (sd) (n 6 in all groups). *P < 0·05, control v. treatment group; #P < 0·05 as indicated.

To elucidate the anti-inflammatory function and antioxidant activity of curcumin, the expression of IL-6 and CCL2 mRNA and DPPH assay were examined. The treatment of primary human preadipocytes with 1 or 10 μM curcumin alone reduced IL-6 and CCL2 mRNA expression levels dose dependently compared with those in the DMSO-treated control (P < 0·05, Figure 6(e)). In addition, curcumin’s scavenging capacity is dose-dependent, meaning the activity increases as the concentration of curcumin increases (P < 0·001, online Supplementary Figure S1).
Effects of curcumin on adipogenic gene expression during human adipogenesis
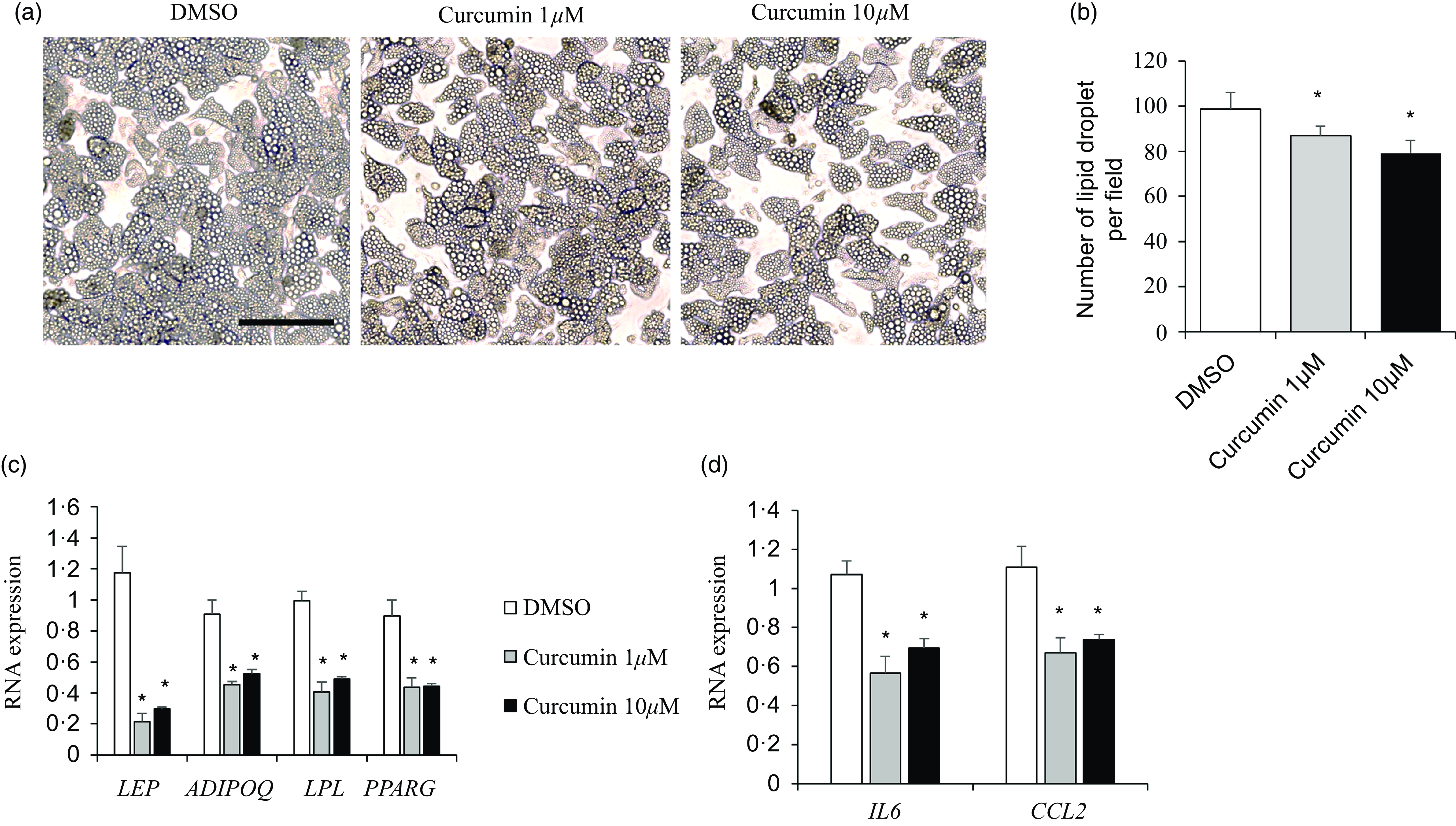
To examine the antiadipogenic effects of curcumin on preadipocyte-to-adipocyte differentiation, human primary adipocytes were incubated with 1 or 10 μM of curcumin for 48 h and then harvested to determine the effects on genes related to adipogenesis in differentiated adipocytes (day 12). Cells treated with 1 or 10 μM curcumin displayed inhibited lipid droplet accumulation (Figure 7(a)). Quantitative real-time PCR demonstrated a significant reduction in the mRNA levels of late adipogenesis markers, including LEP, ADIPOQ, LPL and peroxisome proliferator-activated receptor gamma (PPARG), in curcumin-treated adipocytes (P < 0·05; Figure 7(b)). Additionally, we investigated the anti-inflammatory effects of curcumin on differentiated adipocytes. Irrespective of concentration, curcumin suppressed IL-6 and CCL2 mRNA levels (P < 0·05; Figure 7(c)).
Curcumin inhibited human adipocyte differentiation. (a–c) Differentiated adipocytes were treated with or without curcumin (1 and 10 µM) for 24 h. (a) Representative image taken on day 14 after differentiation. Data representative of the three independent experiments is shown. (b) The mRNA expression of adipogenic markers (LEP, ADIPOQ, LPL and PPARG) was quantified. (c) The mRNA expression of inflammatory markers (IL-6 and CCL2) was quantified. Data are presented as mean (sd), n 3. *P < 0·05, control v. treatment group. Scale bars are 100 μm.

Discussion
Previously, we found that ferulic acid, as a natural bioactive compound, reduces the size of adipocytes and enhances blood glucose homeostasis and lipid profiles in mice with obesity induced by HFD. Furthermore, it was found that an HFD supplemented with ferulic acid preserved ADMSC populations (Scal-1 + CD 45-) in mice compared with those fed an HFD alone(Reference Cho and Park24).
In this study, we consistently observed that mice with HFD-induced obesity treated with curcumin exhibited lesser weight gain, better blood glucose homeostasis and lipid profiles than the mice fed HFD alone. Curcumin effectively attenuated hepatic steatosis as evidenced by a significant reduction in hepatic TG accumulation and NAS score. Evidence from previous studies suggests that curcumin has a potential therapeutic effect against obesity and obesity-related metabolic complications(Reference Mokgalaboni, Ntamo and Ziqubu18,Reference Hassan, Awadalla and Abd El-Kader19) . Curcumin is known to exert its anti-obesity effects by enhancing cellular antioxidant capacity, reducing inflammatory signalling and suppressing the preadipocytes differentiation(Reference Hassan, Awadalla and Abd El-Kader19,Reference Ferguson, Nam and Morrison26) . In metabolic disorders like fatty liver disease, curcumin has similarly been associated with decreased lipid levels, reduced hepatic fat accumulation and lower liver enzyme markers including Alanine Transaminase (ALT) and Aspartate Aminotransferase (AST)(Reference Mokgalaboni, Ntamo and Ziqubu18,Reference Jiménez-Flores, López-Briones and Macías-Cervantes27) .
Furthermore, mice on HFD with curcumin had increased ADMSC (Sca-1 + CD45-) populations than the mice fed on HFD alone. In addition, our data showed that curcumin enhanced the pCHK1(S345) protein expression in HFD-induced obese mice and CHK2 protein expression in human preadipocytes. These results suggested that curcumin may involve playing a regulatory role in obesity-related diseases by activating the cellular DNA repair signalling.
CHK1/2 (serine/threonine kinases) are necessary for the activation of DNA repair mechanisms, checkpoint-mediated cell cycle arrest and apoptosis in response to DNA damage(Reference Xu, Barnes and Yang28,Reference Guilherme, Virbasius and Puri29) . It negatively modulates cell cycle progression during the S phase of cycles and promotes the transcription of genes associated with HR in DNA repair. We observed that HFD + CUR mice had higher pCHK1(S345) protein levels in the fat tissues than HFD and SC mice. We also speculate that curcumin may influence cell cycle progression in mouse adipose tissues and enhance DNA repair via HR.
Chronic inflammation is a hallmark of obesity(Reference Xu, Barnes and Yang28,Reference Guilherme, Virbasius and Puri29) . Inflammation is crucial for adipocyte function. Obesity, due to its low antioxidant capacity and abnormal inflammatory response, may lead to oxidative stress, inflammation, DNA damage and repair signalling. During the inflammatory response, adipocytes release various endocrine factors, such as LEP, adiponectin and cytokines. Cytokine release regulates a variety of cellular signals that can prevent or aggravate metabolic complications such as obesity(Reference Xu, Barnes and Yang28,Reference Guilherme, Virbasius and Puri29) . For example, as adipocytes increase, the production of inflammatory cytokines including IL-6 and CCL2, is accelerated. We previously showed that damaged cells increase secretion of inflammatory cytokines, including IL-6 and CCL2 and persistent DNA damage lesions bearing DNA double-strand breaks(Reference Lee, Wu and Fried1,Reference Mokgalaboni, Ntamo and Ziqubu18,Reference Cho and Park24) . We examined IL-6 and CCL2 levels in H2O2-treated preadipocytes to investigate the relationship between inflammation and DNA repair signalling. Our data showed that curcumin reduced the expression of inflammatory genes, including IL-6 and CCL2 mRNA levels, in preadipocyte cultures with H2O2 treatment, as an oxidative stress inducer, enhanced DNA repair response signaling and increased adipocyte differentiation. Additionally, curcumin reduced the expression of the adipogenic genes LEP, ADIPOQ, LPL and PPARG during preadipocyte-to-adipocyte differentiation. Curcumin consumption decreased serum LEP levels, which is consistent with the reported anti-inflammatory properties of curcumin. These findings suggest that curcumin may participate in the link between reduced HFD-induced oxidative stress, anti-inflammatory response and enhanced DNA repair response in mice with HFD-induced obesity in vitro.
Mesenchymal stem cells are adult stem cells and a type of progenitor cell that can proliferate and later differentiate into osteogenic cells(Reference Lumeng, Bodzin and Saltiel4). Mesenchymal stem cells can be isolated from various tissues, particularly adipose tissue and bone marrow. Adipose-derived mesenchymal stem cells can be isolated and differentiated into mature adipocytes in vitro. Primary preadipocyte cultures may better reflect the context of in vivo adipose function and are an appropriate cell system to confirm data obtained from preadipocyte lineages. Primary preadipocytes do not undergo continuous passage and therefore remain diploid, which better reflects the in vivo situation. This study showed that curcumin reduced the ADMSC population in HFD models*** in vivo and inhibited adipogenesis in primary preadipocytes in vitro.
However, this study has both strengths and limitations. First, we showed that curcumin reduced the pro-inflammatory response and enhanced DNA repair signalling in mice with HFD-induced obesity and in preadipocytes. Hyperglycemia-induced oxidative stress is a major contributor to HFD-induced obesity(Reference Lee, Wu and Fried1,Reference Cho and Park24) . Excessive oxidative stress can cause the dysregulation of inflammation, DNA damage and adipocyte repair. Our study supports the beneficial effects of curcumin on the link between reduced oxidative stress, anti-inflammation and enhanced DNA repair responses in mice with HFD-induced obesity, ADMSC system and preadipocyte culture systems, both in vivo and in vitro.
Second, we suggest a role for ADMSCs in obesity that does not involve curcumin-induced maintenance of ADMSC via a direct mechanism. Curcumin exerts protective effects on the proliferation and differentiation of stem cells(Reference Wang, Yan and Xu30). The effects of ADMSC, as determined using mouse models, include weight loss and mitigation of obesity-related phenotypes, such as dyslipidemia, glucose metabolism homeostasis, inflammation and non-alcoholic fatty liver disease(Reference Cho and Park24,Reference Ong, Tan and Chan31,Reference Pincu, Huntsman and Zou32) and regulation of anti-inflammatory effects in response to the microenvironment(Reference Uccelli, Moretta and Pistoia33).
Third, curcumin exerts its pharmacological effects by modifying several targets with anti-inflammatory properties, in a non-cell-autonomous manner and with poor bioavailability(Reference Ferguson, Nam and Morrison26,Reference Wang, Yan and Xu30) . We show that curcumin reduces hepatic TAG lipid accumulation and improves insulin sensitivity in mice fed a HFD. Furthermore, we observed that curcumin enhances the DNA repair response of mouse adipose tissue by pCHK1, but we could not confirm whether this was direct or indirect manner, or whether it was bioavailable. Therefore, we need further studies on the cell autonomous mode of curcumin effects.
In terms of bioavailability, one study reported that when curcumin was orally administered at 400 mg in rodents, absorption was detected at 5 mg/l in the hepatic portal vein up to 24 h after oral injection(Reference Ravindranath and Chandrasekhara34,Reference Xue, Cheng and Xu35) . In this study, we observed that mice fed curcumin at a dose of 2·5 g/kg diet showed improved weight loss, glucose homeostasis and lipid profiles. In clinical studies, curcumin bioactivity has shown that doses as low as 3·6 g/d are sufficient to decrease the responsiveness of circulating leukocytes to lipopolysaccharides(Reference Sharm, Euden and Platton36). The curcumin dose used in this study may have anti-inflammatory effects in human subjects based on a reasonable dosing schedule. Furthermore, advances in the field of nanoparticles are also one way to develop curcumin–based complexes as effective therapeutic agents. Despite these limitations, this is the first study to provide in vivo and in vitro evidence that curcumin modulates DNA damage response signalling via CHK1/CHK2 activation in obesity.
Conclusions
Our findings showed that curcumin suppressed weight gain in mice with HFD-induced obesity and reduced DNA damage in the adipose tissue in vivo and in vitro. These findings suggest that curcumin has potential applications in DNA repair-related therapy for obesity.
Supplementary material
For supplementary material/s referred to in this article, please visit https://doi.org/10.1017/S000711452510353X
Acknowledgements
The authors wish to thank Dr. Susan Fried and Dr. Mi-jeong Lee for sharing the valuable protocol of adipocyte differentiation. Also, we thank Hyuna Lee and Hyelim Park for helping with the animal experiments.
This study was supported by the 2024 Hannam University Research Fund (H2024, E.P.) and the National Research Foundation Grant funded by the Korean Government (NRF-2017R1A2B4007357, J.C.).
E.P. and J.C. contributed to the study conceptualisation and design. E.P. and J.C. analysed the data. J.C. wrote the original draft of the manuscript. E.P. revised the manuscript. All authors have agreed and read the published version of the manuscript.
The authors have no conflict of interest to declare.