


Decades of research examining the association between endogenous testosterone and human aggression have concluded that the relationship is positive but small in effect size (Archer, Reference Archer2006; Geniole et al., Reference Geniole, Bird, McVittie, Purcell, Archer and Carré2020). Meta-analyses also suggest that this effect tends to be slightly stronger in men than in women (Geniole et al., Reference Geniole, Bird, McVittie, Purcell, Archer and Carré2020). To explain these findings, several theoretical models have proposed that the influence of testosterone on aggression is moderated by other variables, such as levels of cortisol (Mehta & Josephs, Reference Mehta and Josephs2010); situational characteristics, such as provocation and victimization (Calvete & Orue, Reference Calvete and Orue2024); and the type of aggression being examined (Armstrong et al., Reference Armstrong, Wells, Boisvert, Lewis, Cooke, Woeckener and Kavish2021; Cima et al., Reference Cima, Meulenbeek, Spagnuolo, Oosterink, Thijssen, Smeijers, Valenzuela Pascual, Mouratidis, Oosterling, Riem and Loheide-Niesmann2025). However, most studies have focused on adults, while research conducted during adolescence remains limited. Adolescence is characterized by a significant rise in testosterone levels, particularly among boys (Shirtcliff et al., Reference Shirtcliff, Allison, Armstrong, Slattery, Kalin and Essex2012), peaking between the ages of 15 and 17 and stabilizing later in early adulthood (Luo et al., Reference Luo, Dashti, Sawyer and Vijayakumar2024).
At the same time, during adolescence there is a shift in relationships with peers, which take on a prominent role in the process of developing autonomy from the family. Thus, adolescents spend more time with their peers and developing friendship bonds (Merrin et al., Reference Merrin, Wang, Kiefer, Jackson, Pascarella, Huckaby, Blake, Gomez and Smith2024). However, peer relationships also involve the risk of exposure to bullying behaviors. Bullying perpetration toward peers is an important form of aggressive behavior during adolescence (Halliday et al., Reference Halliday, Gregory, Taylor, Digenis and Turnbull2021). Bullying has been defined by the Centers for Disease Control and Prevention as “unwanted aggressive behavior(s) by another youth or group of youth who are not siblings or current dating partners that involves an observed or perceived power imbalance and is repeated multiple times or is highly likely to be repeated” (Gladden et al., Reference Gladden, Vivolo-Kantor, Hamburger and Lumpkin2014, p. 7). Bullying perpetration can take many forms, including physical aggression (e.g., hitting), verbal aggression (e.g., threatening and insulting), and relational aggression (e.g., spreading negative comments about the victim). Bullying perpetration typically peaks in adolescence (Dake et al., Reference Dake, Price and Telljohann2003) and decreases between early and late adolescence (Kretschmer et al., Reference Kretschmer, Veenstra, Dekovic and Oldehinkel2017). Forms of bullying also evolve during adolescence, such that physical aggression tends to give way to more complex relational aggression than that observed at earlier ages (Merrin et al., Reference Merrin, Wang, Kiefer, Jackson, Pascarella, Huckaby, Blake, Gomez and Smith2024). There is also a strong reciprocal relationship between bullying perpetration and peer victimization, as experiences of victimization can provoke aggressive reactions that, in turn, heighten the risk of subsequent bullying (Morea & Calvete, Reference Morea and Calvete2022). Therefore, analyzing the relationship between testosterone and bullying perpetration, as well as the factors that modulate this link, is especially relevant during this developmental stage. This study investigates the role of testosterone in bullying perpetration in adolescents, with a focus on the moderating influence of cortisol levels and peer victimization experiences.
Cortisol as a moderator of the testosterone–aggression link
Testosterone and cortisol are produced by the hypothalamic–pituitary–gonadal axis and the hypothalamic–pituitary–adrenal axis, respectively (Terburg et al., Reference Terburg, Morgan and van Honk2009), and are associated with different behavioral mechanisms. Testosterone has been linked to the behavioral activation system and to reward-seeking behaviors, such as social status and dominance. By contrast, cortisol has been linked to the behavioral inhibition system, stress responses, negative emotions, sensitivity to punishment, and social withdrawal (Arnett, Reference Arnett1997; Terburg et al., Reference Terburg, Morgan and van Honk2009).
Given these contrasting behavioral correlates, the dual hormone hypothesis (DHH; Mehta & Josephs, Reference Mehta and Josephs2010) posits that the effects of testosterone on aggression and other risk-related behaviors are moderated by cortisol levels. Specifically, the hypothesis suggests that high testosterone levels lead to aggressive behavior when cortisol levels are low, while high cortisol levels inhibit the effect of testosterone on aggression due to cortisol’s association with fear, social withdrawal, and punishment sensitivity.
Empirical evaluations of the DHH have primarily employed two statistical approaches. The first involves calculating the testosterone/cortisol ratio, under the assumption that a higher level of testosterone relative to cortisol predicts greater aggression (Korpel et al., Reference Korpel, Varkevisser, Hoppenbrouwers, Van Honk and Geuze2019; Platje et al., Reference Platje, Popma, Vermeiren, Doreleijers, Meeus, van Lier, Koot, Branje and Jansen2015). The second uses the testosterone × cortisol interaction term. The latter approach has been argued to offer certain advantages. For instance, the testosterone/cortisol ratio cannot differentiate between combinations in which both hormones are similarly high or low (as the ratio approaches 1), whereas the interaction term better captures the context-sensitive, conditional effects of each hormone (Calvete & Orue, Reference Calvete and Orue2024; Cima et al., Reference Cima, Meulenbeek, Spagnuolo, Oosterink, Thijssen, Smeijers, Valenzuela Pascual, Mouratidis, Oosterling, Riem and Loheide-Niesmann2025).
Regardless of the method used, empirical support for DHH remains mixed. A meta-analysis by Dekkers et al. (Reference Dekkers, van Rentergem, Meijer, Popma, Wagemaker and Huizenga2019) reported a very small effect size for the testosterone × cortisol interaction on status-relevant behaviors, with most included studies focusing on adult populations. However, it has been proposed that the hormonal dynamics between testosterone and cortisol might differ during adolescence, a developmental stage marked by profound hormone changes (Mazur & Booth, Reference Mazur and Booth1998; Shirtcliff et al., Reference Shirtcliff, Allison, Armstrong, Slattery, Kalin and Essex2012), potentially altering their behavioral effects (Cima et al., Reference Cima, Meulenbeek, Spagnuolo, Oosterink, Thijssen, Smeijers, Valenzuela Pascual, Mouratidis, Oosterling, Riem and Loheide-Niesmann2025).
In a systematic review, Cima et al. (Reference Cima, Meulenbeek, Spagnuolo, Oosterink, Thijssen, Smeijers, Valenzuela Pascual, Mouratidis, Oosterling, Riem and Loheide-Niesmann2025) identified only seven studies that examined the interaction between testosterone and cortisol in adolescent samples. Only three of these studies – each using cross-sectional designs – found support for the DHH, showing significant associations between testosterone and aggression at low cortisol levels but not at high cortisol levels. Two of these studies used incarcerated samples (Dabbs et al., Reference Dabbs, Jurkovic and Frady1991; Popma et al., Reference Popma, Vermeiren, Geluk, Rinne, van den Brink, Knol, Jansen, van Engeland and Doreleijers2007), and one used a healthy community sample (Tackett et al., Reference Tackett, Herzhoff, Harden, Page-Gould and Josephs2014). Two other studies reported nonsignificant testosterone × cortisol interactions (Platje et al., Reference Platje, Popma, Vermeiren, Doreleijers, Meeus, van Lier, Koot, Branje and Jansen2015; Ren et al., Reference Ren, Zhao, Yu, Zhao, Zhang, Lin and Zhang2020). Notably, in a large mixed-gender sample of 259 adolescents, Platje et al. (Reference Platje, Popma, Vermeiren, Doreleijers, Meeus, van Lier, Koot, Branje and Jansen2015) compared both the interaction and ratio methods. They found that the testosterone × cortisol interaction was not significantly associated with aggression, whereas the testosterone/cortisol ratio was – but only in girls.
Other studies found significant testosterone × cortisol interactions with patterns different from those proposed by the DHH. For instance, Nguyen et al. (Reference Nguyen, Jones, Elgbeili, Monnier, Yu, Laplante and King2018) reported that in a small adolescent sample, testosterone was not associated with aggression when cortisol was low but was negatively associated when cortisol was high. In a longitudinal study, Susman et al. (Reference Susman, Peckins, Bowes and Dorn2017) observed that boys with either low diurnal testosterone and low cortisol reactivity or high testosterone and high cortisol reactivity showed increased conduct problems and antisocial behavior. No significant findings emerged for girls. Their results are somewhat similar to those obtained by Armstrong et al. (Reference Armstrong, Wells, Boisvert, Lewis, Cooke, Woeckener and Kavish2021) in adults. They found that both low–low and high–high baseline levels of testosterone and cortisol were linked to higher reactive aggression in response to social stress, although these interactions were not statistically significant in men.
These mixed findings have led some authors to suggest that both low and high cortisol levels may relate to aggression. Given cortisol’s role as a marker of stress system activation (Armstrong et al., Reference Armstrong, Boisvert, Wells, Lewis, Cooke, Woeckener, Kavish, Vietto and Harper2022; Raine, Reference Raine2002), both hypo-arousal (low cortisol and low stress reactivity) and hyper-arousal (high cortisol, high stress, and negative affect, such as anger) may facilitate aggression, although possibly through different mechanisms (Cima et al., Reference Cima, Meulenbeek, Spagnuolo, Oosterink, Thijssen, Smeijers, Valenzuela Pascual, Mouratidis, Oosterling, Riem and Loheide-Niesmann2025).
Situational moderators of the testosterone–Cortisol effect
Findings regarding reactive aggression support the view that hormonal effects on behavior may depend on situational moderators, such as peer victimization. Some studies have examined hormonal responses specifically in contexts of provocation and victimization. For example, in a study with undergraduate women, higher testosterone levels were associated with revenge responses to provocation but only when cortisol levels were high (Denson et al., Reference Denson, Mehta and Ho Tan2013). This is consistent with the idea that high testosterone and cortisol levels predict heightened aggression in threatening social contexts.
In a large longitudinal study with 577 adolescents and repeated measures every 6 months, Calvete and Orue (Reference Calvete and Orue2024) found a significant three-way interaction between testosterone, cortisol, and peer victimization. Interestingly, this interaction did not align with the DHH but was similar to the patterns found by Susman et al. (Reference Susman, Peckins, Bowes and Dorn2017). Specifically, adolescents with either high testosterone and high cortisol or low testosterone and low cortisol exhibited the highest levels of bullying and cyberbullying perpetration in response to victimization. By contrast, testosterone/cortisol ratio effects were only significant among girls, with high ratios associated with higher cyberbullying perpetration as a reaction to peer victimization. These findings underscore the importance of contextual moderators, aligning with the General Aggression Model (GAM; Allen et al., Reference Allen, Anderson and Bushman2018; Anderson & Bushman, Reference Anderson and Bushman2002). According to the GAM, aggression arises from the interaction between situational factors (e.g., peer victimization) and personal characteristics (e.g., hormonal levels).
The current study
Despite the longstanding interest in the role of testosterone in aggression, studies in humans have generally found weak associations (Geniole et al., Reference Geniole, Bird, McVittie, Purcell, Archer and Carré2020). The DHH has provided a valuable framework to explain this, proposing that testosterone’s effects on aggression are moderated by cortisol (Mehta & Josephs, Reference Mehta and Josephs2010; Mehta & Prasad, Reference Mehta and Prasad2015). However, empirical evidence has been inconsistent, and significant testosterone × cortisol interactions often deviate from DHH predictions (Cima et al., Reference Cima, Meulenbeek, Spagnuolo, Oosterink, Thijssen, Smeijers, Valenzuela Pascual, Mouratidis, Oosterling, Riem and Loheide-Niesmann2025; Dekkers et al., Reference Dekkers, van Rentergem, Meijer, Popma, Wagemaker and Huizenga2019). Methodological challenges in this field include the need for larger samples to ensure sufficient statistical power for the testosterone × cortisol interaction (e.g., ∼400 participants for 80% power) and the implementation of longitudinal designs with multiple waves (Grebe et al., Reference Grebe, Del Giudice, Thompson, Nickels, Ponzi, Zilioli, Maestripieri and Gangestad2019).
Within this context, the current study aims to advance the understanding of the interplay between testosterone and cortisol in bullying perpetration during adolescence – a developmental period marked by profound hormonal shifts (Mazur & Booth, Reference Mazur and Booth1998; Shirtcliff et al., Reference Shirtcliff, Allison, Armstrong, Slattery, Kalin and Essex2012). Adolescence is also a stage of heightened peer sensitivity, in which experiences of bullying victimization can act as powerful triggers of bullying perpetration (Morea & Calvete, Reference Morea and Calvete2022). Grounded in the GAM (Allen et al., Reference Allen, Anderson and Bushman2018; Anderson & Bushman, Reference Anderson and Bushman2002), we hypothesized that bullying victimization (1) would increase the association between testosterone and bullying perpetration and (2) would strengthen the combined influence of testosterone and cortisol on adolescents’ bullying perpetration.
From a methodological standpoint, this study sought to address some of the mentioned previous limitations by using a large adolescent sample and a longitudinal design. Moreover, in contrast to most prior research, we employed a random-intercept cross-lagged panel model (Hamaker et al., Reference Hamaker, Kuiper and Grasman2015), which included reciprocal longitudinal associations between hormones, bullying victimization, and bullying perpetration over time. Animal studies have suggested bidirectional relationships between testosterone and aggression, depending on whether analyses are conducted at the between-individual or within-individual level (George & Rosvall, Reference George and Rosvall2022). Furthermore, in humans, cortisol is well known to increase in response to social stress (Goodman et al., Reference Goodman, Janson and Wolf2017), and increases in testosterone have been observed in adolescents exposed to social stressors (Rodgers & Kuhlman, Reference Rodgers and Kuhlman2023). Thus, we explored whether both testosterone and cortisol could increase as a consequence of peer victimization. However, as there were not previous studies examining these associations, no hypotheses were stated in this study.
Methods
Participants
The sample consisted of 471 adolescents (Mage = 14.97, SD = 0.70 at Wave 1), of whom 47.1% identified as female, 51.2% as male, 0.8% as nonbinary, and 0.8% as “other.” The participants were from five high schools in Bizkaia (Basque Country, Spain). The Spanish National Institute of Statistics’ occupational criteria were used to measure sociodemographic status. For this purpose, the adolescents were asked about their parents’ occupations, which were distributed as follows: 7.64% were directors and managers; 27.28% were scientific or intellectual professionals; 7.96% were support technicians and professionals; 6.48% were administrative employees; 12.74% were catering, personal protection, and retail sales workers; 0.11% were skilled workers in agriculture and fishing; 10.62% were skilled workers in the manufacturing and construction industries; 4.03% were plant and machine operators and assemblers; 4.03% were unskilled workers; 1.70% were housekeepers; 1.06% were unemployed individuals; and 16.35% did not answer.
Measures
Salivary samples were collected in the classrooms using the Salivette® system (Sarstedt, Nümbrecht, Germany), a widely used and validated method for noninvasive hormone assessment in adolescent populations. Sample collection was scheduled approximately 2h after the adolescents woke up (M = 9.27 AM; SD = 1.32 min) to minimize the impact of diurnal variation on hormonal levels. The participants were instructed to abstain from eating, drinking (except water), exercising, smoking, or brushing their teeth for at least 30 min prior to collection. Two samples were eliminated because they did not meet the criteria for analysis.
Each participant was provided with a sterile Salivette® containing a cotton swab. Under supervision, they placed the swab in their mouth and held or gently chewed it for 1–2 min until adequately saturated. The swabs were then returned to their respective tubes, which were immediately stored at approximately 4°C. The exact time of collection was recorded for each participant. The samples were transported to the laboratory within 2 h and immediately frozen at −25°C and −30°C until analysis at the Reference Laboratory (Barcelona, Spain). Prior to the assay, the samples were thawed and centrifuged to separate clear saliva from the swab material.
Salivary cortisol and testosterone concentrations were assessed using competitive enzyme-linked immunosorbent assay (ELISA) kits from DiaMetra (Perugia, Italy), specifically calibrated for salivary matrices. Cortisol was quantified with the Cortisol Saliva ELISA kit (code DKO020), featuring a sensitivity of 0.12ng/mL and an assay range of 0.5–100ng/mL. Testosterone measurement employed the Testosterone Saliva ELISA kit (code DKO021), characterized by a sensitivity of 3.28pg/mL and an analytical range of 10–1,000pg/mL. Both assays employed a competitive colorimetric immunoenzymatic method and were performed according to the manufacturer’s protocols. The European Bullying Intervention Project questionnaire (EBIPQ; Ortega-Ruiz et al., Reference Ortega-Ruiz, Del Rey and Casas2016) was used to assess the subscales of bullying perpetration and bullying victimization (seven items each). The two subscales include physical, verbal, and social aggression (direct or indirect), as well as psychological aggression, using a five-point Likert scale ranging from 0 (never) to 4 (more than once a week) in the last 3 months. Sample items include “Someone has hit, kicked, or pushed me” (victimization) and “I have insulted or said mean things to another student” (perpetration). Responses to the items were averaged to obtain the scores, with higher scores indicating greater involvement in bullying behaviors. The EBIPQ has demonstrated excellent psychometric properties across adolescent samples. Supporting its convergent validity, previous studies have reported significant correlations between EBIPQ and other bullying and cyberbullying measures (Del Rey et al., Reference Del Rey, Casas, Ortega-Ruiz, Schultze-Krumbholz, Scheithauer, Smith, Thompson, Barkoukis, Tsorbatzoudis, Brighi, Guarini, Pyżalski and Plichta2015). In addition, previous research has shown good internal consistency for both subscales across Spanish (Pichel et al., Reference Pichel, Feijóo, Isorna, Varela and Rial2022) and other European samples (Del Rey et al., Reference Del Rey, Casas, Ortega-Ruiz, Schultze-Krumbholz, Scheithauer, Smith, Thompson, Barkoukis, Tsorbatzoudis, Brighi, Guarini, Pyżalski and Plichta2015). In the present study, the Cronbach’s coefficients were 0.71, 0.78, and 0.80 for bullying perpetration and 0.76, 0.78, and 0.79 for victimization at Wave 1 (W1), Wave 2 (W2), and Wave 3 (W3), respectively.
Procedure
The study protocol was approved by the ethical committee of University of Deusto (Ref. ETK-28/22-23). The adolescents and their parents received information about the study and were invited to give informed consent. Parental acceptance was 82.17%, and all adolescents whose parents consented agreed to participate (100%). The participants provided saliva samples in the classroom before completing the self-report questionnaires via Qualtrics®.
Statistical analysis
Preliminary analyses
Little’s test for Missing Completely at Random was not statistically significant (χ2(139) = 155, p = .173), indicating that the data were likely missing at random. Therefore, full information maximum likelihood estimation was used to handle missing data. Sixty-two outliers were identified in testosterone and/or cortisol and were winsorized at 3 SD above the mean. Winsorization was used to reduce the influence of outliers while maintaining the overall data structure, ensuring more stable and representative estimates of testosterone and cortisol levels. All variables were log-transformed to reduce skewness and kurtosis. Testosterone and cortisol values were standardized into T-scores (M = 50, SD = 10) prior to computing the testosterone/cortisol ratio.
Descriptive statistics and correlations were obtained using SPSS-28. Longitudinal associations among the study variables were examined using RI-CLPMs (Hamaker et al., Reference Hamaker, Kuiper and Grasman2015; Orth et al., Reference Orth, Clark, Donnellan and Robins2021), estimated with maximum likelihood in Mplus 8.11. A key advantage of RI-CLPMs over traditional CLPMs is their ability to disentangle within-person processes from between-person stable differences. This modeling approach accounts not only for temporal stability but also for time-invariant, trait-like stability by including a random intercept for each variable (Hamaker et al., Reference Hamaker, Kuiper and Grasman2015). Two sets of models were estimated: one based on the interaction between testosterone and cortisol, and the other based on the testosterone/cortisol ratio.
Models based on the testosterone × cortisol interaction
The testosterone × cortisol interaction models included bullying perpetration, bullying victimization, testosterone, and cortisol, along with the interaction terms testosterone × cortisol, testosterone × victimization, cortisol × victimization, and testosterone × cortisol × victimization. For each set of models, we first tested the measurement models and next the RI-CLPM models. We describe below the steps for the interaction models.
The measurement models
The confirmatory factor analysis (CFA) model, also known as the measurement model, represents a critical first step because it assesses the adequacy of the hypothesized relationships between the observed indicators (e.g., item parcels) and their underlying latent variables. The measurement model serves as the foundation for evaluating both the measurement properties of each construct and the overall fit of the CFA model. In this study, multiple indicators were used for latent variables, except for the hormonal variables (testosterone and cortisol) and their interaction (testosterone × cortisol), which were modeled using a single indicator. For latent constructs with multiple indicators, three item-parcels were created. Items were assigned to parcels based on an exploratory factor analysis to ensure that each parcel contained items with balanced factor loadings (Little et al., Reference Little, Cunningham, Shahar and Widaman2002). We conducted an exploratory factor analysis with the bullying perpetration items to obtain a single-factor solution. The items were ordered according to their factor loadings, from highest to lowest. Then, the item with the highest factor loading was included in the first parcel, the next in the second parcel, the third in the third parcel, and so on. The same parcel structure was used for parallel bullying victimization items. Latent interactions (e.g., testosterone × bullying victimization) were represented by three product terms (e.g., testosterone × victimization parcel 1, testosterone × victimization parcel 2, and testosterone × victimization parcel 3), following the double mean-centering procedure (Lin et al., Reference Lin, Wen, Marsh and Lin2010). The syntax indicating which items were included in each parcel is available as supplementary material (Table S1).
Following the recommendations outlined by (Osborne & Little, Reference Osborne, Little and Little2024), the initial measurement model (Model 1) was evaluated using effect coding to identify first-order latent variables (bullying, victimization, testosterone × victimization, cortisol × victimization, and testosterone × cortisol × victimization). Effect coding is a method of scaling (Little et al., Reference Little, Slegers and Card2006) that maintains the metric of the latent variables’ indicators. It consists of constraining the factor loadings to an average of 1.0 (instead of arbitrarily setting one loading to be 1). This constraint gives an estimate of the construct variance as the average contribution of each indicator. Similarly, the estimates of the intercepts are constrained to an average of 0. This constraint gives an estimate of the latent construct mean as the average mean of the indicators.
In the initial step to test the measurement model, a configural invariance model was estimated in which each first-order latent variable in each wave was separately parameterized using effect coding. Error terms for the same indicators across time were allowed to correlate, including parallel bullying perpetration and victimization parcels. Phantom or rescaling constructs were added to place all estimated relationships on a common standardized metric. With this aim, latent means were freely estimated, and variances were fixed to zero to define second-order latent variables (phantom constructs). Phantom variables were defined by regressing first-order latent variables onto their respective second-order constructs. These phantom variables were modeled with a mean of 0 and a variance of 1 to maintain metric consistency and facilitate interpretation. For single-indicator latent variables (i.e., hormonal indicators), phantom constructs were estimated directly from the indicator using the same constraints (mean = 0, variance = 1). The measurement errors of the single indicators were fixed to 0. In the configural model, phantom variables were allowed to correlate with themselves over time, but cross-time covariances among first-order latent variables, as well as covariances between second-and first-order latent variables, were fixed to zero.
As this study was longitudinal, checking whether the measurement model was invariant over time was necessary. Therefore, in the second step, we estimated a more restrictive metric invariance model (Model 2), which imposed that factor loadings were equal across waves. In the final step, we estimated the scalar invariance model (Model 3), which constrained intercepts to be equal across waves.
The structural models
Once the final measurement model was established, it served as the baseline for comparison with more parsimonious structural models. The structural models followed the RI-CLPM framework, partitioning variance in the variables into between-person and within-person components by including random intercepts for all eight variables.
Contrary to the approach of (Mulder & Hamaker, Reference Mulder and Hamaker2021), who constrained all RI factor loadings to 1, we followed the method proposed by (Osborne & Little, Reference Osborne, Little and Little2024). In this approach, factor loadings were freely estimated but constrained to be equal across indicators, and RI variances were fixed to 1. This approach places all RIs on a common metric. When RIs were included in the model, previously defined phantom variables became the within-person latent variables.
In this structural model (Model 4), following the approach of Osborne and Little (Reference Osborne, Little and Little2024), cross-time correlations among within-person latent variables (phantom variables) were removed, although their cross-sectional correlations were retained. The model included autoregressive and cross-lagged paths from all phantom variables at time t–1 to the corresponding constructs at time t. The RI-CLPM was initially estimated as a nonstationary process (allowing autoregressive and cross-lagged paths to vary over time) and compared with the measurement model. Next, since hormonal levels, as well as bullying perpetration and victimization, may be influenced by the age and sex of the participants, these two variables were included as covariates in the model and regressed on the RIs (Model 4b). With this aim, values other than boy and girl were considered missing. Finally, we tested a more parsimonious stationary model in which longitudinal paths were constrained to be time invariant (Model 5).
Models based on the testosterone/Cortisol ratio
Models based on the testosterone/cortisol ratio followed the same estimation procedures but included only four variables: bullying perpetration, bullying victimization, testosterone/cortisol ratio, and the testosterone/cortisol × victimization interaction term.
Model fit indicators
Model fit was evaluated using multiple indices: the comparative fit index (CFI), the Tucker–Lewis index (TLI), the root mean square error of approximation (RMSEA), and the standardized root mean square residual (SRMR). A good fit was defined as CFI and TLI values ≥ 0.95, RMSEA < 0.06, and SRMR < 0.08 (Little, Reference Little2024). For nested model comparisons, changes in model fit were assessed using the cutoff criteria of ∆CFI < 0.01 (Cheung & Rensvold, Reference Cheung and Rensvold2002) and ∆RMSEA < 0.015 (Chen, Reference Chen2007). We also examined whether the RMSEA point estimate from a more restrictive model fell within the confidence interval of the less restrictive model.
Results
Descriptive analyses and correlation coefficients between variables
Descriptive analyses and Spearman correlation coefficients between variables are included as supplementary material (Table S2). Cortisol and testosterone levels showed low to moderate positive correlations across waves, except that W1 cortisol was not significantly correlated with testosterone levels at W2 or W3. Most correlations between hormonal levels and self-reported victimization and bullying perpetration were small or nonsignificant. Victimization and bullying perpetration were moderately to strongly correlated within and across waves (range = 0.36 to 0.62), indicating stability and co-occurrence over time.
Models based on the testosterone × cortisol interaction
Table 1 summarizes the fit indices for the models estimated based on the testosterone × cortisol interaction. The measurement model was first evaluated. Model 1 (configural invariance) yielded moderate CFI and TLI values but excellent RMSEA and SRMR values. All factor loadings were statistically significant. Next, Model 2 (metric invariance), which constrained factor loadings to be equal across the three waves, was estimated. Comparison of Model 2 with Model 1 indicated that this constraint was acceptable according to the RMSEA criterion; however, the change in CFI slightly exceeded the ∆CFI < 0.01 cutoff. Inspection of the factor loadings suggested that these loadings might differ across waves for the latent variable testosterone × cortisol × victimization. Therefore, a revised model (Model 2b, partial metric invariance) was estimated, assuming metric invariance for all variables except testosterone × cortisol × victimization. This model met the criteria for acceptable changes in both CFI and RMSEA.
Fit indices for the testosterone × cortisol interaction models

Table 1 Long description
The table presents fit indices for models based on the testosterone cortisol interaction. It has 5 rows and 4 columns. The columns are labeled Fit indices, Comparisons, and Pass?. The rows are labeled with different models: Configural invariance, Metric invariance, Partial metric invariance, Scalar invariance, Nonstationary, Nonstationary with age and sex as covariates, and Stationary with age and sex as covariates. Each row contains specific values for chi-square, RMSEA, CFI, TLI, SRMR, and comparisons. The table indicates whether each model passed the criteria for acceptable changes in CFI and RMSEA.
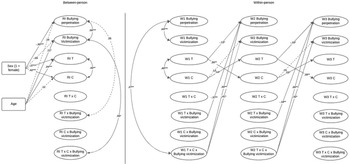
Finally, Model 3 (scalar invariance) added equality constraints on intercepts across time. This model resulted in a small increase in CFI and a reduction in RMSEA and was adopted as the baseline for the next phase of the analysis. In the next phase, the structural RI-CLPM was estimated in both nonstationary and stationary forms. Model 4 (nonstationary), compared to the scalar invariance model, showed a reduction in CFI below the cutoff, while the RMSEA remained similar. Figure 1 represents the measurement model of the within-person variables at W1.
Identification of the measurement model for within-person variables at wave 1 using a phantom variable approach. Note: Ovals represent the latent constructs whereas rectangles represent the manifest indicators (e.g., item-parcels and measures of hormones). Curved double-headed lines represent variance estimates and straight single-headed lines are factor loadings. T = Testosterone, C = Cortisol, BV = Bullying victimization, BP = Bullying perpetration. Item parcels are represented by p1, p2, and p3. *Indicates the parameter is fixed at the given value.

We then included age and sex as covariates of the random intercepts (RIs) for bullying perpetration, victimization, and hormones (Model 4b). Female sex was significantly associated with lower levels of bullying perpetration and testosterone. It was also marginally associated with lower levels of victimization and with higher levels of cortisol. Higher age was significantly associated with higher levels of victimization and testosterone, and marginally with higher levels of bullying perpetration and cortisol. This model served as the baseline for Model 5 (stationary), which imposed stability on the longitudinal paths.
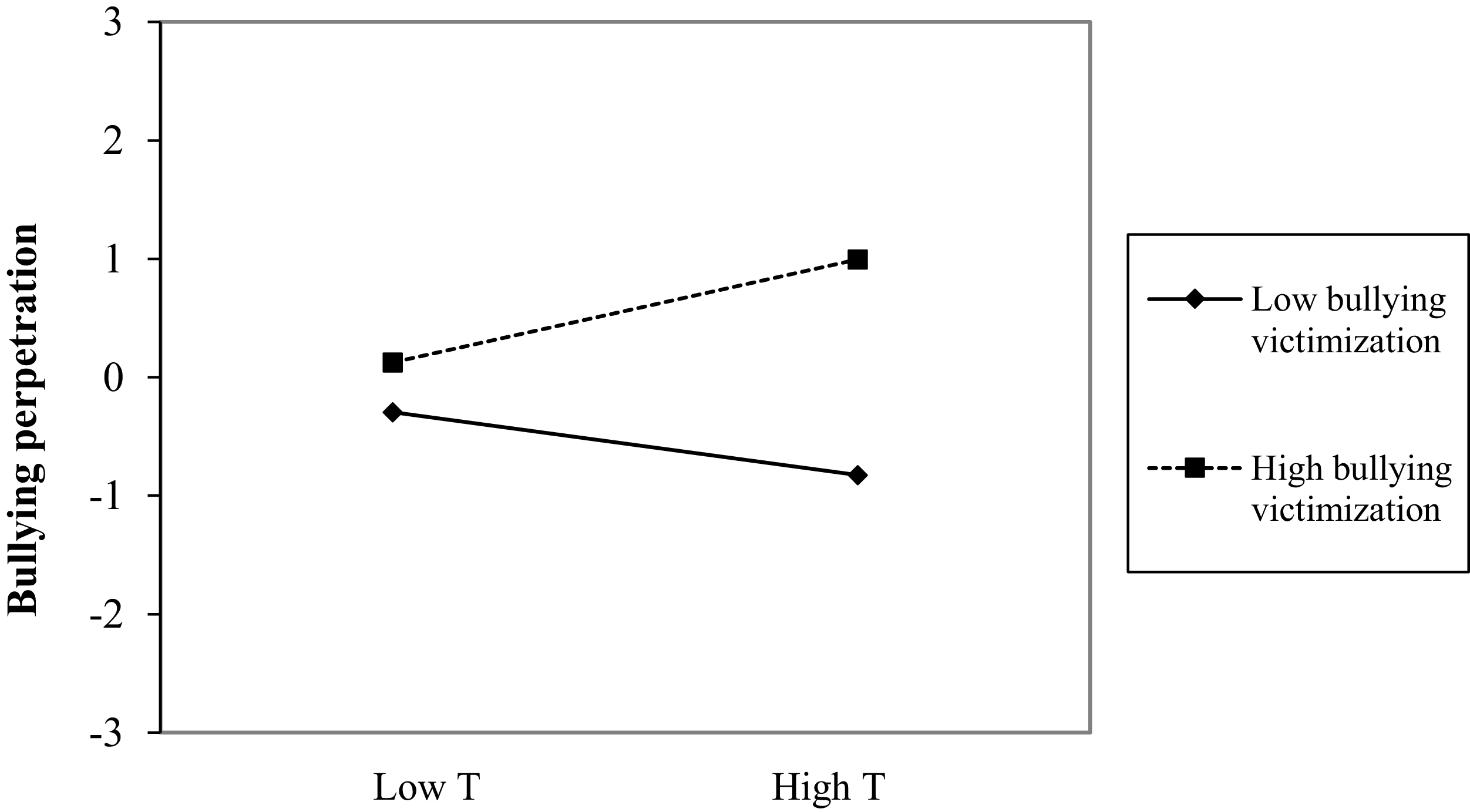
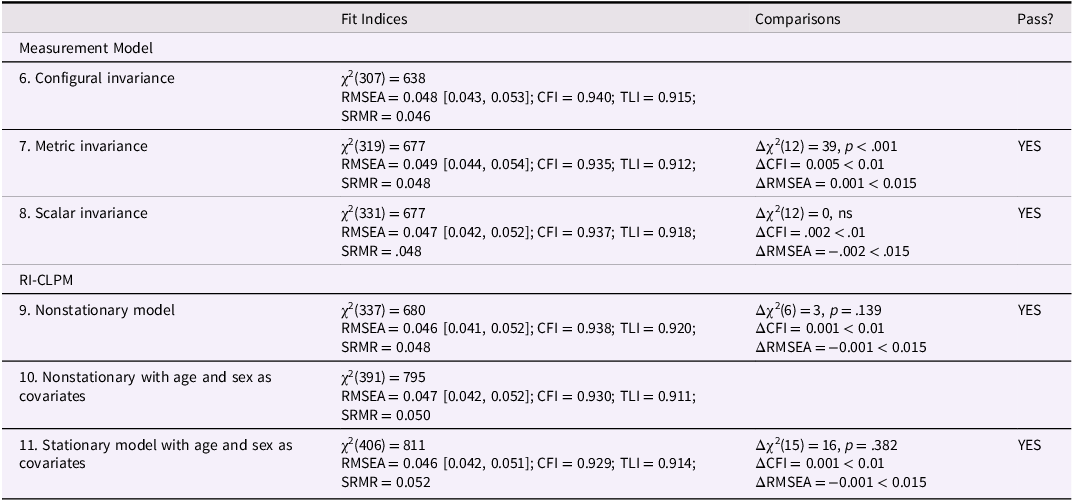
Figure 2 displays the structural paths of Model 5, and Supplementary Table S3 displays the coefficients of the model. At the between-person level, the RI for bullying perpetration was significantly associated with the RI for victimization (0.56, p < .001) and marginally significantly with the RI for testosterone × victimization (0.35, p = .052). Figure 3 shows the form of the interaction. For adolescents who usually experience high levels of victimization, levels of testosterone were marginally associated with levels of bullying perpetration (0.44, t = 1.93, p = .054). By contrast, for adolescents with low levels of victimization, testosterone levels were not associated with bullying perpetration (−0.27, t = −1.30, p = .193).
RI-CLPM model for testosterone × cortisol interaction. Note: For clarity, paths to interaction terms are not included. Dotted lines indicate marginally significant associations. * p < .05, ** p < .01; *** p < .001. T = testosterone, C = cortisol, W = wave.
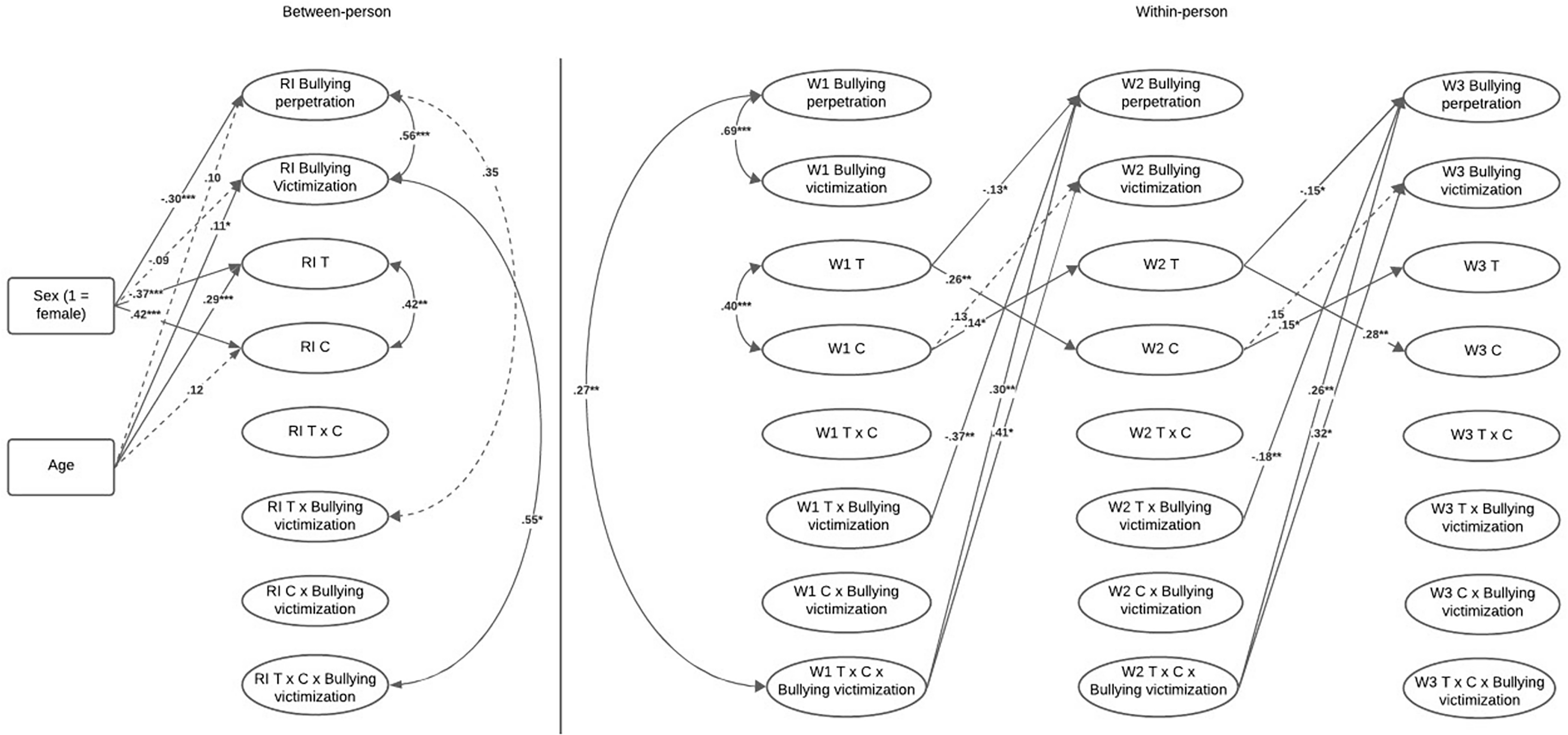
Figure 2 Long description
A diagram illustrating the relationships between testosterone, cortisol, bullying perpetration, and victimization in adolescents. Panel A: Between-person relationships. This panel shows the relationships between various factors at a single point in time. Key components include RI Bullying perpetration, RI Bullying victimization, RI Testosterone (T), RI Cortisol (C), and their interactions. Arrows indicate the direction of influence between these factors, with solid lines representing significant associations and dotted lines representing marginally significant associations. Age and sex are also included as influencing factors. Panel B: Within-person relationships. This panel shows the changes within individuals over three waves (W1, W2, W3). It includes similar components as Panel A but tracks their changes over time. Arrows indicate the direction of influence within individuals across the waves. The diagram uses specific labels and values to denote the strength and significance of these relationships.
Testosterone × victimization interaction for bullying perpetration at the between-person level. Note: T = testosterone.
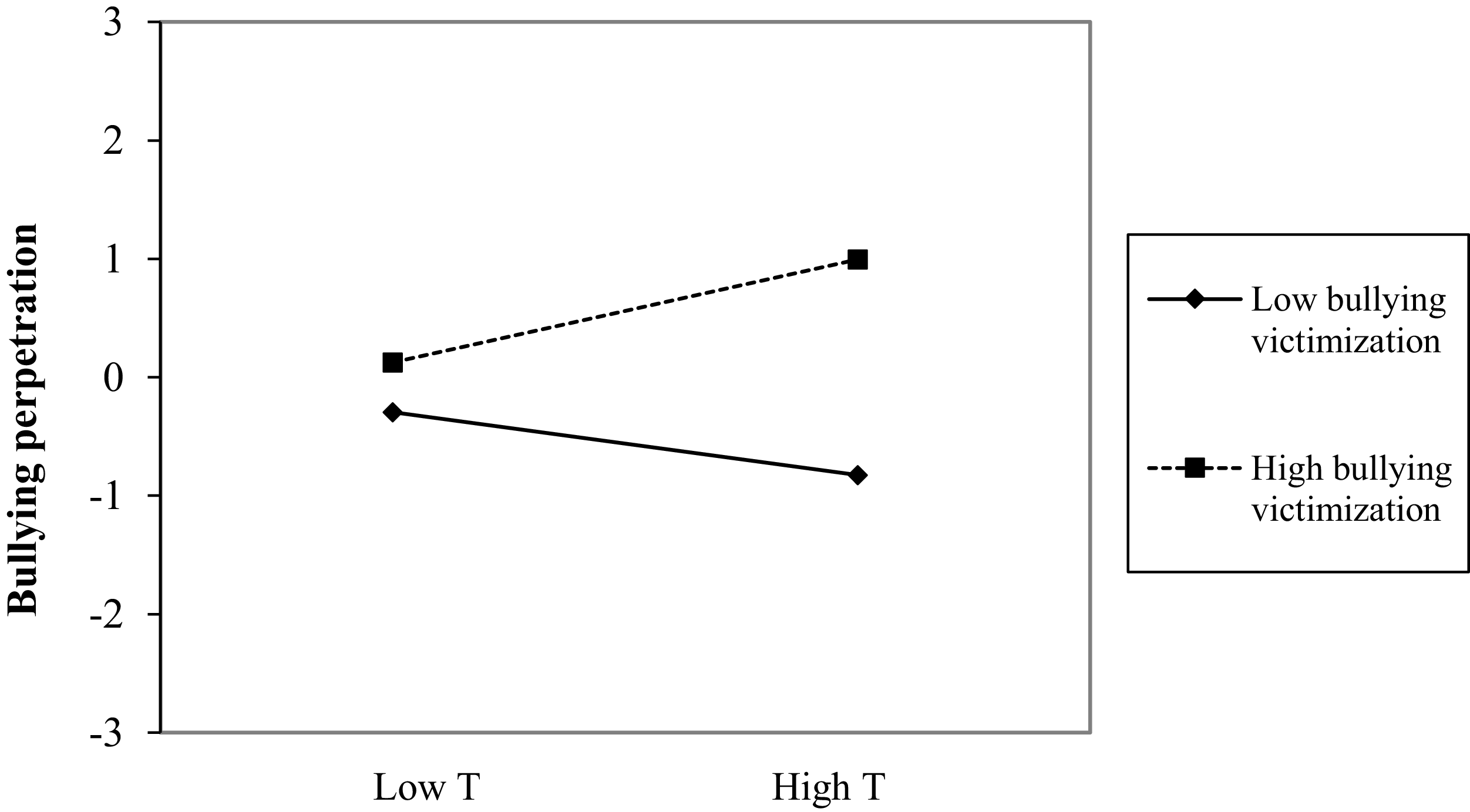
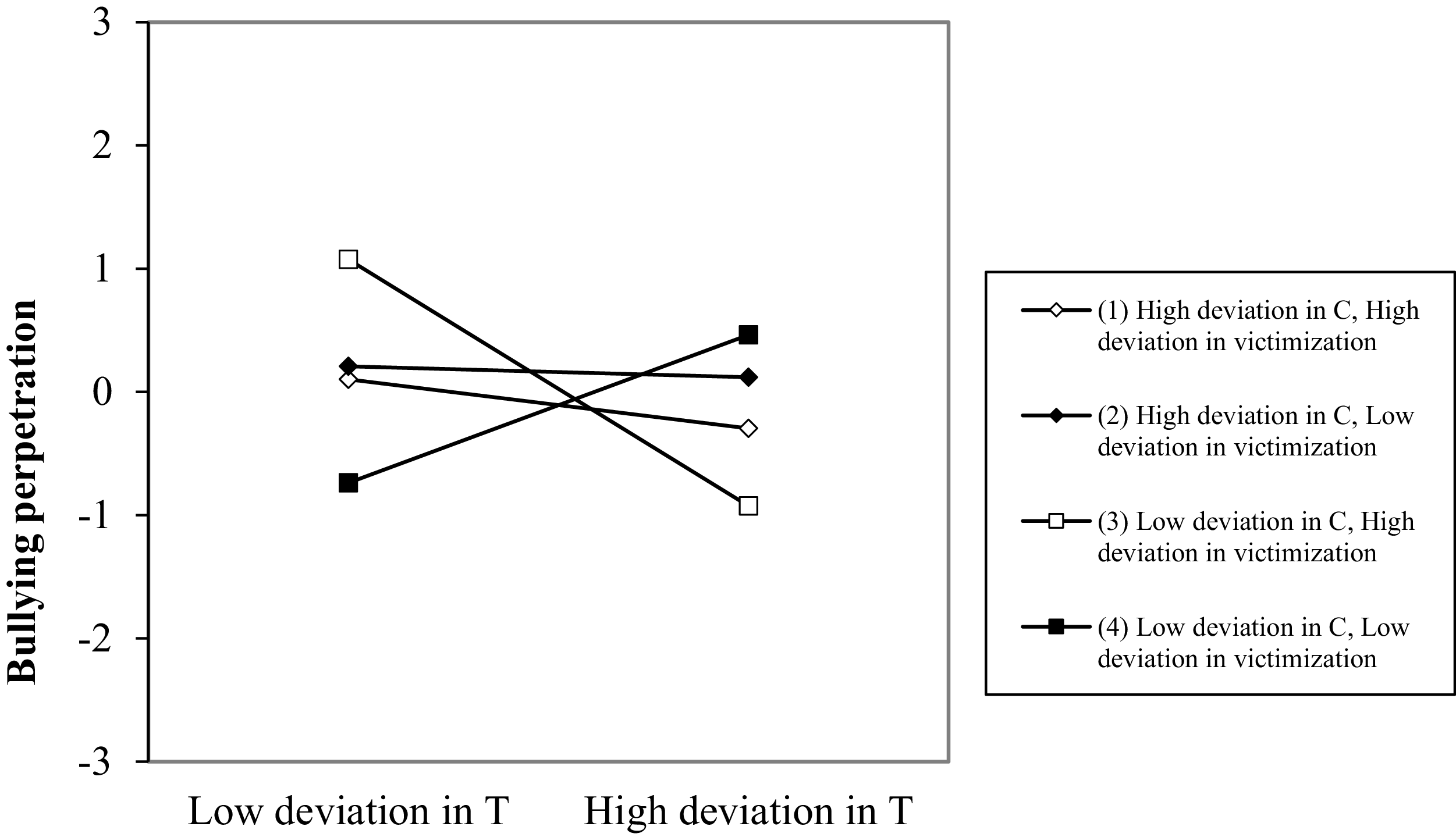
At the within-person level, deviations in testosterone predicted decreases in bullying perpetration. However, this main effect was moderated by cortisol and victimization, as both the testosterone × victimization and testosterone × cortisol × victimization interaction terms significantly predicted deviations in bullying perpetration in the subsequent wave. The graphical representation of the testosterone × cortisol × victimization interaction (Figure 4) showed that temporary deviations above usual levels of testosterone do not predict deviations in bullying perpetration when cortisol deviates above usual levels (−0.20, t = −1.03, p = .303; and −0.04, t = −0.17, p = .863, respectively, for deviations above and below usual levels in victimization). By contrast, when cortisol levels deviate below usual levels, deviations in testosterone predict different bullying perpetration outcomes depending on victimization levels. Specifically, when both cortisol and victimization levels are below usual, increases in testosterone predict an increase in bullying perpetration (0.60, t = 2.85, p = .005). By contrast, when cortisol is below usual but victimization is above usual, increases in testosterone predict lower bullying perpetration (−0.99, t = 5.52, p <. 001). All slopes differed significantly from one another, except for the two corresponding to above-usual deviations in cortisol.
Testosterone × cortisol × victimization interaction for bullying perpetration at the within-person level. Note: T = testosterone, C = cortisol.

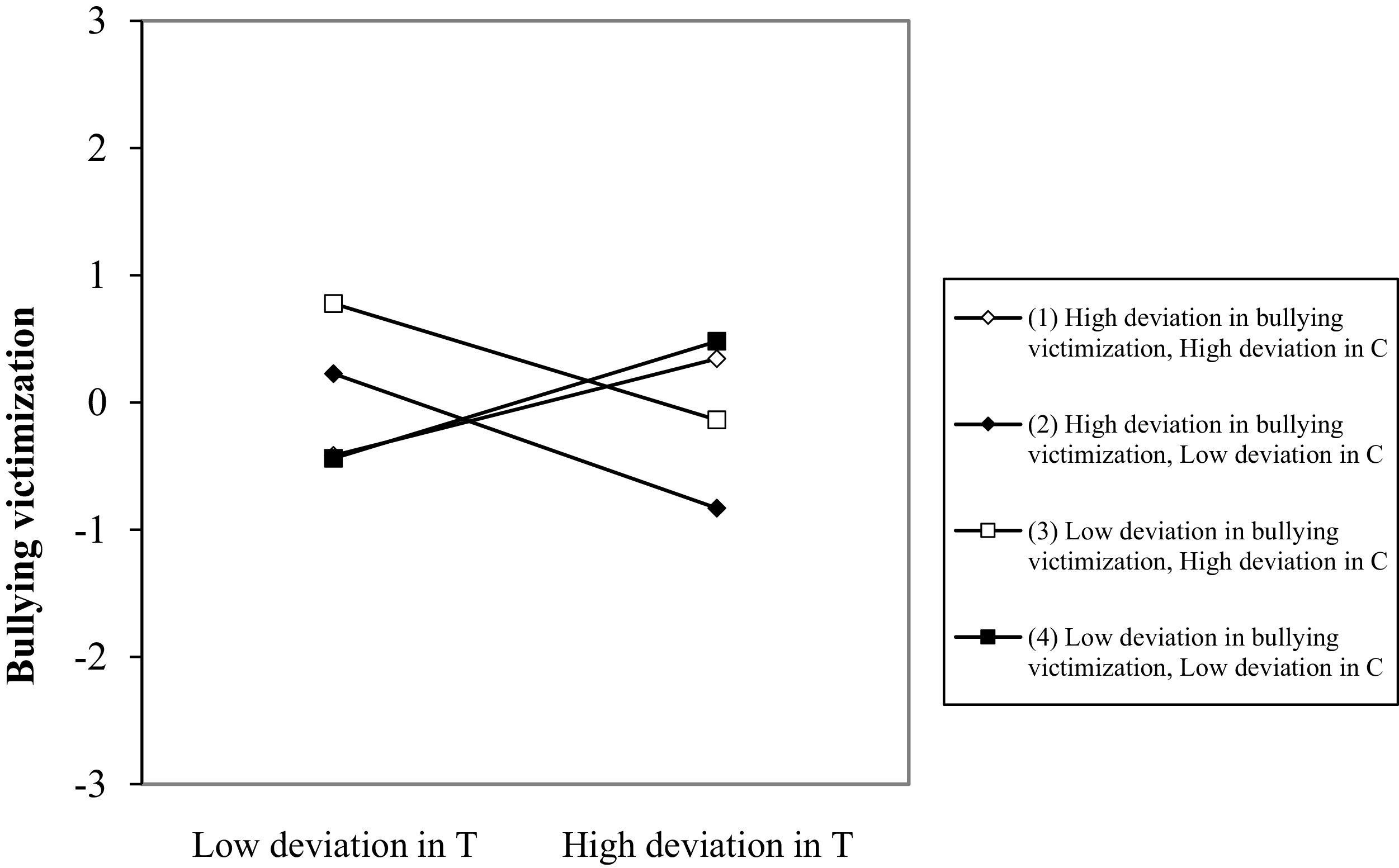
Although it was not the objective of this study, the model also showed that the testosterone × cortisol × victimization interaction covaried with victimization at the between-person level and predicted changes in victimization at the within-person level. Figure 5 shows the role of deviations in testosterone in victimization for low and high levels of cortisol and victimization. Increases in testosterone and reductions in cortisol relative to the adolescents’ usual levels predicted deviations in victimization in the next wave. Importantly, the shape of that prediction also depended on the changes experienced in victimization in the previous wave. Specifically, when the adolescent had experienced an increase in victimization experiences, the combination of an increase in testosterone and a reduction in cortisol predicted a decrease in future victimization (–0.53, t = –2.84, p = .005). By contrast, when the adolescent had experienced lower-than-usual levels of victimization, the same combination of increased testosterone and reduced cortisol predicted an increase in victimization in the next wave (0.46, t = 2.02, p = .044). The slopes were not statistically significant when cortisol increased, although the slope for increased cortisol and victimization was marginally significant (0.38, t = 1.71, p = .087).
Testosterone × cortisol × victimization interaction for victimization at the within-person level. Note: T = testosterone, C = cortisol.

Models based on the testosterone/Cortisol ratio
The models based on testosterone/cortisol ratio included measures of bullying perpetration and victimization, testosterone/cortisol ratio, and the testosterone/cortisol ratio × victimization interaction in the three waves. These models were specified following the same procedures described for the models based on the testosterone × cortisol interaction. Table 2 presents the fit indices for the measurement model and the RI-CLPM structural model. Model 6 (configural invariance) showed a good fit, and the constraints for metric and scalar invariance (Models 7 and 8) did not result in a loss of fit quality, as changes in both CFI and RMSEA remained within the established thresholds. Similarly, the nonstationary RI-CLPM model (Model 9) showed adequate fit indices, with changes within these criteria. The factor loadings of the RIs were statistically significant. Subsequently, paths from age and sex to bullying perpetration, victimization, and testosterone/cortisol ratio were included (Model 10). Consistent with the findings from the testosterone × cortisol interaction models, female sex was associated with lower testosterone/cortisol ratio scores and lower bullying perpetration scores, whereas age was positively associated with testosterone/cortisol ratio and victimization. Stationarity constraints were then imposed on this model, which proved acceptable, as changes in CFI and RMSEA values remained within the cutoff criteria (Model 11).
Fit indices for the testosterone/Cortisol models
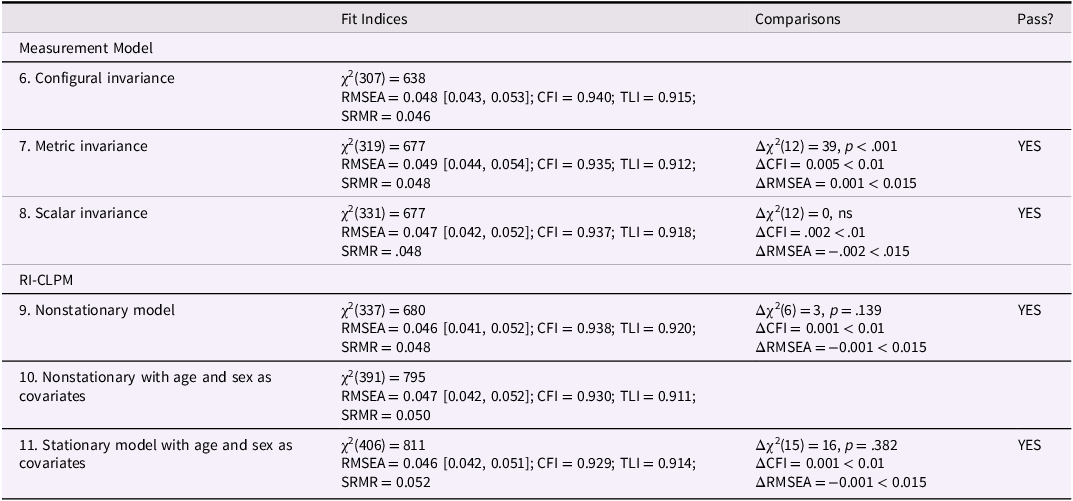
Table 2 Long description
The table presents fit indices for various models related to testosterone/Cortisol ratios. It includes columns for Fit Indices, Comparisons, and Pass?. The rows are labeled with different models: Configural invariance, Metric invariance, Scalar invariance, Nonstationary model, Nonstationary with age and sex as covariates, and Stationary model with age and sex as covariates. Each row contains values for chi-square, RMSEA with confidence intervals, CFI, TLI, SRMR, and comparisons with delta chi-square, delta CFI, and delta RMSEA. The table indicates whether each model passed the fit criteria.
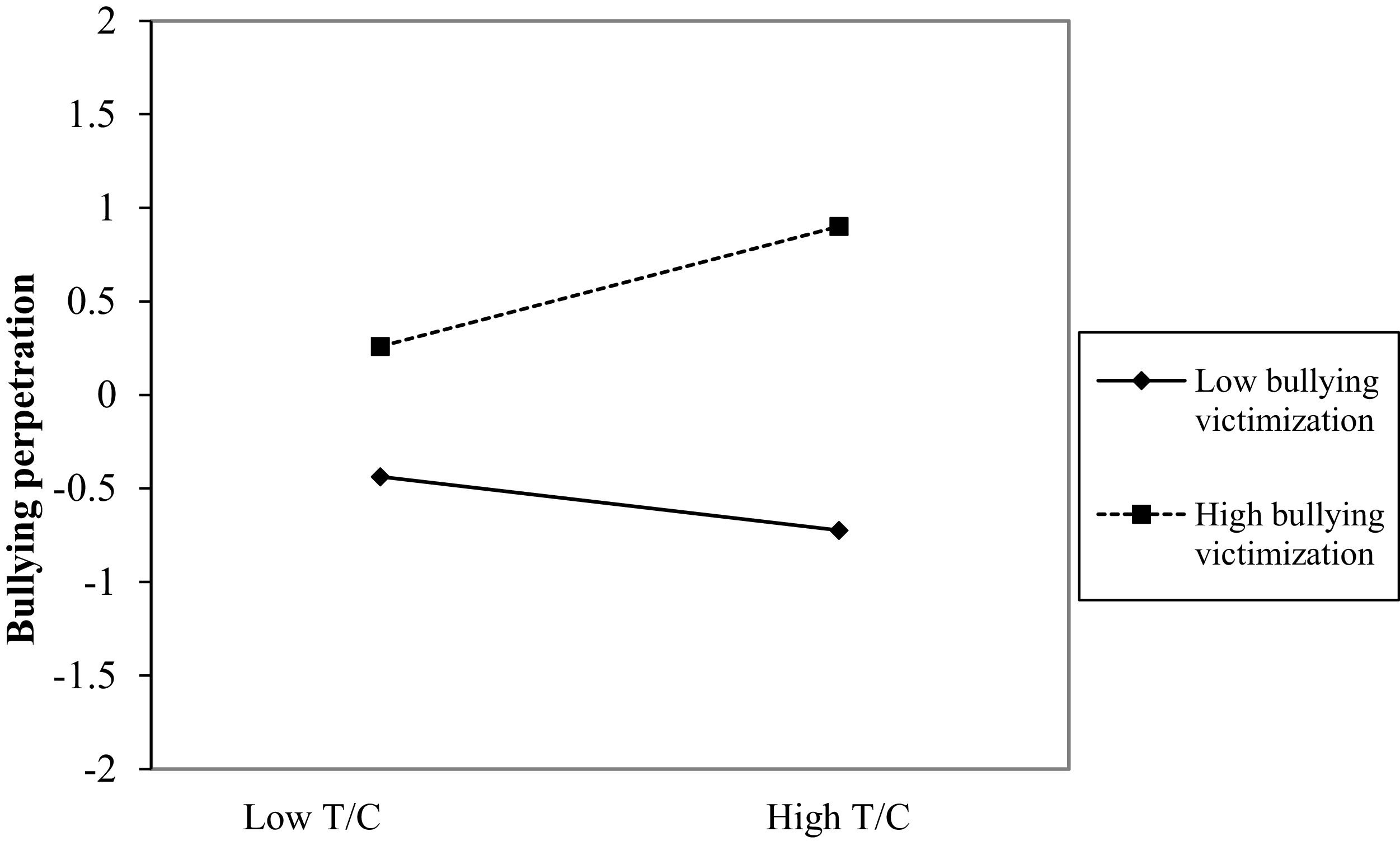
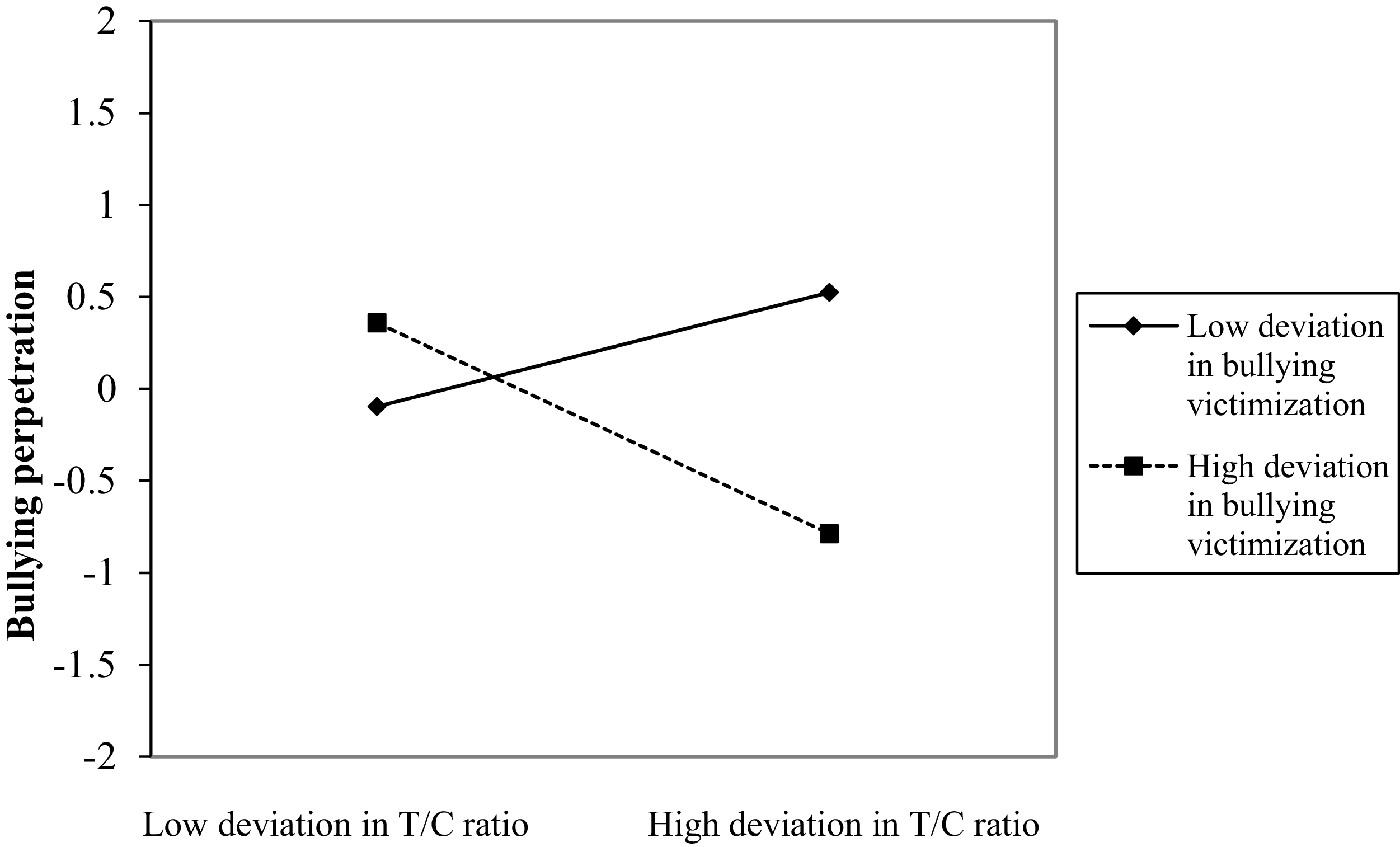
Figure 6 displays the associations between the latent variables in Model 11, and Table S4 includes all the longitudinal coefficients of the model. At the between-person level, the RI for bullying perpetration was significantly associated with victimization and with the testosterone/cortisol × victimization interaction. Figure 7 illustrates that bullying perpetration levels are higher among adolescents generally exposed to higher victimization levels. Additionally, the testosterone/cortisol ratio is associated with greater bullying perpetration at high levels of victimization (0.32, t = 2.08, p = .038). When general levels of victimization are low, the association between testosterone/cortisol ratio and bullying perpetration is not statistically significant (−0.14, t = −0.92, p = .356). At the within-person level, the testosterone/cortisol × victimization interaction predicted bullying perpetration from t–1 to t. Figure 8 shows this interaction. When both testosterone/cortisol and victimization deviated above usual levels, bullying perpetration decreased in the next wave (−0.57, t = −5.72, p < .001). Conversely, this association was positive when victimization deviated below the usual level (0.31, t = 3.14, p = .002).
Structural testosterone/cortisol model. Note: The figure reports standardized regression coefficients and covariances. Dotted lines represent marginally significant regression paths. *p < .05; **p < .001. T = testosterone, C = cortisol, W = wave.

Testosterone/cortisol ratio × victimization interaction for bullying perpetration at the between-person level. Note: T = testosterone, C = cortisol.

Testosterone/cortisol ratio × victimization interaction for bullying perpetration at the within-person level. Note: T = testosterone, C = cortisol.

Post Hoc power analysis
We used Monte Carlo simulation with MPLUS (Muthén & Muthén, Reference Muthén and Muthén2002) to examine the power of the main coefficients of the study. Essentially, this strategy involves specifying the coefficients obtained in the sample as if they represented true population values, and then drawing a large number of samples of the same size from that population. The coefficients are then estimated in each simulated sample. Based on this information, a probability statement can be made regarding the proportion of all replications in which a given path coefficient reached statistical significance at the 0.05 level. This proportion represents the empirical power estimate. The Monte Carlo procedure was applied to both the model based on the testosterone × cortisol interaction and the model based on the testosterone/cortisol ratio. Using the estimates from the final model of the study (Model 5), we specified a model as true for the population and generated 600 replications. We then estimated the proportion of replications in which the path coefficients for the testosterone × bullying victimization interaction (β = −0.44) and the testosterone × cortisol × victimization interaction (β = 0.36) reached significance at the 0.05 level. According to this procedure, the power for these parameters was 97% and 99%, respectively. We followed the same procedure for the final testosterone/cortisol ratio model (Model 11). The estimated power for the testosterone/cortisol ratio × victimization interaction predicting bullying perpetration (−0.22) was 96%.
Discussion
Grounded in the GAM framework (Allen et al., Reference Allen, Anderson and Bushman2018; Anderson & Bushman, Reference Anderson and Bushman2002), this study investigated the joint role of testosterone and cortisol in adolescents’ bullying perpetration toward peers. The central hypotheses proposed that experiences of peer victimization would amplify the influence of testosterone, as well as the combined effect of testosterone and cortisol, on aggressive bullying behaviors. Unlike previous research, this study adopted a RI-CLPM methodology (Hamaker et al., Reference Hamaker, Kuiper and Grasman2015), which allows for the separation of between-person (i.e., trait-like) effects from within-person effects.
Overall, the results obtained at the between-person level supported our hypotheses and were highly consistent for the testosterone × victimization and testosterone/cortisol ratio × victimization interactions. In both cases, it was observed that adolescents generally characterized by higher levels of testosterone or higher testosterone/cortisol ratios – that is, by a predominance of testosterone relative to their low cortisol levels – and by high levels of victimization tended to exhibit higher levels of bullying perpetration. By contrast, both testosterone level and testosterone/cortisol ratio were not associated with bullying perpetration among adolescents who were generally not exposed to victimization.
However, the within-person results provided mixed support for the study’s hypotheses and provided a complementary view of the dynamics that take place between these variables over time. Although deviations in testosterone above the adolescents’ typical levels predicted decreases in bullying perpetration in the next wave, the interaction results qualified this finding by showing that the role of testosterone was moderated by levels of cortisol and victimization. Thus, findings at the within-person level supported the general hypothesis that victimization experiences create contexts in which combinations of testosterone and cortisol may have different effects on aggression. However, these mechanisms involved complex patterns of associations among the variables, which did not completely match our predictions. The triple interaction testosterone × cortisol × victimization revealed that when cortisol increased relative to the adolescents’ usual levels, the increase in testosterone did not predict changes in bullying perpetration in the next wave. This finding is consistent with the proposal of the DHH (Mehta & Josephs, Reference Mehta and Josephs2010), which states that testosterone is not associated with aggression when cortisol is high. By contrast, when cortisol decreased below the adolescents’ typical levels, an increase in testosterone predicted increases in bullying perpetration in the next wave. However, this effect was moderated by experiences of bullying victimization. When victimization levels were below the adolescents’ average, an increase in testosterone predicted more bullying perpetration in the next wave. Conversely, when victimization levels were above average, an increase in testosterone predicted less bullying perpetration in the next wave.
A tentative interpretation of the findings above points to a profile of cold aggression toward peers. This profile would involve adolescents experiencing increases in testosterone (i.e., greater motivation for social status and reward seeking) and decreases in cortisol (i.e., lower levels of negative affect and stress), which – when peer victimization is relatively low – are more likely to reinforce their role as bullies. The same hormonal profile, in situations of relatively increased peer victimization, might reduce aggression because adolescents could strategically assess that such bullying perpetration behaviors are not advantageous, aiming to avoid negative consequences. In other words, the combination of increased testosterone and decreased cortisol in the context of rising victimization by others might activate regulatory or inhibitory processes that dampen aggression. This could reflect a strategic adaptation in which adolescents refrain from escalating aggression in already threatening contexts, potentially to avoid further retaliation or social costs. By contrast, in low-threat contexts, the same hormonal changes may increase the likelihood of engaging in aggressive behavior.
The results obtained for the testosterone/cortisol ratio at the within-person level were very similar. They showed that when bullying victimization was below usual levels, a relative increase in the testosterone/cortisol ratio (i.e., a relative predominance of testosterone over cortisol) predicted an increase in bullying perpetration in the next wave. However, when victimization was above usual levels, the increase in testosterone/cortisol ratio predicted a reduction in bullying perpetration in the next wave. Thus, although the testosterone × cortisol model has been proposed as preferable over the testosterone/cortisol ratio model (Calvete & Orue, Reference Calvete and Orue2024; Cima et al., Reference Cima, Meulenbeek, Spagnuolo, Oosterink, Thijssen, Smeijers, Valenzuela Pascual, Mouratidis, Oosterling, Riem and Loheide-Niesmann2025), in the current study, both methods obtained similar findings.
Moreover, although our study did not specifically aim to examine how changes in hormones predicted changes in bullying victimization, we found that the within-person effect observed for victimization in the testosterone × cortisol model was parallel to that observed for bullying perpetration. The triple interaction testosterone × cortisol × victimization predicted changes in future victimization. When the adolescents experienced an increase in both testosterone and cortisol relative to usual levels, the increase in testosterone did not predict changes in victimization. However, when the relative increase in testosterone was accompanied by a reduction in cortisol, the effect on future victimization depended on prior victimization levels. Specifically, when victimization had recently decreased below usual levels, the increase in testosterone predicted higher victimization in the next wave. This may be due to the adolescents also increasing their aggressive behaviors, which, in turn, may provoke defensive or retaliatory reactions from others. By contrast, when the increase in testosterone and reduction in cortisol occurred in the context of increased victimization, the increase in testosterone predicted fewer victimization experiences in the next wave. This may be due to a reduction in the adolescents’ aggression in that highly threatening context, which then leads to fewer retaliatory reactions from peers.
The similarities between the mechanisms observed for bullying perpetration and bullying victimization are consistent with the well-established overlap between these two variables (Morea & Calvete, Reference Morea and Calvete2022; Park & Cho, Reference Park and Cho2021). In this study, we found a strong association between aggression and victimization at the between-person level. The contemporaneous association between both variables at W1 at the within-person level was also high.
Comparing the findings of the present study with those of previous research is difficult, as this is one of the few longitudinal studies conducted with adolescents to test the DHH (see, for exceptions, Calvete & Orue, Reference Calvete and Orue2024; Susman et al., Reference Susman, Peckins, Bowes and Dorn2017) and, to our knowledge, the first to use an RI-CLPM approach. As mentioned previously, RI-CLPM offers a key advantage over traditional CLPM, namely, the ability to disentangle within-person processes from between-person stable differences (Hamaker et al., Reference Hamaker, Kuiper and Grasman2015; Osborne & Little, Reference Osborne, Little and Little2024). In this study, fluctuations above or below an adolescent’s typical level of victimization experiences modulated the effects of deviations in testosterone and cortisol on future aggressive behavior. Future research using RI-CLPM is needed to examine whether these findings can be replicated in other adolescent samples.
Finally, although the gender invariance of the models could not be tested, some differences were identified as a function of adolescents’ gender. Girls showed lower levels of T than boys, which is consistent with the greater increase in T observed among boys during adolescence (Harden et al., Reference Harden, Kretsch, Tackett and Tucker-Drob2014; Shirtcliff et al., Reference Shirtcliff, Allison, Armstrong, Slattery, Kalin and Essex2012). In addition, in line with previous studies (Cosma et al., Reference Cosma, Bjereld, Elgar, Richardson, Bilz, Craig, Augustine, Molcho, Malinowska-Cieślik and Walsh2022; Kretschmer et al., Reference Kretschmer, Veenstra, Dekovic and Oldehinkel2017), girls reported a lower frequency of bullying perpetration. These gender differences in bullying perpetration have been attributed to macrosocial factors. For example, Kretschmer et al. (Reference Kretschmer, Veenstra, Dekovic and Oldehinkel2017) proposed that bullying perpetration is normatively masculine and is considered deviant among girls. Furthermore, a cross-national study examining gender differences in bullying found that such differences in traditional bullying perpetration were larger in countries with greater gender inequality (Cosma et al., Reference Cosma, Bjereld, Elgar, Richardson, Bilz, Craig, Augustine, Molcho, Malinowska-Cieślik and Walsh2022). One possible explanation is that a central motivation for bullying perpetration is the enhancement of social status and dominance, which may be less socially valued for girls in societies where gender inequality persists.
Regarding the marginally significant findings, girls tended to show higher C levels than boys, which may reflect gender differences in stress reactivity during adolescence, although findings regarding C differences are not entirely consistent (Ordaz & Luna, Reference Ordaz and Luna2012). In addition, girls reported a lower frequency of victimization, which is consistent with their lower scores on bullying perpetration and the reciprocity between perpetration and victimization (Morea & Calvete, Reference Morea and Calvete2022). These findings provide additional context for understanding gender differences in the biological and social correlates of bullying involvement, but they should be interpreted with caution given their marginal significance.
Strengths, limitations, and implications for future research
This study has several strengths, including the novel application of an RI-CLPM approach in this area of research and the participation of a large adolescent sample across three waves. However, the study also presents limitations that provide opportunities for future research. A major limitation concerns the time interval between assessments. We selected a 3-month interval as a compromise between theoretical relevance and feasibility. Hormonal activity and social experiences in adolescence can vary substantially over several weeks to months, and this timeframe was expected to be sensitive enough to detect within-person changes while reducing the participant burden of responding many times. Nevertheless, although our intervals were shorter than the 6-month intervals used in other prior longitudinal studies (e.g., Calvete & Orue, Reference Calvete and Orue2024; Susman et al., Reference Susman, Peckins, Bowes and Dorn2017), it may still be too long to capture meaningful short-term fluctuations in the study variables (Grebe et al., Reference Grebe, Del Giudice, Thompson, Nickels, Ponzi, Zilioli, Maestripieri and Gangestad2019). Therefore, future studies should consider examining within-person changes in hormonal levels, victimization, and aggression using much shorter intervals – for example, by employing techniques such as ecological momentary assessment (Stone et al., Reference Stone, Schneider and Smyth2023). While the use of intensive longitudinal methods poses challenges, especially when it involves the collection of salivary samples, such an approach offers more valuable insights by capturing short-term dynamics across variables. Similarly, experimental studies in which participants are exposed to conditions of victimization and/or provocation while their aggressive behaviors and hormonal levels are monitored could also be valuable for clarifying causal relationships between variables (Bochon et al., Reference Bochon, Bird and Watson2023).
Another important limitation is the potential omission of key variables that may moderate the effects of testosterone and cortisol. For example, the triple imbalance hypothesis of reactive aggression (Van Honk et al., Reference Van Honk, Harmon-Jones, Morgan and Schutter2010) proposes three types of imbalances that contribute to explaining reactive aggressive behavior. The first is a subcortical imbalance between testosterone and cortisol, which is consistent with the DHH. This subcortical imbalance may lead to a cortical–subcortical imbalance, weakening the connections between the cortical and subcortical brain regions. The third imbalance is cortical and, according to the model, indicates that reactive aggression differs from proactive aggression in being associated with lower serotonergic function and more pronounced left-sided frontal brain asymmetry, especially in contexts of threat or provocation. Thus, future studies should consider assessing cortical-level changes, which may provide complementary explanations for the results of the present study.
The fourth limitation is the lack of peer-reported measures. In this study, both victimization and bullying perpetration were measured exclusively by means of self-reports. Although the anonymity of the responses was expected to promote disclosure, future studies should combine self-reports with peer reports.
Finally, it would have been important to assess whether the associations between hormones and behaviors were similar for boys and girls. Sex differences in both bullying perpetration and victimization may shape how testosterone and cortisol relate to these behaviors. Boys seem to be more prone to direct aggression, whereas girls show marginally higher – but small – levels of indirect aggression (Casper & Card, Reference Casper and Card2017), indicating that indirect aggression is largely comparable across sexes. During adolescence, boys also show higher levels of testosterone (Susman et al., Reference Susman, Peckins, Bowes and Dorn2017), while girls often present higher cortisol responses to interpersonal stress (Hollanders et al., Reference Hollanders, van der Voorn, Rotteveel and Finken2017). These differences suggest that the interplay between testosterone, cortisol, and bullying involvement might not operate identically in boys and girls. However, in this study, the size of the male and female subgroups was not large enough to test the model in each subgroup. Future research should aim to use larger samples that allow for multiple-group comparisons.
In conclusion, this study advances our understanding of how testosterone and cortisol jointly influence adolescent aggression, suggesting that, consistent with the GAM, situational experiences of victimization modulate their effects on bullying perpetration. By integrating both between-and within-person perspectives, the study demonstrates that increases in cortisol levels weaken the effect of testosterone on subsequent peer aggression. At the same time, the effect of decreases in cortisol depends on adolescents’ experiences of bullying victimization. Specifically, under conditions of reduced cortisol, testosterone predicted greater bullying perpetration when victimization was low, and lower bullying perpetration when victimization increased. These findings suggest the existence of a cold aggression profile toward peers, characterized by flexible strategies that vary depending on the social context.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0954579426101503.
Data availability statement
Data and code are available upon reasonable request from the authors.
Acknowledgments
The authors thank the schools and adolescents who participated in the study.
Funding statement
This research was supported by a grant from the Knowledge Generation Projects 2022 of the Spanish State Research Agency (ref. PID2022-140773NB-I00) and the Basque Country Government (IT1532-22).
Competing interests
The authors declare that they have no competing interests.
Pre-registration statement
This study was not pre-registered. The research questions and main analyses were defined prior to data analysis; however, no formal pre-registration document was created. We report all measures, conditions, and analyses conducted.





 Open access
Open access